induction of an antigen-specific immune response and partial protection of cattle against challenge...
TRANSCRIPT

Induction of an antigen-specific immuneresponse and partial protection of cattle againstchallenge infection with foot-and-mouth diseasevirus (FMDV) after lipopeptide vaccination withFMDV-specific B-cell epitopes
Bettina-Judith Hohlich,13 Karl-Heinz Wiesmuller,2 Bernd Haas,1
Wilhelm Gerner,1 Roberto Correa,3 Hans-Robert Hehnen,3
Tobias Schlapp,3 Eberhard Pfaff1 and Armin Saalmuller1
Correspondence
Armin Saalmuller
1Institut fur Immunologie, Bundesforschungsanstalt fur Viruskrankheiten der Tiere,Paul-Ehrlichstr. 28, D-72076 Tubingen, Germany
2EMC microcollections GmbH, Sindelfinger Str. 3, D-72070 Tubingen, Germany
3Bayer Animal Health, Building 6210, D-51368 Leverkusen, Germany
Received 22 May 2003
Accepted 19 August 2003
To evaluate the potential of chemically synthesized lipopeptides for vaccination against
foot-and-mouth disease (FMD), seven lipopeptides containing the immunostimulating principle
of bacterial lipoproteins and linear B-cell epitopes of FMDV strain O1Kaufbeuren (O1K) were used
to immunize cattle (n=7). Animals were vaccinated once and 21 days after immunization animals
were infected with the homologous virus. Four animals were protected. After vaccination, as
well as after challenge infection, B- and T-cell responses were examined. Sera were tested for
virus- and peptide-specific antibodies and showed after vaccination only a weak antibody
response. After challenge infection, an increase in antibody titre was obvious but there was no
correlation between antibody titre and protection. The reactivity of the cellular immune system was
detected by analyses of PBMCs for virus- and peptide-specific T-lymphocytes. With regard to
the virus-specific T-lymphocytes, a heterogeneous reactivity could be shown. No correlation
between virus-specific T-cell proliferation and protection was found. Obvious was the fact that
all protected animals showed after vaccination a strong T-cell response against at least one of
the peptides used for immunization. These results suggest a correlation between the onset of
an antigen-specific T-cell reaction and protection.
INTRODUCTION
Foot-and-mouth disease (FMD), a devastating disease oflivestock, is caused by FMD virus (FMDV), a member ofthe genus Aphthovirus within the family Picornaviridae.Although animals will generally recover from the acuteclinical phase of the disease, usually there is a permanentloss of performance, due mostly to chronic lameness,permanent drop in milk yield and poor weight gain. FMDis the most contagious virus disease of animals (Pereira,1981) and is endemic in many regions of the world,including much of Africa, Asia and some countries inSouth America. In FMD-endemic regions, the major costof the disease is associated with reduced livestock produc-tivity and reduced access to international markets for
livestock and livestock products. In many situations, regularvaccination is an essential part of the disease controlstrategy. Regions free of FMD include Europe, Australia,Japan and North and Central America. Here, controlis based upon prevention of introduction of the virusthrough rigorous importation regulation and quarantine.In case of a disease introduction into these areas, emergencyvaccination may have to support movement controls andstamping out in order to eradicate the disease.
Current FMD vaccines are produced by infecting culturedcells with virulent FMDV, followed by inactivation andpurification of the newly synthesized virus particles.Whereas the resulting ‘conventional’ vaccines have beenapplied successfully for decades, they have a number ofdisadvantages. Handling FMDV is inherently hazardous andnecessitates high security facilities (Barteling & Vreeswijk,1991). To ensure reliable inactivation, FMD vaccine plants
3Present address: Miltenyi Biotec, Friedrich-Ebert-Str. 68, D-51429Bergisch Gladbach, Germany.
0001-9366 G 2003 SGM Printed in Great Britain 3315
Journal of General Virology (2003), 84, 3315–3324 DOI 10.1099/vir.0.19366-0

need a special design and complex operating proceduresthat have to be strictly monitored. A conventional FMDvaccine is a biological product that is very difficult tostandardize. To ensure the required potency, animalexperiments are necessary, not only for vaccine registrationbut also for batch control (Barteling & Vreeswijk, 1991). Inaddition, vaccines will quickly lose potency without aconsistent cold chain from the producer to the veterinarianin the field. Furthermore, protection induced by conven-tional vaccines is limited mostly to one or a few topotypeswithin one serotype (Pereira, 1981). New strains are con-stantly evolving in endemically infected countries, makingit difficult for vaccine producers to keep pace. Protection isshort-lived, often making frequent revaccination indispens-able for an effective disease control (Barteling & Vreeswijk,1991). Whereas conventional vaccines can prevent clinicalsigns and further spread of disease in vaccinated popula-tions, they are not able to protect virus-exposed animalsfrom becoming persistently infected virus carriers.
To overcome these problems, a lot of work was performedon alternatives to conventional vaccines. Synthetic peptideswere early found to be promising vaccine candidates (Pfaffet al., 1982; DiMarchi et al., 1986; Doel et al., 1990). Majorantigenic regions of the virus were found on the structuralprotein 1D (Pfaff et al., 1982; Strohmaier et al., 1982;Acharya et al., 1989; Bittle et al., 1982). The antigenic sitecomprised of aa 140–160 of 1D (containing the receptor-binding site of the virus) is usually referred to as ‘site A’,whereas ‘site C’ contains aa 200–213 of 1D. In initialexperiments, peptide vaccines consisting of site A or ofconstructs combining sites A and C were used. Whereasthese vaccines induced considerable titres of neutralizingantibodies in guinea pigs, cattle and swine and sometimesalso good protection in vivo (Pfaff et al., 1982; Bittle et al.,1982; DiMarchi et al., 1986; Morgan & Moore, 1990), ingeneral, their immunogenicity was lower than that ofconventional vaccines. This may be due to the now widelyaccepted thesis: that the immune response against FMDV isnot only B-cell but also T-cell dependent (Domingo et al.,1990; Collen et al., 1991; Brown, 1992; Collen, 1994; Sobrinoet al., 1999). Because sites A and C contain mainly B-cellepitopes, in a recent study these immunogenic sites werecombined with a further 1D peptide containing a T-cellepitope (Taboga et al., 1997). This resulted in a minorincrease in protection rates.
The aim of this study was to evaluate the potential of lipo-peptides containing FMDV-specific B-cell epitopes fromstructural and non-structural (NS) proteins to induce anFMDV-specific immune response in cattle. A special focuswas laid on the variance of the immune response of differ-ent randomly selected outbred cattle against the peptidesused for the immunizations. In addition, the potency ofthe B-cell epitopes in the induction of a peptide and/orFMDV-specific T-cell reaction was studied. Furthermore,the capacity of the lipopeptides to induce protection againsta homologous challenge infection was investigated.
METHODS
Animals. All cattle used for vaccination experiments were approx-imately 1 year old and free of antibodies against FMDV, as con-firmed by liquid-phase blocking sandwich ELISA (LPBE) (Hamblinet al., 1986). Sera of cattle infected with FMDV strain O1Lausanne(O1L) were kindly provided by M. Parkhouse, Institute for AnimalHealth, Pirbright, UK. Sera of cattle infected with FMDV strainsA5Bernbeuren (A5B), Asia1Shamir (A1S) and SAT1Zimbabwe(SAT1Z) were obtained from infected animals between 10 and28 days after challenge infection.
Viruses. FMDV strain O1Kaufbeuren (O1K) was grown on mono-layers of BHK cells, clone Tubingen. Virus stocks used for prolifera-tion assays were stored as clarified tissue culture harvest material(3000 g for 20 min) at 270 uC. For LPBE (Hamblin et al., 1986),virus was inactivated with binary ethyleneimine and additionallyincubated at 32 uC overnight.
Virus used for challenge infections was passaged in cattle and eachanimal was infected with 265000 p.f.u. intralingually.
Peptides. Peptides were synthesized by conventional solid-phasemethodology (Merrifield & Stewart, 1965) and dissolved in a con-centration of 10 mg ml21 in DMSO. Overlapping peptide amides (14or 15 aa in length, overlapping each other by 10 aa) were syn-thesized based on the sequence of FMDV O1K (Forss et al., 1984) usingFmoc chemistry (Multiple Peptide Synthesizer SyRo, MultiSyntech).Synthesized peptides were analysed by analytical high-pressure liquidchromatography (System Gold, Beckman Coulter) on a NucleosilC18 column (Grom) and ion-spray mass spectroscopy (VG QuattroII Triple-Quatrupol Ionspray-MS, Micromass). The purities of crudepeptides were, in general, higher than 70%. For immunization pur-poses, peptides were synthesized as Pam3Cys peptides, as describedpreviously (Wiesmuller et al., 1989).
Vaccination and infection of animals. Vaccination was carriedout intramuscularly with 2 mg peptide per animal. Lipopeptideswere dissolved in PBS and emulsified with an equal volume ofMontanide ISA 9. A 1?5 ml volume per dose was administered.Challenge infections were carried out according to the methodolo-gies described by the European Pharmacopoeia for the determina-tion of the potency of conventional vaccines. A total of 104 p.f.u.FMDV O1K was inoculated intradermally into two sites on theupper surface of the tongue (0?1 ml per site). After 8 days, animalswere sedated with Rompun and examined carefully for FMD-specificlesions. Animals that showed lesions at sites other than the tongue(secondary lesions) at 8 days post-infection were considered non-protected. To control the challenge infection, two control animalswere included in the experiment. Both control animals developedlesions: one animal showed FMDV-caused lesions on all four extre-mities, the other one only on two extremities.
LPBE. LPBE was performed as described (Hamblin et al., 1986),with slight modifications. Briefly, ELISA plates (Nunc) were coatedovernight with rabbit anti-FMDV O1K serum. Twofold dilutionseries of each test serum were mixed with equal volumes of antigen(cell culture supernatant) of a predetermined dilution and kept at4 uC overnight in a dummy plate. After washing, each well of thecoated ELISA plates received 50 ml antigen/serum mixture. Controlwells received antigen/buffer mixture (antigen control) or antigen/standard serum mixture, respectively. After incubation for 1 h at37 uC, plates were washed and incubated with guinea pig anti-FMDV O1K serum. After incubation for 1 h at 37 uC, plates werewashed again and incubated for a further 1 h at 37 uC with goatanti-guinea pig gamma-globulin-peroxidase conjugate (Biozol).After the final washing step, the chromogen orthophenylenediamine(OPD) (Sigma) with H2O2 was added. The reaction was stopped
3316 Journal of General Virology 84
B.-J. Hohlich and others

after 15 min with H2SO4 and plates were read in an automaticmicroplate reader at 492 nm. Antibody titres were expressed as thedilution at which the test sera showed greater than 50% inhibitionof the absorbance value recorded for the antigen control wells. Titresbelow 1 : 40 (1?6 log10) were considered positive.
Peptide ELISA. Plates (Immunoplate Maxisorb, Nunc) were coatedwith 10 mg peptide ml21 in 100 ml H2O per well and air-dried at37 uC. Peptide-coated plates were blocked for 2 h at 37 uC with 3%BSA/PBS and then incubated with sera diluted in 3% BSA/PBS at a1 : 100 dilution for 1 h at 37 uC. Specific antibodies were detectedwith horseradish peroxidase-conjugated goat anti-bovine immuno-globulin antibodies (Dianova). Colour development was obtainedafter addition of the substrate (OPD with 0?2 ml H2O2 ml21). Thereaction was stopped with 2 M H2SO4. All plates were read in anautomatic microplate reader (Titertek).
Results are expressed as indices (average absorbance value of dup-licate wells of the sample/average absorbance value of the duplicatewells of the negative controls). As negative controls, three FMDV O1Kpeptides that did not contain linear B-cell epitopes were used. Theindices of all animals were below 2 before the first vaccination.
Lymphocyte proliferation assay. Bovine PBMCs were isolated byHistopaque (Histopaque 1083, Sigma) centrifugation of heparinizedblood. Cultivation of PBMCs (56105 ml21) was performed inMEM-a (Gibco-BRL) supplemented with 10% FCS, 2 mM L-glutamine,561025 M 2-mercaptoethanol, 10 mM HEPES buffer, 100 IU peni-cillin ml21 and 0?1 mg streptomycin sulphate ml21. For the pro-liferation assay, PBMCs were incubated for 5 days at 37 uC with 5%CO2 in 200 ml microcultures in round-bottomed microtitre plates(Greiner). PBMCs from vaccinated animals were restimulated witheither FMDV O1K (log2 titration, starting with 56106 p.f.u. perwell) or synthetic peptides (25 and 2?5 mg ml21). As negative con-trols, cell culture supernatants from mock-infected cells or irrelevant15-mer peptides derived from hepatitis C virus dissolved in DMSO(25 and 2?5 ml ml21) were used.
To quantify the proliferative response, 1 mCi [3H]thymidine(Amersham) per well was added for an additional 16–18 h. After themicroplates were freeze-thawed, the cells were harvested using a multi-harvester system (Wallac) and [3H]thymidine uptake was measuredin a b-counter (Wallac).
All results are expressed as stimulation indices (SI), which can bedefined as geometric mean c.p.m. of the triplicate microcultures of thesample/geometric mean c.p.m. of the triplicate microcultures of therespective negative controls. The SI determined before vaccinationwas below 2 for all animals.
RESULTS
Identification of FMDV-specific linear B-cellepitopes
FMDV-specific linear B-cell epitopes were identified byscreening sera of FMDV O1K-infected cattle for peptide-specific antibodies in a peptide ELISA, as mentioned above.Fifteen of 442 tetra- or pentadeca peptides, which over-lapped in 10 aa and spanned the whole ORF of FMDV O1K,could be identified as linear B-cell epitopes. As demon-strated in Fig. 1, these epitopes were distributed over fiveNS proteins (2B, 2C, 3A, 3B and 3D) and were also locatedin the RGD region of structural protein 1D (Fig. 1). Thisregion was characterized as the receptor-binding site ofFMDV and is known as the major immunogenic site of thevirus. The sequences of the peptides identified as FMDV-specific linear B-cell epitopes are presented in Fig. 2.
Analysis of the reactivity of the linear B-cellepitopes derived from FMDV O1K with serafrom cattle infected with other FMDV serotypes
Because of the fact that most of the peptides identified aslinear B-cell epitopes for strain O1K derived from highlyconserved regions of FMDV, their reactivities with serafrom 21 animals infected with other FMDV sub- andserotypes were studied (Fig. 3). In these experiments, inaddition to sera of FMDV O1K-infected cattle, sera ofcattle infected with FMDV strains O1L, A5B, A1S and SAT1Zwere examined also.
Fig. 1. Localization of FMDV O1K-specific linear B-cell epitopes for cattle. Schematic map of the FMDV ORF. Indicated arethe two NS protein regions encoding the NS proteins L/L9, 2A–2C and 3A–3D. The structural protein region encoding thecapsid proteins 1A–1D is also shown. Within the ORF the localization of the peptides, identified as linear B-cell epitopes(bars), is shown. Epitopes are numbered consecutively from the N-terminal end of the sequence. Synthetic peptides used forthe identification of the linear B-cell epitopes were 14 to 15 aa long with an overlap of 10 aa and represent the amino acidsequence of FMDV O1K.
http://vir.sgmjournals.org 3317
FMDV-specific B-cell epitopes

As expected, all FMDV strains were able to induceantibodies against several linear B-cell epitopes identifiedbefore. All sera tested so far shared common linear B-cellepitopes. But the B-cell response against epitopes derivedfrom the regions of the structural protein 1D, and the NSproteins 2B, 2C, 3A and 3D (peptides 1–9 and 15), showedwithin the group of animals great virus strain-independentanimal-to-animal variation. This could be demonstrated forthe group of FMDVO1K-infected animals (Fig. 3, lines 1–4)as well as for all other groups. Interestingly, most epitopesderived from the 3B region (peptides 10–14) were recog-nized more consistently within the groups. In this context, itshould be mentioned that protein 3B consists of three non-identical tandem repeats of the viral genome-associatedprotein (VPg) and the peptides derived from this proteinshare parts of their sequence. Peptides 10 and 11 belongto the first tandem repeat and show an overlap in 9 aa(PLERQKPLK), peptides 12 and 13 are based on theequivalent structures of the second repeat and share 9 aaalso (PMERQKPLK). Interestingly, peptide 13, which showedno difference between FMDV O1K and O1L in its sequence,was recognized less by the sera from the FMDV O1L-infected animals. Peptide 14, recognized by a more limitednumber of sera, is part of the third repeat, being equivalentto peptides 11 and 13. This third repeat in the 3B regionis the most heterogeneous part of the protein. The mostremarkable fact revealed by this comparison of the immuneresponse against FMDV-specific linear B-cell epitopes isthat each serum derived from an infected animal that wastested elicited antibodies against both, or, in the case ofthe animals #38 and #296, at least against one, of the
overlapping peptides 10 and 11. The peptides belongingto the second and third VPg repeat were recognized lessefficiently.
Immunization of cattle with lipopeptides anddetermination of their antibody responses
To determine the ability of the identified linear B-cellepitopes to induce an in vivo immune response and toprotect immunized animals from challenge infection,peptides identified as linear B-cell epitopes (Fig. 2) wereselected, synthesized and N-terminally elongated by theimmunostimulating lipoamino acid Pam3Cys. To reducethe number of peptides for the immunization experiments,only seven peptides were selected out of the 15 identified aslinear B-cell epitopes (Figs 1 and 2), namely peptides 1, 3, 4,6, 7, 11 and 15. Peptide 1 derived from the 1D structuralprotein contained the RGD region responsible for theattachment of the virus to the cellular receptor. The epitopecorresponding to peptide 3 is located in the C-terminal partof NS protein 2B. Peptides 4 and 6 describe two B-cellepitopes located in the N terminus of NS protein 2C.Peptide 7 was derived from protein 3A and peptide 11 fromthe first tandem repeat of protein 3B. Peptide 15 was partof the N terminus of protein 3D. The sequences of allthese peptides used for the immunization experimentsare summarized in Fig. 2. As mentioned above for thevaccination experiments, these peptides were synthesizedas lipopeptides and applied in an oil emulsion. Seven cattlewere immunized with a dose of 2 mg for each peptide peranimal. Blood samples were taken from each animal 21 days
Fig. 2. Sequence of FMDV O1K-specific linearB-cell epitopes. The amino acid sequences ofpeptides that contain linear B-cell epitopes forFMDV O1K are shown. Additionally, the viralprotein from which the sequence derived isindicated. The numbers of the peptides refer toFig. 1. For immunization experiments, only thepeptides indicated as being positive (+) wereused.
3318 Journal of General Virology 84
B.-J. Hohlich and others

later and the induction of specific antibodies against FMDVand against the synthetic peptides used for the immuniza-tion was determined. Animals were challenged with FMDVO1K, from which the sequence of the synthetic peptideshad been derived. In this experiment, four of seven animalswere protected (Fig. 4b).
To characterize the effects of lipopeptide vaccination andinfection on the immune response of the respective animals,the humoral immune response was investigated 21 daysafter vaccination and 17–22 days after challenge infection(Fig. 4). The specific antibody reactivity of non-protected(Fig. 4a) and protected (Fig. 4b) animals is compared inFig. 4. After vaccination, significant antibody titres againstwhole virus could be detected in neither the sera of unpro-tected (Fig. 4a) nor the sera of protected animals (Fig. 4b).After the challenge infection, all animals showed a clearantibody titre against FMDV in the LPBE. Interesting isthe fact that the antibody titres of the protected animals(Fig. 4b) were, on average, lower compared to the titres ofthe non-protected animals (Fig. 4a).
Investigating the sera derived after the first immunizationprior to the challenge infection for antibodies againstpeptides (Fig. 4, closed bars), the sera of most animalsshowed a very low reaction in the peptide ELISA and onlyagainst a few of the peptides that were included in thevaccine (A492>2). The serum of animal #69064 showed aresponse to peptide 15 and the serum of animal #99219showed a response against peptides 1 and 7. However, therewere also animals that did not elicit a detectable amount ofantibodies against any of the peptides, for example, theprotected animal #66671.
In short, the humoral immune response after vaccinationrevealed no conclusive pattern. About 3 weeks afterchallenge (Fig. 4, open bars), the sera of most animalsshowed an increased antibody level not only against thepeptides they had already responded to after the vaccina-tion but also against peptides they did not show anyresponse to before the vaccination. Protected animal #15879had developed antibodies against peptides 7 and 15 aftervaccination and additionally against peptides 1, 3, 6 and 11
Fig. 3. Reactivity of linear B-cell epitopes from FMDV O1K with sera from cattle infected with different FMDV sub- andserotypes. The existence of antibodies against peptides containing linear B-cell epitopes in sera of cattle infected with FMDVstrains O1K, O1L, A1S, A5B and SAT1Z is shown. Sera were examined in a peptide ELISA (seeMethods).Serumsamplesweredeemed positive for a peptide when the sample reached an SI of at least 2.
http://vir.sgmjournals.org 3319
FMDV-specific B-cell epitopes
after challenge infection. An exception was animal #43912,which showed, in contrast to all other animals, almost nodetectable antibody response against any peptide in thepeptide ELISA. This animal had also a very low antibodytitre in the LPBE after the first immunization, but afterthe challenge infection the antibody titre was quite high(titre>1 : 2400).
A comparison of the antibody responses of the fourprotected and the three non-protected animals showed noclear difference, neither with regard to the reactivity ofthe sera to FMDV in the LPBE nor in the peptide ELISA. Aminor point might be that the protected animals showedon average a higher antibody level against the peptidesafter challenge than the non-protected animals, but animal#71775 was not protected in spite of a good antibodyresponse to several peptides.
A summary of the data about the humoral immune res-ponse of lipopeptide-vaccinated cattle against FMDV andagainst synthetic peptides as described in detail aboveappears to be difficult and it seems to be impossible to makea clear correlation between antibody response and protec-tion against a challenge infection. Within the group ofprotected and also within the group of non-protectedanimals, animals with high and low antibody levels againstpeptides as well as FMDV could be found.
In vitro analysis of the cellular immuneresponse
Because no clear correlation between the humoral immuneresponse and protection could be recognized in the experi-ments described above, further investigations were directedtowards the second branch of the immune system – the
Fig. 4. B-cell response of cattle after vaccination with synthetic lipopeptides and after challenge infection. Sera of sevencattle vaccinated with a single dose of synthetic lipopeptides (see Fig. 2) were obtained 21 days after vaccination and 17(animals #43912 and #69064), 21 (animals #15879, #38715 and #99219) or 22 (animals #66671 and #71775) days afterchallenge infection. Sera were tested for antibodies against whole virus and against the peptides that were contained in thevaccine. The titres of antibodies against whole virus are presented on the right of the respective individual diagrams. Theupper rows marked by (a) indicate the antibody titre before and the numbers in the second rows marked by (b) indicatethe antibody titre after infection. The results obtained in the peptide ELISA (closed bars, after vaccination; open bars, afterchallenge) are presented as diagrams. The horizontal lines represent A492=1. All analyses are based on three independentexperiments with two replicates. SD values in all experiments were less than 10%. The animals were divided into two groups:non-protected (a) and protected (b) animals.
3320 Journal of General Virology 84
B.-J. Hohlich and others
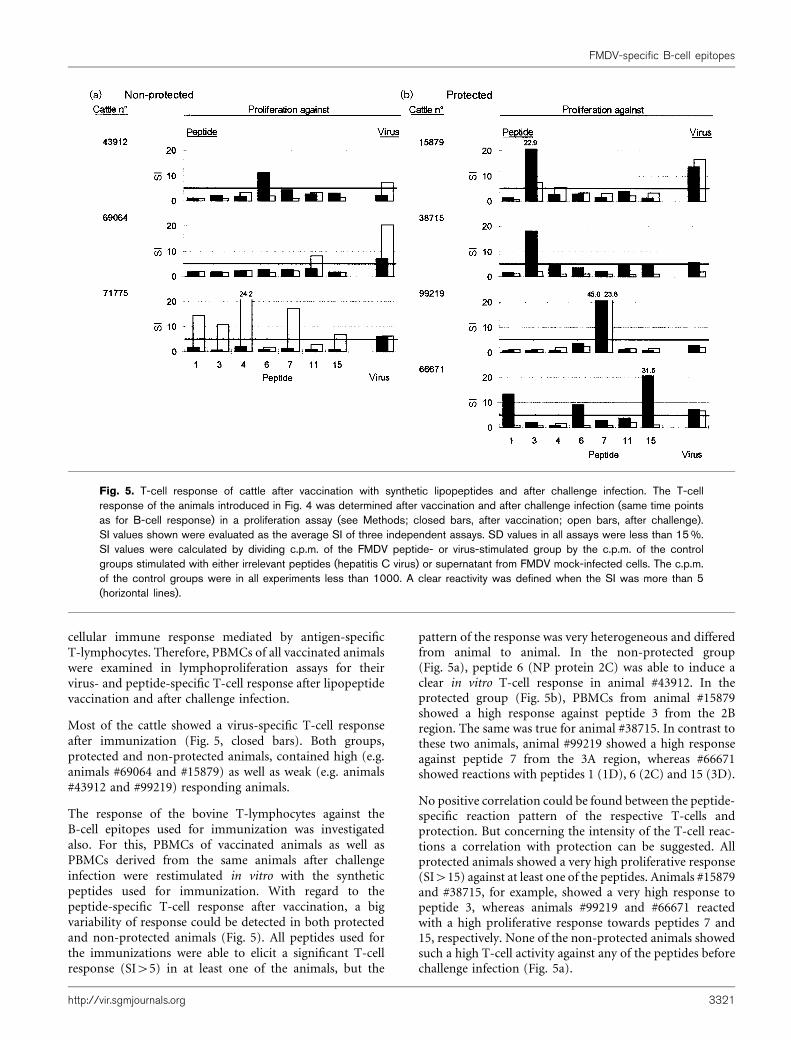
cellular immune response mediated by antigen-specificT-lymphocytes. Therefore, PBMCs of all vaccinated animalswere examined in lymphoproliferation assays for theirvirus- and peptide-specific T-cell response after lipopeptidevaccination and after challenge infection.
Most of the cattle showed a virus-specific T-cell responseafter immunization (Fig. 5, closed bars). Both groups,protected and non-protected animals, contained high (e.g.animals #69064 and #15879) as well as weak (e.g. animals#43912 and #99219) responding animals.
The response of the bovine T-lymphocytes against theB-cell epitopes used for immunization was investigatedalso. For this, PBMCs of vaccinated animals as well asPBMCs derived from the same animals after challengeinfection were restimulated in vitro with the syntheticpeptides used for immunization. With regard to thepeptide-specific T-cell response after vaccination, a bigvariability of response could be detected in both protectedand non-protected animals (Fig. 5). All peptides used forthe immunizations were able to elicit a significant T-cellresponse (SI>5) in at least one of the animals, but the
pattern of the response was very heterogeneous and differedfrom animal to animal. In the non-protected group(Fig. 5a), peptide 6 (NP protein 2C) was able to induce aclear in vitro T-cell response in animal #43912. In theprotected group (Fig. 5b), PBMCs from animal #15879showed a high response against peptide 3 from the 2Bregion. The same was true for animal #38715. In contrast tothese two animals, animal #99219 showed a high responseagainst peptide 7 from the 3A region, whereas #66671showed reactions with peptides 1 (1D), 6 (2C) and 15 (3D).
No positive correlation could be found between the peptide-specific reaction pattern of the respective T-cells andprotection. But concerning the intensity of the T-cell reac-tions a correlation with protection can be suggested. Allprotected animals showed a very high proliferative response(SI>15) against at least one of the peptides. Animals #15879and #38715, for example, showed a very high response topeptide 3, whereas animals #99219 and #66671 reactedwith a high proliferative response towards peptides 7 and15, respectively. None of the non-protected animals showedsuch a high T-cell activity against any of the peptides beforechallenge infection (Fig. 5a).
Fig. 5. T-cell response of cattle after vaccination with synthetic lipopeptides and after challenge infection. The T-cellresponse of the animals introduced in Fig. 4 was determined after vaccination and after challenge infection (same time pointsas for B-cell response) in a proliferation assay (see Methods; closed bars, after vaccination; open bars, after challenge).SI values shown were evaluated as the average SI of three independent assays. SD values in all assays were less than 15%.SI values were calculated by dividing c.p.m. of the FMDV peptide- or virus-stimulated group by the c.p.m. of the controlgroups stimulated with either irrelevant peptides (hepatitis C virus) or supernatant from FMDV mock-infected cells. The c.p.m.of the control groups were in all experiments less than 1000. A clear reactivity was defined when the SI was more than 5(horizontal lines).
http://vir.sgmjournals.org 3321
FMDV-specific B-cell epitopes

A strategy to overcome confusion about immune responseand protection might be a combinatorial view on B- andT-cell responses of the protected and non-protectedanimals. Maybe a summary of the B- and T-cell responses,as presented in Fig. 6, can give an explanation for acorrelation between the FMDV-specific immune responseand protection.
Looking at Fig. 6, in which the antigen-specific B- and T-cellresponses of all animals are summarized, it becomes moreand more obvious that all animals showing a high T-cellresponse after the peptide immunization were protected,independent of their B-cell response.
DISCUSSION
For this study, the whole ORF of FMDV O1K was examinedfor linear B-cell epitopes. Sections of 14 or 15 aa (over-lapping in 10 aa) were analysed and 15 linear B-cell epitopeslocated in the structural as well as in the NS protein regionof FMDV could be detected. For several of the peptides abroad reactivity with sera from cattle infected with differentother sero- and subtypes of FMDV could be shown. Thiscan be explained easily by the fact that most of the peptides(2–15) originate from NS proteins. Compared to the viral
capsid-forming structural proteins, the NS proteins showa considerably higher conservation in their amino acidsequence. The most consistent results in the tests for thereactivity of the antibodies could be found for peptides 10and 11, derived from the 3B protein. All FMDV-infectedcattle tested so far showed an antibody response againstat least one of these two overlapping peptides. The otherpeptides showed a higher inconsistency in their reactivitybut were also recognized by sera of cattle infected withFMDV serotypes others than O1K, which served as basisfor the peptide synthesis. This may lead to the conclusionthat mainly individual differences between animals areresponsible for the various recognition patterns of linearB-cell epitopes and not only the FMDV serotype with whichthey were infected.
These results indicate that if it should be possible to for-mulate a protective vaccine containing these B-cell epitopes,a good chance would exist that this vaccine would showprotection against several or most of the FMDV sero- andsubtypes.
The idea to use peptides for the development of a FMDVvaccine already existed in the eighties of the last century.Different B- and T-cell epitopes were used for protectionexperiments with cattle (DiMarchi et al., 1986; Morgan &Moore, 1990; Taboga et al., 1997; Volpina et al., 1999). Theresults varied strongly. The ratio of protected animals insome experiments was quite high in spite of the fact that inmost cases no more than two different peptides were usedfor vaccination. But there were already hints that the use ofonly few B- and T-cell epitopes supports the selection ofescape variants (Taboga et al., 1997). As in all of these earlyexperiments, all peptides were derived from the structuralproteins of FMDV, a heterotypic protection against differentFMDV serotypes appeared to be highly unlikely.
In contrast to these experiments, the study presented hereused a vaccine with seven different epitopes (six of seven)derived mainly from the NS protein region.
Regarding the peptide-specific B-cell response, vaccinatedanimals showed a reaction pattern against single peptidesthat was highly variable. Such variations in antigen recog-nition are already known for FMDV immunizations(McCullough et al., 1992). Even if a B-cell response is notMHC-restricted, quite often animal-to-animal variationsare observed for the same antigen (Riley, 1996).
In contrast to peptide-specific antibodies that could bedetected in the sera of most vaccinated cattle, only in fewanimals could antibodies against FMDV be determined withthe ELISA technique used commonly for FMDV diagnosis(Hamblin et al., 1986).
When looking at the cellular immune response, most cattleshowed a good reaction against single peptides after vac-cination. Astonishing was the extremely good proliferativeresponse that could be shown for some of the protected
Fig. 6. Summary of the peptide-specific T- and B-cell res-ponse of cattle induced by lipopeptides. The peptide-specificT- and B-cell response of the cattle introduced in Figs 4 and5, before challenge infection, in simplified terms. The B-cellresponse was regarded as negative (”) when the serum con-tained not enough antibodies against any peptide, contained inthe vaccine, to reach an SI of 3 (see Methods). The T-cellresponse of an animal was then regarded as negative when thePBMCs did not show an SI over 5 to any peptide, contained inthe vaccine, in the proliferation assay. Could a SI over 5 orover 15 be detected for at least one peptide the response wasdetermined as weak (+) or good (++), respectively. In addi-tion to the B- and T-cell response, it is indicated if the animalwas protected (+) or non-protected (”).
3322 Journal of General Virology 84
B.-J. Hohlich and others

animals (SI>15). As mentioned above, a correlation withthe antibody titre could not be presented in these animals.But despite the absence of detectable antibody titres againstFMDV, these animals were protected from challengeinfection. This is most probably due to an effective T-cellreaction that was able to support the formation of veryeffective neutralizing antibodies. On the other hand, thisprotection by T-lymphocytes might be explained by aneffective cellular immune response, including the activity ofT-helper cells as well as cytolytic T-lymphocytes. Furtherstudies might elucidate these speculations.
It was also interesting to note with regards to the T-cellresponse against the synthetic peptides that five of theseven peptides used for the immunization experimentsseemed to contain T-cell epitopes responsible for the in vitrorestimulation of the respective T-lymphocytes. Lipopep-tides are synthetic analogues of membrane lipoproteins ofbacteria and activate cells for cytokine release by theirinteraction with Toll-like receptors, which indicates amolecular link between host defence mechanisms andmicrobial products. (Aliprantis et al., 1999). Syntheticanalogues of the N-terminal part of these lipoproteinsconstitute potent immunoadjuvants in vitro and in vivo(Wiesmuller et al., 1992). One might also speculate thatthe Pam3Cys-coupled peptides were caught by the specificB-cell receptors, the surface immunoglobulin molecules,internalized into the B-cells and presented by B-cell MHCmolecules to the respective T-cell receptors of the specificT-lymphocytes. At this stage, it is not clear whether thepresentation is due mainly to MHC class II molecules asit might be expected or whether MHC class I moleculesare involved also. Nothing is known to date whether thePam3Cys-anchor molecule can be involved in a kind ofcross-presentation. These questions will be the content offurther studies.
Another peculiarity without clear explanation was thereduced proliferation against peptides after challengeinfection. The reason for this decrease of the proliferativecapacity is most probably due to the fact that the abilityof the T-cells to become restimulated was inhibited due tothe strong stimulation before. This could be a momentaryeffect, as described similarly for the mouse (De Mattia et al.,1999), but such a decreased reactivity of antigen-specificT-helper cells after restimulation could also occur afterrestimulation with the same antigen after consecutivevaccinations.
In general, it can be concluded that it seems that animalswere protected (four of seven) when a high T-cell activity(SI>15) could be shown for at least one peptide.
This information is important for a further optimization ofa potential peptide vaccine, and after a detailed analysis ofthis ex vivo data, it might be possible in future experimentsto avoid challenge infections. This would considerablyreduce the cost and time needed for further investigations.
ACKNOWLEDGEMENTS
The authors thank Bayer Leverkusen for funding, Andreas Moss forhelp with the animal experiments and Gabriele Kuebart and ThorstenDecker for technical help.
REFERENCES
Acharya, R., Fry, E., Stuart, D., Fox, G., Rowlands, D. J. & Brown, F.(1989). The three-dimensional structure of foot-and-mouth diseasevirus at 2?9 A resolution. Nature 337, 709–716.
Aliprantis, A. O., Yang, R. B., Mark, M. R., Suggett, S., Devaux, B.,Radolf, J. D., Klimpel, G. R., Godowski, P. & Zychlinsky, A. (1999).Cell activation and apoptosis by bacterial lipoproteins through Toll-like receptor-2. Science 285, 736–739.
Barteling, S. J. & Vreeswijk, J. (1991). Developments in foot-and-mouth disease vaccines. Vaccine 9, 75–88.
Bittle, J. L., Houghten, R. A., Alexander, H., Shinnick, T. M., Sutcliffe,J. G., Lerner, R. A., Rowlands, D. J. & Brown, F. (1982). Protectionagainst foot-and-mouth disease by immunization with a chemicallysynthesized peptide predicted from the viral nucleotide sequence.Nature 298, 30–33.
Brown, F. (1992). New approaches to vaccination against foot-and-mouth disease. Vaccine 10, 1022–1026.
Collen, T. (1994). Foot-and-mouth disease virus (aphthovirus):viral T cell epitopes. In Cell-Mediated Immunity in Ruminants,pp. 173–197. Edited by B. M. I. Goddeeris & I. Morrison. BocaRaton: CRC Press.
Collen, T., DiMarchi, R. & Doel, T. R. (1991). A T cell epitope in VP1of foot-and-mouth disease virus is immunodominant for vaccinatedcattle. J Immunol 146, 749–755.
De Mattia, F., Chomez, S., Van Laethem, F., Moulin, V., Urbain, J.,Moser, M., Leo, O. & Andris, F. (1999). Antigen-experienced T cellsundergo a transient phase of unresponsiveness following optimalstimulation. J Immunol 163, 5929–5936.
DiMarchi, R., Brooke, G., Gale, C., Cracknell, V., Doel, T. & Mowat, N.(1986). Protection of cattle against foot-and-mouth disease by asynthetic peptide. Science 232, 639–641.
Doel, T. R., Gale, C., Do Amaral, C. M. C. F., Mulcahy, G. &DiMarchi, R. (1990). Heterotypic protection induced by syntheticpeptides corresponding to three serotypes of foot-and-mouth diseasevirus. J Virol 64, 2260–2264.
Domingo, E., Mateu, M. G., Martınez, M. A., Dopazo, J., Moya, A. &Sobrino, F. (1990). Applied Virology Research, vol. 2, pp. 233–266.Edited by E. Kurstak, R. G. Marusyk & B. Murphy. New York:Plenum.
Forss, S., Strebel, K., Beck, E. & Schaller, H. (1984). Nucleotidesequence and genome organization of foot-and-mouth disease virus.Nucleic Acids Res 12, 6587–6601.
Hamblin, C., Barnett, I. T. & Hedger, R. S. (1986). A new enzyme-linked immunosorbent assay (ELISA) for the detection of antibodiesagainst foot-and-mouth disease virus. I. Development and method ofELISA. J Immunol Methods 93, 115–121.
McCullough, K. C., Bruckner, L., Schaffner, R., Fraefel, W., Muller,H. K. & Kihm, U. (1992). Relationship between the anti-FMD virusantibody reaction as measured by different assays, and protectionin vivo against challenge infection. Vet Microbiol 30, 99–112.
Merrifield, R. B. & Stewart, J. M. (1965). Automated peptidesynthesis. Nature 207, 522–523.
Morgan, D. O. & Moore, D. M. (1990). Protection of cattle and swineagainst foot-and-mouth disease, using biosynthetic peptide vaccines.Am J Vet Res 51, 40–45.
http://vir.sgmjournals.org 3323
FMDV-specific B-cell epitopes

Pereira, H. G. (1981). Virus Diseases of Food Animals, pp. 333–363.Edited by E. P. Gibbs. London, UK: Academic Press.
Pfaff, E., Mussgay, M., Bohm, H. O., Schulz, G. E. &Schaller, H. (1982). Antibodies against a preselected peptiderecognize and neutralize foot-and-mouth disease virus. EMBO J1, 869–874.
Riley, E. M. (1996). The role of MHC- and non-MHC-associatedgenes in determining the human immune response to malariaantigens. Parasitology 112, 39–51.
Sobrino, F., Blanco, E., Garcia-Briones, M. & Ley, V. (1999).Synthetic peptide vaccines: foot-and-mouth disease virus as amodel. Dev Biol Stand 101, 39–43.
Strohmaier, K., Franze, R. & Adam, K. H. (1982). Location andcharacterization of the antigenic portion of the FMDV immunizingprotein. J Gen Virol 59, 295–306.
Taboga, O., Tami, C., Carrello, E. & 13 other authors (1997). A
large-scale evaluation of peptide vaccines against foot-and-mouth
disease: lack of solid protection in cattle and isolation of escape
mutants. J Virol 71, 2606–2614.
Volpina, O. M., Surovoy, A. Y., Zhmak, M. N., Kuprianova, M. A.,
Koroev, D. O., Chepurkin, A. V., Toloknov, A. S. & Ivanov, V. T.
(1999). A peptide construct containing B-cell and T-cell epitopes
from the foot-and-mouth disease viral VP1 protein induces efficient
antiviral protection. Vaccine 17, 577–584.
Wiesmuller, K.-H., Jung, G. & Hess, G. (1989). Novel low-molecular-
weight synthetic vaccine against foot-and-mouth disease containing a
potent B-cell and macrophage activator. Vaccine 7, 29–33.
Wiesmuller, K.-H., Bessler, W. G. & Jung, G. (1992). Solid phase
peptide synthesis of lipopeptide vaccines eliciting epitope-specific B-,
T-helper and T-killer cell response. Int J Pept Protein Res 40, 255–260.
3324 Journal of General Virology 84
B.-J. Hohlich and others