inhibition of vasopressin v1b receptor translation by upstream open reading frames in the...
TRANSCRIPT

ORIGINAL ARTICLE
Inhibition of Vasopressin V1b Receptor Translation By Upstream OpenReading Frames in the 5¢-Untranslated RegionC. Rabadan-Diehl,* A. Martı́nez,� S. Volpi,* S. Subburaju* and G. Aguilera*
*Section on Endocrine Physiology, Developmental Endocrinology Branch, National Institute of Child Health and Human Development, National Institutes of
Health, Bethesda, MD, USA.
�Department of Neuroanatomy and Cell Biology, Instituto Cajal, Madrid, Spain.
Vasopressin (VP) secreted into the pituitary portal circulation from
hypothalamic parvicellular neurones is an important regulator of
adrenocorticotrophic hormone (ACTH) secretion (1, 2). The peptide
binds to VP V1b receptors (V1bR) coupled to protein kinase C-cal-
cium dependent mechanisms located in the pituitary corticotroph
(3, 4). Previous studies showing a good correlation between pituit-
ary ACTH responses and the number of VP receptors in the pituitary
suggest that regulation of V1bR may be a critical determinant of
corticotroph responsiveness during adaptation of the hypothalamic-
pituitary-adrenal (HPA) axis to stress (5). The mechanisms control-
ling the number of V1bR in the pituitary are complex, involving
transcriptional and post-transcriptional events (6). In a number of
situations, such as glucocorticoid administration, adrenalectomy
and osmotic stimulation, a dissociation exists between the levels of
V1bR mRNA and the number of receptors, with binding down-regu-
lation in spite of elevated mRNA levels (7, 8). This indicates that
post-transcriptional mechanisms play a major role in regulating the
number of V1bR, and suggests the presence of mechanisms capable
of actively inhibiting V1bR translation.
There is a body of evidence indicating that the 5¢-UTR of the
mRNA plays an important role in determining the translational effi-
ciency and protein synthesis (9). The conventional mode of initi-
ation of translation involves binding of the translation initiation
complex to a cap structure at the 5¢ end of the 5¢ untranslated
region (5¢-UTR), followed by scanning of the mRNA and recruitment
of ribosomal subunits. High complexity of the 5¢-UTR represses
Journal ofNeuroendocrinology
Correspondence to:
Dr Greti Aguilera, Section on
Endocrine Physiology, Developmental
Endocrinology Branch, NICHD, NIH
Bldg 10 Rm10N262, Bethesda, MD
20892, USA (e-mail:
The 5¢-UTR of the vasopressin V1b receptor (V1bR) mRNA contains small open reading frames
(ORF) located upstream (u) of the main ORF encoding the V1bR. The ability of the three proximal
uORFs to be translated into peptides and their influence on V1bR translation was examined
using fusion constructs of uORFs and V5 epitope, or ATG ⁄ ATA uORF mutations in the V1bR
cDNA. In vitro translation and western blot analysis after transfection of uORF1-V5 or uORF2-
V5 into cells revealed that uORF1 can be translated. As predicted by computer analysis, in vitro
translation using a rabbit reticulocyte ⁄ canine microsome system, immunohistochemistry and
western blot in membranes of transfected cells with uORF1-V5 revealed translocation of the
uORF1 peptide into membrane fractions. In vitro translation of V1bR cDNA with mutations of
the two uORFs proximal to the initiating methionine, uORFs 1 and 2 (Mut 1–2), or uORF2 (Mut
2) showed significantly increased translation of a 46 kDa band corresponding to the V1bR, com-
pared with wild-type (WT) V1bR, an effect that was attenuated by cotranslation of uORF1-V5.
Consistently, VP-induced inositol phosphate formation was higher in Chinese hamster ovay cells
transfected with Mut 1–2 than with WT V1bR. Immunohistochemical and western blot analysis,
using an antibody against uORF1, revealed peptide immunoreactivity in rat pituitary but not in
liver. Pituitary uORF immunoreactivity increased following glucocorticoid administration. The pre-
sent study shows that uORFs in the 5¢-UTR of the V1bR mRNA inhibit V1bR translation, and
suggests that translation of a 38-amino acid membrane peptide encoded by uORF1 exerts tonic
inhibition of V1bR translation.
Key words: vasopressin V1b receptor, translational control, 5¢-untranslated region, minicistron,
in vitro translation.
doi: 10.1111/j.1365-2826.2007.01533.x
Journal of Neuroendocrinology 19, 309–319
ª 2007 The Authors. Journal Compilation ª 2007 Blackwell Publishing Ltd
No claim to original US government works

cap-dependent translation, and many mRNAs with structured
5¢-UTRs are poorly translated using this mechanism (9–11). It has
been shown that the presence of short open reading frames (ORF)
or minicistrons in the 5¢-UTR of the mRNA can affect translation of
the main ORF in a number of transcripts (9). This feature is uncom-
mon in vertebrates but it has been described for low abundance
proteins, including some members of the G-protein coupled recep-
tor family, such as muscarinic, adrenergic, serotonergic, substance
P, substance K, angiotensin II and corticotrophin-releasing hormone
(CRH) receptors (9, 12, 13).
The rat V1bR 5¢-UTR consists of 826 bases, and has a complex
structure. Sequence analysis reveals five ORFs upstream of the initi-
ating methionine of the ORF coding for the V1bR protein. Studies
using constructs of the 5¢-UTR of the V1bR and luciferase reporter
gene have shown that mutations of these uORFs stimulate lucif-
erase activity (14), suggesting that they can inhibit translation. The
aims of the present study were to examine the influence of the
upstream ORFs in the 5¢-UTR of the V1bR mRNA, and to determine
whether the minicistrons can be translated into peptides, which, in
turn, could modulate the translational regulation of the V1bR.
Materials and methods
Animal procedures
Male Sprague-Dawley rats, weighing 200–250 g, were anaesthetised with
ketamine ⁄ xylazine, perfused transcardially with 100 ml of phosphate-buf-
fered saline (PBS) to remove blood followed by 300 ml of 4% paraformalde-
hyde in phosphate buffer. The pituitary was dissected and placed in a jar
with the same fixative for an additional period of 3 h before processing for
immunohistochemistry.
For western blot analysis, groups of rats subjected to sham operation or
bilateral adrenalectomy received subcutaneous pellets of 100% cholesterol
(controls) or 100% corticosterone (corticosterone). Pellets were prepared by
dipping 1 cm2 GF-B filter paper into melted cholesterol (as a control) or cor-
ticosterone and were implanted at the time of adrenalectomy or sham sur-
gery. Seven days later, rats were killed by decapitation, the pituitary
removed, dissected from the neural lobe, rapidly frozen on dry ice and
stored at )80 �C until processed for western blot. Plasma corticosterone
levels were undetectable in adrenalectomised rats, and reached levels of
20.3 ± 7 in sham-operated receiving cholesterol pellets, and
250.8 ± 17 ng ⁄ ml in sham-operated or adrenalectomised rats implanted
with corticosterone pellets.
Cell culture and transfections
MCF-7 breast cancer cells, which express V1bRs, and Chinese hamster ovary
(CHO) carcinoma cells were purchased from ATCC (Manassas, VA, USA) and
cultured at 37 �C under 5% CO2 atmosphere in medium supplemented with
10% foetal bovine serum, glutamine and penicillin ⁄ streptomycin. MCF-7 and
CHO cells were cultured in DMEM and alpha-MEM (Life Technologies Inc.,
Gaithersburg, MD, USA), respectively. Cells were cultured in 75-cm2 flasks
(for western blot) or slide wells (for immunohistochemistry) until reaching
about 80% confluence before transfection. A total amount of 10 lg DNA per
flask or 0.4 lg per well were transfected using Lipofectamine plus (Life Tech-
nologies, Inc.), according to the manufacturer’s instructions. After transfec-
tion, MCF-7 cells were cultured for 48 h before cell fractionation for western
blot, and CHO cells for 24 h before processing for immunohistochemistry.
DNA cloning of DNA fragments
A 2.7-Kb BamH1 fragment excised from the V1bR cDNA clone, rAP9-1 (3)
was subcloned into pAlter-Max (Promega, Madison, WI, USA) by blunt end
ligation into the SmaI site, after blunting ends of the fragment and vector
with Klenow (BioServe-Biotechnologies, Laurel MD, USA). This construct, pAl-
ter-V1bR, was used for in vitro translation studies and to generate muta-
tions ATG to ATA of the translation initiation codon of uORF 2 (Mut 2), or
mutations of the initiating codon of uORFs 2 and 1 (Mut 1–2). Mutant
clones were obtained using Altered Sites�II Mammalian Mutagenesis Sys-
tem (Promega). For Mut 2, single stranded DNA (ssDNA) from pALTER-V1bR
was isolated and annealed at 75 �C for 5 min with the complementary
phosphorylated mutagenic oligonucleotide, 5¢-GGA GCA GCT GCT ATC TCC
CAA ATG-3¢, and the mutant strand was synthesised using T4 DNA polym-
erase. DNA was transformed into competent ES1301 mutS cells to avoid
selection against the desired mutation. Plasmid DNA was purified and trans-
formed into JM109 cells to segregate WT from mutant plasmids. Mut 1–2
was generated using ssDNA from Mut 2 and the complementary oligonucle-
otide, 5¢-AAC AGG ACC AAT ATC CCT GAC CCA-3¢. The accuracy of the
mutants was verified by sequence analysis (Veritas, Rockville, MD, USA).
Fusion constructs for the uORFs 1 and 2 with the V5 epitope were gener-
ated by polymerase chain reaction (PCR) on pAlter-V1bR using primers FW-
1; 5¢-GGG TCA GGG ATG TTG GTC CT-3¢ and RW-1; 5¢-GAG GTG GAG AGA
AGG GAG GT-3¢ for uORF1, and FW-2; 5¢-ATT TGG GAG ATG GCA GCT GC-3¢and RW-2; 5¢-GTG TGG GAA GGT TGG AGT GG-3¢ for uORF2. PCR products
were TA cloned into pcDNA3.1 ⁄ V5-His TOPO vector (Invitrogen, Carlsbad, CA,
USA) and the accuracy of the constructs was verified by sequence analysis
(Veritas, Rockville, MD, USA).
In vitro translation
One lg of pAlter-V1bR or its mutants linearised with NotI, or uORF1-V5
and uORF2-V5 plasmids linearised with PmeI, were transcribed in vitro,
using a RiboprobeR in vitro transcription system (Promega) and a T7 RNA
polymerase. Equal amounts of in vitro-transcribed mRNA were in vitro
translated in a Wheat Germ cell free System (Promega) in the presence of35S methionine. Cold in vitro translation for western blot analysis was per-
formed in the absence of radiolabelled methionine. Aliquots of one-fifth of
the in vitro translated product were centrifuged through a 3% sucrose
cushion at 100 000 g for 1 h at 4 �C, the supernatant was acetone preci-
pitated, re-suspended in SDS sample buffer (Quality Biological, Inc., Gai-
thersburg, MD, USA), separated in a 15% SDS-PAGE, and exposed to a
BIOMAX film (Kodak, Rochester, NY, USA) at room temperature overnight.
Light transmittance of the autoradiographic bands for V5-fusion proteins
was quantified using a computerised imaging system (Imaging Research,
ST Catherine, Ontario, Canada), using the public domain NIH Image pro-
gramme (developed at the US National Institutes of Health and available:
http://rsb.info.nih.gov/nih-image). The product of the cold in vitro transla-
tion was processed for western blot analysis with an antibody against the
V5 epitope, as described below. To determine whether uORF1 is potentially
targeted to membrane, uncapped uORF1 RNA was in vitro translated using
a rabbit reticulocyte system in the presence of canine microsomes (TNT
Coupled Reticulocyte Lysate System, Promega). Aliquots of 0.5 lg of
in vitro transcribed uORF1-V5 mRNA were denatured at 65 �C for 3 min
and incubated for 1 h at 30 �C with 12.5 ll of rabbit reticulocyte lysate,
1.8 ll of canine microsomes and [35S]methionine in a final volume of
25 ll. Five microlitres were centrifuged at 10 000 g through a 3% sucrose
cushion for 1 h at 4 �C. The supernatant was discarded and the pellet
containing the canine microsomes with the translocated newly synthesised
membrane protein was re-suspended in 10 ll in SDS sample buffer and
run in a 15% SDS gel as described above.
310 C. Rabadan-Diehl et al.
ª 2007 The Authors. Journal Compilation ª 2007 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 19, 309–319
No claim to original US government works

Western blot
Western blot analysis of the upstream peptide-V5 epitope fusion protein
was performed in total cell extracts or in 30 000 g membrane-rich frac-
tions of MCF-7 cells tranfected with uORF1-V5 or uORF2-V5 DNA. Five
million cells were plated in 75-cm2 flasks and transfected with 10 lg of
DNA. For total cell protein, cells were lysed with a T-PER tissue protein
extraction reagent (Pierce, Rockford, IL, USA) supplemented with proteinase
inhibitors, 24 h after transfection. For membrane fractions, cells were
homogenised in 50 mM Tris-HCl pH 7.4, 2 mM EDTA, 5 mM MgCl2, 1 mM
DTT supplemented with proteinase inhibitors. After centrifugation at
1000 g to separate nuclei and unbroken cells, membrane fractions were
pelleted at 30 000 g at 4 �C for 30 min and re-suspended in 100 ll of
homogenisation buffer containing 1% Triton X-100. After shaking for
90 min at 4 �C and centrifugation for 30 min at 14 000 g, supernatants
were collected for protein determination and western blot. Twenty lg of
cell or membrane protein were loaded and separated in a 6% SDS-PAGE.
For western blot analysis of the cold in vitro translation product, 5 ll of
the reaction were loaded in a 15% SDS-PAGE gel. Proteins were trans-
ferred from the gel to a PDVF membrane (Amersham Pharmacia Biotech,
Piscataway, NJ, USA), incubated with 5% blocking agent in 1 · PBST (PBS
plus 0.1% Tween-20) for 1 h and incubated with anti-V5 antibody (Invitro-
gen) at a 1 : 5000 dilution overnight. After washing in 1 · PBST, mem-
branes were incubated for one hour with peroxidase-linked anti-mouse
IgG at a 1 : 50 000 dilution. Detection of immunoreactive uORF1-V5
fusion construct band was performed by using ECL Plus TM reagents
(Amersham Pharmacia Biotech) and exposure to Hyper film (Amersham
Pharmacia Biotech) for one minute. Light transmittance was quantified as
described for in vitro translation.
The presence of endogenous uORF1 peptide in rat tissues was exam-
ined by western blot using a polyclonal antibody generated against the
aminoterminus of the uORF peptide. The antibody was produced in rab-
bits by immunisation with the synthetic peptide, LTC FYL HPI VTP HLP
SLH L, conjugated with keyhole limpet haemocyanin (Covance, Vienna, VA,
USA). The antiserum was subjected to affinity purification using a CNBr
Sepharose column coupled to the peptide (TherImmune, Gaithersburgh,
MD, USA).
Pools of two to four pituitaries or equivalent amounts of liver frag-
ments were washed in ice-cold PBS in a 14-ml Falcon tube and homoge-
nised in 1 ml of PBS containing protease inhibitors (Pierce, Rockford, IL,
USA) using a mechanical homogeniser (Teckmar, Cincinnati, OH, USA) for
10 s and transferred to 1.5-ml microfuge tubes. After centrifugation at
1000 g, pellets were re-suspended in 30 ll of lysis buffer (Pierce) con-
taining protease inhibitors, vortexed for 15 s and centrifuged at 16 000 g
for 10 min. Thirty lg of solubilised proteins were separated by electro-
phoresis in a 17% SDS-polyacrylamide gel and transferred to PDVF mem-
brane (Amersham Pharmacia Biotech). Western blot was performed as
described above but using a 1 : 200 dilution of affinity purified anti-
uORF1 peptide antibody (0.74 mg ⁄ ml). For specificity tests, preabsorption
of the antibody (at 1 : 200 dilution) with the synthetic peptide (1 : 5
weight ⁄ weight) was performed at 4 �C overnight, in 10 ml of PBS con-
taining 0.1% Tween-20.
Immunohistochemistry
The subcellular location of the uORF-1 peptide was studied by immunohisto-
chemistry in cells transfected with uORF1-V5 constructs using either V5
antibodies (Invitrogen) or the uORF1 peptide polyclonal antibody. CHO cells
transfected with uORF-V5 constructs and cultured in glass slide wells were
fixed for 10 min in either neutral buffered formalin at room temperature or
in 50% acetone)50% methanol at )20 �C. Non-specific binding was
blocked by incubation in 3% normal goat serum in PBS for 30 min at room
temperature. The primary antibodies against the V5 epitope or uORF1 pep-
tide were applied overnight at 4 �C at dilutions of 1 : 1000 and 1 : 75,
respectively, in PBS. After several washes in PBS, Bodipy conjugated goat
anti-rabbit (for anti-uORF) or goat anti-mouse (for anti-V5) IgG (Molecular
Probes, Eugene, OR, USA) were applied at a dilution 1 : 200 for 1 h at room
temperature. Nuclei were counterstained with DAPI (Molecular Probes) for
10 min, and images obtained with a Zeiss Laser Scanning Microscope 510,
equipped with four lasers. To demonstrate the specificity of the staining, the
antibody was preincubated with the synthetic uORF peptide for 2 h at room
temperature before applying the mixture to the slide. To characterise the
subcellular distribution of the uORF immunoreactivity, the cells were treated
with different organelle markers before immunocytochemistry. These inclu-
ded Mito Tracker Red (for mitochondria), Lyso Tracker Red (for lysosomes)
and Red Bodipy-brefeldin A (for endoplasmic reticulum and Golgi) (Molecu-
lar Probes).
Rat pituitaries were embedded in paraffin following standard protocols
and 5-lm sections were obtained. After dewaxing, pituitary sections were
exposed to a mixture of the primary antibodies (rabbit anti-uORF 1 : 100;
mouse monoclonal anti-ACTH 1 : 1000) overnight at 4 �C. The next day,
sections were thoroughly washed in PBS and incubated for 1 h with a mix-
ture of the labelled secondary antibodies (Texas Red-goat anti-mouse IgG
and Bodipy-goat anti-rabbit IgG, both at 1 : 200; Molecular Probes) at room
temperature. After several washes, sections were mounted in Gel-Mount
(Biomeda, Foster City, CA, USA) and observed with a laser scanning micro-
scope.
Inositol phosphate formation
Transiently transfected CHO cells with pAlter-V1bR, mut 2 or mut 1–2
were cultured in 24-well plates and labelled with 2.5 lCi ⁄ ml of myo-[3H]
inositol per well for 24 h, washed with media containing 0.1% bovine
serum albumin and 10 mM LiCl, and then incubated for 15 min under the
conditions indicated in results and figure legends. Incubations were
stopped by addition of one volume of cold stop solution (1 M KOH, 18 mM
sodium tetraborate, 3.8 mM EDTA, 7.6 mM NaOH) followed by neutralisation
with 7.5% HCl. Total inositol phosphates were separated by anion
exchange chromatography as previously described (15) and measured in a
liquid scintillation counter.
Statistical analysis
Data are expressed as the mean ± SE of the number of experiments indica-
ted. The statistical significance of the differences between experimental
groups was calculated by one-way ANOVA followed by Fisher’s PLSD test.
P < 0.05 was considered statistically significant.
Results
The proximal upstream ORF is translated
The rat V1bR has a complex 5¢UTR, 826 bp length, containing five
short ORFs. Three major ones, located 500 bp from the initiating
methionine, encode potential peptides containing 38 (uORF1), 26
(uORF2) and ten amino acids (uORF3) (Fig. 1). The potential of the
putative peptides for translation was determined by in vitro transla-
tion of the two fusion constructs, uORF1-V5 and uORF2-V5, using35S methionine labelling of the product, or western blot analysis of
cold product. As shown in Fig. 2(B), no radiolabelled band was
observed after in vitro translation of uORF2-V5, while uORF1-V5
Vasopressin V1b receptor translation 311
ª 2007 The Authors. Journal Compilation ª 2007 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 19, 309–319
No claim to original US government works
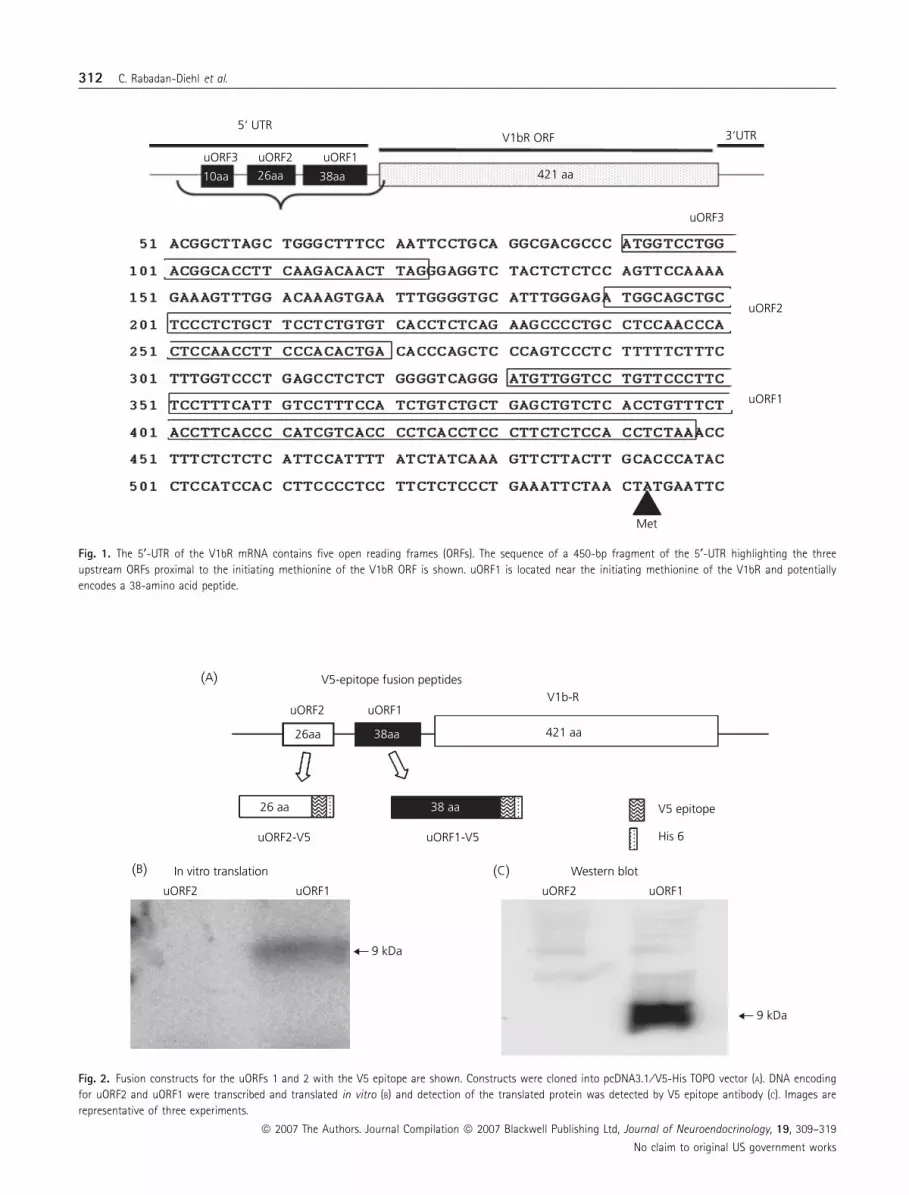
5‘ UTR
uORF1uORF2uORF3
38aa 421 aa
Met
uORF1
uORF2
uORF3
3‘UTR
26aa10aa
V1bR ORF
Fig. 1. The 5¢-UTR of the V1bR mRNA contains five open reading frames (ORFs). The sequence of a 450-bp fragment of the 5¢-UTR highlighting the three
upstream ORFs proximal to the initiating methionine of the V1bR ORF is shown. uORF1 is located near the initiating methionine of the V1bR and potentially
encodes a 38-amino acid peptide.
(B)
(A)
(C)
V5-epitope fusion peptides
26aa
Western blot
9 kDa
In vitro translation
His 6
V5 epitope
421 aa
V1b-R
uORF1-V5uORF2-V5
26 aa
uORF1
uORF1
uORF2
uORF2 uORF1uORF2
38aa
38 aa
9 kDa
Fig. 2. Fusion constructs for the uORFs 1 and 2 with the V5 epitope are shown. Constructs were cloned into pcDNA3.1 ⁄ V5-His TOPO vector (A). DNA encoding
for uORF2 and uORF1 were transcribed and translated in vitro (B) and detection of the translated protein was detected by V5 epitope antibody (C). Images are
representative of three experiments.
312 C. Rabadan-Diehl et al.
ª 2007 The Authors. Journal Compilation ª 2007 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 19, 309–319
No claim to original US government works
yielded a 9-kDa band corresponding to the molecular size of the
upstream peptide-V5 fusion protein. Consistent with the radiola-
belled band, western blot analysis of cold in vitro translated prod-
uct of uORF1-V5 with a V5 antibody, revealed a similar 9-kDa
band, confirming the identity of the band as the fusion protein
(Fig. 2C).
The upstream peptide-1 is a membrane protein
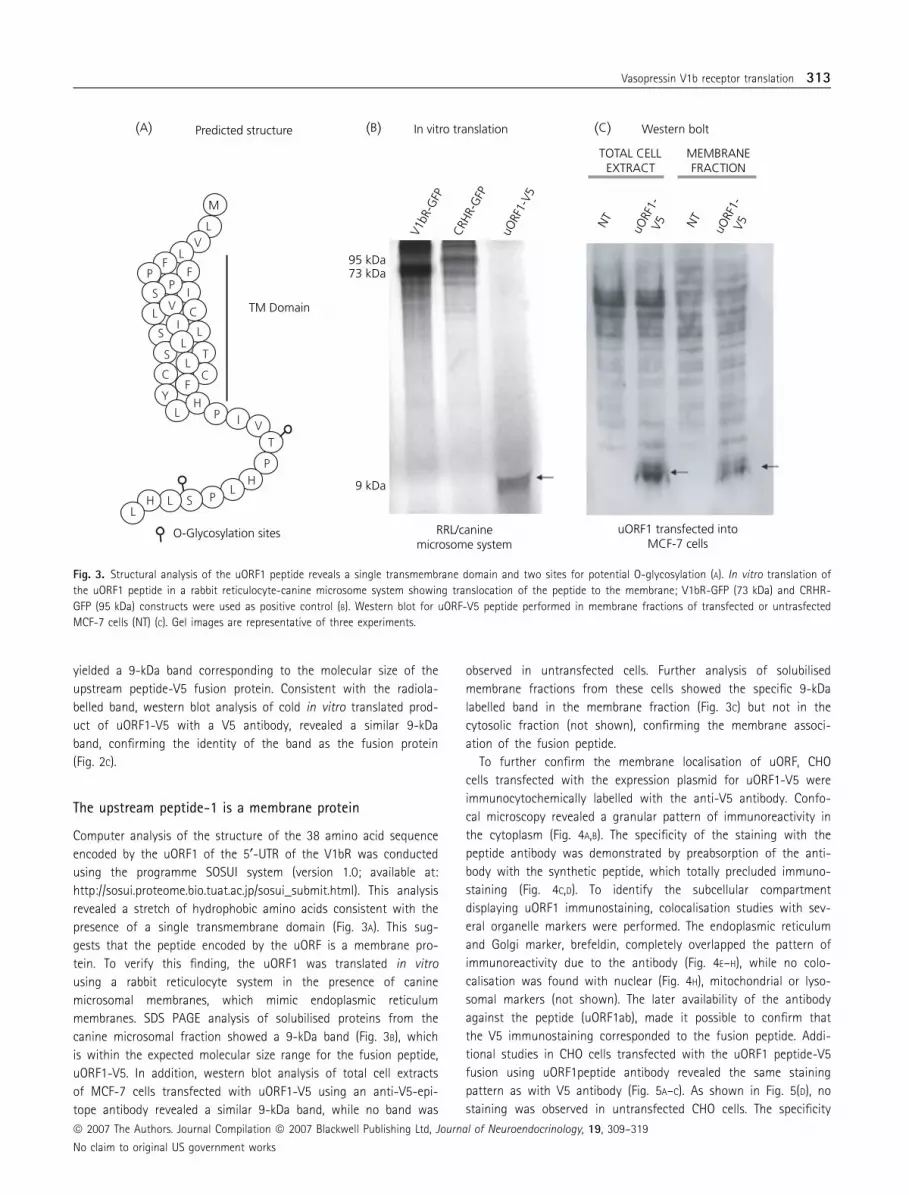
Computer analysis of the structure of the 38 amino acid sequence
encoded by the uORF1 of the 5¢-UTR of the V1bR was conducted
using the programme SOSUI system (version 1.0; available at:
http://sosui.proteome.bio.tuat.ac.jp/sosui_submit.html). This analysis
revealed a stretch of hydrophobic amino acids consistent with the
presence of a single transmembrane domain (Fig. 3A). This sug-
gests that the peptide encoded by the uORF is a membrane pro-
tein. To verify this finding, the uORF1 was translated in vitro
using a rabbit reticulocyte system in the presence of canine
microsomal membranes, which mimic endoplasmic reticulum
membranes. SDS PAGE analysis of solubilised proteins from the
canine microsomal fraction showed a 9-kDa band (Fig. 3B), which
is within the expected molecular size range for the fusion peptide,
uORF1-V5. In addition, western blot analysis of total cell extracts
of MCF-7 cells transfected with uORF1-V5 using an anti-V5-epi-
tope antibody revealed a similar 9-kDa band, while no band was
observed in untransfected cells. Further analysis of solubilised
membrane fractions from these cells showed the specific 9-kDa
labelled band in the membrane fraction (Fig. 3C) but not in the
cytosolic fraction (not shown), confirming the membrane associ-
ation of the fusion peptide.
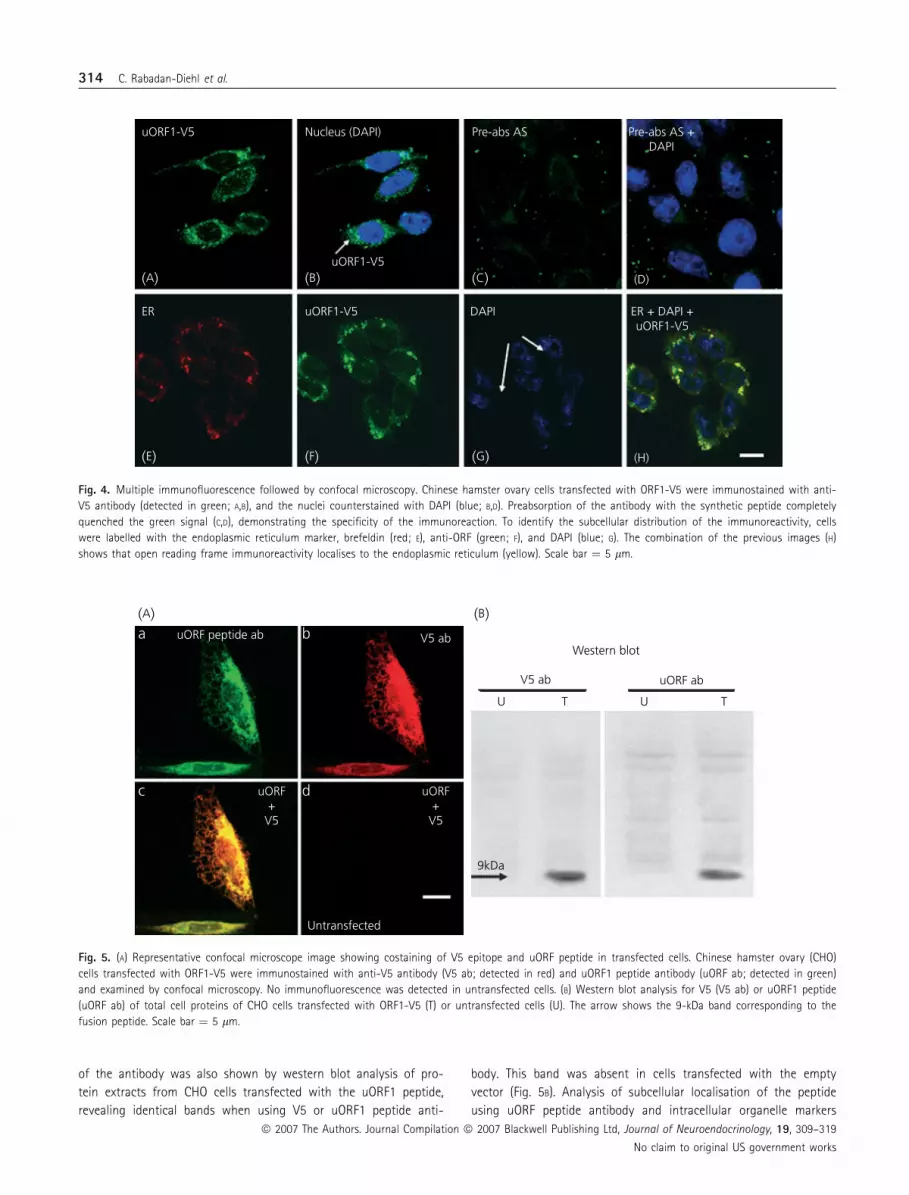
To further confirm the membrane localisation of uORF, CHO
cells transfected with the expression plasmid for uORF1-V5 were
immunocytochemically labelled with the anti-V5 antibody. Confo-
cal microscopy revealed a granular pattern of immunoreactivity in
the cytoplasm (Fig. 4A,B). The specificity of the staining with the
peptide antibody was demonstrated by preabsorption of the anti-
body with the synthetic peptide, which totally precluded immuno-
staining (Fig. 4C,D). To identify the subcellular compartment
displaying uORF1 immunostaining, colocalisation studies with sev-
eral organelle markers were performed. The endoplasmic reticulum
and Golgi marker, brefeldin, completely overlapped the pattern of
immunoreactivity due to the antibody (Fig. 4E–H), while no colo-
calisation was found with nuclear (Fig. 4H), mitochondrial or lyso-
somal markers (not shown). The later availability of the antibody
against the peptide (uORF1ab), made it possible to confirm that
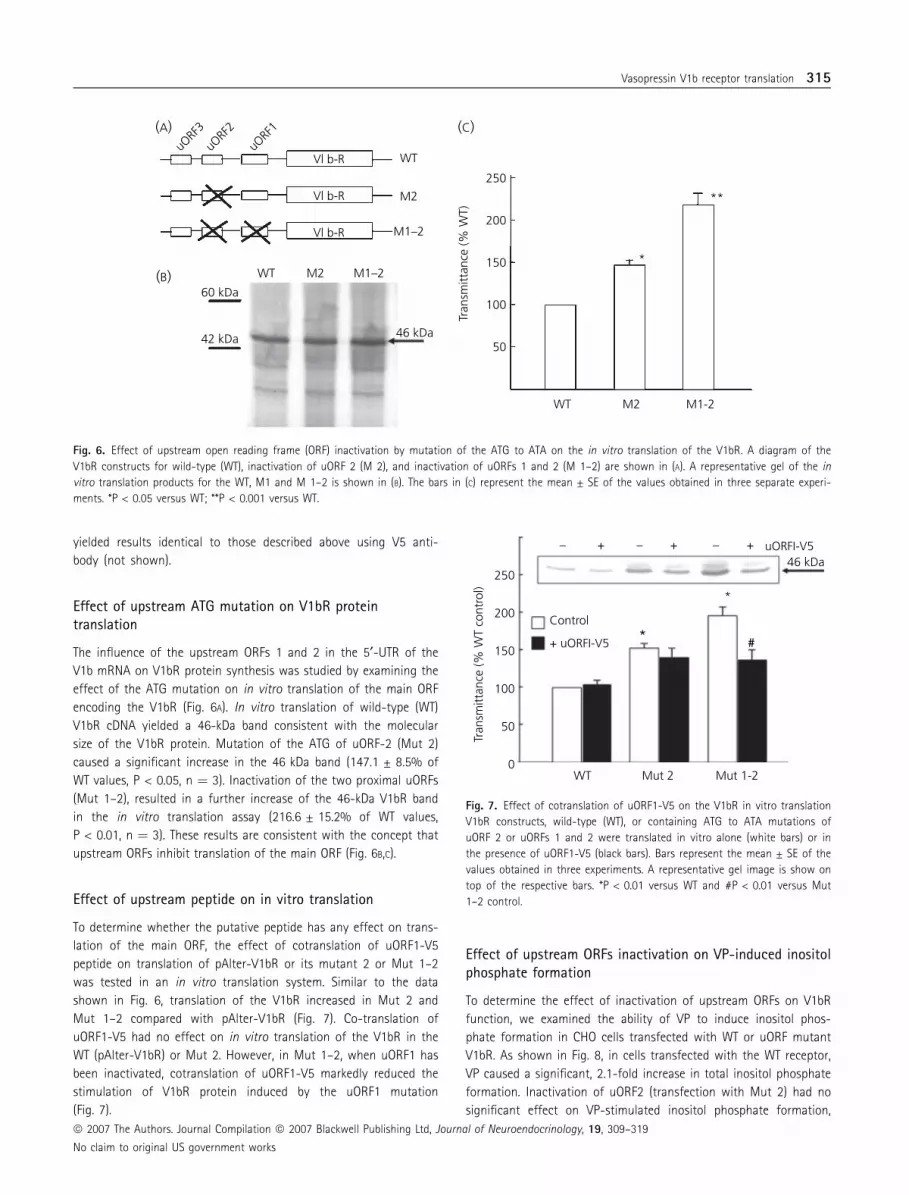
the V5 immunostaining corresponded to the fusion peptide. Addi-
tional studies in CHO cells transfected with the uORF1 peptide-V5
fusion using uORF1peptide antibody revealed the same staining
pattern as with V5 antibody (Fig. 5A–C). As shown in Fig. 5(D), no
staining was observed in untransfected CHO cells. The specificity
(A) (B) (C)
V1bR
-GFP
uORF
1-V5
uORF
1-V5 uO
RF1-
V5NT NT
CRHR
-GFP
9 kDa
RRL/caninemicrosome system
uORF1 transfected intoMCF-7 cells
73 kDa95 kDa
In vitro translation Western bolt
MEMBRANEFRACTION
TOTAL CELLEXTRACT
Predicted structure
O-Glycosylation sites
TM Domain
LV
L
L
L
I
I
V C
FC C
YL
HI V
T
H
LT
L
LL
LH S
S
S
S
FF
PP
P
P
P
M
Fig. 3. Structural analysis of the uORF1 peptide reveals a single transmembrane domain and two sites for potential O-glycosylation (A). In vitro translation of
the uORF1 peptide in a rabbit reticulocyte-canine microsome system showing translocation of the peptide to the membrane; V1bR-GFP (73 kDa) and CRHR-
GFP (95 kDa) constructs were used as positive control (B). Western blot for uORF-V5 peptide performed in membrane fractions of transfected or untrasfected
MCF-7 cells (NT) (C). Gel images are representative of three experiments.
Vasopressin V1b receptor translation 313
ª 2007 The Authors. Journal Compilation ª 2007 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 19, 309–319
No claim to original US government works
of the antibody was also shown by western blot analysis of pro-
tein extracts from CHO cells transfected with the uORF1 peptide,
revealing identical bands when using V5 or uORF1 peptide anti-
body. This band was absent in cells transfected with the empty
vector (Fig. 5B). Analysis of subcellular localisation of the peptide
using uORF peptide antibody and intracellular organelle markers
uORF1-V5
ER
(A) (B)
(F)
(C)
(G)
(D)
(H)(E)
ER + DAPI +uORF1-V5
uORF1-V5
uORF1-V5
DAPI
Nucleus (DAPI) Pre-abs AS Pre-abs AS +DAPI
Fig. 4. Multiple immunofluorescence followed by confocal microscopy. Chinese hamster ovary cells transfected with ORF1-V5 were immunostained with anti-
V5 antibody (detected in green; A,B), and the nuclei counterstained with DAPI (blue; B,D). Preabsorption of the antibody with the synthetic peptide completely
quenched the green signal (C,D), demonstrating the specificity of the immunoreaction. To identify the subcellular distribution of the immunoreactivity, cells
were labelled with the endoplasmic reticulum marker, brefeldin (red; E), anti-ORF (green; F), and DAPI (blue; G). The combination of the previous images (H)
shows that open reading frame immunoreactivity localises to the endoplasmic reticulum (yellow). Scale bar ¼ 5 lm.
uORF peptide ab
Untransfected
uORF+
V5
uORF+
V5
V5 ab
uORF ab
9kDa
V5 ab
U T U T
Western blot
(A)a b
c d
(B)
Fig. 5. (A) Representative confocal microscope image showing costaining of V5 epitope and uORF peptide in transfected cells. Chinese hamster ovary (CHO)
cells transfected with ORF1-V5 were immunostained with anti-V5 antibody (V5 ab; detected in red) and uORF1 peptide antibody (uORF ab; detected in green)
and examined by confocal microscopy. No immunofluorescence was detected in untransfected cells. (B) Western blot analysis for V5 (V5 ab) or uORF1 peptide
(uORF ab) of total cell proteins of CHO cells transfected with ORF1-V5 (T) or untransfected cells (U). The arrow shows the 9-kDa band corresponding to the
fusion peptide. Scale bar ¼ 5 lm.
314 C. Rabadan-Diehl et al.
ª 2007 The Authors. Journal Compilation ª 2007 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 19, 309–319
No claim to original US government works
yielded results identical to those described above using V5 anti-
body (not shown).
Effect of upstream ATG mutation on V1bR proteintranslation
The influence of the upstream ORFs 1 and 2 in the 5¢-UTR of the
V1b mRNA on V1bR protein synthesis was studied by examining the
effect of the ATG mutation on in vitro translation of the main ORF
encoding the V1bR (Fig. 6A). In vitro translation of wild-type (WT)
V1bR cDNA yielded a 46-kDa band consistent with the molecular
size of the V1bR protein. Mutation of the ATG of uORF-2 (Mut 2)
caused a significant increase in the 46 kDa band (147.1 ± 8.5% of
WT values, P < 0.05, n ¼ 3). Inactivation of the two proximal uORFs
(Mut 1–2), resulted in a further increase of the 46-kDa V1bR band
in the in vitro translation assay (216.6 ± 15.2% of WT values,
P < 0.01, n ¼ 3). These results are consistent with the concept that
upstream ORFs inhibit translation of the main ORF (Fig. 6B,C).
Effect of upstream peptide on in vitro translation
To determine whether the putative peptide has any effect on trans-
lation of the main ORF, the effect of cotranslation of uORF1-V5
peptide on translation of pAlter-V1bR or its mutant 2 or Mut 1–2
was tested in an in vitro translation system. Similar to the data
shown in Fig. 6, translation of the V1bR increased in Mut 2 and
Mut 1–2 compared with pAlter-V1bR (Fig. 7). Co-translation of
uORF1-V5 had no effect on in vitro translation of the V1bR in the
WT (pAlter-V1bR) or Mut 2. However, in Mut 1–2, when uORF1 has
been inactivated, cotranslation of uORF1-V5 markedly reduced the
stimulation of V1bR protein induced by the uORF1 mutation
(Fig. 7).
Effect of upstream ORFs inactivation on VP-induced inositolphosphate formation
To determine the effect of inactivation of upstream ORFs on V1bR
function, we examined the ability of VP to induce inositol phos-
phate formation in CHO cells transfected with WT or uORF mutant
V1bR. As shown in Fig. 8, in cells transfected with the WT receptor,
VP caused a significant, 2.1-fold increase in total inositol phosphate
formation. Inactivation of uORF2 (transfection with Mut 2) had no
significant effect on VP-stimulated inositol phosphate formation,
Vl b-RuO
RF3
uORF
2
uORF
1
Vl b-R
Vl b-R
WT
60 kDa
42 kDa 46 kDa
WT
250
(A) (C)
(B)150
50
200
100
M2
M2
M1–2
WT
Tran
smitt
ance
(% W
T)
M2
*
**
M1-2
M1–2
Fig. 6. Effect of upstream open reading frame (ORF) inactivation by mutation of the ATG to ATA on the in vitro translation of the V1bR. A diagram of the
V1bR constructs for wild-type (WT), inactivation of uORF 2 (M 2), and inactivation of uORFs 1 and 2 (M 1–2) are shown in (A). A representative gel of the in
vitro translation products for the WT, M1 and M 1–2 is shown in (B). The bars in (C) represent the mean ± SE of the values obtained in three separate experi-
ments. *P < 0.05 versus WT; **P < 0.001 versus WT.
Control
+ uORFI-V5
uORFI-V5–– – +++46 kDa
Mut 1-2
#
*
*
Mut 2WT
250
150
50
0
200
100
Tran
smitt
ance
(% W
T co
ntro
l)
Fig. 7. Effect of cotranslation of uORF1-V5 on the V1bR in vitro translation
V1bR constructs, wild-type (WT), or containing ATG to ATA mutations of
uORF 2 or uORFs 1 and 2 were translated in vitro alone (white bars) or in
the presence of uORF1-V5 (black bars). Bars represent the mean ± SE of the
values obtained in three experiments. A representative gel image is show on
top of the respective bars. *P < 0.01 versus WT and #P < 0.01 versus Mut
1–2 control.
Vasopressin V1b receptor translation 315
ª 2007 The Authors. Journal Compilation ª 2007 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 19, 309–319
No claim to original US government works
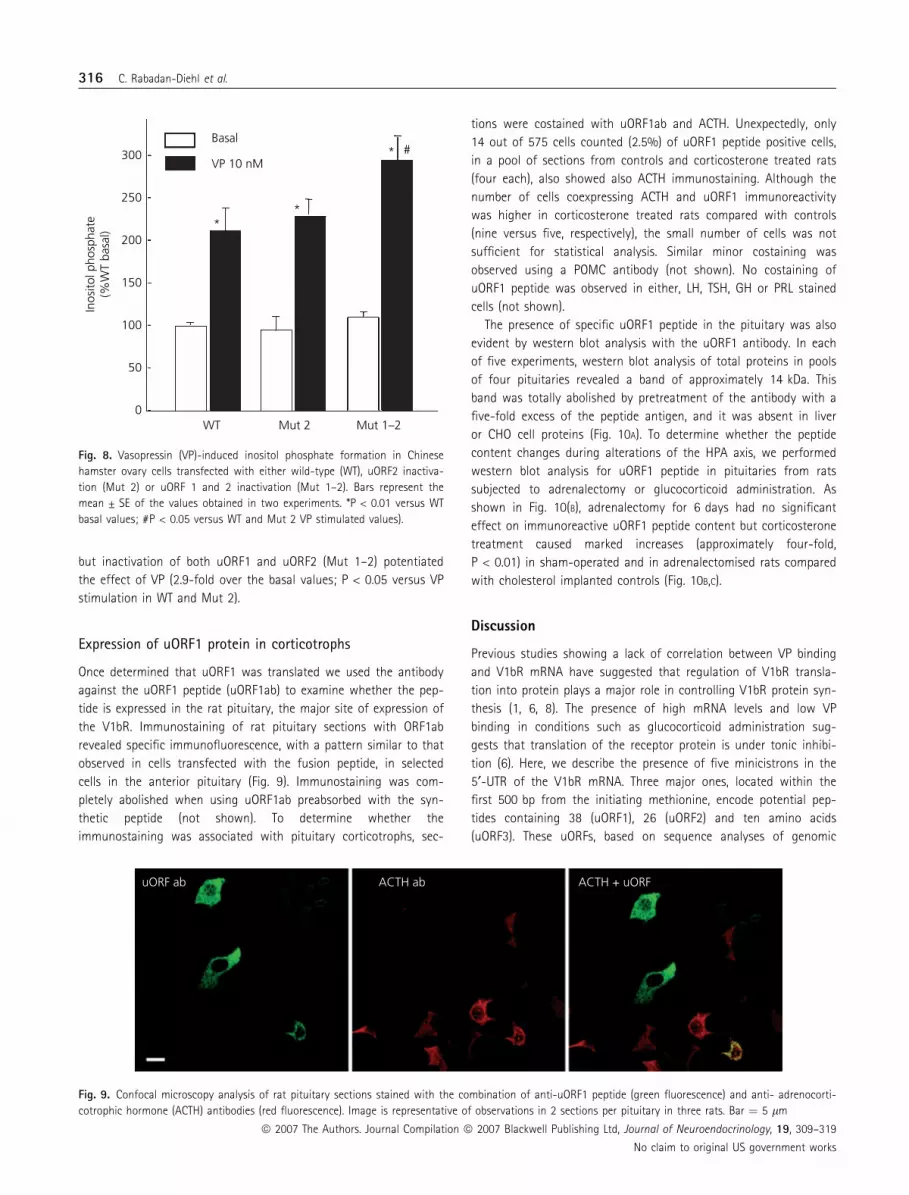
but inactivation of both uORF1 and uORF2 (Mut 1–2) potentiated
the effect of VP (2.9-fold over the basal values; P < 0.05 versus VP
stimulation in WT and Mut 2).
Expression of uORF1 protein in corticotrophs
Once determined that uORF1 was translated we used the antibody
against the uORF1 peptide (uORF1ab) to examine whether the pep-
tide is expressed in the rat pituitary, the major site of expression of
the V1bR. Immunostaining of rat pituitary sections with ORF1ab
revealed specific immunofluorescence, with a pattern similar to that
observed in cells transfected with the fusion peptide, in selected
cells in the anterior pituitary (Fig. 9). Immunostaining was com-
pletely abolished when using uORF1ab preabsorbed with the syn-
thetic peptide (not shown). To determine whether the
immunostaining was associated with pituitary corticotrophs, sec-
tions were costained with uORF1ab and ACTH. Unexpectedly, only
14 out of 575 cells counted (2.5%) of uORF1 peptide positive cells,
in a pool of sections from controls and corticosterone treated rats
(four each), also showed also ACTH immunostaining. Although the
number of cells coexpressing ACTH and uORF1 immunoreactivity
was higher in corticosterone treated rats compared with controls
(nine versus five, respectively), the small number of cells was not
sufficient for statistical analysis. Similar minor costaining was
observed using a POMC antibody (not shown). No costaining of
uORF1 peptide was observed in either, LH, TSH, GH or PRL stained
cells (not shown).
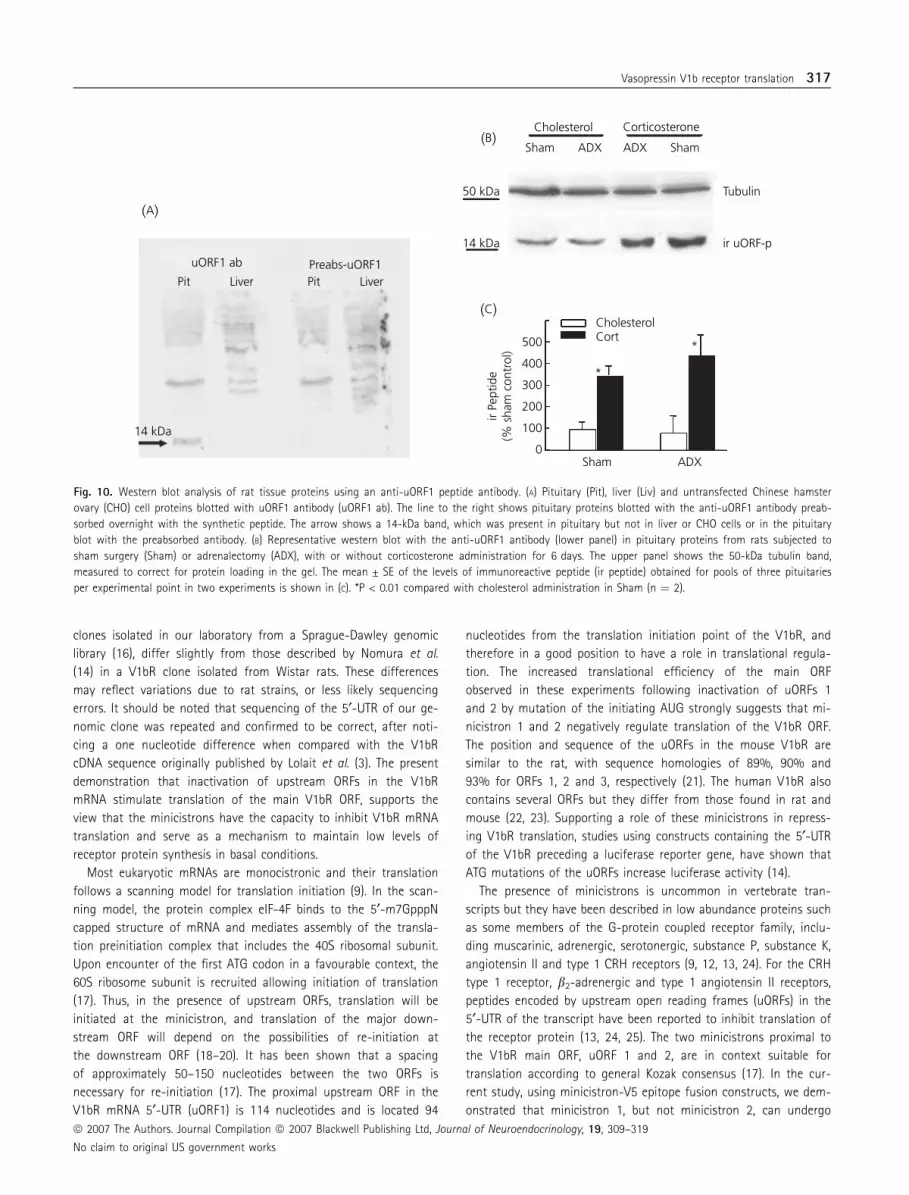
The presence of specific uORF1 peptide in the pituitary was also
evident by western blot analysis with the uORF1 antibody. In each
of five experiments, western blot analysis of total proteins in pools
of four pituitaries revealed a band of approximately 14 kDa. This
band was totally abolished by pretreatment of the antibody with a
five-fold excess of the peptide antigen, and it was absent in liver
or CHO cell proteins (Fig. 10A). To determine whether the peptide
content changes during alterations of the HPA axis, we performed
western blot analysis for uORF1 peptide in pituitaries from rats
subjected to adrenalectomy or glucocorticoid administration. As
shown in Fig. 10(B), adrenalectomy for 6 days had no significant
effect on immunoreactive uORF1 peptide content but corticosterone
treatment caused marked increases (approximately four-fold,
P < 0.01) in sham-operated and in adrenalectomised rats compared
with cholesterol implanted controls (Fig. 10B,C).
Discussion
Previous studies showing a lack of correlation between VP binding
and V1bR mRNA have suggested that regulation of V1bR transla-
tion into protein plays a major role in controlling V1bR protein syn-
thesis (1, 6, 8). The presence of high mRNA levels and low VP
binding in conditions such as glucocorticoid administration sug-
gests that translation of the receptor protein is under tonic inhibi-
tion (6). Here, we describe the presence of five minicistrons in the
5¢-UTR of the V1bR mRNA. Three major ones, located within the
first 500 bp from the initiating methionine, encode potential pep-
tides containing 38 (uORF1), 26 (uORF2) and ten amino acids
(uORF3). These uORFs, based on sequence analyses of genomic
Basal
VP 10 nM
**
* #
Mut 1–2Mut 2WT
250
300
150
50
0
200
100
Inos
itol p
hosp
hate
(%W
T ba
sal)
Fig. 8. Vasopressin (VP)-induced inositol phosphate formation in Chinese
hamster ovary cells transfected with either wild-type (WT), uORF2 inactiva-
tion (Mut 2) or uORF 1 and 2 inactivation (Mut 1–2). Bars represent the
mean ± SE of the values obtained in two experiments. *P < 0.01 versus WT
basal values; #P < 0.05 versus WT and Mut 2 VP stimulated values).
uORF ab ACTH ab ACTH + uORF
Fig. 9. Confocal microscopy analysis of rat pituitary sections stained with the combination of anti-uORF1 peptide (green fluorescence) and anti- adrenocorti-
cotrophic hormone (ACTH) antibodies (red fluorescence). Image is representative of observations in 2 sections per pituitary in three rats. Bar ¼ 5 lm
316 C. Rabadan-Diehl et al.
ª 2007 The Authors. Journal Compilation ª 2007 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 19, 309–319
No claim to original US government works
clones isolated in our laboratory from a Sprague-Dawley genomic
library (16), differ slightly from those described by Nomura et al.
(14) in a V1bR clone isolated from Wistar rats. These differences
may reflect variations due to rat strains, or less likely sequencing
errors. It should be noted that sequencing of the 5¢-UTR of our ge-
nomic clone was repeated and confirmed to be correct, after noti-
cing a one nucleotide difference when compared with the V1bR
cDNA sequence originally published by Lolait et al. (3). The present
demonstration that inactivation of upstream ORFs in the V1bR
mRNA stimulate translation of the main V1bR ORF, supports the
view that the minicistrons have the capacity to inhibit V1bR mRNA
translation and serve as a mechanism to maintain low levels of
receptor protein synthesis in basal conditions.
Most eukaryotic mRNAs are monocistronic and their translation
follows a scanning model for translation initiation (9). In the scan-
ning model, the protein complex eIF-4F binds to the 5¢-m7GpppN
capped structure of mRNA and mediates assembly of the transla-
tion preinitiation complex that includes the 40S ribosomal subunit.
Upon encounter of the first ATG codon in a favourable context, the
60S ribosome subunit is recruited allowing initiation of translation
(17). Thus, in the presence of upstream ORFs, translation will be
initiated at the minicistron, and translation of the major down-
stream ORF will depend on the possibilities of re-initiation at
the downstream ORF (18–20). It has been shown that a spacing
of approximately 50–150 nucleotides between the two ORFs is
necessary for re-initiation (17). The proximal upstream ORF in the
V1bR mRNA 5¢-UTR (uORF1) is 114 nucleotides and is located 94
nucleotides from the translation initiation point of the V1bR, and
therefore in a good position to have a role in translational regula-
tion. The increased translational efficiency of the main ORF
observed in these experiments following inactivation of uORFs 1
and 2 by mutation of the initiating AUG strongly suggests that mi-
nicistron 1 and 2 negatively regulate translation of the V1bR ORF.
The position and sequence of the uORFs in the mouse V1bR are
similar to the rat, with sequence homologies of 89%, 90% and
93% for ORFs 1, 2 and 3, respectively (21). The human V1bR also
contains several ORFs but they differ from those found in rat and
mouse (22, 23). Supporting a role of these minicistrons in repress-
ing V1bR translation, studies using constructs containing the 5¢-UTR
of the V1bR preceding a luciferase reporter gene, have shown that
ATG mutations of the uORFs increase luciferase activity (14).
The presence of minicistrons is uncommon in vertebrate tran-
scripts but they have been described in low abundance proteins such
as some members of the G-protein coupled receptor family, inclu-
ding muscarinic, adrenergic, serotonergic, substance P, substance K,
angiotensin II and type 1 CRH receptors (9, 12, 13, 24). For the CRH
type 1 receptor, b2-adrenergic and type 1 angiotensin II receptors,
peptides encoded by upstream open reading frames (uORFs) in the
5¢-UTR of the transcript have been reported to inhibit translation of
the receptor protein (13, 24, 25). The two minicistrons proximal to
the V1bR main ORF, uORF 1 and 2, are in context suitable for
translation according to general Kozak consensus (17). In the cur-
rent study, using minicistron-V5 epitope fusion constructs, we dem-
onstrated that minicistron 1, but not minicistron 2, can undergo
Cholesterol
500
400
300
ir Pe
ptid
e(%
sha
m c
ontr
ol)
200
100
0
14 kDa
uORF1 ab
Pit Liver Pit LiverPreabs-uORF1
Cort*
*
Sham ADX
50 kDa
14 kDa
Tubulin
ir uORF-p
(B)
(A)
(C)
Sham ADX ShamADX
CorticosteroneCholesterol
Fig. 10. Western blot analysis of rat tissue proteins using an anti-uORF1 peptide antibody. (A) Pituitary (Pit), liver (Liv) and untransfected Chinese hamster
ovary (CHO) cell proteins blotted with uORF1 antibody (uORF1 ab). The line to the right shows pituitary proteins blotted with the anti-uORF1 antibody preab-
sorbed overnight with the synthetic peptide. The arrow shows a 14-kDa band, which was present in pituitary but not in liver or CHO cells or in the pituitary
blot with the preabsorbed antibody. (B) Representative western blot with the anti-uORF1 antibody (lower panel) in pituitary proteins from rats subjected to
sham surgery (Sham) or adrenalectomy (ADX), with or without corticosterone administration for 6 days. The upper panel shows the 50-kDa tubulin band,
measured to correct for protein loading in the gel. The mean ± SE of the levels of immunoreactive peptide (ir peptide) obtained for pools of three pituitaries
per experimental point in two experiments is shown in (C). *P < 0.01 compared with cholesterol administration in Sham (n ¼ 2).
Vasopressin V1b receptor translation 317
ª 2007 The Authors. Journal Compilation ª 2007 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 19, 309–319
No claim to original US government works

translation. The inability of uORF2 to be translated in spite of the
good theoretical translation context could result from the complex
secondary structure predicted for the V1bR 5¢-UTR. Translation of
the proximal minicistron-V5 fusion peptide was evident by in vitro
translation, as well as by western blot in cells transfected with an
expression vector containing the fusion peptide. The ability of the
upstream peptide to inhibit V1bR translation when cotranslated with
the V1bR cDNA, suggests that the peptide is part of the mechanism
by which upstream ORF1 inhibits V1bR translation. It should be
noted that uORF1 inhibited in vitro translation of the V1bR ORF only
when the native uORF1 was inactivated by mutation of the ATG. This
suggests that uORF1 produced by the WT construct exerts full inhi-
bitory effect and that additional production by the transfected con-
struct has no additional effect. This supports the view that
translation of the upstream ORF1 mediates tonic inhibition of trans-
lation of the main ORF encoding the V1bR protein.
An unexpected finding was the membrane localisation of the
upstream peptide. This was first suspected from the predicted
structure of the peptide, with a stretch of hydrophobic amino acids
consistent with the presence of a single transmembrane domain.
The present experiments using the V5 epitope tagged peptide, dem-
onstrate that the peptide is translated and subsequently translocat-
ed into the membrane, in vitro as well as in transfected cells.
Although it is possible that the peptide remains partially in the
lumen of the endoplasmic reticulum, this is highly unlikely because
no radiolabelled peptide was found in the supernatant fraction
after membrane purification, and some leakage would be expected
if the peptide is not membrane incorporated. Moreover, in addition
to the computer prediction of a membrane protein, the immunoh-
istochemical evidence is consistent with endoplasmic reticulum
localisation. An important finding, made possible by the availability
of the uORF1 peptide antibody, was the identification of immunore-
active native peptide in the rat pituitary but not in the liver, an
organ devoid of V1bR. The molecular size of the immunoreactive
peptide shown by western blot was about twice the expected size
for the peptide. Since the peptide sequence reveals two glycosyla-
tion sites, it is possible that glycosylation accounts for the larger
molecular size. Alternatively, the higher molecular size could corres-
pond to detergent resistant dimers. However, this is less likely
because dimerisation could not account for the larger than expec-
ted molecular size of the fusion uORF1-V5 peptide observed after
in vitro translation in canine microsomes or transfection in CHO
cells (9 kDa versus the expected size of approximately 7 kDa).
Although further studies involving degycosylation, affinity purifica-
tion and sequencing of the immunoreactive band are needed to
confirm that the immunoreactive material in the pituitary corres-
ponds to the uORF1 peptide, the present findings strongly suggests
that the immunoreactivity detected in the pituitary contains the
uORF1 peptide sequence. In addition, computer analysis of the nuc-
leotide sequence encoding the amino terminus of the peptide used
for the antibody production identified only the rat V1bR region and
no homology with any other known sequence.
The minor colocalisation of uORF1 immunoreactivity in ACTH
containing cells in the anterior pituitary was quite unexpected
because the major site of expression of the V1b receptor is the
anterior pituitary corticotroph. In addition, the total lack of colocali-
sation with other pituitary hormones, suggest that the cells
expressing the peptide correspond to undifferentiated cells or
non-endocrine cells in the pituitary. An important question to be
resolved is whether these cells expressing uORF1 peptide immuno-
reactivity also express the V1b receptor. Using a V1b receptor anti-
body, Hernando et al. (26) reported an approximate 90%
colocalisation of V1b receptor immunoreactivity and ACTH cells.
However, in double staining in situ hybridisation studies, it appears
that V1b receptor autoradiographic grains also concentrate overlay-
ing areas void of POMC staining (3), raising the possibility that V1b
other cell types in the pituitary express the V1b receptor. Also sup-
porting an effect of VP in undifferentiated cells are recent studies
showing that administration of a VP V1 receptor antagonist
prevents the increase in the number of cells incorporating bromo
deoxy uridine (BrdU) in the anterior pituitary, but the BrdU stained
nuclei do not correspond to ACTH- or other pituitary hormone-
stained cells (27).
The present study also provides evidence that the expression of
the immunoreactive-like peptide in the pituitary is regulated by
glucocorticoids. It is noteworthy that long-term glucocorticoid
administration is associated with an increase in immunoreactive
peptide in the pituitary. We have previously shown that glucocorti-
coid administration for 6 days leads to VP binding down regulation,
but increased VPmRNA levels in the pituitary suggesting inhibition
of translation. The present demonstration that coexpression of the
peptide decreases V1bR translation and that the levels of immunore-
active peptide increases in a condition associated with translation
inhibition, supports the hypothesis that uORF peptide contributes to
V1bR down-regulation by inhibiting translation. A potential mechan-
ism by which the peptide could inhibit V1bR translation involves the
binding of proteins to the 5¢-UTR (28). For a number of genes with
complex 5¢-UTR such as CRHR1, angiotensin type-1 and IGF I recep-
tors, it has been shown that RNA binding proteins to the 5¢-UTR
regulate translation (29–31). In the case of the CRHR1, alterations
of the HPA axis regulate the binding of protein complexes to the
5¢-UTR of the receptor mRNA (31). Binding of proteins or other fac-
tors could directly inhibit translation of the main ORF, they could
also modulate positive regulatory mechanisms. In this regard, it has
been demonstrated that the 5¢-UTR of the V1bR mRNA can posi-
tively regulate translation through an internal ribosome entry site
(IRES) (32). Activation of protein kinase C by the phorbol ester, PMA,
increased IRES-mediated translation, suggesting that this mechan-
ism contributes to V1bR mRNA up-regulation induced by PKC-cou-
pled regulators, such as VP itself (32). The mechanisms regulating
the balance between translational inhibition and stimulation by the
5¢-UTR of the V1bR mRNA will be the subject of future studies.
It is evident from the in vitro translation and western blot stud-
ies that the uORF is involved in translational inhibition. The mech-
anism by which a membrane targeted peptide may influence the
rate of translation remains to be elucidated, but since the endo-
plasmic reticulum is the site for receptor translation, the peptide
could exert direct or indirect effects on ribosome function. Other
single transmembrane proteins encoded by upstream ORFs, such as
receptor-activity-modifying proteins, are known to determine the
318 C. Rabadan-Diehl et al.
ª 2007 The Authors. Journal Compilation ª 2007 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 19, 309–319
No claim to original US government works

specificity of the calcitonin-receptor-like receptor for either calcito-
nin-gene-related peptide or adrenomedullin depending on the type
of protein associated to the receptor (33). Inactivation of the V1bR
uORF1 potentiated the effect of VP on inositol phosphate forma-
tion. However, it is more likely that the potentiation is due to an
increase in receptor synthesis because the mutation also increases
receptor protein content in the cell.
In conclusion, this study demonstrates that upstream ORFs in
the 5¢-UTR of the V1bR can act as translational repressors for the
V1bR ORF. The in vitro data in conjunction with the presence of
regulated levels of immunoreactive uORF1 peptide in the rat pituit-
ary provides strong evidence that ORF1 is translated into a peptide,
which is likely to exert inhibitory actions on V1bR translation. Cur-
rent knowledge indicates that the 5¢-UTR of the V1bR has dual
effects on mRNA translation. Although it has been shown that an
IRES can mediate positive regulation, upstream ORFs are likely to
be part of the mechanism for tonic inhibition of V1bR translation
under resting conditions.
Accepted 10 January 2007
References
1 Aguilera G. Regulation of pituitary ACTH secretion during chronic stress.
Front Neuroendocrinol 1994; 15: 321–350.
2 Antoni FA. Vasopressinergic control of pituitary adrenocorticotropin
secretion comes of age. Front Neuroendocrinol 1993; 14: 76–122.
3 Lolait SJ, O’Carroll AM, Mahan LC, Felder CC, Button DC, Young WS III,
Mezey E, Brownstein MJ. Extrapituitary expression of the rat V1b vaso-
pressin receptor gene. Proc Natl Acad Sci USA 1995; 92: 6783–6787.
4 Jard S, Barberis C, Audigier S, Tribollet E. Neurohypophyseal hormone
receptor systems in brain and periphery. Prog Brain Res 1987; 72: 173–187.
5 Aguilera G, Pham Q, Rabadan-Diehl C. Regulation of pituitary vasopres-
sin receptors during chronic stress: relationship to corticotroph respon-
siveness. J Neuroendocrinol 1994; 6: 299–304.
6 Aguilera G, Volpi S, Rabadan-Diehl C. Transcriptional and post-transcrip-
tional mechanisms regulating the rat pituitary vasopressin V1b receptor
gene. J Mol Endocrinol 2003; 30: 99–108.
7 Rabadan-Diehl C, Makara G, Kiss A, Lolait S, Zelena D, Ochedalski T,
Aguilera G. Regulation of pituitary V1b vasopressin receptor messenger
ribonucleic acid by adrenalectomy and glucocorticoid administration.
Endocrinology 1997; 138: 5189–5194.
8 Rabadan-Diehl C, Lolait SJ, Aguilera G. Regulation of pituitary vasopres-
sin V1b receptor mRNA during stress in the rat. J Neuroendocrinol 1995;
7: 903–910.
9 Kozak M. Initiation of translation in prokaryotes and eukaryotes. Gene
1999; 234: 187–208.
10 Shatkin AJ. Capping of eucaryotic mRNAs. Cell 1976; 9: 645–653.
11 Sachs AB, Sarnow P, Hentze MW. Starting at the beginning, middle, and
end: translation initiation in eukaryotes. Cell 1997; 89: 831–838.
12 Kobilka BK, MacGregor C, Daniel K, Kobilka TS, Caron MG, Lefkowitz RJ.
Functional activity and regulation of human beta 2-adrenergic receptors
expressed in Xenopus oocytes. J Biol Chem 1987; 262: 15796–15802.
13 Xu G, Rabadan-Diehl C, Nikodemova M, Wynn P, Spiess J, Aguilera G.
Inhibition of corticotropin releasing hormone type-1 receptor translation
by an upstream AUG triplet in the 5¢ untranslated region. Mol Pharma-
col 2001; 59: 485–492.
14 Nomura A, Iwasaki Y, Saito M, Aoki Y, Yamamori E, Ozaki N, Tachikawa
K, Mutsuga N, Morishita M, Yoshida M, Asai M, Oiso Y, Saito H. Involve-
ment of upstream open reading frames in regulation of rat V(1b) vaso-
pressin receptor expression. Am J Physiol Endocrinol Metab 2001; 280:
E780–E787.
15 Rabadan-Diehl C, Aguilera G. Glucocorticoids increase vasopressin V1b
receptor coupling to phospholipase C. Endocrinology 1998; 139: 3220–
3226.
16 Rabadan-Diehl C, Lolait SJ, Aguilera G. Isolation and characterization of
the promoter region of the rat vasopressin V1b receptor gene. . J Neuro-
endocrinol 2000; 12: 437–444.
17 Kozak M. Pushing the limits of the scanning mechanism for initiation of
translation. Gene 2002; 299: 1–34.
18 Geballe AP, Morris DR. Initiation codons within 5¢-leaders of mRNAs as
regulators of translation. Trends Biochem Sci 1994; 19: 159–164.
19 Hunt T. False starts in translational control of gene expression. Nature
1985; 316: 580–581.
20 Hinnebusch AG. Translational regulation of yeast GCN4. A window on
factors that control initiator-trna binding to the ribosome. J Biol Chem
1997; 272: 21661–21664.
21 Ventura MA, Rene P, de Keyzer Y, Bertagna X, Clauser E. Gene and cDNA
cloning and characterization of the mouse V3 ⁄ V1b pituitary vasopressin
receptor. J Mol Endocrinol 1999; 22: 251–260.
22 de Keyzer Y, Auzan C, Lenne F, Beldjord C, Thibonnier M, Bertagna X,
Clauser E. Cloning and characterization of the human V3 pituitary vaso-
pressin receptor. FEBS Lett 1994; 356: 215–220.
23 Sugimoto T, Saito M, Mochizuki S, Watanabe Y, Hashimoto S, Kawashima
H. Molecular cloning and functional expression of a cDNA encoding the
human V1b vasopressin receptor. J Biol Chem 1994; 269: 27088–27092.
24 Mori Y, Matsubara H, Murasawa S, Kijima K, Maruyama K, Tsukaguchi H,
Okubo N, Hamakubo T, Inagami T, Iwasaka T, Inada M. Translational
regulation of angiotensin II type 1A receptor. Role of upstream AUG tri-
plets. Hypertension 1996; 28: 810–817.
25 Parola AL, Kobilka BK. The peptide product of a 5¢ leader cistron in the
beta 2 adrenergic receptor mRNA inhibits receptor synthesis. J Biol
Chem 1994; 269: 4497–4505.
26 Hernando F, Schoots O, Lolait SJ, Burbach JP. Immunohistochemical
localization of the vasopressin V1b receptor in the rat brain and pituit-
ary gland: anatomical support for its involvement in the central effects
of vasopressin. Endocrinology 2001; 142: 1659–1668.
27 Subburaju S, Aguilera G. Vasopressin mediates pituitary mitogenesis but
not increases in the number of ACTH containing corticotrophs. 6th Inter-
national Congress of Neuroendocrinology, Pittsburgh, PA. Front Neuro-
endocrinol 2006; 27: 57.
28 McCarthy JE, Kollmus H. Cytoplasmic mRNA–protein interactions in euk-
aryotic gene expression. Trends Biochem Sci 1995; 20: 191–197.
29 Krishnamurthi K, Verbalis JG, Zheng W, Wu Z, Clerch LB, Sandberg K.
Estrogen regulates angiotensin AT1 receptor expression via cytosolic pro-
teins that bind to the 5¢ leader sequence of the receptor mRNA. Endo-
crinology 1999; 140: 5435–5438.
30 Giraud S, Greco A, Brink M, Diaz JJ, Delafontaine P. Translation initiation
of the insulin-like growth factor I receptor mRNA is mediated by an
internal ribosome entry site. J Biol Chem 2001; 276: 5668–5675.
31 Wu Z, Ji H, Hassan A, Aguilera G, Sandberg K. Regulation of pituitary
corticotropin releasing factor type-1 receptor mRNA binding proteins by
modulation of the hypothalamic-pituitary-adrenal axis. J Neuroendocri-
nol 2004; 16: 214–220.
32 Rabadan-Diehl C, Volpi S, Nikodemova M, Aguilera G. Translational regu-
lation of the vasopressin v1b receptor involves an internal ribosome
entry site. J Mol Endocrinol 2003; 17: 1959–1971.
33 McLatchie LM, Fraser NJ, Main MJ, Wise A, Brown J, Thompson N,
Solari R, Lee MG, Foord SM. RAMPs regulate the transport and ligand
specificity of the calcitonin-receptor-like receptor. Nature 1998; 393:
333–339.
Vasopressin V1b receptor translation 319
ª 2007 The Authors. Journal Compilation ª 2007 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 19, 309–319
No claim to original US government works