insight into the microbial multicellular lifestyle via flow-cell technology and confocal microscopy
TRANSCRIPT

Insight into the Microbial Multicellular Lifestyle via
Flow-Cell Technology and Confocal Microscopy
Sunje Johanna Pamp,* Claus Sternberg, Tim Tolker-Nielsen
� AbstractBiofilms are agglomerates of microorganisms surrounded by a self-produced extracellu-lar matrix. During the last 10 years, there has been an increasing recognition of biofilmsas a highly significant topic in microbiology with relevance for a variety of areas in oursociety including the environment, industry, and human health. Accordingly a numberof biofilm model systems, molecular tools, microscopic techniques, and image analysisprograms have been employed for the study of biofilms under controlled and reproduc-ible conditions. Studies using confocal laser scanning microscopy (CLSM) of biofilmsformed in flow-chamber experimental systems by genetically color-coded bacteria haveprovided detailed knowledge about biofilm developmental processes, cell differentia-tions, spatial organization, and function of laboratory-grown biofilms, in some casesdown to the single cell level. In addition, the molecular mechanisms underlying theincreased tolerance that biofilm cells often display towards antibiotic treatment arebeginning to be unravelled. ' 2008 International Society for Advancement of Cytometry
� Key termsbiofilm; confocal microscopy; fluidic device; fluorescent reporter; subpopulation
BIOFILMS: THE MICROBIAL MULTICELLULAR LIFESTYLEMicroorganisms constitute the most successful form of life on earth, in terms of
total number, phylogenetic diversity and extent of habitats colonized. They impact
on human existence and well-being, either directly by influencing human develop-
ment, health and disease, or indirectly by carrying out processes in the natural envir-
onment or man-made environments (1–3). Our present state of knowledge on the
biology of microbial cells is, due to historical reasons, largely a result of empirical
research work on microbial cells living in suspension in a liquid growth medium.
However, it is now generally acknowledged that the majority of microbial cells on
earth are living in spatially distinct communities, referred to as biofilms. Biofilm cell
populations in many cases exhibit distinct features compared to planktonic cell
populations (1–5). Therefore, research work aimed at unraveling characteristics of
the microbial multicellular lifestyle will provide a more complete understanding and
definition of microbial life, and ultimately also human life, in nature.
Microbial Multicellular Communities
Biofilms come in a great variety of sizes and shapes. Some of the most common
types contain mushroom-like, pillar-like, hilly, or flat multicellular structures. These
structures are formed by cells that are held together by interconnecting compounds,
such as self-produced polysaccharides, proteins, extracellular DNA, and cell lysis pro-
ducts, as well as matter from the immediate surrounding environment, which alto-
gether constitute the so-called ‘‘matrix.’’ This allows cells to form long-term relation-
ships, interact with each other and establish metabolic cooperations (6,7). Biofilms
in nature are generally beneficial and frequently established on hydrous solid and
semi-solid surfaces, such as soil, rock material, or surfaces of animals and plants. Bac-
Department of Systems Biology,Technical University of Denmark, Lyngby,Denmark
Received 19 August 2008; Accepted3 November 2008
Grant sponsor: Danish research councils.
*Correspondence to: S€unje JohannaPamp, Department of Microbiology andImmunology, Stanford University Schoolof Medicine, VAPAHCS, 154T, Building101, B4-185, 3801 Miranda Avenue, PaloAlto, CA 94304, USA.
Email: [email protected]
Published online 2 December 2008 inWiley InterScience (www.interscience.wiley.com)
DOI: 10.1002/cyto.a.20685
© 2008 International Society forAdvancement of Cytometry
Review Article
Cytometry Part A � 75A: 90�103, 2009

terial communities play key roles in food webs in nature.
Many of the underlying processes are interdependent and
require cooperation between various bacterial species with dif-
ferent metabolic capacities (3,4,6,8,9). The fact that in biofilms
the participating microbial members are situated in close
proximity seems to be advantageous, since metabolites can
easily be transferred and metabolized further. In cases of
adverse conditions such as desiccation, osmotic shock, or ex-
posure to toxic compounds, UV radiation, or predators, the
microbial community as whole can provide protection. More-
over, multicellular communities provide ideal conditions for
horizontal gene transfer, which is important for microbial evo-
lution and genetic diversity (4,6,10,11). However, in many
man-made environments, such as industrial or medical set-
tings, the formation of sessile microbial communities, for
example, in production lines or on indwelling medical devices
is unwanted and detrimental. Resultant energy losses, corro-
sion, fouling, persistent infections, and potential death of
humans cause an enormous socioeconomic burden each year
worldwide (6,12).
Microbial Communities as Inhabitants of Humans:
Impact on Health and Disease
Microbial communities natively populate human mucous
membranes and epithelial surfaces like the gastrointestinal
tract, oral cavity, and skin. Each of the body sides is colonized
with a mixed microbial community of characteristic composi-
tion (2,13,14). Intriguingly, for most of our lifespan we do not
suffer from harboring these microbial communities. In fact,
they are important and beneficial to us as they can degrade
nutrients and thereby making them accessible to us, and
synthesize some vitamins, which we have not evolved to
synthesize on our own. Moreover, these communities play key
roles in the development of our immune system and anatomy
of the mucosal surfaces and exert protective functions against
exogenous pathogens (2,15,16).
The relationship between the host and its microbial com-
munities is delicately balanced but under certain conditions, it
can break down and result in infectious diseases. These infec-
tions can be caused either by members of the indigenous
human microbial community or by microorganisms from the
environment (1,2,6). Under conditions where the host is
impaired, for example immunocompromized, injured, or suf-
fering from cancer or cystic fibrosis, harmful biofilms can de-
velop at different body sides and cause persistent infections.
Among those infections are various device-related infections,
pulmonary infections, periodontitis, wound infections, otitis
media, osteomyelitis, infective endocarditis, and chronic pros-
tatitis (5,17–19). Bacteria, which have been found to be
involved in human biofilm-related infections, are for example
Pseudomonas aeruginosa, Staphylococcus spp., Escherichia coli,
Salmonella spp., Enterococcus spp., Streptococcus spp., Proteus
mirabilis, Klebsiella spp., Enterobacter spp., and Haemophilus
influenza. Biofilm-related infections are in many cases persis-
tent, that is, they evidently cannot be eradicated by the host
immune system and they are difficult to eradicate by antimi-
crobial chemotherapy (5,17–20).
Cultivation and Analysis of Multicellular Communities
Under Laboratory Conditions
Biofilms are intriguing societies of microbes and it is of
general interest to unravel the processes involved in their de-
velopment, physiology, and adaptation to perturbations. Em-
pirical research work on biofilm biology will ultimately reveal
new strategies that contribute to maintenance and restoration
of human health through well-informed manipulations of mi-
crobial communities. However, due to their complexity, nat-
ural microbial communities have been challenging objects of
investigation. In addition, biofilms are often located at places
that are difficult to access, which makes direct and continuous
examinations difficult. Various factors impact on biofilm biol-
ogy and many of those factors are interrelated. To reduce com-
plexity and facilitate investigations in the laboratory under
controlled and reproducible conditions, a number of ‘‘simple’’
biofilm model systems have been established. These include
flow-cell-grown biofilms, colony biofilms, microtiter dish-
grown biofilms, and pellicle biofilms (21–23). To discover
genetic determinants and regulatory pathways impacting on
the biofilm mode of life, major focus relies on well-character-
ized and genetically tractable microorganisms, such as Pseudo-
monas aeruginosa. The gold standard in biofilm research is an
approach, which involves flow-cell technology in combination
with confocal laser scanning microscopy (CLSM). Unlike
other techniques, this particular methodology allows getting
insight into details of developmental processes, spatial organi-
zation and function of laboratory-grown biofilms in real-time
under continuous and non-invasive conditions down to the
single-cell level (21,24,25).
CULTIVATION OF BIOFILM CELLS IN A FLUIDIC DEVICE
Biofilm flow-cell setups allow the cultivation of biofilms
under continuous hydrodynamic conditions. The biofilm
flow-cell system consists of five major components: A medium
reservoir, a multichannel peristaltic pump, bubble traps, flow-
cells, and an effluent reservoir. All parts are consecutively con-
nected via silicone tubings, splitters and connectors (Fig. 1A)
(22).
The Biofilm Flow-Cell
A central component of the biofilm flow-cell system is
the flow-cell, providing chambers for biofilm cultivation. A
number of different flow-cell designs exist. Widely used is the
flow-cell described here, which is a modified version of a flow-
cell originally developed by Wolfaardt et al. (26). The flow-cell
is designed so that it can be mounted on nearly any optical
microscope. It consists of two parts, a flow-cell base (Fig. 1B)
and a conventional microscopy glass coverslip. The flow-cell
base is made of a polycarbonate part in which parallel chan-
nels with individual dimensions of 40 3 4 3 1 mm have been
drilled. In the flow-cell presented here three individual chan-
nels have been drilled, meaning that in one flow cell three
individual (parallel) biofilm experiments can be performed.
To both ends of each channel, ports of 1 mm diameter have
been drilled, which serve as medium inlet and effluent outlet,
REVIEW ARTICLE
Cytometry Part A � 75A: 90�103, 2009 91

respectively (22). A microscopy glass coverslip (50 3 24 mm)
is placed on top of the flow-cell base and thereby covering the
open side of the channels to form closed channels. The cover-
slip is glued with silicone on the flow-cell base by applying sili-
cone glue as thin strings on top of the base along the perimeter
of the channels. The microscopy coverslip basically fulfills
three functions: it serves as one wall of the channels thereby
forming a closed flow channel, it serves as substratum for bio-
film formation, and it is optically compatible with micro-
scopic examination techniques (21,22,24,27).
The Biofilm Flow-Cell System
To eventually continuously supply the biofilm cells with
nutrients, the flow-cell needs to be connected with the remain-
ing components of the flow-cell system (Fig. 1A). Before the
medium reservoir is connected, possible contaminants are
removed from the entire system. This is achieved by disinfec-
tion of the system with sodium hypochloride or ethanol, or by
sterilization with ethylene gas. Most components are also com-
patible with autoclaving at 1218C. Subsequently the reservoir
with sterile nutrient medium is connected to the system. The
composition of the medium is chosen dependent on the
requirements of the organism(s) of interest. The system is
filled with medium using a peristaltic pump. Conventional
peristaltic pumps do not deliver an entirely pulseless flow, in
contrast to multiroller peristaltic pumps. If the carbon source
in the medium has a high hydrophobicity then medium is run
through the system for an extended period to equilibriate the
silicone tubings, before introducing the organisms of interest
(21,22,24,27).
The organisms of interest are introduced at desired initial
optical density into the flow chambers via a syringe while me-
dium flow is paused. The syringe is inserted upstream of the
flow-cell through the silicon tubing into the inlet of the flow
chamber, the cell suspension of microorganisms is injected
and the resulting fine hole in the silicone tubing is sealed with
a drop of silicone glue. Microbial cells are allowed to attach to
the substratum for 1 h, whereupon medium flow is resumed
and adjusted to laminar flow. For the biofilm system described
here, the flow rate is often set to 0.2 mm/s (approximately 3
ml/h/channel) (21,22,24,27). Universal equations for the cal-
culation of hydrodynamic parameters in a flow-cell can be
found elsewhere in the literature (e.g., 28). While fresh me-
dium is continuously transported into the flow cells to allow
biofilm development and differentiation, effluent is trans-
ported out and collected in the reservoir placed at the most
downstream part of the system.
Under certain circumstances air bubbles might arise in
the system, which can remove biofilm cells from the substra-
tum while passing through a flow channel. To prevent this,
bubble traps have been designed and are mounted between
the peristaltic pump and the flow-cells. Bubble traps are com-
posed of three parts: a bubble trap base made of a polycarbo-
nate part (Fig. 1C), a syringe cylinder, and a lid to close the
top of the syringe cylinder. Any air bubbles that might pass
into the bubble trap will float to the top of the syringe cylin-
der, and are prevented from passing through the downstream
part of the system. Note that the inlet of the bubble trap base
is designed so that it is situated higher than the outlet part
(Fig. 1C).
The biofilm setup is compatible with various visualiza-
tion and quantification techniques (see following text). Cell–
cell interactions in a biofilm can be studied between (i) cells of
a single strain (e.g., 29), (ii) cells of a number of different
Figure 1. The biofilm flow-cell system allows the cultivation and
analysis of biofilm cells under continuous hydrodynamic condi-
tions. The system consists of five major components: a medium
reservoir, amultichannel peristaltic pump, bubble traps, flow-cells,
and an effluent reservoir. All parts are consecutively connected via
silicone tubings, splitters and connectors (A). Working drawing of
the flow cell base (design copyright DTU-Biosys). Reproduced
from Curr Protoc Microbiol 1B.2.1—1B.2.15 with permission fromJohnWiley& Sons, Inc. (B).Working drawing of the advanced bub-
ble trap base (design copyright DTU-Biosys). Reproduced from
Curr Protoc Microbiol 1B.2.1—1B.2.15 with permission from John
Wiley & Sons, Inc. (C). [Color figure can be viewed in the online
issue, which is available at www.interscience.wiley.com.]
REVIEW ARTICLE
92 Insight Into Biofilms via Confocal Microscopy

strains (e.g., mutants) of a particular single species (e.g.,
30,31), (iii) cells belonging to different bacterial species (e.g.,
32–35), and (iv) cells belonging to different domains, such as
bacterial cells and eukaryotic cells (e.g., 36). Activities in nat-
ural multicellular microbial consortia might also be studied in
flow-cells after harvesting samples from the natural environ-
ment, and transplanting the microbial cells into flow-cham-
bers where they are subjected to further analysis (e.g., 26,37).
The setup might be modified, for example, to study the effect
of substratum coatings on biofilm development and physiol-
ogy, or to study the effect of oxygen up- or down-shifts on
biofilm cells (38–40). Biofilm cells might also be harvested
from the flow chambers and subjected to further analysis, for
example global transcriptional analysis, fluorescence activated
cell sorting (FACS), or c.f.u. determinations (31,41,42). The
entire biofilm flow-cell system can be placed on a rolling table
to facilitate secure transport of the system for example
between an incubator room with a constant temperature (of-
ten 308C or 378C) and the room in which microscopic exami-
nations of the biofilms are taking place.
ANALYSIS OF BIOFILM CELLS
Because biofilms are complex 3-dimensional structures
the analysis of them is not trivial. While microbial single cells
easily can be monitored using a conventional microscope, bio-
films require additional resolution in the direction vertical to
the substratum (the z-axis). Here we describe the use of confo-
cal laser scanning microscopy (CLSM), fluorescent labeling of
biofilm cells, and image analysis in biofilm research.
Confocal Laser Scanning Microscopy (CLSM)
The use of the confocal laser scanning microscope has
helped overcoming the apparent shortcomings of the conven-
tional light microscope (e.g., the presence of out-of-focus
light) by introducing point illumination and a pinhole, which
allows optical sectioning of the specimen. The individual opti-
cal sections are subsequently assembled by aid of advanced
computer software (some examples are listed in the following
text) to a virtual 3D image (e.g., Figs. 2A and 2B and Fig. 3).
Typically a biofilm with a thickness of more than �150 lmcannot be rendered with a reasonable detail due to physical
factors. The implementation of multiphoton excitation is a
major step forward. Using a pulsed laser it is possible to guide
two (or more) photons to excite a fluorophore simultaneously.
This means that the energy of the photons is combined to
excite the target molecule. Using this technique the depth re-
solution (i.e., the minimum distance to resolve two points) is
many fold increased, from around 500–700 nm for standard
confocal microscopy to 100 nm for two-photon microscopy
(43). Neu and co-workers demonstrated two-photon confocal
imaging of thick z-sections with very high resolution (43,44).
Recently a new technology, stimulated emission depletion
(STED) has been developed into commercial products,
increasing the optical resolution even further by using two-
photon excitation in combination with quenching of near-by
fluorescence which otherwise could deteriorate the image
(45,46). STED, however, is currently of limited use since
only few fluorophores are suitable for this particular laser
excitation.
Fluorescent Labeling of Biofilm Cells
Confocal microscopy and derived methods require the
specimen to be fluorescent. The biofilm must therefore either
be autofluorescent by means of indigenous fluorescent mole-
cules, or the biofilm cells must express a fluorescent protein
(e.g., the green fluorescent protein, Gfp (47)), or individual
biofilm cells or other components of the multicellular struc-
ture must be stained. Early biofilm studies by the Caldwell
group employed a simple, yet efficient way of detecting the
biomass in flow cells: The void volume, that is, the liquid
phase was supplemented with a solution of fluorescein iso-
thiocyanate (FITC), leaving the biomass unstained. The result-
ing images were ‘‘negatives’’ and the biofilm could be rendered
as the dark portions of the images (48,49). This gave suffi-
ciently high resolution to determine for example cell sizes and
spatial relations. More recently developed stains, such as the
Syto stains (Invitrogen, Carlsbad, CA), can efficiently stain
cells in virtually any color of the rainbow. In combination with
propidium iodide (PI) it is further possible to specifically stain
live and dead cells. The dye Syto 9 will stain all cells green
regardless if they are dead or alive, while it is generally
assumed that only cells with a damaged membrane will be
stained by the red PI dye, indicating dead cells. Recent results
suggest that propidium iodide might be of limited use as cell
viability indicator for some environmental bacterial species
(50). Therefore, for each individual bacterial species a fine-
tuning of the dye combinations is recommended prior to per-
forming the actual experiments using mixtures of live and
dead cells of known ratios (51). Recently the assumption that
propidium iodide only targets dead cells was confirmed also
for biofilm cells that had been exposed to a membrane dama-
ging agent, using cell sorting of harvested biofilm cells and
re-growth test of the separated green (Gfp-tagged) and red
(PI-stained) cells on nutrient agar. Only the cells that were not
labeled red with PI were able to grow (42, and unpublished
results). Stains targeting the extracellular matrix such as lectins
(52,53) or calcofluor white (54,55) can also be employed to
visualize the surrounding of the biofilm cells. In addition, the
extracellular DNA component of the matrix can be visualized by
the use of different DNA-binding fluorophores (e.g., Ref. 56).
If genetic manipulation of the biofilm cells is possible,
chromosomal tagging of cells with a gene cassette encoding
the green fluorescent protein (Gfp) can be a useful option
(e.g., 57). Alternatively, plasmids encoding for the Gfp might
be introduced into the cells prior to biofilm examinations.
Depending on the construction, this fluorescent tagging can
be used as a simple labeling to verify the location of the cells
in a biofilm, or, by selecting suitable variants of gfp genes and
promoters, it can be used for monitoring gene expression in
biofilms. Such tagging of biofilm cells has been done to moni-
tor metabolic/physiological activity in biofilms by introducing
constructs encoding for Gfp derivatives with a short half-life,
placed under transcriptional control of a ribosomal promoter
(58). For example, the gfp[AGA] gene, encoding for a green
REVIEW ARTICLE
Cytometry Part A � 75A: 90�103, 2009 93
fluorescent protein (Gfp) with a short half life, was placed
under transcriptional control of the ribosomal promoter
rrnBP1 and introduced into either E. coli or Pseudomonas spp.
wild type strains. Cells which have a high metabolic/physio-
logical activity can be expected to exhibit high Gfp[AGA]
expression and emit a high fluorescent signal, whereas cells
which have a low metabolic/physiological activity can be
expected to exhibit a low or no expression of the fluorescent
protein (41,58). Further, using Gfp variants with different
emission spectra, such as the Cfp (cyan fluorescent protein),
Yfp (yellow fluorescent protein), and Rfp (red fluorescent
protein), the spatial distribution of either cells in a multi spe-
cies biofilm can be determined, or of a number of (mutant)
strains of a single species tagged with different colors
(30,31,59,60).
Another way of fluorescently labeling biofilm cells is
through the use of fluorescent in situ hybridization (FISH),
where specific probes hybridize to the 16S rRNA in the cells.
For FISH a DNA probe is designed to match a distinct region
of the cell’s ribosomal RNA. The probes can be conjugated to
a fluorescent dye, such as fluorescein isothiocyanate (FITC) or
Rhodamine, or to an enzyme (e.g., horseradish peroxidase),
which deposits fluorescent molecules. It might be challenging
to introduce larger conjugates such as the horseradish peroxi-
dase into cells of thicker biofilms without destroying biofilm
cells due to harsh permeabilization procedures. Therefore
FISH involving probes with larger conjugates might preferen-
tially be applied on thin sections of thick biofilms. The num-
ber of ribosomes present in a given cell is proportional to the
growth potential of the cell, and FISH labeling can conse-
quently also be used to determine the growth status of a bio-
film cell (61). However, under certain conditions, for example,
stress, cells might have increased numbers of ribosomes,
although their actual growth rate is low. The probe design can
be adjusted so that the probe only labels a single species by tar-
geting a so called variable rRNA region, or a probe can label
all cells belonging to the same domain or phylum by choosing
a more conserved region. An example of such a probe is the
widely used EUB338, which can hybridize to virtually all bac-
teria (62). However, the growing knowledge on ribosomal
RNA encoding sequences has revealed that probes formerly
believed to be universal fail to be able to hybridize to species
or entire phyla of microorganisms in the realm they originally
were thought to cover completely (63).
Image Analysis
The recorded microscopic images can be used immedi-
ately or processed further for presentation or quantitative
analysis. The images that originate from a confocal micro-
scope are usually grayscale bitmap images, one from each focal
plane and one for each detection channel (color). An image of
a 30-lm-thick biofilm recorded with a step size of 0.5 lm in
three channels will result in 30/0.5 3 3 5 180 individual
images. Most microscope software pack the images in contain-
ers such as the LIF file format for Leica and the LSM format
for Zeiss microscopes. Furthermore the images will be rela-
tively large. In the example above the standard resolution of
256 gray tones and 512 3 512 pixels will result in a file with
the size of 45 MB. Special software is required to handle these
files and to render the beautiful biofilm representations. While
several packages are available, a few seem to dominate the
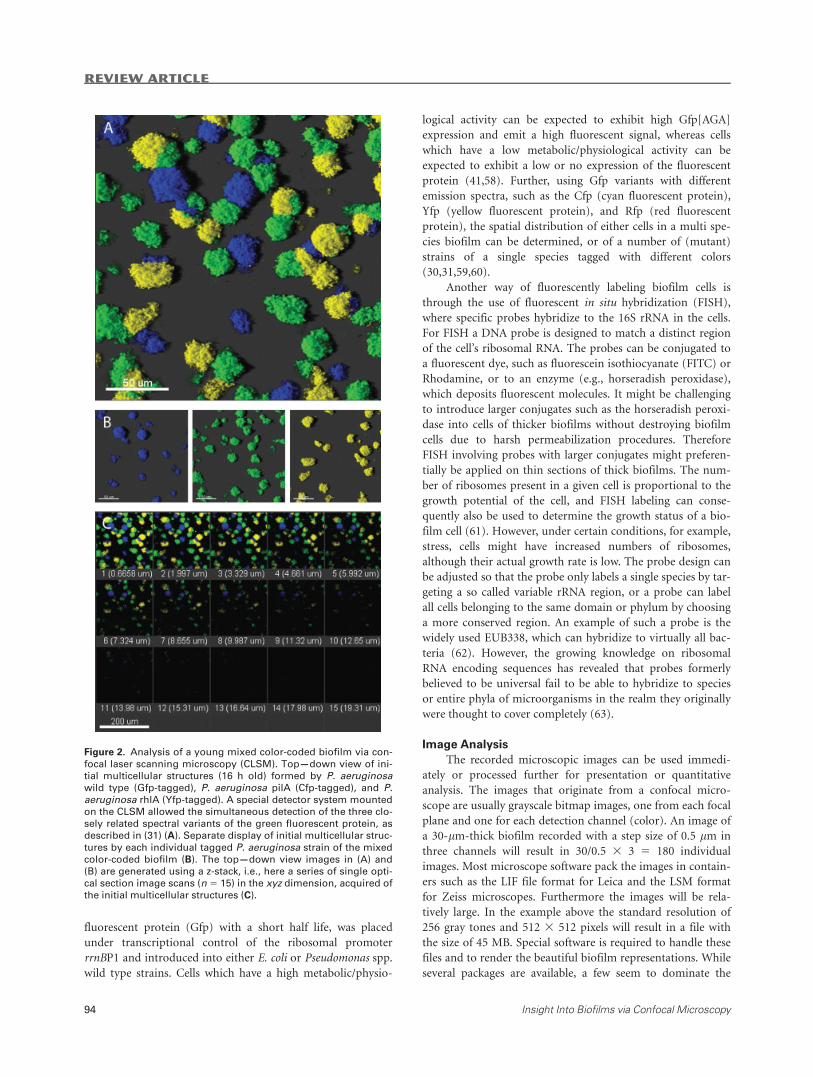
Figure 2. Analysis of a young mixed color-coded biofilm via con-
focal laser scanning microscopy (CLSM). Top—down view of ini-
tial multicellular structures (16 h old) formed by P. aeruginosawild type (Gfp-tagged), P. aeruginosa pilA (Cfp-tagged), and P.aeruginosa rhlA (Yfp-tagged). A special detector system mounted
on the CLSM allowed the simultaneous detection of the three clo-
sely related spectral variants of the green fluorescent protein, as
described in (31) (A). Separate display of initial multicellular struc-
tures by each individual tagged P. aeruginosa strain of the mixedcolor-coded biofilm (B). The top—down view images in (A) and
(B) are generated using a z-stack, i.e., here a series of single opti-
cal section image scans (n 5 15) in the xyz dimension, acquired ofthe initial multicellular structures (C).
REVIEW ARTICLE
94 Insight Into Biofilms via Confocal Microscopy
market: Imaris (Bitplane, Bern, Switzerland), Amira (Visage
Imaging, Carlsbad, CA), Volocity (Improvision, Coventry, the
UK), Voxblast (Vaytek, Fairfield, IA), and Metamorph (Molec-
ular Devices, Sunnyvale, CA) (see 43 for a review). Designed
for the visualization of larger eukaryotic cells, most 3D presen-
tation packages are not optimized for the small cell sizes
involved in microbial biofilms. The average size for a Gram-
negative bacterium is 1 lm by 2–3 lm. The optical resolution
of a confocal image recorded with one-photon excitation is at
best 0.48 lm (64). This means that a sampling of images with
a smaller step size than this will not provide more informa-
tion. Consequently, a single bacterium will not appear in more
than one or two independent focal plane images, making a 3D
reconstruction of the single cell difficult. However, the biofilm
as a whole is much larger and can be rendered in 3D, although
it may not be possible to locate the individual cells. Typically
a step size of 1 lm is used for bacterial biofilms (e.g., Ref. 30).
The visualization software packages also include tools for
cleaning up the recordings, such as filters for noise and cross-
talk (the situation where one fluorophore is recorded in the
detection channel of another fluorophore). The main features
of these softwares are, however, their capability to visualize
the spatial organization of the recorded data. The rendering
can be in perspective 3D, or as 2D images in all three axes,
x-y, x-z and y-z. Sequential recordings over time can also be
rendered, and animated, providing a four-dimensional data-
set, x-y-z-t. Such data sets can quickly be very large (some-
times several gigabytes) and it sets new requirements for the
software and hardware. Special detector systems mounted on
the confocal microscope might also facilitate the simultane-
ous separation of a number of fluorescent spectra (lambda
mode configurations) originating from different fluorescent
molecules or proteins with overlapping emission spectra,
increasing the possibilities for multi fluorescent labeling of
biofilms (Fig. 2) (31).
Quantitative analysis of 3D images can be challenging
and several groups have developed special software packages
for this purpose. The algorithms start by determining the
extent of the biomass, by thresholding each focal plane
image. This step is crucial and much effort has been put into
optimization of it. It can be done either manually, semi- or
fully automated. Some of the first attempts to provide robust
quantification software were done by Yang et al. (65) and
Heydorn et al. (66) with the programs ISA and COMSTAT,
respectively. Both extract a number of parameters, which can
be used to characterize the biofilm: biomass (pixels occupied
by biomass), biofilm height, height distribution, roughness
coefficient, and diffusion distances, to mention a few. Both
programs are developed in MATLAB (MathWorks, Natik,
MA). COMSTAT utilizes a command-line interface within
the MATLAB shell, which is required for operation, whereas
ISA and its successor ISA3D are compiled programs that do
not require the MATLAB package. Other quantification soft-
ware is available, for example, the web-based PHILIP (67), a
program that has a higher level of automation than the ISA
and COMSTAT packages. Further developments of PHILIP
have taken the automation and robustness towards the
threshold function to a new level (68,69). COMSTAT is now
available in a complete reprogrammed version 2, which is
running on the platform independent software foundation
Java (unpublished results). This new version of COMSTAT
software uses the same thresholding algorithms as the PHI-
LIP software and also incorporates a number of new features
such as a user programmable plug-in interface for end user
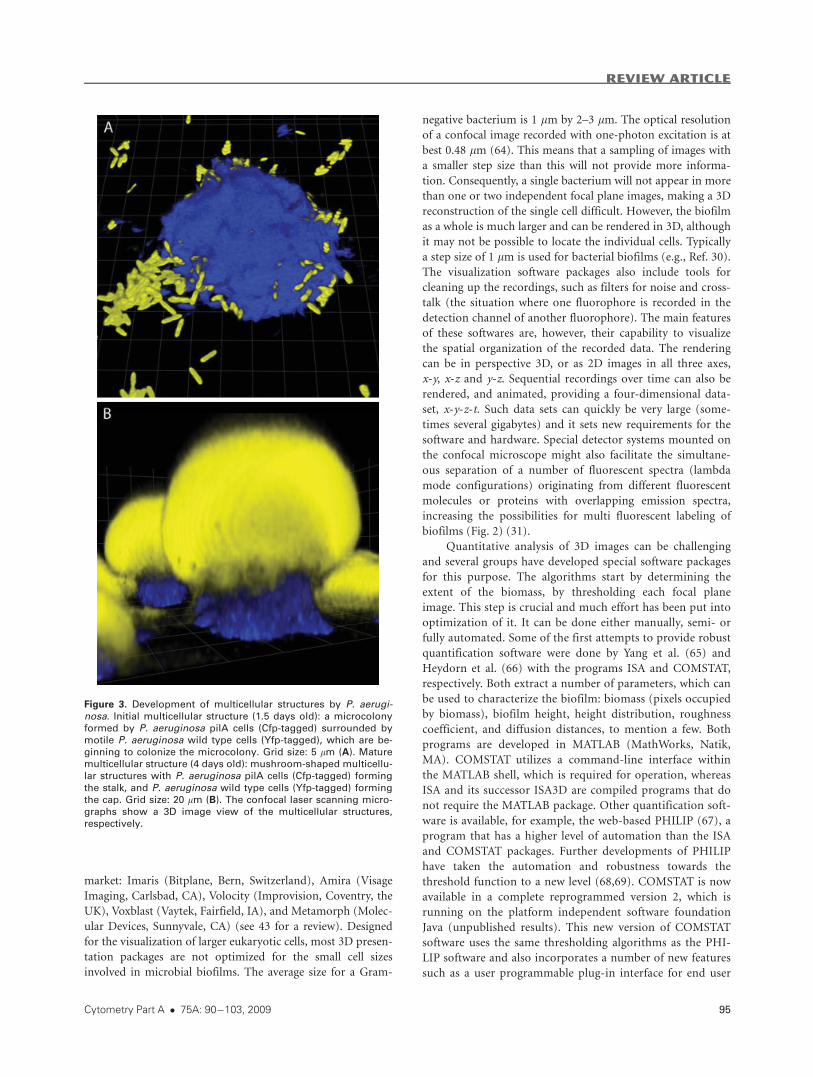
Figure 3. Development of multicellular structures by P. aerugi-nosa. Initial multicellular structure (1.5 days old): a microcolonyformed by P. aeruginosa pilA cells (Cfp-tagged) surrounded by
motile P. aeruginosa wild type cells (Yfp-tagged), which are be-ginning to colonize the microcolony. Grid size: 5 lm (A). Mature
multicellular structure (4 days old): mushroom-shaped multicellu-
lar structures with P. aeruginosa pilA cells (Cfp-tagged) formingthe stalk, and P. aeruginosa wild type cells (Yfp-tagged) formingthe cap. Grid size: 20 lm (B). The confocal laser scanning micro-
graphs show a 3D image view of the multicellular structures,
respectively.
REVIEW ARTICLE
Cytometry Part A � 75A: 90�103, 2009 95

defined image processing functions, and a wider range of
image formats.
DEVELOPMENT OF MULTICELLULAR STRUCTURES IN
PSEUDOMONAS AERUGINOSA BIOFILMS
Studies involving flow-chamber technology and CLSM
have provided knowledge about biofilm formation of numer-
ous bacterial species of both environmental and medical rele-
vance. The studies have, among many other things, given
information about the mechanisms, environmental cues and
underlying genetic elements involved in attachment of cells to
surfaces, formation of multicellular structures, and dispersal
of cells from multicellular structures. In the present section we
present examples of work done on these topics with biofilms
formed by the opportunistic pathogen P. aeruginosa.
Transport of Cells to the Surface
The bacterial cells can reach a surface prior to coloniza-
tion by means of passive motility mediated by brownian
motion or vortex currents, or by active swimming motility
mediated by flagella rotation (70). Flagella-driven motility has
been reported to enhance the efficiency of surface colonization
by P. aeruginosa (71,72). The dependence on flagella is condi-
tional however, as non-flagellated P. aeruginosa mutants and
the isogenic wild type were shown to attach equally well to a
surface in flow-chambers under some conditions (29).
Attachment of Cells to the Surface
P. aeruginosa appears to possess a number of different
adhesins that can function in attachment to a surface. The
conditional dependence on flagella for surface colonization
described above may, besides a role in bacterial transport to
the surface, be because of adhesion properties of the flagella. P.
aeruginosa cells were shown to attach apically to the glass sur-
face in flow-chambers and rotate, indicating that the initial
attachment occurred by means of flagella (72). The initial
attachment was reversible, but cells became irreversibly
attached by progressing from apical to longitudinal attach-
ment. The gene sadB (PA5346) was shown to be required for
the progression from apical to longitudinal attachment (73).
Non-flagellated mutants of P. aeruginosa attached longitudin-
ally (72), indicating that flagella are not required for attach-
ment per se but may enhance the process. Type IV pili were
shown to be important for attachment of P. aeruginosa to vari-
ous surfaces (74–76), and hyperpiliated variants of P. aerugi-
nosa were shown to rapidly initiate formation of strongly
adherent biofilms (75). The dependence on type IV pili for
cell attachment appears to be conditional, however, as non-
piliated P. aeruginosa mutants and the isogenic wild type were
shown to attach equally well to surfaces in flow-chambers
under some conditions (29). In addition to flagella and type
IV pili, fimbrial appendages termed Cup fimbria were shown
to play a role in surface-attachment of P. aeruginosa (77), and
evidence has been provided that extracellular DNA plays a role
in surface attachment of P. aeruginosa cells under some condi-
tions (78).
Formation of Initial Multicellular Structures
Shortly after attachment to the surface in flow-chambers
the P. aeruginosa cell population consist of a non-motile sub-
population, and a motile subpopulation that moves on the
surface via type IV pili or flagella activity (29,30,79,80).
Experiments with mixtures of Cfp-tagged and Yfp-tagged cells
provided evidence that microcolonies develop by clonal
growth in the flow-chambers (30). The initial microcolonies
are formed by cells, which do not display motility and there-
fore proliferate at fixed positions, while the cells that move on
the surface do not participate in formation of the initial
microcolonies (29,30). Conditions that promote extensive
motility of P. aeruginosa have been shown to prevent micro-
colony formation in flow-chambers. Interestingly, this could
be a mechanism which prevents biofilm formation in humans
since lactoferrin, a component of the innate immune system,
was shown to induce extensive type IV-driven motility in P.
aeruginosa (79).
The cellular adhesiveness mediated by the factors
involved in surface attachment probably also plays a role as
cell-to-cell adhesin in the earliest phase of microcolony forma-
tion. For example, the small microcolonies formed by P. aeru-
ginosa in the initial phase of biofilm formation could be dis-
persed by exogenous DNase activity indicating a role for extra-
cellular DNA as matrix component in the early phase of P.
aeruginosa biofilm formation (78).
Formation of Mature Multicellular Structures
Although the relative importance of the mechanisms
involved in initial formation of multicellular structures
appears to vary dependent on the conditions, the outcome of
initial biofilm formation seems to be similar under various
conditions. The early P. aeruginosa biofilm generally consists
of a subpopulation of non-motile cells forming small microco-
lonies, and a subpopulation of motile cells, which move on
the surface between the small microcolonies. After formation
of the initial microcolonies structural biofilm development by
P. aeruginosa depends on the prevailing conditions. For exam-
ple, flat biofilms are formed in flow-chambers irrigated with
citrate minimal medium (29), while heterogeneous biofilms
containing mushroom-shaped multicellular structures are
formed in flow-chambers irrigated with glucose minimal
medium (30).
Formation of the flat P. aeruginosa biofilm in flow-cham-
bers irrigated with citrate medium was shown to occur via ex-
pansive surface-migration of cells from the initial microcolo-
nies (29). CLSM time-lapse microscopy indicated that a shift
from non-motile to migrating cells occurred when the initial
microcolonies reached a certain size, suggesting that the shift
may by induced by a limitation arising in the initial microco-
lonies (For representative movies the reader is referred to Ref.
81). Migration of the bacteria appeared to cease with matura-
tion of the citrate-grown biofilm. Since biofilm formation by a
P. aeruginosa pilA mutant (deficient in biogenesis of type IV
pili) occurred without the expansive phase, and resulted in
discrete protruding microcolonies, it was suggested that the
REVIEW ARTICLE
96 Insight Into Biofilms via Confocal Microscopy

expansive migration of the cells on the surface was type IV
pili-driven (29).
Formation of the mushroom-shaped structures in glu-
cose-grown P. aeruginosa biofilms evidently occurs in a se-
quential process where the initial microcolonies formed by the
non-motile subpopulation become colonized by cells from the
migrating subpopulation that subsequently form mushroom
cap-like structures on top of the initial microcolonies, which
then become mushroom stalks (for representative movies the
reader is referred to Ref. 82) (30,42). Growth of the initial
microcolonies in the glucose-grown biofilms continue past the
point where spreading by type IV-driven motility prevents fur-
ther microcolony formation in the citrate-grown biofilms. In
glucose-grown biofilms containing a mixture of Cfp-tagged
and Yfp-tagged P. aeruginosa wild type cells, mushroom-
shaped structures are formed that have single-color stalks and
two-color caps, in concordance with the stalks being formed
by proliferation of non-motile cells, and the caps being formed
via aggregation of motile cells (30). In biofilms containing a
mixture of P. aeruginosa wild type and P. aeruginosa pilA mu-
tant, the pilA mutants can only form stalks whereas the wild-
type cells form the caps, suggesting that type IV pili are neces-
sary for cap-formation (30). Figure 3A shows a microcolony
formed by Cfp-tagged pilA mutants surrounded by Yfp-tagged
motile wild type cells, which are beginning to colonize the
microcolony. Figure 3B shows mature mushroom-shaped
structures with Cfp-tagged pilA mutants in the stalk and Yfp-
tagged wild type cells in the cap. In addition to type IV pili,
formation of the mushroom cap evidently also depends on fla-
gella, as P. aeruginosa fliM mutants (deficient in biogenesis of
flagella) were shown to be deficient in cap-formation (83).
Evidence was provided that flagellum-driven surface-asso-
ciated motility is involved in cap-formation, whereas the de-
pendence of cap-formation on type IV pili may be due to
binding of these pili to extracellular DNA which is particularly
abundant on the microcolonies that become colonized during
mushroom-structure formation (as described in the following
text). In accordance with a role of type IV pili-driven motility
in the early phase of biofilm formation and a role of flagella-
driven motility in the later phase of biofilm formation, fla-
gella-driven surface associated motility (swarming) is known
to depend on quorum sensing (e.g., 84), that is, a mechanism
by which bacteria can monitor their cell population density
through the extracellular accumulation of signaling molecules
and express genes when the cell density is high. Production of
biosurfactants, which is under quorum sensing control, was
shown to facilitate formation of the mushroom cap structures
(31). Migration of the bacteria appears to cease with matura-
tion of the glucose-grown biofilm.
Experiments involving the fluorescent reporter rrnBP1-
gfp[AGA] (see preceding text) provided evidence that meta-
bolic/physiological activity is highest in cells forming the cap-
subpopulation, and lowest in cells forming the stalk-subpopu-
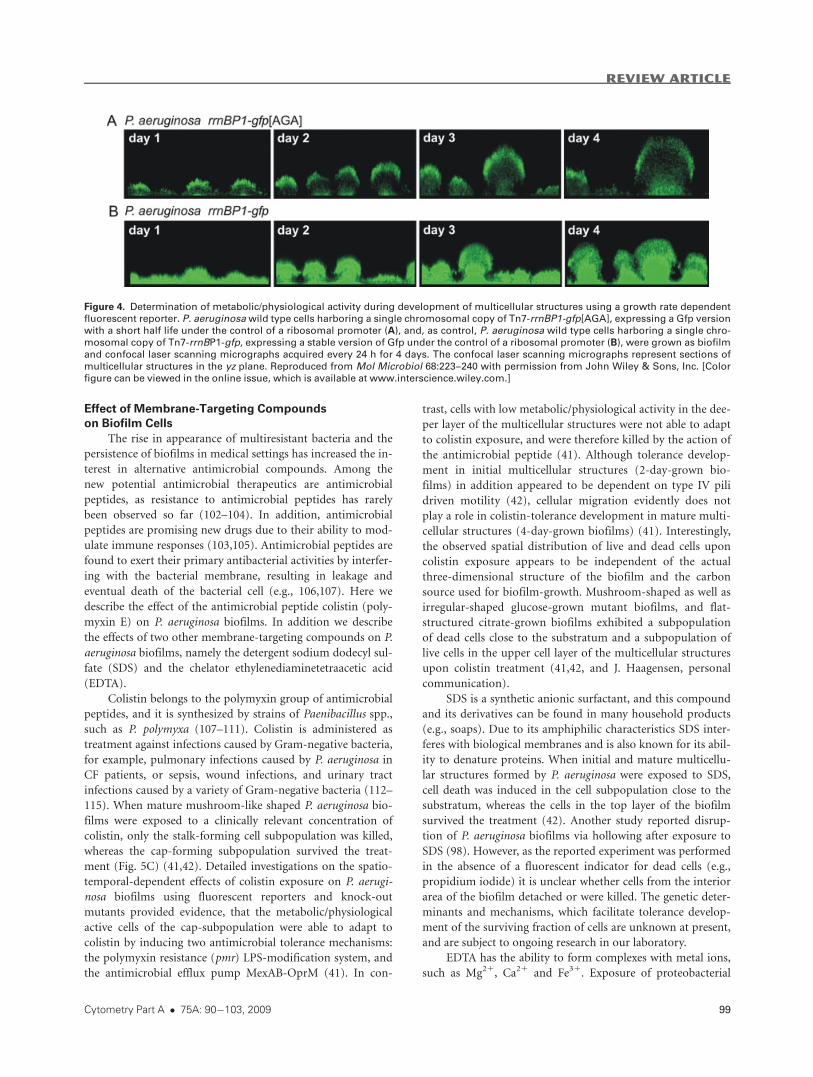
lation (41). As can be seen in Figure 4A, the cells of the cap-
forming subpopulation exhibit a high fluorescent signal, indi-
cating high metabolic activity, whereas cells of the stalk-form-
ing subpopulation exhibit a low fluorescent signal, indicating
low metabolic activity (41). In the control biofilm formed by a
P. aeruginosa wild type strain expressing the stable version of
Gfp, all cells exhibit similar levels of fluorescence emission
(Fig. 4B). This spatial distribution of high and low metaboli-
cally active cells seems plausible as cells in the cap part can
obtain oxygen and nutrients from the bulk liquid to drive
metabolic processes such as replication, transcription and
translation, in contrast to the cells in the stalk part where con-
centrations of dissolved oxygen and nutrients are likely to be
low. It might be speculated that higher levels of oxygen and
nutrients in the bulk liquid are sensed by some cells during
biofilm development, and that this results in an attraction,
driving cells to migrate on top of microcolonies formed by
non-motile cells. In support of this, P. aeruginosa strains de-
fective in chemosensory systems show defects in cap-forma-
tion [(83) and S.J.P and T.T.N., unpublished].
The multicellular structures in P. aeruginosa biofilms are
stabilized by a matrix consisting of exopolysaccharides, extra-
cellular DNA, and proteins. The alg operon (PA3540-PA3551)
encoding alginate polysaccharide appears not to be expressed
in P. aeruginosa flow-chamber biofilms (85), but evidently
plays a role in biofilm formation by P. aeruginosa in particular
in the lungs of cystic fibrosis patients (86). The psl genes
(PA2231-PA2245) encode production of a mannose rich exo-
polysaccharide, which was shown to play a role in P. aerugi-
nosa biofilm formation (87–89). The pel (PA3058–PA3064)
genes encode production of a glucose rich matrix component,
which was shown to facilitate biofilm formation of some P.
aeruginosa strains (90). Extracellular DNA was shown to be
present in high concentrations particularly on the microcolo-
nies in young P. aeruginosa biofilms and between the stalk-
forming and cap-forming subpopulations in mature biofilms
(56). Type IV pili bind to DNA (91,92), and evidence has been
presented that the high concentration of extracellular DNA on
the mushroom stalks may cause accumulation of the migrat-
ing piliated cells and thereby facilitate formation of the mush-
room caps (83). Production of large amounts of extracellular
DNA in P. aeruginosa biofilms has been shown to be depend-
ent on the PQS quorum-sensing system (56). Evidence is
accruing that Cup fimbria in addition to their role in initial
biofilm formation also play a role as matrix components in
mature biofilms. The sadARS genes (PA3946-3948, also
termed rocARS) were shown to regulate biosynthesis of Cup
fimbria (93), and mutations in any of these genes resulted in
biofilms with an altered mature structure (94).
Dissemination of Cells fromMulticellular Structures
In addition to the mechanisms involved in biofilm for-
mation, bacteria also possess mechanisms to reduce their
adhesiveness and to break down or modulate the biofilm ma-
trix. Emigration of cells from biofilm communities is neces-
sary to spawn novel communities at new locations, and it may
be induced if the biofilm cells face unfavorable conditions
(e.g., 95,96). Migration of cells may also allow sessile commu-
nities to change spatial organization in response to changing
environments (e.g., 34). After prolonged biofilm development
of P. aeruginosa in flow-chambers local dispersion was
REVIEW ARTICLE
Cytometry Part A � 75A: 90�103, 2009 97

observed as a hollowing out of the mature microcolonies (97).
Through careful microscopic inspection it was observed that
two subpopulations existed in the mature multicellular struc-
tures. The outer parts of these structures contained a wall-
forming subpopulation of non-motile cells, whereas a motile
rapidly moving cell subpopulation was present inside the mul-
ticellular structures. The motile subpopulation coordinately
evacuated the multicellular structures from local break out
points resulting in structures with a central void. This phe-
nomenon has been termed ‘‘seeding dispersal,’’ and was shown
to be dependent on the mature multicellular structures reach-
ing a critical size, suggesting that it may be induced by sub-
strate limitation or accumulation of signal molecules or waste
products. Biosurfactants appears to have multiple roles in
P. aeruginosa biofilm development, as production of large
amounts of rhamnolipid biosurfactant has been associated
with dispersal of cells from P. aeruginosa biofilms (98).
ANTIMICROBIAL ACTION ON BIOFILM CELLS
Biofilms in the environment as well as those associated
with animals and plants or present in man-made environ-
ments are frequently exposed to antimicrobial compounds,
both of natural and synthetic origin. Flow-chamber-grown
biofilms appear to be a useful model system to study antimi-
crobial action in biofilms, for example, as the spatial appear-
ance and distribution of dead and surviving cells in a biofilm
upon antimicrobial attack can be followed in real time. Here
we will give an overview of the spatial antimicrobial suscepti-
bility and tolerance phenotypes of P. aeruginosa biofilms living
in flow-chambers exposed to a number of different anti-
microbial compounds.
As described in the previous section, P. aeruginosa mush-
room-shaped biofilms are commonly found to be composed
of two major subpopulations, a subpopulation situated close
to the substratum and a subpopulation on top. Intriguingly, it
appears that antimicrobial compounds seem to exert their
antimicrobial effects on only one of the two subpopulations,
whereas the other subpopulation survives the treatment. In
most cases the surviving subpopulation of cells exhibits phe-
notypic tolerance and not resistance, as surviving biofilm cells
harvested from antimicrobial-treated biofilms exhibit the
same antimicrobial susceptibility phenotype as the cells, which
were used to initiate the biofilm (e.g., Refs. 41,42).
Effect of Conventional Antibiotics on Biofilm Cells
Most conventional antimicrobial agents used to treat bac-
terial infections in humans and animals, interfere with funda-
mental physiological processes of bacterial cells, such as repli-
cation, transcription or translation processes. Here we describe
the effect of three such conventional antibiotics on mush-
room-like shaped P. aeruginosa biofilms, namely ciprofloxacin,
tetracycline, and tobramycin.
The fluoroquinolone ciprofloxacin induces bacterial cell
death by interfering with the replication process due to inhibi-
tion of the DNA gyrase. Ciprofloxacin is administered for
treatment of various infections caused by Gram-negative and
Gram-positive bacteria. Exposure of P. aeruginosa flow-cell-
grown biofilms was found to preferentially induce cell death
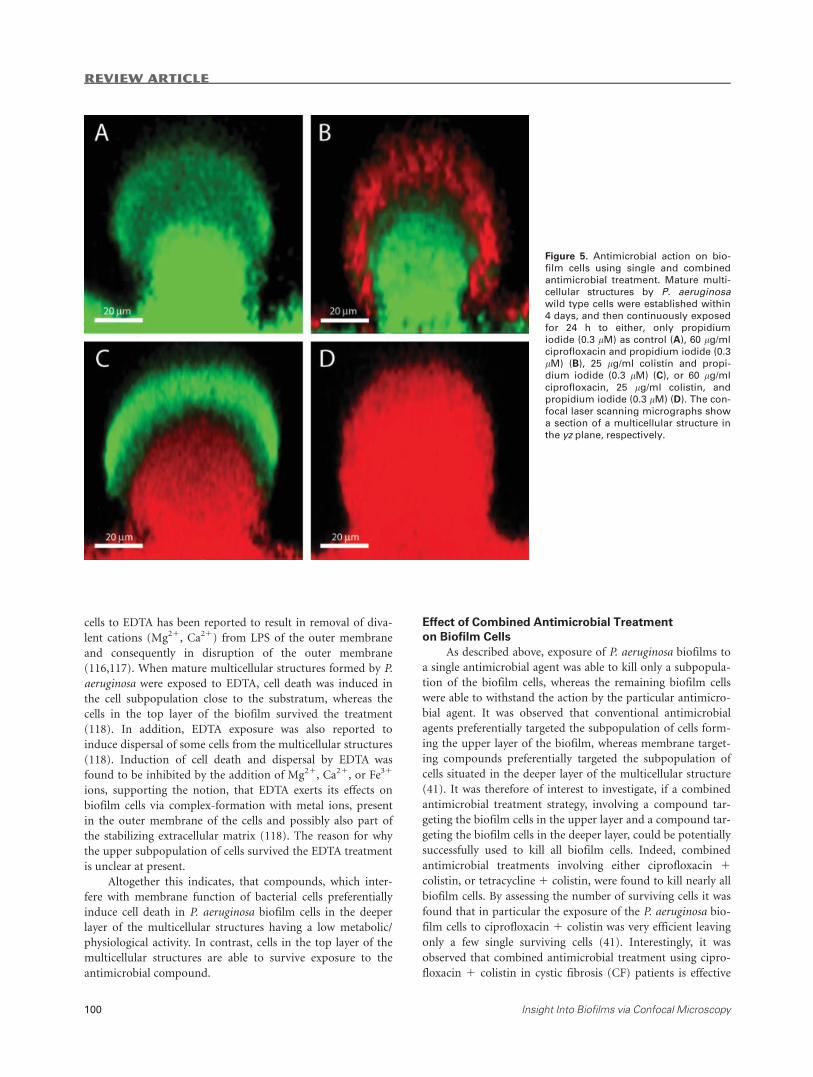
in the cap-forming cell subpopulation (Fig. 5B) (41). Using in
situ gene expression analysis, involving a growth activity-de-
pendent fluorescent reporter (see details about the fluorescent
reporter above), it was found that ciprofloxacin specifically
targets the P. aeruginosa biofilm cells exhibiting high metabolic
activity in the upper part of the multicellular structures. By
contrast, the biofilm cells in the deeper layers exhibiting low
metabolic activity survive ciprofloxacin treatment (Fig. 5B)
(41). A similar phenotype with respect to the distribution of
growth activity and ciprofloxacin-induced cell death was
observed for a P. aeruginosa colony biofilm (99).
Tetracycline is an antimicrobial agent, which originates
from secondary metabolites produced by Streptomyces spp. It
can inhibit bacterial protein synthesis by preventing attach-
ment of aminoacyl-tRNA to the ribosomal acceptor site (A-
site) and thereby induce cell death. When mature P. aeruginosa
wild type biofilms were exposed to tetracycline, the cells of the
cap-forming subpopulation were killed, whereas the cells
situated in the deeper areas survived the treatment (41). An
experiment involving treatment of a biofilm formed by a
strain, which harbors the growth activity-dependent fluores-
cent reporter fusion, indicated that tetracycline preferentially
kills the cells in the upper area, which exhibit high metabolic
activity. By contrast, cells in the deeper areas of the biofilm,
which exhibit a lower metabolic activity, were not killed by
tetracycline (S.J.P. and T.T.N., unpublished observation).
The aminoglycoside tobramycin is a secondary metabo-
lite derived from Streptomyces spp. and it can inhibit protein
synthesis in Gram-negative bacteria by preventing transloca-
tion of peptidyl-tRNA from the A-site to the P-site of the ribo-
some, and thereby induce cell death. When mature P. aerugi-
nosa biofilms were exposed to tobramycin, the cells situated in
the upper area of the multicellular structures were killed,
whereas the cells situated in the deeper areas survived the
treatment (36,100). The efficiency of tobramycin-induced kill-
ing in P. aeruginosa biofilms was increased by co-administra-
tion of furanone C-30, a compound, which was identified to
inhibit quorum sensing regulated gene expression (100). A
biofilm formed by a mutant strain, which was defective in
las- and rhl-mediated cell-to-cell-communication showed
increased sensitivity to tobramycin, indicating a possible role
of quorum sensing in tolerance towards tobramycin (36). Mah
et al. presented results, which indicate that in biofilm cells of
strain P. aeruginosa PA14 periplasmatic glucans might seques-
ter tobramycin and hence prevent the interaction of tobramy-
cin with its target (101). An ndvB-mutant, which is deficient
in the synthesis of periplasmatic glucans, exhibited increased
sensitivity to tobramycin in biofilms (101).
Altogether this indicates, that conventional antimicrobial
compounds, which interfere with fundamental physiological
processes of bacterial cells preferentially induce cell death in
P. aeruginosa biofilm cells that have a high metabolic/physio-
logical activity in the top layer of the multicellular structures.
In contrast, cells that have a low metabolic/physiological
activity are able to survive exposure to the antimicrobial
compound.
REVIEW ARTICLE
98 Insight Into Biofilms via Confocal Microscopy
Effect of Membrane-Targeting Compounds
on Biofilm Cells
The rise in appearance of multiresistant bacteria and the
persistence of biofilms in medical settings has increased the in-
terest in alternative antimicrobial compounds. Among the
new potential antimicrobial therapeutics are antimicrobial
peptides, as resistance to antimicrobial peptides has rarely
been observed so far (102–104). In addition, antimicrobial
peptides are promising new drugs due to their ability to mod-
ulate immune responses (103,105). Antimicrobial peptides are
found to exert their primary antibacterial activities by interfer-
ing with the bacterial membrane, resulting in leakage and
eventual death of the bacterial cell (e.g., 106,107). Here we
describe the effect of the antimicrobial peptide colistin (poly-
myxin E) on P. aeruginosa biofilms. In addition we describe
the effects of two other membrane-targeting compounds on P.
aeruginosa biofilms, namely the detergent sodium dodecyl sul-
fate (SDS) and the chelator ethylenediaminetetraacetic acid
(EDTA).
Colistin belongs to the polymyxin group of antimicrobial
peptides, and it is synthesized by strains of Paenibacillus spp.,
such as P. polymyxa (107–111). Colistin is administered as
treatment against infections caused by Gram-negative bacteria,
for example, pulmonary infections caused by P. aeruginosa in
CF patients, or sepsis, wound infections, and urinary tract
infections caused by a variety of Gram-negative bacteria (112–
115). When mature mushroom-like shaped P. aeruginosa bio-
films were exposed to a clinically relevant concentration of
colistin, only the stalk-forming cell subpopulation was killed,
whereas the cap-forming subpopulation survived the treat-
ment (Fig. 5C) (41,42). Detailed investigations on the spatio-
temporal-dependent effects of colistin exposure on P. aerugi-
nosa biofilms using fluorescent reporters and knock-out
mutants provided evidence, that the metabolic/physiological
active cells of the cap-subpopulation were able to adapt to
colistin by inducing two antimicrobial tolerance mechanisms:
the polymyxin resistance (pmr) LPS-modification system, and
the antimicrobial efflux pump MexAB-OprM (41). In con-
trast, cells with low metabolic/physiological activity in the dee-
per layer of the multicellular structures were not able to adapt
to colistin exposure, and were therefore killed by the action of
the antimicrobial peptide (41). Although tolerance develop-
ment in initial multicellular structures (2-day-grown bio-
films) in addition appeared to be dependent on type IV pili
driven motility (42), cellular migration evidently does not
play a role in colistin-tolerance development in mature multi-
cellular structures (4-day-grown biofilms) (41). Interestingly,
the observed spatial distribution of live and dead cells upon
colistin exposure appears to be independent of the actual
three-dimensional structure of the biofilm and the carbon
source used for biofilm-growth. Mushroom-shaped as well as
irregular-shaped glucose-grown mutant biofilms, and flat-
structured citrate-grown biofilms exhibited a subpopulation
of dead cells close to the substratum and a subpopulation of
live cells in the upper cell layer of the multicellular structures
upon colistin treatment (41,42, and J. Haagensen, personal
communication).
SDS is a synthetic anionic surfactant, and this compound
and its derivatives can be found in many household products
(e.g., soaps). Due to its amphiphilic characteristics SDS inter-
feres with biological membranes and is also known for its abil-
ity to denature proteins. When initial and mature multicellu-
lar structures formed by P. aeruginosa were exposed to SDS,
cell death was induced in the cell subpopulation close to the
substratum, whereas the cells in the top layer of the biofilm
survived the treatment (42). Another study reported disrup-
tion of P. aeruginosa biofilms via hollowing after exposure to
SDS (98). However, as the reported experiment was performed
in the absence of a fluorescent indicator for dead cells (e.g.,
propidium iodide) it is unclear whether cells from the interior
area of the biofilm detached or were killed. The genetic deter-
minants and mechanisms, which facilitate tolerance develop-
ment of the surviving fraction of cells are unknown at present,
and are subject to ongoing research in our laboratory.
EDTA has the ability to form complexes with metal ions,
such as Mg21, Ca21 and Fe31. Exposure of proteobacterial
Figure 4. Determination of metabolic/physiological activity during development of multicellular structures using a growth rate dependent
fluorescent reporter. P. aeruginosa wild type cells harboring a single chromosomal copy of Tn7-rrnBP1-gfp[AGA], expressing a Gfp versionwith a short half life under the control of a ribosomal promoter (A), and, as control, P. aeruginosa wild type cells harboring a single chro-mosomal copy of Tn7-rrnBP1-gfp, expressing a stable version of Gfp under the control of a ribosomal promoter (B), were grown as biofilmand confocal laser scanning micrographs acquired every 24 h for 4 days. The confocal laser scanning micrographs represent sections of
multicellular structures in the yz plane. Reproduced from Mol Microbiol 68:223--240 with permission from John Wiley & Sons, Inc. [Color
figure can be viewed in the online issue, which is available at www.interscience.wiley.com.]
REVIEW ARTICLE
Cytometry Part A � 75A: 90�103, 2009 99
cells to EDTA has been reported to result in removal of diva-
lent cations (Mg21, Ca21) from LPS of the outer membrane
and consequently in disruption of the outer membrane
(116,117). When mature multicellular structures formed by P.
aeruginosa were exposed to EDTA, cell death was induced in
the cell subpopulation close to the substratum, whereas the
cells in the top layer of the biofilm survived the treatment
(118). In addition, EDTA exposure was also reported to
induce dispersal of some cells from the multicellular structures
(118). Induction of cell death and dispersal by EDTA was
found to be inhibited by the addition of Mg21, Ca21, or Fe31
ions, supporting the notion, that EDTA exerts its effects on
biofilm cells via complex-formation with metal ions, present
in the outer membrane of the cells and possibly also part of
the stabilizing extracellular matrix (118). The reason for why
the upper subpopulation of cells survived the EDTA treatment
is unclear at present.
Altogether this indicates, that compounds, which inter-
fere with membrane function of bacterial cells preferentially
induce cell death in P. aeruginosa biofilm cells in the deeper
layer of the multicellular structures having a low metabolic/
physiological activity. In contrast, cells in the top layer of the
multicellular structures are able to survive exposure to the
antimicrobial compound.
Effect of Combined Antimicrobial Treatment
on Biofilm Cells
As described above, exposure of P. aeruginosa biofilms to
a single antimicrobial agent was able to kill only a subpopula-
tion of the biofilm cells, whereas the remaining biofilm cells
were able to withstand the action by the particular antimicro-
bial agent. It was observed that conventional antimicrobial
agents preferentially targeted the subpopulation of cells form-
ing the upper layer of the biofilm, whereas membrane target-
ing compounds preferentially targeted the subpopulation of
cells situated in the deeper layer of the multicellular structure
(41). It was therefore of interest to investigate, if a combined
antimicrobial treatment strategy, involving a compound tar-
geting the biofilm cells in the upper layer and a compound tar-
geting the biofilm cells in the deeper layer, could be potentially
successfully used to kill all biofilm cells. Indeed, combined
antimicrobial treatments involving either ciprofloxacin 1colistin, or tetracycline 1 colistin, were found to kill nearly all
biofilm cells. By assessing the number of surviving cells it was
found that in particular the exposure of the P. aeruginosa bio-
film cells to ciprofloxacin 1 colistin was very efficient leaving
only a few single surviving cells (41). Interestingly, it was
observed that combined antimicrobial treatment using cipro-
floxacin 1 colistin in cystic fibrosis (CF) patients is effective
Figure 5. Antimicrobial action on bio-
film cells using single and combined
antimicrobial treatment. Mature multi-
cellular structures by P. aeruginosawild type cells were established within
4 days, and then continuously exposed
for 24 h to either, only propidium
iodide (0.3 lM) as control (A), 60 lg/mlciprofloxacin and propidium iodide (0.3
lM) (B), 25 lg/ml colistin and propi-
dium iodide (0.3 lM) (C), or 60 lg/mlciprofloxacin, 25 lg/ml colistin, andpropidium iodide (0.3 lM) (D). The con-focal laser scanning micrographs show
a section of a multicellular structure in
the yz plane, respectively.
REVIEW ARTICLE
100 Insight Into Biofilms via Confocal Microscopy

in reducing the onset of chronic P. aeruginosa infection (119),
and hence this particular treatment strategy was recently
implemented in the recommended early intervention and pre-
vention therapy in CF (120,121).
CONCLUSION AND FUTURE PROSPECTS
The biofilm mode of living is assumed to be the predomi-
nant bacterial life style in nature. These complex microbial
communities carry out processes in the natural environment as
well as man-made environments, and thereby contribute to
human development, health and disease. Investigations of bio-
films established under controlled conditions in laboratory set-
ups have provided fascinating insights into the fundamental
capacities of bacteria to form multicellular structures. Studies of
model systems, such as flow-chamber-grown biofilms, have
revealed a set of inherent elements of the participating microbes
that can facilitate their organization into multicellular commu-
nities. Among these factors are for example the production of
matrix compounds, cell-surface bound proteins, the secretion
of biosurfactants, cellular migration, and regulatory elements
such as signal transduction systems, and intra- and extra-cellu-
lar signal messenger molecules. CLSM monitoring of fluores-
cently color-coded bacteria, grown in mixed-strain biofilms
under continuous hydrodynamic conditions, has provided in-
triguing insights into the spatiotemporal developmental pro-
cesses, in some cases down to the single cell level (e.g., 30,31).
Detailed microscopic examinations have revealed that, even in
monospecies biofilms, a number of physiologically distinct cell
subpopulations exist and differentiate during biofilm develop-
ment (e.g., 41). Intriguingly, recent investigations have revealed
that these distinct cell subpopulations exhibit differential sensi-
tivity towards antimicrobial agents (e.g., 41,42). One cell subpo-
pulation, which exhibited high metabolic/physiological activity,
was sensitive towards conventional antimicrobial agents,
whereas the second subpopulation was refractory (41). How-
ever, the second cell subpopulation exhibited increased sensitiv-
ity towards a number of membrane-targeting compounds, and
hence a combined antimicrobial treatment using a conventional
antibiotic and a membrane-targeting compound was able to kill
both cell subpopulations of the biofilm (41). These, and other
studies highlight the importance of studying the characteristics
of biofilm cells at the subpopulation, and single cell level.
Future studies, facilitated by sophisticated approaches
and new technologies, will increase our understanding of mi-
crobial life in multicellular communities. Investigations of spa-
tiotemporal gene expression in biofilms by the use of fluores-
cent reporter genes and CLSM will continue, and expand
through the use of combinations of gene expression markers.
Cell subpopulations will be isolated from complex commu-
nities, for example, by the use of microdissection, microfluidic
devices or cell sorting, and subjected to further analysis, such
as RT-PCR measurements of specific gene expression or DNA
array analysis of global gene expression (e.g., Ref. 122). More-
over, new technologies are emerging that might enable analysis
of global transcription profiles of single biofilm cells
(123,124). Studies of the metabolic functions of subpopula-
tions of biofilm cells or single cells in a biofilm community,
might be done by the use of NanoSIMS (nanometer-scale sec-
ondary-ion mass spectrometry) or related techniques, which
have recently been successfully employed on microbial cells
from the environment and microbial cells associated with ani-
mals (e.g., 125,126). Insight into the features of distinct cell
subpopulations and single cells in microbial communities will
provide a more complete understanding of the microbial mul-
ticellular lifestyle, and open up new strategies to manipulate
harmful biofilms to restore and maintain human well-being.
ACKNOWLEDGMENTS
The authors acknowledge the scientific contributions of
their colleagues at the Technical University of Denmark, and
elsewhere.
LITERATURE CITED
1. Costerton JW, Lewandowski Z, Caldwell DE, Korber DR, Lappin-Scott HM. Micro-bial biofilms. Annu Rev Microbiol 1995;49:711–745.
2. Wilson M. Microbial Inhabitants of Humans: Their Ecology and Role in Health andDisease. Cambridge: Cambridge University Press; 2005.
3. Marshall K. Interfaces in Microbial Ecology. Cambridge: Harvard University Press;1976.
4. Davey ME, O’Toole GA. Microbial biofilms: From ecology to molecular genetics.Microbiol Mol Biol Rev 2000;64(4):847–867.
5. Hall-Stoodley L, Costerton JW, Stoodley P. Bacterial biofilms: From the naturalenvironment to infectious diseases. Nat Rev Microbiol 2004;2(2):95–108.
6. Costerton JW, Cheng KJ, Geesey GG, Ladd TI, Nickel JC, Dasgupta M, Marrie TJ.Bacterial biofilms in nature and disease. Annu Rev Microbiol 1987;41:435–464.
7. Pamp SJ, Gjermansen M, Tolker-Nielsen T. The biofilm matrix—A sticky frame-work. In: Kjelleberg S, Givskov M, editors. The Biofilm Mode of Life. Norfolk: Hori-zon Bioscience; 2007. pp 37–69.
8. Flint HJ, Bayer EA, Rincon MT, Lamed R, White BA. Polysaccharide utilization bygut bacteria: Potential for new insights from genomic analysis. Nat Rev Microbiol2008;6(2):121–131.
9. Flint HJ, Duncan SH, Scott KP, Louis P. Interactions and competition within the mi-crobial community of the human colon: Links between diet and health. EnvironMicrobiol 2007;9(5):1101–1111.
10. Marsh PD, Bowden GHW. Microbial community interactions in biofilms. In: AllisonDG, Gilbert P, Lappin-Scott HM, Wilson M, editors. Community Structure and Co-operation in Biofilms. Cambridge: Cambridge University Press; 2000. pp 167–198.
11. Molin S, Tolker-Nielsen T. Gene transfer occurs with enhanced efficiency in biofilmsand induces enhanced stabilisation of the biofilm structure. Curr Opin Biotechnol2003;14(3):255–261.
12. Marshall KC. Biofilms: An overview of bacterial adhesion, activity, and control atsurfaces. ASM News 1992;58(4):202–207.
13. Dethlefsen L, McFall-Ngai M, Relman DA. An ecological and evolutionary perspec-tive on human-microbe mutualism and disease. Nature 2007;449(7164):811–818.
14. Macfarlane S, Dillon JF. Microbial biofilms in the human gastrointestinal tract.J Appl Microbiol 2007;102(5):1187–1196.
15. Berg RD. The indigenous gastrointestinal microflora. Trends Microbiol 1996;4(11):430–435.
16. Noverr MC, Huffnagle GB. The ‘‘microflora hypothesis’’ of allergic diseases. ClinExp Allergy 2005;35(12):1511–1520.
17. Costerton JW, Stewart PS, Greenberg EP. Bacterial biofilms: A common cause ofpersistent infections. Science 1999;284(5418):1318–1322.
18. Costerton W, Veeh R, Shirtliff M, Pasmore M, Post C, Ehrlich G. The application ofbiofilm science to the study and control of chronic bacterial infections. J Clin Invest2003;112(10):1466–1477.
19. Parsek MR, Singh PK. Bacterial biofilms: An emerging link to disease pathogenesis.Annu Rev Microbiol 2003;57:677–701.
20. Fux CA, Costerton JW, Stewart PS, Stoodley P. Survival strategies of infectious bio-films. Trends Microbiol 2005;13(1):34–40.
21. Christensen BB, Sternberg C, Andersen JB, Palmer RJ, Nielsen AT, Givskov M, SørenM. Molecular tools for study of biofilm physiology. In: Doyle RJ, editor. Methods inEnzymology, Vol. 310: Biofilms. San Diego: Academic Press; 1999. pp 20–42.
22. Sternberg C, Tolker-Nielsen T. Growing and Analyzing Biofilms in Flow Cells. In:Coico R, Kowalik T, Quarles J, Stevenson B, Taylor R, editors. Current Protocols inMicrobiology, Vol. 1B. 2. New Jersey: John Wiley; 2005. pp 1B. 2.1–1B. 2.15.
23. Branda SS, Vik S, Friedman L, Kolter R. Biofilms: The matrix revisited. TrendsMicrobiol 2005;13(1):20–26.
24. Palmer RJ. Microscopy flow cells: Perfusion chambers for real-time study of bio-films. In: Doyle RJ, editor. Methods in Enzymology, Vol.310: Biofilms. San Diego:Academic Press; 1999. pp 160–166.
25. Palmer RJ, Haagensen JA, Neu TR, Sternberg C. Conforcal microscopy of biofilms—statiotemporal approaches. In: Pawley JB, editor. Handbook of Biological ConfocalMicroscopy. New York: SpringerScience1Business Media; 2006. pp 882–900.
REVIEW ARTICLE
Cytometry Part A � 75A: 90�103, 2009 101

26. Wolfaardt GM, Lawrence JR, Robarts RD, Caldwell SJ, Caldwell DE. Multicellularorganization in a degradative biofilm community. Appl Environ Microbiol1994;60(2):434–446.
27. Center for Systems Microbiology. Biofilm flow-cell setup. Lyngby, Denmark: Centerfor Systems Microbiology, Technical University of Denmark; 2005. http://www.dtu.dk/centre/CSM/English/Instrument%20Center/Resources/Biofilm%20Setup.aspx.
28. Zinn MS, Kirkegaard RD, Palmer RJ, White DC. Laminar flow chamber for continu-ous monitoring of biofilm formation and succession. In: Doyle RJ, editor.Methodsof Enzymology, Vol. 310: Biofilms. San Diego: Academic Press; 1999. pp 224–232.
29. Klausen M, Heydorn A, Ragas P, Lambertsen L, Aaes-Jorgensen A, Molin S, Tolker-Nielsen T. Biofilm formation by Pseudomonas aeruginosa wild type, flagella and typeIV pili mutants. Mol Microbiol 2003;48(6):1511–1524.
30. Klausen M, Aaes-Jorgensen A, Molin S, Tolker-Nielsen T. Involvement of bacterialmigration in the development of complex multicellular structures in Pseudomonasaeruginosa biofilms. Mol Microbiol 2003;50(1):61–68.
31. Pamp SJ, Tolker-Nielsen T. Multiple roles of biosurfactants in structural biofilmdevelopment by Pseudomonas aeruginosa. J Bacteriol 2007;189(6):2531–2539.
32. Hansen SK, Rainey PB, Haagensen JA, Molin S. Evolution of species interactions ina biofilm community. Nature 2007;445(7127):533–536.
33. Christensen BB, Haagensen JA, Heydorn A, Molin S. Metabolic commensalism andcompetition in a two-species microbial consortium. Appl Environ Microbiol2002;68(5):2495–2502.
34. Nielsen AT, Tolker-Nielsen T, Barken KB, Molin S. Role of commensal relationshipson the spatial structure of a surface-attached microbial consortium. Environ Micro-biol 2000;2(1):59–68.
35. Breugelmans P, Barken KB, Tolker-Nielsen T, Hofkens J, Dejonghe W, Springael D.Architecture and spatial organization in a triple-species bacterial biofilm synergisti-cally degrading the phenylurea herbicide linuron. FEMS Microbiol Ecol2008;64(2):271–282.
36. Bjarnsholt T, Jensen PO, Burmolle M, Hentzer M, Haagensen JA, Hougen HP,Calum H, Madsen KG, Moser C, Molin S, Høiby N, Givskov M. Pseudomonas aeru-ginosa tolerance to tobramycin, hydrogen peroxide and polymorphonuclear leuko-cytes is quorum-sensing dependent. Microbiology 2005;151 (Pt 2):373–383.
37. Wolfaardt GM, Lawrence JR, Robarts RD, Caldwell DE. The role of interactions, ses-sile growth and nutrient amendments on the degradative efficiency of a microbialconsortium. Can J Microbiol 1994;40(5):331–340.
38. Hansen SK, Haagensen JA, Gjermansen M, Jorgensen TM, Tolker-Nielsen T, MolinS. Characterization of a Pseudomonas putida rough variant evolved in a mixed-spe-cies biofilm with Acinetobacter sp. strain C6. J Bacteriol 2007;189(13):4932–4943.
39. Kuhl M, Rickelt LF, Thar R. Combined imaging of bacteria and oxygen in biofilms.Appl Environ Microbiol 2007;73(19):6289–6295.
40. Landry RM, An D, Hupp JT, Singh PK, Parsek MR. Mucin-Pseudomonas aeruginosainteractions promote biofilm formation and antibiotic resistance. Mol Microbiol2006;59(1):142–151.
41. Pamp SJ, Gjermansen M, Johansen HK, Tolker-Nielsen T. Tolerance to the antimi-crobial peptide colistin in Pseudomonas aeruginosa biofilms is linked to metaboli-cally active cells, and depends on the pmr and mexAB-oprM genes. Mol Microbiol2008;68(1):223–240.
42. Haagensen JA, Klausen M, Ernst RK, Miller SI, Folkesson A, Tolker-Nielsen T, MolinS. Differentiation and distribution of colistin- and sodium dodecyl sulfate-tolerantcells in Pseudomonas aeruginosa biofilms. J Bacteriol 2007;189(1):28–37.
43. Neu TR, Lawrence JR. One-photon versus two-photon laser scanning microscopyand digital image analysis of microbial biofilms. MethodsMicrobiol 2005;34:89–136.
44. Neu TR, Walczysko P, Lawrence JR. Two-photon imaging for studying the microbialecology of biofilm systems. Microbes Environ 2004;19:1–6.
45. Dyba M, Keller J, Hell SW. Phase filter enhanced STED-4Pi fluorescence microscopy:Theory and experiment. New J Phys 2005;134:1–21.
46. Harke B, Keller J, Ullal CK, Westphal V, Schonle A, Hell SW. Resolution scaling inSTED microscopy. Opt Exp 2008;16(6):4154–4162.
47. Chalfie M, Tu Y, Euskirchen G, Ward WW. Green fluorescent protein as a markerfor gene expression. Science 1994;263(5148):802–805.
48. Caldwell DE, Korber DR, Lawrence JR. Imaging of bacterial cells by fluorescenceexclusion using scanning confocal laser microscopy. J Microbiol Methods1992(15):249–261.
49. Lawrence JR, Korber DR, Hoyle BD, Costerton JW, Caldwell DE. Optical sectioningof microbial biofilms. J Bacteriol 1991;173(20):6558–6567.
50. Shi L, Gunther S, Hubschmann T, Wick LY, Harms H, Muller S. Limits of propi-dium iodide as a cell viability indicator for environmental bacteria. Cytometry A2007;71A:592–598.
51. Molecular Probes. Sheet mp07007.http://probes.invitrogen.com/media/pis/.
52. Lawrence JR, Neu TR, Swerhone GDW. Application of multiple parameter imagingfor the quantification of algal, bacterial and exopolymer components of microbialbiofilms. J Microbiol Methods 1998;32:253–261.
53. Strathmann M, Wingender J, Flemming HC. Application of fluorescently labelledlectins for the visualization and biochemical characterization of polysaccharides inbiofilms of Pseudomonas aeruginosa.. J Microbiol Methods 2002;50(3):237–248.
54. Yang L, Haagensen JAJ, Jelsbak L, Johansen HK, Sternberg C, Høiby N, Molin S. Insitu growth rates and biofilm development of Pseudomonas aeruginosa populationsin chronic lung infections. J Bacteriol 2008;190(8):2767–2776.
55. Stewart PS, Murga R, Srinivasan R, de Beer D. Biofilm structural heterogenecityvisualized by three microscopic methods. Water Res 1995;8:2006–2009.
56. Allesen-Holm M, Barken KB, Yang L, Klausen M, Webb JS, Kjelleberg S, Molin S,Givskov M, Tolker-Nielsen T. A characterization of DNA release in Pseudomonas aer-uginosa cultures and biofilms. Mol Microbiol 2006;59(4):1114–1128.
57. Lambertsen L, Sternberg C, Molin S. Mini-Tn7 transposons for site-specific taggingof bacteria with fluorescent proteins. Environ Microbiol 2004;6(7):726–732.
58. Sternberg C, Christensen BB, Johansen T, T. NA, Andersen JB, Givskov M, Molin S.Distribution of bacterial growth activity in flow-chamber biofilms. Appl EnvironMicrobiol 1999;65(9):4108–4117.
59. Tolker-Nielsen T, Brinch UC, Ragas PC, Andersen JB, Jacobsen CS, Molin S. Devel-opment and dynamics of Pseudomonas sp. Biofilms. J Bacteriol 2000;182(22):6482–6489.
60. Kjaergaard K, Schembri MA, Ramos C, Molin S, Klemm P. Antigen 43 facilitates for-mation of multispecies biofilms. Environ Microbiol 2000;2(6):695–702.
61. Møller S, Pedersen AR, Poulsen LK, Arvin E, Molin S. Activity and three-dimen-sional distribution of toluene-degrading Pseudomonas putida in a multispecies bio-film assessed by quantitative in situ hybridization and scanning confocal laser mi-croscopy. Appl Environ Microbiol 1996;62(12):4632–4640.
62. Stahl DA, Amann R. Development and application of nucleic acid probes. In: Stack-ebrandt E, Goodfellow M, editors. Nucleic Acid Techniques in Bacterial Systematics.New York: John Wiley; 1991. pp 205–248.
63. Amann R, Fuchs BM. Single-cell identification in microbial communities byimproved fluorescence in situ hybridization techniques. Nat Rev Microbiol 2008;6(5):339–348.
64. Hell SW, Reiner G, Cremer C, Stelzer EHK. Aberrations in confocal fluorescence mi-croscopy induced by mistakes in refractive index. J Microsc 1993;169:391–405.
65. Yang X, Beyenal H, Harkin G, Lewandowski Z. Quantifying biofilm structure usingimage analysis. J Microbiol Methods 2000;39(2):109–119.
66. Heydorn A, Ersbøll BK, Hentzer M, Parsek MR, Givskov M, Molin S. Experimentalreproducibility in flow-chamber biofilms. Microbiology 2000;146:2409–2415.
67. Xavier JB, White DC, Almeida JS. Automated biofilm morphology quantificationfrom confocal laser scanning microscopy imaging. Water Sci Technol 2003;47(5):31–37.
68. Merod RT, Warren JE, McCaslin H, Wuertz S. Toward automated analysis of biofilmarchitecture: Bias caused by extraneous confocal laser scanning microscopy images.Appl Environ Microbiol 2007;73(15):4922–4930.
69. Mueller LN, de Brouwer JF, Almeida JS, Stal LJ, Xavier JB. Analysis of a marinephototrophic biofilm by confocal laser scanning microscopy using the new imagequantification software PHLIP. BMC Ecol 2006;6:1.
70. Mils AL, Powelson DK. Bacterial interactions with surfaces in soil. In: Fletcher M,editor. Bacterial Adhesion: Molecular and Ecological Diversity. New York: JohnWiley; 1996. pp 25–57.
71. O’Toole GA, Kolter R. Flagellar and twitching motility are necessary for Pseudomo-nas aeruginosa biofilm development. Mol Microbiol 1998;30(2):295–304.
72. Sauer K, Camper AK, Ehrlich GD, Costerton JW, Davies DG. Pseudomonas aerugi-nosa displays multiple phenotypes during development as a biofilm. J Bacteriol2002;184(4):1140–1154.
73. Caiazza NC, O’Toole GA. SadB is required for the transition from reversible to irre-versible attachment during biofilm formation by Pseudomonas aeruginosa PA14.J Bacteriol 2004;186(14):4476–4485.
74. Ramphal R, Sadoff JC, Pyle M, Silipigni JD. Role of pili in the adherence of Pseu-domonas aeruginosa to injured tracheal epithelium. Infect Immun 1984;44(1):38–40.
75. Deziel E, Comeau Y, Villemur R. Initiation of biofilm formation by Pseudomonasaeruginosa 57RP correlates with emergence of hyperpiliated and highly adherentphenotypic variants deficient in swimming, swarming, and twitching motilities. JBacteriol 2001;183(4):1195–1204.
76. Chiang P, Burrows LL. Biofilm formation by hyperpiliated mutants of Pseudomonasaeruginosa.. J Bacteriol 2003;185(7):2374–2378.
77. Vallet I, Olson JW, Lory S, Lazdunski A, Filloux A. The chaperone/usher pathwaysof Pseudomonas aeruginosa: Identification of fimbrial gene clusters (cup) and theirinvolvement in biofilm formation. Proc Natl Acad Sci USA 2001;98(12):6911–6916.
78. Whitchurch CB, Tolker-Nielsen T, Ragas PC, Mattick JS. Extracellular DNA requiredfor bacterial biofilm formation. Science 2002;295(5559):1487.
79. Singh PK, Parsek MR, Greenberg EP, Welsh MJ. A component of innate immunityprevents bacterial biofilm development. Nature 2002;417(6888):552–555.
80. Shrout JD, Chopp DL, Just CL, Hentzer M, Givskov M, Parsek MR. The impact ofquorum sensing and swarming motility on Pseudomonas aeruginosa biofilm forma-tion is nutritionally conditional. Mol Microbiol 2006;62(5):1264–1277.
81. Movies: Flat-structured P. aeruginosa biofilm. http://www.dtu.dk/centre/CSM/English/Instrument%20Center/Supplementary%20Data/2003%20-%20Klausen%201.aspx.
82. Movies: Mushroom-like structured P. aeruginosa biofilm. http://www.dtu.dk/centre/CSM/English/Instrument%20Center/Supplementary%20Data/2003%20-%20Klausen%202.aspx.
83. Barken KB, Pamp SJ, Yang L, Gjermansen M, Bertrand JJ, Klausen M, Givskov M,Whitchurch CB, Engel JN, Tolker-Nielsen T. Roles of type IV pili, flagellum-mediatedmotility and extracellular DNA in the formation of mature multicellular structuresin Pseudomonas aeruginosa biofilms. Environ Microbiol 2008;10(9):2331–2343.
84. Kohler T, Curty LK, Barja F, van Delden C, Pechere JC. Swarming of Pseudomonasaeruginosa is dependent on cell-to-cell signaling and requires flagella and pili. J Bac-teriol 2000;182(21):5990–5996.
85. Wozniak DJ, Wyckoff TJ, Starkey M, Keyser R, Azadi P, O’Toole GA, Parsek MR. Al-ginate is not a significant component of the extracellular polysaccharide matrix ofPA14 and PAO1 Pseudomonas aeruginosa biofilms. Proc Natl Acad Sci USA2003;100(13):7907–7912.
86. Govan JR, Deretic V. Microbial pathogenesis in cystic fibrosis: Mucoid Pseudomonasaeruginosa and Burkholderia cepacia. Microbiol Rev 1996;60(3):539–574.
REVIEW ARTICLE
102 Insight Into Biofilms via Confocal Microscopy

87. Jackson KD, Starkey M, Kremer S, Parsek MR, Wozniak DJ. Identification of psl, alocus encoding a potential exopolysaccharide that is essential for Pseudomonas aeru-ginosa PAO1 biofilm formation. J Bacteriol 2004;186(14):4466–4475.
88. Matsukawa M, Greenberg EP. Putative exopolysaccharide synthesis genes influencePseudomonas aeruginosa biofilm development. J Bacteriol 2004;186(14):4449–4456.
89. Friedman L, Kolter R. Two genetic loci produce distinct carbohydrate-rich structuralcomponents of the Pseudomonas aeruginosa biofilm matrix. J Bacteriol2004;186(14):4457–4465.
90. Friedman L, Kolter R. Genes involved in matrix formation in Pseudomonas aerugi-nosa PA14 biofilms. Mol Microbiol 2004;51(3):675–690.
91. Aas FE, Wolfgang M, Frye S, Dunham S, Lovold C, Koomey M. Competence for nat-ural transformation in Neisseria gonorrhoeae: Components of DNA binding anduptake linked to type IV pilus expression. Mol Microbiol 2002;46(3):749–760.
92. van Schaik EJ, Giltner CL, Audette GF, Keizer DW, Bautista DL, Slupsky CM, SykesBD, Irvin RT. DNA binding: A novel function of Pseudomonas aeruginosa type IVpili. J Bacteriol 2005;187(4):1455–1464.
93. Kulasekara HD, Ventre I, Kulasekara BR, Lazdunski A, Filloux A, Lory S. A noveltwo-component system controls the expression of Pseudomonas aeruginosa fimbrialcup genes. Mol Microbiol 2005;55(2):368–380.
94. Kuchma SL, Connolly JP, O’Toole GA. A three-component regulatory system regu-lates biofilm maturation and type III secretion in Pseudomonas aeruginosa.. J Bacter-iol 2005;187(4):1441–1454.
95. Gjermansen M, Ragas P, Sternberg C, Molin S, Tolker-Nielsen T. Characterization ofstarvation-induced dispersion in Pseudomonas putida biofilms. Environ Microbiol2005;7(6):894–906.
96. Gjermansen M, Ragas P, Tolker-Nielsen T. Proteins with GGDEF and EAL domainsregulate Pseudomonas putida biofilm formation and dispersal. FEMS Microbiol Lett2006;265(2):215–224.
97. Purevdorj-Gage B, Costerton WJ, Stoodley P Phenotypic differentiation and seedingdispersal in non-mucoid and mucoid Pseudomonas aeruginosa biofilms. Microbiol-ogy 2005;151 (Pt 5):1569–1576.
98. Boles BR, Thoendel M, Singh PK. Rhamnolipids mediate detachment of Pseudomo-nas aeruginosa from biofilms. Mol Microbiol 2005;57(5):1210–1223.
99. Walters MC, 3rd, Roe F, Bugnicourt A, Franklin MJ, Stewart PS. Contributions ofantibiotic penetration, oxygen limitation, and low metabolic activity to tolerance ofPseudomonas aeruginosa biofilms to ciprofloxacin and tobramycin. AntimicrobAgents Chemother 2003;47(1):317–323.
100. Hentzer M, Wu H, Andersen JB, Riedel K, Rasmussen TB, Bagge N, Kumar N,Schembri MA, Song Z, Kristoffersen P, Manefield M, Costerton JW, Molin S, EberlL, Steinberg P, Kjelleberg S, Høiby N, Givskov M. Attenuation of Pseudomonas aeru-ginosa virulence by quorum sensing inhibitors. EMBO J 2003;22(15):3803–3815.
101. Mah TF, Pitts B, Pellock B, Walker GC, Stewart PS, O’Toole GA. A genetic basis forPseudomonas aeruginosa biofilm antibiotic resistance. Nature 2003;426(6964):306–310.
102. Boman HG. Peptide antibiotics and their role in innate immunity. Annu Rev Immu-nol 1995;13:61–92.
103. Hancock RE. Cationic peptides: Effectors in innate immunity and novel antimicro-bials. Lancet Infect Dis 2001;1(3):156–164.
104. Peschel A, Sahl HG. The co-evolution of host cationic antimicrobial peptides andmicrobial resistance. Nat Rev Microbiol 2006;4(7):529–536.
105. Hancock RE, Sahl HG. Antimicrobial and host-defence peptides as new anti-infec-tive therapeutic strategies. Nat Biotechnol 2006;24(12):1551–1557.
106. Hancock RE, Chapple DS. Peptide antibiotics. Antimicrob Agents Chemother1999;43(6):1317–1323.
107. Storm DR, Rosenthal KS, Swanson PE. Polymyxin and related peptide antibiotics.Annu Rev Biochem 1977;46:723–763.
108. Ainsworth GC, Brown AM, Brownlee G. Aerosporin, and antibiotic produced byBacillus aerosporus Geer. Nature 1947;160:263.
109. Benedict RG, Langlykke AF. Antibiotic activity of Bacillus polymyxa. J Bacteriol1947;54:24–25.
110. Stansly PG, Shepherd RG, White HJ. Polymyxin: A new chemotherapeutic agent.Bull Johns Hopkins Hosp 1947;81:43–54.
111. Suzuki T, Hayashi K, Fujikawa K, Tsukamoto K. The chemical structure of poly-myxin E: The identities of polymyxin E1 with colistin a and of polymyxin E2 withcolistin B. J Biochem 1965;57:226–227.
112. Levin AS, Barone AA, Penco J, Santos MV, Marinho IS, Arruda EA, Manrique EI,Costa SF. Intravenous colistin as therapy for nosocomial infections caused by multi-drug-resistant Pseudomonas aeruginosa and Acinetobacter baumannii.. Clin InfectDis 1999;28(5):1008–1011.
113. Garnacho-Montero J, Ortiz-Leyba C, Jimenez-Jimenez FJ, Barrero-Almodovar AE,Garcia-Garmendia JL, Bernabeu-Wittel IM, Gallego-Lara SL, Madrazo-Osuna J.Treatment of multidrug-resistant Acinetobacter baumannii ventilator-associatedpneumonia (VAP) with intravenous colistin: A comparison with imipenem-suscep-tible VAP. Clin Infect Dis 2003;36(9):1111–1118.
114. Markou N, Apostolakos H, Koumoudiou C, Athanasiou M, Koutsoukou A, Alama-nos I, Gregorakos L. Intravenous colistin in the treatment of sepsis from multiresis-tant Gram-negative bacilli in critically ill patients. Crit Care 2003;7(5):R78–R83.
115. Karabinis A, Paramythiotou E, Mylona-Petropoulou D, Kalogeromitros A, KatsarelisN, Kontopidou F, Poularas I, Malamou-Lada H. Colistin for Klebsiella pneumoniae-associated sepsis. Clin Infect Dis 2004;38(1):e7–e9.
116. Hancock RE. Alterations in outer membrane permeability. Annu Rev Microbiol1984;38:237–264.
117. Tamber S, Hancock RE. The outer membranes of Pseudomonads. In: Ramos JL, edi-tor. Pseudomonas, Vol. 1. New York: Kluwer Academic; 2004. pp 575–601.
118. Banin E, Brady KM, Greenberg EP. Chelator-induced dispersal and killing of Pseudo-monas aeruginosa cells in a biofilm. Appl EnvironMicrobiol 2006;72(3):2064–2069.
119. Valerius NH, Koch C, Høiby N. Prevention of chronic Pseudomonas aeruginosa colo-nisation in cystic fibrosis by early treatment. Lancet 1991;338(8769):725–726.
120. Doring G, Høiby N. Early intervention and prevention of lung disease in cysticfibrosis: A European consensus. J Cyst Fibros 2004;3(2):67–91.
121. Hoiby N, Frederiksen B, Pressler T. Eradication of early Pseudomonas aeruginosainfection. J Cyst Fibros 2005;4 (Suppl 2):49–54.
122. Lenz AP, Williamson KS, Pitts B, Stewart PS, Franklin MJ. Localized gene expressionin Pseudomonas aeruginosa biofilms. Appl Environ Microbiol 2008;74(14):4463–4471.
123. Zhong JF, Chen Y, Marcus JS, Scherer A, Quake SR, Taylor CR, Weiner LP. A micro-fluidic processor for gene expression profiling of single human embryonic stem cells.Lab Chip 2008;8(1):68–74.
124. Marcus JS, Anderson WF, Quake SR. Microfluidic single-cell mRNA isolation andanalysis. Anal Chem 2006;78(9):3084–3089.
125. Behrens S, Losekann T, Pett-Ridge J, Weber PK, Ng WO, Stevenson BS, HutcheonID, Relman DA, Spormann AM. Linking microbial phylogeny to metabolic activityat the single-cell level by using enhanced element labeling-catalyzed reporter deposi-tion fluorescence in situ hybridization (EL-FISH) and NanoSIMS. Appl EnvironMicrobiol 2008;74(10):3143–3150.
126. Lechene CP, Luyten Y, McMahon G, Distel DL. Quantitative imaging of nitrogen fixa-tion by individual bacteria within animal cells. Science 2007;317(5844):1563–1566.
REVIEW ARTICLE
Cytometry Part A � 75A: 90�103, 2009 103