integrative proteomic analysis of the nucleus accumbens in rhesus monkeys following cocaine...
TRANSCRIPT

ORIGINAL ARTICLE
Integrative proteomic analysis of the nucleus accumbensin rhesus monkeys following cocaine self-administrationNS Tannu1, LL Howell2,3 and SE Hemby1,4
1Department of Physiology and Pharmacology, Wake Forest University School of Medicine, Winston-Salem, NC, USA;2Neuroscience Division, Yerkes National Primate Research Center, Emory University, Atlanta, GA, USA; 3Department ofPsychiatry and Behavioral Sciences, Emory University School of Medicine, Atlanta, GA, USA and 4Department of Psychiatryand Behavioral Medicine, Wake Forest University School of Medicine, Winston-Salem, NC, USA
The reinforcing effects and long-term consequences of cocaine self-administration have beenassociated with brain regions of the mesolimbic dopamine pathway, namely the nucleusaccumbens (NAc). Studies of cocaine-induced biochemical adaptations in rodent models haveadvanced our knowledge; however, unbiased detailed assessments of intracellular alterationsin the primate brain are scarce, yet essential, to develop a comprehensive understanding ofcocaine addiction. To this end, two-dimensional difference in gel electrophoresis (2D-DIGE)was used to compare changes in cytosolic protein abundance in the NAc between rhesusmonkeys self-administering cocaine and controls. Following image normalization, spots withsignificantly differential image intensities (P < 0.05) were identified, excised, trypsin digestedand analyzed by matrix-assisted laser-desorption ionization time-of-flight time-of-flight(MALDI-TOF-TOF). In total, 1098 spots were subjected to statistical analysis with 22 spotsfound to be differentially abundant of which 18 proteins were positively identified by massspectrometry. In addition, approximately 1000 protein spots were constitutively expressed ofwhich 21 proteins were positively identified by mass spectrometry. Increased levels of proteinsin the cocaine-exposed monkeys include glial fibrillary acidic protein, syntaxin-binding protein3, protein kinase C isoform, adenylate kinase isoenzyme 5 and mitochondrial-related proteins,whereas decreased levels of proteins included b-soluble N-ethylmaleimide-sensitive factorattachment protein and neural and non-neural enolase. Using a complimentary proteomicsapproach, the differential expression of phosphorylated proteins in the cytosolic fraction ofthese subjects was examined. Two-dimensional gel electrophoresis (2DGE) was followed bygel staining with Pro-Q Diamond phosphoprotein gel stain, enabling differentiation ofapproximately 150 phosphoprotein spots between the groups. Following excision and trypsindigestions, MALDI-TOF-TOF was used to confirm the identity of 15 cocaine-alteredphosphoproteins. Significant increased levels were detected for c-aminobutyric acid type Areceptor-associated protein 1, 14-3-3 c-protein, glutathione S-transferase and brain-typealdolase, whereas significant decreases were observed for b-actin, Rab GDP-dissociationinhibitor, guanine deaminase, peroxiredoxin 2 isoform b and several mitochondrial proteins.Results from these studies indicate coordinated dysregulation of proteins related to cellstructure, signaling, metabolism and mitochondrial function. These data extend andcompliment previous studies of cocaine-induced biochemical alterations in human post-mortem brain tissue, using an animal model that closely recapitulates the human conditionand provide new insight into the molecular basis of the disease and potential targets forpharmacotherapeutic intervention.Molecular Psychiatry (2010) 15, 185–203; doi:10.1038/mp.2008.53; published online 27 May 2008
Keywords: cocaine; protein expression; nucleus accumbens; phosphorylation; monkey
Introduction
Cocaine abuse remains a significant health concern inthe United States and abroad.1,2 Approximately 2.4
million Americans, 12 years and older, currently usecocaine. This represents approximately 0.97% ofthe population and a 20% increase in the number ofusers since 2002.1 The propensity to use cocaine isinfluenced by both positive (euphoric, pleasurableeffects) and negative (withdrawal, depressed moodstates and drug cravings) consequences, including thedevelopment of neuroadaptive changes in specificbrain regions.3 Understanding the neurobiologicalmechanisms that contribute to cocaine abuse is
Received 19 December 2007; revised 12 March 2008; accepted 10April 2008; published online 27 May 2008
Correspondence: Dr SE Hemby, Department of Physiology andPharmacology, Wake Forest University School of Medicine,Medical Center Boulevard, Winston-Salem, NC 27157, USA.E-mail: [email protected]
Molecular Psychiatry (2010) 15, 185–203& 2010 Nature Publishing Group All rights reserved 1359-4184/10 $32.00
www.nature.com/mp

critical for developing new pharmacotherapies andmatching clinical diagnoses with appropriate treat-ment strategies.
The majority of research designed to understandthe addictive properties of cocaine has focusedlargely on the neural circuit that mediates motiva-tional processes, the mesolimbic dopamine pathway.Projections from the ventral tegmental area (VTA) tothe nucleus accumbens (NAc) comprise part of themesolimbic dopamine pathway and have been iden-tified as a critical substrate in the reinforcing effects ofcocaine in humans and animal models. Indeed,cocaine has been shown to increase extracellulardopamine concentrations within this pathway, aneffect attributed to the drug’s abuse liability. Further-more, there is ample evidence that repeated cocaineuse leads to biochemical adaptations in mesolimbicbrain regions and these adaptations appear to berelevant to the processes of sensitization, craving,withdrawal and relapse.4 Studies in rodent modelsindicate cocaine-induced biochemical alterations inregions associated with the mesolimbic pathwayincluding upregulation of the cyclic AMP (cAMP)pathway,5–8 activator protein 1 family members9–11
and glutamate signaling.12–14
Genomics-based analyses, such as microarrays,have revealed novel mechanisms of drug-inducedneuronal and non-neuronal dysregulation in humanpostmortem brain tissue15,16 and rodent models.17–21
Although these studies have been highly informativein furthering our understanding of drug-inducedtranscriptional regulation contributing to long-termchanges in cellular function, research determiningcoordinate changes in the expression of multipleproteins following cocaine exposure has beenscarce.22 To comprehend the intricate neuroadaptivemachinery implicated in the development and ex-pression of cocaine abuse, it is desirable to comple-ment the global gene expression analyses with studiesexamining the corresponding proteomes. Currentunderstanding of stimulant-induced neurobiologicalalterations (including expression and functionalgenomics/proteomics, epigenetic modifications, andso on) is based predominantly on rodent models ofhuman drug taking; however, the direct determina-tion of protein expression status and factors regulat-ing protein expression in primate brain has beenlacking, yet it is essential for understanding theconsequential molecular pathology of cocaine addic-tion in humans.
Recently, we interrogated a portion of the NAcproteome in individuals following cocaine overdoseusing two-dimensional difference in gel electrophor-esis (2D-DIGE) combined with tandem mass spectro-metry (MS/MS) analysis.23 The abundances ofproteins from several families were found to besignificantly altered, including proteins belonging tocell structure, synaptic plasticity/signal transduction,mitochondrial function and metabolic pathways.Interestingly, several of the proteins were functionallyassociated with N-methyl-D-aspartate (NMDA) and
a-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid(AMPA) receptors, previously shown to be increased inthe VTA and NAc of cocaine overdose victims.16,24
In addition to assessing changes in the abundanceof native proteins, changes in the abundance of post-translational modifications represent an importantmeans of activation/deactivation of various proteinsand thereby significantly influencing cellular func-tion. Phosphorylation, the most prevalent covalentmodification of proteins in eukaryotic cells, affectsapproximately one-third of all proteins at any giventime.25 Phosphorylation is catalyzed by a variety ofprotein kinases including protein kinase A, one of themost well studied in cocaine abuse,4 which phos-phorylates serine and threonine residues on multipletargets including cAMP response element-bindingprotein.26,27 Detecting changes in post-translationallymodified proteins in human postmortem tissue isdifficult at best; however, similar assessments innonhuman primate models of cocaine abuse arepossible and have the advantage of recapitulatingbehavioral aspects of human drug intake and offeringneuroanatomical and biochemical similarities com-pared with other species used as models.28
Historically, research examining the abundance ofproteins and post-translational modifications as afunction of cocaine abuse has been restricted to theserial analysis of individual proteins. With the adventof high-throughput separation and MS-based analysisstrategies, it is possible to provide a broad andunbiased coverage of the proteome to delineate themultitude of neurobiological effects of abuseddrugs.22,29 In the present study, the proteome andphosphoproteome of the NAc was examined follow-ing chronic cocaine self-administration in rhesusmonkeys. Proteins contained in the cytosolic fractionwere assessed by 2D-DIGE and a phospho-specificdye for native and phosphorylated proteins, respec-tively. Proteins that were differentially expressedbetween the cocaine and control groups were isolatedand followed by de novo identification by matrix-assisted laser-desorption ionization time-of-flighttime-of-flight (MALDI-TOF-TOF) MS. These strate-gies provide new insight into the neurobiological/neuropathological changes associated with chronicintake and may yield novel targets for future drugdevelopment.
Materials and methods
Surgery and self-administration proceduresEight male adult rhesus monkeys (Macaca mulatta)were singly housed with standard enrichment, in-cluding social enrichment, human interaction, varietyin diet and age appropriate objects as dictated by theAnimal Welfare Act and the Emory University Policyfor Environmental Enhancement. Each subject wasfed Purina monkey chow (Ralston Purina, St Louis,MO, USA), fruits and vegetables. Food and waterwere available ad libitum. Animal care proceduresstrictly followed the National institutes of Health
Neuroproteome of cocaine in nonhuman primatesNS Tannu et al
186
Molecular Psychiatry

Guide for the Care and Use of Laboratory Animals andwere approved by the Institutional Animal Care andUse Committee of Emory University and Wake ForestUniversity Schools of Medicine. Four monkeys weresurgically prepared with chronically indwelling ve-nous catheters using procedures described pre-viously.30,31 Under appropriate anesthesia, eitherisoflurane alone or ketamine in combination withdiazepam, and under aseptic conditions, one end of asilicone catheter was passed by way of a jugular orfemoral vein to the level of the right atrium and venacava, respectively. The distal end of the catheter waspassed under the skin and attached to a vascularaccess port (Access Technologies, Skokie, IL, USA),which remained subcutaneous in the center of theback for easy access. The 0.25 ml unit was accessedduring testing with special right-angle Huber needles(Access Technologies) that minimize damage to theport membrane and allow for repeated punctures overa year or more. Catheters were flushed after daily testsessions with heparinized saline (0.9%). Daily ex-perimental sessions were conducted within a venti-lated, sound-attenuating chamber with each monkeyseated in a standard primate chair of the typecommercially available (Primate Products, RedwoodCity, CA, USA). A panel equipped with a responselever and stimulus lights was mounted on the front ofthe chair. The vascular access port was connectedthrough polyvinyl-chloride tubing to a motor-drivesyringe located outside the test chamber to yield aprecise injection volume of 2.0 ml during drug self-administration experiments with dose determined bythe concentration of drug solution in the syringe.Experimental procedures were controlled online by amicroprocessor and electromechanical programmingsystems, and data were monitored and recordedduring daily sessions.
Four monkeys were trained to self-administercocaine by pressing a response key while seated in aprimate chair. Responding was initiated using a 1-response fixed-ratio schedule (FR 1) so that eachresponse in the presence of a red light produced anintravenous drug injection and the brief illuminationof a white light followed by a timeout. The ratio valuewas increased gradually as responding increased.When the schedule value reached FR 20, druginjection no longer followed completion of each FRand, instead, was arranged to follow an increasingnumber of FR components. Ultimately, the schedulewas a second-order schedule of FR 20 componentswith drug injection following the first componentcompleted after 10 min had elapsed (fixed-interval(FI) 600-s (FR 20:S)). A 2-s white light was presentedupon completion of each FR 20 component. Drugadministration was accompanied by a change in thestimulus light from red to white for 15 s, followed by a1-min timeout. Daily sessions consisted of fiveconsecutive 10-min intervals. The unit dose ofcocaine remained constant at 0.1 mg kg�1 per injec-tion.30,32 Use of this second-order procedure andlimiting the daily session to approximately 1 h
enabled the standardization of total drug intake to0.5 mg kg�1 per session.
In total, 18–24 h after the last drug self-administra-tion studies, monkeys were anesthetized with Telazol(4.0 mg kg�1; i.v.), administered intravenous heparinfollowed by an overdose of intravenous sodiumpentobarbital. This time point was selected to ensurethat no cocaine was present at the time of killing andto avoid any effects associated with cocaine with-drawal. After the confirmed absence of brain stemreflexes was established, the monkeys were transcar-dially perfused with ice-cold phosphate-bufferedsaline (PBS; pH 7.2–7.4), the brain was removed andplaced in 4 1C PBS for 5 min. Brains were blockedusing a rhesus monkey brain matrix that allows 4 mmcoronal blocks at various AP locations (ElectronMicroscopy Sciences, Ft Washington, PA, USA).Tissue was frozen at �80 1C within 40 min of necropsyand stored at �80 1C until further processing.
Protein isolation and fractionationThe NAc was dissected from the rostral pole to thebeginning of the anterior commissure, using theinternal capsule and lateral ventricle as landmarks33
(Supplementary Figure). A steel mortar and pestlechilled in dry ice were used to pulverize the frozenbrain tissue into a dry homogenate in the presence ofliquid nitrogen. Tissue proteins were fractionated asdescribed previously.13,23,24,34 Fractionation enablesthe enrichment of low abundance proteins in distinctcellular fractions compared to total cell/tissue homo-genate and therefore allows increased coverage of theanalyzed proteome.
Cytosolic fractions from the NAc were evaluated inthe present study to compliment and extend similaranalysis conducted in postmortem brain of humancocaine overdose victims.23 It is important to note thatthe membrane fraction was not included in thecurrent analysis primarily due to the limitations of2DG for analyzing membrane proteins.35 Most hydro-phobic (membrane-bound) proteins are insoluble innondetergent sample buffer used for isoelectricfocusing (IEF) and the ones which are solubleprecipitate at their respective isoelectric points (pI).Second, the pIs of hydrophobic proteins are generallyalkaline and even with the use of extended pHgradients, the proteins are difficult to resolve well atthe basic end. Liquid chromatography (LC)-MSmethodologies can overcome issues of hydrophobicmembrane protein separation. Studies are currentlyunderway in our lab utilizing multidimensionalliquid chromatographic separation along with MS toexplore the membrane proteome in these subjects.
Experiment 1: Differential native protein expressionusing 2D-DIGE
Cyanine dye labeling. Minimal labeling of the lysineresidues was achieved by reaction with cyanine dyes,as described previously.23,36,37 Briefly, a normalization(pooled) sample was prepared by combining 50 mg
Neuroproteome of cocaine in nonhuman primatesNS Tannu et al
187
Molecular Psychiatry

from each sample. The labeling of 50mg of proteinsample (each sample) was optimized by labelingwith 200 pmol of appropriate dye (suspended in> 99.5% pure dimethylformamide). On each gel,electrophoresis was carried out on a pooled samplelabeled with Cy2 and also individuals samples fromthe cocaine and control groups labeled with eitherCy3 or 5.
Two-dimensional polyacrylamide gel electrophoresis. Two-dimensional polyacrylamide gel electrophoresis (2D-PAGE) was executed as described previously.23,36,37
Aliquots of cytosolic proteins were diluted in400ml of rehydration buffer and increased to a finalvolume of 450ml with DeStreak rehydration buffer(GE Healthcare, Piscataway, NJ, USA). Isoelectricfocusing was performed using Immobiline DryStrips(240�3� 0.5 mm, pH 4-7 linear; GE Healthcare) onan Ettan IPGphor apparatus (GE Healthcare).23,36,37
Gel image analysis. Image analysis was conducted asdescribed previously,23,36,37 using a Typhoon 9400scanner (GE Healthcare) to scan all gels at 100mmresolution. The photomultiplier tube was set toensure maximum pixel intensity of 85 000–95 000for all the images in every gel. Image analysis wasperformed using DeCyder 5.01 software (GEHealthcare). The Difference In-gel Analysis (DIA)mode of DeCyder was employed for protein spotdetection and also for normalization of cocaine andcontrol gel images to the pooled sample gel image.The photomultiplier tube was set to ensure maximumpixel intensity of 85 000–95 000 for all the images inevery gel. After spot detection, the abundancechanges are represented by the normalized volumeratio of cocaine or control to the pooled sample. Thefollowing parameters were used for spot filtering;slope > 1.0, area < 350, peak height < 350 andvolume < 100 000. The spots were authenticatedmanually for all the gels. Spot maps from all thegels were first matched by manual landmarks andthen in automatic mode by DeCyder BiologicalVariation Analysis (BVA). Protein spot matches wereconfirmed manually for all the gels. The average ratioand also the corresponding student’s t-test value foreach protein spot was calculated based on all gelimages in the DeCyder BVA mode.37 Selected spotswere isolated using the Ettan Spot HandlingWorkstation (GE Healthcare) and picked proteinswere prepared for MS analysis as described in thesection below entitled in-gel trypsin digestion.
Experiment 2: Phosphoproteome gel staining andimage analysisAliquots of cytosolic protein fractions were separatedby 2D gels in the same manner as the samples forexperiment 1 and as described above in the subsec-tion 2D-PAGE. Following, gels were stained forproteins containing phosphorylated moieties accord-ing to the instructions provided by the manufacturer(Molecular Probes, Eugene, OR, USA). Gels were
fixed in 500 ml of 50% methanol and 10% trichlor-oacetic acid for 15 h and then sequentially washedthrice with 500 ml distilled water for 15 min, followedby incubation with 500 ml Pro-Q Diamond phospho-protein stain for 2 h in the dark. Gels were then de-stained with 20% acetonitrile, 50 mM sodium acetate(pH = 4) for 90 min and washed twice for 5 min eachin distilled water. All gels were scanned at 100 mmresolution of Typhoon 9400 scanner (GE Healthcare),using the green laser (532 nm) for excitation and610 nM band pass emission filter for visualization ofproteins with phospho-specific moieties. The photo-multiplier tube was set to ensure maximum pixelintensity of 80 000–90 000 for all the gels. ImageQuantV5.2 and DeCyder 5.01 (GE Healthcare) were used toremove extraneous areas to the scanned gel imagesand for performing image analysis, respectively. Spotswere authenticated manually for all the gels duringthe differential in-gel analysis (DeCyder DIA). Spotmaps from all gels were analyzed by manuallyassigning landmarks and then by automatic mode byDeCyder BVA, which enables the matching of multi-ple 2DGE gels for comparison and statistical analysisof protein abundance changes. The protein spotmatches performed in the automatic mode were alsoconfirmed manually for all the gels.37 To visualize theproteome from this specific pH and mass range, gelswere stained with Sypro Ruby stain overnight. Theexcess stain was removed by 10% methanol and 6%glacial acetic acid for 20 min. The gels were imagedand analyzed, in a similar fashion to the phospho-protein stain using Typhoon 9400 scanner andDeCyder image analysis software, respectively. Stu-dent’s t-test was used to determine differentialabundance of the phosphoproteins between groups.Individual protein spots from the 2D gels wereexcised with 1.5 mm diameter gel cutter (The GelCompany, San Francisco, CA, USA).
In-gel trypsin digestionThe excised gel spots from experiments 1 and 2 werewashed for 20 min, twice in 100ml of solution of50 mM ammonium bicarbonate, 50% methanol (v/v)in distilled water and once in 75% acetonitrile indistilled water for 30 min or until the gel plugs turnedopaque. Lyophilized trypsin (20mg) (883 pmol; Pro-mega, Madison, WI, USA) was reconstituted in 1 ml of20 mM ammonium bicarbonate and incubated for15 min at 37 1C. The gel fragments were dried byvacuum centrifugation and then incubated overnightwith 10 ml (200 ng) of trypsin at 37 1C. The supernatantfrom trypsin digest was transferred to a low retention96-well plate. Peptides from the gel pieces weresequentially extracted twice in 100ml of extractionbuffer (50% (v/v) acetonitrile, 0.1% (v/v) trifluoroa-cetic acid in distilled water). The original trypticsupernatant and the supernatants from two sequentialextractions were combined and dried in a vacuumcentrifuge. The dried peptides from each gel plugwere dissolved in 5 ml of 50% (v/v) acetonitrile, 0.1%trifluoroacetic acid in distilled water and 0.5 ml
Neuroproteome of cocaine in nonhuman primatesNS Tannu et al
188
Molecular Psychiatry

deposited on the stainless-steel MALDI target plate.After drying, the spot residue was mixed with 0.5 mlof 5 mg ml�1 of a-cyano-4-hydroxy-cinnamic acid(Sigma-Aldrich, St Louis, MO, USA) in 50% (v/v)acetonitrile, 0.1% trifluoroacetic acid in distilledwater.23,37
MALDI-TOF-TOFMass spectrometry analyses were performed using theApplied Biosystems 4700 Proteomics Analyzer (MAL-DI-TOF-TOF; Foster City, CA, USA) in reflector modefor positive ion detection. The laser wavelength andthe repetition rate were 355 nm and 200 Hz, respec-tively. All the MS spectra resulted from accumulationof at least 2000 laser shots. The peak detection criteriaused were: minimum S/N of 8, local noise windowwidth mass/charge (m/z) of 200 and minimum full-width half-maximum (bins) of 2.9. The mass spectrawere calibrated using the three trypsin auto digestproducts: fragment 100–107 ([MþH]þ = 842.51 Da),fragment 90–99 ([MþH]þ = 1045.556 Da) and frag-ment 50–69 ([MþH]þ = 2211.105 Da). A maximum ofthe 10 strongest precursor ions per sample werechosen for MS/MS analysis. The following mono-isotopic precursor selection criteria were used for theMS/MS: minimum S/N filter of 10, excluding themost commonly observed peptide peaks for trypsinand keratin, and excluding the precursors within 150resolution. In the TOF1 stage, all ions were acceler-ated to 1 kV under conditions promoting metastablefragmentation. The peak detection criteria used were:S/N of 8 and local noise window width of 250 (m/z).23
Protein identificationThe peak lists generated by the 4000 Series Explorersoftware (version 3.6) were submitted to GPS Explorer(Applied Biosystems) to search against the NationalCenter for Biotechnology Information nonredundantfor protein characterization using the limited M.mulatta database.23,38 The following parameters wereused: one allowed missed cleavage, ±50 p.p.m. for m/z error for MS and 0.1 Da (m/z) error for MS/MS,partial modification of cysteine (carbamidomethyl-cysteine), methionine (oxidized) and phosphorylationof serine, threonine and tyrosine. Peptide massfingerprint (PMF) and MS/MS spectra were inter-preted with the MASCOT software (Matrix ScienceLtd, London, UK).39,40 Database searches, throughMASCOT, using combined PMF and MS/MS datasetswere performed through GPS Explorer software. Thecriterion for identification was a MASCOT confidenceinterval greater than 95%.
In order to reduce false-positive identification ofproteins, an open access (http://www.matrixscience.-com/help/decoy_help.html) Perl script ‘decoy.pl.gz.’was used to randomize the database entries from theforward database (target database) and create a newdatabase (decoy database).41 Randomization, insteadof reversal, of peptide sequence was used to establishthe decoy database as it is more suitable for PMFincorporated with the MS/MS searches in the current
study and also the fact that a reverse decoy databasewill have at least half of the tryptic peptide massvalues unchanged. The decoy database is comprisedof random sequences of the same length and has thesame average amino-acid composition as the forwarddatabase. As no matches are expected to be generatedagainst the decoy database, the number of matchesfound provides a good estimate of the number of falsepositives present from the real database. The originalsearch was repeated using identical search para-meters against the decoy database (randomizedsequences).
De novo sequencingThe method for the de novo peptide sequencing for M.mulatta has been published recently by our group andwas adapted for the current study,42 using the PEAKSStudio 4.0 (Bioinformatics Solutions, Waterloo, ON,Canada) de novo sequencing software. One potentialobstacle to comprehensive assessment of proteinalterations in rhesus monkey is the relative paucityof available protein annotations for this speciesmaking it difficult to identify proteins. The use ofstandard database search engines (for example,MASCOT) has a limitation in that ‘broad speciesdatabase’ searches are needed resulting in less thanoptimal protein annotation. This limitation can beovercome in some respects using a de novo sequen-cing strategy, in which partial or complete amino-acidsequence information is obtained using either manualor automated de novo peptide sequence analysisusing PEAKS software. Briefly, when positive char-acterization was not obtained for all spectra, spectrawere subjected to PEAKS de novo analysis. The denovo sequencing parameters used were as follows:parent- and fragment-mass error tolerance of 0.08 U;trypsin as the protease with one maximum missedcleavage allowed; deconvolution of the charge state inthe spectra to generate a spectra in which eachmonoisotopic peak becomes singly charged; partialmodification of cysteine (carbamidomethyl-cysteine)and methionine (oxidized). The most abundantpeptide fragments ‘b- and y-ions’; the less abundantpeptide fragments ‘a-ions’; the neutral losses of waterand ammonia for b- and y-ions; and also theimmonium ions were utilized to develop confidentand complete peptide sequences de novo from MS/MS spectra. The sequences generated from eachspectrum were used for protein identification bysequence homology in the mammalian database usingeither the PEAKS or SPIDER software (SoftwareProtein Identifier). Protein identification was con-firmed by protein mass and pI accuracy.
Results
Behavioral dataThree of four subjects readily acquired drug self-administration during the first month of training. Thefourth subject required approximately 4 monthsestablishing reliable drug self-administration and
Neuroproteome of cocaine in nonhuman primatesNS Tannu et al
189
Molecular Psychiatry

drug intake was very erratic during this extendedtraining period. Once stable self-administration beha-vior was established, all subjects reliably received allscheduled injections during daily sessions. Total drugintake for the group of four subjects over the 18-month period was 37.9±4.6 mg kg�1. Previously, wehave reported significant alterations in ionotropicglutamate receptor subunits in the NAc from thesubjects used in the present study.24
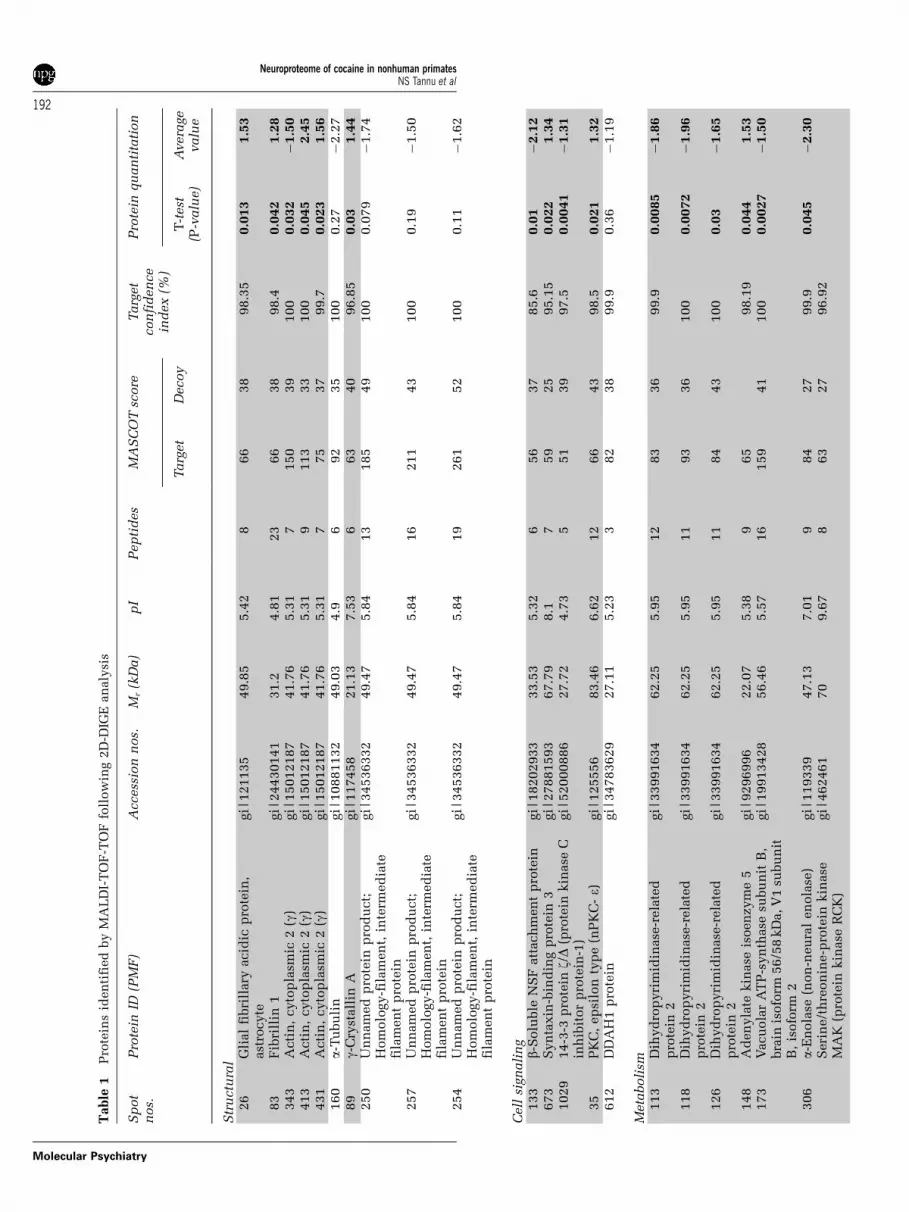
2D-DIGECytosolic fractions of NAc from CSA rhesus monkeysand controls were compared using the DIGE proteo-mics strategy to determine differences in the abun-dance levels of proteins in a pH range of 4–7 toelucidate the molecular mechanisms by which pre-vious exposure to cocaine alters the cytosolic pro-teome of NAc (Figure 1 and Table 1). The totalnumbers of protein spots subjected to statisticalanalysis were 1098. The DeCyder image analysis ofthe images representing CSA rhesus monkeys andcontrol NAc cytosol proteome elucidated the putativeCSA-specific protein spots. Quantification of theindividual protein spots revealed that the vastmajority of B1000 gene products relegated to struc-tural and housekeeping functions were quantitativelysimilar. Image analysis (Figure 1) of fluorescentlylabeled NAc cytosol lysates loaded on 2D gelsidentified differential distributions of 22 proteinspots (t-test: P < 0.05) characterized by 18 proteinsin CSA rhesus monkeys (Figures 2 and 3). Notably,the expression of glial fibrillary acidic protein(GFAP, spot 26) was increased significantly in humanNAc cytosol after chronic cocaine exposure asshown in Figure 2. Even though there have beenmany studies reporting the cocaine-induced altera-tions in gene expression, this is the first follow-upstudy which utilizes a noncandidate approach todecipher the involvement of novel proteins andpathways.
A number of proteins extracted from 2D gels wereconstitutively expressed in the control and CSAgroups and identified by MALDI-TOF-MS and -MS/MS. The proteins which were constitutively ex-pressed belonged to the following groups: inter- andintracellular signaling, cell morphology, protein fold-ing and stability, cell proliferation and apoptosis fullyrealizing that such a classification is somewhatarbitrary as a number of proteins could be assignedto more than one functional category. It should benoted that although some of the differentially ex-pressed proteins seen here are already known to beeither directly or indirectly involved in cocaineintake, a number of gene products (for example,syntaxin-binding protein 3 (Figure 2)) with unprece-dented involvement in cocaine intake were alsoidentified in the current study. The more conservativestrategy of comparing results from the decoy andtarget databases resulted in three false-positive pro-tein identifications: actin, cytoplasmic 2 (g)—spots291 and 320, and isocitrate dehydrogenase subunit-a,
mitochondrial precursor-spot 594. The proteins iden-tified from the respective spots were not included forsubsequent analysis.
Pro-Q diamond for staining phosphoproteinsThe specificity, quantitative assessment and thelinear dynamic range of the Pro-Q Diamond stainingfor phosphoproteins has been rigorously establishedpreviously.43–47 The specificity of Pro-Q Diamond wasre-validated in the current study by examining thedifferential staining of known phosphorylated andnonphosphorylated proteins. Standard proteins andNAc cytosolic proteins from control and cocaine-treated monkeys were electrophoresed and the gelsstained with Pro-Q Diamond and Sypro-Ruby. Thestaining of specific proteins, ovalbumin and b-casein,with documented phosphorylation sites was observedin the protein standards stained with Pro-Q Diamond.As anticipated bovine serum albumin, avidin andlysozyme were not stained by Pro-Q Diamond due tothe lack of phosphorylation sites on these proteins.Specific protein band staining from cocaine andcontrol samples was also observed with Pro-QDiamond stain.
NAc phosphoproteomes in cytosolic fractionsTo determine changes in protein phosphorylation inNAc with cocaine self-administration, 2D gels werestained with the Pro-Q Diamond. DeCyder software-based comparisons of gels revealed that 123 and 152spots were resolved in the control and cocainegroups, respectively, and spots were consistentlymatched in at least three of four subjects in eachgroup. DeCyder-based image analysis was performedto quantify staining intensities of the putativephosphoproteins. Representative cytosolic proteinspots with significant differences in phosphorylationabundance between control and cocaine groups areshown in the Figure 4. Targeted phosphoprotein spotswere identified by MALDI-TOF-TOF (Tables 2 and 3).Although the specificity of the Pro-Q Diamond forstaining phosphoproteins has been rigorously estab-lished,43–47 a note of caution in the interpretation ofthese data. The identification of a spot, stained withPro-Q Diamond, as a phosphoprotein by MS may bebiased if an abundant protein (which may or may notbe a phosphoprotein) co-migrates with this particularspot.
Thirty-six proteins, matched in at least three of foursubjects in each group, which were identified by Pro-Q Diamond as phosphorylated proteins betweencontrol and cocaine groups. The proteins were identi-fied by MALDI-TOF MS and were corroborated bypeptide sequencing using MALDI-TOF-TOF for selectpeptides. For example, mu crystallin protein wasidentified by MALDI-MS (Figure 5a) initially andconfirmed by amino-acid sequence analysis of itstryptic peptides by MALDI-MS/MS (Figures 5b andc), unequivocally determining the protein’s identity.
The MS analysis of tryptic digest identified KIAA1258 protein from protein spot 173 and a BLAST
Neuroproteome of cocaine in nonhuman primatesNS Tannu et al
190
Molecular Psychiatry

search of the sequence returned guanine deaminase.The identification was confirmed by the followingreturning parameters for the search: CD-length = 429residues, 100.0% aligned score = 524 bits (1351),expect = 9e-150. The list of 34 phosphoproteinsidentified by 2D-PAGE, MS and bioinformatics meth-ods are presented in Table 2 along with an elaborationof the protein identification parameters, MASCOTscores (independent searches against target and decoydatabases), spot intensity ratios of all the phospho-
proteins and the corresponding P-value. Of the 34proteins identified by PMF, the identities of 15proteins was further confirmed by MALDI-MS/MS.We have attempted to organize the differentiallyexpressed proteins into functional categories, fullyrealizing that such a classification is somewhatarbitrary as proteins could be assigned to more thanone functional category. Comparison of results fromthe decoy and target databases resulted in false-positive protein identification of g-enolase (spot 180).
Figure 1 Two-dimensional difference in gel electrophoresis (2D-DIGE)-based comparison of control and cocaine groups. Arepresentative 2D-DIGE image of cytosolic proteins from the nucleus accumbens (NAc) (a). Proteins were fractionated in thefirst dimension by a 4–7 linear pH gradient (isoelectric points, pI) and in the second dimension by an 8–15% gradient SDS–polyacrylamide gel electrophoresis (PAGE) gel (molecular weight, kDa). Areas of the gel containing spots of interest aremarked with boxes and shown separately for control (b–e) and cocaine groups (b*, c*, d*, e*). The numbered protein spotscorrespond with numbers in Figures 2 and 3, and Table 1.
Neuroproteome of cocaine in nonhuman primatesNS Tannu et al
191
Molecular Psychiatry
Table
1P
rote
ins
iden
tifi
ed
by
MA
LD
I-T
OF
-TO
Ffo
llow
ing
2D
-DIG
Ean
aly
sis
Sp
ot
nos.
Pro
tein
ID(P
MF
)A
ccess
ion
nos.
Mr
(kD
a)
pI
Pep
tid
es
MA
SC
OT
score
Targ
et
con
fid
en
ce
ind
ex
(%)
Pro
tein
qu
an
tita
tion
Targ
et
Decoy
T-t
est
(P-v
alu
e)
Avera
ge
valu
e
Str
uctu
ral
26
Gli
al
fibri
llary
acid
icp
rote
in,
ast
rocyte
gi|
121135
49.8
55.4
28
66
38
98.3
50.0
13
1.5
3
83
Fib
rill
in1
gi|
24430141
31.2
4.8
123
66
38
98.4
0.0
42
1.2
8343
Acti
n,
cyto
pla
smic
2(g
)gi|
15012187
41.7
65.3
17
150
39
100
0.0
32
�1.5
0413
Acti
n,
cyto
pla
smic
2(g
)gi|
15012187
41.7
65.3
19
113
33
100
0.0
45
2.4
5431
Acti
n,
cyto
pla
smic
2(g
)gi|
15012187
41.7
65.3
17
75
37
99.7
0.0
23
1.5
6160
a-T
ubu
lin
gi|
10881132
49.0
34.9
692
35
100
0.2
7�
2.2
789
g-C
ryst
all
inA
gi|
117458
21.1
37.5
36
63
40
96.8
50.0
31.4
4250
Un
nam
ed
pro
tein
pro
du
ct;
Hom
olo
gy-f
ilam
en
t,in
term
ed
iate
fila
men
tp
rote
in
gi|
34536332
49.4
75.8
413
185
49
100
0.0
79
�1.7
4
257
Un
nam
ed
pro
tein
pro
du
ct;
Hom
olo
gy-f
ilam
en
t,in
term
ed
iate
fila
men
tp
rote
in
gi|
34536332
49.4
75.8
416
211
43
100
0.1
9�
1.5
0
254
Un
nam
ed
pro
tein
pro
du
ct;
Hom
olo
gy-f
ilam
en
t,in
term
ed
iate
fila
men
tp
rote
in
gi|
34536332
49.4
75.8
419
261
52
100
0.1
1�
1.6
2
Cell
sign
ali
ng
133
b-S
olu
ble
NS
Fatt
ach
men
tp
rote
ingi|
18202933
33.5
35.3
26
56
37
85.6
0.0
1�
2.1
2673
Syn
taxin
-bin
din
gp
rote
in3
gi|
27881593
67.7
98.1
759
25
95.1
50.0
22
1.3
41029
14-3
-3p
rote
inz/D
(pro
tein
kin
ase
Cin
hib
itor
pro
tein
-1)
gi|
52000886
27.7
24.7
35
51
39
97.5
0.0
041
�1.3
1
35
PK
C,
ep
silo
nty
pe
(nP
KC
-e)
gi|
125556
83.4
66.6
212
66
43
98.5
0.0
21
1.3
2612
DD
AH
1p
rote
ingi|
34783629
27.1
15.2
33
82
38
99.9
0.3
6�
1.1
9
Meta
boli
sm113
Dih
yd
rop
yri
mid
inase
-rela
ted
pro
tein
2gi|
33991634
62.2
55.9
512
83
36
99.9
0.0
085
�1.8
6
118
Dih
yd
rop
yri
mid
inase
-rela
ted
pro
tein
2gi|
33991634
62.2
55.9
511
93
36
100
0.0
072
�1.9
6
126
Dih
yd
rop
yri
mid
inase
-rela
ted
pro
tein
2gi|
33991634
62.2
55.9
511
84
43
100
0.0
3�
1.6
5
148
Ad
en
yla
tekin
ase
isoen
zym
e5
gi|
9296996
22.0
75.3
89
65
98.1
90.0
44
1.5
3173
Vacu
ola
rA
TP
-syn
thase
subu
nit
B,
bra
inis
ofo
rm56/5
8kD
a,V
1su
bu
nit
B,
isofo
rm2
gi|
19913428
56.4
65.5
716
159
41
100
0.0
027
�1.5
0
306
a-E
nola
se(n
on
-neu
ral
en
ola
se)
gi|
119339
47.1
37.0
19
84
27
99.9
0.0
45
�2.3
0S
eri
ne/t
hre
on
ine-p
rote
inkin
ase
MA
K(p
rote
inkin
ase
RC
K)
gi|
462461
70
9.6
78
63
27
96.9
2
Neuroproteome of cocaine in nonhuman primatesNS Tannu et al
192
Molecular Psychiatry

Table
1C
on
tin
ued
Sp
ot
nos.
Pro
tein
ID(P
MF
)A
ccess
ion
nos.
Mr
(kD
a)
pI
Pep
tid
es
MA
SC
OT
score
Targ
et
con
fid
en
ce
ind
ex
(%)
Pro
tein
qu
an
tita
tion
Targ
et
Decoy
T-t
est
(P-v
alu
e)
Avera
ge
valu
e
313
a-E
nola
se(n
on
-neu
ral
en
ola
se)
gi|
119339
47.1
37.0
16
44
35
87.8
0.0
33
�1.7
5323
g-E
nola
se(n
eu
ral
en
ola
se)
(NS
E)
gi|
119348
47.2
64.9
95
46
37
93.1
0.0
46
�1.7
3318
NS
Egi|
930063
47.1
24.9
47
53
36
97.7
0.0
5�
1.3
9319
NS
Egi|
930063
47.1
24.9
46
72
44
99.5
0.5
91.0
9315
Gu
an
ine
deam
inase
gi|
4758426
50.9
75.4
45
71
30
99.2
0.7
0�
1.0
5689
Lacta
ted
eh
yd
rogen
ase
Bgi|
4557032
36.6
15.7
15
78
40
99.8
0.5
2�
1.1
5
Mit
och
on
dri
al
226
AT
Psy
nth
ase
,Hþ
tran
sport
ing,
mit
och
on
dri
al
F1
com
ple
x,b-
subu
nit
pre
cu
rsor
gi|
32189394
56.5
25.2
612
122
35
100
0.0
14
2.3
0
139
Su
ccin
yl-
CoA
:3-k
eto
acid
CoA
tran
sfera
se1
gi|
10280560
56.1
27.1
42
92
38
100
0.2
11.4
1
Mis
cell
an
eou
s190
Heat
shock
70
kD
ap
rote
in8
isofo
rm2
gi|
24234686
53.4
85.6
27
75
37
98.8
20.0
17
1.1
9
327
Bra
incre
ati
ne
kin
ase
gi|
17939433
42.6
25.3
46
88
38
99.9
0.0
47
�1.5
4434
Bra
incre
ati
ne
kin
ase
gi|
21536286
42.6
15.3
49
201
40
100
0.4
3�
1.1
6668
T-c
ell
recep
torb-
ch
ain
pre
cu
rsor
Vre
gio
n(8
.3)
gi|
88712
12.9
86.0
76
59
35
86.9
0.7
51.0
6
Abbre
via
tion
s:A
TP,
ad
en
osi
netr
iph
osp
hate
;D
DA
H1,
dim
eth
yla
rgin
ine
dim
eth
yla
min
oh
yd
rola
se1;
nos.
,n
um
bers
;N
SE
,n
eu
ron
-sp
ecif
icen
ola
se;
NS
F,
N-
eth
ylm
ale
imid
e-s
en
siti
ve
facto
r;p
I,is
oele
ctr
icp
oin
ts;
PK
C,
pro
tein
kin
ase
C;
PM
F,
pep
tid
em
ass
fin
gerp
rin
t.S
tati
stic
all
ysi
gn
ific
an
td
iffe
ren
ces
inp
rote
inabu
nd
an
ce
betw
een
the
gro
up
sare
shad
ed
.M
AS
CO
Tsc
ore
issh
ow
nfo
rth
ese
arc
hes
again
stth
eta
rget
an
dals
oth
ed
ecoy
data
base
s.A
vera
ge
rati
os
are
pro
vid
ed
as
cocain
e:c
on
trol.
Posi
tive
an
dn
egati
ve
rati
os
ind
icate
that
the
pro
tein
abu
nd
an
ce
isgre
ate
rin
the
cocain
egro
up
an
dcon
trol
gro
up
,re
specti
vely
.C
OC
/0re
fers
top
rote
ins
pre
sen
tin
thre
eor
fou
rcocain
esu
bje
cts
an
dn
on
eof
the
con
trol
subje
cts
.0/C
TR
refe
rsto
pro
tein
sp
rese
nt
inth
ree
or
fou
rcon
trol
subje
cts
an
dn
on
eof
the
cocain
esu
bje
cts
.
Neuroproteome of cocaine in nonhuman primatesNS Tannu et al
193
Molecular Psychiatry

As noted previously, such proteins were excludedfrom subsequent analysis.
Discussion
The present study was undertaken to interrogate thecytosolic proteome of the NAc in primate brain as a
function of cocaine intake. Previous studies inrodents and human postmortem tissue have demon-strated that cocaine administration can involvetransitory and also enduring biochemical alterationsin specific brain regions. Furthermore, these changesmay be the seat of persistent drug-seeking behavior,craving and also relapse.3,4 This is the first study toprovide an unbiased simultaneous assessment of
COCCTRL
COCCTRL
COCCTRL
COCCTRLCOCCTRL
COCCTRL
COCCTRL
COCCTRL
-0.25
Sta
ndar
dize
d Lo
gA
bund
ance
Sta
ndar
dize
d Lo
gA
bund
ance
Sta
ndar
dize
d Lo
gA
bund
ance
Sta
ndar
dize
d Lo
gA
bund
ance
Sta
ndar
dize
d Lo
gA
bund
ance
Sta
ndar
dize
d Lo
gA
bund
ance
Sta
ndar
dize
d Lo
gA
bund
ance
Sta
ndar
dize
d Lo
gA
bund
ance
-0.2-0.15-0.1
-0.050
0.050.1
-0.12-0.1
-0.08-0.06-0.04-0.02
00.020.04 Spot 118: Dihydropyrimidinase-related protein 2Spot 35: nPKC-epsilon
-0.3-0.25-0.2
-0.15-0.1
-0.050
0.050.1
0.150.2
Spot 673: Syntaxin binding protein 3
-0.06-0.04-0.02
00.020.040.060.08
0.10.120.14
Spot 1029: KCIP 1
-0.1
-0.05
0
0.05
0.1
Spot 89: Gamma crystallin A
-0.3-0.25
-0.2-0.15
-0.1-0.05
00.05
0.10.15
0.2Spot 133: Beta SNAP
-0.1-0.08-0.06-0.04-0.02
00.020.040.060.080.1
Spot 26: GFAP0.180.160.140.12
0.10.080.060.040.02
0
Spot 83: Fibrillin 1
Figure 2 Differentially expressed proteins between the control and cocaine groups. Each panel depicts differentiallyexpressed proteins as determined by two-dimensional difference in gel electrophoresis (2D-DIGE). Panels show individualdata from the control (CTRL: red circles) and cocaine (COC: blue circles) groups and mean standardized log abundance of thetwo groups (cross symbols with blue line) for differentially expressed proteins. A three-dimensional image of each proteinspot is provided from a representative control and COC subject. All proteins shown are significantly different between thetwo groups (P < 0.05).
Neuroproteome of cocaine in nonhuman primatesNS Tannu et al
194
Molecular Psychiatry

multiple protein abundances and also the post-translational modifications in an animal model ofdrug abuse and compliments a previous studyinvestigating the cytosolic proteome of the NAc incocaine overdose victims.23 The use of proteomictechnology provides a unique format to query nativeand post-translationally modified proteomes anddetermine the proteomic phenotype of cocaine abuse
in the primate brain. As in our previous study,subcellular fractionation enabled the quantificationof low abundance proteins that would not have beenpossible by analysis of whole-cell protein homo-genates. In the present study, DIGE analysis revealedthe differential expression of 18 polypeptides fromseveral protein families including cell structure,synaptic plasticity/signal transduction, metabolism
COCCTRLCOCCTRL
COCCTRL
COCCTRL
COCCTRL
COCCTRL
COCCTRL
COCCTRL
-0.02-0.01
00.010.020.030.040.050.06
Sta
ndar
dize
d Lo
gA
bund
ance
Sta
ndar
dize
d Lo
gA
bund
ance
Sta
ndar
dize
d Lo
gA
bund
ance
Sta
ndar
dize
d Lo
gA
bund
ance
Sta
ndar
dize
d Lo
gA
bund
ance
Sta
ndar
dize
d Lo
gA
bund
ance
Sta
ndar
dize
d Lo
gA
bund
ance
Sta
ndar
dize
d Lo
gA
bund
ance
Spot 190: Heat shock 70 kDa protein 8 isoform 2Spot 226: ATP synthase, H+ transporting, mitochondrial F1
-0.25-0.2
-0.15-0.1
-0.050
0.050.1
0.150.2
0.250.3
-0.25-0.2
-0.15-0.1
-0.050
0.050.1
0.15Spot 318: Neuron-specific enolase
0.15
0.1
0.05
0
-0.05
-0.1
SPot 594: NDA (+)- specific ICDH
-0.3-0.25
-0.2-0.15
-0.1-0.05
00.05
0.10.15
0.20.25
0.3 Spot 306: Alpha enolase & Protein kinase RCKSpot 173: Vacuolar H+ ATPase B2
-0.05
0
0.05
0.1
0.15
-0.3-0.25-0.2
-0.15-0.1
-0.050
0.05
Spot 118: Dihydropyrimidinase-related protein 2
-0.05
0
0.05
0.1
0.15
Spot 148: Adenylate kinase isoenzyme 5
Figure 3 Differentially expressed proteins between the control and cocaine groups. Each panel depicts differentiallyexpressed proteins as determined by two-dimensional difference in gel electrophoresis (2D-DIGE). Panels show individualdata from the control (CTRL: red circles) and cocaine (COC: blue circles) groups and mean standardized log abundance of thetwo groups (cross symbols with blue line) for differentially expressed proteins. A three-dimensional image of each proteinspot is provided from a representative control and cocaine subject. All proteins shown are significantly different between thetwo groups (P < 0.05).
Neuroproteome of cocaine in nonhuman primatesNS Tannu et al
195
Molecular Psychiatry

and mitochondrial function. Concurrent analysisof the phosphorylation modifications to the NAccytosolic proteome of the same subjects revealedsignificant alterations in complimentary proteinsof the aforementioned categories. Notwithstanding,the results of this study provide the first proteomicanalysis of chronic cocaine exposure in nonhumanprimates and a significant extension of our knowledgeof the effects of cocaine in the primate brain.
StructuralThe detected isoform of GFAP was significantlyincreased (53%) in monkeys self-administering co-caine compared to controls. The data compliment aprevious report from this laboratory in which GFAPlevels in the NAc of cocaine overdose victims wereincreased on average 40 and 60% above controlslevels,23,24 suggesting that such an effect in humans ismost likely attributable to the effects of cocaineexposure. GFAP is an intermediate filament proteinspecific to mature astrocytes and elevated levels ofthis protein are indicative of astrocytic activation orastrogliosis. Whereas astrocytic activation is known tocontribute to neuronal plasticity,48 several studieshave suggested that astrocytes may contribute tosynaptic plasticity associated with abused drugs.49,50
For example, previous studies have demonstratedelevated GFAP levels in the NAc, ventral tegmental
area and hippocampus following various withdrawalperiods from cocaine.11,50,51 Alternatively, elevatedGFAP levels may reflect a compensatory response,such as stabilization of astrocytic processes52,53 to theeffects of cocaine administration. Acute and chroniccocaine administration in mice resulted in increasedGFAP immunoreactivity in the dentate gyrus accom-panied by changes in astrocyte proliferation and cellmorphology.54 The biochemical mechanisms involvedin reactive astrocytic gliosis remain to be determined;however, one possible means is activation of dopa-mine and glutamate receptors on astrocytes leading tostimulation of adenylyl cyclase activity55 and subse-quent phosphorylation of transcription factors thatbind to the cAMP response elements located on theGFAP promoter region.56 By means of this interactionbetween astrocytes and neurons,52,57 it is likely thatastrocytic cells actively contribute to long-termneuronal changes in response to psychostimulantdrugs. Whether these changes are indicative ofneurotoxicity or neuroplasticity remains to be deter-mined; however, further understanding of the me-chanisms involved in reactive astrocytic gliosis mayprovide insight into whether such changes areassociated with repair and reversal of neuronaldamage associated with cocaine abuse.
In addition to increased levels of GFAP, cocaine self-administration also resulted in reduced phosphorylation
76
pl
54CocaineControl
CocaineControl
CocaineControl
CocaineControl CocaineControl
Peroxiredoxin 2 isoform b: 293
14-3-3 gamma protein: 256
Neurofilament 68 kDa: 111
Rab GDP dissociation inhibitor alpha: 315 Crystallin, mu: 232
160105
7550
35
kDa
30
25
15
A
Figure 4 Comparison of the phosphoproteins between control and cocaine groups. Representative gel of spots visualizedfollowing Pro-Q Diamond staining identified as differentially abundant between the two groups. Proteins were fractionatedand electrophoresed by a linear 4–7 pH gradient in first dimension and by 12.5% SDS–polyacrylamide gel electrophoresis(PAGE) in second dimension. The identified protein name is followed by the spot number. The numbered protein spotscorrespond with numbers in Tables 2 and 3.
Neuroproteome of cocaine in nonhuman primatesNS Tannu et al
196
Molecular Psychiatry

Table
2P
hosp
ho-p
rote
ins
iden
tifi
ed
by
MA
LD
I-T
OF
-TO
F
Sp
ot
nos.
Pro
tein
ID(P
MF
)A
ccess
ion
nos.
Mr
(kD
a)
pI
Pep
tid
es
MA
SC
OT
score
CI
(%)
Pro
tein
qu
an
tita
tion
Targ
et
Decoy
T-t
est
(P-v
alu
e)
Avera
ge
rati
o
Str
uctu
ral
204
Acti
n,b
Q96H
G5
40.9
85.5
21
243
41
100
0.0
19
�2.0
8205
AC
TG
1p
rote
in(g
-acti
n)
P63261
41.7
65.3
18
185
49
100
0.9
95
1.2
1183
Acti
n,
cyto
pla
smic
1P
60709
41.7
15.3
20
72
43
100
0.3
15
�1.3
0191
Acti
n,
cyto
pla
smic
1P
60709
41.7
5.3
23
116
46
100
0.1
22
5.8
8190
Acti
n,
cyto
pla
smic
1P
60709
41.7
5.3
21
96
35
100
0.9
16
�1.5
6186
Acti
n,
cyto
pla
smic
1P
60709
41.7
5.3
16
102
39
100
0.3
15
�1.3
0193
Acti
n,
cyto
pla
smic
1P
60709
41.7
5.3
16
66
37
100
0.4
95
�1.0
4111
Neu
rofi
lam
en
t,li
gh
tp
oly
pep
tid
e68
kD
aP
07196
61.7
44.7
16
66
39
100
0.0
32
�3.5
7
7N
eu
rofi
lam
en
ttr
iple
tM
pro
tein
160
kD
aP
07197
102.3
84.9
25
91
38
100
0.7
28
�1.5
8
Cell
sign
ali
ng
274
GA
BA
Are
cep
tor-
ass
ocia
ted
pro
tein
1(G
EC
1)
Q9H
0R
814.0
38.7
555
39
100
N/A
CO
C/0
315
Rab
GD
Pd
isso
cia
tion
inh
ibit
or-a
P31150
50.5
55
17
54
41
100
0.0
008
�1.5
4119
Rab
GD
Pd
isso
cia
tion
inh
ibit
or-a
P31150
50.5
55
17
69
38
100
0.1
92
�1.7
2256
14-3
-3g-
pro
tein
P61981
28.3
64.7
13
92
46
99.9
90.0
57
4.7
9255
14-3
-3g-
pro
tein
P61981
28.3
64.7
11
86
37
99.9
9N
/AC
OC
/0264
r-G
DP
dis
socia
tion
inh
ibit
or-a
P52565
23.2
5.0
669
40
100
0.2
41
2.2
2
Meta
boli
sm286
Glu
tath
ion
eS
-tra
nsf
era
sep
i—M
acaca
mu
latt
aQ
28514
23.4
25.9
11
107
46
100
N/A
CO
C/0
267
Glu
tath
ion
eS
-tra
nsf
era
sep
i—h
um
an
P09211
23.3
75.4
754
39
100
N/A
CO
C/0
293
Pero
xir
ed
oxin
2is
ofo
rmb
P32119
159.7
96.1
10
141
44
100
0.0
14
�3.3
7219
Bra
in-t
yp
eald
ola
seP
09972
39.4
26.4
17
92
46
100
N/A
CO
C/0
115
Bra
in-t
yp
eald
ola
seP
09972
39.4
36.4
16
93
44
100
N/A
CO
C/0
245
Dim
eth
yla
rgin
ine
dim
eth
yla
min
oh
yd
rola
se2
Q5S
RR
829.6
35.7
859
38
100
N/A
CO
C/0
172
Gu
an
ine
deam
inase
Q9Y
2T
352.8
5.6
22
95
44
100
N/A
0/C
TR
217
Fru
cto
se-b
isp
hosp
hate
ald
ola
seC
P09972
39.4
36.4
15
82
39
100
0.5
96
�2.3
8
Mit
och
on
dri
al
166
AT
P-s
yn
thase
b-ch
ain
,m
itoch
on
dri
al
pre
cu
rsor
P06576
56.5
25.3
20
103
51
100
0.5
69
�3.6
9
164
AT
P-s
yn
thase
b-ch
ain
,m
itoch
on
dri
al
pre
cu
rsor
P06576
56.5
25.3
15
86
41
100
0.1
65
�1.7
6
325
Ubiq
uit
inC
-term
inal
hyd
roxyla
se12
O75317
24.7
35.3
12
120
44
100
0.3
55
�1.1
9
Neuroproteome of cocaine in nonhuman primatesNS Tannu et al
197
Molecular Psychiatry

states of neurofilament (NF) (light polypeptide68 kDa; �3.57) and b-actin (�2.08). The NF, neuron-specific intermediate filaments (L, M and H corre-sponding to 68, 160 and 200 kDa) regulate neuronalplasticity by affecting the dynamics and function ofcytoskeletal elements such as microtubules and actinthereby regulating neurite outgrowth, axonal caliberand transport. Studies have shown that NR1 and NF-Lare colocalized and NF-L appears to be involved inNMDA receptor anchoring and localization.58 Morerecently, NF-L was demonstrated to increase NMDAreceptor expression and prevent NMDA receptorubiquination.59 Although the role of phosphorylatedNF-L on NMDA receptor expression has not beenstudied, NF-L phosphorylation prevents subsequentNF assembly and induces disassembly in existing NF-L formed filaments.60–62 Nixon and Shiag63 suggestthat in postmitotic cells, NF-L phosphorylationprevents polymerization immediately after synthesis(that is increased turnover)—allowing proper integra-tion of NF-M and -H subunits. Therefore, decreasedNF-L phosphorylation may be expected to increaseNF assembly and stabilize existing filaments—there-by stabilizing NF-L/NR1 interactions. Such an effectwould be parsimonious with our recent studydemonstrating increased NR1 subunit protein levelsin the NAc of cocaine overdose victims and inmonkeys self-administering cocaine and increasedNR1 subunit phosphorylation in these monkeys aswell.24 The lack of detectable change in NF-Hphosphorylation is not surprising given that thisisoform is the most extensively phosphorylatedprotein in brain.64 Nonetheless, decreased phosphor-ylation of NF-L and its known interaction withNMDA receptors provides a mechanism for increasedmembrane stabilization of NMDA receptors that wehave observed following chronic cocaine administra-tion in the NAc of human and nonhuman primatebrain.24
MetabolismSeveral metabolic proteins were identified as differ-entially expressed in the NAc following cocaine self-administration subjects in the present study, ofparticular interest are the proteins involved in theglycolytic pathway and those involved in oxidativeregulation (Table 1). Glucose is the principal source ofenergy for the brain and the oxidation of glucose topyruvic acid through the glycolytic pathway gener-ates adenosinetriphosphate (ATP) and nicotinamideadenine dinucleotide, sources of energy for cellularfunction and culminates in the production of pyr-uvate, which is used in the citric acid cycle foraerobic respiration. Brain-type aldolase C (ALDOC) isinvolved in the preparatory stage of glycolysis con-verting fructose 1,6-biphosphate to glyceraldehyde-3phosphate. Increased levels of phospho-ALDOC ob-served in the present study suggest a means ofenzyme activation that would catalyze the abovereaction to produce increased amounts of glyceralde-hyde-3 phosphate for the energy-yielding phase of theT
able
2C
on
tin
ued
Sp
ot
nos.
Pro
tein
ID(P
MF
)A
ccess
ion
nos.
Mr
(kD
a)
pI
Pep
tid
es
MA
SC
OT
score
CI
(%)
Pro
tein
qu
an
tita
tion
Targ
et
Decoy
T-t
est
(P-v
alu
e)
Avera
ge
rati
o
Mis
cell
an
eou
s232
Cry
stall
in,m
Q14894
33.7
55.1
8140
43
100
0.0
012
1.6
8173
KIA
A1258
pro
tein
Q9U
LG
253.4
5.5
17
91
41
100
0.4
44
�1.1
5214
GL
UL
pro
tein
P15104
42.1
16.4
15
55
100
0.3
51
�1.4
4197
Cre
ati
ne
kin
ase
B-t
yp
eP
12277
42.5
95.3
25
149
39
100
0.1
87
�2.1
1196
Cre
ati
ne
kin
ase
,bra
inQ
6F
G40
42.6
15.3
14
113
43
100
0.7
53
1.4
3114
Heat-
shock
60
kD
pro
tein
1P
10809
61.0
15.7
25
142
46
100
0.1
92
�1.7
2331
60
kD
ah
eat-
shock
pro
tein
,m
itoch
on
dri
al
pre
cu
rsor
(Hsp
60)
P10809
61.0
15.7
17
66
35
100
0.7
30
1.2
6
101
Heat-
shock
70
kD
ap
rote
in8
isofo
rm1
Q53G
Z6
70.8
55.4
28
118
53
100
0.2
49
�1.4
8
Abbre
via
tion
s:A
CT
G1,g-
acti
n1;
CI,
con
fid
en
ce
inte
rval;
GA
BA
A,g-
am
inobu
tyri
c-a
cid
typ
eA
;n
os.
,n
um
bers
;p
I,is
oele
ctr
icp
oin
ts;
PM
F,
pep
tid
em
ass
fin
gerp
rin
t.S
tati
stic
all
ysi
gn
ific
an
td
iffe
ren
ces
inp
rote
inabu
nd
an
ce
betw
een
the
gro
up
sare
shad
ed
.M
AS
CO
Tsc
ore
issh
ow
nfo
rth
ese
arc
hes
again
stth
eta
rget
an
dals
oth
ed
ecoy
data
base
s.A
vera
ge
rati
os
are
pro
vid
ed
as
cocain
e:c
on
trol.
Posi
tive
an
dn
egati
ve
rati
os
ind
icate
that
the
pro
tein
abu
nd
an
ce
isgre
ate
rin
the
cocain
egro
up
an
dcon
trol
gro
up
,re
specti
vely
.C
OC
/0re
fers
top
rote
ins
pre
sen
tin
thre
eor
fou
rcocain
esu
bje
cts
an
dn
on
eof
the
con
trol
subje
cts
.0/C
TR
refe
rsto
pro
tein
sp
rese
nt
inth
ree
or
fou
rcon
trol
subje
cts
an
dn
on
eof
the
cocain
esu
bje
cts
.
Neuroproteome of cocaine in nonhuman primatesNS Tannu et al
198
Molecular Psychiatry

Table
3P
hosp
ho-p
rote
ins
iden
tifi
ed
by
am
ino
acid
sequ
en
cin
gu
sin
gM
AL
DI-
TO
F-M
S/M
S
Sp
ot
#P
rote
inID
(MS
MS
)A
ccess
ion
#P
ep
tid
ese
qu
en
ce
Pre
cu
rsor
mass
Mass
err
or
(pp
m)
MA
SC
OT
score
Con
fid
en
ce
inte
rval
(%)
101
Heat
shock
70
kD
ap
rote
in8
isofo
rm1
Q53G
Z6
FE
EL
NA
DL
FR
1253.6
162
029
97.9
5T
VT
NA
VV
TV
PA
YF
ND
SQ
R1981.9
982
035
99.4
9
114
Heat
shock
60
kD
pro
tein
1P
10809
AA
VE
EG
IVL
GG
GC
AL
LR
1684.9
053
085
100
LV
QD
VA
NN
TN
EE
AG
DG
TT
TA
TV
LA
R2560.2
588
429
98.4
9
166
AT
Psy
nth
ase
beta
ch
ain
,m
itoch
on
dri
al
pre
cu
rsor
P06576
AIA
EL
GIY
PA
VD
PL
DS
TS
R1988.0
354
142
99.9
1
172
Gu
an
ine
deam
inase
Q9Y
2T
3F
QN
IDF
AE
EV
YT
R1631.7
726
248
99.9
8
196
Bra
incre
ati
ne
kin
ase
Q6F
G40
DL
FD
PII
ED
R1232.6
084
638
99.6
6V
LT
PE
LY
AE
LR
1303.7
258
034
99.2
6
197
Bra
incre
ati
ne
kin
ase
P12277
VLT
PE
LY
AE
LR
1303.7
256
054
99.9
8L
AV
EA
LS
SL
DG
DL
AG
R1586.8
346
233
98.6
9
204
Acti
n,
beta
Q96H
G5
QE
YD
ES
GP
SIV
HR
1516.6
962
473
100
SY
EL
PD
GQ
VIT
IGN
ER
1790.8
821
564
99.9
9V
AP
EE
HP
VL
LT
EA
PL
NP
K1954.0
564
449
99.9
6
205
AC
TG
1p
rote
inP
63261
QE
YD
ES
GP
SIV
HR
1516.6
975
348
99.9
7S
YE
LP
DG
QV
ITIG
NE
R1790.8
89
272
100
232
Cry
stall
in,
mu
Q14894
LV
TF
YE
DR
1042.5
265
635
99.3
1V
PA
FL
SA
AE
VE
EH
LR
1667.8
766
197
100
255
14-3
-3gam
ma
pro
tein
P61981
NV
TE
LN
EP
LS
NE
ER
1643.7
914
357
100
264
Rh
oG
DI
alp
ha
P52565
SIQ
EIQ
EL
DK
DD
ES
LR
1917.9
427
135
99.1
7
267
Glu
tath
ion
eS
-tra
nsf
era
sep
iH
um
an
P09211
PP
YT
VV
YF
PV
R1337.7
222
266
100
286
Glu
tath
ion
eS
-tra
nsf
era
sep
im
acaca
mu
latt
aQ
28514
PP
YT
VV
YF
PV
R1337.7
238
131
98.6
3
FQ
DG
DLT
LY
QS
NT
FL
R1917.9
314
129
97.8
8
293
Pero
xir
ed
oxin
2is
ofo
rmb
P32119
QIT
VN
DL
PV
GR
1211.6
733
149
99.9
7K
EG
GL
GP
LN
IPL
LA
DV
TR
1863.0
67
229
97.7
8
325
Ubiq
uit
incarb
oxyl-
term
inal
est
era
seL
1O
75317
NE
AIQ
AA
HD
AV
AQ
EG
QC
R1967.8
951
3101
100
Neuroproteome of cocaine in nonhuman primatesNS Tannu et al
199
Molecular Psychiatry

glycolytic pathway. Interestingly, we found signifi-cant downregulation of two members of the enolasefamily, the family of enzymes involved in thecatalysis of phosphoglycerate to phosphoenolpyru-vate in the energy-yielding phase of the glycolyticpathway. Levels of g-enolase, also known as neuron-specific enolase (NSE) and a-enolase, ubiquitouslyexpressed in cell cytoplasm, were significantly de-creased in the NAc—in contrast to previous findingsin human cocaine overdose victims.23 NSE formshomodimers or heterodimers with a-enolase. Thereductions in NSE and a-enolase levels may representa compensatory mechanism to elevated ALDOC tocontrol glucose utilization in this brain region. Theseresults support the results of a previous studydemonstrating decreased glucose utilization in theNAc of rhesus monkeys self-administering cocaine65
and may provide a potential mechanism of thiseffect. Although not statistically significant, thephosphorylation status of NSE was decreased 1.9-fold in the same monkeys, further suggesting reducedglycolysis in this region following cocaine self-administration.
The phosphorylation status of two proteins in-volved in oxidative metabolism and identified asdifferentially expressed in the NAc of monkeys self-administering cocaine were PRDX2 (�3.37) andglutathione S-transferase pi (GSTP1, exclusively ex-pressed in cocaine group). PRDX2 is a highlyabundant cytosolic protein primarily involved inredox regulation, which is accomplished by decreas-ing intracellular peroxides with reducing equivalentsfrom the thioredoxin system. PRDX2 is expressed inneurons, specifically those vulnerable to oxidativestress,66 and functions as the primary regulator ofH2O2 generated by cell surface receptors,67 therebyprotecting protein and lipids against oxidative stress68
and regulating apoptosis through peroxide elimina-tion.69 Interestingly, cocaine administration in ro-dents increases lipid peroxidation,70 and also thepresence of reactive oxygen species in striatum andfrontal cortex71—both of which indicate the like-lihood of H2O2 accumulation and resultant oxidativestress. The present finding of decreased phosphoryla-tion of PRDX2 b-isoform in monkeys corroborates ourprevious report of decreased protein expression of thePRDX2 a-isoform in the NAc of human cocaineoverdose victims.23
Increased levels of phosphorylated GSTP1 isoformmay compensate for the reduced phosphorylation ofPRDX2. GST activation is one of the principalmechanisms of cellular detoxification catalyzing theconjugation of hydrophobic electrophilic compoundswith reduced glutathione and thereby countering theeffect of reactive oxygen metabolites.72 Thus, in-creased phosphorylation of GSTP1 in the NAc ofmonkeys self-administering cocaine may be a meansof protecting cells against potential oxidative damageinduced by cocaine. These data compliment a recentstudy demonstrating increased frequency of a splicevariant of GSTP1, which confers increased enzymatic
0
500
1000
1500
2000
Inte
nsity
Inte
nsity
Inte
nsity
0
500
1000
1500
2000
2500
3000
0
2000
4000
6000
8000
1000800600Mass (m/z)
Mass (m/z)
400200
1600140012001000800600400200
30002500200015001000
Mass (m/z)
MALDI-TOF-MS of Crystallin, mu
1667.8
*
*1042.5
1535.71681.8** * * * *
1973.92176.1
2256.1 2970.5
MALDI-TOF-MS/MS of [4-18] VPAFLSAAEVEHLR(m/z: 1667.8)
H
P
a2
b3
b2
y1
y3
y2
R
y3-NH3a4 b4
y4
y6
y5 y9 y10 y13
LF
L, a1
y1
PY
F
DT
MALDI-TOF-MS/MS of [76-83] LVTFYEDR(m/z: 1045.5)
a2
b2
y1-NH3
y2b3
y2-NH3y3 y4-NH3
y5
Figure 5 Mass spectrometry identification of mu crystal-lin. (a) Peptide mass fingerprint of mu crystallin withpeptides marked with ‘*’ matched by MASCOT searchagainst the National Center for Biotechnology Information(NCBI) primate database. The x and y axes show the mass/charge (m/z) ratio and the % abundance of the trypticpeptide fragments, respectively. (b) Matrix-assisted laser-desorption ionization time-of-flight tandem mass spectro-metry (MALDI-TOF-MS/MS) analysis of peptide fragmentfrom mu crystallin with m/z of 1667.8 was sequenced asVPAFLSAAEVEEHLR. The corresponding y- and b-seriesions and also the immonium ions are shown. (c) A similarMALDI-TOF-MS/MS analysis of peptide fragment with m/z1042.5 was sequenced as LVTFYEDR.
Neuroproteome of cocaine in nonhuman primatesNS Tannu et al
200
Molecular Psychiatry

activity and is correlated with cocaine dependence.73
Together, these results indicate that glutathioneconjugation is likely a predominant mechanism toattenuate oxidative stress in the NAc induced bycocaine.
Cell signalingIn the present study, four proteins were identified assignificantly differentially expressed following co-caine self-administration: b-soluble N-ethylmalei-mide-sensitive factor (NSF) attachment protein (b-SNAP; �2.12), syntaxin-binding protein 3 (þ 1.34),14-3-3 protein z/D (�1.31) and protein kinase Cepsilon isoform (þ1.32). b-SNAP is one of threesoluble SNAPs, the b-form of which is brain speci-fic.74 Targets of NSF-SNAP complexes include SNAPreceptors such as synaptobrevin 2, syntaxin, SNAP25,and so on. Disassembly of SNARE complexes on themembrane surface is driven by an ATP-dependentprocess that requires the NSF-SNAP complex.75 Inaddition to this more classical function, two addi-tional targets of NSF-SNAP include one in whichtargets bind directly to NSF (for example, GluR2) andalso targets which appear to be isoform specific suchas the binding of b-SNAP to synaptotagmin76 or a- andb-SNAP to Pick1 complexed with GluR2.77 Theinteraction of b-SNAP with GluR2 is particularlyintriguing given that previously we have demon-strated increased GluR2 protein levels in membranefractions from the NAc of monkeys self-administeringcocaine.24 Hanley et al.24 have proposed a model ofNSF-SNAP regulation of Pick1-AMPA receptor inter-actions in AMPA receptor cycling. GluR2 subunitsthat are bound to Pick1 are contained in a receptorpool in which low levels of a/b-SNAP bindingdisassemble the GluR2/Pick1 complex rendering aportion of GluR2 subunits available for anchoring tothe membrane by ABP/GRIP and conversely; highlevels of b-SNAP inhibit the NSF-mediated disrup-tion of GluR2/Pick1 interactions leading to internali-zation of the receptor. Thus, the present findings ofdownregulation of b-SNAP suggests reduced GluR2endocytosis in the cocaine monkeys, which may leadto increased levels of the GluR2 subunit at themembrane surface, as reported previously.24
These data are complimented by the identificationof two proteins whose phosphorylation status wasaltered following cocaine self-administration. g-Ami-nobutyric-acid type A (GABAA) receptor-associatedprotein-like 1 (GABARAPL1/GEC1) was recentlyidentified as a member of the protein family thatincludes GABARAP—sharing 87% identity and 96%similarity. Like GABARAP, GEC1 binds tubulin andexhibits similar subcellular localization.78 GABARAPand GEC1 are biochemical linkers between theGABAA receptor and microtubules and contribute tothe intracellular trafficking and membrane insertionof GABAA receptors thereby important in synapticplasticity.78,79 Given the prevalence of GABAergicsynapses in the NAc and their role in the beha-vioral effects of cocaine, further studies evaluating
the mechanisms of GABAA subunit trafficking follow-ing cocaine administration are warranted.
Rab proteins are a group of membrane-associatedproteins that localize to discrete subcellular compart-ments and are found in two conformations: bound toGTP in association with membrane and bound to GDPin association with guanine-dissociation inhibitor(GDI). Rab proteins exert their effects on the secretorypathway in which proteins bind Rab proteins speci-fically in the GTP-bound state.80 In the current study,the phosphorylated form of Rab GDIa was found to bedecreased in NAc following cocaine self-administra-tion. Rab GDIa is found predominantly in neural andsensory tissues and acts as a general regulator of allRab small G proteins implicated in intracellularvesicle trafficking.81,82 Rab GDIa regulates the GDP/GTP exchange reaction of most Rab proteins byinhibiting the dissociation of GDP and the subsequentbinding of GTP, implicated in neurotransmission.83 Inaddition, Rab GDIa has an important in Rab3Arecycling to suppress hyperexcitability through mod-ulation of presynaptic forms of plasticity in thehippocampus.84 In the NAc, increased phosphoryla-tion of Rab GDIa may serve as a compensatorymechanism to offset hyperexcitability induced byionotropic glutamate receptors and increased phos-phorylation of NR1 following cocaine self-adminis-tration in primates.24
Conclusion
In our continuing effort to understand the neurobiol-ogy of cocaine addiction, the current study delineatesa preliminary functional proteome of the NAc inrhesus monkeys following chronic self-administra-tion of cocaine and compliments a recent proteomicstudy of the NAc in human cocaine overdosevictims.23 The current study documents quantitativedifferences in expression and differential phosphor-ylation of several proteins in the NAc followingcocaine self-administration including those involvedin cytoskeleton structure, mitochondrial function,energy metabolism and cell signaling. The use of anonhuman primate model of human cocaine intake tostudy proteomic alterations associated with long-termcocaine use and abuse offers new perspectives on theassociated neuropathology and may provide targetsfor functional assessments in other models. Determi-nation of the orchestrated changes in proteins, relatedpost-translational modifications and biochemicalpathways altered by chronic cocaine intake is essen-tial for providing a comprehensive understanding ofthe neuropathophysiology associated with drug abuseand necessary for the development of novel pharma-cotherapies targeting these proteins.
Acknowledgments
This work was supported by National Institutes ofHealth grants P50 DA006634 (SEH), DA003628 (SEH),DA012498 (SEH), DA016589 (LLH), DA000517 (LLH)
Neuroproteome of cocaine in nonhuman primatesNS Tannu et al
201
Molecular Psychiatry

and RR000165. We are thankful to Dr Jim Smith foreditorial suggestions.
References
1 SAMHSA. Results from the 2006 National Survey on Drug Use andHealth: National Findings. Substance Abuse and Mental HealthServices Administration: Rockville, MD, 2007.
2 WHO. Neuroscience of Psychoactive Substance Use and Depen-dence. World Health Organization: Geneva, 2004.
3 Koob GF, Le Moal M. Drug addiction, dysregulation of reward, andallostasis. Neuropsychopharmacology 2001; 24: 97–129.
4 Nestler EJ. Molecular basis of long-term plasticity underlyingaddiction. Nat Neurosci Rev 2001; 2: 119–128.
5 Miserendino MJ, Nestler EJ. Behavioral sensitization to cocaine:modulation by the cyclic AMP system in the nucleus accumbens.Brain Res 1995; 674: 299–306.
6 Self DW, Genova LM, Hope BT, Barnhart WJ, Spencer JJ, Nestler EJ.Involvement of cAMP-dependent protein kinase in the nucleusaccumbens in cocaine self-administration and relapse of cocaine-seeking behavior. J Neurosci 1998; 18: 1848–1859.
7 Pliakas AM, Carlson RR, Neve RL, Konradi C, Nestler EJ,Carlezon Jr WA. Altered responsiveness to cocaine and increasedimmobility in the forced swim test associated with elevated cAMPresponse element-binding protein expression in nucleus accum-bens. J Neurosci 2001; 21: 7397–7403.
8 Carlezon Jr WA, Thome J, Olson VG, Lane-Ladd SB, Brodkin ES,Hiroi N et al. Regulation of cocaine reward by CREB. Science 1998;282: 2272–2275.
9 Hope B, Kosofsky B, Hyman SE, Nestler EJ. Regulation ofimmediate early gene expression and AP-1 binding in the ratnucleus accumbens by chronic cocaine. Proc Natl Acad Sci USA1992; 89: 5764–5768.
10 Hiroi N, Brown JR, Haile CN, Ye H, Greenberg ME, Nestler EJ. FosBmutant mice: loss of chronic cocaine induction of Fos-relatedproteins and heightened sensitivity to cocaine’s psychomotor andrewarding effects. Proc Natl Acad Sci USA 1997; 94: 10397–10402.
11 Haile CN, Hiroi N, Nestler EJ, Kosten TA. Differential behavioralresponses to cocaine are associated with dynamics of mesolimbicdopamine proteins in Lewis and Fischer 344 rats. Synapse 2001;41: 179–190.
12 White FJ, Hu XT, Zhang XF, Wolf ME. Repeated administration ofcocaine or amphetamine alters neuronal responses to glutamate inthe mesoaccumbens dopamine system. J Pharmacol Exp Ther1995; 273: 445–454.
13 Hemby SE, Horman B, Tang W. Differential regulation ofionotropic glutamate receptor subunits following cocaine self-administration. Brain Res 2005; 1064: 75–82.
14 Tang W, Wesley M, Freeman WM, Liang B, Hemby SE. Alterationsin ionotropic glutamate receptor subunits during binge cocaineself-administration and withdrawal in rats. J Neurochem 2004; 89:1021–1033.
15 Albertson DN, Pruetz B, Schmidt CJ, Kuhn DM, Kapatos G,Bannon MJ. Gene expression profile of the nucleus accumbens ofhuman cocaine abusers: evidence for dysregulation of myelin.J Neurochem 2004; 88: 1211–1219.
16 Tang WX, Fasulo WH, Mash DC, Hemby SE. Molecular profilingof midbrain dopamine regions in cocaine overdose victims.J Neurochem 2003; 85: 911–924.
17 Hemby SE. Morphine-induced alterations in gene expression ofcalbindin immunopositive neurons in nucleus accumbens shelland core. Neuroscience 2004; 126: 689–703.
18 Ang E, Chen J, Zagouras P, Magna H, Holland J, Schaeffer E et al.Induction of nuclear factor-kappaB in nucleus accumbens bychronic cocaine administration. J Neurochem 2001; 79: 221–224.
19 Yuferov V, Nielsen D, Butelman E, Kreek MJ. Microarray studies ofpsychostimulant-induced changes in gene expression. Addict Biol2005; 10: 101–118.
20 Ahmed SH, Lutjens R, van der Stap LD, Lekic D, Romano-Spica V,Morales M et al. Gene expression evidence for remodeling oflateral hypothalamic circuitry in cocaine addiction. Proc NatlAcad Sci USA 2005; 102: 11533–11538.
21 Backes E, Hemby SE. Discrete cell gene profiling of ventraltegmental dopamine neurons after acute and chronic cocaine self-administration. J Pharmacol Exp Ther 2003; 307: 450–459.
22 Hemby SE. Assessment of genome and proteome profiles incocaine abuse. Prog Brain Res 2006; 158: 173–195.
23 Tannu N, Mash DC, Hemby SE. Cytosolic proteomic alterations inthe nucleus accumbens of cocaine overdose victims. Mol Psychia-try 2007; 12: 55–73.
24 Hemby SE, Tang W, Muly EC, Kuhar MJ, Howell L, Mash DC.Cocaine-induced alterations in nucleus accumbens ionotropicglutamate receptor subunits in human and non-human primates.J Neurochem 2005; 95: 1785–1793.
25 Zolnierowicz S, Bollen M. Protein phosphorylation and proteinphosphatases. De Panne, Belgium, September 19–24, 1999. EMBOJ 2000; 19: 483–488.
26 Yamamoto KK, Gonzalez GA, Biggs III WH, Montminy MR.Phosphorylation-induced binding and transcriptional efficacy ofnuclear factor CREB. Nature 1988; 334: 494–498.
27 Gonzalez GA, Yamamoto KK, Fischer WH, Karr D, Menzel P,Biggs III W et al. A cluster of phosphorylation sites on the cyclicAMP-regulated nuclear factor CREB predicted by its sequence.Nature 1989; 337: 749–752.
28 Creely H, Khaitovich P. Human brain evolution. In: Hemby SE,Bahn S (eds). Progress in Brain Research: Functional Genomicsand Proteomics in the Clinical Neurosciences. Elsevier:Amsterdam, 2006, pp 295–309.
29 Matsumoto I, Alexander-Kaufman K, Iwazaki T, Kashem MA,Matsuda-Matsumoto H. CNS proteomes in alcohol and drug abuseand dependence. Expert Rev Proteomics 2007; 4: 539–552.
30 Howell LL, Wilcox KM. Intravenous drug self-administration innonhuman primates. In: JJ B (ed). Methods of Behavior Analysis inNeuroscience. CRC Press: Boca Raton, FL, 2001, pp 91–110.
31 Byrd LD. The behavioral effects of cocaine: rate dependency orrate constancy. Eur J Pharmacol 1979; 56: 355–362.
32 Lindsey KP, Wilcox KM, Votaw JR, Goodman MM, Plisson C,Carroll FI et al. Effects of dopamine transporter inhibitorson cocaine self-administration in rhesus monkeys: relationshipto transporter occupancy determined by positron emissiontomography neuroimaging. J Pharmacol Exp Ther 2004; 309:959–969.
33 Paxinos G, Huang X-F, Toga A. The Rhesus Monkey Brain in StereotaxicCoordinates, 1st edn. Academic Press: San Diego, CA, 1999.
34 Tang W-X, Fasulo WH, Mash DC, Hemby SE. Molecular profilingof midbrain dopamine regions in cocaine overdose victims.J Neurochem 2003; 85: 911–924.
35 Churchward MA, Butt RH, Lang JC, Hsu KK, Coorssen JR.Enhanced detergent extraction for analysis of membrane pro-teomes by two-dimensional gel electrophoresis. Proteome Sci2005; 3: 5.
36 Tannu NS, Hemby SE. Methods for proteomics in neuroscience.In: Hemby SE, Bahn S (eds). Progress in Brain Research:Functional Genomics and Proteomics in the Clinical Neuros-ciences. Elsevier: Amsterdam, 2006, pp 41–82.
37 Tannu N, Hemby SE. Two dimensional fluorescence difference gelelectrophoresis (2D-DIGE) for comparative proteomics profiling.Nat Protoc 2006; 1: 1732–1742.
38 Henzel WJ, Billeci TM, Stults JT, Wong SC, Grimley C,Watanabe C. Identifying proteins from two-dimensional gels bymolecular mass searching of peptide fragments in proteinsequence databases. Proc Natl Acad Sci USA 1993; 90: 5011–5015.
39 Mann M, Wilm M. Error-tolerant identification of peptides insequence databases by peptide sequence tags. Anal Chem 1994;66: 4390–4399.
40 Yates III JR. Database searching using mass spectrometry data.Electrophoresis 1998; 19: 893–900.
41 Elias JE, Gygi SP. Target-decoy search strategy for increasedconfidence in large-scale protein identifications by mass spectro-metry. Nat Methods 2007; 4: 207–214.
42 Tannu NS, Hemby SE. De novo protein sequence analysis ofMacaca mulatta. BMC Genomics 2007; 8: 270.
43 Martin K, Steinberg TH, Cooley LA, Gee KR, Beechem JM, PattonWF. Quantitative analysis of protein phosphorylation status andprotein kinase activity on microarrays using a novel fluorescentphosphorylation sensor dye. Proteomics 2003; 3: 1244–1255.
Neuroproteome of cocaine in nonhuman primatesNS Tannu et al
202
Molecular Psychiatry

44 Steinberg TH, Agnew BJ, Gee KR, Leung WY, Goodman T,Schulenberg B et al. Global quantitative phosphoprotein analysisusing multiplexed proteomics technology. Proteomics 2003; 3:1128–1144.
45 Schulenberg B, Goodman TN, Aggeler R, Capaldi RA, Patton WF.Characterization of dynamic and steady-state protein phosphory-lation using a fluorescent phosphoprotein gel stain and massspectrometry. Electrophoresis 2004; 25: 2526–2532.
46 Tannu NS, Rao VK, Chaudhary RM, Giorgianni F, Saeed AE, Gao Yet al. Comparative proteomes of the proliferating C(2)C(12)myoblasts and fully differentiated myotubes reveal the complexityof the skeletal muscle differentiation program. Mol Cell Proteomics2004; 3: 1065–1082.
47 Steinberg TH, Agnew BJ, Gee KR, Leung WY, Goodman T,Schulenberg B et al. Global quantitative phosphoprotein analysisusing multiplexed proteomics technology. Proteomics 2003; 3:1128–1144.
48 Ullian EM, Christopherson KS, Barres BA. Role for glia insynaptogenesis. Glia 2004; 47: 209–216.
49 Narita M, Miyatake M, Shibasaki M, Shindo K, Nakamura A,Kuzumaki N et al. Direct evidence of astrocytic modulation in thedevelopment of rewarding effects induced by drugs of abuse.Neuropsychopharmacology 2006; 31: 2476–2488.
50 Bowers MS, Kalivas PW. Forebrain astroglial plasticity is inducedfollowing withdrawal from repeated cocaine administration. Eur JNeurosci 2003; 17: 1273–1278.
51 Beitner-Johnson D, Guitart X, Nestler EJ. Glial fibrillary acidicprotein and the mesolimbic dopamine system: regulation bychronic morphine and Lewis-Fischer strain differences in the ratventral tegmental area. J Neurochem 1993; 61: 1766–1773.
52 Haydon PG. Glia: listening and talking to the synapse. Nat RevNeurosci 2001; 2: 185–193.
53 Weinstein DE, Shelanski ML, Liem RK. Suppression by antisensemRNA demonstrates a requirement for the glial fibrillary acidicprotein in the formation of stable astrocytic processes in responseto neurons. J Cell Biol 1991; 112: 1205–1213.
54 Fattore L, Puddu MC, Picciau S, Cappai A, Fratta W, Serra GP et al.Astroglial in vivo response to cocaine in mouse dentate gyrus:a quantitative and qualitative analysis by confocal microscopy.Neuroscience 2002; 110: 1–6.
55 Porter JT, McCarthy KD. Astrocytic neurotransmitter receptorsin situ and in vivo. Prog Neurobiol 1997; 51: 439–455.
56 Laping NJ, Teter B, Nichols NR, Rozovsky I, Finch CE. Glialfibrillary acidic protein: regulation by hormones, cytokines, andgrowth factors. Brain Pathol 1994; 4: 259–275.
57 Fields RD, Stevens-Graham B. New insights into neuron-gliacommunication. Science 2002; 298: 556–562.
58 Ehlers MD, Fung ET, O’Brien RJ, Huganir RL. Splice variant-specific interaction of the NMDA receptor subunit NR1 withneuronal intermediate filaments. J Neurosci 1998; 18: 720–730.
59 Ratnam J, Teichberg VI. Neurofilament-light increases the cellsurface expression of the N-methyl-D-aspartate receptor andprevents its ubiquitination. J Neurochem 2005; 92: 878–885.
60 Nakamura Y, Takeda M, Angelides KJ, Tanaka T, Tada K,Nishimura T. Effect of phosphorylation on 68 KDa neurofilamentsubunit protein assembly by the cyclic AMP dependent proteinkinase in vitro. Biochem Biophys Res Commun 1990; 169:744–750.
61 Gill SR, Wong PC, Monteiro MJ, Cleveland DW. Assemblyproperties of dominant and recessive mutations in the smallmouse neurofilament (NF-L) subunit. J Cell Biol 1990; 111:2005–2019.
62 Sihag RK, Nixon RA. Identification of Ser-55 as a major proteinkinase A phosphorylation site on the 70-kDa subunit of neuro-filaments. Early turnover during axonal transport. J Biol Chem1991; 266: 18861–18867.
63 Nixon RA, Sihag RK. Neurofilament phosphorylation: a new look atregulation and function. Trends Neurosci 1991; 14: 501–506.
64 Petzold A. Neurofilament phosphoforms: surrogate markersfor axonal injury, degeneration and loss. J Neurol Sci 2005; 233:183–198.
65 Porrino LJ, Lyons D, Smith HR, Daunais JB, Nader MA. Cocaineself-administration produces a progressive involvement of limbic,association, and sensorimotor striatal domains. J Neurosci 2004;24: 3554–3562.
66 Sarafian TA, Verity MA, Vinters HV, Shih CC, Shi L, Ji XD et al.Differential expression of peroxiredoxin subtypes in human braincell types. J Neurosci Res 1999; 56: 206–212.
67 Rhee SG, Chae HZ, Kim K. Peroxiredoxins: a historical overview andspeculative preview of novel mechanisms and emerging concepts incell signaling. Free Radic Biol Med 2005; 38: 1543–1552.
68 Yim MB, Chae HZ, Rhee SG, Chock PB, Stadtman ER. On theprotective mechanism of the thiol-specific antioxidant enzymeagainst the oxidative damage of biomacromolecules. J Biol Chem1994; 269: 1621–1626.
69 Kim H, Lee TH, Park ES, Suh JM, Park SJ, Chung HK et al. Role ofperoxiredoxins in regulating intracellular hydrogen peroxide andhydrogen peroxide-induced apoptosis in thyroid cells. J Biol Chem2000; 275: 18266–18270.
70 Kloss MW, Rosen GM, Rauckman EJ. Biotransformation ofnorcocaine to norcocaine nitroxide by rat brain microsomes.Psychopharmacology (Berl) 1984; 84: 221–224.
71 Dietrich JB, Mangeol A, Revel MO, Burgun C, Aunis D, Zwiller J.Acute or repeated cocaine administration generates reactiveoxygen species and induces antioxidant enzyme activity indopaminergic rat brain structures. Neuropharmacology 2005; 48:965–974.
72 Landi S. Mammalian class theta GST and differential suscepti-bility to carcinogens: a review. Mutat Res 2000; 463: 247–283.
73 Guindalini C, O’Gara C, Laranjeira R, Collier D, Castelo A, ValladaH et al. A GSTP1 functional variant associated with cocainedependence in a Brazilian population. Pharmacogenet Genomics2005; 15: 891–893.
74 Whiteheart SW, Griff IC, Brunner M, Clary DO, Mayer T,Buhrow SA et al. SNAP family of NSF attachment proteinsincludes a brain-specific isoform. Nature 1993; 362: 353–355.
75 Jahn R, Lang T, Sudhof TC. Membrane fusion. Cell 2003; 112:519–533.
76 Schiavo G, Gmachl MJ, Stenbeck G, Sollner TH, Rothman JE. Apossible docking and fusion particle for synaptic transmission.Nature 1995; 378: 733–736.
77 Hanley JG, Khatri L, Hanson PI, Ziff EB. NSF ATPase and alpha-/beta-SNAPs disassemble the AMPA receptor-PICK1 complex.Neuron 2002; 34: 53–67.
78 Mansuy V, Boireau W, Fraichard A, Schlick JL, Jouvenot M,Delage-Mourroux R. GEC1, a protein related to GABARAP,interacts with tubulin and GABA(A) receptor. Biochem BiophysRes Commun 2004; 325: 639–648.
79 Wang H, Olsen RW. Binding of the GABA(A) receptor-associatedprotein (GABARAP) to microtubules and microfilaments suggestsinvolvement of the cytoskeleton in GABARAPGABA(A) receptorinteraction. J Neurochem 2000; 75: 644–655.
80 Stenmark H, Olkkonen VM. The Rab GTPase family. Genome Biol2001; 2: REVIEWS3007.
81 Pfeffer SR, Dirac-Svejstrup AB, Soldati T. Rab GDP dissociationinhibitor: putting rab GTPases in the right place. J Biol Chem 1995;270: 17057–17059.
82 Novick P, Zerial M. The diversity of Rab proteins in vesicletransport. Curr Opin Cell Biol 1997; 9: 496–504.
83 Takai Y, Sasaki T, Shirataki H, Nakanishi H. Rab3A smallGTP-binding protein in Ca(2þ )-dependent exocytosis. GenesCells 1996; 1: 615–632.
84 Ishizaki H, Miyoshi J, Kamiya H, Togawa A, Tanaka M, Sasaki Tet al. Role of rab GDP dissociation inhibitor alpha in regulatingplasticity of hippocampal neurotransmission. Proc Natl Acad SciUSA 2000; 97: 11587–11592.
Supplementary Information accompanies the paper on the Molecular Psychiatry website (http://www.nature.com/mp)
Neuroproteome of cocaine in nonhuman primatesNS Tannu et al
203
Molecular Psychiatry