investigating the effects of polychlorinated biphenyls on circulating oxytocin levels, area of the...
TRANSCRIPT

INVESTIGATING THE EFFECTS OF POLYCHLORINATED BIPHENYLS ON CIRCULATING OXYTOCIN LEVELS, AREA OF THE PARAVENTRICULAR NUCLEUS
AND SOCIAL BEHAVIOR IN JUVENILE MALE RATS
Banafsheh Jolous-Jamshidi
A Thesis
Submitted to the Graduate College of Bowling Green State University in partial fulfillment of
the requirements for the degree of
MASTER OF SCIENCE
August 2007
Committee:
Lee Meserve, Advisor
Howard Casey Cromwell
Roudabeh Jamasbi

ii
ABSTRACT
Lee Meserve, Advisor
Social recognition is a class of behavior that is responsible for pair bonding and
attachment to conspecifics. Neural circuits responsible for bringing about this behavior are
present in the central and medial amygdala as well as the ventromedial hypothalamus (VMH)
which contains oxytocin receptors. Oxytocinergic neurons from the paraventricular nucleus
(PVN) and pre-optic area of the brain project to these areas and have been shown to modulate
social recognition. Poly-chlorinated biphenyls (PCB) are a class of environmental toxicants that
can exert effects at both the cellular and behavioral levels. They exert their toxic effects on
almost every part of the body including the brain. In the present study, the effect of a mixture of
PCB 47 (2, 2’, 4, 4’- tetrachlorobiphenyl) and PCB 77 (3, 3’, 4, 4’- tetrachlorobiphenyl) on
circulating oxytocin levels, change in the area of the PVN and social recognition in juvenile male
Spraque-Dawley rats was examined. Rat offspring were exposed to low doses of PCB (12.5 and
25 ppm) via the maternal diet perinatally and postnatally through lactation. Social recognition
experiments were conducted in juvenile rats under red light on PND 21 using connected social
and non-social boxes. The difference in investigation time between social and non-social box
suggests that doses of PCB 12.5 do not cause significant changes in social recognition as
compared to that in controls. However, PCB 25 results in a significant difference as compared to
controls in the second trial for mean social investigation time and mean difference in
investigation time between social and non-social box. Neither dose of PCB significantly altered
either PVN area or circulating level of oxytocin. These results could indicate that low doses of a

iii
mixture of PCB 47 and 77 do not significantly affect the morphology of brain areas and
peripheral oxytocin influencing social recognition.
Key Words: polychlorinated biphenyls (PCB), PVN, social recognition, oxytocin levels

iv
DEDICATION
To my family, who has always been a source of strength and support.

v
ACKNOWLEDGMENTS
Special thanks to Drs. Lee A. Meserve, Howard Casey Cromwell, Roudabeh J. Jamasbi, who
have always guided and supported me in my research endeavors. Thanks to my labmates
specially Avanti Desai, Dena Krishnan and Ashley McFarland for the help rendered during
various phases of my research.

vi
TABLE OF CONTENTS
Page
REVIEW OF LITERATURE ................................................................................................ 1
SUMMARY........................................................................................................................... 19
MATERIALS AND METHODS........................................................................................... 20
RESULTS…………… .......................................................................................................... 26
RESULTS SUMMARY......................................................................................................... 32
DISCUSSIONS……….......................................................................................................... 33
FUTURE DIRECTIONS ....................................................................................................... 43
REFERENCES……… .......................................................................................................... 45
APPENDIX…………............................................................................................................ 52

vii
LIST OF FIGURES
Figure Page
1 The structure of polychlorinated biphenyl................................................................. 3
2 Different kinds of Social Recognition Paradigms used to measure social recognition...12
3 Proposed mechanisms for the social recognition in the brain.................................... 12
4 A sagittal section of the rat brain showing the location of the PVN.......................... 14
5 Test set-up for measurement of social recognition .................................................... 22
6 The area of the PVN measured using the Motic Image software .............................. 23
7 Mean investigation time in 21-day-old Sprague-Dawley rats between control, PCB 12.5,
and PCB 25 groups in first, second, and third trials .................................................. 27
8 Mean difference in investigation time between social-non social box across trials in
control, PCB 12.5, and PCB 25 Sprague Dawley rats at the age of 21 days ............. 27
9 The normalized PVN area for the control (n=10), PCB 12.5 (n=10), and PCB 25 (n=10)
fed rats ............................................................................................................ 30
10 The curve showing the value B/Bo against the standard oxytocin concentration
(pg/mL)………. ......................................................................................................... 31

viii
LIST OF TABLES
Table Page
1 Mean investigation time (±SEM) in non-social box for trials with unfamiliar
(Trial 1), familiar (Trial 2) and second unfamiliar (Trial 3) conspecifics in
control (n = 18), PCB 12.5 (n = 15), and PCB 25 (n = 15)…………………………….28
2 Mean and the mean±SEM for the number of entries into the social box for all three trials
within control (n = 18), PCB 12.5 (n = 15), and PCB 25 (n = 15) ............................ 28
3 Mean and the mean±SEM for the number of entries into the non- social box for all three
trials…….................................................................................................................... 29
4 Mean and the mean±SEM for the number of entries into the social + non- social box for
all three trials ...................................................................................................……. 29

REVIEW OF LITERATURE
Polychlorinated biphenyls (PCBs) are a class of persistent environmental chemicals that
are known to be severe health hazards for animals and humans (Hellbach et al., 1998). There are
no known natural sources of PCB. These are man-made chemicals, which were introduced into
the environment as early as 1929 (Lind et al., 1999), and from that time till 1977, were being
released into the environment by industrial concerns. In 1977, the manufacture of PCB was
banned in United States because of increasing awareness of their health hazards. PCBs are either
oily liquids or solids that are colorless to light yellow. Some PCB can exist as vapor in air. They
are without taste or smell. As a result, their entry into the body goes unnoticed. Many
commercial PCB mixtures are known by their commercial designation Aroclor (Lind et al.,
1999), and they found commercial applications in electrical appliances because they show good
insulating properties. PCB has been used widely for 40 years in compressors and carbonless
carbon paper, and as insulators for transformers and capacitors (Hellbach et al., 1998). PCB
enters air, water, and soil during manufacture and continues to enter through improper disposal
of industrial wastes and accidental spills. PCB resists degradation in the environment and
therefore, persists for a very long time. They can travel long distances in the air and be deposited
in areas far away from where they were released. In water most PCB adhere to organic
compounds and bottom sediments. PCB can be consumed by fish and other aquatic animals and
can become a part of the complex food chain. As a result, humans can be directly or indirectly
affected by these toxicants.
PCB is a complex organic molecule formed by substituting varying numbers of chlorine
atoms on biphenyl rings (linked by a single carbon to carbon bond at the 1 and 1’ positions) (Lim

2and Young, 2006). There are ten hydrogen atoms on the biphenyl rings upon which can be
substituted chlorine atoms (with ten positions of chlorine substitution). Chlorine substitution may
occur in any or all of the 2-6 and 2’-6’ positions (Fig. 1). The number and position of chlorine
atoms can result in 209 individual chlorinated compounds, known as congeners. Of these 209
congeners, 135 congeners have been detected in environmental samples such as surface soil,
groundwater, and marine organisms (Safe, 1990). However, the number of congeners that fall
into the category of having both toxic properties and abundance in animal tissue is less than 50.
In addition, less than 25 of these congeners belong to the category which is mostly responsible
for PCB induced damage in invertebrate, fish, bird, and mammal tissue (McFarland and Clarke,
1989). Toxicological properties of these individual congeners depend upon the number and
position of chlorine atoms. The degree of increased lipophilicity depends upon increased
chlorination, which in turn also leads to less hydroxylation of PCB. Hydroxylation is a major
process whereby these congeners are metabolized in the body, and is selective for para and meta
positions (Safe, 1990). Some congeners have one or no chlorine atoms substituted in ortho
positions, and as a result, their biphenyl rings line up in a single plane. Their number is
approximately twelve. These coplanar configurations are similar to that of dioxins, and hence,
they are called dioxin-like chemicals, with toxicological profiles similar to those of dioxins
(Sanchez-Alsono et al., 2003).
Dioxins are a group of chemicals that are produced during industrial processes such as
waste incineration, chemical and pesticide manufacturing, and pulp and paper bleaching
(Tuomisto, 2004). The most toxic of all the dioxins is 2, 3, 7, 8- tetrachlorodibenzo-p-dioxin
(TCDD), the major contaminant in the herbicide, Agent Orange. TCDD is used as a frame of
reference to measure the toxicity level of the remaining dioxins (Van den Berg et al., 1998).

3Dioxin is widespread in terms of its distribution and causes endocrine disruption, reproductive
dysfunction, liver toxicity, and cancer.
Figure 1. The structure of polychlorinated biphenyl. It contains the two aromatic hydrocarbon rings with the various ortho (2,2’,6,6’), meta (3,3’5,5’), and para (4,4’) positions. (From: de.geocities.com/image/pcb-molecule.gif). PCB exerts its toxicity by three different mechanisms (McKinney and Waller, 1994).
These mechanisms might not act independently of each other, but they might be correlated. In
the first mechanism, PCB can reversibly interact with receptors, enzymes, or other regulatory
molecules. The second mechanism is by irreversibly binding covalently to target molecules like
proteins and DNA. The third mechanism is accumulation of metabolically stable PCB in adipose
tissues via their lipophilic properties. Toxicity of the PCB depends upon their reactivity which in
turn depends upon its structure. The SAR (structure activity relationship) has been used as an
approach to understand reversible mechanisms of binding of PCB congeners to specific
molecules (McKinney and Waller, 1994). Broadly speaking, PCB could be either coplanar or
non-coplanar. Coplanar PCB exerts a dioxin like activity (McKinney and Singh, 1981). In this
mechanism, there is a stacking kind of interaction between PCB and other aromatic molecules
such as the heme system in heme proteins (Ishida et al., 1986). Such interactions are also called
charge transfer or donor-acceptor interactions. In layman terms, they can be defined as “Velcro-
type” interactions because planar rings facilitate the sticking together of two pieces of the

4molecule. Another type of interaction is cleft-type interaction. Chlorine atoms in meta and para
position are highly polarized and they can undergo cleft-type of interaction. This interaction
happens between the highly polarizable lateral halogen and hydrophobic interior of the cleft of
the protein provided by amino acid side chains that converge on the halogen substituents. For
example, the chlorine atom in this position can interact with hydrophobic interior of the cleft that
contains amino acid side chains. Another structural feature could be the involvement of a vicinal
unsubstituted position in the molecules that can provide the sites for oxidative metabolism (Kato
et al., 1980). In this kind of mechanism areas of high electron density are places in the molecule
that are involved in chemical reactivity that control the rate and regional specificity of oxidative
attack. This leads to the formation of reactive intermediary metabolites. These reactive
metabolites can exhibit the same toxicity as PCB and can form covalently bond residues with
biomolecules (McKinney and Waller, 1994).
Modulation of biochemical and neurochemical functions during development under the
influence of PCB is a major area of research worldwide. PCB commonly affects adipose tissue,
skin, liver and muscle (Carpenter, 2006). Human exposure to PCB has resulted largely from
consumption of food, but occupational exposure also results from inhalation and absorption
through skin (Lind et al., 1999). Other targets for the entry of PCB include gastrointestinal tract,
immune system, and nervous system (Lind et al., 1999). PCB is also known to contaminate
ovarian follicular fluid and serum samples in women undergoing in-vitro fertilization, which in
turn affect the reproductive outcome (Jarrell et al., 1993). Some PCB congeners can act as
carcinogens in animals and they can enhance cancer causing properties of other chemicals.
PCB can act as general cancer promoters, for example they are not directly cancer
causing, but they enhance the effect of other carcinogenic agents by generation of free radicals

5and induction of expression of a number of genes (Tharappel et al., 2002). Sustained exposure to
PCB causes chromosomal aberrations (Silberhorn et al., 1990), which are mediated by the
metabolites of PCB. There are studies which provide evidence to the fact that dioxin-like PCB
congeners and dioxin itself can cause oxidative DNA damage (Oakley et al., 1996). There are
primarily two types of studies of cancer in human populations – occupational studies of workers
and case-control studies of individuals with cancer of a specific type (Carpenter, 2006). PCB
have officially been identified as “probable human carcinogens” by the World Health
Organization (WHO), based on the results obtained from animals and humans. Dioxins are
classified as “known human carcinogens” (Carpenter, 2006). However, there is no concrete
evidence for the involvement of PCB in carcinogenicity, because PCB can accumulate with
many other fat-soluble contaminants in the body. As a result, the cause of cancer could be PCB,
DDT, or any other fat-soluble chemicals present in the body. The problem with performing
occupational studies is that the sample size is relatively small. Therefore, the incidence in the
population will not be large, even if the risk is elevated owing to genetic predisposition to
carcinogenicity (Carpenter, 2006). Studies have shown prevalence of cancers of liver, gall
bladder, leukemia, gastrointestinal, skin (especially malignant melanoma), lymphoma, lung and
pancreas (Longnecker et al., 2003). Elevation in incidence is not equal for all kinds of cancers,
possibly because PCB interacts with different tissues differently (Carpenter, 2006). It has also
been reported that there is an elevated incidence of brain cancer in workers in capacitor plants
(Sink et al, 1992). There have been studies that correlate the prevalence of breast cancer with the
doses of PCB (Moysich et al., 1999). A correlation has been found between certain PCB
congeners and risk of breast cancer in individuals of certain racial ethnicity and from certain
gene pools (Carpenter, 2006).

6Mono-ortho congeners like PCB 28 and 118 show an increased risk of colorectal cancers
(Howsam et al., 2004). Animals exposed to PCBs have been reported to be diagnosed with liver
and biliary cancers (Kimbrough et al., 1975; Mayes et al., 1998). Aroclor 1254 and other PCBs
of similar chlorine content makes the lung susceptible to cancer (Beebe et al., 1993). Malignant
melanoma has also been observed in PCB-exposed workers (Longnecker et al., 2003).
Exposure to PCB in utero can impair the intellectual growth in infants and young
children. There have been studies by Jacobson and Jacobson (1996) to monitor whether the
effect of prenatal exposure to PCB in infants persists through school age. The sample size was
212 children from birth to 11 years of age. The effect was monitored by giving IQ and
achievement tests. The results revealed a reduction in the IQ of children fed breast milk
containing PCB at a concentration of 1,250 ppb (lipid adjusted). The performance was poor in
reading skill in children whose mothers had 1,000 ppb PCB or greater in their breast milk.
Hence, the survey concluded that PCB causes an irreversible decrement of IQ. Other than these
abnormalities, PCB also has the potential to induce hypothyroidism, infertility and reproductive
system disorders, cardiovascular disease (CVD) and elevated serum lipids, hypertension,
diabetes, liver disease, asthma and arthritis (Carpenter, 2006).
Concentration of PCB in living systems could reach as much as 1000 times greater than
that found in water. The most commonly observed health effects in adult humans exposed to
large amounts of PCB are skin conditions such as acne and rashes (Safe, 1994). Exposed workers
in factories show changes in blood and urine chemistry indicating liver damage. PCB exposure
in the general population is not likely to result in skin and liver effects. Most of the health
hazards observed from PCB exposure have been in women and children. Being lipophilic in
nature, they are readily transferred post-natally through breast milk. Women who are exposed to

7relatively high levels of PCB in the workplace or consume large amount of contaminated fish
have babies that weigh slightly less than those of women who are not exposed. PCB can cross
the placenta from mother to fetus, and exposure in the womb to chemicals could produce serious
and irreversible injury to the brain. Babies exposed in utero also show abnormal responses in
terms of their behavior. They demonstrate problems with motor skills and a decrease in short
term memory, lasting for several years (Freund–Mercier et al., 1988). Hence, there is a need to
investigate the mode of entry of PCB into the brain and the various areas that are damaged under
the effect of varying doses of PCB. Research on these areas could allow discovery of a new
treatment for people who have been exposed to acute or chronic doses of PCB.
Observations in animals have shown that there is increasing evidence that a number of
persistent organochlorine pollutants can alter endocrine homeostasis, thereby resulting in a toxic
effect in a particular organism (De Felip et al., 2004). In the case of classes of different
compounds (for example pesticides) or different congeners (e.g., PCB), there is a correlation
between the exposure to these toxicants and their effects. This correlation of damage with these
compounds is further complicated because of different structure-dependent endocrine activity.
For instance, PCB is known to show either estrogenic or antiestrogenic properties (Safe and
Krishnan, 1995). For example, coplanar PCB like 3, 3’, 4, 4’-tetrachlorobiphenyl (PCB 77; TCB)
exerts dioxin-like activity by interacting with the aryl hydrocarbon receptor (AhR) and the AhR-
dependent signal transduction pathways (Hemming et al., 1992; Safe, 1994), and are mainly
characterized by estrogenic activity (Seegal et al., 2005). Because of its resemblance to dioxin,
coplanar non-ortho substituted PCB congeners have a high toxic equivalent rating and are known
to have more toxicity than their non-coplanar counterparts (Safe, 1994; Van der Berg et al.,

81998). The toxicity results in neurodegeneration, estrogenicity, or anti-estrogenicity and tumor
production.
However, there are other studies, which conversely indicate the antiestrogenic activity of
coplanar PCB (Ramamoorthy et al., 1999). Non-coplanar PCB like 2, 2’, 4, 4’-
tetrachlorobiphenyl (PCB 47) induce Ah receptor independent mechanisms (Safe, 1994). Studies
have shown that non-coplanar congeners like PCB 47 reduce dopamine level and have
antiestrogenic activity (Seegal et al., 2005). These ortho-substituted PCB congeners are more
abundant in human tissues, and are known to affect the endocrine function in various ways,
could be of particular significance in determining overall interference with the hormone
homeostasis of an intact organism. They are endocrine disruptors, causing wide reaching
alterations in several endocrine systems which lead to the production of estrogens, androgens,
thyroid hormones, retinoids and corticosteroids (Brouwer et al., 1998). Cognitive impairment
and hyperactivity in infants exposed to PCB and related toxins have also been reported in many
studies. There is a hypothesis proposed by Kuroda et al., (2004) and Jacobson and Jacobson
(1996) that spatio-temporal disruption of developing neuronal circuits by PCB exposure can
cause the co-morbidity of learning disorders (LD), attention deficit hyperactivity disorder
(ADHD), and autism with the co-exposure to other environmental chemicals. The neurotoxic
effects of PCB on the infant and the adult brain have been shown by monitoring the effects of
PCB on the synaptosomal preparations from the various areas of the brain. Alteration of
concentration of various kinds of neurotransmitters like dopamine (DA) and its metabolites has
been detected in tissues using HPLC (Seegal et al., 2005). The neurotoxic effects of PCB are
exerted not alone, but in synergy with other toxins such as methyl mercury (MeHg) (Lind et al.,
1999).

9Social recognition is the ability to recognize a conspecific as being familiar. Depending
upon the kind of activity that the individual indulges in with a conspecific, it might be required to
remember gender and reproductive status of the conspecifc at times or important details like the
conspecific’s kinship and its position in the social hierarchy (Ferguson et al., 2002). The second
type of information is required for all animals that live in complex social systems where they
must encode and recall specific information about their conspecifics. Humans and other primates
rely on visual and auditory cues for individual recognition. Subjects with lesions to areas like the
fusiform gyrus, a subdivision in the brain critical for face recognition, show prospagnosia, or an
inability to recognize faces (Grill-Spector et al., 2004).
In monkeys, neurons in the temporal cortex fire in relation to the familiarity of an
individual (Perrett et al., 1984). It is the olfactory and pheromonal signals which play an
important role in social recognition in most mammals, though one cannot deny that the auditory
and visual sensory modalities play an important role as well (Popik et al., 1991).
Regardless of the kind of sensory modality responsible for social recognition, the
common outcome of this behavior is enhancement of reproductiveness and survival of individual
species. Social recognition could be both long term and short term. Rodents can form short term
memories of recently encountered individuals. The short term memory for juvenile rats is around
eight minutes and that for adult rats is around thirty to sixty minutes (Thor and Holloway, 1982).
Social memory is a process which involves introduction of an unfamiliar conspecifc for
the first time into the home cage of a rat or mouse. It is observed that a male rat vigorously
investigates a novel rat. Introduction of a novel animal for the second time into the cage triggers
an anogential response from a male rat, which is the same as that in the first trial if the same
animal is introduced to a resident rat in a second encounter. The investigation time of a male rat

10in the second trial is less than that in first trial (Thor and Holloway, 1981). Hence, the differences
in investigation times during repeated pairing with a novel animal can be used as an index for the
short term memory in rat. In adult male rats, if the inter-exposure interval exceeds 30-60
minutes, then, there will be no change in investigation time of male rat towards either a familiar
or a novel rat indicating that the resident male rat is unable to recognize the stimulus animal
because of the disruption of short term memory of adult rat. The same event, if performed on a
juvenile rat will show the disruption after a span of around eight minutes. If a novel animal is
presented repeatedly, the duration of the memory can be extended. It can also be impaired if
another novel animal is introduced between the encounters to the original con-specific (Dantzer
et al., 1987; Sekiguchi et al., 1991). The specificity of the effect of social recognition can be
tested by presenting a novel stimulus. This will ensure that the treatment shall specifically
change social recognition rather than change the investigation time or motivation to interact with
con-specific (Ferguson et al., 2002). This type of interaction is just one aspect of social
recognition. Recognition of mates and kin involves long term memories which might last days,
weeks or even months. These processes may involve the cognitive and neural systems that are
distinct from those involved in short-term memory recognition. Three other variations of social
recognition have been suggested (Fig. 2). In the first social recognition paradigm, a male rat is
introduced to a novel juvenile during the first 5 min trial and the duration of olfactory
investigation is measured. The second type of social recognition paradigm is when the subject is
given a choice between the previously encountered and a novel conspecific. The difference is
monitored in the time spent investigating between the familiar and novel stimulus animal. The
original paradigm seems to work best with retired breeders; the modified paradigm is more
applicable to assess recognition between sexually active males and females. Another paradigm

11that seems to work best with rapid high-throughput pharmacological or phenotypic screening
studies is the habituation-dishabituation paradigm (Dluzen and Kreutzberg, 1993; Winslow and
Camacho, 1995). In this paradigm, the test subject is repeatedly exposed to the same restrained
animal for 1 min with 10 min intertrial interval. The familiarity is detected by reduction in
investigation time of the test rat toward the conspecific on each trial. After the fourth exposure, a
novel stimulus is introduced to rule out the possibility that the reduced investigation is not
caused by habituation or fatigue. Since, the effect of the drug on the behavior is manifested after
some time; this paradigm works best for such studies.
A neural mechanism (Fig. 3) has been proposed for social recognition (Choleris et al.,
2004). Based on this mechanism, the volatile and non-volatile olfactory cues from the individual
conspecific are taken up by the vomeronasal organ and the main olfactory bulb system. These
sensory stimuli cause depolarization of the afferent neurons at the olfactory bulb which synapse
at amygdala. The estrogen from blood stream binds to estrogen receptor alpha in the amygdala
and estrogen receptor beta in the hypothalamus, paraventricular nucleus (PVN) and supraoptic
nucleus (SON). This leads to the synthesis of oxytocin from these areas which eventually lead to
binding of oxytocin to the oxytocin receptors in amygdala. The whole cascade brings about
social recognition. This mechanism could apply to all three kinds of social recognition paradigm
discussed above.

12
Figure 2. Different kinds of Social Recognition Paradigms used to measure social recognition. (a) This paradigm was described by Thor and Holloway (1982). (b) The social recognition paradigm described by Engelmann. (c) The habituation-dishabituation paradigm described by Winslow and Dluzen (Ferguson et al., 2002).
SOCIAL
RECOGNITION
Figure 3. Proposed mechanisms for the social recognition in the brain (Choleris et al., 2004).

13 The paraventricular nucleus (PVN) is a triangular group of large neurons in the
periventricular zone of the anterior half of the hypothalamus and is functionally associated with
the posterior lobe of the pituitary (Kenney et al., 2003). It is an aggregation of neurons in the
hypothalamus, located adjacent to the third ventricle. The PVN is highly vascularized, but is
inside the blood brain barrier, although the neuroendocrine neurons extend their projections to
the sites which are outside the blood brain barrier. The PVN contains magnocellular
neurosecretory cells whose axons extend into the posterior pituitary, parvocellular
neurosceretory cells that project to the median eminence, and several populations of peptide-
containing cells that project to many different brain regions (Kenney et al., 2003). The chief
function of magnocellular cells in the PVN is to produce the peptide hormones/neuromodulators
oxytocin and vasopressin. These peptide hormones are processed from large precursors while
packaged in large dense core vesicles. These large dense core vesicles are transported down the
axons and released into the circulation from the neurosecretory nerve terminals in the posterior
pituitary gland. Similar magnocellular neurons are found in the supraoptic nucleus. Parvocellular
neurosecretory neurons of the PVN project axons to the median eminence at the base of the
brain. At the median eminence, neurosecretory nerve terminals release hypophysiotropic
peptides into the blood vessels of the hypothalamo-hypophyseal portal system. These vessels
carry the peptides to the anterior pituitary gland, where they regulate hormone secretion into the
systemic circulation (Freund-Mercier et al., 1988). Parvocellular neurosecretory cells include
cells that produce corticotrophin releasing hormone, which stimulates ACTH secretion from
anterior pituitary gland, and cells that produce thyrotropin releasing hormone which stimulates
TSH secretion.

14
Figure 4. A sagittal section of the rat brain showing the location of the PVN (From: http://meds.queensu.ca/~ferguson/FergusonLab3/images/pvn.ht3.gif)
Oxytocinergic neurons are present in the paraventricular nucleus of the hypothalamus and
become activated by estrus female odor and sexual contact in sexually naïve and experienced rats
(Nishitani et al., 2004). Oxytocin is a neuropeptide also synthesized in the supraoptic nucleus of
the brain. Oxytocin plays an important role in the reproductive behavior and physiology of
mammals. This hormone facilitates behavioral phenomenon of nest building and pup retrieval in
rats, acceptance of offspring in sheep, and the formation of adult pair-bonds in prairie voles. In
mammals, oxytocin stimulates milk ejection during lactation, uterine contraction during birth,
and is also released during sexual orgasm in both man and women. A central role of oxytocin
systems is maternal affiliation in preweaning rats. Some forms of social behaviors that are
mutually solicited and cohesive, that is essential for survival of individuals and continuation of
species, are also influenced by oxytocin levels. The oxytocin of parvocellular cells of the PVN is
responsible for many behavioral and cognitive functions such as parental behavior, aggression,
learning and memory (Ferguson et al., 2001 b). Regulation of oxytocin synthesis is through the
binding of estrogen on estrogen receptor beta (Patisaul et al., 2003).

15 The PVN also contains interneurons and populations of neurons that project centrally.
The centrally projecting neurons include parvocellular oxytocin cells that project to the
brainstem (involved in gastric reflexes) and spinal cord (involved in penile erection). The
parvocellular vasopressin cells project to areas of the hypothalamus and the limbic system, as
well as to the brain stem and spinal cord, and are involved in blood pressure regulation and
thermoregulation. Finally, the parvocellular CRF neurons are thought to be involved in stress-
associated behaviors (Kenney et al., 2003).
Afferent inputs to the PVN include neurons in structures adjacent to the anterior wall of
the third ventricle (“AV3V region”) carrying information about electrolyte composition of the
blood, and about circulating concentrations of hormones such as angiotensin and relaxin to
regulate magnocellular neurons. Inputs from the brain stem nucleus of the solitary tract and the
ventrolateral medulla carry information from the heart and stomach. The stress response
regulators are the inputs from the hippocampus to the CRF neurons (Kenney et al., 2003).
Oxytocin is produced by the supraoptic nucleus and PVN of the hypothalamus.
Physiological actions of oxytocin, such as facilitation of milk-ejection reflex or induction of
behavioral changes are mediated via receptors, which have been localized in vitro in the adult rat
brain by an autoradiography technique (Ferguson et al., 2000). Also, oxytocin receptors are
found in other parts of the body like thymus, spleen and lymphocytes (Elands et al., 1990).
Oxytocin containing neurons have been demonstrated in various brain areas in which oxytocin is
thought to act as a neurotransmitter or a neuromodulator.
The evidence that oxytocin (OT) plays a role in social recognition is more
straightforward and well explained as compared to the role played by vasopressin. The oxytocin
neurons from PVN send projections to many regions within the brain, including the

16hippocampus, amygdala and hypothalamus where oxytocin is involved in the affiliative
behaviors, the behaviors which involve close interaction or attachment with conspecific (Bale et
al., 2001). These affiliative behaviors are grouped into one of the many subdivisions of social
behaviors known so far. Oxytocin receptors (OTRs) are found in many regions of the brain,
including the central nucleus of the amygdala (cAmyg) and the ventromedial nucleus of the
hypothalamus (VMH). These areas of the brain are very important components for the neural
circuits which regulate distinctive behavioral processes. Central amygdala is a critical component
of the limbic system that induces fear, anxiety responses and pair bonding. Experiments by
Ferguson et al., (2001) has shown the induction of Fos-IR in the amygdala of the wild type
mice but not in oxytocin knock out mice during initial exposure to the social recognition task,
suggesting an involvement of this region for the initial pair bonding.
The initial thinking was that OT and AVP have opposing effects on social memory. The
AVP lead to facilitation and OT caused attenuation of learning and memory. There was
attenuation in social recognition when OT was administered peripherally in high doses.
Administering physiological levels of OT demonstrated that the peripheral administration of OT
at low doses facilitated social recognition in male rats. This suggests an inverted U-shaped dose
response curve for the effect of oxytocin on social recognition. The same kind of curve has also
been obtained when OT was administered centrally (icv) in male rats. It has also been
documented in previous studies that the facilitating or attenuating effects of oxytocin depend
upon the part of the molecule that binds to the receptor. Hence, OT has different mechanisms to
facilitate and attenuate social recognition. The role of OT in social recognition in females is
different from that in males. Central administration of OT in females did not facilitate social
memory. Hence, OT in males facilitates social recognition but not in females. This suggests a

17sexual dimorphism with respect to the roles of oxytocin in males and female (Isadora et al.,
2004).
Individual recognition is a common trait of all mammals. Social memory unlike other
forms of memory has a neural basis. Mammals are skillful in recognizing signatures of their
conspecifics through one or more of their sensory modality such as vision, olfaction and acoustic
stimuli. In rodents, it is the olfactory signature that predominantly determines social recognition.
The ability to determine healthy conspecifics determines the reproductive success of the
individual (Ferguson et al., 2000). Studies have shown that in the brain oxytocin (OT) and
vasopressin (AVP) seem to modulate a range of social behaviors from maternal care to mate
guarding. Injecting doses of AVP tends to facilitate social memory whereas injecting of oxytocin
may have a facilitatory effect or an inhibitory effect on social memory depending on the dose,
route and paradigm. The same studies have shown that the male mice mutant for the oxytocin
gene (Oxt -/-) failed to develop social memory, whereas wild type (OXT +/+) mice showed
intact social memory. The measurement of olfactory foraging and olfactory habituation tasks
indicated that the olfactory detection in non-social stimuli is intact in Oxt -/- mice. The
somatosensory stimulation during breast feeding or suckling increases the maternal oxytocin
levels. It could also be released by non-noxious stimuli such as touch and warm temperature in
plasma and cerebrospinal fluid (Moberg, 1998). In both male and female rats, oxytocin exerts
potent physiological anti-stress effects. Oxytocin released in response to social stimuli may be
part of a neuroendocrine substrate which underlies the benefits of positive social experience.
This could be an explanation of health promoting effects of oxytocin such as positive social
interaction and emotions. Owing to the relation of oxytocin to the psychological states or
imagery, oxytocin may also mediate the benefits caused owing to hypnosis and meditation.

18There have been studies in the past which suggest the involvement of environmental
toxicants including PCB on the social behaviors in various kinds of animals. Studies done in fish
on the Japanese medaka (Oryzias latipes) have shown that the environmental toxicant Kanechlor
400 significantly decreases the swimming velocity in a dose dependent manner, increases the
nearest neighbor distance (NND), and increases the fractal dimensions of trajectory and turning
angle in the highest PCB-exposure group. It also alters the schooling behavior of these fish
(Nakayama et al., 2005). Similarly there are studies on the carp (Cyrinus carpio) showing an
alteration in its swimming behavior and biotransformation activity (GST) (Schmidt et al., 2004).
Hence, polychlorinated biphenyls are known to alter behavior and change the fixed action
patterns. Similarly, there have been studies on the tree swallows (Tachycineta bicolor) breeding
in an area of the Hudson River with high levels of PCB contamination. When ornamental traits
like plumage color were compared with those of swallows from other parts of the species range,
it was shown that non-contaminated individuals had significantly more adult-type blue-green
coloring than females from rest of the species range. This pattern of plumage anomaly is
consistent with endocrine disruption resulting in the early expression of a trait (Mc Carty and
Secord, 2000), which in turn determines the reproductive success in the future. Hence, PCB
appears to influence a wide spectrum of factors that impact social interaction in the animal
kingdom.

19SUMMARY
Polychlorobiphenyls (PCBs) are environmental toxicants that can be potential health
hazards. These toxicants can be taken into the body either through occupational exposure or
through dietary intake. These aromatic congeners can exert their toxicity by three basic
mechanisms: binding reversibly to the receptors, irreversibly to molecules like protein and DNA
and thirdly, accumulating in the adipose tissue via their lipophilic properties. The toxicity of
PCB depends upon its structure. The toxicological effects can be manifested in many areas of the
body like the adipose tissue, skin liver, muscles, gastrointestinal tract, the immune system, and
the nervous system. These toxicants also affect the proper functioning of the brain and therefore
impairing the higher brain functions like learning and memory. The aim of the current study is to
examine the effect of these toxicants on social recognition using rat as an animal model. In the
present study, the effect of low doses of PCB 47/77 has been investigated with regard to the
concentration of oxytocin in the blood, the area of PVN, and the social behavior of juvenile male
rats.

20MATERIALS AND METHODS
Animals:
Juvenile male Spraque-Dawley rats were used for these experiments. All rats were
housed and treated at Bowling Green State University, Animal Facility. All animals were
obtained from Harlan-Sprague Dawley (Indianapolis, IN, USA). Maintenance and procedures
were in accordance with the Bowling Green State University and NIH guide for the Care and
Use of Laboratory animals, and were approved by the BGSU Institutional Animal Care and Use
Committee, Protocol # 04-015.
PCBs:
Two PCB congeners, 47 (2, 2’, 4, 4’-tetrachlorobiphenyl) and 77 (3, 3’, 4, 4’-
tetrachlorobiphenyl), were obtained from AccuStandard, Inc., New Haven, CT, USA. Stock PCB
was dissolved in absolute ethanol, mixed with 100 g of rat chow (Mowlan Teklad, Madison, WI,
USA), and the ethanol was allowed to evaporate to obtain rat chow with PCB mixed in Equal
amounts of PCB 47 and PCB 77 containing diet were mixed together and formulation of 12.5
ppm and 25 ppm doses was done by adding an appropriate weight of this concentrated mixture to
plain diet to a final weight of 1000 g. The diet was mixed thoroughly. Control animals were fed
mash diet, which did not contain PCB. Females weighing 225-275 g were mated to males of the
same strain. Pregnancy of the females was determined by the presence of the sperm in the
vaginal smear which indicated gestational day 1. These pregnant females were caged separately
and fed either control or PCB containing diet. Exposure to PCB was provided to the rat pups
through maternal diet during gestation and lactation. Litters were standardized to 10 pups (5
males/5 females) on day 3 of age. Male pups were used for the purpose of experiment. The pups

21were separated at 20 days of age, habituated to the social recognition setup for 2 min and isolated
for 18-24 hours before the experiment.
Behavioral assay:
Social recognition test was done using a social recognition arena which is composed of
two rectangular chambers, a social box and a non-social box. The dimensions of each of the
social and non-social boxes were 25 × 20 ×13 cm. These two chambers were connected to each
other through a transparent tube. The social box contained a partition to provide a space for the
restrained animal (Fig. 4). The effect of PCB on social recognition was examined by measuring
the amount of time spent by the rest animal in social investigation as well as the number of
entrances into a social box. Juvenile litter mate male Sprague-Dawley rats (21 days old) were
used for these studies. One rat was restrained in the social cage. The freely moving rat was
placed inside the transparent tube which was closed at both ends with barriers. Then the barriers
were removed and the rat was allowed to move from one cage to another. The time spent in the
vicinity of the restrained rat and the numbers of crosses from one cage to the other cage in a 5
min test trial were recorded. The experiment was processed in three trials. In the second trial the
same restrained rat was exposed to the test rat and in the third trial a novel restrained rat was
introduced. The inter-exposure interval time was 3 minutes. Pairs of rats for the above tests
received the same dietary treatment: CON-CON, PCB 12.5-PCB 12.5, or PCB 25-PCB 25.

22
Figure 5. Test set-up for measurement of social recognition. A social (left) and a non-social box (right) are connected to each other with plastic transparent pipe. The restrained rat was restricted to a small area in the social box.
PVN area measurement:
For the purpose of PVN area measurements, 29-day-old male rats were anesthetized with
a lethal dose of sodium pentobarbital and decapitated. Brains were removed, weighed and placed
in 10% formalin overnight in the refrigerator. After four days, they were placed in 30% sucrose
in 10% formalin for 48 hours. Brains were mounted on a freezing microtome and cut in a coronal
plane with slice thickness of 40 µm. From each brain 8 sections of PVN were taken between
bregma -1.80 and -2.30. Sections were collected and mounted on a slide. Brain sections were
stained with cresyl violet. The slices were examined microscopically using 40X and 100 X
magnifications. Using appropriate software (Motic Image, Version 1.3, Motic China Group Co.,
Ltd), the PVN was accentuated with proper contrast conditions. The boundary of the PVN was
traced in each of the slices and the procedure was repeated for both normal and PCB fed litter
rats.
SSOOCCIIAALL BBOOXX NNOONN--SSOOCCIIAALL BBOOXX

23
.
Figure 6. The area of the PVN measured using the Motic Image software. The left
and the right side bordered by the light blue lines indicate the left and the right side of the PVN.
Enzyme Immunoassay:
Juvenile male rats were anesthetized with sub-lethal doses of sodium pentobarbital at 29
days of age and decapitated. Serum was separated from clotted blood by centrifugation and
commercial ELISA Kit (Assay Designs, Inc. 5777 Hines Drive Ann Arbor, MI 48108 USA) was
used to determine the circulating level of oxytocin hormone. For this purpose an equal volume of
0.1 % trifluoro acetic acid (TFA) in water was added to the serum sample (~ 500 µl). The
mixture was then centrifuged at 17,000 x g for 15 minutes at 4 ºC and the supernatant was saved.
The supernatant was added to a Sep-Pak column and washed with 10-20 mL of 0.1 % TFA in
water. The wash was discarded, and the sample was eluted slowly by applying 3 mL of
acetonitrile: 0.1 % TFA in water 60:40. The eluant was collected in a plastic tube and was
evaporated to dryness using a centrifugal concentrator (Savant Instruments Inc. Model VG-5)
under vacuum. The sample was stored at -20 degrees to be used for enzyme immunoassay.
Oxytocin enzyme immunoassay kit (96 well) was used to estimate the concentration of the

24unknown oxytocin sample. The ELISA was done using the kit (Assay Designs, Inc. 5777 Hines
Drive Ann Arbor, MI 48108 USA) to determine the concentration of oxytocin in control and
PCB samples. The details of the procedure have been included in the appendix.
The results of ELISA experiments were calculated as follows:
a. The average net Optical Density (OD) for the standard and the sample was
calculated by subtracting the average non- specific binding (NSB) OD from
the average OD bound: Average Net OD = Average Bound OD – Average
NSB OD.
b. The binding of each pair of standard wells was calculated as a percentage
of maximum binding wells (Bo), using the formula: Percent Bound = ( Net
OD/Net Bo OD) * 100
c. The Percent bound versus concentration of Oxytocin was plotted using the
Logit-Log paper. A straight line was approximated through the points. The
concentration of the oxytocin was determined by interpolation.
Statistical tools:
Four statistical tools were used to analyze the results of the experiments. In behavioral
analysis repeated measure ANOVA was used to reveal the differences between three groups
(control, PCB 12.5, and PCB 25). If there was any significant interaction between dependent
variables (social time, non-social time, social entry, non-social entry, differences in social-
nonsocial time, and total entry) and groups, then independent sample‘t’ test and student paired‘t’
test were used. Independent sample‘t’ test was used to observe the differences between two
groups and student paired‘t’ test was applied to see the differences within each group across

25trials. For PVN area measurement and enzyme immunoassay (ELISA), one way ANOVA was
used to determine the differences between three groups.

26RESULTS
Social Recognition Test:
Statistical analysis with repeated measure ANOVA revealed that there was no significant
main effect between control, PCB 12.5, and PCB 25 groups in investigation time in social box (p
= 0.27-0.49). However, there was a significant interaction effect between the investigation time
in the social box and the groups (F (4/90) = 2.98, P<0.05). The independent sample‘t’ test was
used to compare the social investigation time between two groups across the trials. These results
showed that there is no significant difference between control group and PCB 12.5 group.
However, there was a significant difference between control and PCB 25 groups for social
investigation time in the second trial (t (31) = -3.01, P<0.01). The mean investigation time was
156 seconds in the second trial for control group and 204 seconds for PCB 25 group. Also,
statistical analysis by student paired‘t’ test showed the difference in investigation times across
the trials within each group. The PCB 25 animals showed the mean investigation time of 172
seconds in social box in the first trial. This value was 204 seconds in second trial and 201
seconds in the third trial. These differences were significant between first and second trials (t
(14) = -2.45, P<0.05) and between first and third trial (t (14) = -2.21, P<0.05). There was no
significant difference across the trials within control group. PCB 12.5 animals showed the same
trend as control (Fig. 7). Also, statistical analysis by repeated measure ANOVA showed a
significant interaction between difference in investigation time in social- and non-social box
(dependent variable), and groups (control, PCB12.5.PCB 25) (F (4/90) =2.62, P<0.05). Statistical
analysis by independent sample‘t’ test shows significance in difference in mean investigation
time in social- and non-social box between control and PCB 25 groups in second trial (t(31) = -

27
* *
120
140
160
180
200
220
240
first trial second trial third trial
Trials
Mea
n so
cial
inve
stig
atio
n tim
e (s
ec.)
CONTROL(n = 18) PCB 12.5 (n= 15) PCB25 (n = 15)
Figure 7. Mean investigation time in 21-day-old Sprague-Dawley rats between control, PCB 12.5, and PCB 25 groups in first, second, and third trials. Each bar represents the Mean±SEM, for control (n=18), PCB 12.5 (n=15), and PCB 25 (n=15).
* *
* *
0
20
40
60
80
100
120
140
160
180
first trial second trial third trial
Trials
Mea
n di
ffere
nce
in s
ocia
l-non
soci
al
inve
stig
atio
n tim
e (s
ec.)
CONTROL (n = 18) PCB 12.5 (n= 15) PCB 25 (n = 15)
Figure 8. Mean difference in investigation time between social-non social box across trials in control, PCB 12.5, and PCB 25 Sprague Dawley rats at the age of 21 days. Each bar represents Mean±SEM for control (n=18), PCB 12.5 (n=15), and PCB 25 (n=15) animals.

282.54, P<0.05). This difference in social- and non-social time in the second trial was 57 seconds
for control group and 133 seconds for PCB 25 groups. There were no significant differences
between control/PCB 12.5 and PCB 12.5/PCB 25 across the trials (Fig. 8).
Also, statistical analysis by student paired‘t’ test showed significant differences between
first and second trial within PCB 25 group (t (14) = -2.34, P<0.05) as well as first and third trial
(t (14) = -2.28, P<0.05). This difference in social and nonsocial time was 61 seconds between
first and second trial, and 62 seconds between first and third trial.
There was no significant difference in the nonsocial time, number of entries in social and
non-social box, and total entries in all groups (Tables 1-4).
GROUPS Trial 1(sec) Trial 2(sec) Trial 3(sec)
CONTROL 89.78±6.84 100.55±8.10 101.11±11.44
PCB (12.5) 74.87±8.87 67.87±15.28 70.67±15.42
PCB (25) 100.40±10.20 71.47±13.89 67.80±14.96
Table.1. Mean investigation time (±SEM) in non-social box for trials with unfamiliar (Trial 1), familiar (Trial 2) and second unfamiliar (Trial 3) conspecifics in control (n = 18), PCB 12.5 (n = 15), and PCB 25 (n = 15)
GROUPS Mean entry1 Mean entry2 Mean entry3
CONTROL 5.72 ±0.36 4.44 ±0.39 4.83 ±0.54
PCB (12.5) 4.73 ±0.56 4.67 ±0.41 4.87 ±0.60
PCB (25) 5.53 ±0.51 4.60 ±0.36 5.20 ±0.78
Table.2. Mean and the mean±SEM for the number of entries into the social box for all three trials within control (n = 18), PCB 12.5 (n = 15), and PCB 25 (n = 15).

29
GROUPS Mean 1 Mean 2 Mean 3
CONTROL 4.89 ±0.33 4.67 ±0.38 4.67±0.50
PCB (12.5) 4.53±0.45 4.47±0.31 5.1±0.57
PCB (25) 5.1±0.39 3.9 ±0.49 4.60±0.67
Table.3. Mean and the mean±SEM for the number of entries into the non- social box for all three trials. Control (n = 18), PCB 12.5 (n = 15), and PCB 25 (n = 15).
GROUPS Mean 1 Mean 2 Mean 3
CONTROL 10.61±0.66 9.1±0.74 9.50±1.01
PCB (12.5) 9.27±0.94 9.13±0.62 10.0±1.07
PCB (25) 10.67±0.85 8.47±0.77 9.80±1.40
Table.4. Mean and the mean±SEM for the number of entries into the social + non- social box for all three trials. Control (n = 18), PCB 12.5 (n = 15), and PCB 25 (n = 15).
PVN area measurement:
Statistical analysis by one way ANOVA showed that there was no significant difference
observed for the PVN areas across the groups. However, there was a trend toward a depression
of PVN area in PCB animals of 0.44 square µm in PCB 12.5 and 2.76 square µm in PCB 25 as
compared to controls (Fig. 9).

30
282
284
286
288
290
292
294
296
1CONTROL PCB(12.5) PCB(25)
NO
RM
ALI
ZED
PVN
ARE
A(sq
mic
rom
eter
)
CONTROL PCB 12.5 PCB 25
Figure 9. The normalized PVN area for the control (n=10), PCB 12.5 (n=10), and PCB 25 (n=10) fed rats. The graph shows that there was not any significant difference between the normalized area of the PVN of control rats, PCB 12.5 and PCB 25.
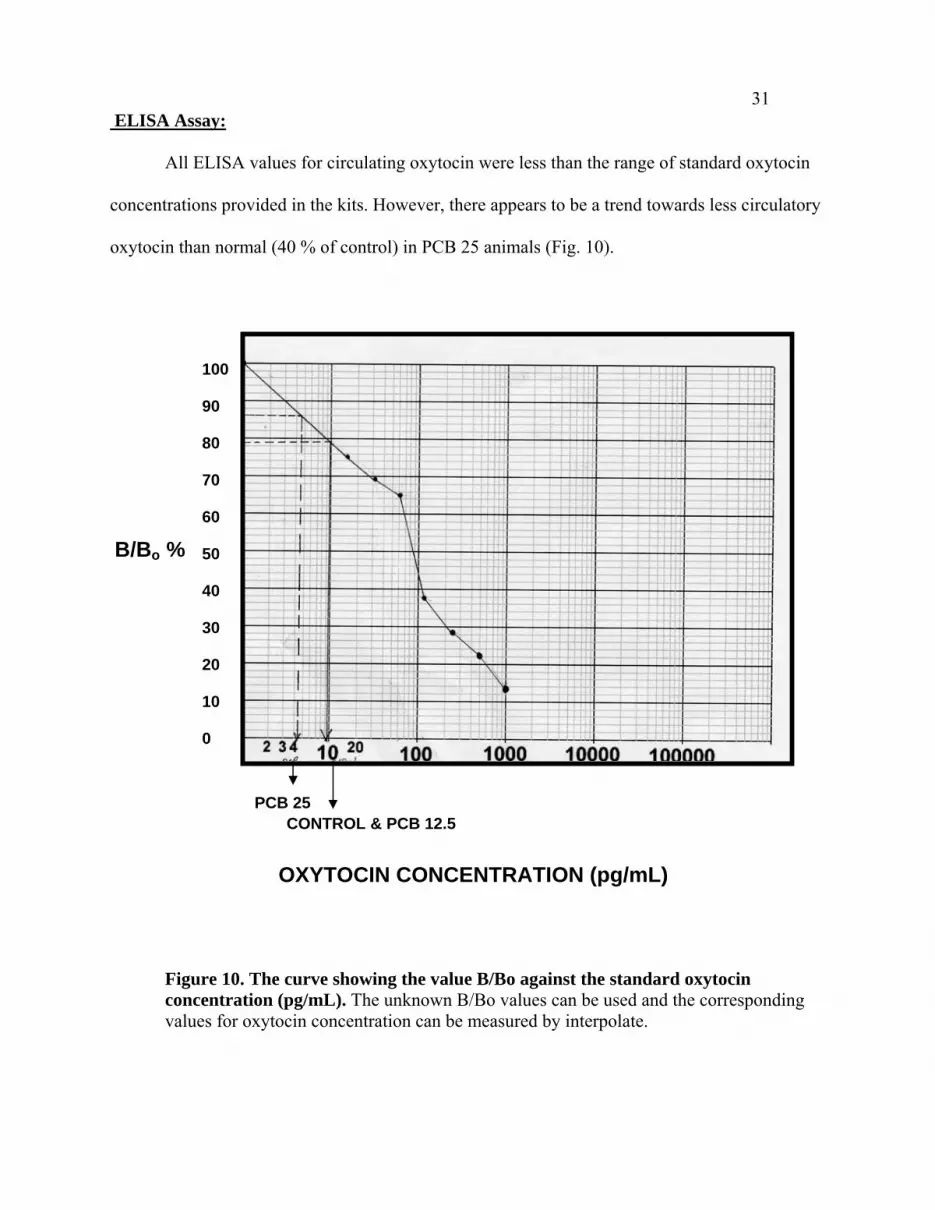
31 ELISA Assay:
All ELISA values for circulating oxytocin were less than the range of standard oxytocin
concentrations provided in the kits. However, there appears to be a trend towards less circulatory
oxytocin than normal (40 % of control) in PCB 25 animals (Fig. 10).
100 90 80 70 60 50 40 30 20 10 0
B/Bo %
PCB 25 CONTROL & PCB 12.5
OXYTOCIN CONCENTRATION (pg/mL)
Figure 10. The curve showing the value B/Bo against the standard oxytocin concentration (pg/mL). The unknown B/Bo values can be used and the corresponding values for oxytocin concentration can be measured by interpolate.

32RESULTS SUMMARY
The results of these studies show that the small amount of PCB 47/77 at concentration of
12.5 ppm does not have any significant effect in altering social recognition in juvenile rats.
However, PCB 25 does show a significant difference in the second trial as compare to controls.
The value of the serum oxytocin concentration for all the three categories was negligible.
Moreover, the area of the PVN does not undergo any significant changes in PCB fed rats but
there is a trend toward a smaller PVN area as compared to that in the control groups. This could
suggest that the concentration of PCB was not sufficient to bring about a robust effect. It might
be possible that the changes in these parameters might be more pronounced if the same study is
performed in adult rats, with an increased concentration of the PCB 74/77.

33DISCUSSION
Social recognition Test:
Previous work investigating social recognition between rat conspecifics has been done
either with adult animals or with juvenile rats that were at least 25–35 days of age (Thor and
Holloway, 1982). In these studies, the second exposure of test rat to a conspecifc resulted in a
robust decrease in the amount of social interaction, whereas exposure to another novel rat
restored the previous degree of interaction. The present study was done with younger (21-day-
old) male rats, to determine whether social recognition occurs at this age, and whether exposure
to PCB 47/77 alters these interactions. While the mean time spent in the social box by a control
test rat decreased on a second exposure to a conspecific, and returned to the original level with
exposure to a novel rat (Figs. 7, 8), the alteration was insufficiently robust to reach statistical
significance. This result suggests that social recognition and affiliation are not robust at the time
of weaning, but develop rapidly as rats mature. PCB 12.5 did not alter social recognition in 21-
day-old rats (Figs. 7, 8). However, PCB 25 rats displayed significantly more social interaction on
second exposure to the same rat (i.e., the familiar rat) and to the second novel rat than did
controls. While the test species is not directly analogous to the present 21 day old rats, prairie
voles have been examined for social recognition after exposure to low doses of the toxicants
DES and MXC (0.2 µg/kg DES or 2 mg/kg MXC) (Engell et al., 2004). While voles exposed to
either toxicant spent the same amount of time as controls with their mates, those exposed to
MXC were less inclined to spend time with a novel vole. While not directly comparable to the
present study, evidence is provided that social recognition can be disrupted in the voles by the
toxicants MXC and in young male rats by PCB 47/77 at the 25 ppm dietary concentration.

34A more directly related study on the influence of low doses of PCB 47/77 in short- term
and long- term memory of Sprague-Dawley rats shows the same trend (Donahue et al., 2004).
When a mixture of PCB 47//77 was administrated at the dietary concentration of 1.25 ppm, 12.5
ppm, or 25 ppm, measurement of short-term or long-term memory in the Morris water maze
revealed a trend towards disruption of both short-term and long-term memory. While the
measurement of memory was made in a setting different from that in the present study, both
support the consequences of PCB 47/77 exposure on development of behaviors relying on
memory.
The dose dependent response of social behavior to PCB is similar to the change in motor
behavior, learning and memory processes and the cholinergic transmitter system in adult mice
observed under the influence of 2,2’,4,4’,5,5’-hexa BDE (polybromodiphenyl ether) (0.9 and 9.0
mg/Kg of body weight) (Viberg et al., 2003). The effect on significant decrease of motor activity
is more pronounced at a greater dose of 9 mg/Kg as compared to the lesser dose. This could
account for the detrimental effect of PCB 25 as compared to PCB 12.5 ppm.
A recent study (Kenet et al., 2007) showed that early exposure to PCB 99 impairs
auditory capabilities of the developing rat. While the auditory cortex of control rats developed
normally, that of rat pups exposed to PCB in utero and through lactation was smaller, and
morphologically and functionally altered. Thus, the current studies on social behavior were done
on juvenile rats to investigate whether the PCB congeners 47/77 can cause significant changes in
behavior during this critical phase of prenatal brain development. The result that differences in
social behavior are not robust could have occurred because lesser amount of PCBs were
administrated in the present study and/or that rats were tested at 21 days of age.

35In the present study, alteration of social recognition of the familiar conspecifc by rats
exposed to 25 ppm PCB 47/77 may result from an effect of the toxicant on expression of
oxytocin receptor in the medial amygdala. In future investigation, such an effect could be
confirmed by receptor antibody binding studies. This effect of PCB 47/77 congeners could be
through aryl hydrocarbon dependent mechanism. Previous studies have shown that aryl
hydrocarbon dependent PCB activates cytocrome P 450, 1A1 AND 1B1, which leads to
disruption of estrogen. (Carpenter, 2006). Therefore, in the current study, it might be possible
that PCB 47/77 reduces the estrogen level, which is involved in oxytocin production through
estrogen receptor beta in PVN and oxytocin receptor expression through estrogen receptor alpha
in medial amygdale (Choleris et al., 2004). Another possibility could be through the effect of
PCB on estrogen receptor expression (Bonefeld-Jorgensen et al., 2001). It might be possible that
PCB 47/77 congener result in under expression of estrogen receptor alpha and beta in the brain. .
Therefore, there will not be sufficient hormones–receptor interactions to mediate social
recognition. Studies have shown that the dopamine activation of the D1 receptors increases the
activity of protein kinase A, which results in increased or tonically maintained levels of oxytocin
receptor binding in central amygdala, which in turn brings about social behavior (Bale et al.,
2001). Hence, the second possible mechanism for disruption of the social behavior by PCB could
be through decrease in the dopamine level, which in turn deactivates protein kinase A, resulting
in decreased levels of oxytocin receptor binding in the central amygdala, thereby impairing
social recognition.
In the present study, absence of significant difference in the cumulative entry times for
social- and non-social box suggests that PCB does not induce hyperactivity in these rats.
Previous studies have shown the behavioral hyperactivity in rats caused by sub-toxic doses of

36PCB 153 and 126 (Holene et al., 1998). Therefore, it could be that the amount of PCB 47/77 fed
in the current study was below threshold doses, and thus had no effect on increasing the
hyperactivity in these rats.
PVN Area Measurement:
The weights of the brains in the present study were not altered by PCB exposure and
averaged for all animals 1.5 g. The area of paraventricular nucleus did not undergo a significant
change under the influence of PCB 12.5 and 25 ppm. This could suggest that the PCB does not
exert an effect on the structure of the PVN, the neurons might not undergo the process of
apoptosis under the effect of the endocrine disruptor but their firing rate might change,
suggesting a functional rather than a structural deterioration. The Sprague-Dawley rats used for
these experiments are juvenile one. It could be possible that the oxytocin producing neurons are
not as abundant in the juvenile rats as those present in adult rats and therefore, there is no
significant change in the area of the PVN even if some or all of the neurons are lost under the
effect of PCB. Another accurate way to measure the effect of PCB 47/77 on the oxytocin
neurons in the PVN is to perform cell counts. If there is a significant loss of the oxytocin neurons
from these areas, then the number of counts will be different for the controls and the PCB fed
rats.
Studies done in human subjects have shown that the disorder in the PVN can lead to
eating disorders and obesity (Swaab et al., 1995). This disease is known as Prader-Willi
Syndrome. The symptoms of this disease are diminished fetal motor activity; sever infant
hypotonia, mental retardation, hypogonadism, and hypogenitalism (Swaab et al., 1995). Also
since the oxytocin neurons exert an inhibitory effect on eating and dietary intake, the atrophy to
the PVN leads to an increased appetite and obesity. Since no such changes were observed in the

37juvenile rats after having been exposed to PCB 47/77 pre- and post-natally, it can be concluded
that this is the reason why the low doses of these endocrine disruptors do not cause a significant
change in the area of the PVN. If these doses were causing a significant change in the area of the
PVN, then there would likely to be a significant effect on the weight of the rats fed on a PCB
diet. A qualitative observation of these rats suggests no change in their body weights.
ELISA Test:
The ELISA test was done on the serum samples obtained from control, PCB 12.5, and
PCB 25 to monitor the circulating oxytocin concentration in each of these serum categories.
Concentrations for control and PCB 12.5 subjects were nearly identical, though the values were
very low. The explanation for the same concentration of the oxytocin in PCB 12.5 and controls
could be that this dose of PCB was insufficient to produce any measurable changes in synthesis
and/or secretion of oxytocin. Also, it is possible that juvenile male rats at the age of 21 days do
not have circulating oxytocin levels measurable with the ELISA Kit employed. While the value
for the oxytocin concentration obtained for PCB 25 animals was approximately 40 % of control,
all values were below the lowest standard oxytocin concentration supplied with the kit, so no
valid conclusions could be drawn.
The concentrations of oxytocin found in the controls, PCB12.5 and PCB 25 suggest that
it is not produced in greater amounts. However, PCB 25 does manifest a decrease in the social
behavior and this could be due to its toxic effect on the central oxytocin level. This is because
social behavior is interplay of the oxytocinergic neurons in the PVN and its projections to the
amygdala, ventromedial nucleus of the hypothalamus, and the number of oxytocin receptors
expressed on them (Bale et al., 2001). It could be possible that juvenile rats have less firing and
number of oxytocin receptors and therefore, shows a lower OT concentration. The number of

38oxytocin receptors expressed in the cAmyg and the VMH might increase as the animal becomes
mature and experiences more exposure to its conspecifc. The present juvenile rats have been
used for social behavior right after weaning and therefore, have not been exposed to many
conspecifcs. As a result the social behavior is not strongly manifested by them. It could be
possible that as the rats get older, they are more exposed to conspecifics and might show stronger
social behavior. This strong social behavior will be correlated with an increase in the
concentration of oxytocin in the serum.
Involvement of other sensory cues (olfaction and auditory):
Social recognition in rodents, unlike mammals is primarily based on olfactory cues
(Keverne and Brennan, 1996). To recall, during a brief social encounter, the male rat investigates
a novel rat by sniffing for a brief period of time. It is assumed that during this process of
investigation, the male rat collects and stores the information about the identity of the
conspecific. If the rat encounters the same conspecific during second encounter, then it
investigates it for only a few seconds and quickly engages in different behaviors. The first step is
the process of acquisition and the second step is the retrieval of the social memory. In the present
study, PCB 25 alters the social recognition as compared to the controls and this is manifested as
an increase in the investigation time in second trial. The difference is also manifested between
first and second, and first and third trials. In addition to correlating the effect of PCB on the
PVN, oxytocin concentration, it could be possible that PCB 25 alters the olfactory sensory
epithelial tissue and therefore, the rat is not receiving olfactory cues from the conspecific. As a
result, afferent fibers do not carry information to amygdala, which leads to an impairment in the
social recognition response. Studies have shown that chlorinated organic molecules exert site-
specific toxicity in olfactory mucosa of rats by causing necrosis of cells (Franzen et al., 2003).

39However, recent studies by Cromwell et al. (2007) have shown the ability of young PCB 12.5
and 25 rats to discriminate between a familiar and an unfamiliar odor.
Another possible way in which PCB can disrupt social behavior in rats is by affecting the
auditory capabilities of the rat. Studies have shown that maternal exposure to a commercial PCB
mixture, Arochlor 1254 (A 1254) results in low-frequency hearing loss in adult offspring
(Goldey et al., 1995). This study hypothesized that the hearing loss resulted from PCB-induced
reduction in circulating thyroid hormone, which is necessary for the normal development of the
cochlea (Uzei, 1986). The idea was supported when postnatal replacement of thyroxin
ameliorated the auditory deficit. Hence, cochlear damage has been shown to be an effect of PCB.
Though the primary sensory modality responsible for social recognition is olfaction, there are
also other senses that play a role in social recognition like auditory cues. Another study which
supports this hypothesis showed that the toxicant disrupted the neural circuitry, which decreased
the capacity of rat to change or learn in response to sound (Kenet et al., 2007). Hence, the
reduction in social investigation time in PCB 25 rat could be caused by hearing impairment. As a
result, rats were not able to recognize their conspecific.
The effect of PCB on motor activity of rats was not evident in the present study because
the cumulative number of entries was same for all the three groups of rats. If there was a deficit
in motor skills, then the PCB 12.5 and 25 rats would have shown fewer entries than controls
(Table 2-4).
Role of neurotransmitters:
Experiments in the present thesis only examine the effect of PCB on the neuropeptide
oxytocin that is directly involved in social recognition. Other than oxytocin, there are other
biochemical agents that might contribute in bringing about social recognition. Few studies have

40suggested the involvement of dopaminergic and noradrenergic mechanism (Guan et al., 1994). It
might be possible that the doses of PCB 12.5 and PCB 25 might not be effecting the
concentration of the oxytocin in the serum significantly, but might disrupt the reuptake of
dopamine and noradrenalin in the brain, which might impair social recognition. Other than this,
as discussed earlier PCB can also act on the dopamine receptors and deactivate protein kinase A.
Involvement of other biochemical agents:
Other than the involvement of oxytocin in social recognition, vasopressin has also been
shown to be involved in a variety of complex social behaviors like territorial defense and
interspecies communication (Dantzer and Bluthe, 1992). In monogamous prairie voles,
vasopressin has been shown to be involved in the development of pair bonds. In the male prairie
voles, low doses of vasopressin can facilitate partner preference formation and mate guarding
without any reduction in the mating behavior (Winslow et al., 1993). Hence, other than the role
of oxytocin in social recognition, vasopressin could also be involved in affiliative behaviors.
Studies done by Prediger and Takahashi, (2003) have shown the involvement of opiod receptors
in mediating the formation of short term social memory in rats when ethanol was administered to
rats intraperitoneally (i.p.) and orally. In our studies, there could be a possibility that these opiod
receptors might be under expressed in PCB 25 animals which might lead to a decrease in social
recognition in juvenile male rats.
Clinical Implications:
This study on social recognition in rats provides a good model for conducting various
kinds of experiments that are not possible on human subjects. The study helps in understanding
the various neural processes that take place in the brain during the social behavior. The study
does not necessarily model any specific human conditions, but it provides a better understanding

41about the social brain from both behavioral and biochemical approach. If there is a clear
understanding as to how the brain processes social information and regulates social behavior,
then it could help to understand the mechanism behind various kinds of neuropsychiatric
disorders in humans like schizophrenia. Various studies have shown that the brain processes
social information using specific neural circuits that are not involved in non-social recognition
(Storm and Tecott, 2005). fMRI studies in humans have shown that the brain processes social
visual stimuli in a different way as compared to nonsocial stimuli. In the study, the researchers
have shown that the left lateral fusiform gyrus is activated to a greater degree when the subjects
view faces than when they view nonface objects (Grill-Spector et al., 2004). Unfortunately, it is
not possible to understand the mechanism in humans at the molecular and biochemical level. In
such situations, these experiments on rats are very useful. Knockout mice can be used with the
oxytocin gene knocked out to monitor the changes in social recognition behavior. If oxytocin is
specifically involved in social memory, then these knock out mice will not habituate to, or
recognize a familiar rats in the second trial, or after repeated exposures. To rule out deficits in
olfactory processing, these rats when exposed to nonsocial scents like those that a cotton ball
scented with lemon extract will show habituation. Hence, these experiments can deductively tell
us the biochemical agents that are involved in social recognition.
Impairment of social recognition in humans can have severe clinical implications. These
clinical implications can be both psychological and non-psychological. The impairment in social
attributes occurs frequently in humans after lesions to the orbitofrontal cortex (Linda et al.,
2004). This impairment is manifested as disinhibition and lack of restraint, impaired sight and
self-monitoring social withdrawal, and inability to respond to interpersonal cues. This evidence
has come after analyzing the fMRI scans of impaired subjects when they were made to perform

42on an Interpersonal Perception Task, and comparing the brain activities with the healthy subjects.
The impaired subjects showed a decrease in neural firing activity in the prefrontal cortex
especially in the dorsolateral prefrontal areas. The decrease in the performance in interpersonal
perception task correlated with the degree of damage to the dorsolateral prefrontal cortex.
Though there might be impairment in social behavior, there was no effect on the intellectual
functions, such as language, memory, perception and performance in the standard
neuropsychological tests (Linda et al., 2004). There is story of a person called Phineas Gage
which throws light on how extensive damage to the prefrontal cortex can impair all the social
attributes of the human brain. Gage was a railroad foreman, and was unearthing the rock when a
thick rod went into his skull. Surprisingly, he did not die but the clinician who was attending him
observed a drastic change in the emotional and rational faculties of the brain. The damage was so
severe that Phineas lost his job eventually. He turned from a person with a very sound
personality to somebody who was rude, impolite and negligent. The consequence of that was he
lost his job. These kinds of damage are also likely to happen through the effect of PCB on the
brain.
The alteration in PVN size has been reported in a disease called Prader-Willi syndrome.
In this disease there is a decrease in the size of the PVN (Swaab et al., 1995). The oxytocin
neurons of the hypothalamic PVN are very important in regulation of food intake. Studies have
shown that lesion to the PVN produces overeating and obesity (Leibowitz et al., 1981). It also
says that the stimulation of the medial parvocellular subdivision of the PVN elicits significant
increases in the gastric acid secretion. Hence, the oxytocin neurons in PVN exert an inhibitory
effect on eating and the body weight.

43FUTURE DIRECTIONS
1. The oxytocin concentration was measured from peripheral circulation and not from
central nervous system. If the brain tissue from the PVN and the central and medial
amygdale was minced and was used to perform ELISA, that would have been a more
accurate test estimating the concentration of CNS oxytocin and the change in the
concentration under the effect of PCB.
2. One of the mechanisms proposed by Choleris et al., (2004) to explain social recognition
is that oxytocin binds to the oxytocin receptors in amygdala to bring about social
recognition. Hence, not only can PCB exert its effect on social behavior by decreasing
the level of oxytocin, it can also show its effect by altering the expression of oxytocin
receptors (OTRS). Oxytocin levels might be normal in the brain, but if there is under
expression of OTRs from the effect of PCB, then the oxytocin cannot play its
contributory effect in bringing about social recognition. Hence, an experiment whereby
oxytocin receptor expression can be monitored should be a good way of understanding
the effect of PCB on social recognition.
3. Results of the experiment show that the area of the PVN might be unaltered under the
effect of PCB, but that does not imply that the parvocellular and magnocellular cells
which produce oxytocin do not undergo any damage under the effect of PCB. To
understand the effect of the PCB on the levels of oxytocin, one of the accurate ways
would be to perform cell counts to determine the number of healthy and apoptotic cells in
the PVN under the effect of PCB 12.5 and 25 and compare it with the cell counts from
the control PVN.

444. The parameter used to measure social recognition is the amount of time that juvenile rat
spends in the social box. The rat might be in the social box but that does not mean that
the rat is actively investigating its conspecific. Hence, nosing and sniffing are the more
accurate parameters to measure social recognition because in that case there is an
accurate measure for active social investigation by rat.
5. Thor and Holloway (1982) have shown that juvenile male rats between 25-35 days of age
are able to acquire and retrieve social memory better than ones which are 21 days of age.
Hence, using rats of this age will provide a more accurate measure for social recognition.

45REFERENCES
Bale, T.L., Davis, A.M., Auger, A.P., Dorsa, D.M., and McCarthy, M.M. 2001. CNS-region specific oxytocin receptor expression: Importance in the regulation of anxiety and sex behavior. The Journal of Neuroscience 21(7): 2546-2552. Beebe, L.E., Kim, Y.E., Amin, S., Riggs, C.W., Kovatch, R.M., and Anderson, L.M. 1993. Comparison of transplacental and neonatal initiation of mouse lung and liver tumors by N- nitrosodimethylamin (NDMA) and 4- (methylnitrosamino)-1-(3-pyridyl)-1-butanone (NNK) and promotability by a polychlorinated biphenyls mixture (Arocolor 1254). Carcinogenesis, 14:1545-1548. Ben-Barak, Y., Russell, J., Whitnall, M., Ozato, K., and Gainer, H. 1985. Neurophysin in the hypothalamo-neurohypophyseal system. I. Production and characterization of monoclonal antibodies. J Neurosci 5:81-97. Bonefels-Jorgensen, E.C., Andersen, H.R., Rasmussen, T.H., and Vinggaard, A.M. 2001. Effect of highly bioaccumulated polychlorinated biphenyl congeners on estrogen and androgen receptor activity. Toxicology 158:141-153. Brouwer, A., Morse, D., Lans, M., Schuur, A., Murk, A., Klasson-Wehler, E., Bergman, A., and Visser, T. 1998. Interactions of persistent environmental organohalogens with the thyroid hormone system: mechanism and possible consequences for animal and human health. Toxicol Ind Health 14(1/2):59-84. Carpenter, D.O. 2006. Polychlorinated Biphenyls (PCBs): routes of exposure and effects on humans and health. Review on environmental health 21(1) pp 1-23. Choleris,E., Kavaliers, M., and Pfaff, D.W. 2004. Functional genomics of social recognition. Journal of Neuroendocrinology (16) 383-389. Cromwell, H.C., Johnson, A., McKnight, L., Horinek, M., Asbrock, C., Burt, S., Jolous-Jamshidi, B., and Meserve, L.A. 2007. Effect of polychlorinated biphenyls on maternal odor conditioning in rat pups. Physiol Behav. April 1 ;( Epub ahead of print). Dantzer, R., Bluthe, R.M., Koob, G.F., Le Moal, M. 1987. Modulation of social memory in male rats by neurohypophyseal peptides. Psychopharmacology 91:363-368. Dantzer, R. and Bluthe, R.M. 1992. Vasopressin involvement in antipyresis, social communication, and social recognition: A synthesis. Critic Rev Neurobiol 6:243-255. De Felip, E., Domenico, A., Miniero, R., Silvestroni, L. 2004. Polychlorobiphenyls and other organochlorine compounds in human follicular fluid. Chemosphere 54: 1445-1449.

46Dluzen, D.E., and Kreutzberg, J.D. 1993. 1-Methyl-4-phenyl-1, 2, 3, 6, tetrahydropyridine (MPTP) disrupts social memory/recognition process in the male mouse. Brain Res 609: 98-102. Donahue, D.A., Dougherty, E.J., and Meserve, L.A. 2004. Influence of a combination of two tetrachlorobiphenyl congeners (PCB 47; PCB77) on thyroid status, choline acetyltransferase (ChAT) activity, and short- and long –term memory in 30-day-old Sprague-Dawley rats. Toxicology 203:99-107. Elands, J., Resink, A., and De Kloet, E.R. 1990. Neurohypophyseal hormone receptors in the rat thymus, spleen, and lymphocytes. Endocrinology (Baltimore) 126(5): 2703-2710.
Engell, M.D., Godwin, J., Young, L.J., and Vanderbergh, J.G.2005. Perinatal exposure to endocrine disrupting compounds alters behavior and brain in the female pine vole. Neurotoxicol Teratol. 28(1):103-110. Ferguson, J., Aldag, J.H., Insel, T.,and Young, L. 2001 b. Oxytocin in the medial amygdala is essential for social recognition in the mouse. J Neurosci. 21:8278-8285. Ferguson, J.N., Mattew Aldag, J., Insel, T.R., and Young, L.J. 2000. Social amnesia in mice lacking the oxytocin gene. Nature Genetics 25(3): 294-288. Ferguson, J.N., Mattew Aldag, J., Insel, T.R., and Young, L.J. 2001. Oxytocin in the medial amygdala is essential for social recognition in the mouse. The Journal of Neuroscience 21(20): 8278-8285. Ferguson, J., Young, L., and Insel, T.R. 2002. The neuroendocrine basis of social recognition. Front. Neuroendocrinol. 23:200-224.
Franzen, A., Carlsson, C., Brandi, I., and Brittebo, E.B. 2003. Isomer-Specific Bioactivation and Toxicity of dichlorophenyl methylsulphone in rat olfactory mucosa. Toxicology Pathology (31) 364-372.
Freund Mercier, M.J., Dietl, M.M., Stoekel, M.E., Palacios, J.M., and Richard, Ph. 1988. Quantitative autoradiographic mapping of neurohypophyseal hormone binding sites in the rat forebrain and pituitary gland – II. Comparative study on the Long Evans and Brattleboro strains. Neuroscience, July; 26(1):273-281. Goldey, E.S., Kehn, L.S., Lau, C., Rehnberg, G.L., and Crofton, K.M. 1995. Developmental exposure to polychlorinated biphenyls (Arochlor 1254) reduce circulating thyroid hormone concentrations and causes hearing deficits in rats. Toxicol. Appl. Pharmacol. 135, 77-88. Grill Spector, K., Knouf, N., and Kanwisher, N. 2004. The fusiform face area subserves face perception, not genetic within-category identification. Nature Neuroscience 7(5): 555-562.

47Guan, X., and Dluzen, D.E. 1994. L-Dopa reverses castration- induced disruption of dishabituation responses to female chemical cues in male rats. Pharmacol Biochem Behav 48:515-519. Guan, X., Blank, J.L., and Dluzen, D.E. 1994. Role of olfactory bulb norepinephrine in the identification and recognition of chemical cues. Physiol Behav 53:947-441. Hellbach, S., Gartner, P., Deicke, J., Fischer, D., Hassan, A.H.S., and Almeida, O.F.X. 1998. Inherent glucocorticoid response potential of isolated hypothalamic neuroendocrine neurons. The FASEB Journal (12) 199-207. Hemming, H., Flodstrom, S., Fransson Steen, R., Warngard, L., and Ahlborg, U. 1992. Inhibitor of intercellular communication in cells in culture by polychlorinated compounds. Chemosphere 25:939-943. Holene, E., Nafstad, I., Skaare, J.U., and Sagvolden, T. 1998. Behavioral hyperactivity in rats following postnatal exposure to sub-toxic doses of polychlorinated biphenyl congeners 153 and 126. Behavioral Brain Research 94(1): 213-24. Howsam, M., Grimalt, J.O., Guino, E., Navarro, M., Marti- Rague, J., and Peinado, M.A. 2004. Organochlorine exposure and colorectal cancer risk. Environ Health Perspect. 112:1460-1466. <http:// de.geocities.com/image/pcb-molecule.gif> [accessed May 25, 2007]
< http://www.atsdr.cdc.gov/tfacts17.html> [accessed May 7, 2006].
<http://en.wikipedia.org/wiki/Paraventricular_nucleus/Magnocellular_neurosecretory_neurons_in_the_ PVN > [accessed May 12, 2006]. Isadora, F.B., and Young, L.J. 2004. Oxytocin, vasopressin, and social recognition in mammals. Peptides 25: 1565-1574. Ishida, T., Kamiichi, K., Kuwahara, A., Doi, M., and Inoue, M. 1986. Stacking and hydrogen bonding interactions between phenylalanine and guanine nucleotide: crystal structure of L-phenylalanine-7- methylguanosine -5’- monophosphate complex. Biochem Biophys Res Cummun 136:294-299. Jacobson, J., and Jacobson, S. 1997. Teratogen Update: Polychlorinated Biphenyls. Teratology 55:338-347. Jacobson, J.L., and Jacobson, S.W. 1996. Intellectual impairment in children exposed to polychlorinated biphenyls in utero. N Eng J Med 335: 783-789. Jarrell, J.F., Villeneuve, D., Franklin, C., Bartlett, S., Wrixon, W., Kohut, J., and Zouves, C. 1993. Contamination of human ovarian follicular fluid and serum by chlorinated organic compound in three Canadian cities. Canadian Medical Association Journal 148:1321-1327.

48 Kato, S., McKinney, J.D., and Matthews, H.B. 1980. Metabolism of hexachlorobiphenyls in the rat. Toxicol Appl Pharmacol 53:389-398. Kenney, M.J., Weiss, M.L., and Haywood, J.R. 2003. The paraventricular nucleus: an important component of the central neurocircuitry regulating sympathetic nerve outflow. Acta Physiol Scan 177:7-15. Kenet, T., Froemke`, R.C., Schreiner, C.E., Pessah, I.N., and Merzenich, M.M. 2007. Perinatal exposure to a noncoplanar polychlorinated biphenyl alters tonotopic, receptive field, and plasticity in rat primary auditory cortex. Proc Natl Acad Sci U S A 104(18): 7646-7651. Keverne, E.B., and Brennan, P.A. 1996. Olfactory recognition memory. J.Physiol. Paris 90:399-401. Kimbrough, R.D., Squire, R.A., Linder, R.E., Strandberg, J.D., Montali, R.J., and Burse, V.W. 1975. Induction of liver tumors in Sherman strain female rats by polychlorinated biphenyl Arocolor 1260. J Natl Cancer Inst 55: 1453-1459. Kuroda, Y., Kimura-Kuroda, J., and Nagata, I. 2004. Developmental disorders of the brain can be caused by PCBs; low doses of hydroxyl-PCBs disrupt thyroid hormone- dependent dendrite formation from Purkinje neurons in culture. Neurotoxicity and Other Effects (66) 3105-11. Leibowitz, S.F., Hammer, N.J., and Chang, K. 1981. Hypothalamic paraventricular nucleus lesions produce overeating and obesity in the rat. Physiol Behav. 27:1031-1040. Lim, M.M., and Young, L.J. 2006. Neuropeptidergic regulation of affiliative behavior and social bonding in animals. Hormones and Behavior 50:506-517. Linda, M., Arnold, M.C., and Grafman, J. 2004. Impairment of social perception associated with lesion of the prefrontal cortex. Am J Psychiatry (161) 1247-1255. Lind, P.M., Eriksen, E.F., Sahlin, L., Edlund, M., and Orberg, J. 1999. Effects of the antiestrogenic environmental pollutant 3, 3’, 4, 4’, 5-pentochlorobiphenyl (PCB#126) in rat bone and uterus: diverging effects in ovariectomized and intact animals. Toxicology and Applied Pharmacology 154:236- 244. Longnecker, M.P., Korrivk, S.A., and Moysich, K.B. 2003. Human health effects of polychlorinated biphenyls. In: Schecter A, Gasiewicz TA, Eds, Dioxins and Health, 2nd Edition. Hoboken, New Jersey, USA: Wiley Interscience. 670- 728. Mayes, B.A., McConnell, E.E., Neal, B.H., Brunner, M.J., Hamilton, S.B., and Sullivan, T.M.,1998. Comparative carcinogenicity in Sprague-Dawley rats of the polychlorinated biphenyl mixtures Aroclors 1016, 1242, 1254 and 1254 and 1260. Toxicol Sci 41: 62-76.

49McCarty, J.P., and Secord, A.L. 2000. Possible effects of PCB contamination on female plumage color and reproductive success in Hudson River tree swallows. BioOne 117(4): 987-995 McFarland, V.A., and Clarke, J.U. 1989. Environmental occurrence, abundance, and potential toxicity of poly chlorinated biphenyl congeners: consideration for congener specific analysis. Environ Health Perspect 81: 225- 239. McKinney, J.D., and Singh, P. 1981. Structure- activity relationships in halogenated biphenyls: unifying hypothesis for structural specificity. Chem- Biol Interact 33:271- 283. McKinney, J., and Waller, C. 1994. Polychlorinated biphenyls as hormonally active structural analogues. Enviorn Health Perspect 102: 290-297. Moberg, K. 1998. Oxytocin may mediate the benefits of positive social interaction and emotions Psychoneuroendocrinology 23(8):819-35. < meds.queensu.ca/ images/pvn.ht3> [accessed May 7, 2006).
Moysich, KB., Shields, P.G., Freudenheim, J.L., Schisterman, E.F., Vena, J.E., Kostyniakn, P. 1999. Polychlorinated biphenyls, cytochrome P4501A1 polymorphism, and postmenopausal breast cancer risk. Cancer Epidemiol Biomark Prev 8: 41-44. Nakayama, K., Oshima, Y., Hiramatsu, K., Shimasaki, Y., and Honjo, T. 2005. Effect of polychlorinated biphenyls on the schooling behavior of Japanese medaka (Oryzias latipes). Environmental Toxicology and Chemistry 24(10): 2588-2593. Nishtani, S., Moriya, T., Kondo, Y., Sakuma, Y., Shinohara, K. 2004. Induction of Fos imunoreactivity in oxytocin neurons in the paraventricular nucleus after female odor exposure in male rats: effects of sexual experience. Cell Mol Neurobiol. 24(2):283-91. Oakley, G.G., Devanaboyina, U., Robertson, L.W., Gupta, R.C. 1996. Oxidative DNA damage induced by activation of polychlorinated biphenyls (PCBS): implications for PCB-induced oxidative stress in breast cancer. Chem Res Toxicol 9: 1285-1292. Patisaul, H.B., Scordalakes, E.M., Young, L.J., and Rissman, E.F. 2003. Oxytocin, but not oxytocin receptor, is regulated by the estrogen receptor beta in the female mouse hypothalamus. Journal of Endocrinology 15:1-7. Perio, A., Terranowa, J.P., Worms, P., Bluthe, R.M., Dantzer, R., and Biziere, K. 1989. Specific modulation of social memory in rats by cholinomimetic and neotropic drugs, by benzodiazepine inverse agonists, but not by psycho stimulants. Psychopharmacology 97:262-268. Perrett, D.J., Smith, P.A., Potter, D.D., Mistlin, A.J., Head, A.S., Milner, A.D., and Jeeves, M.A. 1984. Neurons responsive to faces in the temporal cortex: studies of functional organization, sensitivity to identity and relation to perception. Hum Neurobiology 3: 197-208.

50Popik, P., Vetulani, J., and van Ree, J. 1991. Low doses of oxytocin facilitate social recognition in rats. Psychopharmacology (106) 71-74. Popik, P., Vetulani, J., Bisaga, A., and Van Ree, J.M. 1991. Recognition cue in the rat’s social memory paradigm. J Basic Clin Physiol Pharmacol 2: 315-327. Prediger, R.D., and Takahashi, R.N. 2003. Ethanol improves short-term social memory in rats. Involvement of opiod and muscarinic receptors. Eur J Pharmacol. Feb 21; 462(1-3): 115-23. Ramamoorthy, K., Gupta, M.S., Sun, G., McDougal, A., and Safe, S.H. 1999. 3, 3’, 4, 4’-Tetrachlorobiphenyl exhibits antiestrogenic and antitumorigenic activity in the rodent uterus and mammary cells and in human breast cancer cells. Carcinogenesis 20(1): 115-123. Sanchez-Alsono, J., Lopez-Aparicio, P., Recio, M., and Perez-Albarasnaz, M. 2003. Apoptosis-mediated neurotoxic potential of a planar (PCB 77) and nonplanar (PCB 153) polychlorinated biphenyl congeners in neuronal cell cultures Toxicol Lett 144:37-349. Safe, S. 1990. Polychlorinated biphenyls (PCBs), dibenzo-p-dioxins (PCDDs), dibenzofurans (PCDFs), and related compounds: environmental and mechanical considerations which support the development of toxic equivalency factors (TEFs). Crit Rev Toxicol 21(1):51-88. Safe, S.H. 1994. Polychlorinated biphenyls (PCBs): environmental impact, biochemical and toxic responses and implications for risk assessment. Crit Rev Toxicol. 24(2): 87-149. Safe, S., and Krishnan,V. 1995. Chlorinated hydrocarbons: estrogens and antiestrogens. Toxicol Lett.82-83: 731-736. Schmidt, K., Steinberg, C.E.W., Pflugmacher, S., and Staaks, G.B.O. 2004. Xenobiotic substances such as PCB mixtures (Aroclor 1254) and TBT can influence swimming behavior and biotransformation activity (GST) of carp (Cyprinus carpio). Environmental Toxicology 19(5):460-470. Seegal, R.F., Brosch, K.O., and Okoniewski, R.J. 2005. Coplanar PCB congeners increase uterine weight and frontal cortical dopamine in the developing rat: implications for developmental neurotoxicity. Toxicol. Sci. 86(1): 125-131. Sekiguchi, R., Wolterink, G., and Van Ree, J.M. 1991. Short duration of retroactive facilitation of social recognition in rats. Physiol Behav 50: 1253-1256. Silberhorn, E.M., Glauert, H.P., and Robertson, L.W. 1990. Carcinogenicity of polychlorinated biphenyls: PCBs and PBBs. Crit Rev Toxicol 20: 439- 496. Sink, T., Steele, G., Smith, A.B., Watkins, K., and Shults, R.A. 1992. Mortality among workers exposed to polychlorinated biphenyls. Am J Epidemiol. 136: 389- 398.

51Storm, E.E., and Tecott LH. (2005). Social Recognition: peptidergic regulation of mammalian social behavior. Neuron 47(4) 483-486. Swaab, D.F., Purba, J.S., and Hofman, M.A. 1995. Alteration in the hypothalamic paraventricular nucleus and its oxytocin neurons (putative satiety cells) in Prader-Willi syndrome: a study of five cases. J. Clin Endocrinol Metab 80: 573-579. Tharappel, J.C., Lee, E.Y., Robertson, L.W., Spear, B.T.,Glauert, H.P. 2002. Regulation of cell proliferation, apoptosis, and transcription factor activities during the promotion of liver carcinogenesis by poly chlorinated biphenyls. Toxicol Appl Pharmacol 179: 172- 184. Thor, D.H., and Holloway, W.R. 1982. Social memory of the male laboratory rat. Journal of Comparative Psychology 96:1000-1006. Thor, D.H., Holloway, W.R. 1981. Persistence of social investigatory behavior in the male rat: Evidence for long-term memory of initial copulatory experience. Anim learn Behav 9:561-565. Tuomisto J. 2004. Dose- additive carcinogenicity of a defined mixture of dioxin- like compounds. Environ Health Perspect 112: 1217-1224. Uzei, A. 1986. Periods of sensitivity to thyroid hormone during the development of the organ of corti. Acta Otolaryngol.Suppl 429: 23-27. Van der Berg, M., Birnbaum, L., Bosveld, A., Brunstrom, B., Cook, P., Feely, M., Girsy, J., Hanberg, A., Hasegawa, R., Kennedy, S., Kubiak, T., Larsen, J., Rolaf van Leeuwen, F., Liem, A., Nolt, C., Peterson, R., Poellinger, L., Safe, S., Schrenk, D., Tillet, D., Tysklind, M., Younes, M., Waern, F., and Zacharewski, T. 1998. Toxic equivalency factors (TEFs) for PCBs, PCDDs, PCDFs for humans and wildlife. Environ Health Perspect 106:1-25. Viberg, H., Fredriksson, A., and Eriksson, P. 2003. Neonatal exposure to polybrominated diphenyl ether (PBDE 153) disrupts spontaneous behavior, impairs learning and memory, and decreased hippocampal cholinergic receptors in adult mice. Toxicology and applied Pharmacology 192 PP 95-106. Winslow, J.T., and Camacho, F. 1995. Cholinergic modulation of a decrement in social investigation following repeated contacts between mice .Psychopharmacology(Berl) 121:164-172. Winslow, T., Hastings, N., Carter, C.S., Harbaugh, C.R., Insel, T.R. 1993. A role for central vasopressin in pair bonding in monogamous prairie voles. Nature 365:545-548. <www.albany.edu/docs.sph/ poster_2005/abstracts/pdfs/shan > [accessed May 10, 2006

52APPENDIX
ELISA Test:
The oxytocin solutions were prepared in serial dilutions. The 10,000 pg/mL oxytocin
standard was allowed to warm at room temperature. The seven 12×75 mm glass tubes were
labeled #1 through #7. One mL of standard diluent was pipetted into tube 1 and 500 μL of the
standard diluent was pipetted into tube #2 through #7. One hundred μL of buffer was removed
from tube #1. 10,000 pg/mL standards were added to tube #1 and vortexed thoroughly, 500 μL of
tube #1 was added to tube #2 and vortexed thoroughly. Five hundred μL of tube #2 was added to
tube #3 and vortexed. The process was continued for tubes #4 through #7. This gives a
concentration of 1000, 500, 250, 125, 62.5, 31.2 and 15.6 pg/mL. The diluted standards were
used within 60 minutes of preparation. The oxytocin conjugate was allowed to warm at the room
temperature. The wash buffer was prepared by diluting 5 mL of the supplied concentrate with 95
mL of deionized water. One hundred μL of standard diluent was pipetted into the NSB and the
Bo (0 pg/mL) wells. One hundred μL of standards #1 through #7 were pipetted into appropriate
wells. Hundred μL of the samples was pipetted into the appropriate wells. Fifty μL of assay
buffer was pipetted into the NSB wells. Fifty μL of the blue conjugate was pipetted into each
well, except the total activity (TA) and blank wells. Fifty μL of the antibody was pipetted into
each well, except the TA and NSB wells. The plate was tapped gently to mix, sealed and
incubated at 4 degrees for 18-24 hours. The contents were emptied from the wells and washed by
adding 400 μL of the wash solution to each well. The washes were done for a total of three times.
The wells were emptied or aspirated after the final wash, and the plate was tapped dry on a paper
towel to remain any remaining wash buffer. Five μL of the blue conjugate was added to the TA
wells. Two hundred μL of the pNpp substrate solution was added to each well and incubated at

53room temperature for 1 hour without shaking. Fifty µl of the stop solution was added to each
well. This process stopped the reaction and the plate was ready to be read by a plate reader. The
plate