invasion of mesenchyme into three-dimensional collagen gels: a regional and temporal analysis of...
TRANSCRIPT

DEVELOPMENTAL BIOLOGY 95, 108-114 (1983)
Invasion of Mesenchyme into Three-Dimensional Collagen Gels: A Regional and Temporal Analysis of Interaction
in Embryonic Heart Tissue
RAYMONDB.RUNYAN'ANDROGERR.MARKWALD Department of Anatomy, Texas Tech University Health Sciences Center, Lubbock, Texas 79450
Received February 16, 1982; accepted in revised form September 10, 1982
In normal heart development the endothelium of the atrioventricular canal, but not the ventricle, produces mes- enchymal cells which seed (invade) into the intervening extracellular matrix toward the myocardium at around 64- 69 hr of development. We have utilized three-dimensional collagen substrates to examine the initiation of seeding by atrioventricular canal endothelia in vitro and to compare and contrast the responses of the ventricular endothelia. Explants of atrioventricular canals and ventricles from staged embryos were placed on the surfaces of collagen gels prior to the onset of seeding in situ. At varied intervals of incubation, the explant was removed, leaving behind a monolayer on the surface of the gel which consisted of endothelial cells. Subsequently, the endothelial outgrowths were examined for seeded cells. The results confirm the regional endothelial differences seen in vivo. They also show that invasion of the collagen gels is due to an alteration in phenotype mediated by interaction with other components of embryonic heart explant. Lastly, the time course of this tissue interaction in vitro mimics the onset of seeding in vivo.
INTRODUCTION
One of the major developmental events in the early heart is the regional acquisition of a mesenchymal phe- notype by the endothelia of the atrioventricular canal. In contrast, the adjacent endothelia of the ventricle remain unaltered. The question of how only one of these heretofore indistinguishable cell populations acquires this capability is an intriguing one. This mechanism is crucial to normal heart development since these mes- enchymal cells are the putative precursors of the valves and membranous septa (Markwald et ab, 1981).
Beginning at stage 17 and continuing through stage 18 (approximately 65-69 hr), some of the endothelial cells of the atrioventricular canal (an epithelial tissue) detach from one another, invade the underlying ECM, and migrate toward the myocardium (Bolender and Markwald, 1979). Adjacent endothelial cells of the ven- tricle area do not demonstrate this ability to seed into the ECM and migrate at this time (Markwald et ah, 1975). Several studies have indicated spatial and tem- poral correlation of the composition and properties of the glycosaminoglycans of the ECM with the events of this mesenchyme formation (Markwald and Adams- Smith, 1972; Markwald et ah, 1977,1978). Other studies with the inhibitor DON, a glutamine analog, have dem-
’ To whom all correspondence should be addressed. ’ Abbreviations used: ECM, extracellular matrix; DON, 6-diazo-5-
0x0-L-norleucine.
onstrated a concomitant impairment of mesenchyme cell formation and migration and the incorporation of radioisotope into glycosaminoglycans (Markwald and Bernanke, 1981; Markwald et ah, 1979). However, the use of inhibitors or subtractive agents such as enzymes cannot totally separate casually or causally related ef- fects.
Bernanke and Markwald (1979, 1982) recently devel- oped an in vitro model of cardiac mesenchyme forma- tion utilizing a repolymerized collagen lattice. They observed that atrioventricular canal explants produce an endothelial monolayer over the surface of collagen gels and that some of these endothelial cells detach and seed into the matrix of the gels in a manner which mimics the formation of the mesenchymal cells in vivo. They also observed, but did not pursue, that ventricular endothelia do not appear to invade the underlying gel. A number of investigators have also observed that other mesenchymal cell types including fibroblasts, tumor cells, and smooth muscle cells have the ability to invade collagen lattices (both natural and reconstituted), while a variety of definitive epithelial cell types do not (Over- ton, 1977, 1979; Schor, 1980; Delvos et al., 1982).
This study was undertaken to determine: (1) whether the temporal onset of seeding seen in situ is mimicked by atrioventricular endothelial cells in culture; (2) whether regional differences in seeding seen in situ are maintained in culture; and (3) whether the acquisition of seeding in vitro is dependent on an interaction with
108 0012-1606/83/010108-07$03.00/O Copyright 0 1983 by Academic Press. Inc. All rights of reproduction in any form reserved.

RUNYAN AND MARKWALD Mesenchymal Seeding into Collagen Gels 109
A 0 A n’ B’ B A Al B’ B
FIG. 1. Diagram of the experimental manipulation. An atrioventricular canal explant was placed at site A and the ventricular explant was placed at site B. After an interval, varying from 8 to 22 hr of incubation, the explant at site A was moved to A’ and the explant at site B was moved to B’. 24 hr later, the collagen gel was examined at all four sites A, A, B, and B for seeding into the lattice.
other components of embryonic heart, or if it is an in- dependent programmed event.
The use of the collagen gel system permits the iso- lation and examination of the phenomena of cell seeding which would be intractable under normal two-dimen- sional culture conditions. Since such epithelial-mes- enchymal transformations (i.e., the acquisition of a migratory phenotype bby an ostensibly epithelial cell) occur throughout the early embryo (Trelstad et ak, 1982), these data may have a more general significance in un- derstanding embryonic development.
MATERIALS AND METHODS
Preparation of collagen lattices. Three-dimensional collagen substrates (1.86 mg protein/ml) of repolymer- ized rat tail collagen were prepared according to the method of Bernanke and Markwald (1979, 1982) in 35 mm tissue culture dishes. The gels were rinsed once with 1 ml medium 199 (Gibco), followed by a rinse with 1 ml medium 199 plus 2% fetal calf serum (Gibco). Fol- lowing a final addition of medium with serum, the cul- ture dishes were incubated in 5% CO, at 37°C for one hour prior to the addition of the explants. The medium on some cultures was supplemented with 5 pug/ml of insulin, 5 pg/ml of transferrin, and 5 ng of selenium (ITS supplement, Collaborative Research). This re- sulted in a greater outgrowth from the explants. There were no observed differences in the percentages of seeded cultures between medium with or without the ITS supplement.
Experimental protocol. Fertilized White Leghorn chicken eggs (Poultry Science Dept., Texas A & M Uni- versity) were incubated at 3’7.5”C for 48-72 hr. Stages 12 to 18 (Hamburger and Hamilton, 1951) were utilized since this period brackets the time from the initial en- largement of the atrioventricular canal ECM to the appearance of a large population of mesenchymal cells. Embryos were collected and placed in sterile Tyrode’s balanced salt solution (Gibco). After careful staging, the hearts were removed and the atria and outflow
tracts cut away. The remaining tissue was separated into atrioventricular canal and ventricular portions and cut longitudinally to expose the central lumen. These portions of the heart were explanted onto the surface of drained collagen gels (i.e., the dishes were decanted so that the only medium remaining, approximately 1 ml, was contained within the collagen latticework). After a period of time, the explants were moved to a new portion of the gel (see Fig. l-AV explant moved from A to A’ and ventricular explant moved from B to B’), leaving behind the initial outgrowth. In order to examine the timing of tissue interaction in culture, the period of explant incubation (at sites A and B) was varied between 8 and 22 hr. Twenty-four hours after explant removal, the cultures were examined with phase microscopy and scored for the presence of seeded cells. The primary criterion for seeding was defined as the presence of cells whose plane of focus was obviously beneath the polygonal endothelial cells that remained on the surface of the gel. Seeded cells also displayed a characteristic fusiform or stellate morphology with ex- tended filopodia. A culture was not scored as “seeded” unless there were at least three cells clearly visible be- neath the gel surface.
The myocardium in both the atrioventricular canal and ventricular explants continued to beat throughout the duration of each culture. This served as a useful marker for monitoring the completeness of removal of the explant since cultures which exhibited beating (myocardial) cells at their original sites (A or B) were excluded from the data.
From these data, scatter plots of time of explant in- cubation vs whether or not the culture was seeded were made. Linear regression analysis of the scatter plot for each stage was performed and the regression lines ob- tained were graphed as the probability of observing a seeded culture x 100 vs the time of explant incubation. The totals for each stage were grouped together and analyzed by Duncan’s multiple range test. Represen- tative cultures were photographed on a Leitz zernicke phase light microscope or on a Wild Photomicroscope.
DEVELOPMENTALBIOLOGY VOLUME 95,1983
-
F , E

RUNYAN AND MARKWALD Mesenchymal Seeding into Collagen Gels 111
RESULTS
Growth of the Explants in Culture
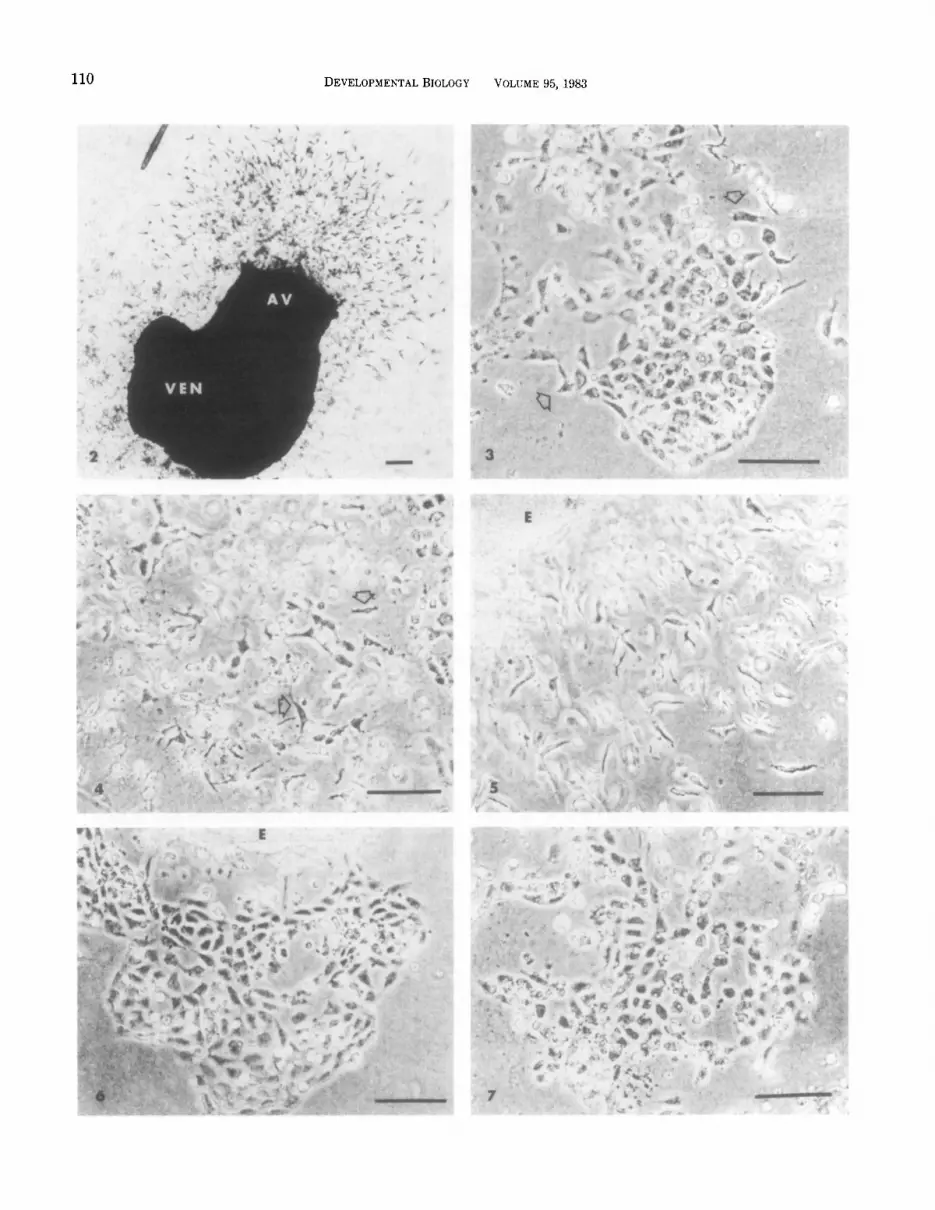
Explants taken from the embryonic heart during stages 12-17 contained only two detectable cell phe- notypes, myocardial cells and endothelial cells (Mana- sek, 1969; Bernanke and Markwald, 1982). The endo- thelial cells grew out as ;a monolayer on the surface of the gel while the myocar’dial portion of the explant re- mained a tightly clustered ball of cells surrounded by a layer of adhering endothelium. As shown in Fig. 2, only the endothelium derived from the atrioventricular canal region of the heart seeded the collagen gel. En- dothelial cells which proliferated from the ventricular region remained on the surface. These cells are out of focus in Fig. 2. The proliferative ability of the endo- thelial cells derived from the ventricular and atrioven- tricular canal explants was consistently different at all stages examined. The ventricular endothelia did not spread out over the surface of the gel to as great an extent as did those of the atrioventricular canal ex- plants. Only 84% of the ventricular explants were able to produce a monolayer (outgrowth at the original site (B) whereas 99% of atrioventricular canal explants pro- duced an endothelial outgrowth (site A). In both the atrioventricular canal and ventricular monolayers, two forms of endothelial cells could be seen on the gel sur- face (Figs. 3, 6, and 7): polygonal cells closely apposed to one another (typical of endothelial cells), and cells with pseudopodia or lamlellipodia extending from their surfaces similar to those :seen in two-dimensional tissue culture. This second type of appearance was found pri- marily among isolated cells and along the free border of cell clusters (arrow, Fig. 3).
Seeding of Atrioventricular Canal Outgrowths
Monolayers derived from stage 12 atrioventricular canal explants (n = 10) in which the explant was re- moved prior to 22 hr in culture (from site A) failed to exhibit seeding of the collagen gel. Continued incuba- tion of the explants at site A’ for another 24 hr resulted
in the appearance of seeded mesenchymal cells beneath the explants (not shown). Older stages showed the ap- pearance of seeding with correspondingly less time of incubation but, by 24 hr after explant transfer to site A’ (Fig. l), an average of 97% of all atrioventricular canal explants, regardless of stage at the time of initial explantation, had initiated seeding of cushion cells into the collagen gel.
Figures 3,4, and 5 depict differences seen with varied periods of explant incubation. An unseeded atrioven- tricular canal monolayer is shown in Fig. 3. In this ex- ample, a stage 14 explant removed after 14 hr in culture exhibited no seeded cells when the persisting endothe- lial monolayer was examined 24 hr later. Figure 4 shows a stage 16 monolayer in which the explant was removed after a similar period of time. In this case, the mono- layer exhibited a number of seeded cells. Finally, Fig. 5 shows the result of extended incubation of the en- dothelium with the myocardium at site A’. The explant is surrounded by a halo of seeded mesenchymal cells. The relationship between the time of explant incubation in culture and the appearance of seeded cultures for each stage is shown in Fig. 8. With increasing time of incubation of the explant at site A, there was an in- crease in the percentage of seeded cultures observed for all stages. Cultures incubated less than 8 hr did not have a sufficient outgrowth from the explant to be scored. Isolated atrioventricular monolayers which were not seeded by 24 hr after the removal of the ex- plant did not appear to acquire this ability later. Un- seeded monolayers were observed for up to 2 weeks without the appearance of seeded cells in the gels.
Statistical confirmation of the difference in seeding ability between stages is shown in Table 1. The grouped data for each stage show a progression toward a greater percentage of seeded cultures with increasing age in ovo prior to explantation.
Seeding of Ventricular Outgrowths
The appearance of a ventricular explant (site B’, Fig. 1) is shown in Fig. 6. The closely knit endothelial out-
FIG. 2. A macroscopic view of heart laid on the surface of a collagen gel. Focus is beneath the surface of the gel. The atrium and outflow tract of this stage 17 heart were cut away to permit access of the endothelial cells to the gel surface. The atrioventricular canal (AV) endothelium can be seen to invade collagen gel while no seeded cells can be seen from the cut end of the ventricle (VEN). Bar = 100 pm.
FIG. 3. A phase light micrograph of an unseeded atrioventricular canal monolayer. Focus is on the gel surface. This monolayer was the outgrowth of a stage 14 heart which was present on the gel surface for 14 hr. Arrows indicate cells with extended cell processes. Bar = 50 firn.
FIG. 4. A phase light micrograph of a seeded atrioventricular canal monolayer. The image was focused beneath the surface of the gel to show seeded, fusiform and stellate cells (arrows). This outgrowth was derived from a stage 16 heart which was present on the surface for 14 hr. Bar = 50 pm.
FIG. 5. A phase light micrograph of a seeded stage 16 atrioventricular canal explant. As in Fig. 4, the image was focused beneath the surface to demonstrate seeded cells (arrow). E, explant. Bar = 50 Frn.
FIG. 6. A phase light micrograph of an unseeded stage 15 ventricular explant (E). Focus is on the gel surface. Bar = 50 Frn. FIG. ‘7. A phase light micrograph of an unseeded ventricular monolayer from a stage 15 heart. Focus is on the gel surface. Bar = 50 pm.

112 DEVELOPMENTAL BIOLOGY VOLUME 95, 1983
growth has fewer free cells on the surface of the gel than are typically seen in an atrioventricular canal out- growth. Figure 7 depicts a ventricular monolayer from site B (Fig. 1). Again, the cells are seen to be generally polygonal in shape with no evidence of seeded fusiform or stellate cells.
The results of observations on seeding by cells derived from ventricular outgrowths are shown in Fig. 9. No relationship between age of the explant and the time of incubation was observed until stage 18. The cumu- lative data for each stage are shown in table 1. Very low percentages of cultures with seeded cells were seen prior to stage 18.
8 IO 12 14 16 18 20 22
a EXPLANT INCUBATION TIME (HR.)
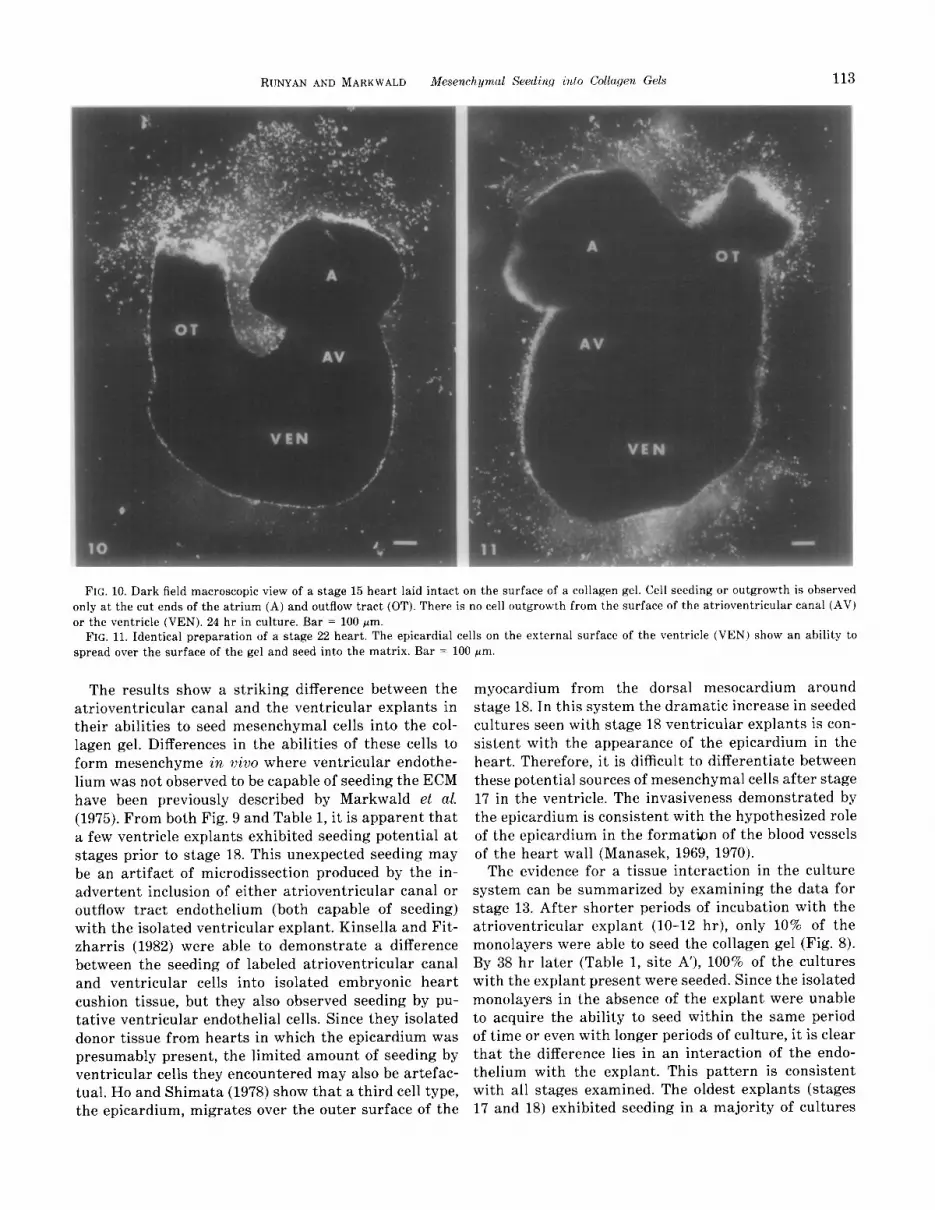
The increased percentage of seeded cultures seen with stage 18 explants may be attributed to the initial ap- pearance of a third cell type (epicardium) on the surface of the ventricle at this stage (Table 1). To test whether the epicardium is capable of seeding, an intact stage 15 (no epicardium present) heart and an intact stage 22 (epicardium present) heart were laid on the gel surface. As shown in Figs. 10 and 11, the younger heart showed outgrowth or seeding only at the cut ends of the atrium and outflow tract while the older heart showed an area of seeded cells apparently derived from the ventricular surface.
DISCUSSION 9 EXPLANT INCUBATION TIME #if?.)
The major events of atrioventricular cushion for- mation were recently shown to occur in three-dimen- sional collagen gel culture in the exact histological se- quence as they occur in situ (Bernanke and Markwald, 1982). One importance of this observation was the op- portunity to experimentally discern causal or casual interrelationships between endothelium (progenitor of mesenchyme) and the myocardium and its multitude
FIGS. 8,9. Linear regression plots of the relationship between seed- ing and incubation time prior to explant removal. The numbers at the ends of the lines indicate the stage of the embryo at the time incu- bation started. Figure 8 shows the data for atrioventricular canal endothelia and Fig. 9 is the corresponding plot for ventricular en- dothelia.
TABLE 1 PERCENTAGE SEEDED CULTURES (CUMULATIVE TOTALS)
AV canal* monolayer AV explant Ven monolayer Ven explant
Stage” (site A) (site A’) (site B) (site B’)
13 [48-521 18 (23) A” loo (17) 15 (12) X,Y 0 (17) 14 [50-531 27 (26) A 97 (36) 0 (20) x 12 (34) 15 [50-551 61 (54) B 94 (64) 0 (41) x 3 (64) 16 [51-561 71 (62) B,C 98 (62) 5 (48) X 11 (62) 17 [52-641 79 (48) B,C 100 (36) 10 (44) x 14 (36) 18 [65-691 87 (28) C 96 (28) 35 (23) Y 39 (28)
of matrical secretory products. Of course, the validity of the collagen gel system as an in vitro model for cush- ion morphogenesis requires more than histological sim- ilarity. This study tested the morphogenetic potential of the model further and found that: (1) chronologically, the in situ timing sequence of endothelial seeding is mimicked in culture; (2) regional variation in the in situ capacity to form mesenchyme is duplicated; and (3) en- dothelial seeding as suggested from in situ inhibitor studies (Markwald and Bernanke, 1979) cannot occur independent of interaction with other tissue compo- nents of the embryonic heart.
a Stage of embryo from which heart explant was taken. Numbers in brackets indicate the approximate age of the embryo in hours (Hamburger and Hamilton, 1951).
b Abbreviations: AV, atrioventricular canal; Ven, ventricle. “Parentheses indicate the number of cultures scored within this
group. Identical capital letters indicate that the totals are not sig- nificantly different. 01 = 0.05.
Seeding was observed in a majority of cultures only after the total chronological age (initial age of explant plus time in culture) of the tissue reached 64-69 hr. Therefore, stage 15 explants required more time in cul- ture to demonstrate a majority of seeded cultures than did older stages. The final evaluation of the cultures 24 hr after explant removal was chosen empirically since the youngest explants (at site A) took that long to manifest seeding.
RUNYAN AND MARKWALD Mesench ymal Seeding Ato Collagen Gels 113
FIG. 10. Dark field macroscopic view of a stage 15 heart laid intact on the surface of a collagen gel. Cell seeding or outgrowth is observed only at the cut ends of the atrium (A) and outflow tract (OT). There is no cell outgrowth from the surface of the atrioventricular canal (AV) or the ventricle (VEN). 24 hr in culture. Bar = 100 pm.
FIG. 11. Identical preparation of a stage 22 heart. The epicardial cells on the external surface of the ventricle (VEN) show an ability to spread over the surface of the gel and seed into the matrix. Bar = 100 pm.
The results show a striking difference between the atrioventricular canal and the ventricular explants in their abilities to seed mesenchymal cells into the col- lagen gel. Differences in the abilities of these cells to form mesenchyme in vivo where ventricular endothe- lium was not observed to be capable of seeding the ECM have been previously described by Markwald et al. (1975). From both Fig. 9 and Table 1, it is apparent that a few ventricle explants exhibited seeding potential at stages prior to stage 18. This unexpected seeding may be an artifact of microdissection produced by the in- advertent inclusion of either atrioventricular canal or outflow tract endothelium (both capable of seeding) with the isolated ventricular explant. Kinsella and Fit- zharris (1982) were able to demonstrate a difference between the seeding of labeled atrioventricular canal and ventricular cells into isolated embryonic heart cushion tissue, but they also observed seeding by pu- tative ventricular endothelial cells. Since they isolated donor tissue from hearts in which the epicardium was presumably present, the limited amount of seeding by ventricular cells they encountered may also be artefac- tual. Ho and Shimata (1978) show that a third cell type, the epicardium, migrates over the outer surface of the
myocardium from the dorsal mesocardium around stage 18. In this system the dramatic increase in seeded cultures seen with stage 18 ventricular explants is con- sistent with the appearance of the epicardium in the heart. Therefore, it is difficult to differentiate between these potential sources of mesenchymal cells after stage 17 in the ventricle. The invasiveness demonstrated by the epicardium is consistent with the hypothesized role of the epicardium in the formattin of the blood vessels of the heart wall (Manasek, 1969, 1970).
The evidence for a tissue interaction in the culture system can be summarized by examining the data for stage 13. After shorter periods of incubation with the atrioventricular explant (lo-12 hr), only 10% of the monolayers were able to seed the collagen gel (Fig. 8). By 38 hr later (Table 1, site A’), 100% of the cultures with the explant present were seeded. Since the isolated monolayers in the absence of the explant were unable to acquire the ability to seed within the same period of time or even with longer periods of culture, it is clear that the difference lies in an interaction of the endo- thelium with the explant. This pattern is consistent with all stages examined. The oldest explants (stages 17 and 18) exhibited seeding in a majority of cultures

114 DEVELOPMENTAL BIOLOGY VOLUME 95, 1983
which was not prevented by the prompt removal of the explant. In these cases, it appears that the maturational interaction(s) had previously occurred in situ, and that the endothelial outgrowth was competent to seed mes- enchyme without the continued presence of the explant.
Order in Growth” (D. Rudnick, ed.), pp. 233-256. Princeton Univ. Press, Princeton, N. J.
The data show that a product or a portion of the myocardial explant promotes seeding of endocardial cushion tissue cells into a three-dimensional collagen lattice. The experimental protocol reported here does not differentiate the source of the tissue interaction. One cannot tell from these data whether the differences seen between the atrioventricular and ventricular en- dothelia are a product of regional differences in the interacting component or components of the explant or are a result of differences in the ability of the endo- thelial cells to respond. Preliminary experiments which would support the second alternative, that the differ- ences lie in the response of the cells, have recently been reported elsewhere (Runyan et al., 1982).
Experiments devised to disrupt the normal produc- tion of the ECM by the use of synthetic inhibitors (Markwald et al., 1979; Markwald and Bernanke, 1981) show a concomitant disruption of cushion tissue for- mation and migration. There is also evidence in a num- ber of other systems that ECM can mediate tissue in- teraction (for reviews see Grobstein, 1955, 1967; Hay, 1977; Hay and Meier, 1978). It seems plausible, for these reasons, that the tissue interaction observed in these studies can be mediated by the ECM, but this remains to be proven. Further studies are underway to clarify this point by attempting to isolate components of the explants and test them against unseeded but potentially receptive monolayers.
GROBSTEIN, C. (1967). Mechanisms of organogenetic tissue interaction. Nat. Cancer Inst. Mmzogr. 26, 279-299.
HAMBURGER, V., and HAMILTON, H. L. (1951). A series of normal stages in the development of the chick embryo. J. Mwphol. 88,49- 92.
HAY, E. D. (1977). Cell-matrix interaction in embryonic induction. In “International Cell Biology 1976-1977” (B. R. Brinkley and K. R. Porter, eds.), pp. 50-5’7. Rockefeller Univ. Press, New York.
HAY, E. D., and MEIER, S. (1978). Concept of embryonic induction; inductive mechanisms in orofacial morphogenesis. In “Textbook of Oral Biology” (J. Shaw, E. Sweeney, C. Cappucino, and S. Meller, ed.), pp. 3-23. Saunders, Philadelphia.
HO, E., and SHIMADA, Y. (1978). Formation of the epicardium studied with the scanning electron microscope. Dev. BioL 66, 579-585.
KINSELLA, M. G., and T. P. FITZHARRIS (1982). Control of cell migra- tion in atrioventricular pads during chick early heart development: Analysis of cushion tissue migration in vitro. Dev. BioL 91, l-10.
MANASEK, F. J. (1969). Embryonic development of the heart. II. For- mation of the epicardium. J. Embryol. Exp. Mwrphol. 22, 233-248.
MANASEK, F. J. (1970). Histiogenesis of the embryonic myocardium. Amer. J. CardioL 25, 149-168.
MANASEK, F. J., REID, M., VINSON, W., SEYER, J., and JOHNSON, R. (1973). Glycosaminoglycan synthesis by the early embryonic chick heart. Dev. BioL 35, 332-348.
MARKWALD, R. R., and ADAMS-SMITH, W. N. (1972). Distribution of mucosubstances in the developing rat heart. J Histochem. Cytv them. 29, 896-907.
MARKWALD, R. R., FITZHARRIS, T. P., and ADAMS-SMITH, W. N. (1975). Structural analysis of endocardial cytodifferentiation. Dev. BioL 42, 160-180.
MARKWALD, R. R., and BERNANKE, D. H. (1981). Structural analyses of 6-diazo-5-oxo-L-norleucine effects on cardiac tissue morphogen- esis. In “Cardiac Morphogenesis and Teratology” (T. Pexieder, ed.). Perspect. Cardiovasc. Res. 5, 237-251.
MARKWALD, R. R., FITZHARRIS, T. P. BANK, H., and BERNANKE, D. H. (1978). Structural analyses on the matrical organization of glycos- aminoglycans in developing endocardial cushion. Dev. BioL 62,292- 316.
This work was supported by NIH Grant HL 19136 and a RCDA grant to Dr. Roger R. Markwald. The preparation of Fig. 1 by Waid Griffin and the assistance of S.D. Prien and Valinda Bradshaw in the preparation of the paper are gratefully acknowledged. We thank Dr. David Bernanke for both technical assistance and fruitful discussions.
MARKWALD, R. R., FITZHARRIS, T. P., and MANASEK, F. J. (1977). Structural development of endocardial cushions. Amer. J. Anat. 148, 85-120.
REFERENCES
BERNANKE, D. H., and MARKWALD, R. R. (1979). Effects of hyaluronic acid on cardiac cushion tissue cells in collagen matrix cultures. Tez Rep. BioL Med. 39, 271-285.
BERNANKE, D. H., and MARKWALD, R. R. (1982). Cardiac cushion morphogenetic events in a three-dimensional collagen lattice cul- ture model. Dev. Biol 91,235-245.
MARKWALD, R. R., FUNDERBURG, F. M., and BERNANKE, D. H. (1979). Glycosaminoglycans: Potential determinants in cardiac morpho- genesis. Tex. Rep. BioL Med 39, 271-285.
MARKWALD, R. R., KROOK, J. M., KITTEN, G. T., and RUNYAN, R. B. (1981). Endocardial cushion tissue development: Structural analysis on the attachment of extracellular matrix to migrating mesenchy- ma1 cell surfaces. SEM 19Sl(II), 261-274.
OVERTON, J. (1977). Response of epithelial and mesenchymal cells to culture on basement lamella observed by scanning microscopy. Exp. Cell Res. 105, 313-323.
BOLENDER, D. L., and MARKWALD, R. R. (1979). Epithelial-mesenchy- ma1 transformation in chick atrioventricular cushion morphogen- esis. SEM 1979(III), 313-321.
DELVOS, U., GAJDUSEK, G., SAGE, H., HARKER, L. A., and SCHWARTZ, S. M. (1982). Interactions of vascular wall cells with collagen gels. Lab. Invest. 46, 61-72.
GROBSTEIN, C. (1955). Tissue interaction in the morphogenesis of mouse embryonic rudiments in vitro. In “Aspects of Synthesis and
OVERTON, J. (1979). Differential response of embryonic cells to culture on tissue matrices. Tissue Cell 11, 89-98.
RUNYAN, R. B., KITTEN, G. T., and MARKWALD, R. R. (1982). Proteins of the extracellular matrix: Regional and temporal correlation with tissue interaction in the heart. In “The Extracellular Matrix” (S. Hawkes and J. Wang, eds.), Academic Press, New York, in press.
SCHOR, S. L. (1980). Cell proliferation and migration on collagen sub- strata in vitro. .I CeZl Sci. 41, 159-175.
TRELSTAD, R. L., HAYASHI, A., HAYASHI, K., and DONAHOE, P. K. (1982). The epithelial-mesenchymal interface of the male rat mul- lerian duct: Loss of basement membrane integrity and ductal regression. Dew. BioL 92, 27-40.