intracellular trafficking of lrp9 is dependent on two acidic cluster/dileucine motifs
TRANSCRIPT

Histochem Cell Biol (2008) 130:315–327
DOI 10.1007/s00418-008-0436-5ORIGINAL PAPER
Intracellular traYcking of LRP9 is dependent on two acidic cluster/dileucine motifs
Rémi Boucher · Heidi Larkin · Julie Brodeur · Hugo Gagnon · Caroline Thériault · Christine Lavoie
Accepted: 21 April 2008 / Published online: 7 May 2008© Springer-Verlag 2008
Abstract LDL receptor-related protein 9 (LRP9) is a dis-tant member of the low-density lipoprotein receptor(LDLR) superfamily. To date, there are no reports on thecellular distribution of LRP9 or the signals responsible forits localization. Here, we investigated the intracellularlocalization and traYcking of LRP9. Using confocalmicroscopy, we demonstrated that LRP9 was not present atthe plasma membrane but co-localized with various mark-ers of the trans-Golgi network (TGN) and endosomes. Thisco-localization was dependent on the presence of twoacidic cluster/dileucine (DXXLL) motifs in the cytoplasmictail of LRP9, which interact with GGA proteins, clathrinadaptors involved in transport between the TGN and endo-somes. LRP9 is the Wrst example of a transmembrane pro-tein with an internal GGA-binding sequence in addition tothe usual C-terminal motif. An inactivating mutation(LL ! AA) in both DXXLL motifs, which completelyinhibited the interaction of LRP9 with GGA proteins, led toan intracellular redistribution of LRP9 from the TGN toearly endosomes and the cell surface, indicating that thetwo DXXLL motifs are essential sorting determinants ofLRP9. In conclusion, our results suggest that LRP9 cyclesbetween the TGN, endosomes and the plasma membranethrough a GGA dependent-traYcking mechanism.
Keywords LDLR related protein 9 (LRP9) · GGA (Golgi-localized, � ear-containing, ADP ribosylation
factor binding protein) · trans-Golgi network · Endosome · Acidic cluster/dileucine (DXXLL) motif · LDL receptor
Introduction
The low-density lipoprotein receptor (LDLR) family com-prises a large number of members that share common struc-tural features as the LDLRA repeats, LDLRB repeats,epidermal growth factor precursor-type repeats and a cyto-plasmic region which includes internalization motifs. Thevariable numbers and combinations of these elements cor-respond with the huge variety of ligands and functions ofthese receptors. A new member of this growing family,termed LDLR-related protein 9 (LRP9), was recently iden-tiWed (Sugiyama et al. 2000). LRP9 is part of a new sub-family of the LDLR superfamily that includes two otherreceptors, LRP3 and LRP12 (previously named st7) (Battleet al. 2003). Like all LDLR family members, this new sub-group contains LDLRA domain repeats in their extracellu-lar domains (Hussain et al. 1999; Strickland et al. 2002)that function as ligand binding sites (Yamamoto et al.1984). However, several unique structural features distin-guish this new LRP subfamily from prototypical membersof the LDLR superfamily (Battle et al. 2003): (1) theabsence of EGF-like repeats and YWTD-containingdomains in their extracellular domain; (2) the presence ofextracellular CUB domains that are found in a diverse arrayof functionally unrelated extracellular proteins (Bork andBeckmann 1993) and that are involved in ligand binding(Christensen and Birn 2002); and (3) a large cytoplasmictail that contains a proline-rich domain.
The important role of many members of the LDLRsubfamily like LDL, LRP, megalin, VDL and ApoERreceptors in endocytosis and lipid metabolism is well
R. Boucher · H. Larkin · J. Brodeur · H. Gagnon · C. Thériault · C. Lavoie (&)Department of Pharmacology, Faculty of Medicine and Health Sciences, Université de Sherbrooke, 3001-12e Avenue Nord, Sherbrooke, QC, Canada J1H 5N4e-mail: [email protected]
123

316 Histochem Cell Biol (2008) 130:315–327
known (Willnow 1999). However, several other biologicalfunctions are becoming more and more evident. Membersof the LDLR family are involved in the regulation of cellsurface protease activity, control of cellular entry of bacte-rial toxins and viruses, the transport and activation of ste-roid hormones, the regulation of Ca2+ homeostasis and alsoin cellular signaling events (Sugiyama et al. 2000; Herz2001; Strickland et al. 2002; Battle et al. 2003; Kikuchiet al. 2007; May et al. 2007)
Little is known about the function of LRP9. It has beenreported to be ubiquitously expressed and to be involved inthe internalization of apolipoprotein E (ApoE)-enriched�VLDL (Sugiyama et al. 2000). Additionally, putative sig-nals for endocytosis and intracellular traYcking (YXX�,dileucine, acidic cluster/dileucine [DXXLL] motifs) arepresent in the cytoplasmic tail of LRP9 (Sugiyama et al.2000). Moreover, several putative signal transductionmotifs (e.g., PDZ-binding domain, proline-rich region) aswell as potential phosphorylation sites have also been iden-tiWed in the cytoplasmic tail of LRP9 (Battle et al. 2003).LRP9 thus has the molecular characteristics of a traYckingand signalling member of the LDLR family.
GGA proteins (Golgi localized, � ear-containing, ADPribosylation factor-binding proteins) are monomeric adap-tor proteins that mediate the transport of cargo proteinsbetween the trans-Golgi Network (TGN) and endosomes(reviewed by Bonifacino 2004; Ghosh and Kornfeld 2004).They have a modular structure consisting of an amino-ter-minal VHS (Vps27, Hrs, STAM) domain, a GAT (GGAand TOM1) domain, a hinge region and a C-terminal �-adaptin “ear” (GAE) domain. The VHS domain binds toacidic cluster/dileucine (DXXLL (where X is any aminoacid)) sorting signals that are present in the cytoplamic tailof various transmembrane proteins such as mannose-6-phosphate receptor (MPR) (Puertollano et al. 2001; Zhuet al. 2001), sortilin (Nielsen et al. 2001), �-secretase (Heet al. 2002) and the LDLR member, LRP3 (Takatsu et al.2001). The GAT domain binds to the Arf family of GTP-binding protein, the hinge region binds to clathrin and theGAE domain binds to several accessory proteins (reviewedby Bonifacino 2004). Through these multiple interactions,GGAs are involved in the sorting of DXXLL-containingreceptors into coated vesicles.
In this study, we describe for the Wrst time the intracellu-lar localization and traYcking of LRP9. We observed byconfocal microscopy that LRP9 was predominantly localizedat the TGN, where it co-localized with GGA proteins, and toa lesser extent, in endosomes. We also demonstrated that theVHS domain of GGA proteins bound to two functionalDXXLL motifs in the cytoplasmic tail of LRP9. LRP9 is theWrst reported example of a transmembrane protein with aninternal GGA-binding sequence in addition to the usualC-terminal motif. Interestingly, mutation (LL ! AA) of
both DXXLL motifs prevented the interaction of LRP9 withGGA proteins and caused a redistribution of LRP9 in earlyendosomes and at the plasma membrane. These data demon-strated that LRP9 cycles between the TGN, endosomes andthe plasma membrane and suggested that GGA proteinsparticipate in the regulation of LRP9 traYcking.
Materials and methods
Antibodies and reagents
Rabbit antibodies against mannosidase II were generouslyprovided by Dr. Marilyn Gist Farquhar (University of Cali-fornia, San Diego, CA, USA). Anti-HA monoclonal anti-bodies (mAbs) were purchased from Covance (Berkley,CA, USA), anti-Myc mAbs from Cell Signaling Technol-ogy (Danvers, MA, USA), anti-FLAG mAbs from Sigma(St Louis, MO, USA) and anti-GM130 mAbs from BDTransduction Laboratories (Franklin Lakes, NJ, USA).Anti-GFP polyclonal antibodies (pAbs) were purchasedfrom Molecular Probes (Eugene, OR, USA), anti-HA pAbsfrom Covance, anti-EEA1 pAbs from AYnity Bioreagents(Golden, CO, USA), and anti-TGN46 pAbs from NovusBiologicals (Littleton, CO, USA). Brefeldin A and monen-sin were obtained from Sigma.
DNA constructs
Expression vectors for GGA1-Myc, GGA2-Myc andGGA3-Myc were kindly provided by Dr. Juan Bonifacino(National Institutes of Health, Bethesda, MD, USA).pcDNA-FLAG-tagged furin was kindly provided by Dr.Richard Leduc (Université de Sherbrooke, QC, Canada).Mammalian expression vector pMKITNeo encoding themurine LRP9-HA was kindly provided by Dr. T. Kitamura(University of Tokyo, Japan). PCR-based mutagenesis wasused to generate the LRP9 mutant constructs. To generatethe LRP9-proximal mutant, Leu692, Leu693 and Leu694 inthe C-terminal sequence DDVLLL were replaced by ala-nines. To generate the LRP9-distal mutant, Leu711 andLeu712 in the C-terminal sequence DEPLLA were replacedby alanines. To generate the double LRP9 mutant, Leu692,Leu693, Leu694, Leu711 and Leu712 in the two DXXLLmotifs in the C-terminal sequences (DDVLLL and DEP-LLA) were replaced by alanines. The sequences of the DNAconstructs generated by PCR were systematically veriWed.
Cell culture and transfection
COS7 cells were obtained from Dr. Klaus Hahn (Universityof North Carolina, Chapel Hill, NC, USA) and HEK293Tcells were obtained from Dr. Alexandra Newton (University
123

Histochem Cell Biol (2008) 130:315–327 317
of California, San Diego, CA, USA). The cells were grownin Dulbecco’s modiWed Eagle’s high glucose medium (Invit-rogen, Carlsbad, CA, USA) containing 10% foetal bovineserum (FBS) (Hyclone Laboratories, Logan, UT, USA),penicillin, and streptomycin. COS7 cells were transfectedusing Fugene6 transfection reagent (Roche Diagnostics,Indianapolis, IN, USA), while HEK cells were transfectedusing Lipofectamine 2000 transfection reagent (Invitrogen),both according to the manufacturers’ instructions.
ImmunoXuorescence
COS7 cells were plated on coverslips. Twelve hours aftertransfection, the cells were Wxed in 3% paraformaldehyde(PFA) in 100 mM phosphate buVer, pH 7.4 for 30 min, per-meabilized with 0.1% Triton X-100 for 10 min, blockedwith 10% goat serum for 30 min, and incubated with pri-mary antibodies for 1 h at RT, followed by Alexa Fluor-594or 488-conjugated antibodies (Molecular Probes) for 1 h atRT. Specimens were visualized using an inverted confocallaser-scanning microscope (FV1000, Olympus, Tokyo,Japan) with a PlanApo 60x/1.42 oil immersion objective(Olympus). Olympus Fluoview software version 1.6a wasused for image acquisition and analysis. The images werefurther processed using Adobe Photoshop (Adobe Systems,San Jose, CA, USA).
Glutathione S-transferase (GST) pull-down assays
GST constructs encoding GGA1-VHSGAT (residues 5–314), GGA1-VHS (residues 5–147), GGA1-GAT (residues148–314), GGA1-Hinge (residues 315–514), GGA1-GAE(residues 515–639) and GGA3-VHS (residues 1–146) weregenerated by standard PCR methods and subcloned intopGEX-KG (Amersham-Pharmacia Biotech, Piscataway, NJ,USA). GST fusion proteins were expressed in E. Coli BL21and puriWed on glutathione-Sepharose 4B beads (GE Health-care, Piscataway, NJ, USA) according to the manufacturer’sinstructions. Cells were lysed in buVer (50 mM Tris pH 7.4,200 mM NaCl, 1% NP40 and complete protease inhibitors(Roche)) for 1 h at 4°C and then centrifuged at 13,000£g for20 min. One mg of cleared lysate was incubated overnight at4°C with 10 �g GST fusion proteins immobilized on beads.Beads were washed three times in lysis buVer and boiled inLaemmli sample buVer. Bound proteins were analyzed bySDS-PAGE and detected by immunoblotting.
Coimmunoprecipitation
HEK cells were plated in 60 mm culture dishes and trans-fected with the various constructs. After 48 h, the cells werelysed in 50 mM Tris buVer (pH 7.4) containing 100 mMNaCl, 1% NP40 and protease inhibitors for 1 h at 4°C and
then centrifuged at 13,000 rpm for 20 min. Cleared super-natants were incubated with primary antibodies overnightat 4° and then with protein A-sepharose (GE Healthcare,Piscataway, NJ, USA) or protein G-Sepharose (Zymed, SanFrancisco, CA, USA) for 1 h. The beads were washed threetimes in lysis buVer and boiled in Laemmli sample buVer.Bound immune complexes were analyzed by SDS-PAGEand immunoblotting.
Immunoblotting
Protein samples were separated on 10% SDS-PAGE gelsand transferred to nitrocellulose membranes (Perkin Elmer,Woodbridge, ON, Canada). Membranes were blocked inTris-buVered saline (20 mM Tris–HCl, pH 7.5, 150 mMNaCl) containing 0.1% Tween 20 and 5% nonfat dry milkand incubated with primary antibodies for 2 h at RT andthen with horseradish peroxidase-conjugated goat anti-rab-bit or anti-mouse IgG (Bio-Rad, Richmond, ON, Canada)and enhanced chemiluminescence detection (Pierce Chemi-cal, Rockford, IL, USA).
Results
LRP9 localizes in the trans-Golgi network and early endosomes in COS7 cells
The cellular distribution of LRP9 has never been investi-gated. To characterize the intracellular localization of thisreceptor, we compared the distribution of LRP9 with severalspeciWc intracellular markers by confocal microscopy. HA-tagged LRP9 expressed in COS7 cells was not detected at thecell surface but was primarily localized in the juxtanuclearcompartment as well as in some scattered vesicles (Fig. 1a, d,g). The juxtanuclear distribution of LRP9 did not signiW-cantly co-localize with �-mannosidase II (Fig. 1a–c) orgalactosyl transferase (Fig. 1d–f), which are markers of themedial- and trans-Golgi cisternae, respectively. However,extensive co-localization with furin (Fig. 1g–i) and TGN46(Fig. 2e–h), which are markers of the TGN, was observed.To determine whether LRP9 is a Golgi resident protein or acargo protein in transit, COS7 cells transfected with LRP9-HA were treated for 1 h with cycloheximide, a protein syn-thesis inhibitor. This treatment did not alter the distribution ofLRP9-HA, which suggested that LRP9, which has a half-lifeof 4 h, remains in the Golgi for a long period of time and isnot just in transit to the plasma membrane (data not shown).
To conWrm that LRP9-HA speciWcally localized to theTGN compartment, experiments were performed withbrefeldin A (BFA) and monensin, two Golgi perturbingagents. BFA is a fungal metabolite known to inhibit coatformation on the Golgi membranes, thereby disrupting the
123

318 Histochem Cell Biol (2008) 130:315–327
cisternal stacks leading to a redistribution of the cis-,medial- and trans-cisternae in the endoplasmic reticulum,leaving the TGN to coalesce into a spherical mass near themicrotubule organizing centre (MTOC; (Banting and Pon-nambalam 1997; Chardin and McCormick 1999; Watsonand Pessin 2000). After BFA treatment, LRP9-HA showeda concentrated vesicular distribution that colocalized withthe TGN marker TGN46 (Fig. 2m–p). This diVers from thecis-Golgi marker GM130 (Fig. 2i–l) which remained asso-ciated with dispersed punctate structures as previouslydescribed (Nakamura et al. 1995; Xu et al. 2002). Monen-sin is an ionophore that disrupts the acidic pH of luminalcompartments and causes the accumulation of some Golgiproteins such as TGN38/46 and GPP130 in swollen vesiclesthat has been deWned as derivative of the TGN (Schaub et al.2006) or endosomes (Linstedt et al. 1997; Puri et al. 2002).Monensin treatment of COS7 cells led to the relocalizationof LRP9-HA and TGN46 to swollen vesicles (Fig. 2u–x)while GM130 staining remained in a characteristic perinu-clear Golgi pattern (Fig. 2q–t). These results support theTGN localization of LRP9. Confocal microscopy analysisof the intracellular localization of LRP9-HA also showed apartial co-localization with EEA1, an early endosomemarker (Fig. 3a–c). These LRP9-EEA1-labelled vesicles
mainly localized in the Golgi area and not at the peripheryof the cells (Fig. 3a–c). The presence of LRP9 in EEA1-labelled endosomes was conWrmed in COS7 cells transfectedwith an established Rab5 GTPase-deWcient mutant (Rab5Q79L)that greatly enlarges early endosomes due to increasedhomo- and heterotypic fusion (Stenmark et al. 1994)(Fig. 3d–f). These observations suggest that LRP9 predom-inantly localizes to the TGN and, to a lesser extent, to earlyendosomes at steady state in COS7 cells
LRP9 co-localizes with GGA proteins in COS7 cells
GGA proteins play a role of molecular adaptors in thetransport and retrieval of cargo proteins between the TGNand endosomal compartments (Puertollano et al. 2001; Zhuet al. 2001; Ghosh et al. 2003; Bonifacino 2004). GGA pro-teins directly bind to acidic cluster/dileucine (DXXLL)motifs located at the extreme end of the C-termini of cargoproteins (Bonifacino 2004; He et al. 2005). Since LRP9localized to the TGN and endosomes (Figs. 1–3) and sincethe C-terminal domain of LRP9 possesses a DXXLL motifsimilar to the one found in transmembrane proteinsreported to bind GGAs (LRP9-distal; Fig. 5a, b), we postu-lated that GGA proteins might interact with LRP9 and
Fig. 1 Comparison of LRP9 distribution with diVerent Golgi markers in COS7 cells. COS7 cells transfected with LRP9-HA alone (a–c) or together with galactosyl transferase-YFP (d–f) or furin-FLAG (G-I) were Wxed, permeabilized and immuno-stained using anti-HA (a, d, g), anti-mannosidase II (b), anti-GFP (e), anti-FLAG (h) antibod-ies. Stained cells were examined by confocal Xuorescence microscopy. Yellow colour in the merged images (c, f, i) indicates co-localization. Scale bar 10 �m.
123

Histochem Cell Biol (2008) 130:315–327 319
consequently regulate its intracellular traYcking. To test thishypothesis, we Wrst examined the co-localization of GGAisoforms with LRP9 by confocal microscopic imaging. Thiswas performed using COS7 cells expressing HA-taggedLRP9 together with Myc-tagged GGA1, GGA2 or GGA3.GGA1-Myc displayed both juxtanuclear and vesicular dis-tributions that substantially overlapped those of LRP9-HA(Fig. 4a–c). A similar distribution pattern was observed forGGA3-Myc (Fig. 4g–i) whereas partial co-localization wasobserved between GGA2-Myc and HA-LRP9 (Fig. 4d–f).
These results demonstrated that GGA proteins broadlyco-localize with LRP9 and suggested that GGA may beinvolved in the TGN-endosome transport of LRP9 througha potential interaction with its C-terminal domain.
GGA proteins interact with two DXXL motifs in the cytoplasmic tail of LRP9
To determine whether GGA proteins bind to LRP9, we Wrsttested whether GGA proteins co-immunoprecipitate with
Fig. 2 EVect of brefeldin A and monensin treatments on LRP9 intracellular localization. COS7 cells transfected with LRP9-HA were incubated in either the ab-sence (a–h) or presence of 10 �g/�l brefeldin A (i–p) or 10 �M monensin (q–x) for 60 min. Cells were Wxed, perme-abilized and immunostained using anti-HA (b, f, j, n, r, v), anti-GM130 (a, i, q) and anti-TGN46 (e, m, u) antibodies. Stained cells were examined by confocal Xuorescence micros-copy. Yellow colour in the merged images indicates co-localization. Last column show £2.5 magniWed views of the boxes. Scale bar 10 �m.
123

320 Histochem Cell Biol (2008) 130:315–327
LRP9 in HEK293 cells transfected with both HA-taggedwild-type LRP9 (WT) and Myc-tagged GGA1, GGA2 orGGA3. When the immunoprecipitations were carried out
using anti-HA IgGs, GGA1, GGA2 and GGA3 co-immuno-precipitated with LRP9-HA (Fig. 5c, lanes 2, 6, 10). How-ever, GGA2-Myc and GGA3-Myc bind better to LRP9-HA
Fig. 3 LRP9 partially co-local-izes with EEA1 on endosomes. COS7 cells were co-transfected with HA-tagged LRP9 alone (a–c) or together with Rab5Q79L, a Rab5 GTPase-deWcient mutant that creates enlarged early endo-somes (d–f). Cells were Wxed, permeabilized and immuno-stained using anti-HA (a, d) and anti-EEA1 (b, e) antibodies. Stained cells were examined by confocal Xuorescence micros-copy. Yellow colour in the merged images (c, f) indicates co-localization. Scale bar 10 �m.
Fig. 4 LRP9 co-localizes with GGA proteins at the TGN in COS7 cells. COS7 cells co-transfected with HA-tagged LRP9 (a, d, g) together with Myc-tagged GGA1 (b), GGA2 (e) or GGA3 (h) were Wxed, per-meabilized and immunostained using anti-HA (a, d, g) or anti-Myc (b, e, h) antibodies. Stained cells were examined by confocal Xuorescence microscopy. Yel-low colour in the merged images (c, f, i) indicates co-localization. Scale bar 10 �m.
123

Histochem Cell Biol (2008) 130:315–327 321
than GGA1-Myc. The same results were obtained when theimmunoprecipitation was carried out using anti-Myc IgGs.HA-LRP9 co-immunoprecipitated with GGA1-Myc,
GGA2-Myc and GGA3-Myc (Fig. 5d, lanes 2, 6, 10), thusconWrming the interaction of GGA proteins and LRP9 incultured cells.
Since an inactivating mutation of the dileucine residuesof the DXXLL sequence has previously been reported toabrogate the interaction with GGA proteins (Nielsen et al.2001; Puertollano et al. 2001; Takatsu et al. 2001; Zhuet al. 2001; He et al. 2002), leucines 711 and 712 werereplaced by alanines in the DXXLL motif at the extremeC-terminus of the cytoplasmic tail of HA-LRP9 (nameddistal DXXLL, Fig. 5a). Interactions between the variousGGA-Myc proteins and this LRP9 mutant were then inves-tigated using the co-immunoprecipitation assays describedabove with either anti-HA (Fig. 5c) or anti-Myc IgGs(Fig. 5d). The interaction between LRP9 and GGA1 wasalmost completely abolished (Fig. 5c, d, lane 4) by thismutation. However, the interactions between LRP9 andGGA2 and GGA3 were not aVected (Fig. 5c, d, lanes 8,12). These results suggested that the distal DXXLL motif isnot the only key element that contributes to the interactionwith GGA proteins. After further examination of the C-ter-minal sequence of LRP9, another DXXLL motif was iden-tiWed thirteen amino acids upstream from the distalDXXLL (Fig. 5a). This new DXXLL motif was composedof two overlapping DXXLL motifs (D689 D690 V691 L692
L693 L694) and was called the proximal DXXLL motif(Fig. 5a, b). Alanines were substituted for Leu692, Leu693
and Leu694 in the proximal DXXLL motif and interactionsbetween GGA proteins and the mutated LRP9 were ana-lyzed by immunoprecipitation using either anti-HA IgGs(Fig. 5c) or anti-Myc IgGs (Fig. 5d). As shown in Fig. 5cand d (lanes 3, 7, 11), the mutations introduced into theproximal DXXLL did not abrogate the interaction of LRP9with GGA1, GGA2 or GGA3. This indicated that both theproximal and distal DXXLL motifs might be responsiblefor the binding of GGA proteins to LRP9. To conWrm this,leucines (Leu692, Leu693, Leu694, Leu711 and Leu712) weresubstituted by alanines in both the proximal and distalDXXLL motifs of LRP9 cytoplasmic tail (Fig. 5a). Thisdouble DXXLL mutation completely inhibited the interac-tion of LRP9 with the three GGA proteins (Fig. 5c, d; lanes5, 9, 13), indicating that the C-terminal domain of LRP9contained two DXXLL motifs that were able to bind toGGA proteins. This discovery is of particular interestbecause LRP9 is the Wrst example of a transmembrane pro-tein with an internal GGA-binding sequence in addition tothe usual C-terminal motif.
We next determined which domain of GGA proteinsmediates the interaction with LRP9 using pull-down assays.Glutathione-S-transferase (GST) fusion proteins compris-ing the various GGA1 domains (Fig. 6a) were incubatedwith lysates from HEK293 cells expressing HA-taggedwild-type LRP9 (Fig. 6b). As a positive control, the pull-
Fig. 5 LRP9 interacts with GGA proteins via a proximal and distalDXXLL motif in its cytoplasmic tail. a Schematic representation of thecytoplasmic tail of LRP9 containing two DXXLL motifs: proximal anddistal (bold face letters). Underlined leucines were substituted by ala-nines. b Alignment of the C-terminal amino acids of transmembraneproteins that contain acidic cluster/dileucine signals in their cytosolictails. The two crucial leucine residues are shown in red and the crucialaspartate is shown in blue. The position of the transmembrane domain(Tm) and the number of residues before and/or after the signals is indi-cated. CD-MPR, cation-dependent mannose-6-phosphate receptor;CI-MPR, cation-independent mannose-6-phosphate receptor; LRP,low-density lipoprotein receptor related protein. c–d Co-immunopre-cipitations of wild-type and mutant LRP9 with GGA proteins. Lysatesfrom HEK cells transfected with wild-type (WT) or DXXLL mutantLRP9-HA [proximal (Pr), distal (Di) or double (Do)] and GGA1-Myc,GGA2-Myc or GGA3-Myc were immunoprecipitated with anti-HA (c,lanes 2–13), anti-Myc (d, lanes 2–13) or control (Ctl) (c, d, lanes 1 and14) antibodies and then immunoblotted with anti-Myc or anti-HA todetect GGA proteins and LRP9, respectively. Asterisk denotes longerexposure to show GGA1-Myc bands with same intensity as GGA3-Myc and GGA2-Myc. It highlights the fact that the interaction betweenLRP9-WT and GGA1-Myc is weaker than with GGA2-Myc andGGA3-Myc. Note that the level of expression of Myc-GGA3 isapproximatively six times the level of endogenous GGA3 proteins.
EDEDDVLLLPLAEPEVWVVEAEDEPLLA
686 713446
DistalProximal
TMA
B
C
D
123

322 Histochem Cell Biol (2008) 130:315–327
down-assay were also performed with lysates from cellsexpressing GFP-tagged cation-dependant mannose-6-phos-phate receptor (CD-M6PR-GFP) that contains a DXXLLmotif reported to interact with the VHS domain of GGA1(Puertollano et al. 2001). Similar to CD-M6PR-GFP,LRP9-HA bound strongly to GST-GGA1-VHS or GST-GGA1-VHSGAT and weakly to GST-GGA1-GAT(Fig. 6b). In contrast, no interaction was observed withGST, GST-GGA1-Hinge and GST-GGA1-GAE (Fig. 6b).Similar results were obtained with GST fusion proteinscomprising the various GGA3 domains (data not shown).These results demonstrated that LRP9 interacts predomi-nantly with the VHS domain of GGA proteins. The impor-tance of the DXXLL motifs in LRP9 interaction with theVHS domain of GGA was next examined. To this end,interactions of GST-GGA1-VHS with the double DXXLLmutants of LRP9-HA were compared to wild-type LRP9-HA. As shown in Fig. 6c, mutation of both DXXLL motifscompletely inhibited the interaction of LRP9 with the VHSdomain of GGA1. Similar results were obtained with GST-GGA3-VHS (data not shown). These results indicate thatthe DXXLL motifs of LRP9 interact speciWcally with theVHS domain of GGA proteins. This is in agreement withprevious studies showing that VHS is the speciWc GGAdomain that binds to the DXXLL motif of various of trans-membrane proteins such as sortilin, LRP3, �-secretase andM6PR (Puertollano et al. 2001; Takatsu et al. 2001; Heet al. 2002; Bonifacino 2004).
Mutations in both the proximal and distal DXXLL motifs induce a redistribution of LRP9 to the plasma membrane and peripheral endosomes
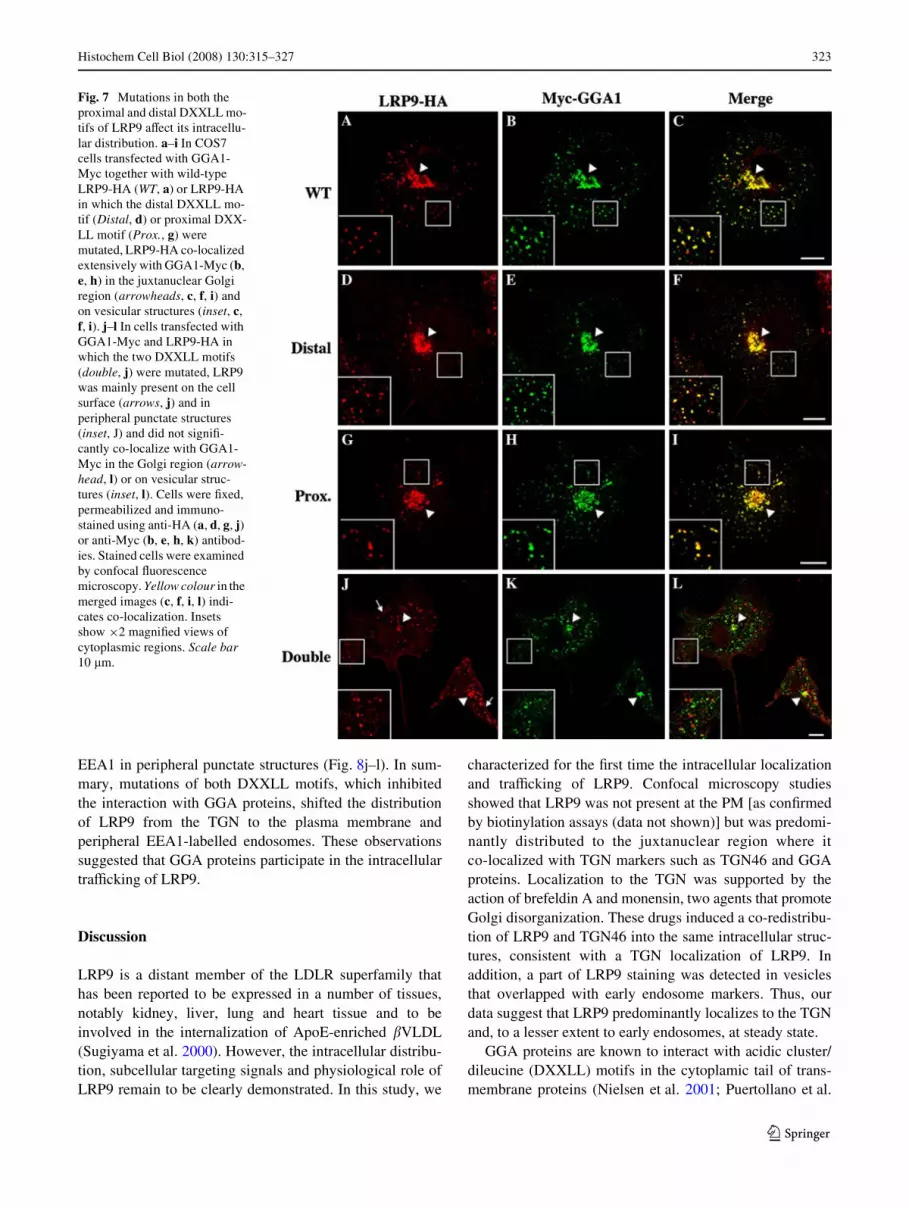
To investigate the role of the DXXLL motifs in LRP9traYcking, the subcellular distributions of the proximal,distal and double DXXLL mutants were compared to wild-type LRP9. COS7 cells were transfected with Myc-taggedGGA1 together with HA-tagged wild-type LRP9 or themutant DXXLLs and were analyzed by confocal micros-copy. The distributions of LRP9-HA mutated in the distal-(Fig. 7d) or proximal-DXXLL (Fig. 7g) motifs were identicalto the wild-type LRP9-HA (Fig. 7a). They all extensivelyco-localized with GGA1 in the juxtanuclear Golgi region(Fig. 7a–i, arrowheads) and in cytoplasmic vesicles (insets,Fig. 7a–i). In contrast, the double DXXLL mutant mainlylocalized at the cell surface and in peripheral punctatestructures (Fig. 7j) and did not co-localize with GGA1(Fig. 7l, inset). Similar data were obtained with cellsexpressing Myc-tagged GGA2 and Myc-tagged GGA3(data not shown). We noticed that the localization of Myc-GGA1 was also modiWed in cells expressing the doubleDXXLL mutant. GGA1 was less concentrated in the Golgiregion in these cells (Fig. 7k) compared to cells expressing
LRP9-WT (Fig. 7b), distal (Fig. 7e) or proximal mutants(Fig. 7h). This could be explained by the fact that cargoproteins help recruit GGAs onto membranes (Hirst et al.2007). Indeed, it has been reported that the overexpressionof CD8 chimeras with cytoplasmic tails containingDXXLL-sorting signals, which bind to GGAs, increases thelocalization of all three GGAs to juxtanuclear membranes(Hirst et al. 2007). In the same vein, we suggest that theoverexpression of LRP9 double DXXLL mutants that can-not bind GGA proteins would decrease GGA labelling inthe Golgi region compared to the cells that overexpressLRP9 that can bind GGA proteins.
To better deWne the subcellular localization of the doubleDXXLL mutant, the cells were transfected with HA-taggedwild-type LRP9 or the DXXLL mutants and were double-labelled with endogenous EEA1, an early endosome marker.Figure 8a–i shows that the wild-type LRP9 (Fig. 8a–c) andthe distal DXXLL (Fig. 8d–f) and proximal DXXLL(Fig. 8g–i) mutants did not co-localize with peripheralEEA1-labelled structures and were not present at theplasma membrane. In contrast, the double DXXLL mutantwas present at the plasma membrane and co-localized with
Fig. 6 LRP9 interacts with the VHS domain of GGA proteins. a Dia-gram of GGA1 showing the VHS (Vps27, Hrs, Stam) domain, theGAT (GGA and TOM) domain, the hinge segment and the GAE (�-adaptin ear) domain (Bonifacino 2004). b–c The indicated GST-fusionproteins immobilized on glutathione beads were incubated with lysatesfrom HEK cells expressing wild-type (WT) LRP9-HA or cation depen-dant mannose-6-phosphate receptor–GFP (CD-MPR-GFP, b) orLRP9-HA in which the two DXXLL motifs were mutated (Double, c).Bound proteins were separated by SDS-PAGE and immunoblottedwith anti-HA or anti-GFP antibodies to detect LRP9 and CD-M6PR,respectively.
123
Histochem Cell Biol (2008) 130:315–327 323
EEA1 in peripheral punctate structures (Fig. 8j–l). In sum-mary, mutations of both DXXLL motifs, which inhibitedthe interaction with GGA proteins, shifted the distributionof LRP9 from the TGN to the plasma membrane andperipheral EEA1-labelled endosomes. These observationssuggested that GGA proteins participate in the intracellulartraYcking of LRP9.
Discussion
LRP9 is a distant member of the LDLR superfamily thathas been reported to be expressed in a number of tissues,notably kidney, liver, lung and heart tissue and to beinvolved in the internalization of ApoE-enriched �VLDL(Sugiyama et al. 2000). However, the intracellular distribu-tion, subcellular targeting signals and physiological role ofLRP9 remain to be clearly demonstrated. In this study, we
characterized for the Wrst time the intracellular localizationand traYcking of LRP9. Confocal microscopy studiesshowed that LRP9 was not present at the PM [as conWrmedby biotinylation assays (data not shown)] but was predomi-nantly distributed to the juxtanuclear region where itco-localized with TGN markers such as TGN46 and GGAproteins. Localization to the TGN was supported by theaction of brefeldin A and monensin, two agents that promoteGolgi disorganization. These drugs induced a co-redistribu-tion of LRP9 and TGN46 into the same intracellular struc-tures, consistent with a TGN localization of LRP9. Inaddition, a part of LRP9 staining was detected in vesiclesthat overlapped with early endosome markers. Thus, ourdata suggest that LRP9 predominantly localizes to the TGNand, to a lesser extent to early endosomes, at steady state.
GGA proteins are known to interact with acidic cluster/dileucine (DXXLL) motifs in the cytoplamic tail of trans-membrane proteins (Nielsen et al. 2001; Puertollano et al.
Fig. 7 Mutations in both the proximal and distal DXXLL mo-tifs of LRP9 aVect its intracellu-lar distribution. a–i In COS7 cells transfected with GGA1-Myc together with wild-type LRP9-HA (WT, a) or LRP9-HA in which the distal DXXLL mo-tif (Distal, d) or proximal DXX-LL motif (Prox., g) were mutated, LRP9-HA co-localized extensively with GGA1-Myc (b, e, h) in the juxtanuclear Golgi region (arrowheads, c, f, i) and on vesicular structures (inset, c, f, i). j–l In cells transfected with GGA1-Myc and LRP9-HA in which the two DXXLL motifs (double, j) were mutated, LRP9 was mainly present on the cell surface (arrows, j) and in peripheral punctate structures (inset, J) and did not signiW-cantly co-localize with GGA1-Myc in the Golgi region (arrow-head, l) or on vesicular struc-tures (inset, l). Cells were Wxed, permeabilized and immuno-stained using anti-HA (a, d, g, j) or anti-Myc (b, e, h, k) antibod-ies. Stained cells were examined by confocal Xuorescence microscopy. Yellow colour in the merged images (c, f, i, l) indi-cates co-localization. Insets show £2 magniWed views of cytoplasmic regions. Scale bar 10 �m.
123

324 Histochem Cell Biol (2008) 130:315–327
2001; Takatsu et al. 2001; Zhu et al. 2001; He et al. 2002;Misra et al. 2002). Interestingly, we noted that the cytoplas-mic tail of LRP9 contained two DXXLL sequences, one atthe extreme end of the cytoplasmic tail (distal motif relativeto the transmembrane domain, residues 708–712) andanother, thirteen amino acids upstream (proximal motifrelative to the transmembrane domain, residues 689–694).Using immunoprecipitation assays, we demonstrated thatLRP9 interacts strongly with GGA2 and GGA3 and moreweakly with GGA1. Simultaneous mutations in bothDXXLL motifs completely inhibited the interaction ofLRP9 with the three GGA proteins. However, the substitu-tion of LL by AA in the proximal DXXLL motif did notabrogate the interaction of LRP9 with the three GGA pro-teins. In addition, the LL ! AA mutation in the distalDXXLL motif did not alter the interaction of LRP9 with
GGA2 and GGA3, although it almost completely abrogatedthe interaction with GGA1. Therefore, both DXXLL motifsbound to GGA proteins but had diVerent aYnities forGGA1. Indeed, the distal motif bound the three GGA pro-teins (albeit more weakly to GGA1) whereas the proximalDXXLL motif bound mainly GGA2 and GGA3. SigniWcantdiVerences in the aYnity of GGA proteins for acidic dileu-cine motifs have been previously reported. Although bothCD-M6PR and CI-M6PR contain similar DXXLL motifsnear the C-terminus of their cytoplasmic tails (Fig. 5b), CI-M6PR binds to GGA proteins with higher aYnity than theCD-M6PR (Doray et al. 2002). There is also a diVerence inthe relative aYnity of the DXXLL motif in the CD-M6PRcytoplasmic tail for the diVerent GGAs. Puertollano et al.(2001) showed that CD-MPR bound well to GGA1, butonly poorly to GGA3, and not at all to GGA2. In contrast,
Fig. 8 Redistribution of LRP9 double DXXLL mutant to the plasma membrane and periphe-ral EEA1-positive compart-ments. a–i In COS7 cells transfected with wild-type LRP9-HA (WT, a–c) or LRP9-HA in which the distal DXXLL motif (Distal, d–f) or proximal DXXLL motif (Prox., g–i) were mutated, LRP9-HA was not present in peripheral EEA1-labelled endosomes (inset, c, f, i). j–l In cells transfected with LRP9-HA in which the two DXXLL motifs were mutated (Double, j–l), LRP9-HA was mainly present on the cell sur-face (arrows, j, l) and in periph-eral punctate structures (arrowheads, j) that signiW-cantly co-localized with EEA1 (inset, l). Cells were Wxed, per-meabilized and immunostained using anti-HA (a, d, g, j) and anti-EEA1 (b, e, h, k) antibod-ies. Stained cells were examined by confocal Xuorescence microscopy. Yellow colour in the merged images indicates co-localization. Merged images (c, f, i, l) show £2 magniWed views of the boxes in a–l. Scale bar 10 �m.
123

Histochem Cell Biol (2008) 130:315–327 325
the cytoplasmic tail of CI-MPR bound to the VHS domainsof all three GGAs with about equal aYnity. Comparison ofthe C-terminal residues of these two M6PR (Fig. 5b)revealed a number of non-conservative diVerences in theacidic cluster/dileucine motifs and the Xanking residues.Mutation analysis revealed that several residues immedi-ately upstream and downstream from the DXXLL motif, aswell as the XX residues of this motif, signiWcantly impactthe speciWcity of the interactions of GGA proteins withthese motifs (Puertollano et al. 2001; Zhu et al. 2001;Doray et al. 2002; He et al. 2002; He et al. 2003). Align-ment of the proximal and distal acidic cluster dileucinemotifs of LRP9 with the amino acid sequences of the cyto-plasmic tail of other transmembrane proteins containing aDXXLL motif (Fig. 5b) showed diVerences in residues inposition +1 and +2 as well as in several residues upstreamand downstream from the proximal and distal DXXLLmotifs. The importance of these residues is now underinvestigation, and should provide new insights into selec-tivity towards speciWc interactions with GGA proteins andthe traYcking mechanisms of DXXLL-mediated sortingsignals.
LRP9 is the Wrst example of a transmembrane proteinwith an internal GGA-binding sequence in addition to theusual C-terminal motif. Using various protein sequencedata banks such as TrEMBL and Swiss-Prot, we wereunable to identify other proteins with two DXXLL motifs,suggesting that LRP9 might be the only transmembraneprotein with two functional DXXLL motifs. The VHSdomain of GGA proteins has been identiWed as the bindingsite of the DXXLL motifs of LRP9. This is in agreementwith previous reports deWning VHS as the speciWc GGAdomain that binds to the DXXLL motif (Bonifacino 2004;He et al. 2002; Nielsen et al. 2001; Puertollano et al. 2001;Takatsu et al. 2001). LRP9 can now be added to the longlist of proteins with DXXLL-type sorting signals that bindto the VHS domain of the GGAs including sortilin, LRP3,�-secretase and M6PR (Bonifacino 2004).
Previous studies have reported that interactions of GGAproteins with a DXXLL motif in the cytoplasmic tail of var-ious receptors mediate their anterograde or retrogradetransport between the TGN and endosomes (Huse et al.2000; Tikkanen et al. 2000; Tortorella et al. 2007). Thesestudies, together with our observation that LRP9 interactedand co-localized with GGA proteins, prompted us to inves-tigate whether GGA proteins were involved in the intracel-lular transport of LRP9. Mutations (LL ! AA) in theproximal or distal DXXLL motif did not aVect the intracel-lular distribution of LRP9. Therefore, proximal and distalDXXLL motifs alone seemed to be suYcient for the propertraYcking of LRP9 since they both had the capacity to bindGGA proteins. In addition, since we did not detect any sub-stantial diVerences between the single DXXLL mutants, we
concluded that these two motifs might play redundant rolesin the traYcking of LRP9. Mutations in both DXXLLmotifs that completely inhibited the interaction of LRP9with the three GGA proteins resulted in a redistribution ofthe receptor from the TGN to EEA1-labelled endosomesand the plasma membrane. Similar mutations in theDXXLL motifs in the cytoplasmic tails of other receptorssuch as MPR and mepapsin (BACE1) have also beenreported to cause a redistribution of these receptors fromthe TGN to endosomes and the plasma membrane (Huseet al. 2000; Tikkanen et al. 2000; He et al. 2005; Tortorellaet al. 2007), thus reinforcing our observations. Further-more, downregulation of GGA proteins or overexpressionof dominant negative GGA mutants cause an accumulationof MPR in endosomal compartments (Ghosh et al. 2003;Puertollano and Bonifacino 2004; He et al. 2005; Wahleet al. 2005). These receptor redistributions have beenexplained by a sorting defect at diVerent levels: (1) TGNsorting: GGA proteins normally retain DXXLL receptors inthe TGN and prevent premature exit from the Golgi appara-tus to early endosomes or from the Golgi apparatus via thesecretory pathway to the cell surface where they would thenbe rapidly internalized into early endosomes or (2) endo-somal sorting: GGA proteins are involved in early endo-some-to-TGN retrieval of receptors containing a DXXLLmotif. These explanations and our results provide supportfor a potential role for GGA adaptor proteins in sortingLRP9 between the TGN and endosomal compartments inboth anterograde and retrograde directions.
Our data thus suggested that LRP9 cycles between theTGN and early endosomes at steady state. LRP9 can alsoreach the cell surface since the double DXXLL mutantre-localized at the plasma membrane. However, since therewas no signiWcant labelling of wild-type LRP9 at theplasma membrane, we propose that either only a minorfraction of wild-type LRP9 is transported to the cell surfaceor its presence at the plasma membrane is very transientbecause the receptor is quickly internalized. Interestingly,Doray et al. (2007) have recently reported that a chimeraprotein containing the cytoplasmic tail of LRP9 undergoesrapid endocytosis. Furthermore, like our observations, theseauthors showed that a double DXXLL mutant (LL ! AA)accumulates at the cell surface, impairing LRP9 internaliza-tion. They reported that the proximal and distal dileucinemotifs of LRP9 bind �/�1 AP1 and �/�2 AP2 hemicom-plexes and that the intracellular distribution of an LRP9mutant that is unable to bind AP1 is not aVected whereasthe LRP9 mutant that is unable to bind to both AP2 andAP1 is redistributed to the plasma membrane (Doray et al.2007). They concluded that there is a correlation between�/�2 AP2 hemicomplex binding and LRP9 chimera recep-tor internalization mediated by dileucine-based sortingsignals. Since current evidence does not support the
123

326 Histochem Cell Biol (2008) 130:315–327
involvement of GGA proteins in the internalization steps,we suggest that the retention of the double DXXLL mutantat the cell surface we observed (Figs. 7, 8) might be due tothe loss of the interaction between LRP9 and AP2. TheDXXLL motifs of LRP9 thus appear to be required for sort-ing at two diVerent steps in LRP9 traYcking: (1) internali-zation from the plasma membrane via an interaction withAP2 and (2) transport between the endosomes and the TGNvia an interaction with the GGA proteins. Further studiesare now underway to characterize the exact traYckingitinerary and kinetics of LRP9 and to identify the precisemolecular machinery involved in the intracellular sorting ofLRP9.
Only a few other members of the LDLR family havebeen reported to bind GGA proteins. LRP3 and LRP12, thetwo other members of the new LRP9 subgroup of theLDLR superfamily, possess a single DXXLL motif in theircytoplasmic tail, suggesting that they have the capacity tointeract with GGA proteins. Indeed, LRP3 has already beenshown to interact with GGA proteins (Takatsu et al. 2001).However, the intracellular traYcking and function of LRP3and LRP12 remain uncharacterized. SorLA (sorting proteinrelated receptor; LR11) is another distant member of theLDLR family shown to mediate the uptake of ApoE-richlipoproteins in vitro (Taira et al. 2001), to interact withGGA1 and GGA2 (Jacobsen et al. 2002) and to shuttlebetween the Golgi and endosomes (Jacobsen et al. 2001;Andersen et al. 2005; OVe et al. 2006). Interestingly,SorLA/LR11 has been recently described as a sortingreceptor that regulates the intracellular transport and pro-cessing of the amyloid precursor protein (APP) (Andersenet al. 2005; Shah and Yu 2006; Schmidt et al. 2007) point-ing to an involvement in Alzheimer’s disease.
To date, the only ligand identiWed for LRP9 is apoE-�VLDL (Sugiyama et al. 2000). However, since LRP9 isnot mainly localized at the cell surface at steady state, it islikely to serve other functions inside the cell. DiVerent linesof evidence give hints towards a possible function of LRP9.First, the intracellular distribution of LRP9 at steady state.We observed that only a minor fraction of the receptor isexpressed on the cell surface, whereas the majority is foundin TGN and endosomes. Secondly, LRP9 cytoplasmicdomain contains signal sequence involved in adaptor pro-tein binding for endocytosis and Golgi-endosome sorting.We and others have recently demonstrated that LRP9 inter-acts with AP1, AP2 (Doray et al. 2007) and GGA proteins(this paper). Thirdly, many receptors containing a DXXLLmotif or a GGA-binding site, such as MPR (Tikkanen et al.2000; Puertollano et al. 2001; Tortorella et al. 2007), Sorti-lin (Nielsen et al. 2001) and SorLA (Andersen et al. 2005;Nielsen et al. 2001), are sorting receptors that traYcbetween the TGN, endosomes and the cell surface. It wouldappear from the above that LRP9 might be a candidate
sorting receptor, targeted for transport by ligands in thesynthetic pathway as well as on the surface membrane.Investigations are presently underway to test this hypothe-sis and identify intracellular cargos for LRP9. Furthermore,there is increasing evidence of a prominent role for certainmembers of the LDLR superfamily in signal transductionpathways and cell physiology regulation (Strickland et al.2002; May et al. 2007). This signalling occurs throughcooperation with other cell surface molecules that associatedirectly or indirectly with LDL receptor family membersand cytoplasmic adaptor molecules that interact to theircytoplasmic tails. The fact that signalling domains are pres-ent in the cytoplasmic tail of LRP9 suggests that it mightalso play a role in signal transduction. This hypothesis issupported by the work of Battle et al. (2003), who reportedthat various proteins involved in signal transduction inter-act with a juxtamembrane region of LRP12 that is con-served in LRP3 and LRP9. Further investigation is neededto clarify the biological functions of LRP9 and the othermembers of this LDLR subfamily.
In conclusion, our study is the Wrst to characterize theintracellular localization and traYcking of LRP9, anunusual member of the LDLR family. Its predominantlysubcellular localization in the TGN and endosomes,together with its capacity to bind GGA proteins via twoDXXLL motifs, suggest that LRP9 cycles between theTGN and endosomes through a GGA dependent-traYckingmechanism. These observations are important Wrst steps inunderstanding the role of this uncharacterized receptor andconstitute evidences pointing to a function of LRP9 in pro-tein sorting and transport.
Acknowledgments We would like to thank Juan Bonifacino, Rich-ard Leduc and T. Kitamura for DNA constructs and Marilyn Gist Far-quhar for the generous gifts of reagents. We would also like to thankEric Chevet, Catherine Denicourt and Richard Leduc for their con-structive comments on the manuscript. This work was supported bygrants from the Canadian Institutes for Health Research and a CanadaResearch Chair to C.L.
References
Andersen OM, Reiche J, Schmidt V, Gotthardt M, Spoelgen R, BehlkeJ, von Arnim CA, BreiderhoV T, Jansen P, Wu X (2005) Neuronalsorting protein-related receptor sorLA/LR11 regulates processingof the amyloid precursor protein. Proc Natl Acad Sci USA102:13461–13466
Banting G, Ponnambalam S (1997) TGN38 and its orthologues: rolesin post-TGN vesicle formation and maintenance of TGN mor-phology. Biochim Biophys Acta 1355:209–217
Battle MA, Maher VM, McCormick JJ (2003) ST7 is a novel low-den-sity lipoprotein receptor-related protein (LRP) with a cytoplasmictail that interacts with proteins related to signal transduction path-ways. Biochemistry 42:7270–7282
Bonifacino JS (2004) The GGA proteins: adaptors on the move. NatRev Mol Cell Biol 5:23–32
123

Histochem Cell Biol (2008) 130:315–327 327
Bork P, Beckmann G (1993) The CUB domain. A widespread modulein developmentally regulated proteins. J Mol Biol 231:539–545
Chardin P, McCormick F (1999) Brefeldin A: the advantage of beinguncompetitive. Cell 97:153–155
Christensen EI, Birn H (2002) Megalin and cubilin: multifunctionalendocytic receptors. Nat Rev Mol Cell Biol 3:256–266
Doray B, Bruns K, Ghosh P, Kornfeld S (2002) Interaction of the cat-ion-dependent mannose 6-phosphate receptor with GGA proteins.J Biol Chem 277:18477–18482
Doray B, Lee I, Knisely J, Bu G, Kornfeld S (2007) The gamma/sigma1 and alpha/sigma2 hemicomplexes of clathrin adaptorsAP-1 and AP-2 harbor the dileucine recognition site. Mol BiolCell 18:1887–1896
Ghosh P, Kornfeld J (2004) The GGA proteins: key players in proteinsorting at the trans-Golgi network. Eur J Cell Biol 83:257–262
Ghosh P, GriYth J, Geuze H, Kornfeld S (2003) Mammalian GGAs acttogether to sort mannose-6-phosphate receptors. J Cell Biol163:755–766
He X, Chang WP, Koelsch G, Tang J (2002) Memapsin 2 (beta-secre-tase) cytosolic domain binds to the VHS domains of GGA1 andGGA2: implications on the endocytosis mechanism of memapsin2. FEBS Lett 524:183–187
He X, Li F, Chang WP, Tang J (2005) GGA proteins mediate the recy-cling pathway of memapsin 2 (BACE). J Biol Chem 280:11696–11703
He X, Zhu G, Koelsch G, Rodgers KK, Zhang XC, Tang J (2003) Bio-chemical and structural characterization of the interaction of me-mapsin 2 (beta-secretase) cytosolic domain with the VHS domainof GGA proteins. Biochemistry 42:12174–12180
Herz J (2001) The LDL receptor gene family: (un) expected signaltransducers in the brain. Neuron 29:571–581
Hirst J, Seaman MN, Buschow SI, Robinson MS (2007) The role ofcargo proteins in GGA recruitment. TraYc 8:594–604
Huse JT, Pijak DS, Leslie GJ, Lee VM, Doms RW (2000) Maturationand endosomal targeting of beta-site amyloid precursor protein-cleaving enzyme. The Alzheimer’s disease beta-secretase. J BiolChem 275:33729–33737
Hussain MM, Strickland DK, Bakillah A (1999) The mammalian low-density lipoprotein receptor family. Annu Rev Nutr 19:141–172
Jacobsen L, Madsen P, Jacobsen C, Nielsen MS, Gliemann J, PetersenCM (2001) Activation and functional characterization of the mo-saic receptor SorLA/LR11. J Biol Chem 276:22788–22796
Jacobsen L, Madsen P, Nielsen MS, Geraerts WP, Gliemann J, SmitAB, Petersen CM (2002) The sorLA cytoplasmic domain inter-acts with GGA1 and -2 and deWnes minimum requirements forGGA binding. FEBS Lett 511:155–158
Kikuchi A, Yamamoto H, Kishida S (2007) Multiplicity of the interac-tions of Wnt proteins and their receptors. Cell Signal 19:659–671
Linstedt AD, Mehta A, Suhan J, Reggio H, Hauri HP (1997) Sequenceand overexpression of GPP130/GIMPc: evidence for saturablepH-sensitive targeting of a type II early Golgi membrane protein.Mol Biol Cell 8:1073–1087
May P, Woldt E, Matz RL, Boucher P (2007) The LDL receptor-relat-ed protein (LRP) family: an old family of proteins with new phys-iological functions. Ann Med 39:219–228
Misra S, Puertollano R, Kato Y, Bonifacino JS, Hurley JH (2002)Structural basis for acidic-cluster-dileucine sorting-signal recog-nition by VHS domains. Nature 415:933–937
Nakamura N, Rabouille C, Watson R, Nilsson T, Hui N, SlusarewiczP, Kreis TE, Warren G (1995) Characterization of a cis-Golgimatrix protein, GM130. J Cell Biol 131:1715–1726
Nielsen MS, Madsen P, Christensen EI, Anders Nykjær Gliemann J,Kasper D, Pohlmann R, Petersen CM (2001) The sortilin cyto-plasmic tail conveys Golgi–endosome transport and binds theVHS domain of the GGA2 sorting protein. EMBO J 20:2180–2190
OVe K, Dodson SE, Shoemaker JT, Fritz JJ, Gearing M, Levey AI, LahJJ (2006) The lipoprotein receptor LR11 regulates amyloid betaproduction and amyloid precursor protein traYc in endosomalcompartments. J Neurosci 26:1596–1603
Puertollano R, Bonifacino JS (2004) Interactions of GGA3 with theubiquitin sorting machinery. Nat Cell Biol 6:244–251
Puertollano R, Aguilar RC, Gorshkova I, Crouch RJ, Bonifacino JS.(2001) Sorting of mannose 6-phosphate receptors mediated by theGGAs. Science 292:1712–1716
Puri S, Bachert C, Fimmel CJ, Linstedt AD (2002) Cycling of earlyGolgi proteins via the cell surface and endosomes upon lumenalpH disruption. TraYc 3:641–653
Schaub BE, Berger B, Berger EG, Rohrer J (2006) Transition of galac-tosyltransferase 1 from trans-Golgi cisterna to the trans-Golginetwork is signal mediated. Mol Biol Cell 17:5153–5162
Schmidt V, Sporbert A, Rohe M, Reimer T, Rehm A, Andersen OM,Willnow TE. (2007) SorLA/LR11 regulates processing of amy-loid precursor protein via interaction with adaptors GGA andPACS-1. J Biol Chem 282:32956–32964
Shah S, Yu G (2006). sorLA: sorting out APP. Mol Interv 6(74–76):58Stenmark H, Parton RG, Steele-Mortimer O, Lutcke A, Gruenberg J,
Zerial M (1994) Inhibition of rab5 GTPase activity stimulatesmembrane fusion in endocytosis. Embo J 13:1287–1296
Strickland DK, Gonias SL, Argraves WS. (2002) Diverse roles for theLDL receptor family. Trends Endocrinol Metab 13:66–74
Sugiyama T, Kumagai H, Morikawa Y, Wada Y, Sugiyama A, YasudaK, Yokoi N, Tamura S, Kojima T, Nosaka T et al (2000) A novellow-density lipoprotein receptor-related protein mediating cellu-lar uptake of apolipoprotein E-enriched beta-VLDL in vitro. Bio-chemistry 39:15817–15825
Taira K, Bujo H, Hirayama S, Yamazaki H, Kanaki T, Takahashi K, IshiiI, Miida T, Schneider WJ, Saito Y (2001) LR11, a mosaic LDLreceptor family member, mediates the uptake of ApoE-rich lipopro-teins in vitro. Arterioscler Thromb Vasc Biol 21:1501–1506
Takatsu H, Katoh Y, Shiba Y, Nakayama K. (2001) Golgi-localizing,gamma-adaptin ear homology domain, ADP-ribosylation factor-binding (GGA) proteins interact with acidic dileucine sequenceswithin the cytoplasmic domains of sorting receptors through theirVps27p/Hrs/STAM (VHS) domains. J Biol Chem 276:28541–28545
Tikkanen R, Obermuller S, Denzer K, Pungitore R, Geuze HJ, von Fig-ura K, Honing S (2000) The dileucine motif within the tail ofMPR46 is required for sorting of the receptor in endosomes.TraYc 1:631–640
Tortorella LL, Schapiro FB, MaxWeld FR. (2007) Role of an acidiccluster/dileucine motif in cation-independent mannose 6-phos-phate receptor traYc. TraYc 8:402–413
Wahle T, Prager K, RaZer N, Haass C, Famulok M, Walter J (2005)GGA proteins regulate retrograde transport of BACE1 from endo-somes to the trans-Golgi network. Mol Cell Neurosci 29:453–461
Watson RT, Pessin JE. (2000) Functional cooperation of two indepen-dent targeting domains in syntaxin 6 is required for its eYcientlocalization in the trans-golgi network of 3T3L1 adipocytes.J Biol Chem 275:1261–1268
Willnow TE (1999) The low-density lipoprotein receptor gene family:multiple roles in lipid metabolism. J Mol Med 77:306–315
Xu Y, Martin S, James DE, Hong W (2002) GS15 forms a SNAREcomplex with syntaxin 5, GS28, and Ykt6 and is implicated intraYc in the early cisternae of the Golgi apparatus. Mol Biol Cell13:3493–3507
Yamamoto TDC, Brown MS, Schneider WJ, Casey ML, Goldstein JL,Russell DW (1984) The human LDL receptor: a cysteine-rich pro-tein with multiple Alu sequences in its mRNA. Cell 39:27–38
Zhu Y, Doray B, Poussu A, Lehto VP, Kornfeld S. (2001) Bindingof GGA2 to the lysosomal enzyme sorting motif of the mannose6-phosphate receptor. Science 292:1716–1718
123