integrin subunit expression associated with epithelial-mesenchymal interactions during murine tooth...
TRANSCRIPT

DEVELOPMENTAL DYNAMICS 205104-113 (1996)
Integrin Subunit Expression Associated With Epithelial-Mesenchymal Interactions During Murine Tooth Development KATRIINA SALMIVIRTA, DONALD GULLBERG, EMILIO HIRSCH, FIORELLA ALTRUDA, AND PETER EKBLOM Department of Animal Physiology, Uppsala University, S-75124 Uppsala, Sweden (K.S., D.G., P-E.); and Dipartimento di Genetica, Biologia e Chimica Medica, Universita di Torino, 10126 Torino, Italy (E.H., F.A.)
ABSTRACT The initial information for pat- terning of early tooth development resides in the epithelium. Later, this is shifted to the mesen- chyme. The process is governed by multiple epi- thelial-mesenchymal interactions. Integrins are cell surface receptors for extracellular matrix components. Expression of the p5 integrin subunit alternates between epithelium and mesenchyme during early tooth development (Yamada et al. 119941 Int. J. Dev. Biol. 38 553-556). By immuno- fluorescence and in situ hybridization we show here a remarkably similar oscillating expression pattern of the u1, inlegrin subunit. This subunit is known to associate with p5, and we therefore sug- gest that integrin is involved in epithelial- mesenchymal interactions during tooth develop- ment. We also demonstrate that the developing tooth epithelium expresses the (Ye, p1 and p4 sub- units. The laminin receptors && and &&4 may thus in part mediate the effect of basement mem- branes on tooth epithelial development. Interest- ingly, the enamel knot region expressed very little a 6 integrin subunit, whereas some expression was seen transiently in the condensing mesenchyme. During early tooth development, integrins pos- sessing the &6 subunit might also be involved in cell-cell interactions independently of laminins. 0 1996 Wiley-Liss, Inc.
Key words: Integrins, Laminin, Epithelial-mesen- chymal interactions, Tooth
INTRODUCTION Tooth development starts by local thickening of the
oral epithelium. The newly formed dental epithelium then forms a bud that grows into the underlying neural crest-derived mesenchyme. This, in turn, leads to the condensation of niesenchymal cells around the epithe- lial bud. The condensation of the dental mesenchyme is followed by rapid cell proliferation during cap and bell stages during which the typical morphology of tooth is established. Odontoblasts, which secrete the extracel- lular matrix of dentine, originate from the mesenchy- ma1 dental papilla cells which are in contact with base- ment membrane, whereas the inner dental epithelium
0 1996 WILEY-LISS. INC.
cells differentiate into ameloblasts which synthesize enamel (Thesleff et al., 1990). The initial information for patterning of tooth development resides in the ep- ithelium (Lumdsen et al., 1988). Later, this odontogen- ic potential shifts to the condensed mesenchyme, now designated dental mesenchyme (Mina and Kollar, 1987). A chain of reciprocal and sequential interactions between epithelial and mesenchymal tissues regulates both morphogenesis and cell differentiation in the de- veloping tooth (Slavkin, 1974; Thesleff and Hurmer- inta, 1981; Ruch, 1987).
Recently, a remarkable oscillating expression pat- lerri of lhe p5 irilegriri rriRNA in the developing loolh was reported. p5 integrin mRNA expression was found to oscillate between the mesenchyme and the epithe- lium during epithelial-mesenchymal interactions (Ya- mada et al., 1994). Since the integrin p5 subunit is known to form complexes with integrin a, subunit (Ya- mada et al., 1994) we have studied the expression of a, integrin in the developing mouse tooth. Integrin a, was found to be coexpressed with the p5 subunit, suggesting that the patterning during epithelial-mesenchymal in- teractions of the developing tooth in part is mediated by the oscillating expression of integrin a,p5.
Integrin a,p5 binds vitronectin and fibronectin (Pas- qualini et al., 19931, but many other extracellular ma- trix components are expressed during tooth develop- ment. Thus, other integrins may be involved in discrete stages of tooth development. Several previous observa- tions suggest that cell-matrix interactions might be in- volved in many aspects of tooth development. Base- ment membrane components are produced during early tooth morphogenesis (Lesot et al., 1981; Thesleff et al., 1981) and experimental transfilter studies have sug- gested that basement membrane components might be required for odontoblast differentiation (Thesleff and Hurmerinta, 1981). Receptors for basement membrane components may thus play a role in epithelial differ- entiation of the developing tooth, but information about such receptors during early tooth development is scanty. In some other embryonic tissues, integrins pos-
Received May 31, 1995; accepted August 25, 1995. Address reprint requestskorrespondence to Peter Ekblom, Depart-
ment of Animal Physiology, Uppsala University, Biomedical Center, Box 596, Husargatan 3, S-75124 Uppsala, Sweden.
INTEGRINS IN DEVELOPING TOOTH 105
sessing the subunit are required for epithelial mor- phogenesis (Sorokin et al., 1990; Kadoya et al., 1995). Integrins containing the a6 subunit are major receptors for laminins, the major glycoproteins of basement membranes (Sonnenberg et al., 1990). The a6 subunit can associate either with the p1 or p4 subunit to form the laminin receptors a6p1 and a6p4 (Sonnenberg et al., 1991; Lee et al., 1992; Giancotti et al., 1992; Tozeren et al., 1994; Spinardi et al., 1995). During terminal odon- toblast differentiation (at E20), integrin as and p4 sub- units have been shown to be expressed in the inner dental epithelium. Integrin was not detected in the developing tooth at this stage (Lesot et al., 1993). How- ever, the expression patterns of the subunits of the in- tegrins and 4 4 during early tooth development are not known. We have therefore examined the ex- pression of integrin (Y6, p1 and p4 subunits during mouse embryonic tooth development by immunohisto- chemistry and by in situ hybridization.
RESULTS Integrin a, Subunit in Developing Tooth
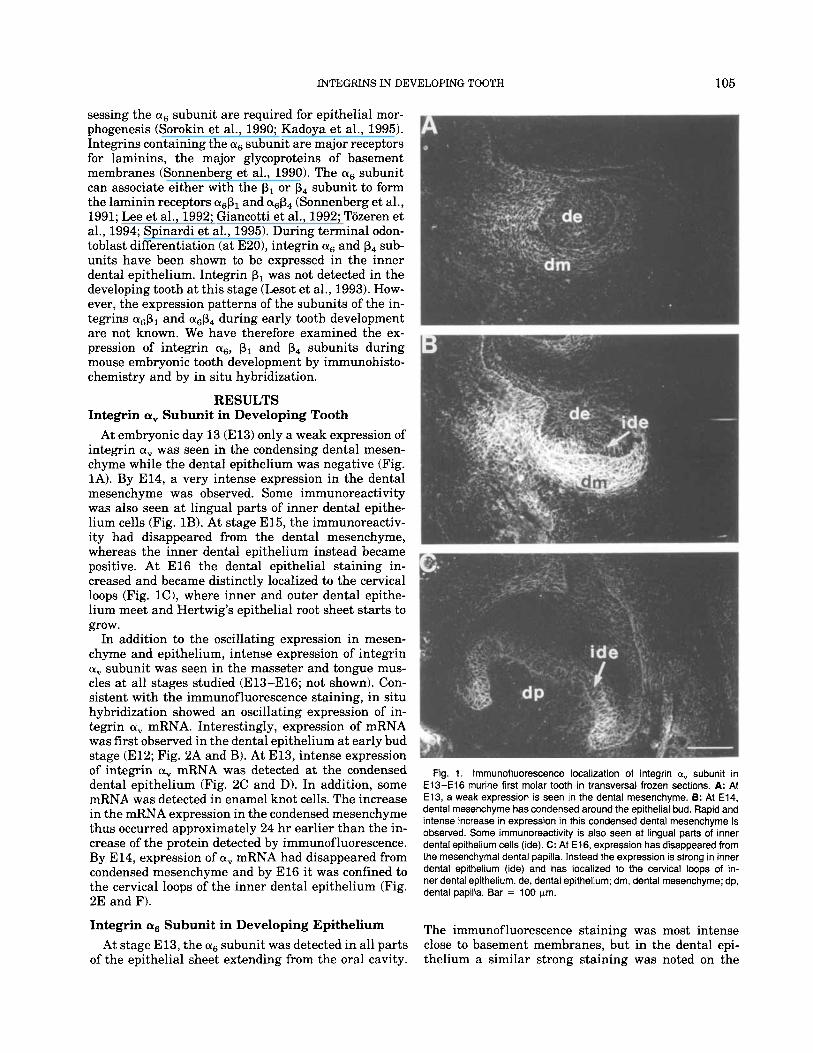
At embryonic day 13 (E13) only a weak expression of integrin a, was seen in the condensing dental mesen- chyme while the dental epithelium was negative (Fig. 1A). By E14, a very intense expression in the dental mesenchyme was observed. Some immunoreactivity was also seen at lingual parts of inner dental epithe- lium cells (Fig. 1B). At stage E15, the immunoreactiv- ity had disappeared from the dental mesenchyme, whereas the inner dental epithelium instead became positive. At E l 6 the dental epithelial staining in- creased and became distinctly localized to the cervical loops (Fig. lC), where inner and outer dental epithe- lium meet and Hertwig’s epithelial root sheet starts to grow.
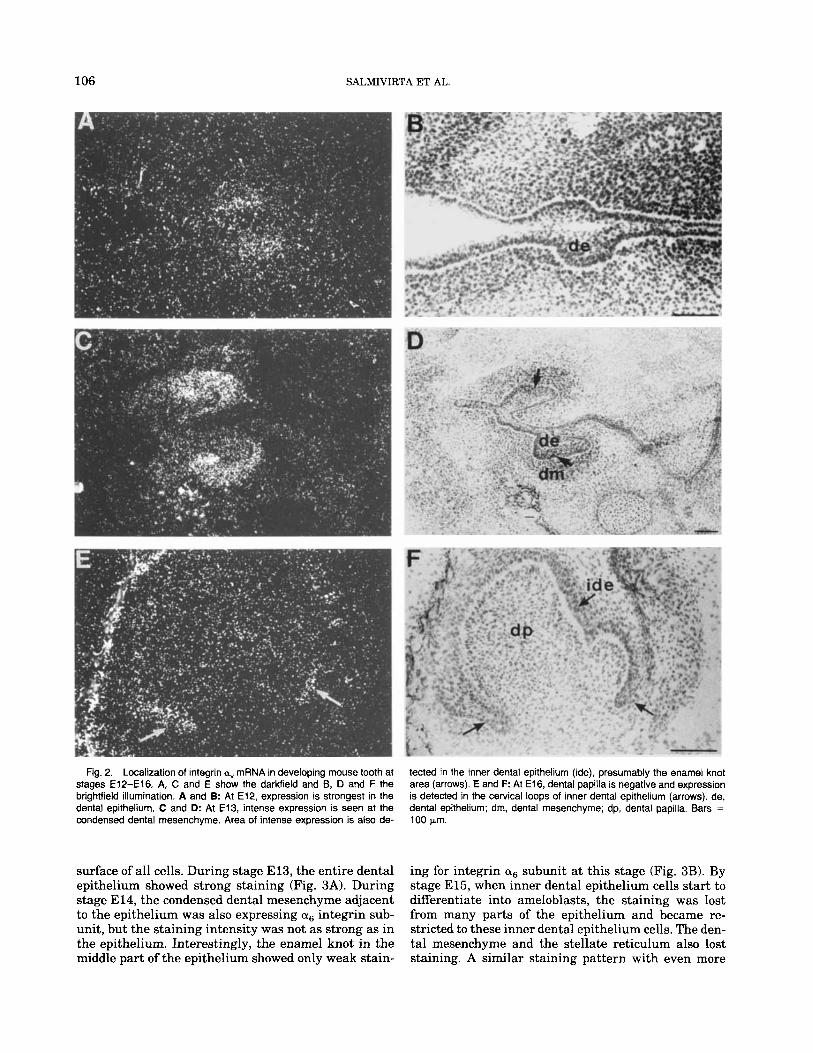
In addition to the oscillating expression in mesen- chyme and epithelium, intense expression of integrin a, subunit was seen in the masseter and tongue mus- cles a t all stages studied (E13-El6; not shown). Con- sistent with the immunofluorescence staining, in situ hybridization showed an oscillating expression of in- tegrin a, mRNA. Interestingly, expression of mRNA was first observed in the dental epithelium at early bud stage (E12; Fig. 2A and B). At E13, intense expression of integrin a, mRNA was detected at the condensed dental epithelium (Fig. 2C and D). In addition, some mRNA was detected in enamel knot cells. The increase in the mRNA expression in the condensed mesenchyme thus occurred approximately 24 hr earlier than the in- crease of the protein detected by immunof luorescence. By E14, expression of a, mRNA had disappeared from condensed mesenchyme and by El6 it was confined to the cervical loops of the inner dental epithelium (Fig. 2E and F).
Integrin as Subunit in Developing Epithelium At stage E13, the a6 subunit was detected in all parts
of the epithelial sheet extending from the oral cavity.
Fig. 1. lmmunofluorescence localization of integrin ay subunit in E13-El6 murine first molar tooth in transversal frozen sections. A: At E13, a weak expression is seen in the dental mesenchyme. 8: At E14, dental mesenchyme has condensed around the epithelial bud. Rapid and intense increase in expression in this condensed dental mesenchyme is observed. Some immunoreactivity is also seen at lingual parts of inner dental epithelium cells (ide). C: At E16, expression has disappeared from the mesenchymal dental papilla. Instead the expression is strong in inner dental epithelium (ide) and has localized to the cervical loops of in- ner dental epithelium. de, dental epithelium; dm, dental mesenchyme; dp, dental papilla. Bar = 100 pm.
The immunof luorescence staining was most intense close to basement membranes, but in the dental epi- thelium a similar strong staining was noted on the
106 SALMIVIRTA ET AL.
Fig. 2. Localization of integrin av mRNA in developing mouse tooth at stages E12-El6. A, C and E show the darkfield and B, D and F the brightfield illumination. A and B: At E12, expression is strongest in the dental epithelium. C and D: At E13, intense expression is seen at the condensed dental mesenchyme. Area of intense expression is also de-
tected in the inner dental epithelium (ide), presumably the enamel knot area (arrows). E and F: At E16, dental papilla is negative and expression is detected in the cervical loops of inner dental epithelium (arrows). de, dental epithelium; dm, dental mesenchyme; dp, dental papilla. Bars = 100 pm.
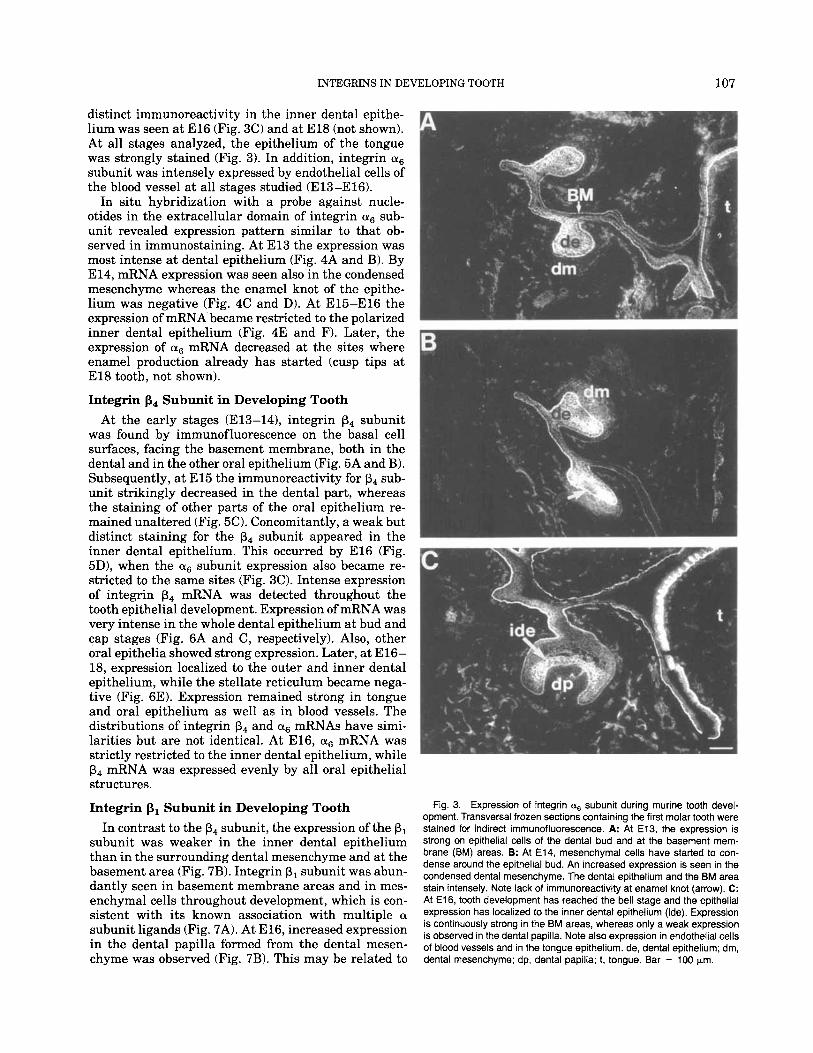
surface of all cells. During stage E13, the entire dental epithelium showed strong staining (Fig. 3A). During stage E 14, the condensed dental mesenchyme adjacent to the epithelium was also expressing as integrin sub- unit, but the staining intensity was not as strong as in the epithelium. Interestingly, the enamel knot in the middle part of the epithelium showed only weak stain-
ing for integrin cis subunit at this stage (Fig. 3B). By stage E15, when inner dental epithelium cells start to differentiate into ameloblasts, the staining was lost from many parts of the epithelium and became re- stricted to these inner dental epithelium cells. The den- tal mesenchyme and the stellate reticulum also lost staining. A similar staining pattern with even more
INTEGRINS IN DEVELOPING TOOTH 107
distinct immunoreactivity in the inner dental epithe- lium was seen at El6 (Fig. 3C) and at El8 (not shown). At all stages analyzed, the epithelium of the tongue was strongly stained (Fig. 3). In addition, integrin subunit was intensely expressed by endothelial cells of the blood vessel at all stages studied (E13-El6).
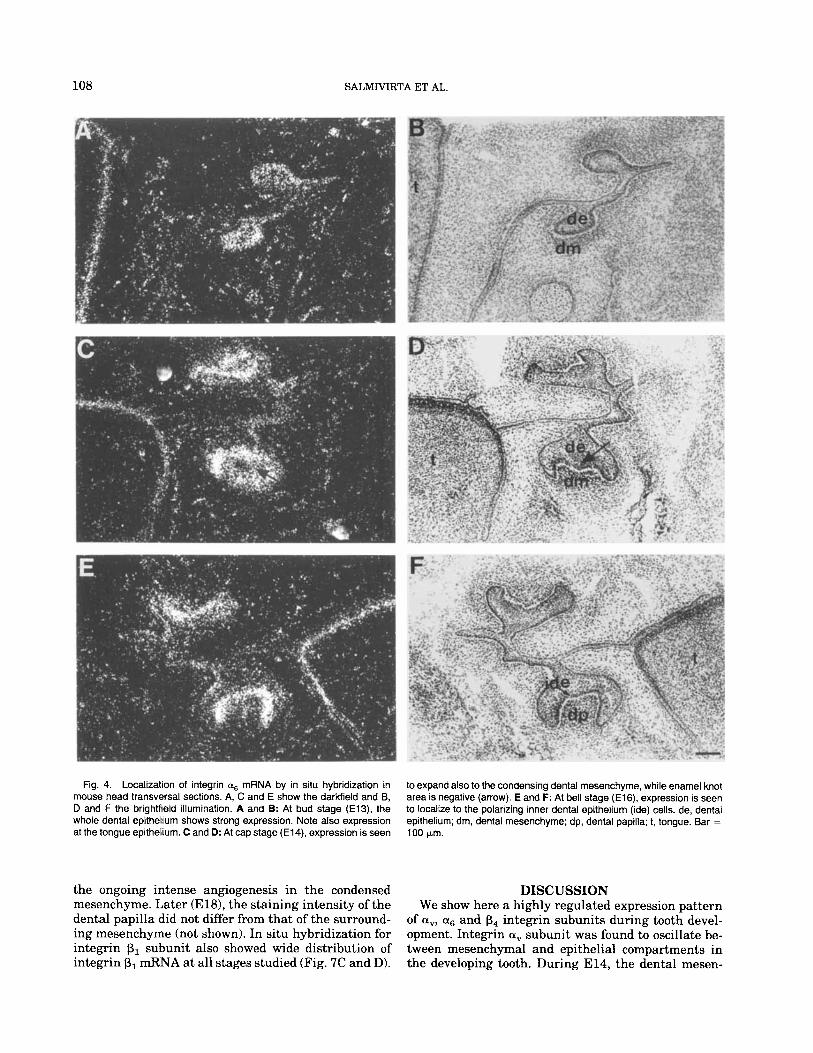
In situ hybridization with a probe against nucle- otides in the extracellular domain of integrin a6 sub- unit revealed expression pattern similar to that ob- served in immunostaining. At El3 the expression was most intense at dental epithelium (Fig. 4A and B). By E14, mRNA expression was seen also in the condensed mesenchyme whereas the enamel knot of the epithe- lium was negative (Fig. 4C and D). At E15-El6 the expression of mRNA became restricted to the polarized inner dental epithelium (Fig. 4E and F). Later, the expression of a6 mRNA decreased at the sites where enamel production already has started (cusp tips a t E l8 tooth, not shown).
Integrin p4 Subunit in Developing Tooth At the early stages (E13-14), integrin p4 subunit
was found by immunofluorescence on the basal cell surfaces, facing the basement membrane, both in the dental and in the other oral epithelium (Fig. 5A and B). Subsequently, at El5 the immunoreactivity for p4 sub- unit strikingly decreased in the dental part, whereas the staining of other parts of the oral epithelium re- mained unaltered (Fig. 5C). Concomitantly, a weak but distinct staining for the p4 subunit appeared in the inner dental epithelium. This occurred by El6 (Fig. 5D), when the subunit expression also became re- stricted to the same sites (Fig. 3C). Intense expression of integrin p4 mRNA was detected throughout the tooth epithelial development. Expression of mRNA was very intense in the whole dental epithelium at bud and cap stages (Fig. 6A and C, respectively). Also, other oral epithelia showed strong expression. Later, a t E16- 18, expression localized to the outer and inner dental epithelium, while the stellate reticulum became nega- tive (Fig. 6E). Expression remained strong in tongue and oral epithelium as well as in blood vessels. The distributions of integrin p4 and a6 mRNAs have simi- larities but are not identical. At E16, 016 mRNA was strictly restricted to the inner dental epithelium, while p4 mRNA was expressed evenly by all oral epithelial structures.
Integrin p1 Subunit in Developing Tooth In contrast to the p4 subunit, the expression of the p1
subunit was weaker in the inner dental epithelium than in the surrounding dental mesenchyme and at the basement area (Fig. 7B). Integrin subunit was abun- dantly seen in basement membrane areas and in mes- enchymal cells throughout development, which is con- sistent with its known association with multiple a subunit ligands (Fig. 7A). At E16, increased expression in the dental papilla formed from the dental mesen- chyme was observed (Fig. 7B). This may be related to
Fig. 3. Expression of integrin a6 subunit during murine tooth devel- opment. Transversal frozen sections containing the first molar tooth were stained for indirect immunofluorescence. A: At E13, the expression is strong on epithelial cells of the dental bud and at the basement mem- brane (BM) areas. B: At E14, mesenchymal cells have started to con- dense around the epithelial bud. An increased expression is seen in the condensed dental mesenchyme. The dental epithelium and the BM area stain intensely. Note lack of immunoreactivity at enamel knot (arrow). C: At E16, tooth development has reached the bell stage and the epithelial expression has localized to the inner dental epithelium (ide). Expression is continuously strong in the BM areas, whereas only a weak expression is observed in the dental papilla. Note also expression in endothelial cells of blood vessels and in the tongue epithelium. de, dental epithelium; dm, dental mesenchyme; dp, dental papilla; t, tongue. Bar = 100 pm.
108 SALMIVIRTA ET AL.
Fig. 4. Localization of integrin as mRNA by in situ hybridization in mouse head transversal sections. A, C and E show the darkfield and B, D and F the brightfield illumination. A and B: At bud stage (E13), the whole dental epithelium shows strong expression. Note also expression at the tongue epithelium. C and D: At cap stage (E14), expression is seen
to expand also to the condensing dental mesenchyme, while enamel knot area is negative (arrow). E and F: At bell stage (E16). expression is seen to localize to the polarizing inner dental epithelium (ide) cells. de, dental epithelium; dm, dental mesenchyme; dp, dental papilla; 1, tongue. Bar =
100 pm.
the ongoing intense angiogenesis in the condensed mesenchyme. Later (El@, the staining intensity of the dental papilla did not differ from that of the surround- ing mesenchyme (not shown). In situ hybridization for integrin p1 subunit also showed wide distribution of integrin p1 mRNA at all stages studied (Fig. 7C and D).
DISCUSSION We show here a highly regulated expression pattern
of a”, a6 and b4 integrin subunits during tooth devel- opment. Integrin a, subunit was found to oscillate be- tween mesenchymal and epithelial compartments in the developing tooth. During E14, the dental mesen-
INTEGRINS IN DEVELOPING TOOTH 109
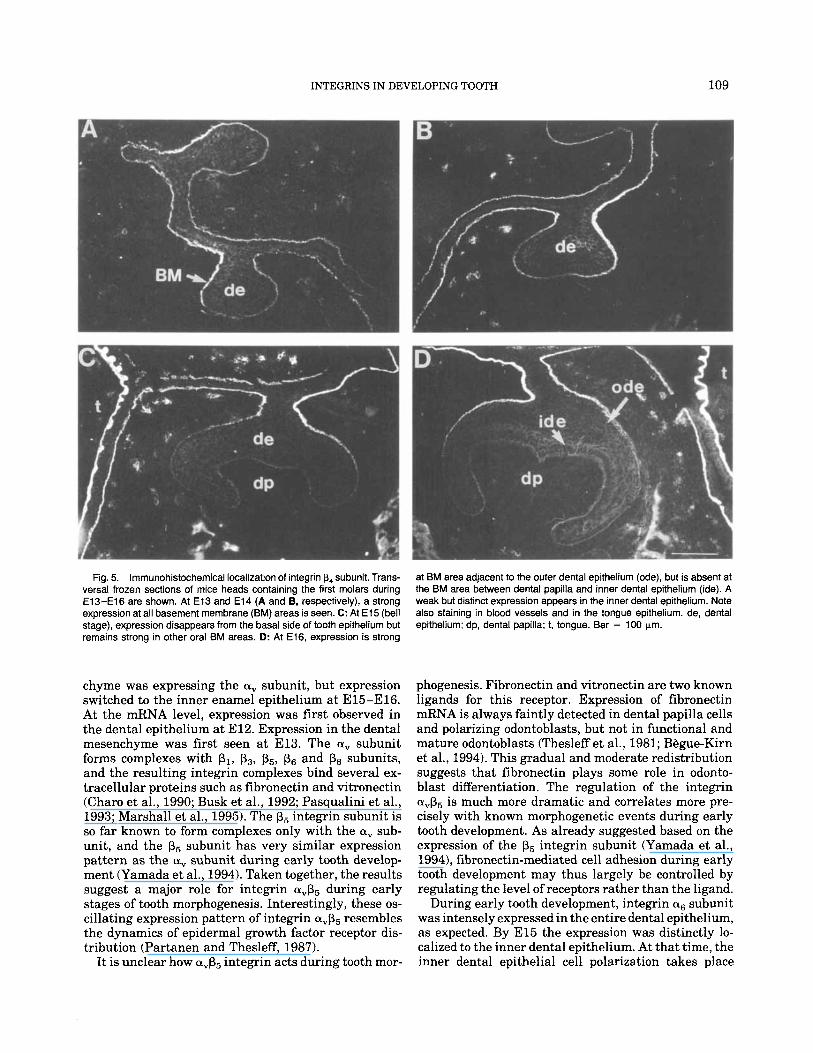
Fig. 5. lmmunohistochemical localization of integrin p4 subunit. Trans- versal frozen sections of mice heads containing the first molars during E13-El6 are shown. At E l 3 and E l 4 (A and B, respectively), a strong expression at all basement membrane (BM) areas is seen. C: At E l 5 (bell stage), expression disappears from the basal side of tooth epithelium but remains strong in other oral BM areas. D: At E16, expression is strong
chyme was expressing the a, subunit, but expression switched to the inner enamel epithelium at E15-El6. At the mRNA level, expression was first observed in the dental epithelium at E12. Expression in the dental mesenchyme was first seen at E13. The a, subunit forms complexes with PI, p3, p5, ps and ps subunits, and the resulting integrin complexes bind several ex- tracellular proteins such as fibronectin and vitronectin (Charo et al., 1990; Busk et al., 1992; Pasqualini et al., 1993; Marshall et al., 1995). The p5 integrin subunit is so far known to form complexes only with the a, sub- unit, and the p5 subunit has very similar expression pattern as the a, subunit during early tooth develop- ment (Yamada et al., 1994). Taken together, the results suggest a major role for integrin a,p5 during early stages of tooth morphogenesis. Interestingly, these os- cillating expression pattern of integrin avp5 resembles the dynamics of epidermal growth factor receptor dis- tribution (Partanen and Thesleff, 1987).
It is unclear how avp5 integrin acts during tooth mor-
at BM area adjacent to the outer dental epithelium (ode), but is absent at the BM area between dental papilla and inner dental epithelium (ide). A weak but distinct expression appears in the inner dental epithelium. Note also staining in blood vessels and in the tongue epithelium. de, dental epithelium; dp, dental papilla; t, tongue. Bar = 100 pm.
phogenesis. Fibronectin and vitronectin are two known ligands for this receptor. Expression of fibronectin mRNA is always faintly detected in dental papilla cells and polarizing odontoblasts, but not in functional and mature odontoblasts (Thesleff et al., 1981; Begue-Kirn et al., 1994). This gradual and moderate redistribution suggests that fibronectin plays some role in odonto- blast differentiation. The regulation of the integrin a,p5 is much more dramatic and correlates more pre- cisely with known morphogenetic events during early tooth development. As already suggested based on the expression of the p5 integrin subunit (Yamada et al., 1994), fibronectin-mediated cell adhesion during early tooth development may thus largely be controlled by regulating the level of receptors rather than the ligand.
During early tooth development, integrin a6 subunit was intensely expressed in the entire dental epithelium, as expected. By El5 the expression was distinctly lo- calized to the inner dental epithelium. At that time, the inner dental epithelial cell polarization takes place
110 SALMIVIRTA ET AL.
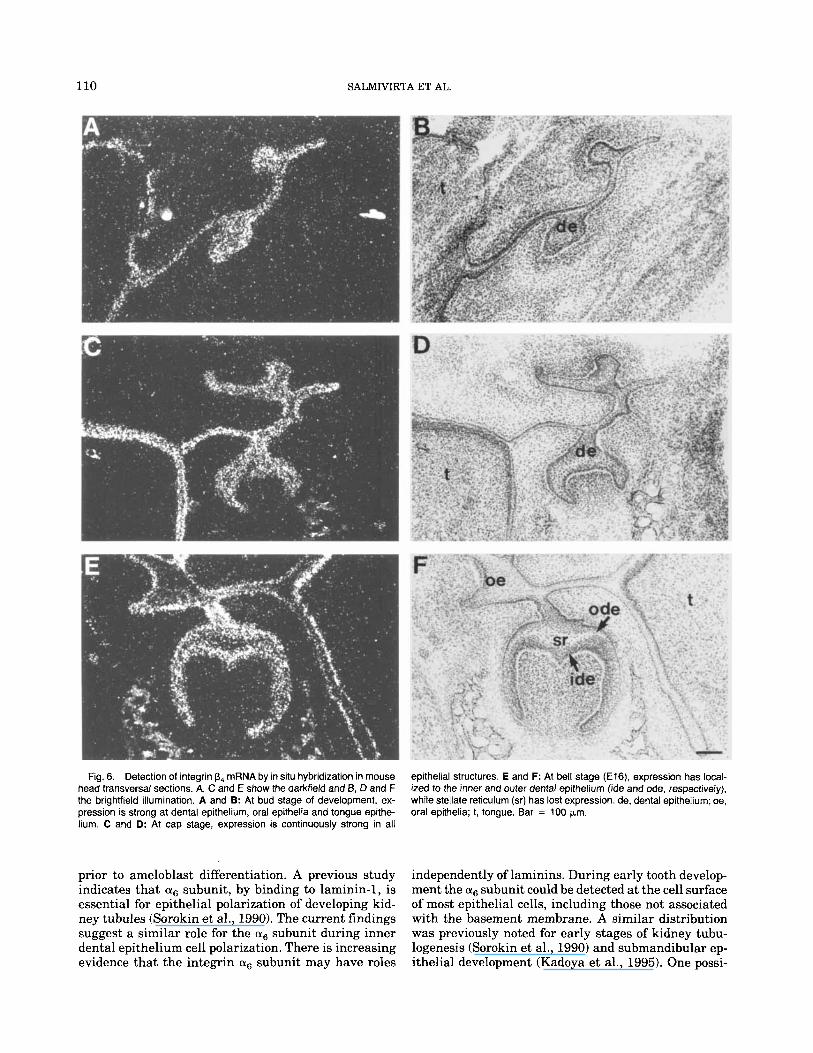
Fig. 6. Detection of integrin p4 mRNA by in situ hybridization in mouse head transversal sections. A, C and E show the darkfield and B, D and F the brightfield illumination. A and 9: At bud stage of development, ex- pression is strong at dental epithelium, oral epithelia and tongue epithe- hum. C and D: At cap stage, expression is continuously strong in all
epithelial structures. E and F: At bell stage (E16), expression has local- ized to the inner and outer dental epithelium (ide and ode, respectively), while stellate reticulum (sr) has lost expression. de, dental epithelium; oe, oral epithelia; 1, tongue. Bar = 100 pm.
prior to ameloblast differentiation. A previous study indicates that subunit, by binding to laminin-1, is essential for epithelial polarization of developing kid- ney tubules (Sorokin et al., 1990). The current findings suggest a similar role for the (Y6 subunit during inner dental epithelium cell polarization. There is increasing evidence that the integrin (Y6 subunit may have roles
independently of laminins. During early tooth develop- ment the a+, subunit could be detected at the cell surface of most epithelial cells, including those not associated with the basement membrane. A similar distribution was previously noted for early stages of kidney tubu- logenesis (Sorokin et al., 1990) and submandibular ep- ithelial development (Kadoya et al., 1995). One possi-
INTEGRINS IN DEVELOPING TOOTH 111
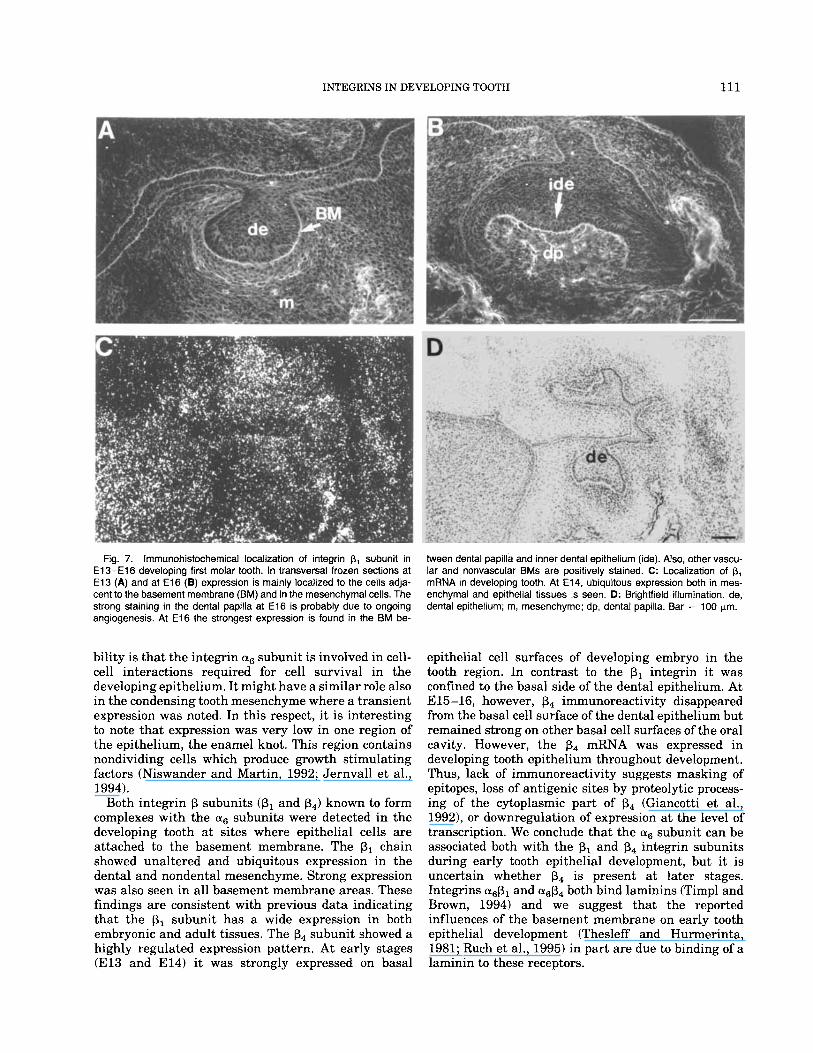
Fig. 7. lrnmunohistochemical localization of integrin p, subunit in E 1 3 - E l 6 developing first molar tooth. In transversal frozen sections at E l 3 (A) and at E l 6 (a) expression is mainly localized to the cells adja- cent to the basement membrane (BM) and in the mesenchymal cells. The strong staining in the dental papilla at E l 6 is probably due to ongoing angiogenesis. At E l 6 the strongest expression is found in the BM be-
Ween dental papilla and inner dental epithelium (ide). Also, other vascu- lar and nonvascular BMs are positively stained. C: Localization of p, mRNA in developing tooth. At E14. ubiquitous expression both in mes- enchymal and epithelial tissues is seen. D: Brightfield illumination. de, dental epithelium; m, mesenchyme; dp, dental papilla. Bar = 100 pn.
bility is that the integrin (Yg subunit is involved in cell- cell interactions required for cell survival in the developing epithelium. It might have a similar role also in the condensing tooth mesenchyme where a transient expression was noted. In this respect, it is interesting to note that expression was very low in one region of the epithelium, the enamel knot. This region contains nondividing cells which produce growth stimulating factors (Niswander and Martin, 1992; Jernvall et al., 1994).
Both integrin p subunits (PI and p4) known to form complexes with the (Yg subunits were detected in the developing tooth at sites where epithelial cells are attached to the basement membrane. The p1 chain showed unaltered and ubiquitous expression in the dental and nondental mesenchyme. Strong expression was also seen in all basement membrane areas. These findings are consistent with previous data indicating that the p1 subunit has a wide expression in both embryonic and adult tissues. The p4 subunit showed a highly regulated expression pattern. At early stages (El3 and E14) it was strongly expressed on basal
epithelial cell surfaces of developing embryo in the tooth region. In contrast to the p1 integrin it was confined to the basal side of the dental epithelium. At E15-16, however, p4 immunoreactivity disappeared from the basal cell surface of the dental epithelium but remained strong on other basal cell surfaces of the oral cavity. However, the p4 mRNA was expressed in developing tooth epithelium throughout development. Thus, lack of immunoreactivity suggests masking of epitopes, loss of antigenic sites by proteolytic process- ing of the cytoplasmic part of p4 (Giancotti et al., 1992), or downregulation of expression a t the level of transcription. We conclude that the as subunit can be associated both with the p1 and p4 integrin subunits during early tooth epithelial development, but it is uncertain whether p4 is present at later stages. Integrins and (Y&4 both bind laminins (Timpl and Brown, 1994) and we suggest that the reported influences of the basement membrane on early tooth epithelial development (Thesleff and Hurmerinta, 1981; Ruch et al., 1995) in part are due to binding of a laminin to these receptors.

112 SALMIVIRTA E T AL.
EXPERIMENTAL PROCEDURES Animals
Mouse embryos were obtained by mating NMRI mice. The day of the vaginal plug was designated as day 0 of embryonic development.
Antibodies Integrin a, was detected by an affinity-purified
monospecific polyclonal antibody raised against a syn- thetic peptide corresponding to the cytoplasmic domain of integrin a, (Hirsch et al., 1994). Integrin was detected with monoclonal antibody GoH3 (Immunotech S. A., Marseilles, France). Integrin p4 was detected with a rat monoclonal antibody against murine p4 in- tegrin (gift from Dr. s. J. Kennel, Biology Division, Oak National Laboratory, TN). Anti-p,-integrin mono- specific polyclonal antibody was as described by Gull- berg et al. (1989). Cy3‘”-conjugated affinity-purified goat anti-rabbit IgG and anti-rat IgG preparations were used as secondary antibodies (Jackson Immuno- Research Laboratories, West Grove, PA).
Oligonucleotide Probes Oligonucleotide probes were selected using OLIGO TM
software and purchased from Scandinavian Gene Syn- thesis AB (Koping, Sweden). To detect integrin a, mRNA, a 42-mer oligonucleotide probe: 5’-TGT GCT GAA CAA TAG GCC CAA CGT CTT CTT CAG TCT CAG GGT-3’, complementary to nucleotides 196-237 of 270 bp sequence of murine integrin a, mRNA, was used. These nucleotides corresponds to nucleotides 2425-2466 of human integrin a, mRNA (Suzuki et al., 1986). In order to obtain a probe encoding the murine a, cDNA, total RNA from adult mouse brain was re- verse-transcribed and amplified using two oligonucle- otides whose sequence was based on regions of high homology between the human and the chick a, cDNA. Two Fg of murine brain total RNA were reverse-tran- scribed in the presence of oligo-dT and amplified with a touchdown protocol ranging from 65°C to 55°C. The oligonucleotides used for amplification (5’-GCTCTA- GACGTTTCAGTGTGCACCAG-3’ and 5’-GCTCTAG- ATCTTGTTCTTCTTGAGGTG-3’) were designed to carry XbaI sites a t their extremity. A fragment of ap- proximately 1 kb was amplified and inserted into Blu- script I1 (Stratagene, La Jolla, CAI. A 700-bp EcoRI- XbaI subclone was partially sequenced on both sides. Using the FastA method (Pearson and Lipman, 1988) to compare sequences of the subclone with the human a, cDNA, an average identity of 85% was obtained. A 42-mer oligonucleotide probe: 5’-TTC CGG ATC ACG
3’, complementary to nucleotides 247-289 of mouse in- tegrin mRNA (Hierck et al., 19931, was used to de- tect integrin 016 mRNA. The probe does not distinguish between different splice variants of a6 integrin sub- unit. For integrin PI mRNA detection a 42-mer oligo- nucleotide probe: 5’-AGG CTC TGC ACT GAA CAC
TTGTCCTCGCGGGTGTCCAGGTTGAAGGCT-
ATT CTT TAT GCT CTG CAC AGA CGC-3’, comple- mentary to nucleotides 1915-1956 of mouse integrin p1 mRNA (Holers et al., 19891, was used. Integrin p4 mRNA was detected using a 45-mer oligonucleotide probe: 5’-CCA TCC GGA GGC CAT CAG AGA AGG AGG GTT TCA GGT GGA TGT TGC-3’, complemen- tary to nucleotides 1588-1633 of mouse integrin p4 mRNA detecting mRNA coding for extracellular do- mains (Kennel et al., 1993).
In Situ Hybridization In situ hybridization was performed according to
Durbeej et al. (1993). Briefly, embryonic mouse heads from stages embryonic day 13 (E13) to El8 were em- bedded in TissueTek (HistoLab, Miles Inc., Elkhart, IN), frozen in liquid nitrogen, cut in 10-pm sections on a cryostat at -20°C and collected on sterile slides coated with poly-L-lysine (50 pg/ml). Sections were fixed in 4% paraformaldehyde, rinsed twice in phos- phate-buffered saline (PBS), pH 7.0 and dehydrated in graded ethanol series including a 5-minute incubation in chloroform. Oligonucleotide probes were labelled at the 3’ end with [a-35 SldATP (Amersham Sweden AB, Solna) using terminal deoxyribonucleotidyl trans- ferase (Scandinavian Diagnostic Services, Falkenberg, Sweden) to specific activity of 1 x lo9 cpdpg. Hybrid- ization was performed in 50% formamide, 4 x SSC (salt sodium citrate), 1 x Denhardt’s solution, 10% dextran sulfate, 0.25 mg/ml yeast tRNA, 0.5 mg/ml sheared salmon sperm DNA, 1% sarcosyl, 0.02M Na,HP04 (pH 7.0), 0.05 M dithiothreitol, using lo7 cpdml respective probe. The sections were hybridized at +42”C for 15-18 hours in a humidified chamber with 0.1 ml of hybridization solution per slide. Slides were subsequently washed four times (15 minutes each) at + 60-65°C in 1 x SSC. As controls for specific hybridization, sections were hybridized with the same amount of labelled probe plus unlabelled probe in ex- cess. The slides were dipped in Kodak NTB-2 photo emulsion (diluted 1:l in water), exposed for 2-4 weeks at + 4“C, developed and counterstained with cresyl vi- olet or hematoxylin. Photographs were taken under brightfield and darkfield illumination.
Immunohistochemistry Methanol-fixed cryostat sections (8 pm) of embryonic
mice heads a t stages E13-El8 were stained for immu- nohistochemistry. Nonspecific binding sites were blocked by incubating the sections with 10% goat se- rum. After 1 hr incubation with primary antibody, slides were washed in PBS (3 x 15 minutes) and incu- bated with secondary antibody for 30-40 minutes, washed in PBS (3 x 15 minutes) and mounted with Vectashield Mounting medium (Vector Laboratories, Burlingame, CAI. All incubations were made at room temperature. Slides were examined using a Zeiss Ax- iophot microscope. Control sections were incubated with 10 % goat serum instead of the primary antibody.

INTEGRINS IN DEVELOPING TOOTH 113
ACKNOWLEDGMENTS This study was supported by grants from the Swed-
ish Cancer Fund, Medicinska Forskningsrbdet, Human Capital and Mobility Program, and Knut och Alice Wallenberg Foundation.
REFERENCES BBgue-Kirn, C., Smith, A.J., Loriot, M., Kupferle, C., Ruch, J.V., and
Lesot, H. (1994) Comparative analysis of TGFbs, BMPs, IGF1, msxs, fibronectin, osteonectin and bone sialoprotein gene expres- sion during normal and in vitro-induced odontoblast differentia- tion. Int. J . Dev. Biol. 38 405-420.
Busk, M., Pytela, R., and Sheppard, D. (1992) Characterization of the integrin a,& as a fibronectin-binding protein. J . Biol. Chem. 267: 5790-5796.
Charo, I.F., Nannizzi, L., Smith, J.W., and Cheresh, D.A. (1990) The vitronectin receptor a& binds fibronectin and acts in concert with asp1 in promoting cellular attachment and spreading on fibronec- tin. J . Cell. Biol. 111: 2795-2800.
Durbeej, M., Sijderstrom, S., Ebendal, T., Birchmeier, C., and Ekblom, P. (1993) Differential expression of neutrophin receptors during re- nal development. Development 119: 977-989.
Giancotti, F.G., Stepp, M.A., Suzuki, S., Engvall, E., andRuoslahti, E. (1992) Proteolytic processing of endogenous and recombinant p4 integrin subunit. J . Cell Biol. 118 951-959.
Gullberg, D., Terracio, L., Borg, T.K., and Rubin, K. (1989) Identifi- cation of integrin-like matrix receptors with affinity for interstitial collagens. J . Biol. Chem. 264 12686-12694.
Hierck, B.P., Thorsteinsdottir, S., Niessen, C.M., Freund, E., VanIpe- ren, L., Feijen, A., Hogervorst, F., Poelmann, R.E., Mummery, C.L., and Sonnenberg, A. (1993) Variants of the laminin receptor in early murine development: Distribution, molecular cloning and chromo- somal localization of the mouse integrin alpha 6 subunit. Cell Adh. Commun. 1: 33-53.
Hirsch, E., Gullberg, D., Balzac, F., Altruda, F., and Silengo, L., and Tarone, G. (1994) a, integrin subunit is predominantly located in nervous tissue and skeletal muscle during mouse development. Dev. Dyn. 201: 108-120.
Holers, V.M., Ruff, T.G., Parks, D.L., McDonald, J.A., Ballard, L.E., and Brown, E.J. (1989) Molecular cloning of a murine fibronectin receptor and its expression during inflammation. J. Exp. Med. 169 1589-1605.
Jernvall, J., Kettunen, P., Karavanova, I., Martin, L.B., and Thesleff, I. (1994) Evidence for the role of the enamel knot as a control center in mammalian tooth cusp formation: Non-dividing cells express growth stimulating Fgf-4 gene. Int. J . Dev. Biol. 38: 463-469.
Kadoya, Y., Kadoya, K., Durbeej, M., Holmvall, K., Sorokin, L., and Ekblom, P. (1995) Antibodies against domain E3 of laminin-1 and integrin as subunit perturb branching epithelial morphogenesis of submandibular gland, but by different modes. J. Cell Biol. 129: 521-534.
Kennel, J., Foote, L.J., Cimino, L., Rizzo, M.G., Chang, L.Y., and Sacchi, A. (1993) Sequence of a cDNA encoding the beta 4 subunit of murine integrin. Gene 130 209-216.
Lee, E.C., Lotz, M.M., Steele, G.D., and Mercurio, A.M. (1992) The integrin asp4 is a laminin receptor. J . Cell Biol. 117: 671-678.
Lesot, H., Osman, M., and Ruch, J.V. (1981) Immunofluorescent lo- calization of collagens, fibronectin and laminin during terminal dif- ferentiation of odontoblasts Dev. Biol. 82: 371-381.
Lesot, H., B6gue-Kirn, C., Kubler, M.D., Meyer, J.M., Smith, A.J., Cassidy, N., and Ruch, J.V. (1993) Experimental induction of odon- toblast differentiation and stimulation during reparative processes. Cells Mater. 3: 201-217.
Lumsden, A.G.S. (1988) Spatial organization of the epithelium and the role of neural crest cells in the initiation of mammalian tooth germ. Development 103: 155-169.
Marshall, J.F., Rutherford, D.C., McCartney, A.C.E., Mitjans, F., Goodman, S.L., and Hart, I.R. (1995) a,& is a receptor for vitronec- tin and fibrinogen, and acts with aspl to mediate spreading on fibronectin. J. Cell Sci. 108: 1227-1238.
Mina, M., and Kollar, E.J. (1987) The induction of odontogenesis in non-dental mesenchyme combined with early murine mandibular arch epithelium. Arch. Oral Biol. 32: 123-127.
Niswander, L., and Martin, G.R. (1992) Fgf-4 expression during gas- trulation, myogenesis, limb and tooth development in the mouse. Development 114: 755-768.
Partanen, A-M., and Thesleff, I. (1987) Localization and quantitation of lZ5I-epidermal growth factor binding in mouse embryonic tooth and other embryonic tissues at different developmental stages. Dev. Biol. 120: 186-197.
Pasqualini, R., Bodorova, J., Song, Y., and Hemler, M.E. (1993) A study of the structure, function and distribution of p, integrin using novel anti-p, monoclonal antibodies. J . Cell Sci. 105: 101-111.
Pearson, W.R., and Lipman, D.J. (1988) Improved tools for biological sequence comparison. Proc. Natl. Acad. Sci. USA 85: 2444-2448.
Ruch, J.V. (1987) Determinisms of odontogenesis. Cell Biol. Rev. 14: 1-112.
Ruch, J.V., Lesot, H., and Begue-Kirn, C. (1995) Odontoblast differ- entiation. Int. J . Dev. Biol. 3 9 51-68.
Slavkin, H.C. (1974) Embryonic tooth formation: A tool for develop- mental biology. Oral Sci. Rev. 4 1-136.
Sonnenberg, A., Linders, C.J.T., Moddeman, P.W., Damsky, C.H., Au- mailley, M., and Timpl, R. (1990) Integrin recognition of different cell-binding fragments of laminin (Pl, P3, E8) and evidence that asp1 but not asp4 functions as a major receptor for fragment E8. J . Cell Biol. 10: 2145-2155.
Surinenberg, A., Calafat, J., Janssen, H., Daams, H., van der kaij- Helmer, L.M.H., Falcioni, R., Kennel, S.J., Aplin, J.D., Baker, J., Loizidou, M., and Garrod, D. (1991) Integrin asp4 complex is located in hemidesmosomes, suggesting a major role in epidermal cell-base- ment membrane adhesion. J. Cell Biol. 113: 907-917.
Sorokin, L., Sonnenberg, A., Aumailley, M., Timpl, R., and Ekblom P. (1990) Recognition of the laminin E8 cell-binding site by an integ- rin possessing the as subunit is essential for epithelial polarization in developing kidney tubules. J. Cell Biol. 111: 1265-1273.
Spinardi, L., Einheber, S., Cullen, T., Milner, T.A., and Giancotti, F.G. (1995) A recombinant tail-less integrin p4 subunit disrupts hemidesmosomes, but does not suppress a&,-mediated cell adhe- sion to laminins. J. Cell Biol. 1 2 9 473-487.
Suzuki, S., Argraves, W.S., Pytela, R., Arai, H., Krusius, T., Piersch- bacher, M.D., and Ruoslahti, E. (1986) cDNA and amino acid se- quences of the cell adhesion protein receptor recognizing vitronec- tin reveal a transmembrane domain and homologies with other adhesion protein receptors. Proc. Natl. Acad. Sci. U.S.A. 83: 8614- 8618.
Thesleff, I., and Hurmerinta, K. (1981) Tissue interactions in tooth development. Differentiation 18: 75-88.
Thesleff, I., Barrach, H.T., Foidart, J.M., Vaheri, A., Pratt, R.M., and Martin G.R. (1981) Changes in the distribution of type IV collagen, laminin, proteoglycan and fibronectin during mouse tooth develop- ment. Dev. Biol. 81: 182-192.
Thesleff, I., Vaahtokari, A,, and Vainio, S. (1990) Molecular changes during determination and differentiation of the dental mesenchy- ma1 cell lineage. J. Biol. Buccale 18: 179-188.
Timpl, R., and Brown J.C. (1994) The laminins. Matrix Biology 14: 275 -28 1.
Tozeren, A., Kleinman, H.K., Wu, S., Mercurio, A.M., and Byers, S.W. (1994) Integrin asp4 mediates dynamic interactions with laminin. J . Cell Sci. 107: 3153-3163.
Yamada, S., Yamada, K.M., and Brown, K.E. (1994) Integrin regula- tory switching in development: Oscillation of ps integrin mRNA expression during epithelial-mesenchymal interactions in tooth de- velopment. Int. J. Dev. Biol. 38: 553-556.