induction of triploidy and gynogenesis in teleost fish with emphasis on marine species
TRANSCRIPT

Genetica 111: 175–195, 2001.© 2001 Kluwer Academic Publishers. Printed in the Netherlands.
175
Induction of triploidy and gynogenesis in teleost fish with emphasis onmarine species
A. Felip1, S. Zanuy1, M. Carrillo1 & F. Piferrer2,∗1Instituto de Acuicultura de Torre la Sal, Consejo Superior de Investigaciones Cientıficas (CSIC), 12595 Riberade Cabanes, Castellon, Spain (Phone: +34-964-319500; Fax: +34-964-319509; E-mails: [email protected],[email protected], [email protected]); 2Institut de Ciències del Mar, CSIC, Passeig Maritim, 37-49, 08003Barcelona, Spain (Phone: +34-93-2309500; Fax: +34-93-2309555; E-mail: [email protected]); ∗Author forcorrespondence
Key words: chromosome set manipulation, Dicentrarchus labrax, gonadal development, growth, gynogenesis,marine teleosts, sex determination, sex ratio, sterility, triploidy
Abstract
The induction of triploidy and gynogenesis by chromosome set manipulation has traditionally been studied moreintensively in freshwater than in marine fish. In the last years, however, several studies have applied these manipula-tions in about a dozen marine species, including mainly sparids, moronids and flatfishes. This paper focuses on themethodologies used to induce, verify, and assess performance of both triploids and gynogenetics of these marinespecies. Since many of them are batch spawners and have small and fragile eggs and larvae, peculiarities relatingto broodstock management, gamete quality and mortality assessment during early larval stages are also taken intoaccount. However, data show that if handling is correct and the treatments are optimized, triploid and gynogeneticrates of 100% can be easily achieved. Survival of triploids with respect to the controls is about 70–80%, whereasin gynogenetics it is generally low and more variable, depending on the species considered. In the marine fishinvestigated so far, triploidy has not resulted in significantly higher growth rates. On the other hand, the inductionof gynogenesis has resulted in the production of both all-female and mix-sex stocks. Throughout the paper, specialreference is made to the European sea bass (Dicentrarchus labrax L.), a species of both basic and applied interest,for which a comprehensive study has been carried out on the induction, verification and performance of triploidsand gynogenetics.
Introduction
Sexual maturation in fish is often accompanied bya reduction in growth, survival and flesh quality. Inaddition, sex related growth differences, usually infavour of females, occur in many cultured species. To-gether, this results in lower profitability (Carrillo et al.,1995a; Piferrer, Felip & Blázquez, 1995; Donaldson,1996). The application of chromosome set manipula-tion techniques (e.g., the induction of triploidy andgynogenesis) have been studied as a means to im-prove the production of many cultured fish species,especially freshwater, including mainly salmonids,cyprinids, cichlids, and ictalurids (Thorgaard, 1983;
Benfey, 1989; Ihssen et al., 1990; Solar, Donald-son & Douville, 1991). Induced triploidy resultingin genetic sterility and gynogenesis can be used toobtain all-female stocks. To date, the only speciesin which triploids are routinely cultured is the rain-bow trout, Oncorhynchus mykiss (Dillon, Schill &Teuscher, 2000). Sterilization can be useful to neg-ate precocious sexual maturation usually in males(Donaldson, 1997; Felip et al., 2001a, b). Moreover,triploidy induction can control unwanted reproductionin culture (Mair, 1993) and can be an advantage inthose species with high mortality during or after thespawning season (Ihssen et al., 1990; Benfey, 1999),as well as in those with a high gonadosomatic index

176
(Knibb, 2000). The production of sterile triploids hasother applications, for example, to prevent the interac-tion between farmed and wild fish (Donaldson, 1996,1997). On the other hand, all-female stocks producedthrough gynogenesis may be exploited for roe pro-duction in sturgeon and salmons (Ihssen et al., 1990;Shelton et al., 1997) and can be used in basic research(see § 6).
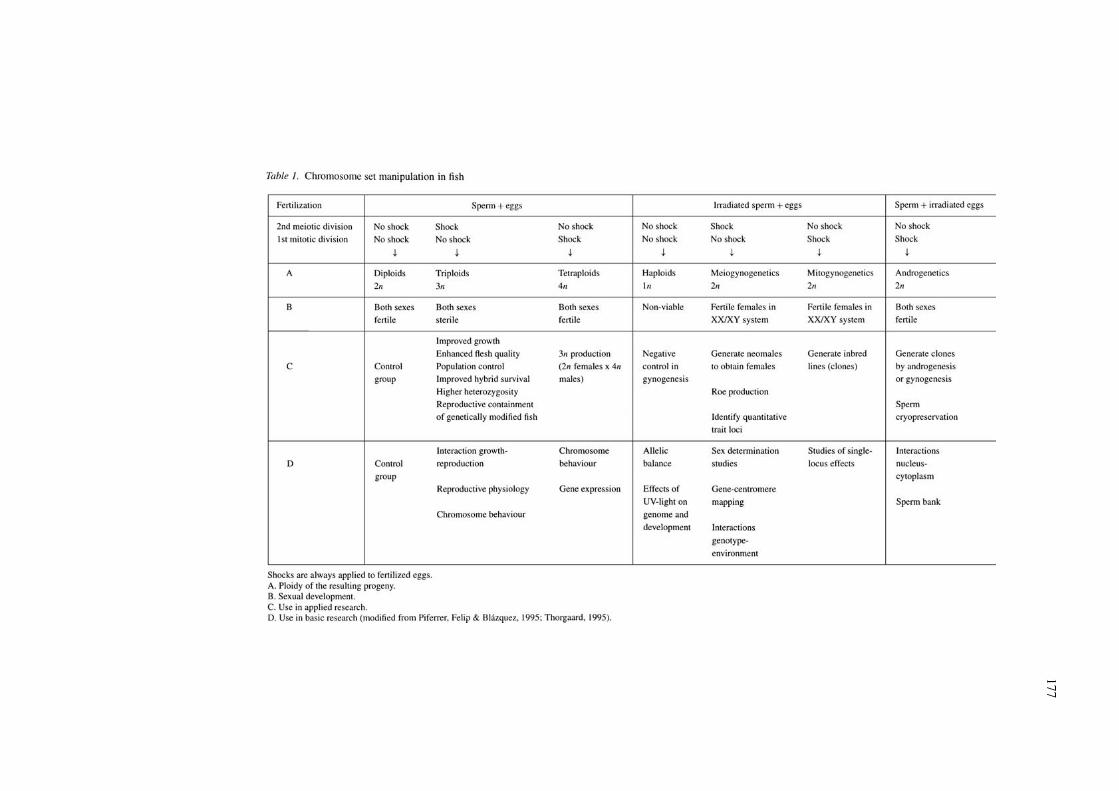
In comparison, fewer attempts on chromosome setmanipulation have been made in marine fish, eventhough pioneering works were carried out to inducetriploidy in flatfishes at the beginning of the 1970’s(see refer. in Lincoln 1981a). Since the late 1980’s,however, research focusing on assessing the condi-tions for ploidy manipulation has been carried out inabout a dozen different marine species, particularlyof the families Pleuronectidae, Sparidae, Paralich-thyidae, Moronidae, Soleidae, Oplegnathidae, andScophthalmidae. Similar to the situation observed infreshwater species, the induction of triploidy and gyn-ogenesis have been the most widely used types ofchromosome set manipulation (Table 1). There areno reports available to date on the induction of tet-raploidy and androgenesis in marine fish. However,tetraploidy has been induced in several freshwater spe-cies (Solar, Hajen & Donaldson, 1992), although theiractual reduced viability and reproductive performancehas limited their use for the production of triploids(Chourrout et al., 1986). The interest for rearing all-male stocks includes those cases where males growfaster than females (Mair et al., 1997) or have a highermarket value than females due to the presence of male-specific secondary sex characters (Kavumpurath &Pandian, 1992; Piferrer & Lim, 1997).
The purpose of this review is to provide a sum-mary of the studies carried out so far dealing withploidy manipulation, focusing on the methodologiesused for the production and subsequent identificationof triploid and gynogenetic fish. Results concerninggrowth performance of triploid and gynogenetic dip-loids, their reproductive activity and the proportion ofsexes are also summarized. Where appropriate or pos-sible, emphasis is given to marine species, in particularthe European sea bass (Dicentrarchus labrax). Thisspecies has great commercial value and is a modelorganism that has been used as a tool in research ongenetics and reproductive physiology (Carrillo et al.,1999), where a considerable amount of research hasalready been carried out concerning various aspectsof chromosome set manipulation (Zanuy et al., 1994;Carrillo et al., 1995b, 1999; Colombo et al., 1995,
1996, 1997; Gorshkova et al., 1995, 1996; Barbaroet al., 1996; Curatolo et al., 1996; Felip et al., 1997,1999a; Peruzzi & Chatain, 2000).
Induction of tripoidy and gynogenesis in fish
Broodstock and gamete quality for ploidymanipulation
A major requirement for ploidy manipulation in sea-water fish is a sound broodstock management programin order to obtain gametes of the best possible qual-ity (Wohlfarth, 1990; Bartley, 1997). Gametes frommarine species can be treated to yield triploid andgynogenetic fish (Table 2). However, hormonal induc-tion, maturation, ovulation and spawning of brood-stock as well as gamete collection, oocyte qualityassessment and sperm storage can be more demand-ing than for freshwater fish (Bromage, 1988; Planas& Cunha, 1999). This is the case because the eggsand newly hatched larvae from marine pelagic spawn-ers are usually smaller and more fragile than thoseof freshwater species (Bromage, 1988; Zohar, 1989;Planas & Cunha, 1999). If eggs are handled correctlyand collected during the optimal period of spawningfrom post-puberal broodfish, the toll on larval sur-vival can be disregarded (Colombo et al., 1995; Felipet al., 1997; Carrillo et al., 2000). However, thetriploidy yield can change among different femalesor experiments depending on the stage of maturationof the eggs (Díaz et al., 1993; Colihueque et al.,1996; Várkonyi et al., 1998), suggesting an importantgenetic component involved in the outcome.
In general, males spermiate throughout the en-tire reproductive season thus making sperm readilyavailable for collection. In the sea bass, they produceabundant sperm of excellent quality (9.4 ml/kg bodyweight, 5.5–60× 109 spermatozoa/ml and motility of70–100%) (Suquet et al., 1994; Sorbera et al., 1996;Fauvel et al., 1999; Felip et al., 1999a). Furthermore,artificial fertilization rates are usually high (Colomboet al., 1995; Felip et al., 1997, 1999a). However,in flatfish, males mature in captivity but the gam-etes sometimes cannot be easily removed by strippingwhich may represent an inconvenience for chromo-some set manipulation (Howell, Baynes & Thompson,1995).
In sea bass, differences in oocyte final matur-ation among females can difficult the productionof abundant good quality gametes (Alvariño et al.,
177

178
Table 2. Chromosome set manipulation in marine teleost fish
TypeaSpecies (common name) Triploidy Gynogenesis
bPleuronectes platessa x Platichthys flesus (plaice x flounder) Purdom, 1972bHippoglossus hippoglossus (Atlantic halibut) Holmefjord and Refstie, 1997cAcanthopagrus schlegeli (Black Sea bream) Arakawa et al., 1987cPagrus major (Red Sea bream) Arakawa et al., 1987 Sugama et al., 1990
Kitamura, Teong and Arakawa, 1991cSparus aurata (gilthead sea bream) Garrido-Ramos et al., 1996; Gorshkov et al., 1998
Gorshkov et al., 1998dParalichthys olivaceus (hirame) Tabata, 1991 Tabata, 1991eDicentrarchus labrax (European sea bass) Seei Seej
eMorone chrysops x Morone saxatilis (hybrid striped bass) Leclerc et al., 1996eMorone chrysops (white bass) Gomelsky et al., 1998fSolea solea (common sole) Howell, Baynes and
Thompson, 1995gOplegnathus fasciatus (Japanese parrot fish) Murata, 1998hScophthalmus maximus (turbot) Piferrer et al., 2000 F. Piferrer et al. (unpublished
observations)
aFamily: bPleuronectidae, cSparidae, dParalichthyidae, eMoronidae, fSoleidae, gOplegnathidae, hScophthalmidae. iCarrillo et al.,1993,1999; Zanuy et al., 1994; Colombo et al., 1995; Gorshkova et al.,1995; Felip et al.,1997. jCarrillo et al.,1993, 1999; Zanuy et al., 1994;Colombo et al., 1995; Gorshkova et al.,1996; Felip et al., 1999a.
1992). Hence, stocked females need to be periodic-ally checked by ovarian biopsy to select those withoocytes in a similar developmental stage for spawninginduction by hormones. Different useful criteria andprocedures for the selection of eggs have been reported(Carrillo et al., 2000). These include batch size, eggdiameter and buoyancy, and development to the firstdivisions (Howell, Baynes & Thompson, 1995; Felipet al., 1997; Gorshkov et al., 1998).
Induction of triploidy
Because fish exhibit external fertilization, their gam-etes may be easily handled for chromosome set manip-ulation. Table 1 shows treatments that interrupt earlycellular divisions of fertilized eggs for the inductionof triploidy and tetraploidy and the production of indi-viduals with uniparental inheritance (gynogenesis andandrogenesis).
Suppressing meiosis II to retain the second po-lar body in fertilized eggs produces triploids. This isachieved by applying a shock, which can be a cold,heat or pressure shock, always taking into accountthree important variables. These are: (a) the time afterfertilization when the shock is initiated, which canbe expressed in minutes or in mitotic intervals (Thor-gaard, 1983; Ihssen et al., 1990); (b) shock intensity;
and (c) shock duration. The degree of oocyte develop-ment and egg size is also important (Díaz et al., 1993;Colihueque et al., 1996; Várkonyi et al., 1998). Cold,heat and pressure shocks have been successfully usedin marine species (Table 3), while chemical shocksare used less often in fish. The use of cytochalasinB in Atlantic salmon, Salmo salar (Allen & Stan-ley, 1979) and colchicine in brook trout, Salvelinusfontinalis (Smith & Lemoine, 1979) resulted in thepresence of mosaic individuals, although mosaicismalso occurred albeit at a lower frequency when usingphysical treatments (Miller et al., 1994; Teplitz et al.,1994).
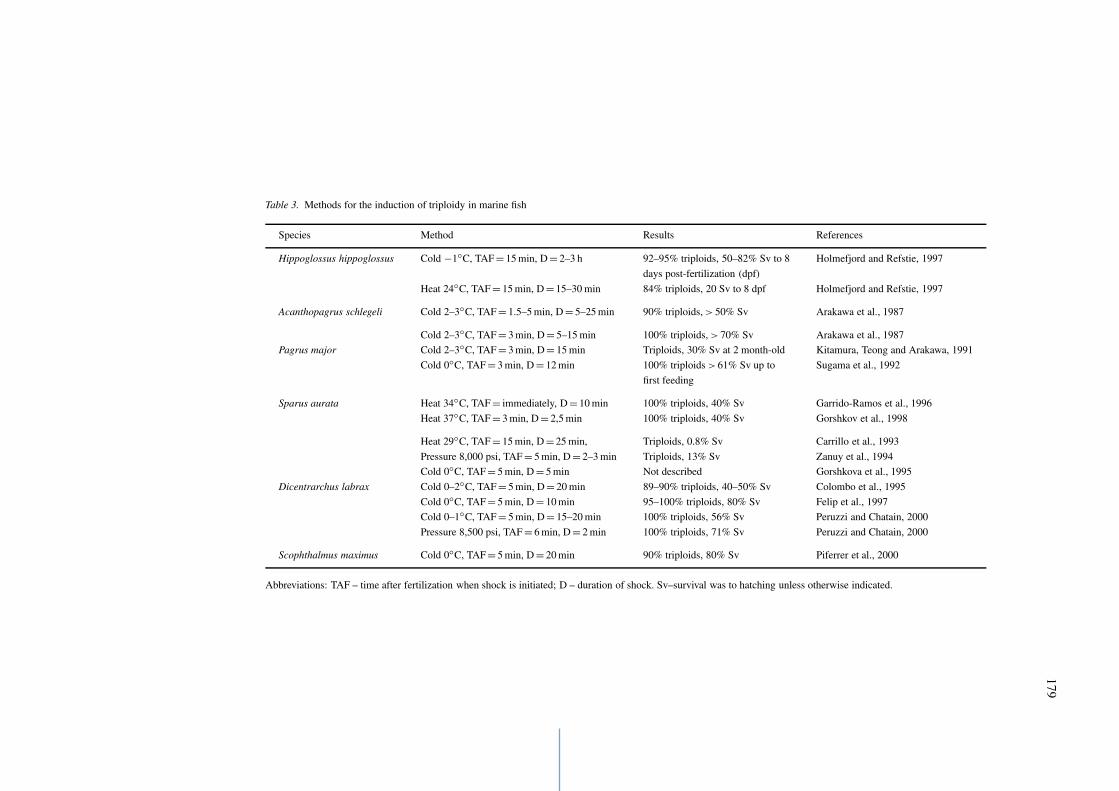
Cold shocks are usually applied in the range from−1◦C to 3◦C, yielding 84–100% triploids (Table 3),and have been the most commonly used to inducetriploidy in marine fish, although heat and pressureshocks also work. However, since high numbers oftriploids are obtained using optimized protocols, it isdifficult to say in general that there is an approach thatworks best for marine fish, although there has been apreference for cold shocks, as mentioned above. Theineffectiveness of pressure shock in the common sole,Solea solea (Howell, Baynes & Thompson, 1995), orcold shock in Atlantic salmon (Lincoln, Aulstad &Grammeltvedt, 1974) may have been due to the lack ofprotocol optimization. The advantage of cold shocks
179
Table 3. Methods for the induction of triploidy in marine fish
Species Method Results References
Hippoglossus hippoglossus Cold −1◦C, TAF = 15 min, D = 2–3 h 92–95% triploids, 50–82% Sv to 8 Holmefjord and Refstie, 1997
days post-fertilization (dpf)
Heat 24◦C, TAF = 15 min, D = 15–30 min 84% triploids, 20 Sv to 8 dpf Holmefjord and Refstie, 1997
Acanthopagrus schlegeli Cold 2–3◦C, TAF = 1.5–5 min, D = 5–25 min 90% triploids, > 50% Sv Arakawa et al., 1987
Cold 2–3◦C, TAF = 3 min, D = 5–15 min 100% triploids, > 70% Sv Arakawa et al., 1987
Pagrus major Cold 2–3◦C, TAF = 3 min, D = 15 min Triploids, 30% Sv at 2 month-old Kitamura, Teong and Arakawa, 1991
Cold 0◦C, TAF = 3 min, D = 12 min 100% triploids > 61% Sv up to Sugama et al., 1992
first feeding
Sparus aurata Heat 34◦C, TAF = immediately, D = 10 min 100% triploids, 40% Sv Garrido-Ramos et al., 1996
Heat 37◦C, TAF = 3 min, D = 2,5 min 100% triploids, 40% Sv Gorshkov et al., 1998
Heat 29◦C, TAF = 15 min, D = 25 min, Triploids, 0.8% Sv Carrillo et al., 1993
Pressure 8,000 psi, TAF = 5 min, D = 2–3 min Triploids, 13% Sv Zanuy et al., 1994
Cold 0◦C, TAF = 5 min, D = 5 min Not described Gorshkova et al., 1995
Dicentrarchus labrax Cold 0–2◦C, TAF = 5 min, D = 20 min 89–90% triploids, 40–50% Sv Colombo et al., 1995
Cold 0◦C, TAF = 5 min, D = 10 min 95–100% triploids, 80% Sv Felip et al., 1997
Cold 0–1◦C, TAF = 5 min, D = 15–20 min 100% triploids, 56% Sv Peruzzi and Chatain, 2000
Pressure 8,500 psi, TAF = 6 min, D = 2 min 100% triploids, 71% Sv Peruzzi and Chatain, 2000
Scophthalmus maximus Cold 0◦C, TAF = 5 min, D = 20 min 90% triploids, 80% Sv Piferrer et al., 2000
Abbreviations: TAF – time after fertilization when shock is initiated; D – duration of shock. Sv–survival was to hatching unless otherwise indicated.

180
over pressure shocks is that no special equipment isneeded and that large volumes of eggs can be treatedat once.
In general, the timing of the shock is the most crit-ical variable (Ihssen et al., 1990; Levanduski, Beck &Seeb, 1990; Felip et al., 1997, 1999a) since even smallvariations result in significant changes in the numberof triploids obtained, as demonstrated in the sea bass(Felip et al., 1997, 1999a). When using temperatureshocks, not only the temperature of the shock but alsothe difference between the pre-shock temperature andthe shock temperature is important and may affect thefinal yields (Chourrout, 1987; Ihssen et al., 1990).Changes in shock temperature are less important thanin shock timing while changes in duration appear tobe the least relevant. Exceptions to this have beendemonstrated in chinook (Levanduski, Beck & Seeb,1990) and pink salmon (Smoker, Crandell & Mat-suoka, 1995) where the duration of the thermal shockwas crucial for the retention of the second polar body.Finally, the outcome of shocks may also be affectedby oocyte quality and the degree of maturation whenthey are obtained from the females (Levanduski, Beck& Seeb, 1990; Díaz et al., 1993; Smoker, Crandell &Matsuoka, 1995; Várkonyi et al., 1998).
Induction of gynogenesis
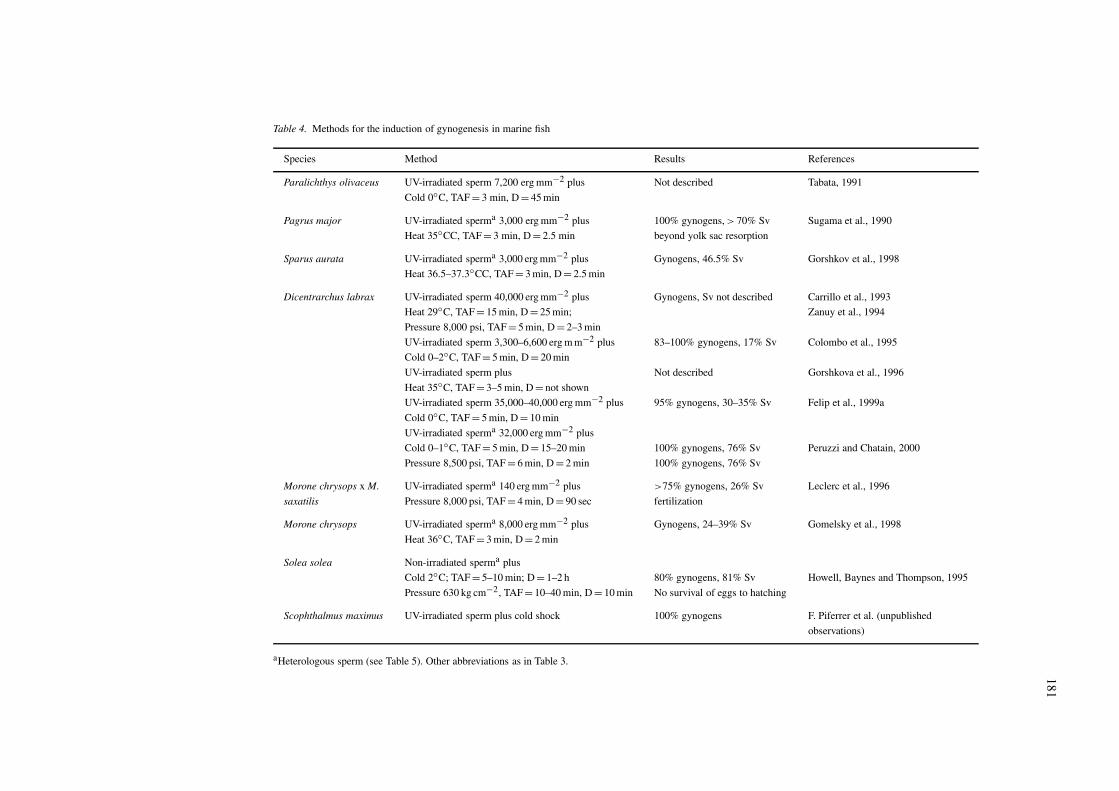
Gynogenetic diploid fish are obtained by first usingirradiated sperm to fertilize normal eggs and then sup-pressing the second meiotic division of these eggs(meiogynogenetic diploids) or the first mitotic divisionof the zygotes (mitogynogenetic diploids) (Table 1).The suppression of these cell divisions is carried outwith shocks similar to those used for the inductionof triploidy (compare Tables 3 and 4). Meiogyno-genetic fish retain significant levels of heterozygositydepending on the degree of crossing-over betweenhomologous chromosomes during meiosis I, whereasmitogynogenetic fish are completely homozygous atevery locus (Thorgaard, 1983; Ihssen et al., 1990).As a result, meiogynogenetic diploids have higherviability than mitogynogenetic diploids, which carrydeleterious alleles that negatively affect their survival(Leary et al., 1985; Don & Avtalion, 1988; Ihssenet al., 1990). Among species, yields are variable butgenerally low (Table 4). In marine teleosts, inductionof mitogynogenetic diploids has only been reportedin the sea bass with results showing a low percentageof viable embryos. However, survival in juvenile and
adult fish still needs to be determined (Colombo et al.,1995).
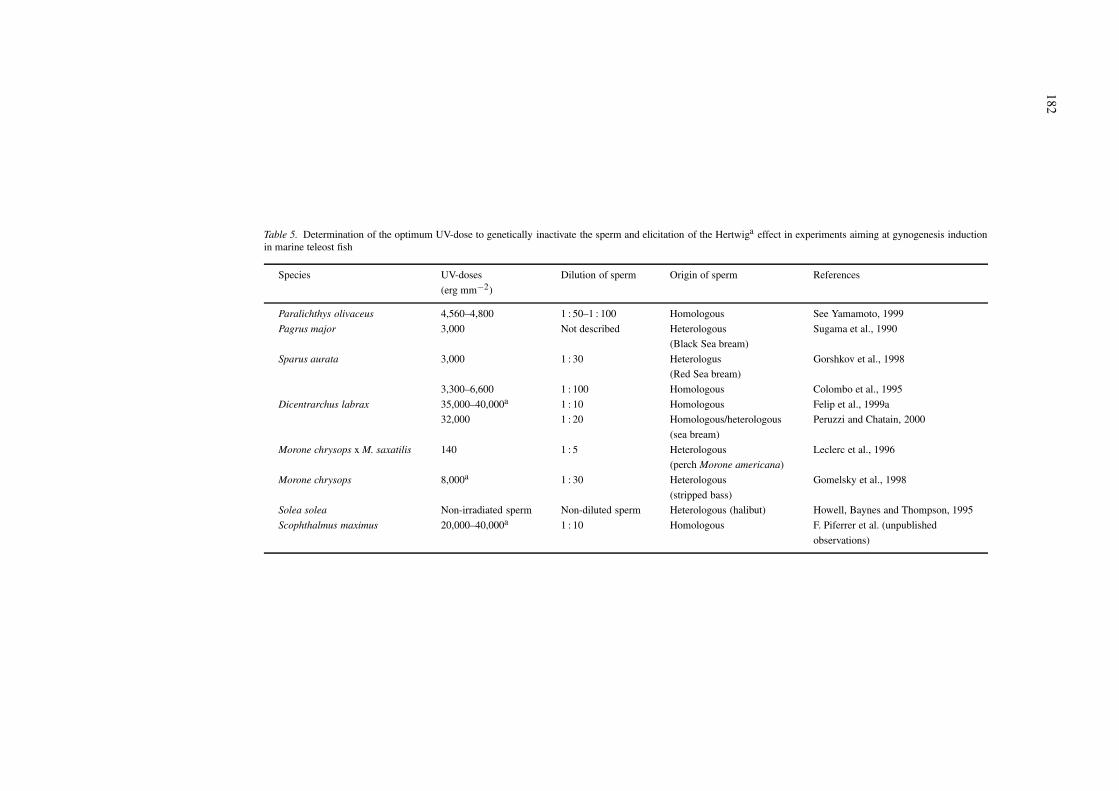
Several types of radiation, for example, ioniz-ing (X-rays, gamma rays) and ultraviolet (UV), andalso chemical treatments have been used to inactivatespermatozoan DNA (Thorgaard, 1983; Ihssen et al.,1990). Unlike ionizing radiation, UV light does notinduce small DNA paternal fragments (minichromo-somes) which may reduce the viability of gynogenet-ics (Thorgaard & Allen, 1987; Ihssen et al., 1990).The effects of UV light can be reversed by expos-ure to visible light, thus, UV- irradiated sperm mustbe protected in darkness until just before fertilizationto avoid a putative photoreactivation process (Ijiri &Egami, 1980). An alternative to sperm irradiation isthe use of sperm from an unrelated species (hetero-logous sperm) (Table 5) which, for added security,can also be UV-irradiated. It facilitates the identifica-tion of non-gynogenetic fish if hybrids are non-viable(Goudie et al., 1995; Smoker, Crandell & Matsuoka,1995; Fujioka, 1998; Gorshkov et al., 1998; Váradiet al., 1999). The appropriate dose of UV light isanother important aspect to effectively inactivate thesperm (Table 5). Thus, a range of doses must be as-sayed. Batches of eggs are fertilized by diluted spermirradiated with increasing doses of UV light inducinga paradoxical phenomenon called the Hertwig effect.This has been elicited in both freshwater (Pongthanaet al., 1995; Lin & Dabrowski, 1996; Palti, Li &Thorgaard, 1997) and marine species (Gomelsky et al.,1998; Felip et al., 1999a). Embryos resulting fromthe fertilization of normal eggs with completely in-activated sperm exhibit a typical ‘haploid syndrome’,for example, inviable embryos or newly-hatched lar-vae with severe body malformations (Pongthana et al.,1995; Lin & Dabrowski, 1996; Felip et al., 1999a).
In many fish species, the irradiation of UV lightdecreases the motility of spermatozoa (Goudie et al.,1995; Felip et al., 1999a). Thus, lower viability at-tributed to gynogenetics could simply be due to lowerfertilization rates as a consequence of reduced num-bers of motile spermatozoa. In sea bass, the exposureof sperm to UV light (2,500–55,000 erg mm−2) re-duced the amount of motile spermatozoa but did notsignificantly affect the duration of motility in the re-maining motile spermatozoa (Felip et al., 1999a). Thisstudy also demonstrated that the thermal shock negat-ively influenced the survival at fertilization, althoughits effects decreased during embryogenesis and disap-peared by hatching, suggesting that the lower viabilityof some triploids is a direct consequence of the shock,
181
Table 4. Methods for the induction of gynogenesis in marine fish
Species Method Results References
Paralichthys olivaceus UV-irradiated sperm 7,200 erg mm−2 plus Not described Tabata, 1991
Cold 0◦C, TAF = 3 min, D = 45 min
Pagrus major UV-irradiated sperma 3,000 erg mm−2 plus 100% gynogens, > 70% Sv Sugama et al., 1990
Heat 35◦CC, TAF = 3 min, D = 2.5 min beyond yolk sac resorption
Sparus aurata UV-irradiated sperma 3,000 erg mm−2 plus Gynogens, 46.5% Sv Gorshkov et al., 1998
Heat 36.5–37.3◦CC, TAF = 3 min, D = 2.5 min
Dicentrarchus labrax UV-irradiated sperm 40,000 erg mm−2 plus Gynogens, Sv not described Carrillo et al., 1993
Heat 29◦C, TAF = 15 min, D = 25 min; Zanuy et al., 1994
Pressure 8,000 psi, TAF = 5 min, D = 2–3 min
UV-irradiated sperm 3,300–6,600 erg m m−2 plus 83–100% gynogens, 17% Sv Colombo et al., 1995
Cold 0–2◦C, TAF = 5 min, D = 20 min
UV-irradiated sperm plus Not described Gorshkova et al., 1996
Heat 35◦C, TAF = 3–5 min, D = not shown
UV-irradiated sperm 35,000–40,000 erg mm−2 plus 95% gynogens, 30–35% Sv Felip et al., 1999a
Cold 0◦C, TAF = 5 min, D = 10 min
UV-irradiated sperma 32,000 erg mm−2 plus
Cold 0–1◦C, TAF = 5 min, D = 15–20 min 100% gynogens, 76% Sv Peruzzi and Chatain, 2000
Pressure 8,500 psi, TAF = 6 min, D = 2 min 100% gynogens, 76% Sv
Morone chrysops x M. UV-irradiated sperma 140 erg mm−2 plus >75% gynogens, 26% Sv Leclerc et al., 1996
saxatilis Pressure 8,000 psi, TAF = 4 min, D = 90 sec fertilization
Morone chrysops UV-irradiated sperma 8,000 erg mm−2 plus Gynogens, 24–39% Sv Gomelsky et al., 1998
Heat 36◦C, TAF = 3 min, D = 2 min
Solea solea Non-irradiated sperma plus
Cold 2◦C; TAF = 5–10 min; D = 1–2 h 80% gynogens, 81% Sv Howell, Baynes and Thompson, 1995
Pressure 630 kg cm−2, TAF = 10–40 min, D = 10 min No survival of eggs to hatching
Scophthalmus maximus UV-irradiated sperm plus cold shock 100% gynogens F. Piferrer et al. (unpublished
observations)
aHeterologous sperm (see Table 5). Other abbreviations as in Table 3.
182
Table 5. Determination of the optimum UV-dose to genetically inactivate the sperm and elicitation of the Hertwiga effect in experiments aiming at gynogenesis inductionin marine teleost fish
Species UV-doses Dilution of sperm Origin of sperm References
(erg mm−2)
Paralichthys olivaceus 4,560–4,800 1 : 50–1 : 100 Homologous See Yamamoto, 1999
Pagrus major 3,000 Not described Heterologous Sugama et al., 1990
(Black Sea bream)
Sparus aurata 3,000 1 : 30 Heterologus Gorshkov et al., 1998
(Red Sea bream)
3,300–6,600 1 : 100 Homologous Colombo et al., 1995
Dicentrarchus labrax 35,000–40,000a 1 : 10 Homologous Felip et al., 1999a
32,000 1 : 20 Homologous/heterologous Peruzzi and Chatain, 2000
(sea bream)
Morone chrysops x M. saxatilis 140 1 : 5 Heterologous Leclerc et al., 1996
(perch Morone americana)
Morone chrysops 8,000a 1 : 30 Heterologous Gomelsky et al., 1998
(stripped bass)
Solea solea Non-irradiated sperm Non-diluted sperm Heterologous (halibut) Howell, Baynes and Thompson, 1995
Scophthalmus maximus 20,000–40,000a 1 : 10 Homologous F. Piferrer et al. (unpublished
observations)

183
Table 6. Methods used to identify triploidy in marine fish
Species Method References
Hippoglossus hippoglossus Flow cytometry Holmefjord and Refstie, 1997
Acanthopagrus schlegeli Not described Arakawa et al., 1987
Pagrus major Karyotyping Kitamura, Teong and Arakawa, 1991
Erythrocyte size Kitamura, Teong and Arakawa, 1991; Sugama et al., 1992
Allozyme markers Sugama et al., 1992
Sparus aurata Karyotyping Garrido-Ramos et al., 1996; Gorshkov et al., 1998
Dicentrarchus labrax Karyotyping Carrillo et al., 1993; Zanuy et al., 1994
Gorshkova et al., 1995; Colombo et al., 1995; Felip et al., 1997
Silver staining Felip et al., 1997
Erythrocyte size Felip et al., 1997
Flow cytometry Carrillo et al., 1993; Zanuy et al., 1994
Colombo et al., 1995; Peruzzi and Chatain, 2000
Scophthalmus maximus Karyotyping
Silver staining Piferrer et al., 2000
not of the triploid condition per se. In contrast, UVlight had an effect more pronounced as developmentprogressed from fertilization onwards, reaching itsmaximum at hatching. Therefore, mortality is higherin gynogenetic fish than in triploid fish. This reflectsthe manipulation of both male and female gametes aswell as the effects of the higher homozigosity com-pared to triploids (Leary et al., 1985; Don & Avtalion,1988). Thus, sperm irradiation rather than the thermalshock has been shown to be the major cause of in-creased mortality in gynogenetics (Lin & Dabrowski,1996; Felip et al., 1999a). Deleterious effects of solarUV radiation affecting survival and development havebeen documented in aquatic ecosystems (Häder, 1986,1993), and in amphibians (Ankley et al., 2000) and fish(Bass & Sistrun, 1997; Béland et al., 1999). In the seabass, the yield of triploids and gynogenetics is similarto that observed in freshwater fish rather than to whathad typically been observed in marine (see Tables 3and 4).
Identification of triploids
Methods to identify triploid fish are presented inTable 6. They include chromosome count or karyotyp-ing (Thorgaard & Disney, 1990), selective stainingof the nucleolar organizer regions (NORs) (Flajshans,Ráb & Dobosz, 1992; Kucharczyk et al., 1999), cel-lular size (Felip et al., 1997, 1999b; Benfey, 1999)
and direct measurement of the nuclear DNA content(Allen, 1983; Ewing & Scalet, 1991; Lecommandeur,Haffray & Philippe, 1994). Comparison of the differ-ent methods can be found in Harrell, Van Heukelemand Kerby (1998). In marine species, karyotyping hasbeen the most commonly used method so far to dis-tinguish between diploids and triploids, for example,48 chromosomes versus 72 chromosomes in sea bass,gilthead sea bream and Red Sea bream, whereas turbothad 44 chromosomes in diploids and 66 in triploids(see refs. in Table 6). Karyotyping, however, is time-consuming and silver staining of NORs can be used asan alternative (Harrell, Van Heukelem & Kerby, 1998).Analysis of NORs is only useful in those species withone nucleolar organizer region per haploid genome,with one or two NORs detected per cell in diploidsand up to three NORs in triploids. Thus, one potentialproblem with this method can be the presence of apolymorphism in the number of NORs, resulting ina lack of correlation between the number of NORsand the ploidy level. In such cases, as for examplein the turbot, it needs to be demonstrated that such apolymorphism affects only a low number of fish andthat it does not interfere with the correct assessment ofploidy (Piferrer et al., 2000).
As cells in triploid fish have 50% more DNA dueto an extra set of chromosomes, cell volume increasesabout 1.5 times in triploids. Thus, cells of triploids arelarger than those of diploids (Benfey, 1999). Hence,

184
Table 7. Methods used to identify gynogenesis in marine fish
Species Methoda References
Pagrus major Allozyme markers Sugama et al., 1990
Sparus aurata Karyotyping Gorshkov et al., 1998
Dicentrarchus labrax External morphology Carrillo et al., 1993; Zanuy et al., 1994;
Colombo et al., 1995; Felip et al., 1999a
Karyotyping Colombo et al., 1995; Gorshkova et al., 1995;
Felip et al., 1997
Silver staining Felip et al., 1997
AFLP markers Felip et al., 2000
Microsatellites Peruzzi and Chatain, 2000
Morone chrysops x M. saxatilis DNA amplification Leclerc et al., 1996
techniques
Morone chrysops External morphology
Morphological characters Gomelsky et al., 1998
Karyotyping
Solea solea External morphology Howell, Baynes and Thompson, 1995
Scophthalmus maximus External morphology
Karyotyping F. Piferrer et al. (unpublished observations)
Silver staining
Microsatellites
aExternal morphology based on haploid syndrome; Karyotyping based on numbers of chromosomes withthe exception of Sparus aurata showing ‘marker’ chromosomes, that is, number of biarmed chromosomes;Morphological characters in Morone chrysops based on lack of melanophores.
the length of the major axis of erythrocytes has beenused to rapidly confirm triploidy in several species,especially if a particle size analyzer is available (Wat-tendorf, 1986). Cytofluorometric analysis is anotheralternative that allows processing large amounts ofsamples in a relatively short period of time but, as inthe previous case, its major drawback is the high costof equipment.
Identification of gynogenetics
Methods to identify gynogenetic fish are summarizedin Table 7. The restoration of diploidy in gynogen-etic fish can easily be confirmed by karyotyping andNOR analysis. However, the true assessment of fishwith exclusively maternal inheritance can only bedemonstrated by using morphological, biochemical ormolecular genetic markers (Johnstone & Stet, 1995;Leclerc et al., 1996; Van Eenennaam et al., 1996; Felipet al., 2000; Peruzzi & Chatain, 2000).
In fish, the number of studies using heritable mor-phological traits for paternity testing is low (Stanley &
Jones, 1976; Nagy et al., 1978; Suzuki, Oshiro & Na-kanishi 1985; Don & Avtalion, 1988). Moreover, evenin the case of species exhibiting these morphologicalvariants, other methods of verification are simultan-eously carried out to determine parental assignment(Don & Avtalion, 1988; Gomelsky et al., 1998). Al-though the ‘haploid syndrome’ is not a morphologicalmarker, it is an external feature in embryos or develop-ing larvae with only one set of chromosomes. Duringgynogenesis induction, the presence of embryos withthis syndrome indicates genetic inactivation of the ir-radiated sperm and thus serves for the identification ofputative gynogenetic fish (see Table 7).
Studies based on protein and/or allozyme variantshave had few applications for paternity testing in fish.The reason may be associated with two main limita-tions of this methodology: the low proportion of poly-morphic loci and the reduced allelic diversity of theseloci (2–3 alleles/locus) making the heterozygosity rateand the genetic variability low (Magoulas, 1997). Incontrast, molecular markers are increasingly being ap-plied because, firstly, DNA genomic sequence variants

185
Table 8. Growth in body weight (BW) of triploid (3n) versus diploid (2n) fish
Environment aSpecies Life stage Growth References
Marine species bPleuronectes platessa x Juveniles 3n = 2n Purdom, 1972
Platichthys flesus
Adults 3n > 2n Lincoln, 1981cPagrus major Juveniles 3n < 2n See Arai, 2001dParalichthys olivaceus Juveniles 3n < 2n See Arai, 2001eDicentrarchus labrax Juveniles 3n = 2n Felip et al., 1999b
Adults 3n < 2n Felip et al., 2001a
Freshwater species fTilapia aurea Juveniles 3n > 2n Valenti, 1975gCyprinus carpio Juveniles 3n = 2n Gervai et al., 1980
Adults 3n < 2n Cherfas et al., 1994gCtenopharyngodon idella Juveniles 3n = 2n Cassani and Caton, 1985
Oliva-Teles and Kaushik, 1990
3n < 2n Friars et al., 2001hIctalurus punctatus Juveniles 3n = 2n Wolters, Libey and Chrisman, 1982
Adults >8 months 3n > 2niOncorhynchus mykiss Juveniles 3n < 2n Thorgaard et al., 1982
Juveniles, 118 days 3n = 2n Myers and Hershberger, 1991
Adults 3n > 2n Chourrout et al., 1986
Guo, Hershberger and Myers, 1990iOncorhynchus kisutch Juveniles, 17 months 3n < 2n Utter et al., 1983
Juveniles, 30 months 3n = 2n Johnson, Dickhoff and Utter, 1986iSalmo salar Juveniles 3n = 2n Benfey and Sutterlin, 1984
McGeachy, Benfey and Friars, 1995iOncorhynchus mykiss x Juveniles 3n < 2n Parsons et al., 1986
O. kisutchiOreochromis mossambicus 21–79 weeks 3n ≤ 2n Penman, Skibinski and Beardmore, 1987iOreochromis aureus 21–79 weeks 3n ≤ 2n Penman, Skibinski and Beardmore, 1987iOreochromis niloticus 21–79 weeks 3n ≤ 2n Penman, Skibinski and Beardmore, 1987
10–40 weeks 3n = 2n Hussain et al., 1995
Adults 3n > 2n Brämick et al., 1995iOncorhynchus gorbuscha Adults 3n = 2n Benfey et al., 1989iOncorhynchus masou 10–13 months 3n = 2n Nakamura et al., 1993iSalmo salar x Salmo trutta Variable results Galbreath and Thorgaard, 1994iHybrids of Oncorhynchus mykiss Adults 3n ≤ 2n Blanc, Vallee and Dorson, 2000jClarias gariepinus 149–205 days 3n = 2n Henken, Brunink and Richter, 1987jClarias macrocephalus 8 months 3n > 2n Fast et al., 1995kPerca flavescens Juveniles 3n < 2n Malison et al., 1993kStizostedium vitreum Variable results Malison and Garcıa-Abiado, 1996lMisgurnus mizolepis 2 months 3n = 2n Kim, Jo and Lee, 1994
3–9 months 3n > 2n Kim, Jo and Lee, 1994mPomoxis annularis x Variable results Parsons and Meals, 1997
P. nigromaculatusnOplegnathus fasciatus Juveniles 3n < 2n See Arai, 2001
aFamily: bPleuronectidae, cSparidae, dParalichthyidae, eMoronidae, fCichlidae, gCyprinidae, hIctaluridae, iSalmonidae, jClariidae,kPercidae, lCobitidae, mCentrarchidae, nOplegnathidae.Data includes both sexes. Diploids and triploids were reared in separate tanks.

186
result in a high polymorphism rate and, secondly,small amounts of tissue are required since DNA can beamplified using the polymerase chain reaction (PCR)(Zhang, Tiersch & Cooper, 1994; Takagi & Tanigu-chi, 1995; Partis & Wells, 1996; Williams, Kazianis& Walter, 1998). Thus, molecular approaches provideunambiguous identification of gynogenetics and arepreferred nowadays.
The technique of DNA fingerprinting, based onvariable numbers of tandem repeats, has been the mostwidely used method to identify gynogenetic fish ina variety of freshwater species (Carter et al., 1991;Volckaert et al., 1994; Johnstone & Stet, 1995; VanEenennaam et al., 1996). To date, the only marinespecies for which true gynogenesis has been proven bymolecular markers (DNA amplification or microsatel-lite analysis) are the hybrid stripped bass (Leclercet al., 1996), European sea bass (Felip et al., 2000;Peruzzi & Chatain, 2000) and turbot (F. Piferrer et al.,unpublished observations) (see Table 7). In the case ofthe European sea bass, the number of true gynogeneticfish produced was 96.5% based on amplified fragmentlength polymorphism (AFLP) markers (Felip et al.,2000).
Performance of triploid fish
Survival and growth
The increased cell size in polyploid fish is balancedwith decreased cell numbers, resulting in fish hav-ing larger cells but not necessarily larger size (Ihssenet al., 1990; Benfey, 1999). Furthermore, triploidymay affect different tissues in a different manner, pro-ducing mosaic individuals. However, no informationis available on the effect of mosaicism on the viabilityand growth performance of these fish (Teplitz et al.,1994).
Most studies on chromosome set manipulation inmarine species have focused on finding the right com-bination of treatment variables to induce triploidy orgynogenesis. Hence, very little research has beenconducted in studying the growth and the reproduc-tion of these fish. This contrasts with the situationin freshwater species where, in addition, a varietyof physiological and behavioral aspects have beenstudied including hematological parameters, oxygenconsumption and aerobic capacity, immunocompet-ence, stress response, osmoregulation, diet utilizationand energetics, sensory perception, and behavior (see
Ihssen et al., 1990; Benfey, 1999, for reviews). Theavailable data published so far revealed that the growthof triploids was dependent on the species concernedand, within the same species, on the age (Thorgaard,1983; Ihssen et al., 1990), rearing conditions (Cherfaset al., 1994; Carter et al., 1994; O’Keefe & Benfey,1997), methodologies used (Thorgaard, 1983; Ihssenet al., 1990; Malison et al., 1993), and reproductivestrategy (Benfey, 1999; Knibb, 2000).
With regard to marine species, the accumulateddata on growth in induced triploids are scarce andsummarized in Table 8, which also includes data onfreshwater species for comparison. Female plaice xmale flounder (Pleuronectes platessa x Platichthysflesus) triploid hybrids grew more than the correspond-ing non-hybrid diploids during the spawning season(Lincoln, 1981). However, diploids reached growthrates similar to those of triploids after sexual matura-tion, thus negating any previous advantage in somaticgrowth. In the sea bass, triploids grew in a patternsimilar to diploids during the juvenile stage. Never-theless, in spite of the gametic sterility, adult triploidsgrew slower than adult diploids even when diploidsmature during the spawning season in winter. Thissuggests that the low growth rate of this species duringwinter depends more on temperature than on the ener-getic cost associated with reproduction (Felip et al.,2001a).
In many species where females grow faster thanmales such as the olive flounder, sea bass and tur-bot, triploid females also show enhanced growth rates,indicating that there is a strong genetic, sex-relatedcomponent in this differential growth. However, sincetriploid females are completely sterile while triploidmales are not, a non genetic component in the formsex-related differences in the energy available forgrowth may also be present. Hence, the establishmentof all-female triploid stocks could benefit the rearingof these species. In freshwater species, this approachhas been considered (Benfey et al., 1989; Piferrer,Benfey & Donaldson, 1994; Galbreath & Thorgaard,1995; Sheehan et al., 1999). Research in rainbow trouthas demonstrated better growth in all-female triploidstocks as compared to those with mixed sex stocks(Sheehan et al., 1999). In contrast, all-female triploidAtlantic salmon appear to have no greater advant-age over mixed-sex cultures (Galbreath et al., 1994;Galbreath & Thorgaard, 1995).
The presence of jaw, operculum and vertebralcolumn deformities has been reported in geneticallymanipulated fish but these deformities have not been

187
related to chromosome set manipulation practices(Sugama et al., 1992; Felip et al., 1999b). Instead, theyare considered to be due to handling during artificialfertilization, inbreeding or chromosomal aberrations,since captive broodstock are typically used. In fact,these deformities are encountered in a similar pro-portions in wild fish as well as in diploid controlsmaintained under similar rearing conditions (Cerdáet al., 1994; Colihueque et al., 1996; Felip et al.,1999b; Tudor & Katavic, 1999). Together, these factssuggest that development and the external morphologyof triploid fish is essentially similar to that of diploids.
Gonadal development
Induced triploidy disrupts gametogenesis with morepronounced effects in females than in males (Ben-fey, 1999). Rudimentary ovaries in triploid femaleshave been observed in salmonids (Benfey & Sutterlin,1984; Johnson, Dickhoff & Utter, 1986; Nakamuraet al., 1987), channel catfish (Wolters, Libey & Chris-man, 1982), yellow perch (Perca flavescens) (Malisonet al., 1993), and mud loach (Misgurnus mizolepis)(Kim, Jo & Lee, 1994). However, in a few other spe-cies, triploid females have managed to develop ovariessimilar to those of diploids, as it is the case of Tilapiasp. (Freund, Hörstgen-Schwark & Holtz, 1995), whitesturgeon (Acipenser transmontanus) (Van Eenennaamet al., 1990), carp (Cyprinus carpio) (Wu et al., 1993;Cherfas et al., 1994), and brook trout (Benfey, 1995).An inappropriate follicular structure in the ovary anda greatly reduced production of steroid hormones areoften blamed for the absence of gonadal developmentin triploid females (Mol et al., 1994; Hussain et al.,1995; Benfey, 1999; Felip et al., 2001b). Althoughno endocrine signs of sexual maturation occur in fe-male triploids, some studies have reported their re-sponse to stimulation with sex steroids (Benfey et al.,1989; Breton & Sambroni, 1996). In contrast, trip-loid males display testes similar to those of diploidsand reach sexual maturation but produce dilute, ab-normal and non-viable aneuploid sperm (Kawamuraet al., 1999; Koedprang & Na-Nakorn, 2000; Tiwary,Kirubbagaran & Ray, 2000; Wills et al., 2000). Mo-reover, since testes in triploid males contain ster-oidogenic cells, they exhibit hormonal profiles andsecondary sexual characters similar to those of dip-loids (Nakamura et al., 1993; Mol et al., 1994; Hussainet al., 1995).
Impaired gametogenesis as a consequence of trip-loidy has been observed in marine fish species such
as plaice x flounder triploid hybrids (Lincoln, 1981),Red Sea bream (Kitamura, Teong & Arakawa, 1991;Sugama et al., 1992) and sea bass (Felip et al., 2001b).The induction of triploidy in the latter species con-ferred functional sterility in males and females, thusproving a good model to describe the effects of trip-loidy on the gonadal development in the two sexes.It was found that triploidy blocked the initial phasesof meiosis in females and the late phases of meiosisin males, resulting in the absence of or a reduction ingonadal development, respectively. Specifically, in fe-males, triploidy blocked meiosis during the zygotene,preventing the pairing of homologous chromosomes.In contrast, in males, triploidy blocked meiosis dur-ing the transformation of secondary spermatocytesinto spermatids, thus preventing spermiogenesis (Felipet al., 2001b). To our knowledge, this is the first casereported in fish where triploidy completely interruptedspermatogenesis and, since males are the predominantsex of this species in captivity, this may have economicbenefits for its aquaculture.
However, the ultimate reason as to why triploidydifferentially affects gonadal development betweensexes is still unknown. Differences associated with mi-totic rates between spermatogonia and oogonia beforethe start of meiosis have been considered (Thorgaard,1983; Krisfalusi & Cloud, 1997). Recent studies deal-ing with meiotic chromosome behavior can be usefulto determine the causes of aberrant meiosis in poly-ploid fish (Gui et al., 1992; Zhang & Arai, 1999).These studies indicate that abnormalities occur amonghomologous chromosomes, affecting the synaptic pro-cesses during the zygotene stage, although the syn-aptonemal complex is still formed (Gui et al., 1992;Carrasco et al., 1998; Zhang & Arai, 1999). Further re-search concerning meiotic chromosomal associationsin Nile tilapia revealed a drastic aberration in chromo-some pairing in oocytes but not in spermatocytes as acause of gonadal sterility in females (Carrasco et al.,1998). In humans, male sterility has been attributedto an abnormal pairing between sex chromosomes andunpaired autosomal chromosomes (Rosenmann et al.,1985).
Proportion of sexes
Changes in genome size or allelic proportions as aconsequence of triploidy induction apparently shouldnot result in deviations in the proportion of sexes infish (Ihssen et al., 1990; Hussain et al., 1995; Felipet al., 2001a). However, altered sex ratios in favor

188
of males have been observed in triploids of Rhodeusocellatus ocellatus (Kawamura et al., 1999), zebrafish,Brachydanio rerio (Kavumpurath & Pandian, 1990),Red Sea bream (Kitamura, Teong & Arakawa, 1991),carp (Cherfas et al., 1994) and Puntius gonionotus(Koedprang & Na-Nakorn, 2000). In contrast, trip-loidy increased the number of females in Oreochromisaureus (Mol et al., 1994), catfish (Goudie et al., 1995)and hybrids of Lepomis spp. (Wills, Sheehan & Allen,2000). In the sea bass, the number of females was 10%higher in triploids than in diploids. The sex ratio intriploids was 2 : 1 (male:female) in comparison to theproportion of sexes found in captive diploid sea bass(3 : 1) in our experiment (Felip et al., 2001a). Accord-ing to Benfey (1999), the abnormalities observed insex proportions in triploids can be due to a differen-tial mortality or the effects of environmental factors.Nevertheless, it remains to be determined if a doublematernal genomic contribution or an extra copy of oneof the sex chromosomes in triploids may influence theexpression of sex determining genes, thus possibly ex-plaining the alterations observed in the sex ratios ofthese fish.
The sex ratios of triploids (and particularly ofgynogenetics) can contribute to elucidate the sex de-termination mechanism of a species (Thorgaard, 1983;Ihssen et al., 1990). Thus, in species with femalehomogamety, the crosses involving XX females withXY males followed by triploidy induction result in asimilar percentage of both sexes, that is, 50% XXXtriploid females and 50% XXY triploid males. Thiswas demonstrated in catfish (Goudie et al., 1995)and Nile tilapia (Hussain et al., 1995). Gynogenesisinduction in these species corroborated a XX-XY sys-tem (Mair et al., 1991a; Goudie et al., 1995). Inthe sea bass, results obtained so far discard an XX-XY system. Firstly, because the sex ratio in triploidsdeviates from a proportion of 1 : 1 and, secondly, be-cause offspring of gynogenetics are of both sexes(Felip, 2000). Meanwhile, in species with male ho-mogamety (ZZ-ZW system), sex ratios in offspringcan be variable depending on the crossing-over, whichcan result in an equal number of males and femalesor an increased number of females (Thorgaard, 1983).In those species with polygenic or environmentalsex determination, the sex ratio will be substantiallyaltered in offspring originating from the same breedersdepending on the balance of sex-determining genesand the influence of environmental factors, respect-ively (Thorgaard, 1983; Yamazaki, 1983; Piferrer,2001).
Performance of gynogenetic diploid fish
Survival and growth
Most studies on gynogenetic diploids stop when thesefish are still quite young probably due to their lowersurvival (Table 4). Thus, information concerningtheir growth performance is available for only a fewspecies. In mud loach (Suzuki, Oshiro & Kakan-ishi, 1985) and Atlantic salmon (Johnstone & Stet,1995), gynogenetics grew less than their correspond-ing controls. In contrast, in Clarias macrocephalus(Na-Nakorn, 1995) and Gnathopogon caerulescens(Fujioka, 1998) gynogenetics had growth rates sim-ilar to those of controls. Comparable growth betweengynogenetics and diploids was found in the two mar-ine species examined so far: Red Sea bream (Sugamaet al., 1990) and sea bass (Felip, 2000). In these spe-cies gynogenetic fish had a survival of 30–35% withrespect to that of the controls one day post-hatching(Felip et al., 1999a, 2000a).
Gonadal development
Gynogenetic females of many species exhibit ovariescontaining undeveloped oocytes or areas filled withconnective tissue. This is likely due to the expressionof deleterious recessive alleles affecting the normalcourse of key reproductive processes (Piferrer, Benfey& Donaldson, 1994; Fujioka, 1998). These alterationshave been demonstrated in carp (Nagy et al., 1978;Komen et al., 1992), coho salmon (Piferrer, Benfey& Donaldson, 1994) and Gnathopogon caerulescens(Fujioka, 1998). In contrast, gynogenetic femalesin mud loach (Suzuki, Oshiro & Nakanishi, 1985)and Clarias macrocephalus (Na-Nakorn, 1995) haveshown normal ovaries and fertility. In the Europeansea bass, histological analysis revealed normal devel-opment of ovaries and testes during the first 2 yearsof age. Furthermore, the fertility and quality of thegametes of these fish was not affected by gynogenesis(Felip, 2000).
The induction of gynogenesis has also been car-ried out for monosex female production (Thorgaard,1983; Price, 1984; Ihssen et al., 1990). To overcometheir low survival, gynogenetic sex-reversed XX fe-males (i.e., phenotypic males) may be produced andcrossed with normal females to produce 100% femaleswith a higher viability (Ihssen et al., 1990; Pandian &Varadaraj, 1990). Crosses between these sex-reversedgynogenetic fish and normal females have been per-formed in several species including Oreochromis

189
Table 9. Gynogenesis induction: percentage of females in offspring and sex determining system
Environment Speciesa References Females (%) Sex determination
Marine species bParalichthys olivaceus Tabata (1991) 97.1 XX-XYm
See Yamamoto, 1999 48–93 XX-XYm
cDicentrarchus labrax Felip (2001b) 50 UnknowndSolea solea Howell, Baynes and Thompson (1995) 40 Unknown
Freshwater species eOreochromis mossambicus Pandian and Varadaraj (1990) 100 XX-XYeOreochromis niloticus Mair et al. (1991a) 100 XX-XYfCtenopharyngodon idella Stanley (1976) 100 XX-XYfCyprinus carpio Nagy et al. (1978) 100 XX-XYfCarassius auratus Oshiro (1987) 0–100 XX-XYm
fPuntius gonionotus Pongthana et al. (1995) 100 XX-XYfGnathopogon caerulescens Fujioka (1998) 87.2 XX-XYm
gIctalurus punctatus Goudie et al. (1995) 100 XX-XYhOncorhynchus kisutch Refstie, Stoss and Donaldson (1982) 49.3–100 XX-XYm
hSalmo salar Johnstone and Stet (1995) 100 XX-XYiClarias macrocephalus Na-Nakorn (1995) 100 XX-XYiClarias gariepinus Galbusera, Volckaert and Ollevier (2000) 100 XX-XYjMisgurnus anguillicaudatus Suzuki, Oshiro and Nakanishi (1985) 94.9 XX-XYm
kTakifugu rubripes See Arai (2001) 77.3–100 XX-XYm
lLimanda yokohamae See Arai (2001) 100 XX-XY
aFamily: bParalichthyidae, cMoronidae, dSoleidae, eCichlidae, fCyprinidae, gIctaluridae, hSalmonidae, iClariidae, jCobitidae,kTetraodontidae, lPleuronectidae. mUnexpected presence of males in species with female homogamety.
mossambicus (Pandian & Varadaraj, 1990), O. niloti-cus (Mair et al., 1991a), O. aureus (Mair et al., 1991b)and Puntius gonionotus (Pongthana et al., 1999)giving all-female progenies.
Proportion of sexes
In species with female homogamety (system XX-XY),the induction of gynogenesis should result in mono-sex female production (Table 9). This has been usedto assess female homogamety in fish, although it canalso be determined by integrating gynogenesis induc-tion with endocrine sex reversal (Hunter & Donaldson,1983; Ihssen et al., 1990; Piferrer, 2001). How-ever, one aspect that needs to be mentioned is that insome species in which the XX-XY scheme operates,a variable proportion of gynogenetic males has beenobserved in the progeny (Table 9). In the sea bass,experiments performed to determine sex ratios in off-spring of sex-reversed females (neomales) (Blázquezet al., 1999) as well as a study to identify the sex ofartificially-induced gynogens (Felip, 2000) have beenunable to conclusively assign any particular type ofsex determining system for this species. Furthermore,Blázquez et al. (1998) and Pavlidis et al. (2000) haveshown that the incubation temperature affects the sex
ratio. In species with male homogamety (system ZZ-ZW), gynogenesis induction renders progenies of bothsexes where ZZ is expected to give males and ZW orWW to give females. In this regard, the proportionof females depends on the viability of WW females(super-females) (Bull, 1983; Yamazaki, 1983). Theproduction of a second generation of gynogenetics us-ing potential viable WW females would result in anall-female progeny.
Prospects on triploid and gynogenetic fish
Besides their potential application in aquaculture, trip-loid and gynogenetic fish possess many traits thatrender them attractive for basic research. Triploid fishcan be used in studies dealing with the allocationof energy between growth and reproduction (i.e., therelationship between somatotropic and gonadotropicaxes) or the physiology and mechanism of fat storageand mobilization (McLean et al., 1991, 1993; Sump-ter et al., 1991; Mol et al., 1994). By virtue of theirsterility, triploids can be used as a negative control instudies examining the steroid activation of the brain-pituitary-gonad axis (Benfey et al., 1989; Breton &Sambroni, 1996, Benfey, 1999). In addition, triploids

190
can be used as models for cytogenetic studies focusingon meiotic chromosome behaviour (Gui et al., 1992;Carrasco et al., 1998) or in studying the relationshipbetween disruption of gonadal function and the genesinvolved in gametogenesis, as it occurs in transgenicanimals (Reventós & Munell, 1997).
On the other hand, gynogenetic fish can be anadvantage for genetic analysis and chromosome map-ping. Gynogenesis results in a very efficient methodfor rapid production of isogenic lines as well as inbredlines (Thorgaard, 1983; Yamazaki 1983; Ihssen et al.,1990). Thus, the production of homozygotes facilit-ates the determination of the genetic parameters of anindividual or population for development of geneticmaps and markers (Nagy & Csányi, 1982; Gervai &Csányi, 1984; Streisinger et al., 1986; Liu, Zhanzhou& Wang, 1996) and the estimation of heterozigosity(Allendorf & Leary, 1984). Recently, a detailed link-age map was reported in three teleost fish (Kocheret al., 1998; Young et al., 1998; Liu et al., 1999a,b; Agresti et al., 2000). Linkages between markerscan be associated with genes in order to carry outcomparative studies of genomic evolution and gen-ome conservation among fish as well as quantitativetrait locus (QTL) analysis (Danzmann, Jackson & Fer-guson, 1999; Sakamoto et al., 1999) or marker-basedselection in the commercial fish species (Ihssen et al.,1990; Young et al., 1998). Another application ofmeiotic gynogenetic diploids is their use to discrim-inate genetic and environmental effects on traits andphysiological aspects while mitotic gynogenetic dip-loids are useful to study single-locus effects (Ihssenet al., 1990). Finally, integration of chromosome setmanipulation can be useful in transgenic technologiesfor gene transfer (Peek et al., 1997).
Acknowledgements
We thank Drs. S. Abe and K. Arai for giving usthe opportunity to write this review, and Drs. M.Blázquez, L.A. Sorbera and two anonymous reviewersfor helpful comments of the manuscript.
References
Agresti, J.J., S. Seki, A. Cnaani, S. Poompuang, E. Hallerman, N.Umiel, G. Hulata, G.A.E. Gall & B. May, 2000. Breeding newstrains of tilapia: development of an artificial center of origin andlinkage map based on AFLP and microsatellite loci. Aquaculture185: 43–56.
Allen, S.K. Jr., 1983. Flow cytometry: assaying experimental po-lyploid fish and shellfish. Aquaculture 33: 317–328.
Allen, S.K. Jr. & J.G. Stanley, 1979. Polyploid mosaics induced bycytochalasin B in landlocked Atlantic salmon Salmo salar. Trans.Am. Fish. Soc. 108: 462–466.
Allendorf, F.W. & R.F. Leary, 1984. Heterozigosity in gynogeneticdiploids and triploids estimated by gene-centromere recombina-tion rates. Aquaculture 43: 413–420.
Alvariño, J.M.R., M. Carrillo, S. Zanuy, F. Prat & E. Mañanós,1992. Pattern of sea bass oocyte development after ovarianstimulation by LHRHa. J. Fish Biol. 41: 965–970.
Ankley, G.T., J.E. Tietge, G.W. Holcombe, D.L. DeFoe, S.A.Diamond, K.M. Jensen & S.J. Degitz, 2000. Effects of labor-atory ultraviolet radiation and natural sunlight on survival anddevelopment of Rana pipiens. Can. J. Zool. 78: 1092–1100.
Arai, K., 2001. Genetic improvement of aquaculture finfish speciesby chromosome manipulation techniques in Japan. Aquaculture197: 205–228.
Arakawa, T., M. Takaya, K. Inoue, I. Takami & K. Yamashita, 1987.An examination of the condition for triploid induction by coldshock in red and black sea breams. Bull. Nagasaki Pref. Inst.Fish. 13: 25–30.
Barbaro, A., P. Belvedere, N. Borgono, G. Bozzato, A. Francescon,A. Libertini, F. Meneghetti, A. Merlin & L. Colombo, 1996.Chromosome set manipulation in the gilthead seabream (Sparusaurata L.) and the European sea bass (Dicentrarchus labrax L.),pp. 227–230 in Sea Bass and Seabream Culture: Problems andProspects. European Aquaculture Society, Oostende, Belgium.
Bartley, D.M., 1997. Genetics and breeding in aquaculture: currentstatus and trends. Cahiers Opt. Méditerran. 34: 13–30.
Bass, E.L. & S.N. Sistrun, 1997. Effect of UV-A radiation on de-velopment and hatching success in Oryzias latipes, the Japanesemedaka. Bull. Environ. Contam. Toxicol. 59: 537–542.
Béland, F., H.I. Browman, C. Alonso Rodríguez & J.F. St-Pierre,1999. Effect of solar ultraviolet radiation (280–400 nm) on theeggs and larvae of Atlantic cod (Gadus morhua). Can. J. Fish.Aquat. Sci. 56: 1058–1067.
Benfey, T.J., 1989. A bibliography of triploid fish, 1943–1988. Can.Tech. Rep. Fish. Aq. Sci. 1682: 33.
Benfey, T.J., 1995. Ovarian development in triploid brook trout(Salvelinus fontinalis), p. 357. in Proceedings of the Fifth In-ternational Symposium on the Reproductive Physiology of Fish,edited by F. W. Goetz & P. Thomas. Austin, Texas, USA.
Benfey, T.J., 1999. The physiology and behavior of triploid fishes.Rev. Fish. Sci. 7: 39–67.
Benfey, T.J., H.M. Dye, I.I. Solar & E.M. Donaldson, 1989. Thegrowth and reproductive endocrinology of adult triploid pacificsalmonids. Fish Physiol. Biochem. 6: 113–120.
Benfey, T.J. & A.M. Sutterlin, 1984. Growth and gonadal develop-ment in triploid landlocked Atlantic salmon (Salmo salar). Can.J. Fish. Aquat. Sci. 41: 1387–1392.
Blanc, J.M., F. Vallée & M. Dorson, 2000. Survival, growth anddressing traits of triploid hybrids between rainbow trout and threecharr species. Aqua. Res. 31: 349–358.
Blázquez, M., M. Carrillo, S. Zanuy & F. Piferrer, 1999. Sex ratiosin offspring of sex-reversed sea bass and the relationship betweengrowth and phenotypic sex differentiation. J. Fish Biol. 55: 916–930.
Blázquez, M., S. Zanuy, M. Carrillo & F. Piferrer, 1998. Effectsof rearing temperature on sex differentiation in the European seabass (Dicentrarchus labrax L.). J. Exp. Zool. 281: 207–216.
Brämick, U., B. Puckhaber, H.J. Langholz & G. Hörstgen-Schwark,1995. Testing of triploid tilapia (Oreochromis niloticus) undertropical pond conditions. Aquaculture 137: 343–353.

191
Breton, B. & E. Sambroni, 1996. Steroid activation of the brain-pituitary complex gonadotropic function in the triploid rain-bow trout Oncorhynchus mykiss. Gen. Comp. Endocrinol. 101:155–164.
Bromage, N., 1988. Propagation and stock improvement, pp. 103–153 in Intensive Fish Farming, edited by J. Shepherd & N.Bromage. BSP Professional Books, Oxford.
Bull, J.J., 1983. Evolution of Sex Determining Mechanism. TheBenjamin/Cummings Publishing Company, Menlo Park, Ca,316 pp.
Carrasco, L.A., S. Doroshov, D.J. Penman & N. Bromage, 1998.Long term, quantitative analysis of gametogenesis in autotriploidrainbow trout, Oncorhynchus mykiss. J. Reprod. Fertility 113:197–210.
Carrillo, M., S. Zanuy, M. Blázquez, J. Ramos, F. Piferrer & E.M.Donaldson, 1993. Sex control and ploidy manipulation in seabass, p. 512. in International Conference of Aquaculture 1993,EAS Spec. Publ. 19.
Carrillo, M., S. Zanuy, M. Blázquez, J. Ramos, F. Piferrer & E.M.Donaldson, 1995b. Sex control and ploidy manipulation in seabass, pp. 125–143 in OECD Documents, Environmental Impactsof Aquatic Biotechnology. OECD, Paris.
Carrillo, M., S. Zanuy, F. Oyen, J. Cerdá, J.M. Navas & J. Ramos,2000. Some criteria of the quality of the progeny as indicators ofphysiological broodstock fitness. Cahiers Opt. Méditerran. 47:61–73.
Carrillo, M., S. Zanuy, F. Piferrer, M. Blázquez, A. Felip, G.Martínez-Rodríguez & J. Ramos, 1999. Sex control in sea bass(Dicentrarchus labrax L.): present state and future prospects, pp.22–32 in Proceedings of International Symposium on Progressand Prospect of Marine Biotechnology (ISPPMB 1998), editedby H.S. Xu & R.R. Colwell. China Ocean Press.
Carrillo, M., S. Zanuy, F. Prat, J. Cerdá, J. Ramos, E. Mañanós& N. Bromage, 1995a. Sea bass (Dicentrarchus labrax), pp.138–168 in Broodstock Managment and Egg and Larval Qual-ity, edited by N.R. Bromage & R.J. Roberts. Blackwell Science,Oxford.
Carter, R.E., G.C. Mair, D.O.F. Skibinski, D.T. Parkin & J.A.Beardmore, 1991. The application of DNA fingerprinting in theanalysis of gynogenesis in tilapia. Aquaculture 95: 41–52.
Carter, C.G., I.D. McCarthy, D.F. Houlihan, R. Johnstone, M.V.Walsingham & A.I. Mitchell, 1994. Food consumption, feedingbehaviour, and growth of triploid and diploid Atlantic salmon,Salmo salar L., parr. Can. J. Zool. 72: 609–617.
Cassani, J.R. & J.R. Caton, 1985. Induced triploidy in grass carp,Ctenopharyngodon idella Val. Aquaculture 46: 37–44.
Cerdá, J., M. Carrillo, S. Zanuy, J. Ramos & M. De la Higuera,1994. Influence of nutritional composition of diet on sea bassDicentrarchus labrax L., reproductive performance and egg andlarvae quality. Aquaculture 128: 345–361.
Cherfas, N.B., B. Gomelsky, N. Ben-Dom, Y. Peretz & G. Hulata,1994. Assessment of triploid common carp (Cyprinus carpio L.)for culture. Aquaculture 127: 11–18.
Chourrout, D., 1987. Genetic manipulations in fish: review ofmethods, pp. 111–126 in Proc. World Symp. On Selection, Hy-bridization, and Genetic Engineering in Aquaculture, Bordeaux,27–30 May, 1986, Vol. II, Berlin.
Chourrout, D., B. Chevassus, F. Krieg, A. Happe, G. Burger &P. Renard, 1986. Production of second generation triploid andtetraploid rainbow trout by mating tetraploid males and dip-loid females-potential of tetraploid fish. Theor. Appl. Genet. 72:193–206.
Colihueque, N., P. Iturra, N.F. Díaz & A. Veloso, 1996. Furtherevidence of chromosome abnormalities in normal and haploid
gynogenetic progenies of rainbow trout, Oncorhynchus mykiss.J. Exp. Zool. 276: 70–75.
Colombo, L., A. Barbaro, A. Libertini, P. Benedetti, A. Francescon& I. Lombardo, 1995. Artificial fertilization and inductionof triploidy and meiogynogenesis in the European sea bass,Dicentrarchus labrax L. J. Appl. Ichthyol. 11: 118–125.
Colombo, L., A. Barbaro, A. Francescon, A. Libertini, P. Bene-detti, L. Dalla Valle, M. Pazzaglia, L. Pugi, F. Argenton, M.Bortolussi & P. Belvedere, 1996. Potential gains through geneticimprovements: chromosome set manipulation and hybridization,pp. 343–462 in Sea Bass and Seabream Culture: Problems andProspects. European Aquaculture Society, Oostende, Belgium.
Colombo, L., A. Barbaro, A. Francescon, A. Libertini, M. Borto-lussi, F. Argenton, L. Dalla Valle, S. Vianello & P. Belvedere,1997. Towards an integration between chromosome set manipu-lation, intergeneric hybridization and gene transfer in marine fishculture. Cahiers Opt. Méditerran. 34: 77–122.
Curatolo, A., F. Gunnella, A. Santulli, N.P. Wilkins & V. D’Amelio,1996. Chromosome set manipulation in the European sea bass(Dicentrarchus labrax L.): preliminary results, pp. 262–265in Sea Bass and Seabream Culture: Problems and Prospects.European Aquaculture Society, Oostende, Belgium.
Danzmann, R.G., T.R. Jackson & M.M. Ferguson, 1999. Epistasis inallelic expresion at upper temperature tolerance QTL in rainbowtrout. Aquaculture 173: 45–58.
Díaz, N.F., P. Iturra, A. Veloso, F. Estay & N. Colihueque, 1993.Physiological factors affecting triploid production in rainbowtrout, Oncorhynchus mykiss. Aquaculture 114: 33–40.
Dillon, J.C., D.J. Schill & D.M. Teuscher, 2000. Relative return tocreel of triploid and diploid rainbow trout stocked in 18 Idahostreams. North Am. J. Fish. Manag. 20: 1–9.
Don, J. & R.R. Avtalion, 1988. Production of F1 and F2 diploidgynogenetic tilapias and analysis of the ‘Hertwig curve’ ob-tained using ultraviolet irradiated sperm. Theor. Appl. Genet. 76:253–259.
Donaldson, E.M., 1996. Manipulation of reproduction in farmedfish. Anim. Reprod. Sci. 42: 381–392.
Donaldson, E.M., 1997. The role of biotechnology in sustainableaquaculture, pp. 101–126 in Sustainable Aquaculture, edited byJ.E. Bardach. Wiley.
Ewing, R.R. & C. Scalet, 1991. Flow cytometric identification oflarval triploid walleyes. Prog. Fish-Cult. 53: 177–180.
Fast, A.W., T. Pewnim, R. Keawtabtim, R. Saijit, F.T. Te & R. Vejar-atpimol, 1995. Comparative growth of diploid and triploid Asiancatfish Clarias macrocephalus in Thailand. J. World Aquacult.Soc. 26: 390–395.
Fauvel, C., O. Savoye, C. Dreanno, J. Cosson & M. Suquet, 1999.Characteristics of sperm of captive seabass in relation to itsfertilization potential. J. Fish Biol., 54: 356–369.
Felip, A., 2000. Inducción de la triploidía y la ginogénesis en la lu-bina (Dicentrarchus labrax L.): Efectos sobre el crecimiento y lafisiología reproductiva. Doctoral Thesis, University of Valencia,263 pp.
Felip, A., G. Martínez-Rodríguez, F. Piferrer, M. Carrillo & S.Zanuy, 2000. AFLP analysis confirms exclusive maternal gen-omic contribution of meiogynogenetic sea bass (Dicentrarchuslabrax L.). Mar. Biotech. 2: 301–306.
Felip, A., F. Piferrer, M. Carrillo & S. Zanuy, 1999a. The rela-tionship between the effects of UV light and thermal shock ongametes and the viability of early developmental stages in a mar-ine teleost fish, the sea bass (Dicentrarchus labrax L.). Heredity83: 387–397.
Felip, A., F. Piferrer, M. Carrillo & S. Zanuy, 2001a. Compara-tive growth performance between diploid and triploid sea bass

192
(Dicentrarchus labrax L.) over the first four spawning seasons.J. Fish. Biol. 58: 76–88.
Felip, A., F. Piferrer, M. Carrillo & S. Zanuy, 2001b. A comparisonof the gonadal development and plasma levels of sex steroid hor-mones in diploid and triploid sea bass, Dicentrarchus labrax L.J. Exp. Zool. 290: 384–395.
Felip, A., S. Zanuy, M. Carrillo, G. Martínez, J. Ramos & F. Piferrer,1997. Optimal conditions for the induction of triploidy in the seabass (Dicentrarchus labrax L.). Aquaculture 152: 287–298.
Felip, A., S. Zanuy, M. Carrillo & F. Piferrer, 1999b. Growth andgonadal development in triploid sea bass (Dicentrarchus lab-rax L.) during the first 2 years of age. Aquaculture 173: 389–399.
Flajshans, M., P. Ráb & S. Dobosz, 1992. Frequency analyses of act-ive NORs in nuclei of artificially induced triploid fishes. Theor.Appl. Genet. 85: 68–72.
Freund, F., G. Hörstgen-Schwark & W. Holtz, 1995. Plasma ster-oid hormones in adult triploid tilapia (Oreochromis niloticus), p.35 in Proc. 5th Intern. Symp. Reproductive Physiology of Fish,edited by F. W. Goetz & P. Thomas. Austin, Texas, USA.
Friars, G.W., I. McMillan, V.M. Quinton, F.M. O’Flynn, S.A.McGeachy & T.J. Benfey, 2001. Family differences in relativegrowth of diploid and triploid Atlantic salmon (Salmo salar L.).Aquacultutre 192: 23–29.
Fujioka, Y., 1998. Survival, growth and sex ratios of gynogeneticdiploid honmoroko. J. Fish Biol. 52: 430–442.
Galbreath, P.F., W.St. Jean, V. Anderson & G.H. Thorgaard,1994. Freshwater performance of all-female diploid and triploidAtlantic salmon. Aquaculture 128: 41–49.
Galbreath P.F. & G.H. Thorgaard, 1994. Viability and freshwaterperformance of Atlantic salmon (Salmo salar) x brown trout(Salmo trutta) triploid hybrids. Can. J. Fish. Aquat. Sci. 51 (1):16–24.
Galbreath P.F. & G.H. Thorgaard, 1995. Saltwater performance ofall- female triploid Atlantic salmon. Aquaculture 138: 77–85.
Galbusera, P., A.M. Volckaert & F. Ollevier, 2000. Gynogenesisin the African catfish Clarias gariepinus (Burchell, 1822). III.Induction of endomitosis and the presence of residual geneticvariation. Aquaculture 185: 25–42.
Garrido-Ramos, M., R. De la Herrán, R. Lozano, S. Cárdenas, C.Ruiz Rejón & M. Ruiz Rejón, 1996. Induction of triploidy inoffspring of gilthead seabream (Sparus aurata) by mean of heatshock. J. Appl. Ichthyol. 12: 53–55.
Gervai, J.S., A. Péter, L. Nagy, L. Horváth & V. Csányi, 1980.Induced triploidy in carp, Cyprinus carpio L. J. Fish Biol. 17:667–671.
Gervai, J. & V. Csányi, 1984. Artificial gynogenesis and mappingof gene-centromere distances in the paradise fish, Macropodusopercularis. Theor. Appl. Genet. 68: 481–485.
Gomelsky, B., N.B. Cherfas, A. Gissis & G. Hulata, 1998. In-duced diploid gynogenesis in white bass. Prog. Fish Cult. 60:288–292.
Gorshkov, S., G. Gorshkova, A. Hadani, H. Gordin & W. Knibb,1998. Chromosome set manipulations and hybridization ex-periments in gilthead seabream (Sparus aurata). Isr. J. Aqua.Bamidgeh 50 (3): 99–110.
Gorshkova, G., S. Gorshkov, H. Gordin & W. Knibb, 1996.Sex control and gynogenetic production in European sea bass,Dicentrarchus labrax, pp. 288–290 in Seabass and SeabreamCulture: Problem and Prospects. European Aquaculture Society,Oostende, Belgium.
Gorshkova, G., S. Gorshkov, A. Hadani, H. Gordin & W. Knibb,1995. Chromosome set manipulation in marine fish. Aquaculture137: 157–158.
Goudie, C.A., B.A. Simco, K.B. Davis & Q. Liu, 1995. Pro-duction of gynogenetic and polyploid catfish by pressure-induced chromosome set manipulation. Aquaculture, 133: 185–198.
Gui, J.F., J. Jia, S.C. Liang & Y.G. Jiang, 1992. Meiotic chro-mosome behaviour in male triploid transparent coloured cruciancarp, Carassius auratus L. J. Fish Biol. 41: 317–326.
Guo, X., W.K. Hershberger & J.M. Myers, 1990. Growth and sur-vival of intrastrain and interstrain rainbow trout (Oncorhynchusmykiss) triploids. J. World Aqua. Soc. 21: 250–256.
Häder, D.P., 1986. The effect of enhanced solar UV-B radiationon motile microorganisms, pp. 223-233 in Stratospheric OzoneReduction, Solar Ultraviolet Radiation and Plant Life, Vol. G8,edited by R.C. Worrest & M.M. Caldwell. NATO ASI Series,Springer, Berlin.
Häder, D.P., 1993. Risks of enhanced solar ultraviolet radiation foraquatic ecosystems, pp. 1-45 in Progress in Phycological Re-search, Vol. 9, edited by F.E. Round & D.J. Chapman. BiopressBristol.
Harrell, R.M., W. Van Heukelem & J.H. Kerby, 1998. A comparisonof triploid induction validation techniques. Prog. Fish Cult. 60:221–226.
Henken, A.M., A.M. Brunink & C.J.J. Richter, 1987. Differencesin growth rate and feed utilization between diploid and triploidAfrican catfish, Clarias gariepinus (Burchell 1822). Aquaculture63: 233–242.
Holmefjord, I. & T. Refstie, 1997. Induction of triploidy in Atlantichalibut by temperature shocks. Aqua. Intern. 5: 169–173.
Howell, B.R., S.M. Baynes & D. Thompson, 1995. Progress to-wards the identification of the sex-determining mechanism of thesole, Solea solea (L.), by the induction of diploid gynogenesis.Aqua. Res. 26: 135–140.
Hunter, G. A. & E.M. Donaldson, 1983. Hormonal sex control andits application to fish culture, pp. 223–275 in Fish Physiology,Vol. IXB, edited by W.S. Hoar, D.J. Randall & E.M. Donaldson.Academic Press, N.Y.
Hussain, M.G., G.P.S. Rao, N.M. Humayun, C.F. Randall, D.J.Penman, D. Kime, N.R. Bromage, J.M. Myers & B.J. Mc-Andrew, 1995. Comparative performance of growth, biochem-ical composition and endocrine profiles in diploid and triploidtilapia Oreochromis niloticus L. Aquaculture 138: 87–97.
Ihssen, P.E., L.R. McKay, I. McMillan & R.B. Phillips, 1990. Ploidymanipulation and gynogenesis in fishes: cytogenetic and fisheriesapplications. Trans. Am. Fish. Soc. 119: 698–717.
Ijiri, K.I. & N. Egami, 1980. Hertwig effect caused by UV-irradiation of sperm of Oryzias latipes (teleost) and its photore-activation. Mutat. Res. 69: 241–248.
Johnson, O.W., W.W. Dickhoff & F.M. Utter, 1986. Comparativegrowth and development of diploid and triploid coho salmon,Oncorhynchus kisutch. Aquaculture 57: 329–336.
Johnstone, R. & R.J.M. Stet, 1995. The prodution of gynogeneticAtlantic salmon, Salmo salar L. Theor. Appl. Genet. 90: 819–826.
Kavumpurath, S. & T.J. Pandian, 1990. Induction of triploidy in thezebrafish, Brachydanio rerio (Hamilton). Aquacult. Fish. Manag.21: 299–306.
Kavumpurath, S. & T.J. Pandian, 1992. The development of all-male sterile triploid fighting fish (Betta splendens regan)by integrating hormonal sex reversal of broodstock andchromosome-set manipulation. Isr. J. Aquacult. Bamidgeh 44(4): 111–119.
Kawamura, K., T. Ueda, K. Aoki & K. Hosoya, 1999. Spermatozoain triploids of the rosy bitterling Rhodeus ocellatus ocellatus. J.Fish Biol. 55: 420–432.

193
Kim, D.S., J.Y. Jo & T.Y. Lee, 1994. Induction of triploidy inmud loach (Misgurnus mizolepis) and its effect on gonadaldevelopment and growth. Aquaculture 120: 263–270.
Kitamura, H., O.Y. Teong & T. Arakawa, 1991. Gonadal develop-ment of artificially induced triploid red sea bream Pagrus major.Nippon Suisan Gakkaishi 57: 1657–1660.
Knibb, W., 2000. Genetic improvement of marine fish – whichmethod for industry?. Aquacult. Res. 31: 11–23.
Kocher, T.D., W.J. Lee, H. Sobolewska, D. Penman & B.McAndrew 1998. A genetic linkage map of a cichlid fish, thetilapia (Oreochromis niloticus). Genetics 148: 1225–1232.
Koedprang, W. & U. Na-Nakorn, 2000. Preliminary study onperformance of triploid Thai silver barb, Puntius gonionotus.Aquaculture 190: 211–221.
Komen, J., G.F. Wiegertjes, V.J.T. Van Ginneken, E.H. Eding &C.J.J. Ritcher, 1992. The effects of heterozygous and homo-zygous gynogenetic inbreeding on sex, gonad development andfertility in common carp (Cyprinus carpio L.). Aquaculture 104:51–66.
Krisfalusi, M. & J.G. Cloud, 1997. Gonadal development in triploidrainbow trout (Oncorhynchus mykiss): testicular but not ovariandevelopment. Biol. Reprod. 56: 105.
Kucharczyk, D., P. Woznicki, M.J. Luczynski, M. Klinger & M.Luczynski, 1999. Ploidy level determination in genetically ma-nipulated northern pike based on the number of active nucleoliper cell. North Am. J. Aquacult. 61: 38–42.
Leary, R.F., F.W. Allendorf, K.L. Knudsen & G.H. Thorgaard,1985. Heterozygosity and developmental stability in gynogeneticdiploid and triploid rainbow trout. Heredity 54: 219–225.
Leclerc, G.M., B. Ely, X.L. Xu & R.M. Harrell, 1996. Gynogenproduction in hybrid striped bass. J. World Aqua. Soc. 27 (1):119–125.
Lecommandeur, D., P. Haffray & L. Philippe, 1994. Rapid flowcytometry method for ploidy determination in salmonid eggs.Aquacult. Fish. Manag. 25: 345–350.
Levanduski, M.J., J.C. Beck & J.E. Seeb, 1990. Optimal thermalshocks for induced diploid gynogenesis in chinook salmon (On-corhynchus tshawytscha). Aquaculture 90: 239–250.
Lin, F. & K. Dabrowski, 1996. Effects of sperm irradiation andheat shock on induction of gynogenesis in muskellunge (Esoxmasquinongy). Can. J. Fish. Aquat. Sci. 53: 2067–2075.
Lincoln, R.F., 1981. The growth of female diploid and triploid plaice(Pleuronectes platessa) and plaice x flounder (Platichthys flesus)hybrids over one spawning season. Aquaculture 25: 259–268.
Lincoln, R.F., D. Aulstad & A. Grammeltvedt, 1974. Attemptedtriploid induction in Atlantic salmon (Salmo salar) using coldshocks. Aquaculture 4: 287–297.
Liu, Z.J., P. Li, B.J. Argue & R.A. Dunham, 1999a. Random ampli-fied polymorphic DNA markers: usefulness for gene mappingand analysis of genetic variation of catfish. Aquaculture 174:59–68.
Liu, Z., P. Li, H. Kucuktas, A. Nichols, G. Tan, X. Zheng, B.J.Argue & R.A. Dunham, 1999b. Development of amplified frag-ment length polymorphism (AFLP) markers suitable for geneticlinkage mapping of catfish. Trans. Am. Fish. Soc. 128: 317–327.
Liu, S., Y. Zhanzhou & Y. Wang, 1996. Sex hormone induction ofsex reversal in the teleost Clarias lazera and evidence for femalehomogamety and male heterogamety. J. Exp. Zool. 276: 432–438.
Magoulas, A., 1997. Application of molecular makers to aquacul-ture and broodstock management with special emphasis onmicrosatellite DNA. Cahiers Opt. Méditerran. 34: 153–168.
Mair, G.C., 1993. Chromosome-set manipulation in tilapia-techni-ques, problems and prospects. Aquaculture 111: 227–244.
Mair, G.C., J.S. Abucay, D.O.F. Skibinski, T.A. Abella & J.A.Beardmore, 1997. Genetic manipulation of sex ratio for the large-scale production of all-male tilapia, Oreochromis niloticus. Can.J. Fish. Aquat. Sci. 54: 396–404.
Mair, G.C., A.G. Scott, D.J. Penman, J.A. Beardmore & D.O.F.Skibinski, 1991a. Sex determination in the genus Oreochromis.1. Sex reversal, gynogenesis and triploidy in O. niloticus (L.).Theor. Appl. Genet. 82: 144–152.
Mair, G.C., A.G. Scott, D.J. Penman, D.O.F. Skibinski & J.A.Beardmore, 1991b. Sex determination in the genus Oreochromis.2. Sex reversal, hybridisation, gynogenesis and triploidy in O.aureus Steindachner. Theor. Appl. Genet. 82: 153–160.
Malison, J.A. & M.A.R. García-Abiado, 1996. Sex control andploidy manipulation in yellow perch (Perca flavescens) andwalleye (Stizostedium vitreum). J. Appl. Ichthyol. 12: 189–194.
Malison, J.A., L.S. Procarione, J.A. Held, T.B. Kayes & C.H.Amundson, 1993. The influence of triploidy and heat and hy-drostatic pressure shocks on the growth and reproductive devel-opment of juvenile yellow perch (Perca flavescens). Aquaculture116: 121–133.
McGeachy, S.A., T.J. Benfey & G.W. Friars, 1995. Freshwaterperformance of triploid Atlantic salmon (Salmo salar) in NewBrunswick aquaculture. Aquaculture 137: 333–341.
McLean, E., E.M. Donaldson, E. Teskeredzic & L.M. Souza, 1993.Growth enhancement following dietary delivery of recombin-ant porcine somatotropin to diploid and triploid coho salmon(Oncorhynchus kisutch). Fish Physiol. Biochem. 11: 363–369.
McLean, E., M.D. Sadar, R.H. Devlin, L.M. Souza & E.M.Donaldson, 1991. Promotion of growth in diploid and triploidcoho salmon with parenteral delivery of a recombinant porcinesomatotropin. Aquat. Liv. Res. 4: 155–160.
Miller, G.D., J.E. Seeb, B.G. Bue & S. Sharr, 1994. Saltwaterexposure at fertilization induces ploidy alterations, includingmosaicism, in salmonids. Can. J. Fish. Aquat. Sci. 51 (1): 42–49.
Mol, K., N. Byamungu, B. Cuisset, Z. Yaron, M. Ofir, Ch. Mélard,M. Castelli & E.R. Kühn, 1994. Hormonal profile of grow-ing male and female diploids and triploids of the blue tilapia,Oreochromis aureus, reared in intensive culture. Fish Physiol.Biochem. 13: 209–218.
Murata, O., 1998. Studies on the breeding of cultivated marinefishes. Bull. Fish. Lab. Kinki Univ. 6: 1–101 (in Japaness withabstract in English).
Myers, J.M. & W.K. Hershberger, 1991. Early growth and survivalof heat-shocked and tetraploid-derived triploid rainbow trout(Oncorhynchus mykiss). Aquaculture 96: 97–107.
Nagy, A. & V. Csányi, 1982. Changes of genetic parameters in suc-cessive gynogenetic generations and some calculations for carpgynogenesis. Theor. Appl. Genet. 63: 105–110.
Nagy, A., K. Rajki, L. Horváth & V. Csányi, 1978. Investigation oncarp, Cyprinus carpio L. gynogenesis. J. Fish Biol. 13: 215–224.
Nakamura, M., Y. Nagahama, M. Iwahashi & M. Kojima, 1987.Ovarian structure and plasma steroid hormones of triploid femalerainbow trout. Nippon Suisan Gakkaishi 53: 1105.
Nakamura, M., F. Tsuchiya, M. Iwahashi & Y. Nagahama, 1993.Reproductive characteristics of precociously mature triploidmale masu salmon, Oncorhynchus masou. Zool. Sci. 10: 117–125.
Na-Nakorn, U., 1995. Comparison of cold and heat shocks to inducediploid gynogenesis in thai walking catfish (Clarias macroceph-alus) and performances of gynogens. Aquat. Living Resour. 8:333–341.
O’Keefe, R.A. & T.J. Benfey, 1997. The feeding response of diploidand triploid Atlantic salmon and brook trout. J. Fish Biol. 51:989–997.

194
Oliva-Teles, A. & S.J. Kaushik, 1990. Growth and nutrient utiliza-tion by 0+ and 1+ triploid rainbow trout, Oncorhynchus mykiss.J. Fish Biol. 37: 125–133.
Oshiro, T., 1987. Sex ratios of diploid gynogenetic progeny derivedfrom five different females of goldfish. Nippon Suisan Gakkaishi53: 10.
Palti, Y., J.J. Li & G.H. Thorgaard, 1997. Improved efficiency ofheat and pressure shocks for producing gynogenetic rainbowtrout. Prog. Fish Cult. 59 (1): 1–13.
Pandian, T.J. & Varadaraj, K., 1990. Development of monosex fe-male Oreochromis mossambicus broodstock by integrating gyno-genetic technique with endocrine sex reversal. J. Exp. Zool. 255:88–96.
Parsons, J.E., R.A. Busch, G.H. Thorgaard & P.D. Scheerer, 1986.Increased resitence of triploid rainbow trout x coho salmon hy-brids to infectious hematopoietic necrosis virus. Aquaculture 57:337–344.
Parsons, G.R. & K. Meals, 1997. Comparison of triploid hybridcrappie and diploid white crappie in experimental ponds. NorthAm. J. Fish. Manag. 17: 803–806.
Partis, L. & R.J. Wells, 1996. Identification of fish species usingrandom amplified polymorphic DNA (RAPD). Mol. Cell Probes10: 435–441.
Pavlidis, M., G. Koumoundouros, A. Sterioti, S. Somarakis,P. Divanach & M. Kentouri, 2000. Evidence of temperature-dependent sex determination in the European sea bass(Dicentrarchus labrax L.). J. Exp. Zool. 287: 225–232.
Peek, A.S., P.A. Wheeler, C.O. Ostberg & G.H. Thorgaard, 1997.A minichromosome carrying a pigmentation gene and brooktrout DNA sequences in transgenic rainbow trout. Genome 40:594–599.
Penman, D.J., D.O.F. Skibinski & J.A. Beardmore, 1987. Survival,growth rate and maturity in triploid tilapia, pp. 277–288 in Se-lection, Hybridization and Genetic Engineering in Aquaculture,edited by K. Tiews. Heenemann Verlags, Berlin.
Peruzzi, S. & B. Chatain, 2000. Pressure and cold shock induc-tion of meiotic gynogenesis and triploidy in the European seabass, Dicentrarchus labrax L.: relative efficiency of methods andparental variability. Aquaculture 189: 23–37.
Piferrer, F., 2001. Endocrine sex control strategies for the feminiza-tion of teleost fish. Aquaculture 197: 229–281.
Piferrer, F., T.J. Benfey & E.M. Donaldson, 1994. Gonadal mor-phology of sex reversed triploid and gynogenetic diploid cohosalmon (Oncorhynchus kisutch). J. Fish Biol. 45: 541–553.
Piferrer, F., R.M. Cal, B. Álvarez-Blázquez, L. Sánchez & P.Martínez, 2000. Induction of triploidy in the turbot (Scophthal-mus maximus). I. Ploidy determination and the effects of coldshocks. Aquaculture 188: 79–90.
Piferrer, F., A. Felip & M. Blázquez, 1995. Control genético yfisiológico de las proporciones de sexos de los teleósteos y suaplicación en acuicultura, pp. 75–109 in Aulas del Mar, Acuicul-tura, Biología Marina, edited by S. Zamora, B. Agulleiro & P.García. Murcia, Spain.
Piferrer, F. & L.C. Lim, 1997. Application of sex reversal tech-nology in ornamental fish culture. Aqua. Sci. Conserv. 1: 113–118.
Planas, M. & I. Cunha, 1999. Larviculture of marine fish: problemsand perspectives. Aquaculture 177: 171–190.
Pongthana, N., D.J. Penman, P. Baoprasertkul, M.G. Hussain,M.S. Islam, S.F. Powell & B.J. McAndrew, 1999. Monosex fe-male production in the silver barb (Puntius gonionotus Bleeker).Aquaculture 173: 247–256.
Pongthana, N., D.J. Penman, J. Karnasuta & B.J. McAndrew, 1995.Induced gynogenesis in the silver barb (Puntius gonionotus
Bleeker) and evidence for female homogamety. Aquaculture 135:267–276.
Price, D.J., 1984. Genetics of sex determination in fishes – abrief review, pp. 77–89 in Fish Reproduction: Strategies andTactis, edited by G.W. Potts & R.J. Wooton. Academic Press,London.
Purdom, C.E., 1972. Induced polyploidy in plaice (Pleuronectesplatessa) and its hybrid with the flounder (Platichthys flesus).Heredity 29: 11–24.
Refstie, T., J. Stoss & E.M. Donaldson, 1982. Production of allfemale coho salmon (Oncorhynchus kisutch) by diploid gyno-genesis using irradiated sperm and cold shock. Aquaculture 29:67–82.
Reventós, J. & F. Munell, 1997. Transgenic animal models in repro-ductive endocrine research. Euro. J. Endocrinol. 136: 566–580.
Rosenmann, A., J. Wahrman, C. Richler, R. Voss, A. Persitz & B.Goldman, 1985. Meiotic association between the XY chromo-somes and unpaired autosomal elements as a cause of humanmale sterility. Cytogenet. Cell Genet. 39: 19–29.
Sakamoto, T., R.G. Danzmann, N. Okamoto, M.M. Ferguson & P.E.Ihssen, 1999. Linkage analysis of quantitative trait loci associ-ated with spawning time rainbow trout (Oncorhynchus mykiss).Aquaculture 173: 33–43.
Sheehan, R.J., S.P. Shasteen, A.V. Suresh, A.R. Kapuscinski &J. Seeb, 1999. Better growth in all-female diploid and triploidrainbow trout. Trans. Am. Fish. Soc. 128: 491–498.
Shelton, W.L., S.D. Mims, J.A. Clark, A.E. Hiott & Ch. Wang,1997. A temperature-dependent index of mitotic interval (τo)for chromosome manipulation in paddlefish and shovelnosesturgeon. Prog. Fish Cult. 59: 229–234.
Smith, L.T. & H.L. Lemoine, 1979. Colchicine-induced polyploidyin brook trout. Prog. Fish Cult. 41: 86–88.
Smoker, W.W., P.A. Crandell & M. Matsuoka, 1995. Second po-lar body retention and gynogenesis induced by thermal shock inpink salmon, Oncorhynchus gorbuscha (Walbaum). Aqua. Res.26: 213–219.
Solar, I.I., E.M. Donaldson & D. Douville, 1991. A bibliographyof gynogenesis and androgenesis in fish (1913–1989). Can.Tech.Rep. Fish. Aq. Sci. No 1788, 41 pp.
Solar, I.I., W.E. Hajen & E.M. Donaldson, 1992. A bibliography oftetraploidy in fish (1964–1991). Can.Tech. Rep. Fish. Aq. Sci.No 1901, 22 pp.
Sorbera, L., C.C. Mylonas, S. Zanuy, M. Carrillo & Y. Zohar,1996. Sustained administration of GnRHa increases milt volumewithout altering sperm counts in the sea bass. J. Exp. Zool. 276:361–368.
Stanley, J.G., 1976. Female homogamety in grass carp (Ctenopha-ryngodon idella) determined by gynogenesis. J. Fish. Res. BoardCan. 33: 1372–1374.
Stanley, J.G. & J.B. Jones, 1976. Morphology of androgeneticand gynogenetic grass carp, Ctenopharyngodon idella (Valen-ciennes). J. Fish Biol. 9: 523–528.
Streisinger, G., F. Singer, C. Walker, D. Knauber & N. Dower, 1986.Segregation analyses and gene-centromere distances in zebrafish.Genetics 112: 311–319.
Sugama, K., N. Taniguchi, S. Seki, H. Nabeshima & Y. Hasegawa,1990. Gynogenetic diploid production in the red sea bream usingUV-irradiated sperm of black sea bream and heat shock. NipponSuisan Gakkaishi 56: 1427–1433.
Sugama, K., N. Taniguchi, S. Seki & H. Nabeshima, 1992. Sur-vival, growth and gonadal development of triploid red sea bream,Pagrus major (Temminck Schlegel): use of allozyme markersfor ploidy and family identification. Aquat. Fish. Manag. 23:149–159.

195
Sumpter, J.P., R.F. Lincoln, V.J. Bye, J.F. Carragher & P.Y. Le Bail,1991. Plasma growth hormone levels during sexual maturation indiploid and triploid rainbow trout (Oncorhynchus mykiss). Gen.Comp. Endocrinol. 83: 103–110.
Suquet, M., R. Billard, J. Cosson, G. Dorange, L. Chauvaud, C.Mugnier & C. Fauvel, 1994. Sperm features in turbot (Scophthal-mus maximus): a comparison with other freshwater and marinefish species. Aquat. Living Resour. 7: 283–294.
Suzuki, R., T. Oshiro & T. Nakanishi, 1985. Survival, growth andfertility of gynogenetic diploids induced in the cyprinid loach,Misgurnus anguillicaudatus. Aquaculture 48: 45–55.
Tabata, K., 1991. Induction of gynogenetic diploid males andpresumption of sex determination mechanisms in the hirameParalichthys olivaceus. Nippon Suisan Gakkaishi 57 (5): 845–850.
Takagi, M. & N. Taniguchi, 1995. Random amplified polymorphicDNA (RAPD) for identification of three species of Anguilla, A.japonica, A. australis and A. bicolor. Fish. Sci. 61 (5): 884–885.
Teplitz, R.L., J.E. Joyce, S.I. Doroshov & B.H. Min, 1994. A pre-liminary ploidy analysis of diploid and triploid salmonids. Can.J. Fish. Aquat. Sci. 51 (1): 38–41.
Thorgaard, G.H., 1983. Chromosome set manipulation and sex con-trol in fish, pp. 405-434 in Fish Physiology, Vol. IXB, edited byW.H. Hoar, D.J. Randall & E.M. Donaldson. Academic Press,N.Y.
Thorgaard, G.H., 1995. Biotechnological approaches to broodstockmanagement, pp. 76-93 in Broodstock Managment and Eggand Larval Quality, edited by N.R. Bromage & R.J. Roberts.Blackwell Science, Oxford.
Thorgaard, G.H. & S.K. Allen, Jr., 1987. Chromosome manip-ulation and markers in fishery management, pp 319–336 inPopulation Genetics and Fishery Management, edited by N.Ryman & F.M. Utter. University of Seattle, Seattle.
Thorgaard, G.H. & J.E. Disney, 1990. Chromosome preparation andanalysis, in Methods for Fish Biology, edited by C.B. Schreck &P.B. Moyle. Bethesda, Maryland, USA. Am. Fish. Soc. 6: 171–190.
Thorgaard, G.H., P.S. Rabinovitch, M.W. Shen, G.A.E. Gall, J.Propp, & F.M. Utter, 1982. Triploid rainbow trout identified byflow cytometry. Aquaculture 29: 305–309.
Tiwary, B.K., R. Kirubbagaran & A.K. Ray, 2000. Gonadal de-velopment in triploid Heteropneustes fossilis. J. Fish Biol. 57:1343–1348.
Tudor, M. & Katavic, 1999. Body abnormalities in relation to sizeand sex of laboratory reared European sea bass Dicentrarchuslabrax L. Acta Adriat. 40: 67–74.
Utter, F.M., O.W. Johnson, G.H. Thorgaard & P.S. Rabinovitch,1983. Measurement and potential applications of induced trip-loidy in Pacific salmon. Aquaculture 35: 125–135.
Valenti, R.J., 1975. Induced polyploidy in Tilapia aurea Stein-dachner by means of temperature shock treatment. J. Fish Biol.7: 519–528.
Van Eenennaam, J.P., R.K. Stocker, R.G. Thiery, N.T. Hagstrom& S.I. Doroshov, 1990. Egg fertility, early developmentand survival from crosses of diploid female x triploid male
grass carp (Ctenopharyngodon idella). Aquaculture 86: 111–125.
Van Eenennaam, A.L., J.P. Van Eenennaam, J.F. Medrano &S.I. Doroshov, 1996. Rapid verification of meiotic gynogen-esis and polyploidy in white sturgeon (Acipenser transmontanusRichardson). Aquaculture 147: 177–189.
Váradi, L., I. Benkó, J. Varga & L. Horváth, 1999. Induction ofdiploid gynogenesis using interspecific sperm and productionof tetraploids in African catfish, Clarias gariepinus Burchell(1822). Aquaculture 173: 401–411.
Várkonyi, E., M. Bercsényi, C. Ozouf-Costaz & R. Billard, 1998.Chromosomal and morphological abnormalities caused by oo-cyte ageing in Silurus glanis. J. Fish Biol. 52: 899–906.
Volckaert, F.A.M., P.H.A.A. Galbusera, B.A.S. Hellemans, C. Vanden Haute, D. Vanstaen & F. Ollevier, 1994. Gynogenesis inthe African catfish (Clarias gariepinus). I. Induction of meio-gynogenesis with thermal and pressure shocks. Aquaculture 128:221–233.
Wattendorf, R.J., 1986. Rapid identification of triploid grass carpwith a coulter counter and channelyzer. Prog. Fish Cult. 48: 125–132.
Williams, D.J., S. Kazianis & R.B. Walter, 1998. Use of randomamplified polymorphic DNA (RAPD) for identification of large-mouth bass subspecies and their intergrades. Trans. Am. Fish.Soc. 127: 825–832.
Wills, P.S., R.J. Sheehan & S.K. Allen, Jr, 2000. Reduced reproduct-ive capacity in diploid and triploid hybrid sunfish. Trans. Am.Fish. Soc. 129: 30–40.
Wohlfarth, G., 1990. The tools of genetics in aquaculture, pp. 213in Mediterranean Aquaculture, edited by R. Flos, L. Tort & P.Torres. Ellis Horwood, N.Y.
Wolters, W., G.S. Libey & C.L. Chrisman, 1982. Effect of triploidyon growth and gonad development of channel catfish. Trans. Am.Fish. Soc. 111: 102–105.
Wu, C., R. Ye, R. Chen & X. Liu, 1993. An artificial multipletriploid carp and its biological characteristics. Aquaculture 111:255–262.
Yamamoto, E., 1999. Studies on sex-manipulation and produc-tion of cloned populations in hirame, (Temminck Schlegel).Aquaculture 173: 235–246.
Yamazaki, F., 1983. Sex control and manipulation in fish. Aquacul-ture 33: 329–354.
Young, W.P., P.A. Wheeler, V.H. Coryell, P. Keim & G.H. Thor-gaard, 1998. A detailed linkage map of rainbow trout producedusing doubled haploids. Genetics 148: 839–850.
Zanuy, S., M. Carrillo, M. Blázquez, J. Ramos, F. Piferrer & E.M.Donaldson, 1994. Production of monosex and sterile sea bass byhormonal and genetic approaches. Publ. Assoc. Devélop. Aquac.119: 409–423.
Zhang, Q. & K. Arai, 1999. Aberrant meiosis and viable aneuploidprogeny of induced triploid loach (Misgurnus anguillicaudatus)when crossed to natural tetraploids. Aquaculture 175: 63–76.
Zohar, Y., 1989. Endocrinology and fish farming: aspects in re-production, growth and smoltification. Fish Physiol. Biochem.7: 395–450.