incessant ovulation, inflammation and epithelial ovarian carcinogenesis: revisiting old hypotheses
TRANSCRIPT

Molecular and Cellular Endocrinology 247 (2006) 4–21
At the Cutting Edge
Incessant ovulation, inflammation and epithelial ovariancarcinogenesis: Revisiting old hypotheses
Jean S. Fleminga,∗, Clare R. Beaugieb, Izhak Havivc,Georgia Chenevix-Trenchd, Olivia L. Tane
a Eskitis Institute for Cell & Molecular Therapies, School of Biomolecular and Biomedical Sciences,Griffith University Nathan Campus, Nathan, Qld 4111, Australia
b Department of Anatomy & Structural Biology, Otago School of Medical Sciences, P.O. Box 913, Dunedin, New Zealandc Peter MacCallum Cancer Centre, Ian Potter Centre for Cancer Genomics & Predictive Medicine, St. Andrews Place,
East Melbourne, Vic. 3002, Australiad Cancer Genetics Laboratory, Queensland Institute of Medical Research, 300 Herston Road, Herston Brisbane, Qld 4029, Australiae Centre for Molecular Biotechnology, School of Life Sciences, Queensland University of Technology, Brisbane, Qld 4001, Australia
Received 8 September 2005; received in revised form 30 September 2005; accepted 11 October 2005
A
rch proceedsa terogeneousc all of whicha arise fromt genesis ae n profiling bym r diagnosis.©
K
C
1 . . 52 . 63 . 84 . . 95 . . 96 . 107 . 108 . 119 . 121 . . 14
. 15. . 15
0d
bstract
Epithelial ovarian cancer (EOC) is often a lethal disease because in many cases early symptoms go undetected. Although reseapace, as yet there are few reliable and specific biomarkers for the early stages of the disease. EOC is an umbrella label for a highly heollection of cancers, which includes tumours of low malignant potential, serous cystadenomas, mucinous and clear cell carcinomas,re likely to arise from a number of epithelial cell types and a variety of progenitor lesions. Many, but not all types of EOC are thought to
he cells lining ovarian inclusion cysts. In this review, we discuss the hypotheses that have driven our ideas on epithelial ovarian carcinondxamine the morphological and genetic evidence for pathways to EOC. The emergence of laser-capture microdissection and expressioicroarray technologies offers the promise of defining these pathways more accurately, as well as providing us with the tools for earlie2005 Elsevier Ireland Ltd. All rights reserved.
eywords: Ovulation; Epithelial ovarian cancer; Inflammation; Ovarian inclusion cyst
ontents
. Epithelial cell types in the ovary. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. Histopathological evidence for the aetiology of EOC. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. Genetic evidence for the aetiology of EOC. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. Incessant ovulation hypothesis. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. Gonadotrophin hypothesis. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. Hormonal hypothesis. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. The inflammation hypothesis. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. Expression profiling of EOC. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. Biomarkers for EOC. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .0. Conclusions. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
Acknowledgements. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .References. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
∗ Corresponding author. Tel.: +61 7 3735 3782; fax: +61 7 3735 37656.E-mail address: [email protected] (J.S. Fleming).
303-7207/$ – see front matter © 2005 Elsevier Ireland Ltd. All rights reserved.oi:10.1016/j.mce.2005.09.014
J.S. Fleming et al. / Molecular and Cellular Endocrinology 247 (2006) 4–21 5
Over 90% of ovarian cancers are described as epithelial inorigin. Epithelial ovarian cancer (EOC) is a highly heteroge-neous group of cancers, which can be classified into four mainsubtypes; serous, mucinous, endometrioid and clear cell, basedlargely on histological evidence (Korner et al., 2005). EOC is thefourth most common cause of death from cancer among womenin the United States (Jemal et al., 2005) and has the highestmortality rate of the gynaecologic cancers (Alvarez et al., 1999;Ho, 2003). The 5-year survival rate is less than 40% (Persson,2000), because of the presentation of the majority of cases atan advanced stage, but the aetiology and precursor lesions ofeven the major subtypes are poorly understood. While geneticstudies are beginning to unravel different pathogenic pathwaysto the different EOC subtypes, our studies on the origin of ovar-ian inclusion cysts in mouse ovaries suggest there may be morethan one epithelial cell of origin for serous ovarian adenomas(Tan et al., 2005). This review revisits the major hypotheses onthe aetiology of EOC, in the light of new evidence from mousemodels, mutation analysis and gene expression profiling.
1. Epithelial cell types in the ovary
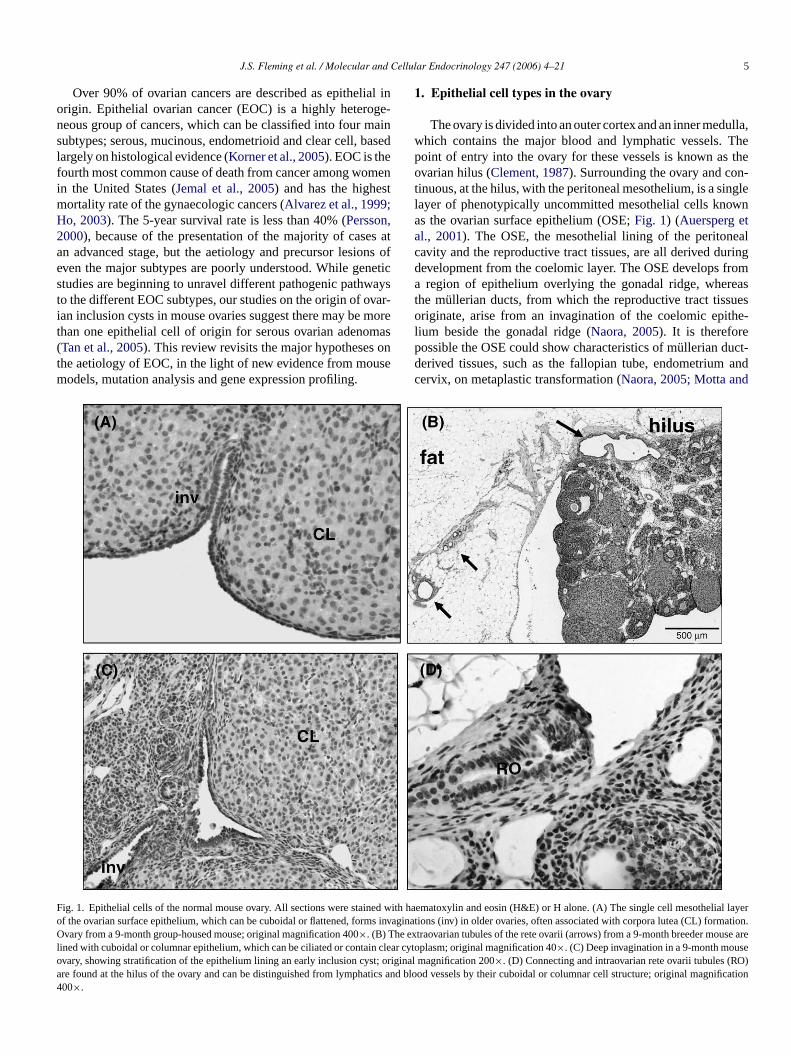
The ovary is divided into an outer cortex and an inner medulla,which contains the major blood and lymphatic vessels. Thepoint of entry into the ovary for these vessels is known as theovarian hilus (Clement, 1987). Surrounding the ovary and con-tinuous, at the hilus, with the peritoneal mesothelium, is a singlelayer of phenotypically uncommitted mesothelial cells knownas the ovarian surface epithelium (OSE;Fig. 1) (Auersperg etal., 2001). The OSE, the mesothelial lining of the peritonealcavity and the reproductive tract tissues, are all derived duringdevelopment from the coelomic layer. The OSE develops froma region of epithelium overlying the gonadal ridge, whereasthe mullerian ducts, from which the reproductive tract tissuesoriginate, arise from an invagination of the coelomic epithe-lium beside the gonadal ridge (Naora, 2005). It is thereforepossible the OSE could show characteristics of mullerian duct-derived tissues, such as the fallopian tube, endometrium andcervix, on metaplastic transformation (Naora, 2005; Motta and
FoOloa4
ig. 1. Epithelial cells of the normal mouse ovary. All sections were stained wf the ovarian surface epithelium, which can be cuboidal or flattened, forms invvary from a 9-month group-housed mouse; original magnification 400×. (B) The ex
ined with cuboidal or columnar epithelium, which can be ciliated or contain cleavary, showing stratification of the epithelium lining an early inclusion cyst; origre found at the hilus of the ovary and can be distinguished from lymphatics a00×.
ith haematoxylin and eosin (H&E) or H alone. (A) The single cell mesothelial layeraginations (inv) in older ovaries, often associated with corpora lutea (CL) formation.traovarian tubules of the rete ovarii (arrows) from a 9-month breeder mouse arer cytoplasm; original magnification 40×. (C) Deep invagination in a 9-month mouseinal magnification 200×. (D) Connecting and intraovarian rete ovarii tubules (RO)nd blood vessels by their cuboidal or columnar cell structure; original magnification

6 J.S. Fleming et al. / Molecular and Cellular Endocrinology 247 (2006) 4–21
Van Blerkom, 1980; Risch, 1998). Epithelial ovarian cancers,specifically serous, endometrioid and mucinous, show histo-logical and secretory properties of the epithelial and stromaltissues of the fallopian tube, endometrium and cervix, respec-tively, supporting this idea (Scully, 1977; Dubeau, 1999). TheOSE cells have the potential to become epithelial or mesenchy-mal cells in response to ovulation or hormonal stimuli (Feeleyand Wells, 2001). When they become cancerous, the cells appearto differentiate to a committed mullerian phenotype, losing theirplasticity and responsiveness to external cues (Feeley and Wells,2001). EOC is different from other cancers in that the epithe-lium becomes more differentiated as it transforms, rather thanless (Auersperg et al., 1997).
For many decades, the OSE was described as the germinalepithelium and its cells were thought to be capable of differenti-ating to form new primordial follicles, complete with oocytes(Allen, 1923). As more became known about fetal develop-ment of the ovary, this idea was discarded and the post-natalovary was regarded as incapable of producing new germ cells(Peters and McNatty, 1980). The recent publication of researchpointing to the existence of ovarian stem or progenitor cells,capable of re-seeding the ovary with new oocytes (Johnson etal., 2004, 2005; Bukovsky et al., 2004), has turned reproductivebiological dogma on its head. Given the increasing evidence thata variety of cancers, including breast cancer, may result fromtransformation of normal stem and progenitor cells (Woodwarde 03;A fm ithe-l re isk cellsi tia-t onst esist e int
oft als woro lopf entm iatedg ulet v,1 eo ulesa elopm le ini osestL
morp rao-v l,1 te isa esosa s, in
the medulla of the ovary. Intraovarian rete are lined by cuboidalepithelia, whereas the connecting and extraovarian rete are linedby ciliated columnar epithelial cells (Wenzel and Odend’hal,1985). The three tubule types are almost always connected.
Morphology of the rete ovarii appears to change with preg-nancy, being described as more luxuriant in pregnant women(Wenzel and Odend’hal, 1985). Changes in the rete ovariithroughout the oestrous cycle and pregnancy have also beenreported in the beagle and cow (Wenzel and Odend’hal, 1985).These results suggest the rete ovarii is under endocrine influence.A secretory role has been hypothesised for the rete ovarii, assecreted materials have been found in the tubule lumen of cows(Archbald et al., 1971) and cats (Gelberg et al., 1984). It has beensuggested the increased size of the rete ovarii during pregnancyin some species is caused by increased secretion from the cellslining the tubules. Morphological studies of ovaries from dogs,cows (Odend’hal et al., 1986) and sheep (Cassali et al., 2000)have revealed a physical connection between the rete ovarii andthe infundibulum of the uterine tube, although the functionalsignificance of this connection is unknown.
Research on the structure and function of the rete ovariihas primarily focused on these structures in animals and con-sequently little is known about the human rete ovarii. Muchof the limited information available on the human rete ovariihas come about because of the tendency for rete ovarii to bemistaken for the relatively common condition of endometriosis( ft stud-ie 04T waysa asso-c age,tw tion(c o nota sa thet erineto dMm therm fluid( edb andb e toa dS tley,2
2
spon-t e 2),o nign
t al., 2005; Liu et al., 2005; Clarke et al., 2005; Li et al., 20nderson and Clarke, 1999; Lochter, 1998), the existence oulti-potent stem cells capable of differentiating to an ep
ial phenotype, should not be disregarded. However, until monown about these proposed ovarian germ-line progenitorncluding their extraovarian or ovarian origin, their differenion pathways, their relationship to OSE cells and their respo factors thought to increase susceptibility to carcinogenhey must remain offstage in this review, awaiting their timhe limelight.
Another source of ovarian epithelial cells is the reticulumubules known as rete ovarii (Fig. 1). The rete ovarii is a normtructure of the adult mammalian ovary and consists of a netf epithelial cell-lined tubules, which are thought to deve
rom the mesonephric tubules. During ovarian developmesonephric cells migrate towards and into the undifferentonad, resulting in the extension of the mesonephric tub
hrough the mesovarian (Czernobilsky et al., 1985; Bysko978; Wenzel and Odend’hal, 1985). The function of the retvarii in the adult ovary is not well characterised and the tubre generally assumed to be a functionless vestige of devent. Byskov suggests the rete ovarii have an active ro
nitiating meiosis in the developing germ cells and prophe rete ovarii is the origin of the granulosa cells (Byskov andintern-Moore, 1973; Byskov et al., 1977).
The rete ovarii tubules exist in all species studied as threehologically distinct groups; the intraovarian rete, the extarian rete and the connecting rete (Wenzel and Odend’ha985). In humans, the major tubule of the extraovarian relso known as the epoophoron, which is located in the mlpinx. The intraovarian rete tubules are found near the hilu
,
e,
k
,
s
-
-
-
Khan et al., 1999; Woolnough et al., 2000). Dilation or cysts ohe rete ovarii have been described in almost all mammalsed, with the incidence of these cysts increasing with age (Clowt al., 2002; Thung et al., 1956; Altinoz and Korkmaz, 20).hese cysts are characterised by their location, which is alt the hilus or outside the ovary in the mesovarian region,iated with the ovarian ligament or adipose tissue. As micehe rete ovarii tubules dilate forming cystic structures (Fig. 2),ith epithelial hyperplasia and increased levels of prolifera
Clow et al., 2002; Thung et al., 1956; Beaugie, 2005). Largeystic rete ovarii may compress adjacent structures, but dppear to affect normal ovarian function (Clow et al., 2002; Sasnd Rehm, 1994). These cysts may form after blockage of
ubules, preventing drainage of secreted fluids into the utube (Odend’hal et al., 1986; Cassali et al., 2000). Cystic retevarii occur frequently in the ovaries of older women (Peters ancNatty, 1980; Rutgers and Scully, 1988; Nogales, 1995). Theorphology of these structures is similar to that seen in oammals, with cuboidal and columnar cells and clear cyst
Rutgers and Scully, 1988). They are usually always diagnosased on a hilar location and are often microscopic in sizeenign (Nogales, 1995). In rare cases, these cysts can give risdenomas and adenocarcinoma of the rete ovarii (Rutgers ancully, 1988; Nogales et al., 1997; Lee et al., 2001; Hea000).
. Histopathological evidence for the aetiology of EOC
EOC appears to arise via one of at least two pathways;aneously and aggressively, with no precursor lesion (Typr by slower development from an inclusion cyst to a be
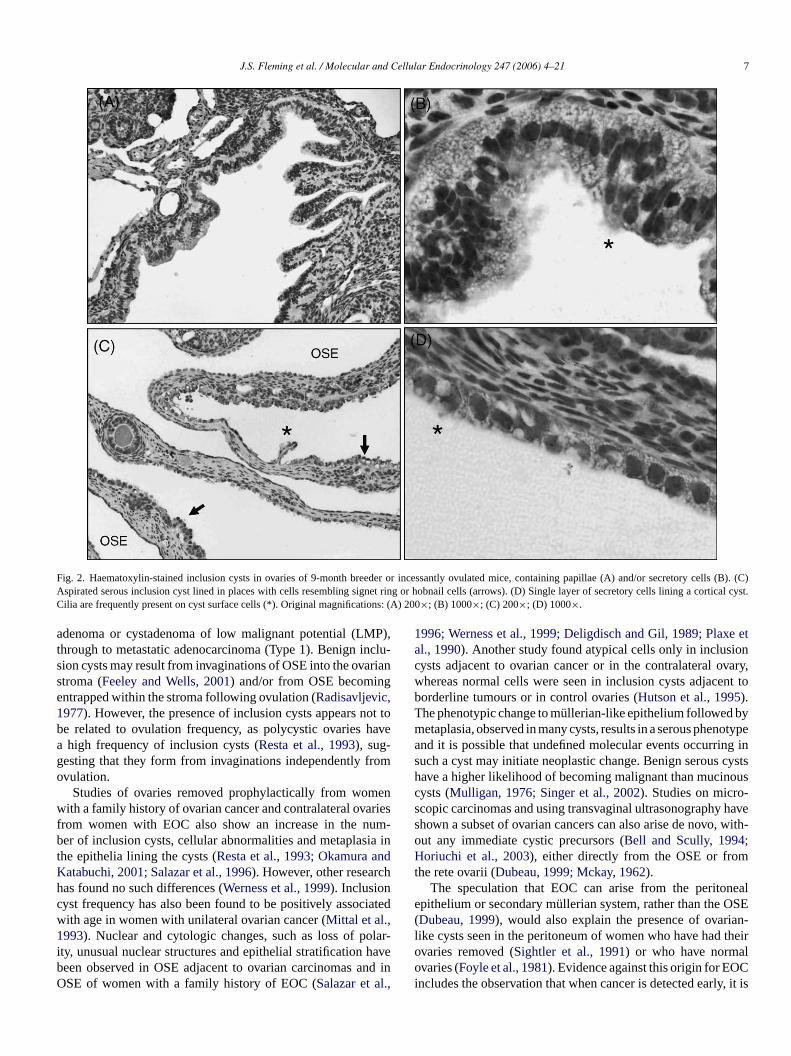
J.S. Fleming et al. / Molecular and Cellular Endocrinology 247 (2006) 4–21 7
Fig. 2. Haematoxylin-stained inclusion cysts in ovaries of 9-month breeder or incessantly ovulated mice, containing papillae (A) and/or secretorycells (B). (C)Aspirated serous inclusion cyst lined in places with cells resembling signet ring or hobnail cells (arrows). (D) Single layer of secretory cells lining a cortical cyst.Cilia are frequently present on cyst surface cells (*). Original magnifications: (A) 200×; (B) 1000×; (C) 200×; (D) 1000×.
adenoma or cystadenoma of low malignant potential (LMP),through to metastatic adenocarcinoma (Type 1). Benign inclu-sion cysts may result from invaginations of OSE into the ovarianstroma (Feeley and Wells, 2001) and/or from OSE becomingentrapped within the stroma following ovulation (Radisavljevic,1977). However, the presence of inclusion cysts appears not tobe related to ovulation frequency, as polycystic ovaries havea high frequency of inclusion cysts (Resta et al., 1993), sug-gesting that they form from invaginations independently fromovulation.
Studies of ovaries removed prophylactically from womenwith a family history of ovarian cancer and contralateral ovariesfrom women with EOC also show an increase in the num-ber of inclusion cysts, cellular abnormalities and metaplasia inthe epithelia lining the cysts (Resta et al., 1993; Okamura andKatabuchi, 2001; Salazar et al., 1996). However, other researchhas found no such differences (Werness et al., 1999). Inclusioncyst frequency has also been found to be positively associatedwith age in women with unilateral ovarian cancer (Mittal et al.,1993). Nuclear and cytologic changes, such as loss of polar-ity, unusual nuclear structures and epithelial stratification havebeen observed in OSE adjacent to ovarian carcinomas and inOSE of women with a family history of EOC (Salazar et al.,
1996; Werness et al., 1999; Deligdisch and Gil, 1989; Plaxe etal., 1990). Another study found atypical cells only in inclusioncysts adjacent to ovarian cancer or in the contralateral ovary,whereas normal cells were seen in inclusion cysts adjacent toborderline tumours or in control ovaries (Hutson et al., 1995).The phenotypic change to mullerian-like epithelium followed bymetaplasia, observed in many cysts, results in a serous phenotypeand it is possible that undefined molecular events occurring insuch a cyst may initiate neoplastic change. Benign serous cystshave a higher likelihood of becoming malignant than mucinouscysts (Mulligan, 1976; Singer et al., 2002). Studies on micro-scopic carcinomas and using transvaginal ultrasonography haveshown a subset of ovarian cancers can also arise de novo, with-out any immediate cystic precursors (Bell and Scully, 1994;Horiuchi et al., 2003), either directly from the OSE or fromthe rete ovarii (Dubeau, 1999; Mckay, 1962).
The speculation that EOC can arise from the peritonealepithelium or secondary mullerian system, rather than the OSE(Dubeau, 1999), would also explain the presence of ovarian-like cysts seen in the peritoneum of women who have had theirovaries removed (Sightler et al., 1991) or who have normalovaries (Foyle et al., 1981). Evidence against this origin for EOCincludes the observation that when cancer is detected early, it is

8 J.S. Fleming et al. / Molecular and Cellular Endocrinology 247 (2006) 4–21
located in the cortex near the OSE, not in the medulla or near thehilus of the ovary (Feeley and Wells, 2001). AlthoughDubeau(1999)argues that accurate evaluation of the anatomical positionof these cysts is difficult, because of their size, the distortion ofthe ovary and lack of serial sectioning,Feeley and Wells (2001)conclude the majority of EOCs originate either from the OSEor from inclusion cysts. Our observation that incessant ovula-tion leads to an increase in non-hilar, cortical inclusion cysts inthe mouse ovary, while suggesting the proportion of cysts fromeach location may vary with species, does support this conclu-sion (Tan et al., 2005).
3. Genetic evidence for the aetiology of EOC
Genetic analyses can help to unravel the origins and het-erogeneity of EOC at many levels. It is well recognised thatfamily history of ovarian or breast cancer is one of the majorrisk factors for EOC (Easton et al., 1996), and some of thehighly penetrant genes that contribute to this genetic suscepti-bility have been identified. Germ-line mutations inBRCA1 andBRCA2 confer an average cumulated risk of EOC by age 70of 39% and 11%, respectively (Antoniou et al., 2003). Invasiveserous EOC, and to a lesser extent, endometrioid invasive EOC,occur at increased frequency inBRCA1 andBRCA2 mutationcarriers, but not in LMPs or other histologic subtypes of EOC(Risch et al., 2001). Mutations in the hereditary non-polyposisc ko ioids ntm therg llelet ociat mos andm k fod l.,2 uedb ot bei -t thep etro ted ite
nferd anyr durino s oE ouc typeo ,1T(( Ca ten ie sup
pressor gene,TP53, is mutated in about 50% of invasive serousEOC, but only rarely in other histological subtypes, or in LMPtumours (Milner et al., 1997; Teneriello et al., 1993). Mutationsof theKRAS oncogene are found in about 30% of LMP seroustumours, but they are rarely observed in invasive serous tumours,which is strong evidence that some or all invasive serous EOCdo not derive from their LMP counterparts (Milner et al., 1997;Teneriello et al., 1993). This would be consistent with the find-ing that BRCA1 mutation carriers do not appear to have anincreased risk of serous LMP EOC, but only serous invasiveEOC (Bjorge et al., 2004). In contrast, identicalBRAF andKRASmutations have sometimes been found in benign epithelium andthe adjacent serous and mucinous ovarian LMP tumours (Hoet al., 2004; Mandai et al., 1998), which suggests thatBRAFandKRAS mutations are very early events in tumour develop-ment, and that serous and mucinous LMP tumours can arise frombenign epithelia. As well as occurring in about 50% mucinousLMPs,KRAS mutations have also been frequently found in inva-sive mucinous EOC (Enomoto et al., 1991; Mok et al., 1993),which suggests that, unlike serous EOC, invasive mucinoustumours may derive from their LMP counterparts (Mandai et al.,1998).
At another level, loss of heterozygosity in LMP tumoursis at a similar level to that in malignant mucinous tumours(Watson et al., 1998) and transitions between tumours of LMPand malignancy are found histologically in 80% of cases. Fur-t diag-n enigna ,2 s inb hem-s rozy-g someL s thes typeso 03T s, asw trioida roughm ,1
ls fort rela-t neticc andm arebe sa fromt thent ast,T highg e notb fromt litiesh ;B
olorectal cancer genes,MSH2 andMLH1, also increase the risf invasive EOC substantially, particularly of the endometrubtype (Watson et al., 1990). As well as the highly penetrautations in these susceptibility genes, it is likely that oenes exist with common, or perhaps rare, polymorphic a
hat have more subtle effects on risk of EOC. Many assion studies have been published evaluating the role of comingle nucleotide polymorphisms (SNPs) and risk of EOC,any of these studies suggest some heterogeneity of risifferent subtypes of EOC (Berchuck et al., 2004; Spurdle et a001a,b). However, association studies of this type are plagy publication of many false positive associations that cann
ndependently replicated (Wacholder et al., 2004). One excepion to this might be the association between a SNP inromoter of the progesterone receptor and risk of the endomid and clear cell subtypes of EOC, which has been replica
wo independent studies since the initial publication (Berchuckt al., 2004).
In addition to the evidence that germ-line variants coifferential risks for different subtypes of EOC, there are meports that the genetic and epigenetic changes that occurvarian tumourigenesis differ among the different subtypeOC. Furthermore, these somatic changes in sporadic tuman help to unravel the relationships between different subf EOC. There are several genes, such asCDKN2A (Shih et al.997), PTEN (Obata et al., 1998), TGFβRI (Chen et al., 2001),GFβRII (Lynch et al., 1998),km23 (Ding et al., 2005),CTNBB1Palacios and Gamallo, 1998; Wright et al., 1999) andPIK3CACampbell et al., 2004), which are occasionally mutated in EOnd for many of these genes the mutations occur most ofndometrioid or clear cell subtypes. By contrast, the tumour
s-n
r
i-n
gfrss
n-
hermore, mucinous tumours with benign components areosed 7–16 years on average earlier than those without breas, implying a temporal relationship (Feeley and Wells001). However, the similar rate of chromosomal lesionenign and LMP conditions, suggests those mutations telves could, sooner or later, lead to cancer. Loss of heteosity analyses also support the concept that at leastMP tumours may originate in benign cystadenomas, aame targets of allelic loss have been reported in bothf EOC (Chenevix-Trench et al., 1997; Thomas et al., 20).here is also evidence from loss of heterozygosity analyseell as numerous histological observations, that endomend clear cell ovarian carcinomas can sometimes arise thalignant transformation of endometrioid lesions (Jiang et al.998).
In summary, there are no detailed and validated modehe development and progression of ovarian tumours, theionship between the various forms, or the sequence of gehanges involved. However, there is clinicopathologicalolecular evidence for a model in which epithelial tumoursroadly divided into two types, Type 1 and Type 2 (Donningert al., 2004; Russell and McCluggage, 2004). Type 1 tumourre low-grade and develop through a hyperplastic process
he OSE to a benign lesion (such as an inclusion cyst), andhrough a LMP tumour and into the invasive form. In contrype 2 tumours are high-grade tumours, predominantlyrade serous carcinoma, for which precursor lesions haveen identified, and so they are thought to develop directly
he OSE. Different mutations and chromosomal abnormaave been associated with the two pathways (Korner et al., 2005ell, 2005).

J.S. Fleming et al. / Molecular and Cellular Endocrinology 247 (2006) 4–21 9
In order to understand the underlying causes and typesof EOC, it is useful to return to the major hypotheses thathave attempted to relate the epidemiology to reproductivephysiology. These include the incessant ovulation hypothesis(Fathalla, 1971), the gonadotrophin hypothesis (Cramer andWelch, 1983) and the hormonal hypothesis (Risch, 1998). Theevidence for and against each of these theories is summarisedbelow.
4. Incessant ovulation hypothesis
The incessant ovulation hypothesis proposes that ovulationtraumatises the ovarian surface, because rupture of the ovulat-ing follicle damages the OSE, requiring immediate repair. Overtime, this process of continuous damage and OSE proliferationto repair the wound, places strain on the OSE, increasing thechance of errors occurring during replication (Fathalla, 1971).Therefore, women with high lifetime ovulation numbers are atan increased risk of developing EOC. Over the last century inci-dence of EOC has increased, as have the average number ofovulations experienced in a lifetime (Banks et al., 1993). A cen-tury ago women often had many children, and thus ovulation wasfrequently suppressed either by pregnancy or lactation. It hasbeen estimated that women living in the 19th century ovulated40–50 oocytes, whereas modern women are thought to ovulate400–500 oocytes in their reproductive lifetime (Banks et al.,1 e ofa umb ,1 z eta ische gri ea 001;M ro r uso ingE atedf cantc theos ’ anc ;C t al2
d byf atem sta ers.H anl ate ac refleo meaS everr Theo C arh oduc
eggs (Fredrickson, 1987; Wilson, 1958), adding further supportfor the incessant ovulation hypothesis.
However, there are some who feel this model is too simplistic(La Vecchia et al., 1983; Siskind et al., 2000). The protectiongained from pregnancy or even short-term oral contraceptive useis far superior to that expected if ovulation number alone wereconsidered (Siskind et al., 2000; Gwinn et al., 1990; Greer et al.,2005). Use of the oral contraceptive pill reduces the relative riskof developing EOC by 7% for each year of use, with protectionpersisting beyond 15 years of exposure (Siskind et al., 2000).Each month of pregnancy has shown to reduce EOC risk by2.6% and each month of breast-feeding by 2.4% (Gwinn et al.,1990). In addition, use of the progestin-only contraceptive pill(which does not suppress ovulation) has been shown to be evenmore protective against EOC than the combined oral contracep-tive pill (Risch, 1998). There is also evidence that women withsome forms of anovulatory infertility, such as polycystic ovariansyndrome, are at increased risk of developing EOC (Schildkrautet al., 1996). Other forms of infertility also appear to increase therisk of developing EOC. It is well documented that nulligravidwomen are at increased risk of developing EOC (Risch et al.,1983, 1994; Riman et al., 2002; Purdie et al., 1995). However,it has been reported that women who have never been preg-nant because of infertility have a 40% higher rate of developingEOC than women who have never attempted to become pregnant(Rodriguez et al., 1998b).
5
ssiveg on ofta ont tingo
anda theap tingp -c rionicgW e ah tio,a ta esisa f theg e ino rtil-i nceh ourd wereg menw lce heri trol
993). Fathalla’s hypothesis is further supported by evidencdecrease in EOC risk with decreased lifetime ovulation ner, through pregnancy or oral contraceptive pill use (Shu et al.989; Whittemore et al., 1992; Whittemore, 1993; Rodriguel., 1998a; Tortolero-Luna et al., 1994; Adami et al., 1994; Rt al., 1983, 1994; Riman et al., 2002; Nasca et al., 1984; Nel., 1991; Akhmedkhanov et al., 2001; Titus-Ernstoff et al., 2odan et al., 2001; Tavani et al., 2000). In addition, the longe
vulation is suppressed (from more pregnancies or longef the oral contraceptive pill), the lower the risk of developOC. When the total lifetime ovulation number was calcul
or women who had EOC and for those that did not, a signifiorrelation between high total lifetime ovulation number andccurrence of cancer was found (Purdie et al., 2003). Similartudies investigating an association between ‘ovulatory ageancer risk have also found this to be the case (Whittemore, 1993asagrande et al., 1979; Moorman et al., 2002; Beard e000).
The incessant ovulation hypothesis is further supporteact that epithelial ovarian tumours are rare in other non-prim
ammals (Land, 1993; Gondos, 1975), possibly because monimals do not normally reach high total ovulation numbumans are one of the few species that continuously cycle. M
arge farm animals are seasonal breeders and will only ovulertain times of the year, whereas some smaller animals arevulators and will only ovulate if mated. Alternatively, sonimals will only cycle in the presence of a male (Austin andhort, 1984). It is therefore thought that other species can n
each an ‘ovulatory age’ high enough to develop cancer.nly species other than humans to frequently develop EOens, specifically those that have been hyperovulated to pr
-
t
e
d
.,
ytx
ee
. Gonadotrophin hypothesis
The gonadotrophin hypothesis states that exceonadotrophin exposure increases oestrogenic stimulati
he OSE, possibly leading to malignant transformation (Cramernd Welch, 1983). Gonadotrophins could act either directly
he OSE, enhancing transformation, or indirectly by stimulaestrogen production (Mohle et al., 1985).
Gonadotrophin levels increase with increasing agere particularly high during menopause, consistent withge-specific rates of EOC (Beltsos and Odem, 1996). Bothregnancy and the oral contraceptive pill lower circulaituitary gonadotrophin levels (Risch, 1998), although signifiant increases in levels of both oestrogen and human choonadotrophin (hCG) occur during pregnancy (Risch, 1998).omen with polycystic ovarian syndrome frequently hav
igh luteinising hormone to follicle stimulating hormone ralong with an elevated risk of developing EOC (Schildkraut el., 1996), lending support for the gonadotrophin hypotht the expense of the incessant ovulation hypothesis. Ionadotrophin theory holds true, a significant increasvarian cancer risk with gonadotrophin use for in vitro fe
sation or infertility would be expected, however the evideere is equivocal. A significantly increased risk for tumevelopment has been reported in infertile women whoiven gonadotrophin treatment, compared with infertile woho were not (Rossing and Daling, 1994). However, actuaases of EOC appear not to differ between these groups (Harrist al., 1992). Rats stimulated with gonadotrophins had a hig
ncidence of ovarian cyst formation compared with the con

10 J.S. Fleming et al. / Molecular and Cellular Endocrinology 247 (2006) 4–21
group, although none of the rats developed EOC (Celik et al.,2004).
Some epidemiological data conflict with the gonadotrophinhypothesis. Lactating women have raised levels of circulatingFSH, yet the epidemiology suggests breast-feeding is protective(Adami et al., 1994; Harris et al., 1992). Conversely, some EOCpatients have been shown to have lower levels of gonadotrophins(Risch, 1998; Ness and Cottreau, 1999) and women with particu-larly low levels of circulating gonadotrophins are at an increasedrisk of developing EOC (Daly and Obrams, 1998).
6. Hormonal hypothesis
The hormonal hypothesis attempts, in part, to resolve theseconflicts and proposes that excess androgen stimulation of theOSE leads to increased risk of cancer, whereas progesteronestimulation of the OSE is protective of EOC (Risch, 1998).Androgens are produced in the ovary by the developing folliclesat higher rates than estrogens (Risch, 1998) and are the prin-cipal sex steroids present within the fluid of growing follicles(McNatty et al., 1979). It has been proposed that epithelia-linedinclusion cysts located near developing follicles may be exposedto high levels of androgens (Risch, 1998). Androgen recep-tors have been detected in OSE and rete ovarii epithelial cellsof canine, primate and human ovaries (Saunders et al., 2000;Vermeirsch et al., 2001; Edmondson et al., 2002) and studiesi on inbH s np r-a ogel C int no e anhe ellk ucti(
velso tinsw roneL thee ;R esn e og e pi( elsa s. Ith ncyt leveA hasb - anp -l tione ,1 ures
resulted in a decrease in basal and oestradiol-17�-stimulatedproliferation rates, as well as up-regulation of p53 tumour sup-presser gene expression (Murdoch, 2002). Progesterone has alsobeen shown to induce apoptosis and up-regulate p53 expressionin human EOC-derived cell lines (Bu et al., 1997), suggestingthe steroid is capable of inhibiting cell division, even after trans-formation. When administered to monkeys, progesterone caninduce apoptosis in the OSE in vivo (Rodriguez et al., 1998a).Hens treated with progesterone have a decreased incidence ofEOC, supporting the hypothesis that progesterone induces apop-tosis of damaged OSE cells (Fredrickson, 1987; Rodriguez et al.,2001).
Reversible formation of ovarian inclusion cysts has beenreported frequently in breast cancer patients receiving tamoxifenchemotherapy (Cohen et al., 2003). Tamoxifen is a non-steroidalanti-oestrogen in breast tissue, but appears to be oestrogenic intissues such as ovary (Mourits et al., 1999; McCluggage andWeir, 2000). Treatment with tamoxifen can raise 17�-oestradiolserum levels significantly and is associated with the developmentof bilateral ovarian cysts in 80% of treated patients (Cohen et al.,1999). Breast cancer patients receiving tamoxifen only developovarian cysts if their ovaries are able to respond to FSH stimu-lation as shown by oestradiol production (Mourits et al., 1999)and the cysts appear to resolve in many patients on cessationof treatment (Cohen et al., 2003; Inal et al., 2005). There is noevidence that tamoxifen exposure is associated with an increasei lasm(
7
tion,w orym nsivet yf rfacefi henu ;M icaa olliclew ndento f tissuer CM)p statics
ep ronerr sultsi ells oft rinef ndo d int tion(
d cells
n guinea pigs have shown androgens stimulate proliferatioth inclusion cysts and OSE (Silva et al., 1997; Bai et al., 2000).owever, androgen stimulation of human OSE cultures haroliferative effect (Karlan et al., 1995). One of the key chacteristics of polycystic ovarian syndrome is elevated andr
evels and these may contribute to increased risk of EOhis condition (Schildkraut et al., 1996). A link has also beebserved between increased EOC risk and a history of acnirsutism, both symptoms of elevated androgen levels (Wyndert al., 1969). Finally, the oral contraceptive pill, which is wnown as protective, suppresses ovarian testosterone prodGaspard et al., 1983).
Not only does the oral contraceptive pill decrease lef androgens, but it also contains high levels of progesith potencies considerably higher than that of progesteong-term exposure to high levels of progestins could bexplanation for the pill’s significant protective effect (Ho, 2003isch, 1998). In addition, the progestin-only pill, which doot suppress ovulation, decreases EOC risk to the samreater level than that seen with the combined contraceptivRosenberg et al., 1994). During pregnancy progesterone levre very high, dwarfing circulating androgen concentrationas been suggested that the protection gained from pregna
hrough the 8–9 months of continuously high progesteronels, rather than through suppression of ovulation (Risch, 1998).n inhibitory effect of progesterone on cell proliferationeen demonstrated in cell cultures of OSE from some preost-menopausal women (Ivarsson et al., 2001), although simi
ar studies have found no effect of progesterone on proliferaither in the rabbit (Bai et al., 2000) or human (Karlan et al.995). Administration of progesterone to sheep OSE cult
o
n
d
on
,.
rll
is-
d
,
n benign or malignant primary or metastatic ovarian neopMcGonigle et al., 1999).
. The inflammation hypothesis
The ovulatory process resembles an inflammatory reacith an infiltration of leukocytes and production of inflammatediators such as cytokines, closely associated with exte
issue remodelling (Bonello et al., 1996). As the pre-ovulatorollicle expands, the theca externa and OSE cells on the surst proliferate to accommodate the growing follicle and tndergo apoptosis as the follicle wall thins (Murdoch, 1995urdoch et al., 1999). The connective tissue layers of the tunlbuginea and theca externa must be weakened before the fall can break open. These changes are probably depen release of collagenases and proteases characteristic oesponse to inflammatory reactions. Extracellular matrix (Eroteolysis is also an important step in the growth and metapread of any tumour (Bonello et al., 1996).
Initiation of follicle wall breakdown stimulated by thre-ovulatory luteinising hormone surge induces progesteeceptor synthesis in granulosa cells (Clemens et al., 1998). Theupture of the ovulatory follicle and release of the oocyte ren an increase in progesterone synthesis by the granulosa che ovulated ovary, now freed from control by oocyte paracactors (Hunter et al., 2005). Production of progesterone arouvulation helps maintain DNA integrity in OSE cells damagehe wave of apoptosis and DNA fragmentation prior to ovulaMurdoch, 1995, 1994).
Inflammatory stimuli induce 11� hydroxysteroidehydrogenase-1 expression and activity in granulosa

J.S. Fleming et al. / Molecular and Cellular Endocrinology 247 (2006) 4–21 11
(Tetsuka et al., 1999) and OSE cells (Yong et al., 2002), causingthe reversible metabolism of cortisone to cortisol (Tanninet al., 1991; Stewart and Mason, 1995). Cortisol may act tocounteract the inflammation arising at ovulation (Yong et al.,2002; Andersen and Hornnes, 1994; Escher et al., 1997; Hillierand Tetsuka, 1998; Rae et al., 2004). The main mediator ofthe inflammatory reaction may be nitric oxide released frominvading leukocytes, which acts as a potent vasodilator andmediates interleukin-1 beta-directed tissue remodelling eventswithin the ovary (Bonello et al., 1996). However, constitutivelyexpressed endothelial nitric oxide synthase (e-NOS) andinducible i-NOS are also present in the stromal cells and thecaof the developing follicles and are up-regulated by hormonalstimulation at ovulation (Zackrisson et al., 1996). Thus, eachround of ovulation is associated with leukocyte invasion, releaseof nitric oxide and inflammatory cytokines, vasodilation, DNArepair and tissue remodelling. Differentiation of the ovulatedfollicle into the new corpus luteum also involves substantialangiogenesis (Zackrisson et al., 1996). The persistence ofgenetic damage caused by inflammatory factors may thereforebe an important factor in transformation of damaged OSE cellsto EOC (Ness et al., 2000).
Hardiman et al. (2000)have suggested that genital tract infec-tions may be one factor that can give rise to both infertility andovarian cancer in women who are unable to conceive despitemedical treatment. Infection, leading to an acute inflammatoryr atorr ithl se ing nseq hinsar pirina for al ove actv trans ntlyd prot heliag nes.T rowtl ;A ytec ytesp t priot
infil-t eta e ofc sa nanc t al.1 f Tc anta ll suv its
regulatory (immune attenuating) T cells (CD4/CD25/FOXP3positive) is associated with worse outcome in ovarian cancer(Curiel et al., 2004). On the other hand, macrophages can stim-ulate tumour growth and/or vascularisation (Malik and Balkwill,1991; Mantovani et al., 1992). COX-2 expression is stimulatedin normal ovary by tumour necrosis factor-alpha, but in EOC thissignalling is uncoupled and COX-2 expression is frequently lost(Yang et al., 2005). Loss of COX-2 also correlates closely withmorphological transformation of the epithelium to a neoplasticphenotype. This suggests COX-2 expression does not contributedirectly to EOC malignancy, unlike in other epithelial cancers(Yang et al., 2005).
There have been numerous epidemiological studies linkingexposure to inflammatory factors such as talc or asbestos parti-cles, to higher EOC risk (Purdie et al., 1995; Ness and Cottreau,1999; Daly and Obrams, 1998; Baker and Piver, 1994; Chen etal., 1992). Direct evidence for such a connection is somewhatharder to find. A study on ovaries from women reporting fre-quent use and no use of perineal powdering with talcum powder,revealed talc particles on the surface of all ovaries, definitivelydemonstrating that the particles can reach the upper genital tractand suggesting post-natal use of talc may have contributed tothe finding (Heller et al., 1996). Two early studies in the ratshowed inflammatory and metaplastic effects on the OSE fromtalc injected under the bursal capsule (Hamilton et al., 1984),as well as demonstrating that talc particles could migrate fromtW to theo y for2 data;F og-i ughe ovar-i se tot
8
ointt g them bil-i ndt elvesu el-e omec s ofb ringe stan-d comet reat-m ofr
pre-h her ina itor-i gnalt 2
esponse, may block the endogenous ovulatory inflammesponse and inhibit ovulation. Primary ovarian failure wack of oestrogen production, may also lead to an increaonadotrophin release from the anterior pituitary and couently a higher exposure of the ovary to both gonadotropnd androgens (Cramer and Welch, 1983). The risk of EOC iseduced in women who are consistent users of low dose ascetaminophen or non-steroidal anti-inflammatory agents
east 6 months (Altinoz and Korkmaz, 2004; Akhmedkhant al., 2001). These anti-inflammatory agents are thought toia a shared pathway, dependent on the suppression ofcription factor NF-kappaB activity, which may subsequeecrease transcription of growth factors, chemokines and
eases, including cyclooxygenase (COX)-2, vascular endotrowth factor (VEGF) and various interleukins and chemokihese can induce angiogenesis, invasion, autocrine g
oops and resistance to apoptosis (Altinoz and Korkmaz, 2004khmedkhanov et al., 2001). Chemokines such as monochemoattractant peptide 1 are potent attractors of leukocarticularly macrophages and T cells and are released jus
o ovulation (Altinoz and Korkmaz, 2004).Ovarian tumours have also been reported to contain
rating macrophages and T cells (Merogi et al., 1997; Negusl., 1997), probably attracted by the production and releashemokines (Burke et al., 1996; Negus et al., 1995). Chemokinere capable of initiating an immune response against maligells (Parmiani, 1990; Rollins and Sunday, 1991; Peoples e993; Goedegebuure et al., 1997) and, indeed, the presence oells in ovarian cancer patients has been shown to be significssociated with increased progression-free time and overaival (Zhang et al., 2003). Consistently, a chemokine that recru
y
-
,t
-
-l
h
,r
t,
lyr-
he perineum to the ovarian surface (Henderson et al., 1986).e have also demonstrated the passage of talc particles
varian surface of mice, after perineal dusting once a damonths (N. Armanasco and J.S. Fleming, unpublishedig. 3), but this study failed to show any significant morphol
cal changes in the OSE in such a short time frame. Althonvironmental factors, such as talc, can clearly access the
an surface, the significance of the inflammatory responheir presence has yet to be demonstrated.
. Expression profiling of EOC
The four hypotheses on the aetiology of EOC may powards causal factors, but the main obstacle to improvinortality rate of this so often fatal disease is our lack of a
ty to identify the cancer early, to fight it more effectively ao prevent its appearance. Microarrays have proved themsseful in classification of tumours into distinct, clinically rvant subtypes and the prediction of clinical outcomes. Slassification studies are now moving from the laboratorieasic investigators to large-scale clinical trials. As measuxpression of all genes in the human genome becomes aard procedure, global gene expression analysis may be
he primary tool by which we can achieve personalised tents. This will be particularly important in the prediction
esponse to chemotherapy (Jazaeri et al., 2005).The advantage of microarrays is that you obtain a com
ensive parallel record of genes that are expressed togetcondition-dependent manner. This in turn allows the mon
ng of groups of genes that respond in parallel to similar siransduction pathways (Segal et al., 2004; Ihmels et al., 200).

12 J.S. Fleming et al. / Molecular and Cellular Endocrinology 247 (2006) 4–21
Fig. 3. Scanning electron micrograph (SEM) of talc particles (arrow) on the ovary of a mouse subjected to a daily dusting of talc on the perineum for 12 weeks. Leftinset: SEM of talcum powder used for dusting mice. Right inset: SEM of talc particles on the OSE of a different ovary, showing evidence of particle engulfment byepithelial cells (arrow).
The heterogeneity of EOC, compared with other types of cancer,makes interpretation of the results extremely difficult, since itis likely that multiple independent molecular mechanisms mayunderlie the same phenotype in different samples. For the mostpart, microarray results are more interpretable, beyond endlessgene lists, when the experiments conducted have clearly definedaims and a strategy that allows a focus on the genes that relate tothese aims. Here we will review recent examples of how genomictechnology can be used to improve our understanding, detectionand treatment of ovarian cancer, with emphasis on studies wherethe aims were clearly outlined and pursued.
9. Biomarkers for EOC
The main cause of fatality in EOC is believed to be the latestage that patients present with symptoms. Therefore, the incen-tive for early detection is enormous. Microarrays are ideallypositioned to provide data on potential biomarkers for this pur-pose and indeed, a number of such genes have been identifiedso far. Since the ultimate aim is a non-invasive test, focus on up-regulated genes encoding transmembrane receptors and secretedproteins is crucial. The main technical obstacles in EOC detec-tion are the ambiguity of the cellular origin of ovarian cancerand the small numbers of possible progenitor cells. The OSEamounts to no more than 300,000 cells in the adult body. Onepossibility would be to compare late and early stage ovariant dedt y ot iffer-e mou( rso LMPa in
et al., 2005; Warrenfeltz et al., 2004). These expression pro-files support the ideas expressed previously, that in some casesbenign and LMP tumours are not precursors of invasive tumours(Chenevix-Trench et al., 1997; Kurman and Trimble, 1993).
In order to compare tumour expression to that in OSE, oneneeds to obtain sufficient numbers of these cells. Since thetumour profile typically is generated from fresh frozen, totalresected mass, generating a comparable sample from normalOSE is difficult and whatever adaptation is chosen for compar-ison will make a fundamental difference to the genes identified(Zorn et al., 2003). One solution is laser-capture microscopy(LCM)-mediated microdissection of OSE and EOC cell popula-tions, combined with SAGE or microarray analysis to generatea list of candidates (Table 1), that are then validated by real-timePCR (Hough et al., 2000, 2001). Another solution is to cor-relate patterns of gene expression in the different histotypes ofEOC, with the normal tissues each histotype resembles. A recentprofiling study compared serous, mucinous, endometrioid andclear cell EOC with normal OSE brushings, as well as fallopiantube, endometrium and colon (Marquez et al., 2005). When com-pared with normal OSE, the changes in gene expression notedin serous EOC correlated with those found in fallopian tube,but not in other normal tissues. Similarly, differences betweenmucinous EOC and normal OSE correlated with those in normalcolon, and differences between both endometrioid and clear cellcancers and OSE correlated with those in normal endometrium( fyt d tot tanth
cul-t cells( nor-
umours, with the rational that the test is only really neeo identify the deadly advanced cases. Surprisingly, manhe expression and genomic profiles obtained identify no dntially expressed genes between early and late stage tuShridhar et al., 2001, 2002) and extremely small numbef coincidentally expressed genes, between tumours ofnd invasive tumours (Gilks et al., 2005; Meinhold-Heerle
f
rs
Marquez et al., 2005). This type of study begins to identihe specific molecular alterations and pathways that leahe different EOC histotypes, as well as identifying imporistotype-specific biomarkers (Table 1).
Alternatively, EOC-specific genes are sought within cellures, which can be obtained for both normal OSE and EOCEuer et al., 2005). Because of the problem of access to the

J.S. Fleming et al. / Molecular and Cellular Endocrinology 247 (2006) 4–21 13
Table 1Genes identified as up-regulated in EOC, and therefore of potential use as biomarkers, in a range of studies using microarray or SAGE techniques
Potential biomarkers Tissues/cells compared References
Apolipoprotein J/clusterin Primary cancers, ovarian surface epithelial cells andcystadenoma cells
Hough et al. (2000, 2001)Apolipoprotein E
Laser-capture cell microdissected EOC and SAGEanalysis
Ceruloplasmin
Claudin-3Claudin-4Epithelial cell adhesion molecule/GA733-2Folate receptor 1Glutathione peroxidase 3 (GPX3)HE4IGF-binding protein 2Kop protease inhibitorMatrix gla proteinMesothelinMucin-1S100A2Secretory leucocyte protease inhibitorSignal transducer and activator of transcription 1Tissue inhibitor of metalloproteinase 3
Cell adhesion molecule L1 CAM 11 ovarian tumour cell lines and 2 immortalisedOSE cell lines
Euer et al. (2005)Notch ligand Jagged 2Secreted polypeptide Neuromedin U
�8 integrin subunit 20 ovarian carcinomas, 17 ovarian carcinomasmetastatic to the omentum and 50 normal ovaries
Hibbs et al. (2004)Bone morphogenetic protein-7Cellular retinoic acid binding protein-1Claudin-4Collagen type IX�2Forkhead box J1S100 calcium-binding protein A1
Prostasin 3 ovarian cancer cell lines and from 3 normal humanOSE cell lines
Mok et al. (2001)
CD24 10 primary OSPC cell lines, 2 established OSPC celllines (UCI-101, UCI-107) and 5 primary normalOSE epithelial cultures
Santin et al. (2004)andShvartsman et al. (2003)Claudin-3Claudin-4Kallikrein 6 (protease M)Kallikrein 10Ladinin 1LamininLipocalin 2Matriptase (TADG-15)OsteopontinS100A2SERPIN2 (PAI-2)StratifinTumour-associated calcium signal transducers 1 and
2 (TROP-1/Ep-CAM, TROP-2)
Kallikrein 6 Epithelial ovarian carcinomas and normal ovariantissue
Welsh et al. (2003)Kallikrein 10
Trefoil factor 1 (TFF1) Normal OSE brushings used as reference standardMarquez et al. (2005)Anterior gradient 2 (AGR2) Mucinous EOC compared with normal colonLectin, galactoside-binding, soluble, 4 (LGALS4)Carcinoembryonic antigen-related cell adhesion
molecule 5 (CEACAM6)
Serous EOC compared with normal fallopian tube
Cathepsin E (CTSE)
Endometrioid and clear cell EOC compared withnormal endometrium

14 J.S. Fleming et al. / Molecular and Cellular Endocrinology 247 (2006) 4–21
mal controls, such studies depend on downstream immunohis-tochemical validation in primary material. L1CAM is expressedpredominantly in ovarian carcinomas, in borderline tumours toa lesser extent and very rarely in ovarian non-epithelial typesof cancer. Similarly, the potential markers found byHibbs et al.(2004)are all over-expressed in EOC (Table 1). Statistical anal-yses showed that the�8 integrin subunit, claudin-4 and S100A1provided the best distinction between ovarian carcinoma andnormal ovary and served as the best candidate tumour markersamong the seven genes identified in one study (Table 1) (Hibbset al., 2004).
Biomarkers may be present at higher levels in the cancertissue as a result of increased signalling of a specific pathway inthe tumour microenvironment (Donninger et al., 2004; Hough etal., 2001), or as a result of a specific chromosomal amplificationthat leads to selective advantage. In such a case, the primaryvalidation may use a combination of expression profiling withcomparative genome hybridization (CGH) (Shridhar et al., 2001;Israeli et al., 2005; Scorilas et al., 2003). Ostensibly, some typicalEOC markers may mark amplified genome loci, not becausethose genes are selectively advantageous, but rather because agene in their genomic vicinity is also amplified. For example,folate receptor, whose selective advantage to EOC has long beensought (Bottero et al., 1993), resides in the centre of an ampliconnext to Cyclin D1 (Shridhar et al., 2001), whose advantage togrowth is easier to explain (Hanahan and Weinberg, 2000).
2 eta ta eo d aa gnac say’s then eduS corrF re ur theiu
OCm ner.S renk 04a
inso them ssiop OCm
OCa ypeso t al.,2 sedg phoo latem SE
but are expressed in different EOC subtypes according to the pat-tern of mullerian-like differentiation of these cancers. Ectopicexpression of Hoxa9 in tumorigenic mouse OSE cells givesrise to papillary tumours resembling serous EOC (Cheng et al.,2005). In contrast, Hoxa10 and Hoxa11 induced morphogene-sis of endometrioid-like and mucinous-like EOC, respectively.Hoxa7 showed no lineage specificity, but promoted the abilitiesof Hoxa9, Hoxa10 and Hoxa11 to induce differentiation alongtheir respective pathways. Therefore, inappropriate activation ofa molecular programme that controls patterning of the reproduc-tive tract could explain the morphologic heterogeneity of EOCand the assumption of mullerian-like features. However, theseHOX genes do not appear to be differentially expressed in clearcell, mucinous, and serous EOC expression profiles (Ono et al.,2000; Zheng et al., 2004; Schwartz et al., 2002). Detecting suchtranscription activators in expression profiles is statistically dif-ficult, because such a low amount of expression is required forthem to convey their corresponding biological effects. Indeed,no such histotype-specific genes were identified in the studycomparing serous, mucinous, endometrioid and clear cell EOCwith normal colon, fallopian tube or endometrium (Marquez etal., 2005).
Some EOC expression profiling studies have offered insightinto ovarian cancer epidemiology. Although, as discussed, pro-gesterone exposure appears to lower risk of EOC, little isknown of its mechanism of action (Ho, 2003; Risch, 1998).A enesw -t Am nsis-t celll linesb , acti-v iverc H2)w s inr
1
OCh f thec cellso n tom bringu sup-p cour-a ges int urredd uref isto-t EOCs e, byu ions.
logyo esise pic-
Other EOC putative markers include prostasin (Mok et al.,001), kallikrein KLK10 (Santin et al., 2004; Shvartsmanl., 2003), claudin-3, claudin-4, TROP-1 and CD24 (Santin el., 2004) and KLK6 (Welsh et al., 2003). Unfortunately, nonf these is observed in 100% of EOC samples tested anre frequently expressed to some degree in the non-malionditions, compromising the prospect of any potential aspecificity and sensitivity. This is a major problem, giveneed for an inexpensive assay as a routine screening procimply combining two inadequate marker assays tends to
upt rather than improve the utility of the test (Mills et al., 2001).urthermore, there is now good evidence some genes aegulated in a variety of cancers, which may compromisese as specific biomarkers for EOC (Welsh et al., 2003).
More specific assays may be found if the putative Earker is alternatively spliced in an EOC specific manuch alternative splice forms have been found for the diffeallikrein genes (Clements et al., 2004; Borgono et al., 20)nd TADG-12D (Sawasaki et al., 2004).
Alternatively, further understanding of the cellular origf the different types of EOC, combined with knowledge ofutations leading to EOC and the use of full genome exprerofiling arrays, may allow the identification of a bone fide Earker with the required specificity and sensitivity.The first attempt to use microarrays to gain insight into E
etiology involved comparison of the histopathological subtf EOC (Ono et al., 2000; Zheng et al., 2004; Schwartz e002). Most interestingly, some of the differentially expresenes agree with independent studies on the cellular morgy of EOC variants. HOX genes, which normally reguullerian duct differentiation, are not expressed in normal O
llnts
re.-
p-r
t
n
l-
,
recent study aimed to identify progesterone-regulated gith potential anti-EOC action (Syed et al., 2005). Transcrip
ional profiling of normal OSE and EOC cell lines with a cDNicroarray identified genes (1) whose expression was co
ently down-regulated in EOC cell lines compared to OSEines, and (2) whose expression was restored in EOC celly progesterone treatment. From the candidates selectedating transcription factor-3 (ATF-3), caveolin-1, deleted in lancer-1 (DLC-1), and non-metastatic clone 23 (NM23-ere re-expressed in normal levels in the tumour cell line
esponse to progesterone treatment (Syed et al., 2005).
0. Conclusions
Despite its huge potential, gene expression profiling of Eas been confounded by lack of clear understanding oellular origin and pathways to EOC. There may be severalf origin and precursor ovarian states, prior to transformatioalignancy. Although each new array experiment seems top a novel subset of new biomarkers and potential tumourressors, results of more recent gene profiling studies are enging, because they appear to be starting to identify chan
he expression of gene families and pathways that have occuring differentiation of the precursor cell to EOC. Our fut
ocus must be to more thoroughly classify the different hypes by their specific biomarkers and to compare eachubtype to a broader range of precursor cells, for examplsing LCM to provide pure reference standard cell populat
The four major hypotheses that arose from the epidemiof EOC may all be partially supported, but no one hypothxplains all we now know about ovarian carcinogenesis. A

J.S. Fleming et al. / Molecular and Cellular Endocrinology 247 (2006) 4–21 15
Fig. 4. Pathways to EOC are influenced by events at ovulation, including hormonal stimulation and inflammation. Post-ovulatory re-epithelialisation involves OSE(blue cells) cell division and possibly migration, to cover the ovulatory lesion (black arrows). Hyperplasia and transformation of the OSE to adenocarcinoma can occurdirectly (red arrow top left). The pre-ovulatory luteinising hormone surge induces increased expression of cytokines and invasion of macrophages and monocytes(orange cells), leading to differentiation of follicle cells into luteal cells. Ovulation stimulates formation of invaginations and the formationof inclusion cysts. Cystcells may differentiate to take on mullerian characteristics under the influence of hormonal or cytokine stimulation and become ciliated (yellow cells) or secretory(tan cells), as shown on the right hand side of the figure. Rete ovarii tubules at the hilus of the ovary, close to the mesothelium to OSE transition (M–E),also containciliated and secretory cells and can dilate to form cysts, at least in rodents. It is still not known whether cells in both cyst types can transform to become cancerous.Furthermore, the role of any proposed ovarian stem or progenitor cells (purple cells) in epithelial ovarian carcinogenesis remains to be elucidated. (For interpretationof the references to colour in this figure legend, the reader is referred to the web version of the article.)
ture is emerging of at least two pathways; a slower developmentof high grade EOC via transformation of the cells lining inclu-sion cysts (Type 1) or a more direct and aggressive developmentof invasive EOC, directly from the OSE or other epithelial source(Type 2;Fig. 4). The key to determining the origins of EOC isan understanding of the very early changes in these two types ofcarcinogenic pathway. Animal studies suggest cysts may formfrom the rete ovarii in some species, so it is possible cysts ofdifferent epithelial origin can form in older human ovaries. Stud-ies on the structure and function of the rete ovarii in the adulthuman are needed before this source of epithelial cell can betotally discounted in EOC. While it is clear incessant ovulationleads to increased rates of invagination and OSE entrapmentin the ovarian stroma (Tan et al., 2005), multiple pathways tocyst formation may explain some of the heterogeneity in thelarge group of serous cystadenocarcinomas. The possibility ofovarian stem cell involvement in EOC also should not be dis-counted. Further studies are needed to confirm the existenceand source of such stem cells and to characterise their responsesto ovulation, hormonal stimulation and possible involvement incarcinogenesis. Given the embryonic origin of the rete ovarii,the possibility that this tubule network is a source of such stemcells should also be investigated. Study of inclusion cyst for-mation under different environmental conditions and endocrinetreatments may also help determine the specific contributions ofoestrogens, androgens and progestins to inclusion cyst formationa ationo sionp us t
determine the molecular basis for transformation of cyst cells inType 1 EOC. Similarly, direct transformation of the OSE in vivo,taking advantage of the bursal capsule surrounding the ovary inthe rodent, may be beneficial in defining the molecular basis ofType 2 EOC. The rate of publication on use of whole genomeexpression profiling in this area has increased rapidly in parallelto access to the technology. Elucidation of the different path-ways to EOC and classification of the numerous subtypes of thisheterogeneous disease will require a true collaboration of funda-mental cell biology and histopathology with molecular genetics.
Acknowledgements
The authors are indebted to the technical expertise ofH. James McQuillan and Melanie Richardson (Departmentof Anatomy & Structural Biology, University of Otago,New Zealand). They also acknowledge the work of NaomiArmanasco (Department of Anatomy & Structural Biology,University of Otago, New Zealand) on the effects of talc onthe mouse ovary. Research on incessant ovulation of mouseovaries was supported by the New Zealand Lottery Grants Board(Health), the HS & JC Anderson Trust (Otago, New Zealand)and the University of Otago.
References
A son,risk
nd transformation of OSE and cyst cells. Incessant ovulf transgenic mice, in which genes identified from expresrofiling studies are over-expressed or deleted, may allow
odami, H.O., Hsieh, C.C., Lambe, M., Trichopoulos, D., Leon, D., PersI., Ekbom, A., Janson, P.O., 1994. Parity, age at first childbirth, andof ovarian cancer. Lancet 344, 1250–1254 (comment).

16 J.S. Fleming et al. / Molecular and Cellular Endocrinology 247 (2006) 4–21
Akhmedkhanov, A., Toniolo, P., Zeleniuch-Jacquotte, A., Kato, I., Koenig,K.L., Shore, R.E., 2001. Aspirin and epithelial ovarian cancer. Prev. Med.33, 682–687.
Allen, E., 1923. Ovogenesis during sexual maturity. Am. J. Anat. 31,439–481.
Altinoz, M.A., Korkmaz, R., 2004. NF-kappaB, macrophage migrationinhibitory factor and cyclooxygenase-inhibitions as likely mechanismsbehind the acetaminophen- and NSAID-prevention of the ovarian cancer.Neoplasma 51, 239–247.
Alvarez, A.A., Krigman, H.R., Whitaker, R.S., Dodge, R.K., Rodriguez, G.C.,1999. The prognostic significance of angiogenesis in epithelial ovariancarcinoma. Clin. Cancer Res. 5, 587–591.
Andersen, C.Y., Hornnes, P., 1994. Intrafollicular concentrations of free cor-tisol close to follicular rupture. Hum. Reprod. 9, 1944–1949.
Anderson, E., Clarke, R.B., 1999. Epithelial stem cells in the mammarygland: casting light into dark corners. Breast Cancer Res. 1, 11–13.
Antoniou, A., Pharoah, P.D., Narod, S., Risch, H.A., Eyfjord, J.E., Hopper,J.L., Loman, N., Olsson, H., Johannsson, O., Borg, A., Pasini, B., Radice,P., Manoukian, S., Eccles, D.M., Tang, N., Olah, E., Anton-Culver, H.,Warner, E., Lubinski, J., Gronwald, J., Gorski, B., Tulinius, H., Thor-lacius, S., Eerola, H., Nevanlinna, H., Syrjakoski, K., Kallioniemi, O.P.,Thompson, D., Evans, C., Peto, J., Lalloo, F., Evans, D.G., Easton, D.F.,2003. Average risks of breast and ovarian cancer associated with BRCA1or BRCA2 mutations detected in case Series unselected for family his-tory: a combined analysis of 22 studies. Am. J. Hum. Genet. 72, 1117–1130.
Archbald, L.F., Schultz, R.H., Fahning, M.L., Kurtz, H.J., Zemjanis, R., 1971.Rete ovarii in heifers: a preliminary study. J. Reprod. Fertil. 26, 413–414.
Auersperg, N., Maines Bandiera, S.L., Dyck, H.G., 1997. Ovarian carcino-genesis and the biology of ovarian surface epithelium. J. Cell. Physiol.173, 261–265.
A var-docr.
A ona
B osia,rolif-
B rian
B T.J.,AM-man
B .D.,can-1991
B ouseesis,
p.B od.
B y of
B alig-
B en-en,roneucedmiol.
B , E.,uta-
ested
Bonello, N., McKie, K., Jasper, M., Andrew, L., Ross, N., Braybon, E.,Brannstrom, M., Norman, R.J., 1996. Inhibition of nitric oxide: effects oninterleukin-1 beta-enhanced ovulation rate, steroid hormones, and ovarianleukocyte distribution at ovulation in the rat. Biol. Reprod. 54, 436–445.
Borgono, C.A., Michael, I.P., Diamandis, E.P., 2004. Human tissuekallikreins: physiologic roles and applications in cancer. Mol. CancerRes. 2, 257–280.
Bottero, F., Tomassetti, A., Canevari, S., Miotti, S., Menard, S., Colnaghi,M.I., 1993. Gene transfection and expression of the ovarian carcinomamarker folate binding protein on NIH/3T3 cells increases cell growth invitro and in vivo. Cancer Res. 53, 5791–5796.
Bu, S.Z., Yin, D.L., Ren, X.H., Jiang, L.Z., Wu, Z.J., Gao, Q.R., Pei, G.,1997. Progesterone induces apoptosis and up-regulation of p53 expressionin human ovarian carcinoma cell lines. Cancer 79, 1944–1950.
Bukovsky, A., Caudle, M.R., Svetilkova, M., Upadhyaya, N.B., 2004. For-mation of new primary follicles in adult human ovaries. Reprod. Biol.Endocrinol. 2, 20.
Burke, F., Relf, M., Negus, R., Balkwill, F., 1996. A cytokine profile ofnormal and malignant ovary. Cytokine 8, 578–585.
Byskov, A.G., 1978. The anatomy and ultrastructure of the rete system inthe fetal mouse ovary. Biol. Reprod. 19, 720–735.
Byskov, A.G., Lintern-Moore, S., 1973. Follicle formation in the immaturemouse ovary: the role of the rete ovarii. J. Anat. 116, 207–217.
Byskov, A.G., Skakkebaek, N.E., Stafanger, G., Peters, H., 1977. Influence ofovarian surface epithelium and rete ovarii on follicle formation. J. Anat.123, 77–86.
Campbell, I.G., Russell, S.E., Choong, D.Y., Montgomery, K.G., Ciavarella,M.L., Hooi, C.S., Cristiano, B.E., Pearson, R.B., Phillips, W.A., 2004.Mutation of the PIK3CA gene in ovarian and breast cancer. Cancer Res.64, 7678–7681.
C erson,173.
C , D.L.,heep52,
C erk,imal
C , J.,eptors. 61,
C Riskiol.
C n, L.,J.A.,Choi,ozy-gical
C delityecify
C C.S.,teroid
C ards,sengerran-e
C 11,
C ssuedis-
uersperg, N., Wong, A.S., Choi, K.C., Kang, S.K., Leung, P.C., 2001. Oian surface epithelium: biology, endocrinology, and pathology. EnRev. 22, 255–288.
ustin, C., Short, R., 1984. Oestrous and menstrual cycles. In: HormControl of Reproduction. Cambridge University Press, Cambridge.
ai, W., Oliveros-Saunders, B., Wang, Q., Acevedo-Duncan, M.E., NicS.V., 2000. Estrogen stimulation of ovarian surface epithelial cell peration. In Vitro Cell. Dev. Biol. 36, 657–666.
aker, T.R., Piver, M.S., 1994. Etiology, biology, and epidemiology of ovacancer. Semin. Surg. Oncol. 10, 242–248.
anks, R.E., Gearing, A.J., Hemingway, I.K., Norfolk, D.R., Perren,Selby, P.J., 1993. Circulating intercellular adhesion molecule-1 (IC1), E-selectin and vascular cell adhesion molecule-1 (VCAM-1) in humalignancies. Br. J. Cancer 68, 122–124.
eard, C.M., Hartmann, L.C., Atkinson, E.J., O’Brien, P.C., Malkasian, GKeeney, G.L., Melton 3rd, L.J., 2000. The epidemiology of ovariancer: a population-based study in Olmsted County, Minnesota, 1935–Ann. Epidemiol. 10, 14–23.
eaugie, C., 2005. Changes with age and ovulation number in the movarian surface epithelium, inclusion cysts and rete ovarii. M.Sc. ThAnatomy & Structural Biology, University of Otago, Dunedin, 165 p
ell, D.A., 2005. Origins and molecular pathology of ovarian cancer. MPathol. 18, S19–S32.
ell, D.A., Scully, R.E., 1994. Early de novo ovarian carcinoma. A studfourteen cases. Cancer 73, 1859–1864.
eltsos, A.N., Odem, R.R., 1996. Ovulation induction and ovarian mnancy. Semin. Reprod. Endocrinol. 14, 367–374.
erchuck, A., Schildkraut, J.M., Wenham, R.M., Calingaert, B., Ali, S., Hriott, A., Halabi, S., Rodriguez, G.C., Gertig, D., Purdie, D.M., KelemL., Spurdle, A.B., Marks, J., Chenevix-Trench, G., 2004. Progestereceptor promoter +331A polymorphism is associated with a redrisk of endometrioid and clear cell ovarian cancers. Cancer EpideBiomarkers Prev. 13, 2141–2147.
jorge, T., Lie, A.K., Hovig, E., Gislefoss, R.E., Hansen, S., JellumLangseth, H., Nustad, K., Trope, C.G., Dorum, A., 2004. BRCA1 mtions in ovarian cancer and borderline tumours in Norway: a ncase–control study. Br. J. Cancer 91, 1829–1834.
l
.
asagrande, J.T., Louie, E.W., Pike, M.C., Roy, S., Ross, R.K., HendB.E., 1979. Incessant ovulation and ovarian cancer. Lancet 2, 170–
assali, G.D., Nogueira, J.C., Nascimento, E.F., Cardosa, J.S., Ferreira2000. Morphological and pathological aspects of the rete ovarii in s(Ovis aries). Arquivo Brasileiro de Medicina Veterinaria e Zootecnia47–52.
elik, C., Gezginc, K., Aktan, M., Acar, A., Yaman, S., Gungor, S., AkyuC., 2004. Effects of ovulation induction on ovarian morphology: an anstudy. Int. J. Gynaecol. Cancer 14, 600–606.
hen, T., Triplett, J., Dehner, B., Hurst, B., Colligan, B., PembertonGraff, J.R., Carter, J.H., 2001. Transforming growth factor-beta rectype I gene is frequently mutated in ovarian carcinomas. Cancer Re4679–4682.
hen, Y., Wu, P.C., Lang, J.H., Ge, W.J., Hartge, P., Brinton, L.A., 1992.factors for epithelial ovarian cancer in Beijing, China. Int. J. Epidem21, 23–29.
henevix-Trench, G., Kerr, J., Hurst, T., Shih, Y.C., Purdie, D., BergmaFriedlander, M., Sanderson, B., Zournazi, A., Coombs, T., Leary,Crawford, E., Shelling, A.N., Cooke, I., Ganesan, T.S., Searle, J.,C., Barrett, J.C., Khoo, S.K., Ward, B., 1997. Analysis of loss of hetergosity and KRAS2 mutations in ovarian neoplasms: clinicopatholocorrelations. Genes Chromosomes Cancer 18, 75–83.
heng, W., Liu, J., Yoshida, H., Rosen, D., Naora, H., 2005. Lineage infiof epithelial ovarian cancers is controlled by HOX genes that spregional identity in the reproductive tract. Nat. Med. 11, 531–537.
larke, R.B., Spence, K., Anderson, E., Howell, A., Okano, H., Potten,2005. A putative human breast stem cell population is enriched for sreceptor-positive cells. Dev. Biol. 277, 443–456.
lemens, J.W., Robker, R.L., Kraus, W.L., Katzenellenbogen, B.S., RichJ.S., 1998. Hormone induction of progesterone receptor (PR) mesribonucleic acid and activation of PR promoter regions in ovarian gulosa cells: evidence for a role of cyclic adenosine 3′,5′-monophosphatbut not estradiol. Mol. Endocrinol. 12, 1201–1214.
lement, P.B., 1987. Histology of the ovary. Am. J. Surg. Pathol.277–303.
lements, J.A., Willemsen, N.M., Myers, S.A., Dong, Y., 2004. The tikallikrein family of serine proteases: functional roles in human

J.S. Fleming et al. / Molecular and Cellular Endocrinology 247 (2006) 4–21 17
ease and potential as clinical biomarkers. Crit. Rev. Clin. Lab. Sci. 41,265–312.
Clow, O.L., Hurst, P.R., Fleming, J.S., 2002. Changes in the mouse ovariansurface epithelium with age and ovulation number. Mol. Cell. Endocrinol.191, 105–111.
Cohen, I., Figer, A., Tepper, R., Shapira, J., Altaras, M.M., Yigael, D.,Beyth, Y., 1999. Ovarian overstimulation and cystic formation in pre-menopausal tamoxifen exposure: comparison between tamoxifen-treatedand nontreated breast cancer patients. Gynecol. Oncol. 72, 202–207.
Cohen, I., Potlog-Nahari, C., Shapira, J., Yigael, D., Tepper, R., 2003.Simple ovarian cysts in postmenopausal patients with breast carci-noma treated with tamoxifen: long-term follow-up. Radiology 227, 844–848.
Cramer, D.W., Welch, W.R., 1983. Determinants of ovarian cancer risk. II.Inferences regarding pathogenesis. J. Natl. Cancer. Inst. 71, 717–721.
Curiel, T.J., Coukos, G., Zou, L., Alvarez, X., Cheng, P., Mottram, P.,Evdemon-Hogan, M., Conejo-Garcia, J.R., Zhang, L., Burow, M., Zhu,Y., Wei, S., Kryczek, I., Daniel, B., Gordon, A., Myers, L., Lackner, A.,Disis, M.L., Knutson, K.L., Chen, L., Zou, W., 2004. Specific recruitmentof regulatory T cells in ovarian carcinoma fosters immune privilege andpredicts reduced survival. Nat. Med. 10, 942–949.
Czernobilsky, B., Moll, R., Levy, R., Franke, W.W., 1985. Co-expressionof cytokeratin and vimentin filaments in mesothelial, granulosa and reteovarii cells of the human ovary. Eur. J. Cell Biol. 37, 175–190.
Daly, M., Obrams, G.I., 1998. Epidemiology and risk assessment for ovariancancer. Semin. Oncol. 25, 255–264.
Deligdisch, L., Gil, J., 1989. Characterization of ovarian dysplasia by inter-active morphometry. Cancer 63, 748–755.
Ding, W., Tang, Q., Espina, V., Liotta, L.A., Mauger, D.T., Mulder, K.M.,2005. A transforming growth factor-beta receptor-interacting protein fre-quently mutated in human ovarian cancer. Cancer Res. 65, 6526–6533.
D rady,ssiontivate
D thethes
E ancetudy.–294
E variaer 86
E 1. K-rarelthol.
E morortiso
E .H.,rian
F lasia?
F alig-
F ours
F Per-
G , E.,new
r des
G . Vet.
Gilks, C.B., Vanderhyden, B.C., Zhu, S., van de Rijn, M., Longacre, T.A.,2005. Distinction between serous tumours of low malignant potential andserous carcinomas based on global mRNA expression profiling. Gynecol.Oncol. 96, 684–694.
Goedegebuure, P.S., Douville, C.C., Doherty, J.M., Linehan, D.C., Lee, K.Y.,Ganguly, E.K., Eberlein, T.J., 1997. Simultaneous production of T helper-1-like cytokines and cytolytic activity by tumor-specific T cells in ovarianand breast cancer. Cell. Immunol. 175, 150–156.
Gondos, B., 1975. Surface epithelium of the developing ovary. Am. J. Pathol.81, 303–320.
Greer, J.B., Modugno, F., Allen, G.O., Ness, R.B., 2005. Short-term oral con-traceptive use and the risk of epithelial ovarian cancer. Am. J. Epidemiol.162, 66–72.
Gwinn, M.L., Lee, N.C., Rhodes, P.H., Layde, P.M., Rubin, G.L., 1990. Preg-nancy, breast feeding, and oral contraceptives and the risk of epithelialovarian cancer. J. Clin. Epidemiol. 43, 559–568.
Hamilton, T.C., Fox, H., Buckley, C.H., Henderson, W.J., Griffiths, K., 1984.Effects of talc on the rat ovary. Br. J. Exp. Pathol. 65, 101–106.
Hanahan, D., Weinberg, R.A., 2000. The hallmarks of cancer. Cell 100,57–70.
Hardiman, P., Nieto, J.J., MacLean, A.B., 2000. Infertility and ovarian cancer.Gynecol. Oncol. 76, 1–2.
Harris, R., Whittemore, A.S., Itnyre, J., 1992. Characteristics relating to ovar-ian cancer risk: collaborative analysis of 12 US case–control studies. III.Epithelial tumours of low malignant potential in white women. Collabo-rative Ovarian Cancer Group. Am. J. Epidemiol. 136, 1204–1211.
Heatley, M.K., 2000. Adenomatous hyperplasia of the rete ovarii. Histopathol-ogy 36, 383–384.
Heller, D., Westhoff, C., Gordon, R., Katz, N., 1996. The relationship betweenperineal cosmetic talc usage and ovarian talc particle burden. Am. J.Obstet. Gynaecol. 174, 1507–1510.
H ths,gina247–
H .M.,A.P.,n of
H ids
H uta-rous
H eprod.
H .,der-clini-
H P.J.,es. 61,
H osen-serial
ovar-
H 005.nim.
H puta-27,
I 002.Nat.
I .,ated
onninger, H., Bonome, T., Radonovich, M., Pise-Masison, C.A., BJ., Shih, J.H., Barrett, J.C., Birrer, M.J., 2004. Whole genome expreprofiling of advance stage papillary serous ovarian cancer reveals acpathways. Oncogene 23, 8065–8077.
ubeau, L., 1999. The cell of origin of ovarian epithelial tumours andovarian surface epithelium dogma: does the emperor have no cloGynecol. Oncol. 72, 437–442.
aston, D.F., Matthews, F.E., Ford, D., Swerdlow, A.J., Peto, J., 1996. Cmortality in relatives of women with ovarian cancer: the OPCS SOffice of Population Censuses and Surveys. Int. J. Cancer. 65, 284
dmondson, R.J., Monaghan, J.M., Davies, B.R., 2002. The human osurface epithelium is an androgen responsive tissue. Br. J. Canc879–885.
nomoto, T., Weghorst, C.M., Inoue, M., Tanizawa, O., Rice, J.M., 199ras activation occurs frequently in mucinous adenocarcinomas andin other common epithelial tumours of the human ovary. Am. J. Pa139, 777–785.
scher, G., Galli, I., Vishwanath, B.S., Frey, B.M., Frey, F.J., 1997. Tunecrosis factor alpha and interleukin 1beta enhance the cortisone/cshuttle. J. Exp. Med. 186, 189–198.
uer, N.I., Kaul, S., Deissler, H., Mobus, V.J., Zeillinger, R., Weidle, U2005. Identification of L1 CAM, Jagged2 and Neuromedin U as ovacancer-associated antigens. Oncol. Rep. 13, 375–387.
athalla, M.F., 1971. Incessant ovulation—a factor in ovarian neopLancet 2, 163.
eeley, K.M., Wells, M., 2001. Precursor lesions of ovarian epithelial mnancy. Histopathology 38, 87–95.
oyle, A., Al-Jabi, M., McCaughey, W.T., 1981. Papillary peritoneal tumin women. Am. J. Surg. Pathol. 5, 241–249.
redrickson, T.N., 1987. Ovarian tumours of the hen. Environ. Healthspect. 73, 35–51.
aspard, U.J., Romus, M.A., Gillain, D., Duvivier, J., Demey-PonsartFranchimont, P., 1983. Plasma hormone levels in women receivingoral contraceptives containing ethinyl estradiol plus levonorgestrel oogestrel. Contraception 27, 577–590.
elberg, H.B., McEntee, K., Heath, E.H., 1984. Feline cystic rete ovariiPathol. 21, 304–307.
d
?
r
.n,
y
l
-
enderson, W.J., Hamilton, T.C., Baylis, M.S., Pierrepoint, C.G., GriffiK., 1986. The demonstration of the migration of talc from the vaand posterior uterus to the ovary in the rat. Environ. Res. 40,250.
ibbs, K., Skubitz, K.M., Pambuccian, S.E., Casey, R.C., Burleson, KOegema Jr., T.R., Thiele, J.J., Grindle, S.M., Bliss, R.L., Skubitz,2004. Differential gene expression in ovarian carcinoma: identificatiopotential biomarkers. Am. J. Pathol. 165, 397–414.
illier, S.G., Tetsuka, M., 1998. An anti-inflammatory role for glucocorticoin the ovaries? J. Reprod. Immunol. 39, 21–27.
o, C.L., Kurman, R.J., Dehari, R., Wang, T.L., Shih Ie, M., 2004. Mtions of BRAF and KRAS precede the development of ovarian seborderline tumours. Cancer Res. 64, 6915–6918.
o, S.M., 2003. Estrogen, progesterone and epithelial ovarian cancer. RBiol. Endocrinol. 1, 73.
oriuchi, A., Itoh, K., Shimizu, M., Nakai, I., Yamazaki, T., Kimura, KSuzuki, A., Shiozawa, I., Ueda, N., Konishi, I., 2003. Toward unstanding the natural history of ovarian carcinoma development: acopathological approach. Gynecol. Oncol. 88, 309–317.
ough, C.D., Cho, K.R., Zonderman, A.B., Schwartz, D.R., Morin,2001. Coordinately up-regulated genes in ovarian cancer. Cancer R3869–3876.
ough, C.D., Sherman-Baust, C.A., Pizer, E.S., Montz, F.J., Im, D.D., Rshein, N.B., Cho, K.R., Riggins, G.J., Morin, P.J., 2000. Large-scaleanalysis of gene expression reveals genes differentially expressed inian cancer. Cancer Res. 60, 6281–6287.
unter, M., Quinn, R., Ferguson, E., Edwards, S., Ashworth, C., 2Oocyte-somatic cell-endocrine interactions in pigs. Domest. AEndocrinol. 29, 371–384.
utson, R., Ramsdale, J., Wells, M., 1995. p53 protein expression intive precursor lesions of epithelial ovarian cancer. Histopathology367–371.
hmels, J., Friedlander, G., Bergmann, S., Sarig, O., Ziv, Y., Barkai, N., 2Revealing modular organization in the yeast transcriptional network.Genet. 31, 370–377.
nal, M.M., Incebiyik, A., Sanci, M., Yildirim, Y., Polat, M., Pilanci, BNayki, C., Camuzcuoglu, H., 2005. Ovarian cysts in tamoxifen-tre

18 J.S. Fleming et al. / Molecular and Cellular Endocrinology 247 (2006) 4–21
women with breast cancer. Eur. J. Obstet. Gynecol. Reprod. Biol. 120,104–106.
Israeli, O., Goldring-Aviram, A., Rienstein, S., Ben-Baruch, G., Korach, J.,Goldman, B., Friedman, E., 2005. In silico chromosomal clustering ofgenes displaying altered expression patterns in ovarian cancer. CancerGenet. Cytogenet. 160, 35–42.
Ivarsson, K., Sundfeldt, K., Brannstrom, M., Janson, P.O., 2001. Productionof steroids by human ovarian surface epithelial cells in culture: possiblerole of progesterone as growth inhibitor. Gynecol. Oncol. 82, 116–121.
Jazaeri, A.A., Awtrey, C.S., Chandramouli, G.V., Chuang, Y.E., Khan, J.,Sotiriou, C., Aprelikova, O., Yee, C.J., Zorn, K.K., Birrer, M.J., Barrett,J.C., Boyd, J., 2005. Gene expression profiles associated with responseto chemotherapy in epithelial ovarian cancers. Clin. Cancer Res. 11,6300–6310.
Jemal, A., Murray, T., Ward, E., Samuels, A., Tiwari, R.C., Ghafoor, A.,Feuer, E.J., Thun, M.J., 2005. Cancer statistics. CA Cancer J. Clin. 55,10–30.
Jiang, X., Morland, S.J., Hitchcock, A., Thomas, E.J., Campbell, I.G., 1998.Allelotyping of endometriosis with adjacent ovarian carcinoma revealsevidence of a common lineage. Cancer Res. 58, 1707–1712.
Johnson, J., Bagley, J., Skaznik-Wikiel, M., Lee, H.-J., Adams, G.B., Niikura,Y., Tschudy, K.S., Canning-Tilly, J., Cortes, M.L., Forkert, R., Spitzer, T.,Iacomini, J., Scadden, D.T., Tilly, J.L., 2005. Oocyte generation in adultmammalian ovaries by putative germ cells in bone marrow and peripheralblood. Cell 122, 303–315.
Johnson, J., Canning, J., Kaneko, T., Pru, J.K., Tilly, J.L., 2004. Germ-linestem cells and follicular renewal in the postnatal mammalian ovary. Nature428, 145–150 (see comment, erratum appears in Nature 430 (August(7003)) (2004) 1062).
Karlan, B.Y., Jones, J., Greenwald, M., Lagasse, L.D., 1995. Steroid hormoneeffects on the proliferation of human ovarian surface epithelium in vitro.
K cep-nd in
K onserousfferen
K f lowl. 12
L noni,h. Int
L Bail-
L case
L , X.,.M.,nts ofrom
L ewal
L lop-8.
L oto,owthr Res
M lled
M nbu,et-
arian4–40
Mantovani, A., Bussolino, F., Dejana, E., 1992. Cytokine regulation ofendothelial cell function. FASEB J. 6, 2591–2599.
Marquez, R.T., Baggerly, K.A., Patterson, A.P., Liu, J., Broaddus, R., Fru-movitz, M., Atkinson, E.N., Smith, D.I., Hartmann, L., Fishman, D.,Berchuck, A., Whitaker, R., Gershenson, D.M., Mills, G.B., Bast Jr.,R.C., Lu, K.H., 2005. Patterns of gene expression in different histotypesof epithelial ovarian cancer correlate with those in normal fallopian tube,endometrium, and colon. Clin. Cancer. Res. 11, 6116–6126.
McCluggage, W.G., Weir, P.E., 2000. Paraovarian cystic endosalpingiosis inassociation with tamoxifen therapy. J. Clin. Pathol. 53, 161–162.
McGonigle, K.F., Vasilev, S.A., Odom-Maryon, T., Simpson, J.F., 1999. Ovar-ian histopathology in breast cancer patients receiving tamoxifen. Gynecol.Oncol. 73, 402–406.
Mckay, D.G., 1962. The origins of ovarian tumours. Clin. Obstet. Gynecol.5, 1181–1198.
McNatty, K.P., Makris, A., DeGrazia, C., Osathanondh, R., Ryan, K.J., 1979.The production of progesterone, androgens, and estrogens by granulosacells, thecal tissue, and stromal tissue from human ovaries in vitro. J.Clin. Endocrinol. Metab. 49, 687–699.
Meinhold-Heerlein, I., Bauerschlag, D., Hilpert, F., Dimitrov, P., Sapinoso,L.M., Orlowska-Volk, M., Bauknecht, T., Park, T.W., Jonat, W., Jacob-sen, A., Sehouli, J., Luttges, J., Krajewski, M., Krajewski, S., Reed, J.C.,Arnold, N., Hampton, G.M., 2005. Molecular and prognostic distinctionbetween serous ovarian carcinomas of varying grade and malignant poten-tial. Oncogene 24, 1053–1065.
Merogi, A.J., Marrogi, A.J., Ramesh, R., Robinson, W.R., Fermin, C.D., Free-man, S.M, 1997. Tumor-host interaction: analysis of cytokines, growthfactors, and tumor-infiltrating lymphocytes in ovarian carcinomas. Hum.Pathol. 28, 321–331.
Mills, J.C., Roth, K.A., Cagan, R.L., Gordon, J.I., 2001. DNA microarraysand beyond: completing the journey from tissue to cell. Nat. Cell Biol.
M ides,86,
M 993.eneo-
M ller,cker,arity,s and45,
M and
M uto,nco-ncy.
M itz,varianInst.
M .F.,f twoncer
M y ofiol-62–
M ma,rianr 79,
M rian
Am. J. Obstet. Gynecol. 173, 97–104.han, M.S., Dodson, A.R., Heatley, M.K., 1999. Ki-67, oestrogen re
tor, and progesterone receptor proteins in the human rete ovarii aendometriosis. J. Clin. Pathol. 52, 517–520.
orner, M., Burckhardt, E., Mazzucchelli, L., 2005. Different proportiof aneusomic cells in ovarian inclusion cysts associated with sborderline tumours and serous high-grade carcinomas support dipathogenetic pathways. J. Pathol. 207, 20–26.
urman, R.J., Trimble, C.L., 1993. The behavior of serous tumours omalignant potential: are they ever malignant? Int. J. Gynecol. Patho120–127.
a Vecchia, C., Franceschi, S., Gallus, G., Decarli, A., Liberati, A., TogG., 1983. Incessant ovulation and ovarian cancer: a critical approacJ. Epidemiol. 12, 161–164.
and, J.A., 1993. Ovulation, ovulation induction and ovarian carcinoma.lieres Clin. Obstet. Gynaecol. 7, 455–472.
ee, E., Kim, D.-S., Kim, I., 2001. Cystadenoma of the rete ovarii—areport. Korean J. Pathol. 35, 555–557.
i, Y., Welm, B., Podsypanina, K., Huang, S., Chamorro, M., ZhangRowlands, T., Egeblad, M., Cowin, P., Werb, Z., Tan, L.K., Rosen, JVarmus, H.E., 2003. Evidence that transgenes encoding componethe Wnt signaling pathway preferentially induce mammary cancersprogenitor cells. Proc. Natl. Acad. Sci. U.S.A. 100, 15853–15858.
iu, S., Dontu, G., Wicha, M.S., 2005. Mammary stem cells, self-renpathways, and carcinogenesis. Breast Cancer Res. 7, 86–95.
ochter, A., 1998. Plasticity of mammary epithelia during normal devement and neoplastic progression. Biochem. Cell Biol. 76, 997–100
ynch, M.A., Nakashima, R., Song, H., DeGroff, V.L., Wang, D., EnomT., Weghorst, C.M., 1998. Mutational analysis of the transforming grfactor beta receptor type II gene in human ovarian carcinoma. Cance58, 4227–4232.
alik, S., Balkwill, F., 1991. Epithelial ovarian cancer: a cytokine propedisease? Br. J. Cancer. 64, 617–620.
andai, M., Konishi, I., Kuroda, H., Komatsu, T., Yamamoto, S., NaK., Matsushita, K., Fukumoto, M., Yamabe, H., Mori, T., 1998. Herogeneous distribution of K-ras-mutated epithelia in mucinous ovtumours with special reference to histopathology. Hum. Pathol. 29, 3
t
,
.
f
.
.
3, E175–E178.ilner, J., Ponder, B., Hughes-Davies, L., Seltmann, M., Kouzar
T., 1997. Transcriptional activation functions in BRCA2. Nature 3772–773.
ittal, K.R., Zeleniuch-Jacquotte, A., Cooper, J.L., Demopoulos, R.I., 1Contralateral ovary in unilateral ovarian carcinoma: a search for prplastic lesions. Int. J. Gynecol. Pathol. 12, 59–63.
odan, B., Hartge, P., Hirsh-Yechezkel, G., Chetrit, A., Lubin, F., BeU., Ben-Baruch, G., Fishman, A., Menczer, J., Ebbers, S.M., TuM.A., Wacholder, S., Struewing, J.P., Friedman, E., Piura, B., 2001. Poral contraceptives, and the risk of ovarian cancer among carriernoncarriers of a BRCA1 or BRCA2 mutation. N. Engl. J. Med. 3235–240.
ohle, J., Whittemore, A., Pike, M., Darby, S., 1985. Gonadotrophinsovarian cancer risk. J. Natl. Cancer Inst. 75, 178–180.
ok, S.C., Bell, D.A., Knapp, R.C., Fishbaugh, P.M., Welch, W.R., MM.G., Berkowitz, R.S., Tsao, S.W., 1993. Mutation of K-ras protoogene in human ovarian epithelial tumours of borderline malignaCancer Res. 53, 1489–1492.
ok, S.C., Chao, J., Skates, S., Wong, K., Yiu, G.K., Muto, M.G., BerkowR.S., Cramer, D.W., 2001. Prostasin, a potential serum marker for ocancer: identification through microarray technology. J. Natl. Cancer93, 1458–1464.
oorman, P.G., Schildkraut, J.M., Calingaert, B., Halabi, S., Vine, MBerchuck, A., 2002. Ovulation and ovarian cancer: a comparison omethods for calculating lifetime ovulatory cycles (United States). CaCauses Control 13, 807–811.
otta, P.M., Van Blerkom, J., 1980. Scanning electron microscopthe mammalian ovary. In: Motta, P.M., Hafez, E.S.E. (Eds.), Bogy of the Ovary. Martinus Nigjhoff Publishers, The Hague, pp. 1175.
ourits, M.J., de Vries, E.G., Willemse, P.H., ten Hoor, K.A., HolleH., Sluiter, W.J., de Bruijn, H.W., van der Zee, A.G., 1999. Ovacysts in women receiving tamoxifen for breast cancer. Br. J. Cance1761–1764.
ulligan, R., 1976. A survey of epithelial inclusion cysts in the ovacortex of 470 patients. J. Surg. Oncol. 8, 61–66.

J.S. Fleming et al. / Molecular and Cellular Endocrinology 247 (2006) 4–21 19
Murdoch, W.J., 1994. Ovarian surface epithelium during ovulatory and anovu-latory ovine oestrous cycles. Anat. Rec. 240, 322–326.
Murdoch, W.J., 1995. Programmed cell death in preovulatory ovine follicles.Biol. Reprod. 53, 8–12.
Murdoch, W.J., Van Kirk, E.A., 2002. Steroid hormonal regulation of prolif-erative, p53 tumor suppressor, and apoptotic responses of sheep ovariansurface epithelial cells. Mol. Cell. Endocrinol. 186, 61–67.
Murdoch, W.J., Wilken, C., Young, D.A., 1999. Sequence of apoptosis andinflammatory necrosis within the formative ovulatory site of sheep folli-cles. J. Reprod. Fertil. 117, 325–329.
Naora, H., 2005. Developmental patterning in the wrong context: the paradoxof epithelial ovarian cancers. Cell Cycle 4, 1033–1035.
Nasca, P.C., Greenwald, P., Chorost, S., Richart, R., Caputo, T., 1984. An epi-demiologic case–control study of ovarian cancer and reproductive factors.Am. J. Epidemiol. 119, 705–713.
Negri, E., Franceschi, S., Tzonou, A., Booth, M., La Vecchia, C., Parazzini,F., Beral, V., Boyle, P., Trichopoulos, D., 1991. Pooled analysis of 3 Euro-pean case–control studies: I. Reproductive factors and risk of epithelialovarian cancer. Int. J. Cancer 49, 50–56.
Negus, R.P., Stamp, G.W., Hadley, J., Balkwill, F.R., 1997. Quantitativeassessment of the leukocyte infiltrate in ovarian cancer and its relationshipto the expression of C–C chemokines. Am. J. Pathol. 150, 1723–1734.
Negus, R.P., Stamp, G.W., Relf, M.G., Burke, F., Malik, S.T., Bernasconi,S., Allavena, P., Sozzani, S., Mantovani, A., Balkwill, F.R., 1995. Thedetection and localization of monocyte chemoattractant protein-1 (MCP-1) in human ovarian cancer. J. Clin. Invest. 95, 2391–2396.
Ness, R.B., Cottreau, C., 1999. Possible role of ovarian epithelial inflamma-tion in ovarian cancer. J. Natl. Cancer Inst. 91, 1459–1467.
Ness, R.B., Grisso, J.A., Cottreau, C., Klapper, J., Vergona, R., Wheeler, J.E.,Morgan, M., Schlesselman, J.J., 2000. Factors related to inflammationof the ovarian epithelium and risk of ovarian cancer. Epidemiology 11,
N enitalathol
N rtin,3.
O , G.,tionsours.
O e and
O arianbility.
O , A.,NAs. 60
P gene58,
P inter-
P erlein34.
P d ovags. J
P urreyP traep
oma
P , M.,ctorstudy.4.
Purdie, D.M., Bain, C.J., Siskind, V., Webb, P.M., Green, A.C., 2003. Ovu-lation and risk of epithelial ovarian cancer. Int. J. Cancer 104, 228–232.
Radisavljevic, S.V., 1977. The pathogenesis of ovarian inclusion cysts andcystomas. Obstet. Gynecol. 49, 424–429.
Rae, M.T., Niven, D., Ross, A., Forster, T., Lathe, R., Critchley, H.O., Ghazal,P., Hillier, S.G., 2004. Steroid signalling in human ovarian surface epithe-lial cells: the response to interleukin-1 alpha determined by microarrayanalysis. J. Endocrinol. 183, 19–28.
Resta, L., Russo, S., Colucci, G.A., Prat, J., 1993. Morphologic precursorsof ovarian epithelial tumours. Obstet. Gynecol. 82, 181–186.
Riman, T., Dickman, P.W., Nilsson, S., Correia, N., Nordlinder, H., Magnus-son, C.M., Persson, I.R., 2002. Risk factors for invasive epithelial ovariancancer: results from a Swedish case–control study. Am. J. Epidemiol. 156,363–373.
Risch, H.A., 1998. Hormonal etiology of epithelial ovarian cancer, with ahypothesis concerning the role of androgens and progesterone. J. Natl.Cancer Inst. 90, 1774–1786 (comment).
Risch, H.A., Marrett, L.D., Howe, G.R., 1994. Parity, contraception, infer-tility, and the risk of epithelial ovarian cancer. Am. J. Epidemiol. 140,585–597.
Risch, H.A., McLaughlin, J.R., Cole, D.E., Rosen, B., Bradley, L., Kwan, E.,Jack, E., Vesprini, D.J., Kuperstein, G., Abrahamson, J.L., Fan, I., Wong,B., Narod, S.A., 2001. Prevalence and penetrance of germ-line BRCA1and BRCA2 mutations in a population series of 649 women with ovariancancer. Am. J. Hum. Genet. 68, 700–710.
Risch, H.A., Weiss, N.S., Lyon, J.L., Daling, J.R., Liff, J.M., 1983. Eventsof reproductive life and the incidence of epithelial ovarian cancer. Am.J. Epidemiol. 117, 128–139.
Rodriguez, C., Tatham, L.M., Calle, E.E., Thun, M.J., Jacobs, E.J., Heath Jr.,C.W., 1998b. Infertility and risk of fatal ovarian cancer in a prospectivecohort of US women. Cancer Causes Control 9, 645–651.
R arian
R .A.,in ontosis?
R vivo11,
R , J.L.,f oralmiol.
R rtile
R rianRAF
R of the
S en-picheno-
810–
S , E.,, E.,ressionvar-rian
S ohr,h on
S D.,000.ndro-
111–117.ogales, F.F., 1995. Mesonephric (Wolffian) tumours of the female g
tract: is mesonephric histogenesis a mirage and trap? Curr. Diag. P94–100.
ogales, F.F., Carvia, R.E., Donne, C., Campello, T.R., Vidal, M., MaA., 1997. Adenomas of the rete ovarii. Hum. Pathol. 28, 1428–143
bata, K., Morland, S.J., Watson, R.H., Hitchcock, A., Chenevix-TrenchThomas, E.J., Campbell, I.G., 1998. Frequent PTEN/MMAC mutain endometrioid but not serous or mucinous epithelial ovarian tumCancer Res. 58, 2095–2097.
dend’hal, S., Wenzel, J.G., Player, E.C., 1986. The rete ovarii of cattldeer communicates with the uterine tube. Anat. Rec. 216, 40–43.
kamura, H., Katabuchi, H., 2001. Detailed morphology of human ovsurface epithelium focusing on its metaplastic and neoplastic capaItal. J. Anat. Embryol. 106, 263–276.
no, K., Tanaka, T., Tsunoda, T., Kitahara, O., Kihara, C., OkamotoOchiai, K., Takagi, T., Nakamura, Y., 2000. Identification by cDmicroarray of genes involved in ovarian carcinogenesis. Cancer Re5007–5011.
alacios, J., Gamallo, C., 1998. Mutations in the beta-catenin(CTNNB1) in endometrioid ovarian carcinomas. Cancer Res.1344–1347.
armiani, G., 1990. An explanation of the variable clinical response toleukin 2 and LAK cells. Immunol. Today 11, 113–115.
eoples, G.E., Schoof, D.D., Andrews, J.V., Goedegebuure, P.S., EbT.J., 1993. T-cell recognition of ovarian cancer. Surgery 114, 227–2
ersson, I., 2000. Estrogens in the causation of breast, endometrial anian cancers—evidence and hypotheses from epidemiological findinSteroid Biochem. Mol. Biol. 74, 357–364.
eters, H., McNatty, K.P., 1980. The Ovary. Granada Publishing Ltd., Slaxe, S.C., Deligdisch, L., Dottino, P.R., Cohen, C.J., 1990. Ovarian in
ithelial neoplasia demonstrated in patients with stage I ovarian carcinGynecol. Oncol. 38, 367–372.
urdie, D., Green, A., Bain, C., Siskind, V., Ward, B., Hacker, N., QuinnWright, G., Russell, P., Susil, B., 1995. Reproductive and other faand risk of epithelial ovarian cancer: an Australian case–control sSurvey of Women’s Health Study Group. Int. J. Cancer 62, 678–68
.,
,
,
r-.
.-.
odriguez, G.C., Carver, D., Anderson, K., et al., 2001. Evaluation of ovcancer preventive agents in the chicken. Gynecol. Oncol. 80, 317.
odriguez, G.C., Walmer, D.K., Cline, M., Krigman, H., Lessey, BWhitaker, R.S., Dodge, R., Hughes, C.L., 1998a. Effect of progestthe ovarian epithelium of macaques: cancer prevention through apopJ. Soc. Gynecol. Investig. 5, 271–276.
ollins, B.J., Sunday, M.E., 1991. Suppression of tumor formation inby expression of the JE gene in malignant cells. Mol. Cell. Biol.3125–3131.
osenberg, L., Palmer, J.R., Zauber, A.G., Warshauer, M.E., Lewis Jr.Strom, B.L., Harlap, S., Shapiro, S., 1994. A case–control study ocontraceptive use and invasive epithelial ovarian cancer. Am. J. Epide139, 654–661.
ossing, M.A., Daling, J.R., 1994. Ovarian tumours in a cohort of infewomen. N. Engl. J. Med. 331, 771–776.
ussell, S.E., McCluggage, W.G., 2004. A multistep model for ovatumorigenesis: the value of mutation analysis in the KRAS and Bgenes. J. Pathol. 203, 617–619.
utgers, J.L., Scully, R.E., 1988. Cysts (cystadenomas) and tumoursrete ovarii. Int. J. Gynecol. Pathol. 7, 330–342.
alazar, H., Godwin, A.K., Daly, M.B., Laub, P.B., Hogan, W.M., Rosblum, N., Boente, M.P., Lynch, H.T., Hamilton, T.C., 1996. Microscobenign and invasive malignant neoplasms and a cancer-prone ptype in prophylactic oophorectomies. J. Natl. Cancer Inst. 88, 11820.
antin, A.D., Zhan, F., Bellone, S., Palmieri, M., Cane, S., BignottiAnfossi, S., Gokden, M., Dunn, D., Roman, J.J., O’Brien, T.J., TianCannon, M.J., Shaughnessy Jr., J., Pecorelli, S., 2004. Gene expprofiles in primary ovarian serous papillary tumours and normal oian epithelium: identification of candidate molecular markers for ovacancer diagnosis and therapy. Int. J. Cancer 112, 14–25.
ass, B., Rehm, S., 1994. Tumours of the ovary. In: Turusov, V.S., MU. (Eds.), Tumours of the Mouse. International Agency for ResearcCancer, Lyon, pp. 493–526.
aunders, P.T., Millar, M.R., Williams, K., Macpherson, S., Harkiss,Anderson, R.A., Orr, B., Groome, N.P., Scobie, G., Fraser, H.M., 2Differential expression of estrogen receptor-alpha and -beta and a

20 J.S. Fleming et al. / Molecular and Cellular Endocrinology 247 (2006) 4–21
gen receptor in the ovaries of marmosets and humans. Biol. Reprod. 63,1098–1105.
Sawasaki, T., Shigemasa, K., Gu, L., Beard, J.B., O’Brien, T.J., 2004. Thetransmembrane protease serine (TMPRSS3/TADG-12) D variant: a poten-tial candidate for diagnosis and therapeutic intervention in ovarian cancer.Tumour Biol. 25, 141–148.
Schildkraut, J.M., Schwingl, P.J., Bastos, E., Evanoff, A., Hughes, C., 1996.Epithelial ovarian cancer risk among women with polycystic ovary syn-drome. Obstet. Gynecol. 88, 554–559.
Schwartz, D.R., Kardia, S.L., Shedden, K.A., Kuick, R., Michailidis, G.,Taylor, J.M., Misek, D.E., Wu, R., Zhai, Y., Darrah, D.M., Reed, H.,Ellenson, L.H., Giordano, T.J., Fearon, E.R., Hanash, S.M., Cho, K.R.,2002. Gene expression in ovarian cancer reflects both morphology andbiological behavior, distinguishing clear cell from other poor-prognosisovarian carcinomas. Cancer Res. 62, 4722–4729.
Scorilas, A., Chiang, P.M., Katsaros, D., Yousef, G.M., Diamandis, E.P.,2003. Molecular characterization of a new gene, CEAL1, encoding fora carcinoembryonic antigen-like protein with a highly conserved domainof eukaryotic translation initiation factors. Gene 310, 79–89.
Scully, R.E., 1977. Ovarian tumours. A review. Am. J. Pathol. 87, 686–720.Segal, E., Friedman, N., Koller, D., Regev, A., 2004. A module map showing
conditional activity of expression modules in cancer. Nat. Genet. 36,1090–1098.
Shih, Y.C., Kerr, J., Liu, J., Hurst, T., Khoo, S.K., Ward, B., Wainwright,B., Chenevix-Trench, G., 1997. Rare mutations and no hypermethylationat the CDKN2A locus in epithelial ovarian tumours. Int. J. Cancer 70,508–511.
Shridhar, V., Lee, J., Pandita, A., Iturria, S., Avula, R., Staub, J., Morrissey,M., Calhoun, E., Sen, A., Kalli, K., Keeney, G., Roche, P., Cliby, W., Lu,K., Schmandt, R., Mills, G.B., Bast Jr., R.C., James, C.D., Couch, F.J.,Hartmann, L.C., Lillie, J., Smith, D.I., 2001. Genetic analysis of early-
S e, J.s inactio
S seds. 49
S S.,, R.,mas.
S ariant the
S nez,elial
ion: a
S verse160
S oral–110
S evix-poly-30,
S evix-rececer.
S min-
S su-C-lated
genes for ovarian cancer cells by gene profiling. Oncogene 24, 1774–1787.
Tan, O.L., Hurst, P.R., Fleming, J.S., 2005. Location of inclusion cysts inmouse ovaries in relation to age, pregnancy and total ovulation number:implications for ovarian cancer? J. Pathol. 205, 483–490.
Tannin, G.M., Agarwal, A.K., Monder, C., New, M.I., White, P.C., 1991.The human gene for 11 beta-hydroxysteroid dehydrogenase. Structure,tissue distribution, and chromosomal localization. J. Biol. Chem. 266,16653–16658.
Tavani, A., Gallus, S., La Vecchia, C., Conti, E., Montella, M., Franceschi,S., 2000. Aspirin and ovarian cancer: an Italian case–control study. Ann.Oncol. 11, 1171–1173.
Teneriello, M.G., Ebina, M., Linnoila, R.I., Henry, M., Nash, J.D., Park, R.C.,Birrer, M.J., 1993. p53 and Ki-ras gene mutations in epithelial ovarianneoplasms. Cancer Res. 53, 3103–3108.
Tetsuka, M., Milne, M., Simpson, G.E., Hillier, S.G., 1999. Expression of11beta-hydroxysteroid dehydrogenase, glucocorticoid receptor, and min-eralocorticoid receptor genes in rat ovary. Biol. Reprod. 60, 330–335.
Thomas, N.A., Neville, P.J., Baxter, S.W., Campbell, I.G., 2003. Geneticanalysis of benign ovarian tumours. Int. J. Cancer 105, 499–505.
Thung, P.J., Boot, L.M., Muhlbock, O., 1956. Senile changes in the oestrouscycle and in ovarian structure in some inbred strains of mice. ActaEndocrinol. 23, 8–32.
Titus-Ernstoff, L., Perez, K., Cramer, D.W., Harlow, B.L., Baron, J.A., Green-berg, E.R., 2001. Menstrual and reproductive factors in relation to ovariancancer risk. Br. J. Cancer 84, 714–721.
Tortolero-Luna, G., Mitchell, M.F., Rhodes-Morris, H.E., 1994. Epidemiologyand screening of ovarian cancer. Obstet. Gynecol. Clin. North Am. 21,1–23.
Vermeirsch, H., Simoens, P., Coryn, M., Van den Broeck, W., 2001.Immunolocalization of androgen receptors in the canine ovary and
122,
W n, N.,roach42.
W Don-ours
W Con-by
W I.G.,is an
W .R.,R.L.,03.
sed in410–
W rature
W ter-oved
cer.
W asiveer 71,
W ovar-s. IV.Can-
W viron-
W mary
versus late-stage ovarian tumours. Cancer Res. 61, 5895–5904.hridhar, V., Sen, A., Chien, J., Staub, J., Avula, R., Kovats, S., Le
Lillie, J., Smith, D.I., 2002. Identification of underexpressed geneearly- and late-stage primary ovarian tumours by suppression subtrhybridization. Cancer Res. 62, 262–270.
hu, X.O., Brinton, L.A., Gao, Y.T., Yuan, J.M., 1989. Population-bacase–control study of ovarian cancer in Shanghai. Cancer Re3670–3674.
hvartsman, H.S., Lu, K.H., Lee, J., Lillie, J., Deavers, M.T., Clifford,Wolf, J.K., Mills, G.B., Bast Jr., R.C., Gershenson, D.M., Schmandt2003. Overexpression of kallikrein 10 in epithelial ovarian carcinoGynecol. Oncol. 90, 44–50.
ightler, S.E., Boike, G.M., Estape, R.E., Averette, H.E., 1991. Ovcancer in women with prior hysterectomy: a 14-year experience aUniversity of Miami. Obstet. Gynecol. 78, 681–684 (comment).
ilva, E.G., Tornos, C., Fritsche Jr., H.A., el Naggar, A., Gray, K., OrdoN.G., Luna, M., Gershenson, D., 1997. The induction of benign epithneoplasms of the ovaries of guinea pigs by testosterone stimulatpotential animal model. Mod. Pathol. 10, 879–883.
inger, G., Kurman, R.J., Chang, H.W., Cho, S.K., Shih Ie, M., 2002. Ditumorigenic pathways in ovarian serous carcinoma. Am. J. Pathol.1223–1228.
iskind, V., Green, A., Bain, C., Purdie, D., 2000. Beyond ovulation:contraceptives and epithelial ovarian cancer. Epidemiology 11, 106
purdle, A.B., Purdie, D.M., Webb, P.M., Chen, X., Green, A., ChenTrench, G., 2001b. The microsomal epoxide hydrolase Tyr113Hismorphism: association with risk of ovarian cancer. Mol. Carcinog.71–78.
purdle, A.B., Webb, P.M., Purdie, D.M., Chen, X., Green, A., ChenTrench, G., 2001a. No significant association between progesteronetor exon 4 Va1660Leu G/T polymorphism and risk of ovarian canCarcinogenesis 22, 717–721.
tewart, P.M., Mason, J.I., 1995. Cortisol to cortisone: glucocorticoid toeralocorticoid. Steroids 60, 143–146.
yed, V., Mukherjee, K., Lyons-Weiler, J., Lau, K.M., Mashima, T., Truo, T., Ho, S.M., 2005. Identification of ATF-3, caveolin-1, DL1, and NM23-H2 as putative antitumorigenic, progesterone-regu
,
n
,
,
.
p-
their relation to sex steroid hormone concentrations. Reproduction711–721.
acholder, S., Chanock, S., Garcia-Closas, M., El Ghormli, L., Rothma2004. Assessing the probability that a positive report is false: an appfor molecular epidemiology studies. J. Natl. Cancer. Inst. 96, 434–4
arrenfeltz, S., Pavlik, S., Datta, S., Kraemer, E.T., Benigno, B., Mcald, J.F., 2004. Gene expression profiling of epithelial ovarian tumcorrelated with malignant potential. Mol. Cancer 3, 27.
atson, J.M., Sensintaffar, J.L., Berek, J.S., Martinez-Maza, O., 1990.stitutive production of interleukin 6 by ovarian cancer cell lines andprimary ovarian tumor cultures. Cancer Res. 50, 6959–6965.
atson, R.H., Neville, P.J., Roy Jr., W.J., Hitchcock, A., Campbell,1998. Loss of heterozygosity on chromosomes 7p, 7q, 9p and 11qearly event in ovarian tumorigenesis. Oncogene 17, 207–212.
elsh, J.B., Sapinoso, L.M., Kern, S.G., Brown, D.A., Liu, T., Bauskin, AWard, R.L., Hawkins, N.J., Quinn, D.I., Russell, P.J., Sutherland,Breit, S.N., Moskaluk, C.A., Frierson Jr., H.F., Hampton, G.M., 20Large-scale delineation of secreted protein biomarkers overexprescancer tissue and serum. Proc. Natl. Acad. Sci. U.S.A. 100, 33415.
enzel, J.G., Odend’hal, S., 1985. The mammalian rete ovarii: a litereview. Cornell Vet. 75, 411–425.
erness, B.A., Afify, A.M., Bielat, K.L., Eltabbakh, G.H., Piver, M.S., Pason, J.M., 1999. Altered surface and cyst epithelium of ovaries remprophylactically from women with a family history of ovarian canHum. Pathol. 30, 151–157.
hittemore, A.S., 1993. Personal characteristics relating to risk of invepithelial ovarian cancer in older women in the United States. Canc558–565.
hittemore, A.S., Harris, R., Itnyre, J., 1992. Characteristics relating toian cancer risk: collaborative analysis of 12 US case–control studieThe pathogenesis of epithelial ovarian cancer. Collaborative Ovariancer Group. Am. J. Epidemiol. 136, 1212–1220.
ilson, J.E., 1958. Adeno-carcinomata in hens kept in a constant enment. Poult. Sci. 37, 1253.
oodward, W.A., Chen, M.S., Behbod, F., Rosen, J.M., 2005. On mamstem cells. J. Cell Sci. 118, 3585–3594.

J.S. Fleming et al. / Molecular and Cellular Endocrinology 247 (2006) 4–21 21
Woolnough, E., Russo, L., Khan, M.S., Heatley, M.K., 2000. An immuno-histochemical study of the rete ovarii and epoophoron. Pathology 32,77–83.
Wright, K., Wilson, P., Morland, S., Campbell, I., Walsh, M., Hurst, T., Ward,B., Cummings, M., Chenevix-Trench, G., 1999. Beta-catenin mutationand expression analysis in ovarian cancer: exon 3 mutations and nucleartranslocation in 16% of endometrioid tumours. Int. J. Cancer 82, 625–629.
Wynder, E.L., Dodo, H., Barber, H.R., 1969. Epidemiology of cancer of theovary. Cancer 23, 352–370.
Yang, W.L., Roland, I.H., Godwin, A.K., Xu, X.X., 2005. Loss of TNF-alpha-regulated COX-2 expression in ovarian cancer cells. Oncogene (July), 25.
Yong, P.Y., Harlow, C., Thong, K.J., Hillier, S.G., 2002. Regulation of 11beta-hydroxysteroid dehydrogenase type 1 gene expression in human ovariansurface epithelial cells by interleukin-1. Hum. Reprod. 17, 2300–2306.
Zackrisson, U., Mikuni, M., Wallin, A., Delbro, D., Hedin, L., Brannstrom,M., 1996. Cell-specific localization of nitric oxide synthases (NOS) in the
rat ovary during follicular development, ovulation and luteal formation.Hum. Reprod. 11, 2667–2673.
Zhang, L., Conejo-Garcia, J.R., Katsaros, D., Gimotty, P.A., Massobrio, M.,Regnani, G., Makrigiannakis, A., Gray, H., Schlienger, K., Liebman,M.N., Rubin, S.C., Coukos, G., 2003. Intratumoral T cells, recurrence,and survival in epithelial ovarian cancer. N. Engl. J. Med. 348, 203–213.
Zheng, M., Simon, R., Kononen, J., Sauter, G., Mihatsch, M.J., Moch, H.,2004. Analysis of gene expression profiles among 3 epithelial ovar-ian tumor subtypes using cDNA and tissue microarrays. Ai Zheng 23,771–776.
Zorn, K.K., Jazaeri, A.A., Awtrey, C.S., Gardner, G.J., Mok, S.C., Boyd, J.,Birrer, M.J., 2003. Choice of normal ovarian control influences deter-mination of differentially expressed genes in ovarian cancer expressionprofiling studies. Clin. Cancer Res. 9, 4811–4818.