interaction study of a novel macrobrachium rosenbergii effector caspase with b2 and capsid proteins...
TRANSCRIPT

lable at ScienceDirect
Fish & Shellfish Immunology 45 (2015) 534e542
Contents lists avai
Fish & Shellfish Immunology
journal homepage: www.elsevier .com/locate/ fs i
Full length article
Interaction study of a novel Macrobrachium rosenbergii effectorcaspase with B2 and capsid proteins of M. rosenbergii nodavirusreveals their roles in apoptosis
Supak Youngcharoen a, Saengchan Senapin b, c, **, Tareerat Lertwimol d,Siwaporn Longyant a, Paisarn Sithigorngul a, Timothy W. Flegel b, c,Parin Chaivisuthangkura a, *
a Department of Biology, Faculty of Science, Srinakharinwirot University, Bangkok 10110, Thailandb Center of Excellence for Shrimp Molecular Biology and Biotechnology, Mahidol University, Bangkok 10400, Thailandc National Center for Genetic Engineering and Biotechnology, National Science and Technology Development Agency, Pathumthani 12120, Thailandd National Metal and Materials Technology Center, National Science and Technology Development Agency, Pathumthani 12120, Thailand
a r t i c l e i n f o
Article history:Received 9 March 2015Received in revised form2 May 2015Accepted 5 May 2015Available online 14 May 2015
Keywords:Macrobrachium rosenbergiiCaspaseApoptosisB2Capsid proteinMrNVProtein interaction
* Corresponding author. Department of Biology, Finwirot University, Bangkok 10110, Thailand. Tel.: þ662 260 0128.** Corresponding author. Center of Excellence for SBiotechnology, Mahidol University, Bangkok 10400, Thfax: þ66 2 354 7344.
E-mail addresses: [email protected] ([email protected] (P. Chaivisuthangkura).
http://dx.doi.org/10.1016/j.fsi.2015.05.0091050-4648/© 2015 Elsevier Ltd. All rights reserved.
a b s t r a c t
Apoptosis is an essential immune response to protect invertebrates from virus infected cells. In shrimp,virus infection has been reported to induce apoptosis. Macrobrachium rosenbergii (Mr) was considered tobe a disease-resistant host when compared to penaeid shrimps. Caspase-3 was classified as an execu-tioner caspase which played a key role in virus-induced apoptosis. In this study, an effector caspase geneof M. rosenbergii (Mrcasp) was cloned and characterized. The open reading frame (ORF) of Mrcasp was957 nucleotide encoding 318 amino acid with a deduced molecular mass of 35.87 kDa. RT-PCR analysisshowed the presence of Mrcasp in all examined tissues. The phylogenetic tree indicated that Mrcasp wasclosely related with caspase 3 of shrimp. The functions of the Mrcasp, B2 and capsid proteins of M.rosenbergii nodavirus (MrNV) were assayed in Sf-9 cells. The results showed thatMrcasp induce apoptoticmorphology cells; however, capsid protein of MrNV could inhibit apoptotic cells whereas B2 couldneither induce nor inhibit apoptotic cells by DAPI staining. The protein interaction between Mrcasp andviral MrNV structure revealed that Mrcasp did not bind to B2 or capsid protein whereas B2 and capsidproteins could bind directly to each other. This study reported a novel sequence of a full-length Mrcaspand its functional studies indicated that Mrcasp could induce apoptotic cells. Our data is the first reportdemonstrating the direct proteineprotein interaction between capsid protein and B2 protein of MrNV.
© 2015 Elsevier Ltd. All rights reserved.
1. Introduction
White tail disease (WTD) caused by Macrobrachium rosenbergiinodavirus (MrNV) and extra small virus (XSV) is a major cause of100% mortality in post-larvae (PL) but not in adult freshwaterprawn M. rosenbergii [1]. MrNV was first reported in the FrenchWest Indies [2] and thereafter in China [3], India [4,5], Taiwan and
aculty of Science, Srinakhar-2 649 5000x18511; fax: þ66
hrimp Molecular Biology andailand. Tel.: þ66 2 201 5889;
Senapin), [email protected],
Thailand [6]. MrNV is a small, icosahedral, non-enveloped virus.The genome is formed by two single stranded RNA (ssRNA) calledRNA1 and RNA2 [2]. MrNV RNA1 is the larger genome (3.2 kb) thatencodes RNA-dependent RNA polymerase (RdRp) and B2 protein.B2 protein is encoded in the sub-genomic RNA3 and its function is asuppressor in post-transcriptional gene silencing whereas theRNA2 (1.2 kb) encodes a capsid protein in size of 43 kDa [7,8]. Thereare several methods for detection of MrNV such as RT-PCR, multi-plex RT-PCR [6], TaqMan realtime RT-PCR, in situ hybridization, dotblot hybridization [9], double antibody sandwich enzyme-linkedimmunosorbant assay [10], monoclonal antibodies based assay[11], loop-mediated isothermal amplification [12,13], and highresolution melt duplex RT-PCR [14].
The crustaceans have an innate immune system to protectthemselves from bacterial, fungal and viral pathogens [15]. Theinnate immune system comprises cellular and humoral immune

S. Youngcharoen et al. / Fish & Shellfish Immunology 45 (2015) 534e542 535
responses. The cellular immune responses include apoptosis,encapsulation, phagocytosis and nodule formulation whereas hu-moral immune responses include the prophenoloxidase system,the clotting cascade, the antimicrobial peptides and antiviral pep-tides [16]. Among the cellular immune responses, apoptosis is alsoan important mechanism for removal of unwanted or dangerouscells such as virus-infected cells [17]. In shrimp, virus infection mayinduce apoptosis based on morphological and biochemical assayssuch as white spot syndrome virus (WSSV) or yellow head virus(YHV) infection can lead to an increased number of apoptotic cells[18e20]. The morphological change of apoptosis such as DNAfragmentation in hemocytes of WSSV-infected Penaeus monodonand Penaeus indicus was observed by using flow cytometry andagarose gel electrophoresis [21]. Thereafter, the majority of cells arebelieved to die via apoptosis pathway. In shrimp, apoptosis wassuggested to be the cause of mortality as a terminal result of viralinfection [22].
The major players of apoptosis which induced morphologicalchanges of cell are a group of caspases, a family of structurallyrelated cysteine proteases [23]. Caspases are divided into twogroups: initiator caspases and effector caspases [23,24]. Usually,caspases are inactive proenzymes or zymogen and become acti-vated under a process of activation during apoptosis [25,26]. Allcaspases can be activated by autoprocessing by other caspasefamily members to produce an enzymatic active form [23,27,28].Caspase-3 is an important terminal effector caspase in apoptosisand involved in virus-induced apoptosis [29,30]. There are severalreports of caspase 3 in shrimp such as banana prawn Penaeus(Fenneropenaeus) merguiensis [28], P. monodon [30] and whitelegshrimp Penaeus (Litopenaeus) vannamei [31]. Wongprasert et al.[20] discovered that caspase-3 activity increased in WSSV-infectedP. monodon.
M. rosenbergii was found to be the host that tolerant to WSSVinfection and able to operate the clearance of WSSV [32]. Moreover,adult M. rosenbergii was resistant to MrNV and XSV but the exactmechanism was still unknown [1]. Recently, Mrcasp3c was identi-fied from the M. rosenbergii transcriptome database using IlluminaSolexa Genome Analyzer technique and was found to be up-regulated after challenge with the infectious hypodermal and he-matopoietic necrosis virus (IHHNV). However, the phylogenetictree of Mrcasp3c revealed the highest similarity to caspases 3C ofHydra vulgaris and H. magnipapillata instead of closely related toshrimp caspases [33]. Recently, a caspase-3-like protein was iso-lated and characterized from a Chinese mitten crab Eriocheirsinensis and the results revealed that EsCaspase-3-like proteinshared the highest similarity to that of Mrcasp3c [34]. This dataindicated that caspase-3 isoformsmay exist in crustacean similar toother invertebrate such as H. vulgaris [35], coral Stylophora pistillata[36], and mussel Mytilus galloprovincialis [37].
The present study aims to isolate an effector caspase ofM. rosenbergii (Mrcasp) and characterize the protein interactionbetweenMrcasp, B2 and capsid proteins ofMrNV by using Sf-9 cells.
2. Materials and methods
2.1. Shrimp RNA preparation
Juvenile M. rosenbergii of approximately 1.5 g each were pur-chased from a local farm in Suphan Buri province, Thailand. TotalRNAs were extracted from various M. rosenbergii tissues (hemo-cytes, hepatopancreas, muscle, gills, heart, pleopods, stomach, andintestine) using the Trizol reagent (Invitrogen) following themanufacturer's manual. RNA concentration was measured byQuant-iTTM RNA as described by the manufacturer's instruction(Molecular Probes) with Fluoroskan Ascent (Labsystems).
2.2. Cloning of the full-length cDNA of Mrcasp
A partial sequence ofMrcaspwas initially obtained by PCR usingM. rosenbergii cDNA generated by SuperScript III First-Strand Syn-thesis kit (Invitrogen) and a pair of degenerate primers called CAP-F2N and CAP-R2N. Briefly, the degenerate primers were designedfrom conserved domains obtained from an alignment of knowncaspase-3 sequences of P. vannamei (ABW69658.1), Penaeus (Fen-neropenaeus) chinensis (ADG84879.1), P. merguiensis (AAX77407.1)and P. monodon (ABI34434.1) using ClustalW2 program. Sequencesof degenerate primers for M. rosenbergii caspase are shown inTable 1. A PCR reaction volume of 50 ml contained; 3 ml of cDNAtemplate, 4 ml of 10 mM dNTPs, 1 ml of 50 mM of each primer, 0.5 mlof Taq DNA polymerase (5 units/ml, Invitrogen), 5 ml of 10 � PCRbuffer, and 35.5 ml of distilled water. The PCR was performed for 1cycle at 94 �C for 5 min and 35 cycles of 94 �C for 1 min, 55 �C for1 min, 72 �C for 1 min and final extension at 72 �C for 10 min. ThePCR products were cloned into pGEM®T Easy Vector System(Promega) and then sequenced.
To obtain the full-length cDNA of Mrcasp, 30 and 50 rapidamplification of cDNA ends (RACE) were performed using a SmarterRACE cDNA amplification kit (Clontech) using the gill RNA as atemplate. Specific primers for 30- and 50-RACE and adapter primersfrom the kit were used (Table 1) in touchdown PCR conditions: 5cycles at 94 �C for 30 s, 72 �C for 1 min; 5 cycles at 94 �C for 30 s,70 �C for 30 s, 72 �C for 1 min; 30 cycles at 94 �C for 30 s, 68 �C for30 s, 72 �C for 1min and cooling at 4 �C. PCR products were purifiedby MinElute®Gel Extraction kit (QIAGEN) and cloned into pGEM®-TEasy Vector System for DNA sequencing. After sequence analysis,the full-length Mrcasp cDNA was amplified by PCR using Pfx DNApolymerase enzyme (Invitrogen) and specific primers Exp_CAS_F-BamHI and Exp_CAS_R-XhoI (Table 1).
2.3. Sequence analysis of the full-length Mrcasp gene
The full-length cDNA and deduced amino acid sequences ofMrcasp were analyzed with the BLAST program (http://www.ncbi.nlm.nih.gov/BLAST/) and the Expasy Translate Tool (http://web.expasy.org/translate/). The protein sequence motifs wereanalyzed with ScanProsite (http://prosite.expasy.org/scanprosite).Multiple sequence alignment was carried out using ClustalW(www.ebi.ac.uk) and phylogenetic trees were constructed by theNeighbor-joining (NJ) method using MEGA 6 software.
2.4. Detection of Mrcasp expression in different tissues ofM. rosenbergii
Total RNA from various tissues of M. rosenbergii were used tostudy expression of Mrcasp by RT-PCR using SuperScript™ III One-Step RT-PCR System (Invitrogen) and specific primers calledCas_ExpFandNcas_ExpR (Table 1), producing anexpectedproduct of444 bp.M. rosenbergii b-actin gene primers generating a partial geneproduct of 337 bp designed based on GenBank accession numberAY626840.1 was used as an internal control. The RT-PCR protocolconsisted50 �C for 30min, 95 �C for 5minand4 �C for 5min followedby 30 cycles at 94 �C for 30 s, 55 �C for 30 s and 72 �C for 30 s and finalextension at 72 �C for 10 min. The amplified RT-PCR products ofMrcasp and b-actinwere analyzed on a 1% agarose gel, stained withethidium bromide and observed under the ultraviolet light.
2.5. Construction of recombinant Mrcasp for polyclonal antibodyproduction
The open reading frame of Mrcasp obtained from PCR usingspecific primers Exp_CAS_F-BamHI and Exp_CAS_R-XhoI

Table 1List of primers used in this study. Underlines indicate restriction sites.
Gene Primer name Primer sequence (50 / 30) Usage
Mrcasp CAP-F2N 50 CCG CGA CCC WSY GCC GCC CAC GAC GCC GA 30 Partial sequence PCRCAP-R2N 50 CCY GGG TAG GAM GCC CAC AT 30 Partial sequence PCR50-Race CAP 50 AAA GCC CGG GTA GGT CGC CCA CAT 30 50-RACE30-RACE CAP New 50 GAG GTT CTC ACG TCG CAG CTC TGG GAG 30 30-RACEb-actin-F 50 CCC AGA GCA AGA GAG GTA 30 Expressionb-actin-R 50 GCG TAT CCT TCG TAG ATG GG 30 ExpressionCas_Exp F 50 ACA TCT TGA AGG CAG CGA GT 30 ExpressionNcas_Exp R 50 CTT CCT CGT CGA GAA CCT TG 30 ExpressionExp_CAS_F-BamHI 50 CGG GAT CCA TGG ATT CTG TGG ATA AGG CTG 30 ExpressionExp_CAS_R-XhoI 50 CCC TCG AGC TAT ACG AAC TTG ACT TTC CTC 30 ExpressionF-MRcaspase3-BamHI 50 CGG GGA TCC ACC ATG GAT TCT GTG GAT AAG GCT 30 Co-IPR-MRcaspase3-NotI 50 CGA GCG GCC GCC TAC GAA CTT GAC TTT CCT CAT 30 Co-IP
B2 FeB2MR-BamHI 50 CGG GGA TCC ACC ATG GCA CAG TGG ACG AAC GTC 30 Co-IPReB2MR-NotI 50 CGA GCG GCC GCC CCA CGT TAT GAG GTC GCT TAG 30 Co-IP
capsid F-MRcapsid-BamHI 50 CGG GGA TCC ACC ATG GCT AGA GGT AAA CAA AAT 30 Co-IPR-MRcapsid-NotI 50 CGA GCG GCC GCC ATT ATT GCC GAC GAT AGC TCT 30 Co-IP
S. Youngcharoen et al. / Fish & Shellfish Immunology 45 (2015) 534e542536
mentioned above was cloned into the pCR®Blunt vector (Invi-trogen). The recombinant plasmid was then excised with BamHIand XhoI to release a 970 bp Mrcasp insert. The digested DNAfragment was subcloned into a BamHI and XhoI restricted pGEX-6P-1 plasmid (Amershame Biosciences) to generate a recombinantplasmid (pGEX-6P-1_Mrcasp) with a glutathione S-transferase(GST) tag at the N terminus. The Escherichia coli strain BL21 trans-formed with pGEX-6P-1_Mrcasp was cultured in LB medium con-taining 100 mg/ml ampicillin until the OD600 reached 0.5e0.7 beforeadding 1 mM isopropyl-b-D-thiogalactopyranoside (IPTG) and in-cubation for 4 h. The bacterial pellet was harvested by centrifuga-tion at 4000 � g at room temperature for 20 min and resuspendedin a lysis buffer B, pH 8.0 [100 mM NaH2PO4, 10 mM TriseHCl, 8 MUrea, 1 mM phenylmethylsulphonylfluoride (PMSF)]. The lysatewas then analyzed by 15% sodium dodecyl sulfate polyacrylamidegel electrophoresis (SDS-PAGE).
Recombinant protein bands of GST-Mrcasp of ~62 kDa from theSDS-PAGE treated with 0.3 M KCl were excised and collected indialysis bags (Sigma). The GST-Mrcasp was eluted with a Transblotapparatus (BioRad) at 70 V for 6 h. The purified protein solutionwasdialyzed and concentrated using a vacuum concentrator. The pro-tein concentrationwas determined by Bradford assay [38] and usedas an antigen. Four ICR mice were intra-peritoneally injected with0.2 mg of the purified protein per mouse in a 1:1 ratio with com-plete Freund's adjuvant (Sigma). Mice were subsequently injectedthree more times with the purified GST-Mrcasp protein mixed withincomplete Freund's adjuvant (Sigma) at 2 week intervals. Oneweek after the forth injection, mouse antisera were collected. TheMrcasp polyclonal antibodywas adsorbedwith purified GSTmoietyprior to western blotting analysis.
2.6. Plasmid construction for co-immunoprecipitation assays
Shrimp Mrcasp sequence and two genes from MrNV encodingB2-like and capsid proteins were subjected for plasmid construc-tion used in co-immunoprecipitation assays. A recombinantplasmid pCR blunt-Mrcasp described above was used as a templateto amplify the shrimp gene while RNA extracted from MrNV-infected post larvae M. rosenbergii was employed for viral geneamplification. With appropriate primers listed in Table 1, PCR andRT-PCR reactions were conducted similar to protocols mentionedabove. Each amplicon was double digested with BamHI and NotIand individually cloned into pDHsp/V5-His (called V5) plasmidand/or pDHsp/FLAG-His (called FLAG) plasmid, expression vectorscontaining a Drosophila heat shock protein 70 promoter [39]. Re-combinant constructs namely Mrcasp/V5, B2/V5, B2/FLAG, and
capsid/FLAG were subsequently sequenced to confirm gene se-quences and correct reading frames.
2.7. Plasmid transfection and protein expression in Sf-9 cells
In this study, we employed an insect cell line Spodoptera frugi-perda Sf-9 to express recombinant proteins. Sf-9 insect cells weremaintained at 27 �C in Sf-900 II SFM medium (Invitrogen) sup-plemented with 10% heat-inactivated fetal bovine serum. The cellswere seeded in six-well plates (9 � 105 cells/well) and incubated at27 �C for 30 min for cell attachment. The transfection mixture wasprepared to consist of 2 mg of each tested plasmid construct andCellfectin®II reagent (Invitrogen) following the supplier's protocol.To induce the expression of inserted genes, the cells were heatshocked at 42 �C for 30 min and then returned to 27 �C and incu-bated for 4e6 h to monitor the expression of protein. The heatshock induced cells were then washed with 1 � PBS buffer, pH 7.4and lysed on ice with 150 ml of NP-40 lysis buffer (50 mM TriseHCl,pH 8.0, 150 mM NaCl, 1% Nonidet P-40, 1 � protease inhibitorcocktail (Sigma)). The lysates were centrifuged at 12,000 � g for10 min and 20 ml of the supernatant was collected for Western blotanalysis using anti-V5 or anti-FLAG antibody (Sigma). After incu-bation with appropriate secondary antibody, the tested proteinstagged with V5 or FLAG were detected using the chem-iluminescence reagent Plus (Perkin Elmer life Sciences) followed byexposure to X-ray film (Kodak Science Imaging Film).
2.8. Co-immunoprecipitation assays
To test protein interaction, co-immunoprecipitation assayswere performed. The remaining supernatant (~110 ml) from theabove step was incubated with 15 ml of anti-FLAG M2 affinity gel(Sigma) and rotated at 4 �C for overnight. The affinity gel beadswere washed with 150 ml of NP-40 lysis buffer for 5 times (10 mineach) with rotation at 4 �C to remove unbound proteins. Then5 � loading buffer (62.5 mM TriseHCl, pH 6.8, 10% (w/v) SDS, 0.2%(v/v) glycerol, 100 mM DTT, 0.1% (w/v) bromphenol blue) wasadded to the gel suspension. The protein complexes were sepa-rated by 12% SDS-PAGE and analyzed by Western blot describedabove.
2.9. Investigation of apoptotic response in Sf-9 cells by DAPIstaining
To observe apoptotic responses between Mrcasp and MrNVproteins, DAPI staining of Sf-9 cells co-transfected with tested

S. Youngcharoen et al. / Fish & Shellfish Immunology 45 (2015) 534e542 537
plasmid pairs was performed. Briefly, cells were seeded on roundglass coverslips in a 24-well plate (1 � 106 cells/well) and plasmidconstructs co-transfected together included empty/V5þempty/FLAG, B2/FLAG þ emptyV5, Mrcasp/V5þemptyFLAG, capsid/FLAG þ emptyV5, Mrcasp/V5þcapsid/FLAG, and Mrcasp/V5þB2/FLAG. After heat shock induction, the cells were gently washedtwice with 1 � PBS (pH 7.4) and then fixed with 4% para-formaldehyde for 15 min and permeabilized with 0.1% Triton X-100for 5 min at 4�C. The cells were then stained with 1 mg/ml of 406-diamidino-2-phenylindole (DAPI) solution according to a previousreport [20]. Cell morphology was visualized by using OlympusConfocal Laser Scanning Microscope (Model FV10i-Liv). Apoptoticcells from each treatment were observed from a total cell numberof 200 cells in triplicate and statistical difference was analyzedusing one-way ANOVA. Sf-9 cells treated with actinomycin D (BioBasic, 150 ng/well) as a potent inducer of apoptosis, were used aspositive controls.
Fig. 1. Nucleotide and deduced amino acid sequences of the full-length Macrobrachiuencoding 318 amino acids contains a conserved active site motif, QACRG (bold italic letters)small subunit domains of Mrcasp, respectively. Bold double underline letters and bold dashof Mrcasp and positions of degenerate primers, respectively. The gray box shows primers u
3. Results and discussion
3.1. Isolation and sequence analysis of a novel Mrcasp cDNA
The novel full-length cDNA sequence of Mrcasp was initiallyamplified from gill RNA of the freshwater prawn M. rosenbergiiusing a degenerate primer pair and finally obtained by 50 and 30-RACEmethods. It was shown thatMrcasp cDNA consisted of 957 bp-open reading frame (ORF) encoding a deduced protein of 318 aminoacids (Fig. 1). This sequence has been submitted to GenBankaccession number KF878972. The predicted molecular weight andthe pI were 35.87 kDa and 5.78, respectively. A putative caspaseactive site, QACRG pentapeptide, found in several shrimp caspasesequences [28,30,31] located at amino acid residues 190 to 194 inthe putative Mrcasp amino acid sequence. ScanProsite analysis toolrevealed that the predicted large subunit domain located at aminoacids from 65 to 196 and the small subunit domain located at
m rosenbergii effector caspase (Mrcasp). An open reading frame of 957 nucleotidesat residues 190-194. Solid-line box and dash-line box represent the predicted large andunderline letters represent primers used to confirm the full-length open reading framesed in analysis of tissue expression of Mrcasp.

Fig. 2. A molecular phylogenetic tree of Mrcasp. The protein caspase sequences were Lvcaspase-3, P. vannamei (ACA00158); LvCAP-3, P. vannamei (ABK88280); Pmercap-3, P.merguiensis (AAX77407); PmCasp-3, P. monodon (ABI34434); Mrcasp, M. rosenbergii (KF878972); Fccaspase, P. chinensis (ADG84879); Lvcaspase4, P. vannamei (AGL61584); Lvcas-pase3, P. vannamei (AGL61582); Pjcaspase, Penaeus (Marsupenaeus) japonicus (ABK62771); Mrcasp3c, M. rosenbergii (AET34920); Lvcaspase2, P. vannamei (AGL61581); Pm caspase, P.monodon (ABO38430); BtCASP-3, Bos taurus (NP_001071308); Rncaspase-3, Rattus norvegicus (NP_037054); Lvcaspase5, P. vannamei (AGL61583); Dlcaspase-3, Dicentrarchus labrax(ABC70997); Trcaspase3, Takifugu rubripes (AAM43816), Slcaspase 3, Spodoptera litura (AFJ04534), and Ggcaspase 3, Gallus gallus (NP990056) based on the Neighbor-joining methodusing MEGA 6.0 software. Percentage bootstrap values with 1000 replications are indicated at each branch.
S. Youngcharoen et al. / Fish & Shellfish Immunology 45 (2015) 534e542538
residues 222 to 318 (Fig. 1). It could then be inferred from theprediction that Mrcasp has a relatively short prodomain (64 aminoacid residues) that is a typical characteristic of effector caspase.BLAST search revealed that the newly identified deduced caspaseamino acid sequence of Mrcasp had significant homology to cas-pase 3 or effector caspases from Penaeus vannamei (GenBankACA00158 and ABK88280) (47%), Penaeus merguiensis (GenBankAAX77407) (42%), and P. monodon (GenBank ABI34434) (40%) aswell as to an initiator caspase from Penaeus chinensis (GenBankADG84879) (41%). Interestingly, Mrcasp showed low homology(25% identity) to a previously identified caspase 3 sequence ofM. rosenbergii called MrCasp3C (GenBank number AET34920).MrCasp3c is a 521-amino acid caspase proposed to be an effectorcaspase in which its gene expression and enzymatic activity wereup-regulated upon infectious hypodermal and hematopoietic ne-crosis virus (IHHNV) infection [33].
Fig. 3. Expression study of Mrcasp and b-actin mRNA of M. rosenbergii in different tiss4 ¼ stomach; 5 ¼ pleopods; 6 ¼ hepatopancreas; 7 ¼ muscle; 8 ¼ hemocytes.
3.2. Phylogenetic analysis of Mrcasp
The phylogenetic tree was constructed based on reported full-length caspase amino acid sequences from all shrimp and caspase3 from various organisms such as fish, chicken, mammals, and in-sects by a Neighbor-joining (NJ) method using MEGA 6 software. Asshown in Fig. 2, the tree based on the selected caspase sequenceswas divided into 2 major clades. Clade 1 includedMrcasp and othercaspase sequences from all of the shrimp except for Lvcaspase5 thatresided in clade 2 containing caspase 3 from selected vertebratesand insects. Supported Blast homology search, the newly isolatedMrcasp had relatively far relationship with a previously identifiedMrCasp3c [33] despite being in the same clade. In addition toprotein domain sequence analysis, enzymatic activity assay ofMrcasp should be further investigated to unveil specific functions ofthis protein.
ues by RT-PCR analysis. Lane M ¼ DNA marker; 1 ¼ intestines; 2 ¼ heart; 3 ¼ gills;
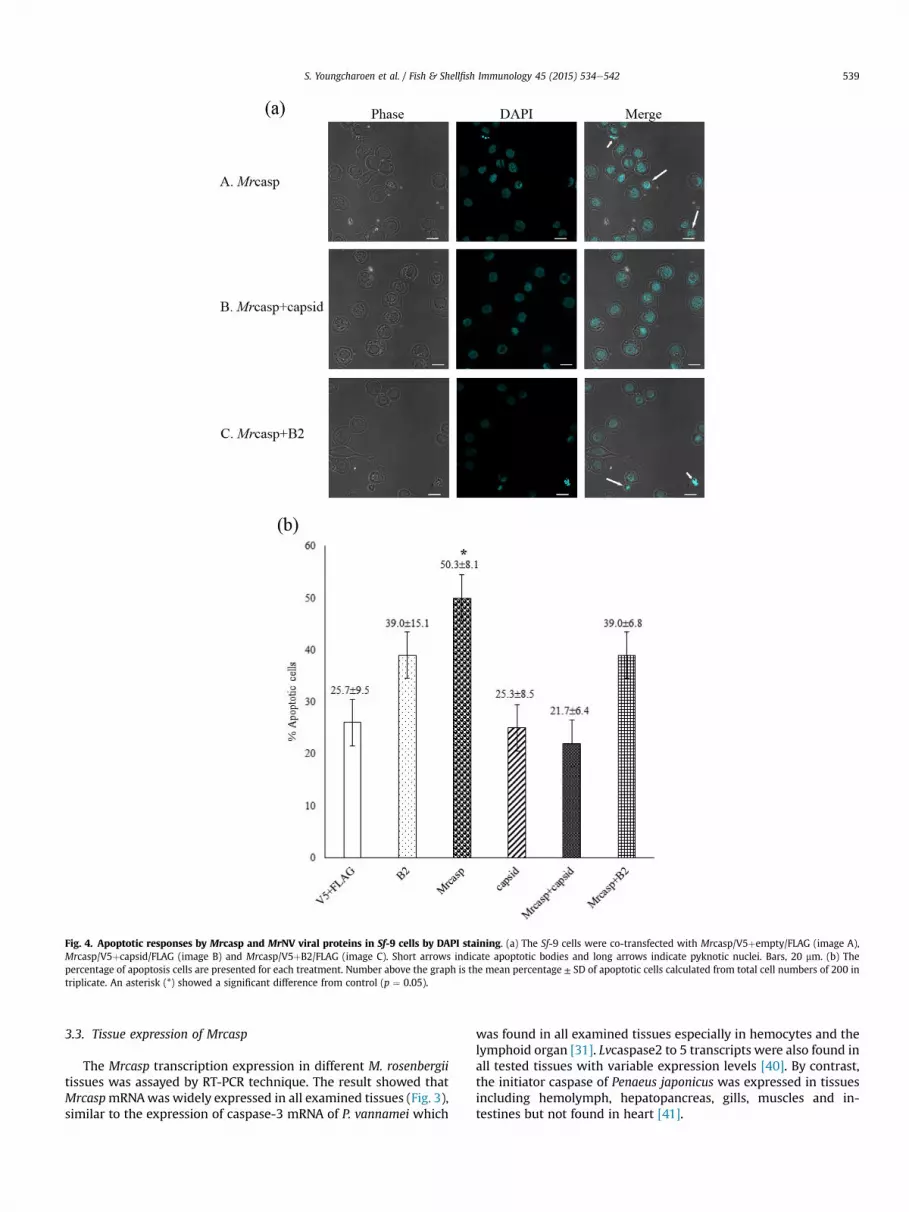
Fig. 4. Apoptotic responses by Mrcasp and MrNV viral proteins in Sf-9 cells by DAPI staining. (a) The Sf-9 cells were co-transfected with Mrcasp/V5þempty/FLAG (image A),Mrcasp/V5þcapsid/FLAG (image B) and Mrcasp/V5þB2/FLAG (image C). Short arrows indicate apoptotic bodies and long arrows indicate pyknotic nuclei. Bars, 20 mm. (b) Thepercentage of apoptosis cells are presented for each treatment. Number above the graph is the mean percentage ± SD of apoptotic cells calculated from total cell numbers of 200 intriplicate. An asterisk (*) showed a significant difference from control (p ¼ 0.05).
S. Youngcharoen et al. / Fish & Shellfish Immunology 45 (2015) 534e542 539
3.3. Tissue expression of Mrcasp
The Mrcasp transcription expression in different M. rosenbergiitissues was assayed by RT-PCR technique. The result showed thatMrcaspmRNAwas widely expressed in all examined tissues (Fig. 3),similar to the expression of caspase-3 mRNA of P. vannamei which
was found in all examined tissues especially in hemocytes and thelymphoid organ [31]. Lvcaspase2 to 5 transcripts were also found inall tested tissues with variable expression levels [40]. By contrast,the initiator caspase of Penaeus japonicus was expressed in tissuesincluding hemolymph, hepatopancreas, gills, muscles and in-testines but not found in heart [41].

Fig. 5. Co-immunoprecipitation of Mrcasp and MrNV proteins. Shrimp and viral genes were constructed in V5 and FLAG plasmids and co-transfected into Sf-9 cells as indicatedin the table. Cell lysates (input) were investigated for expression of the tested proteins (marked by arrows) using western blot assays probing with indicated antibodies. Theremaining lysate was then immunoprecipitated with anti-FLAG M2 affinity resins and immunoblotted with anti-Mrcasp or anti-V5 antibody (co-IP, bottom left). IgG indicated heavyand light chains of immobilized M2 antibody dissociated from the resins. * represents non-specific bands whereas # denotes degraded product of capsid/FLAG protein.
S. Youngcharoen et al. / Fish & Shellfish Immunology 45 (2015) 534e542540
3.4. Apoptotic responses of Mrcasp was suppressed by MrNV capsidprotein
Previous studies in the black tiger shrimp P. monodon, haveshown that two types of caspase could induce cell apoptosis in Sf-9cells observed by DAPI nuclear staining [39,42]. In addition, specificinhibition of caspase-induced apoptosis by white spot syndromevirus (WSSV) proteins was also revealed [39,42,43]. This studytherefore aimed to investigate apoptotic responses induced byMrcasp in the samemanner. Moreover, proteins fromM. rosenbergiinodavirus, a major pathogen of M. rosenbergii, were also subjectedfor investigation of potential anti-apoptotic effects. The MrNV viralproteins used in the study included B2-like protein (located in theRNA1 genome) and capsid protein (located in the RNA2 genome).
Plasmid constructs containing shrimp caspase and viral geneswere then tested for apoptotic responses in Sf-9 cells. The resultsrevealed that Sf-9 cells expressing Mrcasp showed apoptotic signsincluding cell membrane blebbing, pyknotic nuclei, and apoptoticbodies (Fig. 4a: image A) which were similar to the control cellsinduced by actinomycin D (data not shown). Percentage ofapoptotic cell induced by Mrcasp (50.3 ± 8.1%) was significantlyhigher than that induced by other plasmid constructs (p ¼ 0.027)(Fig. 4b). Specifically, cells expressing MrNV B2-like protein andcapsid protein yielded cell apoptosis rates of 39.0 ± 15.1% and25.3 ± 8.5%, respectively that were not significantly different fromempty plasmid control (25.7 ± 9.5%) (Fig. 4b). These resultsdemonstrated functional activity of Mrcasp in apoptosis inductionsimilar to previous findings of two caspase proteins fromP. monodon [39,42]. DAPI staining further revealed that over-expression of MrNV capsid protein could significantly reduce cellapoptosis induced by Mrcasp (21.7 ± 6.4%) more than that of co-expression between Mrcasp and MrNV B2-like protein(39.0 ± 6.8%) (Fig. 4a, b). The findings indicated that the capsidprotein but not B2-like protein of MrNV was a potential anti-apoptotic protein and functioned against Mrcasp. Mechanism by
which MrNV capsid protein inhibited the apoptosis induced byMrcasp required further study. However, structural proteins fromother viruses have been previously demonstrated to suppressapoptosis induced by caspase or non-caspase proteins from hostcells. For example, VP36A (or WSSV134) and VP38B (or WSSV449)from WSSV can inhibit apoptosis induced by P. monodon caspases[39,42,43]. In addition, rubella virus capsid protein was shown tobind to Bax and block Bax-induced apoptosis in mammalian celllines [44]. The gene encoding an RNA-dependent RNA polymerases(RdRp) of MrNV was not included in the present study. It is notknown if MrNV RdRp targets any host proteins or functionsinvolved in anti-apoptosis process. Moreover, it may beworthwhileto explore theMrNV protein functions interplaying withMrCasp3c,another type of caspase previously identified [33].
3.5. Protein interaction between Mrcasp and MrNV proteins by co-immunoprecipitation
Ability of MrNV capsid protein to inhibit Sf-9 cell apoptosisinduced by Mrcasp led to investigation of their protein interactionby co-immunoprecipitation (co-IP) assay.MrNV B2 protein that didnot inhibit Mrcasp activity was also included as a control. Proteinexpression of shrimp and viral gene products constructed in V5and/or FLAG plasmids transfected into Sf-9 cells was assayed bywestern blotting probing with anti-V5 and anti-FLAG antibodies,respectively. In addition, expression of Mrcasp/V5 protein was alsoinvestigated by polyclonal antibody against Mrcasp produced fromthis study. The results depicted in Fig. 5 revealed that Mrcasp/V5protein was detected at ~42 kDa using either anti-Mrcasp poly-clonal antibody (top left panel) or anti-V5 antibody (not shown).B2/V5 and B2/FLAG fusion proteins were detected at ~23 and21 kDa, respectively while capsid/FLAG protein with its possibledegraded product were visualized at ~49 and 25 kDa, respectively.
After verification of the protein expression, the protein com-plexes were then immunoprecipitated with anti-FLAG M2 affinity

S. Youngcharoen et al. / Fish & Shellfish Immunology 45 (2015) 534e542 541
beads followed by western blotting using appropriate antibody. InFig. 5 lanes 1e3, B2/FLAG, capsid/FLAG, and empty FLAG wasindividually co-expressed with Mrcasp/V5, respectively. The re-sults showed that no tested FLAG protein could pulldown theMrcasp/V5 protein evidenced by the absence of a 42 kDa Mrcaspprotein band using either anti-Mrcasp polyclonal antibody (bot-tom left panel) or anti-V5 antibody (not shown). The resultsindicated that Mrcasp did not directly interact with either MrNVcapsid or B2 proteins. Immunoreactive bands present in the blotwere heavy and light chains of IgG dissociated from the beads.When capsid/FLAG was co-transfected with B2/V5, the resultshowed that B2 protein could bind to capsid protein since the B2protein was observed using anti-V5 (Fig. 5, lane 4; bottom leftpanel). Whereas the control experiment using the empty FLAG didnot yield the B2 immunoreactive band (Fig. 5, lane 5; bottom leftpanel).
Our data is the first report showing the direct proteineproteininteraction between capsid protein and B2 protein of MrNV.Therefore, we suspected that this evidence may be associated withpackaging process of the viral genome similar to the capsid proteinof Flock house virus (FHV) involving with the packaging of the viralgenome, but the mechanism of FHV copackages between RNA1 andRNA2 into progeny virions is still unknown [45,46].
Since B2 protein is a suppressor in post-transcriptional genesilencing (PTGS) and capsid protein was shown to directly interactwith B2; therefore, we postulated that MrNV capsid protein mayalso involve in anti-PTGS. It has been shown that the capsid proteinof the Turnip crinkle virus (TCV) is a suppressor of the RNAsilencing system to overcome the host immune responses [47].
The anti-apoptosis proteins fromWSSV, for example: WSSV134,WSSV322 and AAP-1(WSSV449) were found to directly bind toPmCasp and inhibit caspase activity [39,43]. However, in the pre-sent study, the MrNV capsid protein, a putative anti-apoptoticprotein, did not directly bind to Mrcasp protein to inhibitapoptosis. This result indicated that MrNV capsid protein may bindto other molecules to suppress apoptosis activity ofMrcasp protein.It has been shown that two small molecules, IL-2 and evodiaminecould enhance apoptotic activtity of Marsupenaeus japonicus cas-pase (PjCaspase) protein to inhibit WSSV infection [48]. In our case,further study is needed to elucidate the molecule that may bind toMrNV capsid protein leading to apoptosis inhibition.
In conclusion, a novel sequence Mrcasp from M. rosenbergii wasisolated and characterized. TheMrcasp expressed in various shrimptissues examined. Phylogenetic tree demonstrated thatMrcasp hadclose relationship with caspases from other shrimps. The functionalstudy of Mrcasp through transfection in Sf-9 cells revealed thatMrcasp could induce apoptosis butMrNV capsid protein function asan anti-apoptotic protein. B2 protein and MrNV capsid proteindirectly interact with each other but the function is still unknown.
Acknowledgments
Financial supported from the Office of Commission on HigherEducation: Strategic Scholarships Fellowships Frontier ResearchNetworks (grant number 129/2550), Thailand to S.Y. and P.C. isacknowledged. The authors would like to thank Assoc. Prof. Dr. J-H.Leu, National Taiwan Ocean University for providing us theexpression vector pDHsp/V5-His and pDHsp/FLAG-His. We wouldlike to especially thank Center of Nanoimaging, Faculty of Science,Mahidol University, Thailand.
References
[1] M. Ravi, A. Nazeer Basha, G. Taju, R. Ram Kumar, A.S. Sahul Hameed, Clearanceof Macrobrachium rosenbergii nodavirus (MrNV) and extra small virus (XSV)
and immunological changes in experimentally injected Macrobrachium rose-nbergii, Fish. Shellfish Immunol. 28 (2010) 428e433.
[2] J.M. Arcier, F. Herman, D.V. Lightner, R.M. Redman, J. Mari, J.R. Bonami, A viraldisease associated with mortalities in hatchery-reared postlarvae of the giantfreshwater prawn Macrobrachium rosenbergii, Dis. Aquat. Org. 38 (1999)177e181.
[3] D. Qian, Z. Shi, S. Zhang, Z. Cao, W. Liu, L. Li, et al., Extra small virus-likeparticles (XSV) and nodavirus associated with whitish muscle disease in thegiant freshwater prawn, Macrobrachium rosenbergii, J. Fish. Dis. 26 (2003)521e527.
[4] A.S. Sahul Hameed, K. Yoganandhan, J. Sri Widada, J.R. Bonami, Studies on theoccurrence of Macrobrachium rosenbergii nodavirus and extra small virus-likeparticles associated with white tail disease ofM. rosenbergii in India by RT-PCRdetection, Aquaculture 238 (2004) 127e133.
[5] M.S. Shekhar, I.S. Azad, K.P. Jithendran, RT-PCR and sequence analysis ofMacrobrachium rosenbergii nodavirus: Indian isolate, Aquaculture 252 (2006)128e132.
[6] K. Yoganandhan, J. Sri Widada, J.R. Bonami, A.S. Sahul Hameed, Simulta-neous detection of Macrobrachium rosenbergii nodavirus and extra smallvirus by a single tube, one-step multiplex RT-PCR assay, J. Fish. Dis. 28(2005) 65e69.
[7] J.R. Bonami, Z. Shi, D. Qian, J. Sri Widada, White tail disease of the giantfreshwater prawn, Macrobrachium rosenbergii: separation of the associatedvirions and characterization of MrNV as a new type of nodavirus, J. Fish. Dis.28 (2005) 23e31.
[8] O. Hayakijkosol, L. Owens, B2 or not B2: RNA interference reduces Macro-brachium rosenbergii nodavirus replication in redclaw crayfish (Cherax quad-ricarinatus), Aquaculture 326e329 (2012) 40e45.
[9] J. Sri Widada, S. Durand, I. Cambournac, D. Qian, Z. Shi, E. Dejonghe, et al.,Genome-based detection methods of Macrobrachium rosenbergii nodavirus, apathogen of the giant freshwater prawn, Macrobrachium rosenbergii: dot-blot,in situ hybridization and RT-PCR, J. Fish. Dis. 26 (2003) 583e590.
[10] B. Romestand, J.R. Bonami, A sandwich enzyme linked immunosorbent assay(S-ELISA) for detection of MrNV in the giant freshwater prawn, Macro-brachium rosenbergii (de Man), J. Fish. Dis. 26 (2003) 71e75.
[11] P. Wangman, S. Senapin, P. Chaivisuthangkura, S. Longyant, S. Rukpratanporn,P. Sithigorngul, Production of monoclonal antibodies specific to Macro-brachium rosenbergii nodavirus using recombinant capsid protein, Dis. Aquat.Org. 98 (2012) 121e131.
[12] D. Pillai, J.R. Bonami, J. Sri Widada, Rapid detection of Macrobrachium rose-nbergii nodavirus (MrNV) and extra small virus (XSV), the pathogenic agentsof white tail disease of Macrobrachium rosenbergii (De Man), by loop-mediated isothermal amplification, J. Fish. Dis. 29 (2006) 275e283.
[13] T. Puthawibool, S. Senapin, T.W. Flegel, W. Kiatpathomchai, Rapid and sen-sitive detection of Macrobrachium rosenbergii nodavirus in giant freshwaterprawns by reverse transcription loop-mediated isothermal amplificationcombined with a lateral flow dipstick, Mol. Cell. Probes 24 (2010) 244e249.
[14] H. Zhang, J. Wang, J. Yuan, L. Li, J. Zhang, J.-R. Bonami, et al., Quantitativerelationship of two viruses (MrNV and XSV) in white-tail disease of Macro-brachium rosenbergii, Dis. Aquat. Org. 71 (2006) 11e17.
[15] M. Sarathi, V.P.I. Ahmed, C. Venkatesan, G. Balasubramanian, J. Prabavathy,A.S.S. Hameed, Comparative study on immune response of Fenneropenaeusindicus to Vibrio alginolyticus and white spot syndrome virus, Aquaculture 271(2007) 8e20.
[16] T.J. Little, D. Hultmark, A.F. Read, Invertebrate immunity and the limits ofmechanistic immunology, Nat. Immunol. 6 (2005) 651e654.
[17] W. Wang, X. Zhang, Comparison of antiviral efficiency of immune responses inshrimp, Fish. Shellfish Immunol. 25 (2008) 522e527.
[18] K. Khanobdee, C. Soowannayan, T.W. Flegel, S. Ubol, B. Withyachumnarnkul,Evidence for apoptosis correlated with mortality in the giant black tigershrimp Penaeus monodon infected with yellow head virus, Dis. Aquat. Org. 48(2002) 79e90.
[19] A.H. Sahtout, M.D. Hassan, M. Shariff, DNA fragmentation, an indicator ofapoptosis, in cultured black tiger shrimp Penaeus monodon infected withwhite spot syndrome virus (WSSV), Dis. Aquat. Org. 44 (2001) 155e159.
[20] K. Wongprasert, K. Khanobdee, S.S. Glunukarn, P. Meeratana,B. Withyachumnarnkul, Time-course and levels of apoptosis in various tissuesof black tiger shrimp Penaeus monodon infected with white-spot syndromevirus, Dis. Aquat. Org. 55 (2003) 3e10.
[21] A.S. Sahul Hameed, M. Sarathi, R. Sudhakaran, G. Balasubramanian, S. SyedMusthaq, Quantitative assessment of apoptotic hemocytes in white spotsyndrome virus (WSSV)-infected penaeid shrimp, Penaeus monodon andPenaeus indicus, by flow cytometric analysis, Aquaculture 256 (2006)111e120.
[22] T.W. Flegel, Active viral accommodation: a new concept for crustaceanresponse to viral pathogen, in: T.W. Flegel (Ed.), Advances in ShrimpBiotechnology, National Center for Genetic Engineering and Biotechnology,Bangkok, 1998, pp. 245e250.
[23] K. Wang, Caspases: a drug discovery perspective, Curr. Opin. drug Discov. Dev.2 (1999) 519e527.
[24] G. Salvesen, V. Dixit, Caspase activation: the induced proximity model, Proc.Natl. Acad. Sci. U. S. A. 96 (1999) 10964e10967.
[25] G.M. C, Caspases: the executioners of apoptosis, Biochem. J. 326 (1997) 1e16.[26] Y. Shi, Mechanisms of caspase activation and inhibition during apoptosis, Mol.
Cell. 9 (2002) 459e470.

S. Youngcharoen et al. / Fish & Shellfish Immunology 45 (2015) 534e542542
[27] A. Fire, S. Xu, M.K. Montgomery, S.A. Kostas, S.E. Driver, C.C. Mello, Potent andspecific genetic interference by double-stranded RNA in Caenorhabditis ele-gans, Nature 391 (1998) 806e811.
[28] A. Phongdara, W. Wanna, W. Chotigeat, Molecular cloning and expression ofcaspase from white shrimp Penaeus merguiensis, Aquaculture 252 (2006)114e120.
[29] T.-J. Fan, L.-H. Han, R.-S. Cong, J. Liang, Caspase family proteases and apoptosis,Acta Biochim. Biophys. Sin. 37 (2005) 719e727.
[30] K. Wongprasert, P. Sangsuriya, A. Phongdara, S. Senapin, Cloning and char-acterization of a caspase gene from black tiger shrimp (Penaeus monodon)-infected with white spot syndrome virus (WSSV), J. Biotechnol. 131 (2007)9e19.
[31] C.-C. Chang, M.-S. Yeh, H.-K. Lin, W. Cheng, The effect of Vibrio alginolyticusinfection on caspase-3 expression and activity in white shrimp Litopenaeusvannamei, Fish. Shellfish Immunol. 25 (2008) 672e678.
[32] A.S. Sahul Hameed, M.X. Charles, M. Anilkumar, Tolerance of Macrobrachiumrosenbergii to white spot syndrome virus, Aquaculture 183 (2000) 207e213.
[33] J. Arockiaraj, S. Easwvaran, P. Vanaraja, A. Singh, R.Y. Othman, S. Bhassu, Effectof infectious hypodermal and haematopoietic necrosis virus (IHHNV) infec-tion on caspase 3c expression and activity in freshwater prawn Macro-brachium rosenbergii, Fish. Shellfish Immunol. 32 (2012) 161e169.
[34] M.H. Wu, X.K. Jin, A.Q. Yu, Y.T. Zhu, D. Li, W.W. Li, et al., Caspase-mediatedapoptosis in crustaceans: cloning and functional characterization ofEsCaspase-3-like protein from Eriocheir sinensis, Fish. Shellfish Immunol. 41(2014) 625e632.
[35] M. Cikala, B. Wilm, E. Hobmayer, A. B€ottger, C.N. David, Identification ofcaspases and apoptosis in the simple metazoan Hydra, Curr. Biol. 9 (1999)959e962.
[36] H. Kvitt, H. Rosenfeld, K. Zandbank, D. Tchernov, Regulation of apoptoticpathways by Stylophora pistillata (Anthozoa, Pocilloporidae) to survive ther-mal stress and bleaching, PLoS ONE 6 (2011) e28665.
[37] A. Romero, N. Est�evez-Calvar, S. Dios, A. Figueras, B. Novoa, New insights intothe apoptotic process in mollusks: characterization of caspase genes inMytilus galloprovincialis, PLoS ONE 6 (2011) e17003.
[38] M. Bradford, A rapid and sensitive method for the quantification of microgramquantities of protein utilizing the principle of protein-dye binding, Anal.Biochem. 72 (1976) 248e254.
[39] J.H. Leu, H.C. Wang, G.H. Kou, C.F. Lo, Penaeus monodon caspase is targeted bya white spot syndrome virus anti-apoptosis protein, Dev. Comp. Immunol. 32(2008) 476e486.
[40] P.H. Wang, D.H. Wan, Y.G. Chen, S.P. Weng, X.Q. Yu, J.G. He, Characterization offour novel caspases from Litopenaeus vannamei (Lvcaspase2-5) and their rolein WSSV infection through dsRNA-mediated gene silencing, PLoS ONE 8(2013) e80418.
[41] L. Wang, B. Zhi, W. Wu, X. Zhang, Requirement for shrimp caspase inapoptosis against virus infection, Dev. Comp. Immunol. 32 (2008) 706e715.
[42] T. Lertwimol, P. Sangsuriya, K. Phiwsaiya, S. Senapin, A. Phongdara,C. Boonchird, et al., Two new anti-apoptotic proteins of white spot syndromevirus that bind to an effector caspase (PmCasp) of the giant tiger shrimpPenaeus (Penaeus) monodon, Fish. Shellfish Immunol. 38 (2014) 1e6.
[43] J.H. Leu, L.L. Chen, Y.R. Lin, G.H. Kou, C.F. Lo, Molecular mechanism of theinteractions between white spot syndrome virus anti-apoptosis protein AAP-1 (WSSV449) and shrimp effector caspase, Dev. Comp. Immunol. 34 (2010)1068e1074.
[44] C.S. Ilkow, I.S. Goping, T.C. Hobman, The rubella virus capsid is an anti-apoptotic protein that attenuates the pore-forming ability of Bax, PLoSPathog. 7 (2011) e1001291.
[45] P.A. Venter, N.K. Krishna, A. Schneemann, Capsid protein synthesis fromreplicating RNA directs specific packaging of the genome of a multipartite,positive-strand RNA Virus, J. Virol. 79 (2005) 6239e6248.
[46] N.K. Krishna, A. Schneemann, Formation of an RNA heterodimer upon heatingof nodavirus particles, J. Virol. 73 (1999) 1699e1703.
[47] F. Qu, T. Ren, T.J. Morris, The coat protein of turnip crinkle virus suppressesposttranscriptional gene silencing at an early initiation step, J. Virol. 77 (2003)511e522.
[48] B. Zhi, W. Tang, X. Zhang, Enhancement of shrimp antiviral immune responsethrough caspase-dependent apoptosis by small molecules, Mar. Biotechnol. 13(2011) 575e583.