in-vivo-methoden zur analyse von muskelstoffwechsel und körperzusammensetzung beim schwein unter...
TRANSCRIPT

In-vivo-Methoden zur Analyse von Muskelstoffwechsel
und Körperzusammensetzung beim Schwein unter besonderer Berücksichtigung genetischer Einflüsse
Dr. agr. Armin M. Scholz
Oberschleißheim, 2002

Aus dem Institut für Tierzucht der Tierärztlichen Fakultät
der Ludwig-Maximilians-Universität München Lehrstuhl für Tierzucht und Allgemeine Landwirtschaftslehre
Univ.-Prof. Dr. Dr. Martin Förster
In-vivo-Methoden zur Analyse von Muskelstoffwechsel und Körperzusammensetzung beim Schwein unter
besonderer Berücksichtigung genetischer Einflüsse
HABILITATIONSSCHRIFT zur Erlangung der Lehrbefugnis
für das Fach Tierzucht und Allgemeine Landwirtschaftlehre an der Tierärztlichen Fakultät
der Ludwig-Maximilians-Universität München
eingereicht von
Dr. agr. Armin M. Scholz Oberschleißheim, 2002
Gutachter: Univ.-Prof. Dr. Dr. habil. M. Förster Univ.-Prof. Dr. Dr. habil. K. Heinritzi Univ.-Prof. Dr. A. Stolle Kolloquium: 03.12.2002


Für Wiebke und Lisanne

Teile der Habilitationsschrift wurden in folgenden Publikationen veröffentlicht:
Scholz, A.M., A.D. Mitchell, P.C. Wang, H. Song and Z. Yan (1995): Muscle metabolismand body composition of pigs with different ryanodine receptor genotypes studied bymeans of 31P nuclear magnetic resonance spectroscopy and 1H magnetic resonanceimaging. Arch. Tierzucht 38: 539-552.
Mitchell, A.D. and A.M. Scholz (1997): Dual-energy X-ray absorptiometry (DXA) analysisof growth and composition of pigs of different ryanodine receptor genotypes. Arch.Tierzucht 40: 47-56.
Mitchell, A.D., A.M. Scholz, and Harry J. Mersmann (2001): Growth and Body Composition(Chapter 6). In: Biology of the Domestic Pig, 2nd edition (Eds.: W. Pond and H.Mersmann) Cornell University Press, Ithaca, NY, USA: 225-308.
Mitchell, A.D. and A.M. Scholz (2001): Techniques for Measuring Body Composition ofSwine (Chapter 41). In: Swine Nutrition 2nd Edition (Ed.: A.J. Lewis and L. LeeSouthern), CRC Press, Boca Raton, FL, USA: 917-960.
Mitchell, A.D., A.M. Scholz, P.C. Wang and H. Song (2001): Body composition analysis ofthe pig by magnetic resonance imaging. J. Anim. Sci. 79: 1800-1813.
Auf weitere Ergebnisse oder Daten aus eigenen Arbeiten oder aus Arbeiten in
Zusammenarbeit mit anderen Wissenschaftlern wird gesondert im Text hingewiesen.

Inhaltsverzeichnis
a
Abkürzungsverzeichnis IDefinition statistischer Parameter VTabellenverzeichnis VIAbbildungsverzeichnis XIIGenehmigung der Tierversuche XVII
1 Einleitung 1
2 Methodenübersicht
2.1 Muskelstoffwechseluntersuchungen in vivo mittelsMagnetresonanz-Spektroskopie (MRS)
4
2.1.1 31P-Magnetresonanz-Spektroskopie 92.1.2 13C-Magnetresonanz-Spektroskopie 15
2.2 Nicht invasive Methoden zur Ermittlung derKörperzusammensetzung - in vivo
2.2.1 Einflussfaktoren auf die Körperzusammensetzung und derenMessung im Rahmen der Leistungsprüfung
18
2.2.2 Physikalische und chemische Grundlagen für die Messungder Körperzusammensetzung – Modelle derKörperzusammensetzung
21
2.2.3 Methoden der Leistungsprüfung2.2.3.1 Subjektive Methoden 312.2.3.2 Physikalische und chemische (objektive) Methoden 332.2.3.2.1 Körpermasse (Wägung) 332.2.3.2.2 Zerlegung und chemische Analyse (invasiv) 342.2.3.2.3 Densitometrie (spezifische Dichte) – hydro- und aerostatische
Methoden 37
2.2.3.2.4 Lineare Messungen 402.2.3.2.5 Radiologische und bildgebende Methoden im Überblick 412.2.3.2.5.1 Computer-Tomographie (CT) 482.2.3.2.5.2 Magnetresonanz-Tomographie (MRT) 532.2.3.2.5.2.1 Statistische Bildanalyse zur Quantifizierung von
Gewebeanteilen61
2.2.3.2.5.3 Magnetresonanz-Spektroskopie (MRS) 682.2.3.2.5.4 Dualenergie-Röntgenabsorptiometrie (DXA) 692.2.3.2.5.5 Ultraschall (US) 842.2.3.2.5.6 Bioelektrische Impedanz (BIA) 912.2.3.2.5.7 Gesamtkörperleitfähigkeit - „Total Body Electrical
Conductivity“ (TOBEC)95
2.2.3.2.5.8 Infrarot-Körperfettanalyse (Infrarot-Interaktanz) 972.2.3.2.5.9 Videobildanalyse und Dreidimensionale Topometrie 982.2.3.2.5.10 Kalium40-Gammaspektrometrie 992.2.3.2.5.11 Neutronenaktivierung 1012.2.3.3 Stoffwechselanalyse 1052.2.3.3.1 Verdünnungsmethoden 1052.2.3.3.2 Kreatininausscheidung 108

Inhaltsverzeichnis
b
2.3 Ergänzende (invasive) Methodik zur Bestimmung vonMuskelfasermerkmalen mittels Biopsie
110
3 Eigene Untersuchungen3.1 Material und Methodik
3.1.1 Muskelstoffwechseluntersuchungen3.1.1.1 31P-Magnetresonanz-Spektroskopie 1133.1.1.2 13C-Magnetresonanz-Spektroskopie 118
3.1.2 Ermittlung der Körperzusammensetzung mittels MRT undDXA
3.1.2.1 1H-Magnetresonanz-Tomographie 3.1.2.1.1 Lebendmassegruppe bis 20 kg 1263.1.2.1.2 Lebendmassegruppen 30 kg, 60 kg, 90 kg 1303.1.2.2 Dualenergie-Röntgenabsorptiometrie 136
3.1.3 Haltung, Fütterung und Transport 141
3.1.4 Referenzschlachtung 142
3.1.5 Bestimmung der Muskelfaserstruktur mittels Schussbiopsie 146
3.1.6 DNA-Analyse für den Ryanodin-Rezeptor-1-Gentest 150
3.2 Ergebnisse
3.2.1 Muskelstoffwechseluntersuchungen3.2.1.1 Phosphorkomponenten - 31P-Magnetresonanz-Spektroskopie 1533.2.1.2 Kohlenstoffkomponenten - 13C-Magnetresonanz-
Spektroskopie 160
3.2.2 Körperzusammensetzung 3.2.2.1 MRT-Lebendmassegruppe bis 20 kg 1673.2.2.2 MRT- Lebendmassegruppen 30 kg, 60 kg, 90 kg 169
3.2.2.3 Genauigkeit der MR-Tomographie 1773.2.2.4 DXA Lebendmassegruppen 10 kg, 30 kg, 60 kg und 90 kg 1863.2.2.5 Genauigkeit der Dualenergie-Röntgenabsorptiometrie 1893.2.2.6 Vergleich der Genauigkeit von Magnetresonanz-
Tomographie und Dualenergie-Röntgenabsorptiometrie zurErmittlung der Körperzusammensetzung
195
3.2.3 Magergewebewachstum und Magergewebefutterverwertung 198
3.2.4 Muskelfaserstruktur und ihre Beziehung zu Parametern ausder 31P-MR-Spektroskopie
206

Inhaltsverzeichnis
c
4 Diskussion
4.1 Ursachen für die Variation in Muskelstoffwechselvorgängen 215
4.2 Genetische Steuerung des Körperfettgehaltes und desWachstums beim Schwein - Konsequenzen für dieZuchtarbeit
238
5 Zusammenfassung 2606 Summary 263
7 Literaturverzeichnis 266
8 Anhang I: Zusätzliche 1H-MRT-Ergebnisse 339II: Zusätzliche Ergebnisse aus der 13C-MR-Spektroskopie 345III: PCR-Protokoll 346IV: Clusteranalyse 352V: Quantitative Trait Loci für Körperfettgehalt 367

Abkürzungsverzeichnis
I
Abkürzungsverzeichnis
Abb. AbbildungACTH Adrenocorticotropes Hormon AMP AdenosinmonophosphatADP AdenosindiphosphatA-FABP Adipocyte fatty acid-binding protein [Adipozyten-
Fettsäurenbindungsprotein]AP AktionspotentialARS Agricultural Research Service [Landwirtschaftliche Forschungsanstalt]ATP AdenosintriphosphatBARC Beltsville Agricultural Research Center [Landwirtschaftliches
Forschungszentrum Beltsville]BIA Bioelektrische ImpedanzBMC Bone Mineral Content (Knochenmineralgehalt), gBMD Bone Mineral Density (Knochenmineraldichte), g/cm2
BMPC Prozentualer Knochenmineralgehalt bp BasenpaareBW Body Weight (Lebendmasse)C KohlenstoffCa Kalziumca. zirkaCAT Computer-Aided-Tomography (teilweise Computer-Axial-
Tomographie)Chem. chemischCK Kreatinkinase (Kreatinphosphokinase)cM Centi-MorganCRC Calcium Release Channel (Kalziumfreigabekanal)cSv 1 cSv = 0,01 Sievert (Sv) = 1 rem Äquivalentdosis
[1Sv = 1 Joule/Kilogramm = 1 Gray (Gy) = 100 rem] Körperdosisoder effektive Dosis der Röntgenstrahlung
CT Computer-TomographieD2O Schweres Wasser (Deuteriumoxid)DE Deutsches EdelschweinDE im Zusammenhang: „digestable energy“ = verdauliche EnergieDFD dark, firm, dry (dunkel, fest, trocken); hoher pH-WertDL Deutsche LandrasseDNA / DNS Desoxyribonucleinacid (Desoxyribonukleinsäure)Du DurocDPA Dual-PhotonenabsorptiometrieDXA (oder DEXA) Dualenergie-RöntgenabsorptiometrieEDTA Ethylen-Diamin-TetraessigsäureETL Economic Trait Loci (ökonomisch wichtige Merkmals-Genorte)F1 Filialgeneration (1. Kreuzungsgeneration)FAL Bundesforschungsanstalt für LandwirtschaftFETTV Volumen von subkutanem Fett + Haut über der Region der Musculi

Abkürzungsverzeichnis
II
longissimus dorsi aus MR-Tomographie (cm³)FGM Fettgewebemasse (DXA)FFM Fettfreie MasseFID Freier Induktionsabfall (Free Induction Decay)FOM Fat-o-Meater (optisches Fett- und Fleischmessgerät)FMR Verhältnis von Fett- und Muskelfläche (M.l.d.) zwischen 13./14.
Brustwirbel der linken SchlachthälfteFMV Verhältnis von Fett- und Muskelvolumen in der Region der Musculi
longissimus dorsi aus MR-Tomographie (10 cm Region)FUA Futteraufnahme (kg/d)FUV Futterverwertung (kg Futter /kg Lebendmassezunahme)GK GesamtkörperGKC Gesamtkörper-KohlenstoffGKCa Gesamtkörper-KalziumGKN Gesamtkörper-StickstoffGKO Gesamtkörper-SauerstoffGLM General Linear Model (Allgemeines Lineares Modell)GMP Glukose-MonophosphatGP Glykolytisches PotentialG6P Glukose-6-Phosphat1H H H-1 Wasserstoff (ein Proton Kernspin = ½ )2H D H-2 Deuterium (ein Proton und ein Neutron Kernspin = 1), schwerer
Wassersoff3H T H-3 Tritium (ein Proton [+ zwei Neutronen] Kernspin = ½); radioaktiv,
überschwerer WasserstoffHa HampshireHAPI Hampshire x Pietrain - KreuzungHU Hounsfield Units (Hounsfield-Einheiten), Maßeinheit für die
RöntgenschwächungIGF Insulin Like Growth Factor (Insulin ähnlicher Wachstumsfaktor)IGFBP Insulin Like Growth Factor Binding Protein (Insulin ähnlicher
Wachstumsfaktor-Bindungsprotein)IMF Intramuskulärer FettgehaltIMP Inosinmonophosphatinsg. insgesamtK KaliumKB Künstliche BesamungkDa Kilodalton (Masseeinheit) = 1000 DaltonKMM Knochenmineralmasse (DXA)LB Landrasse B (Belgische Landrasse)Lc Leicoma (Schweinerasse Leipzig, Cottbus, Magdeburg; Duroc-
Genanteil ca. 45 %)LEP Leptin-GenortLF1 Leitfähigkeit 85/145 min post mortemLF24 Leitfähigkeitswert 24 Stunden post mortemLIM Spezielle „Zinkfinger-Protein“-Familie (KHURANA, 2000)

Abkürzungsverzeichnis
III
LM Lebendmasse (kg)LR US-LandrasseLW Large WhiteLVG Lehr- und Versuchsgut Oberschleißheim der Tierärztlichen Fakultät der
Ludwig-Maximilians-Universität MünchenM. MusculusMm.l.d Musculi longissimi dorsi (linke und rechte Seite)MFUV Magergewebefutterverwertung (kg Futter /kg Magergewebezunahme)MGM Magergewebemasse (kg; DXA)MGWR Magergewebewachstumsrate (g/d)MHS Malignes Hyperthermie SyndromMLDV Volumen der Mm. longissimi dorsi (cm³)MR MagnetresonanzmRNA „Messenger RNA“ – Boten-RibonukleinsäureMRS Magnetresonanz-SpektroskopieMRT / MRI Magnetresonanz-Tomographie / Imaging (Bildgebung)N StickstoffNa NatriumNAA Neutronenaktivierungs-AnalyseNIR Nahinfrarot-ReflektionsmessungNIT Nahinfrarot-TransmissionsmessungNN homozygot „normal“ am RyR1-GenortNn siehe auch Np heterozygot am RyR1-Genortnn siehe auch pp homozygot „defekt“ am RyR1-GenortNMR /MR Nuklear-Magnetische ResonanzNp heterozygot am RyR1-GenortPCr Phosphokreatin (Phosphocreatine) = KreatinphosphatPCR Polymerase Chain Reaction (Polymerase Kettenreaktion)31P Phosphor-Isotop 31PDE PhosphodiesterpH negativ dekadischer Logarithmus der WasserstoffionenkonzentrationPi Im Zusammenhang: Anorganisches Phosphat (inorganic Phosphate)PI PietrainPI x Ha Pietrain x HampshirePixel picture elements (Bildpunkte)p.m. post mortemPME Phosphomonoesterpp homozygot „defekt“ (mutiert) am RyR1-Genortppm Parts Per MillionPSE pale, soft, exudative (blass, weich, wässrig), zu niedriger pH-Wert, zu
schneller GlykogenabbaupST porcine Somatotropin (porcines Wachstumshormon)QTL Quantitative Trait Loci (Genorte für quantitative Merkmale)R-Wert Im Zusammenhang: R-Wert (spezifische Weichgewebeverhältniszahl
aus DXA in Abhängigkeit von der Röntgenschwächung)

Abkürzungsverzeichnis
IV
RF RadiofrequenzRN Rendement Napole-Genort
RN - Rendement Napole, (Kochschinkenverlust-Gen), dominantes „Defekt“-Allel (”Hampshirefaktor”)
rn+ normales Allel am Rendement-Napole-GenortROI Region of Interest (interessierende Region)RP RohproteinRS Ohmscher WiderstandRyR1 Ryanodin-Rezeptor-1-GenortS Spotted (US-Amerikanische Schweinerasse)SFETTV Volumen der subkutanen Fettauflage + Haut am Teilstück Schinken
(cm³)SGV Gesamtvolumen des Schinkens (cm³)SMGV Volumen des Schinkenmagergewebes (cm³)SFMV Verhältnis von Fett- und Magergewebevolumen am SchinkenSubst. SubstanzSPA Mono-Photonenabsorptiometrie (Single Photon Absorptiometry)SR Sarkoplasmatisches RetikulumT TeslaTab. TabelleT1-Relaxationszeit Longitudinale Relaxationszeit Dauer des Vorgangs für die
Polarisation der Kernspins längs der Magnetfeldlinien des äußerenMagnetfeldes (Spin-Gitter-Relaxationszeit)
T2-Relaxationszeit Transversale Relaxationszeit Dauer des Vorgangs für dieDephasierung der Kernspins durch ihre Wechselbeziehunguntereinander (erfolgt quer zum äußeren Magnetfeld; auch: Spin-Spin-Relaxationszeit)
TE Time between Echos Echoausleseverzögerung (Zeitverzögerungzwischen der Anregung der Kernspins und der Echoauslesung – in einerSpin-Echo-Sequenz: die Zeit zwischen einem 90° Puls und demMaximum der Signalintensität im Echo)
TNF Tumor-Nekrose-FaktorTR Time of Repetition Pulswiederholzeit (Dauer zwischen zwei
Hochfrequenz-Anregungsimpulsen)TZ Tageszunahme (g)USDA United States Department of Agriculture (US-
Landwirtschaftsministerium)Vol. oder -vol. Volumen (cm³)VOI Volume of Interest (interessierendes Volumen)Xc ReaktanzZ ScheinwiderstandZNS Zentrales NervensystemZMP Zentrale Markt- und Preisberichtstelle für Erzeugnisse der Land-, Forst-
und Ernährungswirtschaft
Weitere nicht häufig wiederkehrende Abkürzungen werden direkt im Text erklärt.

Abkürzungsverzeichnis
V
Definition statistischer Parameter:
b Regressionskoeffizient C.V. Variationskoeffizient: Standardabweichung / StichprobenmittelDF Degrees of Freedom (Freiheitsgrade)ln Natürlicher LogarithmusLSM Siehe nächste ZeileLSQ-Means Least Squares Means (Kleinste Quadrate Mittelwerte)MSE Mean Squares Error = se
2 (Fehlervarianz bzw. Mittlere Quadrate Fehleroder Restfehler)(√MSE = Root Mean Squares Error = SEE)
n.s. nicht signifikant (Die Signifikanzgrenze für den Fehler 1. Art liegt im eigenen Materialimmer bei p ≤ 0,05.)
p Irrtumswahrscheinlichkeit: Signifikanzniveau, wenn der Effekt derModellgleichung hinzugefügt wird. Die verbleibende Summe derQuadrate (Rest-Summe der Quadrate) kann nach Erweiterung derSchätzgleichung um weitere Schätzvariablen weiter reduziert werden,wenn das Niveau von „p“ signifikant bleibt.
r Korrelationskoeffizient: Maßzahl für die Stärke der Beziehung zwischenzwei Variablen (Merkmalen) r = -1 ≥ 0 ≤ +1 ; r = 0 keine Beziehung;r = |± 1| vollständige Abhängigkeit
R2 Bestimmtheitsmaß (Anteil der durch das Modell erklärten Varianz ander Gesamtvarianz)
RMSE = SEE =√MSE
Wurzel aus dem Restfehler (Mittlere Quadrate Fehler):Standardabweichung der Residuen (gemessener Wert –Erwartungswert); Standardschätzfehler, (Abweichung von derRegressionsgeraden bei Regressionsanalyse)
RSD Reststandardabweichung (SEE/SD) oder (SEP/SD)SD Standard deviation (Standardabweichung)SEE Standard Error of Estimation (Standardschätzfehler, Wurzel aus dem
Mittleren Quadrate Fehler = √MSE = RSME)SE % (oder S.E.) Prozentuale Einheiten der ReststandardabweichungSEP Standard Error of Prediction (Standardfehler der Vorhersage)
(y y ) / ni i2− ∧ ; Parameter aus der Kreuzvalidierung
∅ oder (x) Durchschnitt (arithmetisches Mittel)|...| Absoluter Betrag einer positiven oder negativen Zahl

Tabellenverzeichnis
VI
Tabellenverzeichnis
Tabelle Titel Seite
2.2.1 Schlachtkörperzusammensetzung von zehn Tieren inunterschiedlichen Maststadien (nach den Angaben von ANDERSON1860, CAMERON, 1868, HALL, 1905)
35
2.2.2 Bewertung „radiologischer“ Methoden zur Bestimmung derKörperzusammensetzung
46
2.2.3 Vor- und Nachteile gebräuchlicher Verfahren zur nicht invasivenBildgebung für die Untersuchung der Körperzusammensetzung
47
2.2.4 Erklärungen zur Interpretation von Bland-Altman-Plots 662.2.5 Wassergehalt von „fettfreiem“ Fleisch unterschiedlicher Tierarten 1063.1.1 In der Literatur verwendete Konstanten zur Berechnung des pHi-
Wertes116
3.1.2 Linienherkunft und Anzahl Probanden für die 1H-MR-Tomographiebis 20 kg
126
3.1.3 Anzahl tomographierter Tiere je RyR1-Genotyp undLebendmassegruppe (30 – 90 kg)
131
3.1.4 Anzahl Referenztiere (für MRT) je RyR1-Genotyp undLebendmassegruppe (30 - 90 kg)
132
3.1.5 Anzahl tomographierter Tiere in Abhängigkeit von Rasse- bzw.Linienzugehörigkeit und Lebendmassegruppe (30-90 kg)
132
3.1.6 Anzahl tomographierter Tiere je Geschlechts- undLebendmassegruppe (30 - 90 kg)
132
3.1.7 Aufteilung der Probanden nach RyR1-Genotyp undLebendmassegruppe (10 - 90 kg) für den DXA-Versuch
136
3.1.8 Aufteilung der Probanden nach Rasse- bzw. Linienzugehörigkeit undLebendmassegruppe (10 - 90 kg) für den DXA-Versuch
136
3.1.9 Aufteilung der Probanden nach Geschlecht und Lebendmassegruppe(10 - 90 kg) für den DXA-Versuch
137
3.1.10 Anzahl Referenztiere je RyR1-Genotyp und Lebendmassegruppe 1433.1.11 Linien- und Geschlechterverteilung innerhalb RyR1-Genotyp für
Referenztiere (10 kg)144
3.1.12 Linien- und Geschlechterverteilung innerhalb RyR1-Genotyp fürReferenztiere (30-90 kg)
145
3.1.13 Tieranzahl in Abhängigkeit von Rasse- bzw. Linienzugehörigkeit undLebendmassegruppe im Wachstumsversuch (DXA)
145
3.2.1 Vergleich von Stoffwechselparametern aus der 31P-MR-Spektroskopie(PCr, pH, ATP, Pi) sowie für die Körpertemperatur nachHalothanbelastung in Schweinen mit unterschiedlichen RyR1-Genotypen
154
3.2.2 Vergleich von Stoffwechselparametern aus der 31P-MR-Spektroskopie(PCr, pH, ATP, Pi) sowie für die Körpertemperatur nachHalothanbelastung in Schweinen mit unterschiedlicher Rasse- bzw.Linienzugehörigkeit
159

Tabellenverzeichnis
VII
3.2.3 Vergleich der Ergebnisse aus der 13C-MR-Spektroskopie für dieRyR1-Genotypen
161
3.2.4 Vergleich der Ergebnisse aus der 13C-MR-Spektroskopie inAbhängigkeit vom Hampshire-Genanteil
164
3.2.5 Volumen der Mm. longissimi dorsi und des darüber liegenden Fettessowie Fett-zu- Muskel-Verhältnis in einem 2,45 cm MRT-Abschnittvon Schweinen mit unterschiedlichen RyR1-Genotypen
167
3.2.6 Körperzusammensetzung von Schweinen unterschiedlicherZuchtrichtung in einem 2,45 cm MRT-Abschnitt
168
3.2.7 Volumen des Schinkenmagergewebes (einschließlich Knochen)(SMGV) und des darüber liegenden Fettes (SFETTV) sowie desgesamten Schinkens (SGV) und das Fett-zu-Muskel-Verhältnis(SFMV) in einer 15 cm MRT-Region für Schweine mitunterschiedlichen RyR1-Genotypen bei ca. 30 kg Lebendmasse
170
3.2.8 Volumen der Mm. longissimi dorsi (MLDV) und des darüberliegenden Fettes (FETTV) sowie das Fett-zu-Muskel-Verhältnis(FMV) in einer 10 cm MRT-Region für Schweine mitunterschiedlichen RyR1-Genotypen bei ca. 30 kg Lebendmasse
170
3.2.9 Volumen des Schinkenmagergewebes (einschließlich Knochen)(SMGV) und des darüber liegenden Fettes (SFETTV) sowie desgesamten Schinkens (SGV) und das Fett-zu-Muskel-Verhältnis(SFMV) in einer 15 cm MRT-Region für Schweine mitunterschiedlichen RyR1-Genotypen bei ca. 60 kg Lebendmasse
171
3.2.10 Volumen der Mm. longissimi dorsi (MLDV) und des darüberliegenden Fettes (FETTV) sowie das Fett-zu-Muskel-Verhältnis(FMV) in einer 10 cm MRT-Region für Schweine mitunterschiedlichen RyR1-Genotypen bei ca. 60 kg Lebendmasse
171
3.2.11 Volumen des Schinkenmagergewebes (einschließlich Knochen)(SMV) und des darüber liegenden Fettes (SFETTV) sowie desgesamten Schinkens (SGV) und das Fett-zu-Muskel-Verhältnis(SFMV) in einer 15 cm MRT-Region für Schweine mitunterschiedlichen RyR1-Genotypen bei ca. 90 kg Lebendmasse
172
3.2.12 Volumen der Mm. longissimi dorsi (MLDV) und des darüberliegenden Fettes (FETTV) sowie das Fett-zu-Muskel-Verhältnis(FMV) in einer 10 cm MRT-Region für Schweine mitunterschiedlichen RyR1-Genotypen bei ca. 90 kg Lebendmasse
172
3.2.13 Volumen des Schinkenmagergewebes (einschließlich Knochen)(SMV) und des darüber liegenden Fettes (SFETTV) sowie desgesamten Schinkens (SGV) und das Fett-zu-Muskel-Verhältnis(SFMV) in einer 15 cm MRT-Region für Schweine mitunterschiedlichen RyR1-Genotypen zwischen 26,4 bis 97,2 kgLebendmasse
173
3.2.14 Volumen der Mm. longissimi dorsi (MLDV) und des darüberliegenden Fettes (FETTV) sowie das Fett-zu-Muskel-Verhältnis(FMV) in einer 10 cm MRT-Region für Schweine mitunterschiedlichen RyR1-Genotypen zwischen 26,4 bis 97,2 kgLebendmasse
174

Tabellenverzeichnis
VIII
3.2.15 Chemische Körperzusammensetzung für Schweine mitunterschiedlichen RyR1-Genotypen zwischen 26,4 bis 97,2 kgLebendmasse
174
3.2.16 DXA-Körperzusammensetzung für Schweine mit unterschiedlichenRyR1-Genotypen zwischen 26,4 bis 97,2 kg Lebendmasse, bei einerdurchschnittlicher DXA-Lebendmasse von 63,06 kg (124,85Lebenstage)
175
3.2.17 Kotelettfläche, Rückenspeckfläche und Speck-zu-Muskelverhältnis(SMV) für Schweine mit unterschiedlichen RyR1-Genotypenzwischen 26,4 bis 97,2 kg Lebendmasse
175
3.2.18 Schinken-, Schulter-, Blatt- und Rückengewicht für Schweine mitunterschiedlichen RyR1-Genotypen zwischen 26,4 bis 97,2 kgLebendmasse
176
3.2.19 Leber-, Herz- und Nierengewichte für Schweine mit unterschiedlichenRyR1-Genotypen zwischen 26,4 bis 97,2 kg Lebendmasse
176
3.2.20 Vergleich der Genauigkeit verschiedener Magnetresonanz-Tomographie-Volumenmessungen in einer 10 cm Sektion beider Mm.longissimi dorsi [Mm.l.d] und der entsprechenden Fettauflage[Mm.l.d-Fett] (Versuch 1: 6,1 – 15 kg Lebendmasse [LM], n=15;Versuch 2: 30-90 kg LM, n=46; Versuch 1+2, n=61) und einer 15 cmSektion des Schinken-Magergewebes [Schinken-Mag.] sowieentsprechender Fettauflage [Schinken-Fett] (allein Versuch 2) zurSchätzung der chemischen Körperzusammensetzung
182
3.2.21 Modelle aus der Stepwise-Regressions-Analyse für Merkmale aus derMR-Tomographie, die am besten die chemischeSchlachtkörperzusammensetzung vorhersagen
183
3.2.22 Genauigkeit der Schätzung der Körperzusammensetzung beimSchwein mittels MRT in vivo
185
3.2.23 Magergewebemasse (MGM), Fettgewebemasse (FGM), undKnochenmineralmasse (KMM) aus der DXA-Analyse in Schweinenmit unterschiedlichen Ryanodin-Rezeptor-1-Genotypen beidurchschnittlich 12,55 kg Lebendmasse und 53,29 Lebenstagen
187
3.2.24 Magergewebemasse (MGM), Fettgewebemasse (FGM), undKnochenmineralmasse (KMM) aus der DXA-Analyse in Schweinenmit unterschiedlichen Ryanodin-Rezeptor-1-Genotypen beidurchschnittlich 32,31 kg Lebendmasse und 84,11 Lebenstagen
187
3.2.25 Magergewebemasse (MGM), Fettgewebemasse (FGM), undKnochenmineralmasse (KMM) aus der DXA-Analyse in Schweinenmit unterschiedlichen Ryanodin-Rezeptor-1-Genotypen beidurchschnittlich 65,09 kg Lebendmasse und 124,75 Lebenstagen
188
3.2.26 Magergewebemasse (MGM), Fettgewebemasse (FGM), undKnochenmineralmasse (KMM) aus der DXA-Analyse in Schweinenmit unterschiedlichen Ryanodin-Rezeptor-1-Genotypen beidurchschnittlich 88,72 kg Lebendmasse und 160,41 Lebenstagen
188
3.2.27 Vergleich von chemischem und DXA-Fettgehalt (%) inunterschiedlichen RyR1-Genotypen innerhalb der 10 kg Gruppe
190
3.2.28 Vergleich von chemischem und DXA-Magergewebegehalt (%) inunterschiedlichen RyR1-Genotypen innerhalb der 10 kg Gruppe
190

Tabellenverzeichnis
IX
3.2.29 Vergleich von chemischem Rohaschegehalt (%) und DXA-Knochenmineralgehalt (%) in unterschiedlichen RyR1-Genotypeninnerhalb der 10 kg Gruppe
190
3.2.30 Vergleich von chemischem und DXA-Fettgehalt (%) inunterschiedlichen RyR1-Genotypen im Lebendmassebereich von30 - 90 kg
191
3.2.31 Vergleich von chemischem und DXA-Magergewebegehalt (%) inunterschiedlichen RyR1-Genotypen im Lebendmassebereich von30 - 90 kg
191
3.2.32 Vergleich von chemischem Rohaschegehalt und DXA-Knochenmineralgehalt in unterschiedlichen RyR1-Genotypen imLebendmassebereich von 30 – 90 kg
192
3.2.33 Genauigkeit der DXA-Messung (in vivo) zur Schätzung derchemischen Zusammensetzung der rechten Schlachthälfte
194
3.2.34 Vergleich der Schätzgenauigkeit für den chemischen Fettgehalt in derrechten Schlachthälfte (%) aus Ergebnissen der MR-Tomographie undDualenergie-Röntgenabsorptiometrie
196
3.2.35 Vergleich der Schätzgenauigkeit für den chemischen Fettgehalt imgesamten Schlachtkörper (%) aus Ergebnissen der MR-Tomographieund Dualenergie-Röntgenabsorptiometrie
196
3.2.36 Vergleich der Schätzgenauigkeit für den chemischenMagergewebegehalt (Wasser + Protein) der rechten Schlachthälfte (%)aus Ergebnissen der MR-Tomographie und Dualenergie-Röntgenabsorptiometrie
197
3.2.37 Tageszunahme (TZ10) und Magergewebezunahme (MZ10) inSchweinen mit unterschiedlichen RyR1-Genotypen von Geburt (1,2kg) bis zur DXA-Messung bei ca. 10 kg Lebendmasse
199
3.2.38 Tageszunahme (TZ1030) und Magergewebezunahme (MZ1030) inSchweinen mit unterschiedlichen RyR1-Genotypen imLebendmasseabschnitt von ca. 10 bis 30 kg
199
3.2.39 Wachstum und Futtereffizienz in Schweinen mit unterschiedlichenRyR1-Genotypen im Lebendmasseabschnitt von ca. 30 bis 65 kg
200
3.2.40 Wachstum und Futtereffizienz bei Schweinen mit unterschiedlichenRyR1-Genotypen im Lebendmassebereich von ca. 65 bis 90 kg
200
3.2.41 Protein-, Fett- und Knochenmineralansatzraten (g/d) in Schweinen mitunterschiedlichen RyR1-Genotypen bei einer Fütterung mit 95% dergeschätzten freiwilligen Futteraufnahme
203
3.2.42 Muskelfaserquerschnittsflächen der Muskelfasertypen I, IIa und IIbsowie „Kotelettflächen“ von Schweinen mit unterschiedlichen RyR1-Genotypen bei einem Durchschnittsgewicht von 83,48 kg und 152,77Lebenstagen
207
3.2.43 Flächenanteile der Muskelfasertypen I, IIa und IIb inunterschiedlichen RyR1-Genotypen
209
3.2.44 Prozentuale Verteilung der Anzahl der Muskelfasertypen I, IIa und IIbin unterschiedlichen RyR1-Genotypen
209
3.2.45 Gesamtanzahl Muskelfasern im M. longissimus dorsi in Abhängigkeitvom RyR1-Genotyp
211

Tabellenverzeichnis
X
4.1.1 Stoffwechselreaktion und Körperzusammensetzung in Schweinenunterschiedlicher RyR1-Genoytpen mit einer Lebendmasse von ca. 10kg
227
4.1.2 Stoffwechselreaktion und Körperzusammensetzung in verschiedenenZuchtlinien
229
5.1 Anzahl Tiere innerhalb der RyR1-Genotypen - Gesamtmaterial 2605.1a Number of animals within RyR1 genotype – total material 263
A1 Volumen des Schinkenmagergewebes einschließlich Knochen (SMV)und des subkutanen Fettes einschließlich Haut (SFETTV) sowie desgesamten Schinkens (SGV) und das Fett-zu-Muskel-Verhältnis(SFMV) in einer 15 cm MRT-Region für Schweine mitunterschiedlicher Rassezusammensetzung bei ca. 30 kg Lebendmasse
339
A2 Volumen der Mm. longissimi dorsi (MLDV) und des subkutanenFettes einschließlich Haut (FETTV) sowie das Fett-zu-Muskel-Verhältnis (FMV) in einer 10 cm MRT-Region für Schweine mitunterschiedlicher Rassezusammensetzung bei ca. 30 kg Lebendmasse
339
A3 Volumen des Schinkenmagergewebes einschließlich Knochen (SMV)und des subkutanen Fettes einschließlich Haut (SFETTV) sowie desgesamten Schinkens (SGV) und das Fett-zu-Muskel-Verhältnis(SFMV) in einer 15 cm MRT-Region für Schweine mitunterschiedlicher Rassezusammensetzung bei ca. 60 kg Lebendmasse
340
A4 Volumen der Mm. longissimi dorsi (MLDV) und des subkutanenFettes einschließlich Haut (FETTV) sowie das Fett-zu-Muskel-Verhältnis (FMV) in einer 10 cm MRT-Region für Schweine mitunterschiedlicher Rassezusammensetzung bei ca. 60 kg Lebendmasse
340
A5 Volumen des Schinkenmagergewebes einschließlich Knochen (SMV)und des subkutanen Fettes einschließlich Haut (SFETTV) sowie desgesamten Schinkens (SGV) und das Fett-zu-Muskel-Verhältnis(SFMV) in einer 15 cm MRT-Region für Schweine mitunterschiedlicher Rassezusammensetzung bei ca. 90 kg Lebendmasse
341
A6 Volumen der Mm. longissimi dorsi (MLDV) und des subkutanenFettes einschließlich Haut (FETTV) sowie das Fett-zu-Muskel-Verhältnis (FMV) in einer 10 cm MRT-Region für Schweine mitunterschiedlicher Rassezusammensetzung bei ca. 90 kg Lebendmasse
341
A7 Volumen des Schinkenmagergewebes einschließlich Knochen (SMV)und des subkutanen Fettes einschließlich Haut (SFETTV) sowie desgesamten Schinkens (SGV) und das Fett-zu-Muskel-Verhältnis(SFMV) in einer 15 cm MRT-Region für männliche Kastraten (M)und weibliche Schweine (W) bei ca. 30 kg Lebendmasse
342
A8 Volumen der Mm. longissimi dorsi (MLDV) und des subkutanenFettes einschließlich Haut (FETTV) sowie das Fett-zu-Muskel-Verhältnis (FMV) in einer 10 cm MRT-Region für männlicheKastraten (M) und weibliche Schweine (W) bei ca. 30 kgLebendmasse
342

Tabellenverzeichnis
XI
A9 Volumen des Schinkenmagergewebes einschließlich Knochen (SMV)und des subkutanen Fettes einschließlich Haut (SFETTV) sowie desSchinkengesamtvolumens (SGV) und das Fett-zu-Muskel-Verhältnis(SFMV) in einer 15 cm MRT-Region für männliche Kastraten (M)und weibliche Schweine (W) bei ca. 60 kg Lebendmasse
343
A10 Volumen der Mm. longissimi dorsi (MLDV) und des subkutanenFettes einschließlich Haut (FETTV) sowie das Fett-zu-Muskel-Verhältnis (FMV) in einer 10 cm MRT-Region für männlicheKastraten (M) und weibliche Schweine (W) bei ca. 60 kgLebendmasse
343
A11 Volumen des Schinkenmagergewebes einschließlich Knochen (SMV)und des subkutanen Fettes einschließlich Haut (SFETTV) sowie desSchinkengesamtvolumens (SGV) und das Fett-zu-Muskel-Verhältnis(SFMV) in einer 15 cm MRT-Region für männliche Kastraten (M)und weibliche Schweine (W) bei ca. 90 kg Lebendmasse
344
A12 Volumen der Mm. longissimi dorsi (MLDV) und des subkutanenFettes einschließlich Haut (FETTV) sowie das Fett-zu-Muskel-Verhältnis (FMV) in einer 10 cm MRT-Region für männlicheKastraten (M) und weibliche Schweine (W) bei ca. 90 kgLebendmasse
344
A13 Prozentuale Konzentrationsveränderung von Glykogen und Kreatinsowie für die Dauer der Halothanzufuhr in Schweinen mitunterschiedlichen RyR1-Genotypen nach Modell I
345
A14 Glykogenniveau im Ruhezustand, prozentuale Konzentrations-veränderung von Glykogen und Kreatin sowie Dauer derHalothanzufuhr in Kreuzungslinien mit unterschiedlichem Hampshire-Genanteil nach Modell II
345
A15 Auswahl potentieller Quantitativer Merkmalsgenorte (QTL) für denKörperfettgehalt bei Schwein, Maus und Mensch
367

Abbildungsverzeichnis
XII
Abbildungsverzeichnis
Abbildung Titel Seite
2.1.1 Eigenschaften wichtiger Atomkerne am Beispiel eines 4,7 TeslaMagnetfeldes
5
2.1.2 Schema zur Ausrichtung der Moleküle (Atomkerne) in einem festenMagnetfeld B0 am Beispiel einer beliebigen Kohlenstoffverbindung.
8
2.1.3 Schematische Darstellung zur Signalverarbeitung am Beispiel eines31P-Magnet-Resonanz-Spektrums
9
2.1.4 31P-Spektrum des M. biceps femoris eines Schweines in vivo 112.1.5 Verlauf der Konzentration von C1-Glykogen (100,5 ppm), Kreatin
(157 ppm) und βC1-Glukose (96,6 ppm) nach Verabreichung einer28 ml Natriumchlorid-Lösung mit 15 g Dextrose und 0,1 g D-Glukose-13C6 bei einem 10,9 kg schweren Schwein (RyR1 = Nn) mit5/8 Duroc-Genanteilen
16
2.2.1 Einflussfaktoren auf die Körperzusammensetzung 18
2.2.2 Entwicklung des Protein-zu-Wasser-Verhältnisses in der fettfreienMasse bzw. Magergewebemasse beim Schwein
22
2.2.3 Abhängigkeit des Wassergehalts im Rückenfettgewebe von derRückenspeckdicke und Rasse bei ∅ 62,5 kg Körpergewicht
24
2.2.4 Nutzbare physikalische und chemische Eigenschaften zur Messungder Körperzusammensetzung
25
2.2.5 Körperkompartimente als Grundlage für verschiedene Modelle derKörperzusammensetzung
27
2.2.6 Geschichtliche Entwicklung der Methoden zur Bestimmung derKörperzusammensetzung bei Nutztieren
33
2.2.7 Veränderung der chemischen Körperzusammensetzung bei Schweinenvon 5 bis 97 kg Lebendmasse
34
2.2.8 Messung der Rückenspeckdicke mittels „Ruler“ oder „Back FatProbe“
41
2.2.9 Frequenzbereiche für elektromagnetische Strahlung 442.2.10 Funktionsweise der Spiral-Röntgen-Computertomographie 532.2.11 Kernspin und Präzession 562.2.12 Bau- und Funktionsprinzip eines MR-Tomographen 562.2.13 Darstellung von MR-„Echo“-Bildern, die sich aus einer „Multi-Slice-
Multi-Echo“-Puls-Sequenz ergeben59
2.2.14 Veränderung von Signalintensität und Bildkontrast während der MR-Bilderzeugung
59
2.2.15 Beispiel für ein MR-Axialschnittbild im Bereich der Nieren 602.2.16 Beispiel für ein MR-Axialschnittbild im Beckenbereich 602.2.17 Schema für den Ablauf der Bildverarbeitung mittels Cluster-Analyse 612.2.18 Übersicht zu statistischen Verfahren für die quantitative
Bildauswertung67
2.2.19 Drei-Komponenten-Modell der Körperzusammensetzung für DXA-Untersuchungen
73

Abbildungsverzeichnis
XIII
2.2.20 Positionierung eines Probanden (Kalb) für einen Ganzkörperscan 742.2.21 Beispiel für das Ergebnis einer DXA-Studie eines Kalbes 752.2.22 Beispiel für DXA-Daten nach einem Ganzkörperscan eines
totgeborenen Ferkels (1,52 kg Körpermasse) mittels Kleintier-Ganzkörper-Modus
78
2.2.23 Einflussfaktoren auf die Variation in der Knochenmasse undKnochendichte
81
2.2.24 Mineralisierungsgrad von Karpal- bzw. Tarsalgelenken vonSchweinen mit unterschiedlichem Gewicht
82
2.2.25 Mineralisierungsgrad einer Landschildkröte mit 677 g Lebendmasse 822.2.26 Gesamt-Körperzusammensetzung einer Landschildkröte mit 677 g
Lebendmasse83
2.2.27 Darstellung der Beziehung zwischen DXA-R-Werten fürWeichgewebe und dem prozentualen DXA-Fettgehalt
83
2.2.28 Beispiel für klinische Anwendung - Fraktur des rechten Femurs einesSchweins – Verwendung des Schenkelhals-Modus
84
2.2.29 Ultraschallscanner „B-Mode“ und Prinzip des Ultraschallverfahrensnach dem „A-Mode“ zur Ermittlung von Fett- und Muskeldicke
86
2.2.30 Entwicklung der „Ultraschall“-Rückenspeckdicke im Verlauf desWachstums bei verschiedenen Schweinerassen
89
2.2.31 Prinzip der Elektrischen Impedanztomographie 942.2.32 Reaktionen der Neutronenaktivierung zur Ermittlung der elementaren
Körperzusammensetzung103
2.3 Montiertes Schussbiopsiegerät LS1 1113.1.1 Platzierung der 31P-MR-Oberflächenspule über dem M. biceps femoris
eines Schweins114
3.1.2 Experimentelle Anordnung von 13C-MR-Doppelspule undGlykogenstandard
119
3.1.3 In situ 13C-MR-Spektrum von reinem Glykogen 1203.1.4 Aufnahmeparameter und Beispiel für ein 13C-Glykogenspektrum in
vivo121
3.1.5 1H-13C-Spulenanordnung über dem M. biceps femoris in vivo 1223.1.6 Beispiel für ein charakteristisches 13C-Spektrum an der C1-Glykogen-
Position123
3.1.7 Beziehung zwischen Glykogenabbau und Hampshire- bzw. Pietrain-Genanteil
125
3.1.8 Versuchsanordnung zur Tomographie der Tiere bis 20 kgLebendmasse mittels 1H-MR-Oberflächenspule
127
3.1.9 1H-MR-Aufnahmeparameter und Beispielbild für die Darstellung derRückenregion mittels Oberflächenspule
128
3.1.10 Darstellung der Regions of Interest (ROI’s) in den 1H-Magnet-Resonanz-Schnittbildern im Bereich 13./14. Brustwirbel für Tiere bis20 kg Lebendmasse
129
3.1.11 Positionierung eines Probanden im Picker-Vista 1,5 Tesla MR-Tomographen
130

Abbildungsverzeichnis
XIV
3.1.12 Lage der Tiere im Magneten mit gekennzeichnetenSchnittbildregionen zur Volumenbestimmung
132
3.1.13 MR-Axialschnittbild zwischen 13./14. Brustwirbel 1333.1.14 Regionen für die MR-Volumenbestimmung im Thoraxbereich (10
Scheiben, 10 cm)133
3.1.15 Anatomische Hilfspunkte und schematische Darstellung der Regionsof Interest (ROI’s) im Becken-/Hintergliedmaßenbereich
134
3.1.16 Lage des Probanden auf dem DXA-Scanner 1383.1.17 Beispiel für DXA-Ganzkörperergebnisse eines Schweins 1393.1.18 Schema für Teilzerlegung 1433.1.19 Verwendetes Schussbiopsiegerät LS1 mit demontiertem Aufsatz und
in Aktion146
3.1.20 Beispiel für ein frisch entnommenes Bioptat aus dem M. longissimusdorsi eines Schweins
147
3.1.21 Im Versuch unterschiedene Muskelfasertypen 1483.1.22 PCR-Ergebnis für die RyR1-Genotypen: 1523.1.23 Prinzip des verwendeten RyR1 -Gentestes 1523.2.1 Beispiel für den zeitlichen Verlauf der 31P-MR-Spektroskopie in
unterschiedlichen RyR1-Genotypen155
3.2.2 Beispiel für die Veränderung der Stoffwechselsituation (in vivo) nachHalothanbelastung in unterschiedlichen RyR1-Genotypen
156
3.2.3 Beispiel für Stoffwechselveränderungen post mortem im Vergleich derdrei RyR1-Genotypen
157
3.2.4 Veränderung des prozentualen Glykogenniveaus nach Halothanzufuhrbei Schweinen unterschiedlicher RyR1-Genotypen
161
3.2.5 Veränderung des prozentualen Kreatinniveaus nach Halothanzufuhrbei Schweinen unterschiedlicher RyR1-Genotypen
162
3.2.6 Zeitlicher Verlauf des absoluten Glykogenabbaus (µmol/g) nachHalothanbelastung in verschiedenen RyR1-Genotypen in vivo
164
3.2.7 Beispiel für den zeitlichen Verlauf der 13C-MR-Spektren vonSchweinen unterschiedlicher RyR1-Genotypen
165
3.2.8 Glykogenabbau in unterschiedlichen Kreuzungsherkünften 1663.2.9 Beispiele für die Variation der MRT-Axialschnitt-Messwerte (1 - 5)
innerhalb eines Tieres178
3.2.10 Beziehung zwischen Schinkenmasse (kg) der linken Schlachthälfteund dem „Schinken“-Gesamtvolumen (cm³) aus der MR-Tomographie
179
3.2.11 Beziehung zwischen Rückenmasse (kg) der linken Schlachthälfte unddem „Rücken“-Volumen (Mm.l.d + Auflagefett, cm³) aus der MR-Tomographie
180
3.2.12 Beziehung zwischen Kotelettfläche (cm2) der linken Schlachthälfte(13./14. Brustwirbel) und dem „Kotelett“-Volumen (cm³) aus der MR-Tomographie
180
3.2.13 Beziehung zwischen Rückenspeckfläche (cm2) der linkenSchlachthälfte (13./14. Brustwirbel) und dem „Rückenspeck“-Volumen (cm³) aus der MR-Tomographie
181

Abbildungsverzeichnis
XV
3.2.14 Bestimmtheitsmaße und „Root Mean Square Errors“ für dieSchätzung von chemischem Fettgehalt im Schlachtkörper (%) undchemischem Magergewebegehalt (Wasser + Protein) imSchlachtkörper (%) beim Vergleich verschiedenerInformationsquellen aus der in vivo MR-Tomographie zwischen 30und 90 kg Lebendmasse (p ≤ 0,05)
184
3.2.15 Beziehung zwischen chemischem Fettgehalt der rechtenSchlachthälfte (%) und DXA-Gesamtkörperfettgehalt (%) imLebendmassebereich 10 – 90 kg
193
3.2.16 Beziehung zwischen DXA-Gesamtkörper-Fettmasse (kg) und MR-Fettvolumen (cm³) in einem 10 cm MR-Abschnitt über den Mm.longissimi dorsi
197
3.2.17 Entwicklung der Tageszunahmen von Schweinen mitunterschiedlichen RyR1-Genotypen zwischen 5 und 80 kgLebendmasse
200
3.2.18 Entwicklung der Magergewebewachstumsrate in Schweinen mitunterschiedlichen RyR1-Genotypen zwischen 5 und 80 kgLebendmasse
201
3.2.19 Durchschnittliche Futteraufnahme (kg/d) undMagergewebewachstumsrate (g/d) in Abhängigkeit vom Prüfabschnittbei Schweinen mit unterschiedlichen RyR1-Genotypen
201
3.2.20 Wachstumsraten für Protein, Fett und Knochenmineral bei Schweinenmit unterschiedlichen RyR1-Genotypen
204
3.2.21 Vergleich der Muskelfaserquerschnitte in Abhängigkeit vom RyR1-Genotyp
206
3.2.22 Beziehung zwischen Lebendmasse und durchschnittlicherMuskelfaserfläche
208
3.2.23 Beziehung zwischen Muskelfaseranzahl und Muskelfaserfläche 2103.2.24 Beziehung zwischen der auf einheitliche 90 kg Lebendmasse
korrigierten Muskelfaserfläche am M. longissimus dorsi und demPhosphokreatinabbau nach Stressauslösung mittels Halothan am M.biceps femoris (bei ca. 10 kg Lebendmasse)
212
3.2.25 Beziehung zwischen Muskelfaseranzahl am M. longissimus dorsi unddem Phosphokreatinabbau nach Stressauslösung mittels Halothan amM. biceps femoris
213
3.2.26 Beziehung zwischen Muskelfaseranzahl am M. longissimus dorsi unddem Phosphokreatinabbau nach Stressauslösung mittels Halothan amM. biceps femoris innerhalb der RyR1-Genotypen
213
4.1.1 Beispiel für den in vivo Zeitverlauf der Phosphokreatin-Konzentration(PCr) und des pH-Wertes von Schweinen mit unterschiedlichenRyanodin-Rezeptor-1-Genotypen nach Halothanbelastung
218
4.1.2 3D-Rekonstruktion des Ryanodin- Rezeptors-1 2244.1.3 Schematische Darstellung der Kontaktregion zwischen T-Tubulus und
terminaler Zisterne des sarkoplasmatischen Retikulums in derSkelettmuskulatur (Trias-Region)
225
4.1.4 Absolutes Glykogenniveau (µmol/g) im Ruhezustand und nachHalothanbelastung in verschiedenen Ryanodin-Rezeptor-1-Genotypen
228

Abbildungsverzeichnis
XVI
4.1.5 Veränderungen in der Körpertemperatur nach Halothanbelastung beiunterschiedlichen Ryanodin-Rezeptor-1-Genoytpen in vivo und postmortem für das Gesamtmaterial (31P + 13C)
228
4.1.6 Körperzusammensetzung und Stoffwechselreaktion am Beispiel dreiverschiedener Zuchtlinien (Modell I)
230
4.1.7 Muskelfaserquerschnittsflächen bei Schweinen mit unterschiedlichenRyanodin-Rezeptor-1-Genotypen
232
4.1.8 Muskelfaserfläche (Typ IIa) and Phosphokreatin-Abbau (%) beiSchweinen mit unterschiedlichen RyR1-Genotypen
232
4.1.9 Muskelfaseranzahl and Phosphokreatin-Abbau (%) bei Schweinen mitunterschiedlichen RyR1-Genotypen
233
4.1.10 Schematischer Vergleich der Muskelkontraktionszyklen vonSchweinen mit ”normaler” Muskulatur und „defekter“ Muskulatur
237
4.2.1 Entwicklung von Muskel- und Fettauflagevolumen in einem 10 cmMRT-Bereich des Rückens bei Schweinen mit unterschiedlichenRyR1-Genotypen
238
4.2.2 Entwicklung des Magergewebewachstums und derLebendmassezunahme bei Schweinen mit unterschiedlichen RyR1-Genotypen
241
4.2.3 Durchschnittlicher Futterverzehr verschiedener Rassen bzw.Kreuzungen während der Leistungsprüfung von ca. 30 – 100 kgLebendmasse
246
4.2.4 Vergleich des DXA-Fettgehaltes (%) bei Tieren der Linie „WeißePietrain“ mit unterschiedlichen RyR1-Genotypen bei a) 30 kg, b) 50kg, c) 70 kg und d) 90 kg Lebendmasse
247
4.2.5 Anstieg des Gehalts an obese mRNA in Abhängigkeit von derFettmasse beim Schwein
248
4.2.6 Beziehungen zwischen Fettgewebe, genetischer Steuerung vonZentren im Hypothalamus und der Futteraufnahmeregulation
257

Tierversuche
XVII
Genehmigung der Tierversuche
Die beschriebenen Experimente am Agricultural Research Service (ARS) des United
States Department of Agriculture (USDA) in Beltsville, MD, USA und an der Howard
University in Washington, DC, wurden in Übereinstimmung mit den durch das “USDA-
ARS, Beltsville Animal Care and Use committee“ bestätigten Versuchsprotokollen
durchgeführt.
Weitere im Text erwähnte Tierversuche, die unter Mitwirkung des Autors am
Lehr- und Versuchsgut Oberschleißheim der Tierärztlichen Fakultät der Ludwig-
Maximilians-Universität München durchgeführt wurden, erhielten im Rahmen des
Tierschutzgesetzes eine Genehmigung durch die Regierung von Oberbayern mit den
folgenden Aktenzeichen:
211-2531.2-57/2000 - Untersuchungen zur Körperzusammensetzung und
Knochenmineraldichte am Kalb mittels Dualenergie-Röntgenabsorptiometrie (DEXA)
und
211-2531.2-21/2000 - Untersuchungen zur Körperzusammensetzung und
Knochenmineraldichte am wachsenden Schwein mittels Dualenergie-
Röntgenabsorptiometrie (DEXA).

Einleitung
1
Fortschritt in der Wissenschaft entsteht dadurch, dass man mühsam einen Stein auf denanderen legt, nicht indem man plötzlich einen herrlichen Palast errichtet.
Advance in science comes by laying brick upon brick, not by sudden erection of fairy palaces.
- Julian S. Huxley -
1 Einleitung
In „Industrieländern“ stellt die Quantität der zu produzierenden Lebensmittel kein
Problem mehr dar (KALM, 1998), so dass das (ideelle) Wohlbefinden der Tiere, die
Produkt- und Prozessqualität sowie die Umweltverträglichkeit zunehmendes Interesse
bei den Konsumenten und damit auch in der Wissenschaft hervorrufen (KALLWEIT,
1992, JAMISON, 2000, WENK, 2000).
Weitreichende Entwicklungen in den Bereichen der Mess- und Computertechnik
sowie der Biotechnologie in Kombination mit bewährten konventionellen Verfahren
ebnen den Weg, bisher nicht zu beantwortende wissenschaftliche Fragestellungen zu
beantworten.
So eröffnet die Kombination von neuesten genetischen Testverfahren mit
hochmodernen nicht invasiven Methoden zur Ermittlung von Gewebezusammensetzung
und Stoffwechselprozessen in vivo ein breites Feld von Möglichkeiten, die Effekte
unterschiedlichster Einflussfaktoren auf morphologische und physiologische Merkmale
in Nutztieren im Detail zu studieren. Im Mittelpunkt stehen neben definierbaren
Umweltfaktoren die Polymorphismen an Genorten, die quantitative und qualitative
Merkmale steuern. Besonderes Augenmerk wird wieder zunehmend - auch im Rahmen
der Leistungsprüfung von Zuchttieren - auf Merkmale gerichtet, die den Zustand bzw.
das Wohlbefinden der Tiere erklären. Sie beeinflussen letztendlich in hohem Maße die
angestrebte Produkt- und/oder Prozessqualität und prägen damit den Nutzen bzw. den
Gebrauchs- und/oder Genusswert für den Menschen (SCHÖNMUTH, 1987, FÖRSTER,
1992, SCHMITTEN, 1993, GLODEK, 1996, GELDERMANN, 1996, HARDGE, 1999,
KALLWEIT, 1992, 1997, VON LENGERKEN, 1997).
Deutschlands Landwirte erzielen ca. 60 Prozent ihres Einkommens durch
Tierproduktion, wobei die Bundesrepublik mit ~19,8 Mrd. € - nach Frankreich mit
~22,2 Mrd. € - das zweitwichtigste Erzeugerland für tierische Produkte in der
Europäischen Union ist (Stand 1998 - WINDHORST, 1998). Obwohl Deutschland mit
enormen Strukturproblemen im Schweinesektor zu kämpfen hat, liegen die deutschen

Einleitung
2
Schweineproduzenten mit einer Jahresproduktion von 3,83 Mio. Tonnen (WEIß, 2001a)
bzw. 40,45 Mio. Schweinen (WEIß, 2001b) im europäischen Vergleich an der Spitze.
Die deutsche Jahresproduktion deckt jedoch nur ca. 85 % des Bedarfs an
Schweinefleisch in Deutschland (ZMP, 2001, RIEU und VAN FERNEIJ, 2001). Wollen die
deutschen Schweineproduzenten weitere Marktanteile zurückgewinnen, muss auf allen
Ebenen vom Erzeuger bis zum Vermarkter ein wettbewerbsfähiges Produkt
gewährleistet werden.
In Abhängigkeit von traditionellen und gesellschaftlichen Gegebenheiten stellt
das Schwein mit seinen Produkten, vor allem dem Fleisch als Proteinquelle (bis zu 5 -
20 % des täglichen Bedarfs) und dem Fett als Energielieferant bzw. Geschmacksträger,
einen wesentlichen Bestandteil der menschlichen Ernährung dar. Es liefert bis zu 20 %
der Nährstoffe, Mineralstoffe und Vitamine des täglichen Bedarfs des Menschen. Da
der Verbrauch an Fett (Energie) durch physisch weniger beanspruchende Arbeit
zurückging, wurden ab Mitte des 20. Jahrhunderts stärker bemuskelte Schweine mit
einem niedrigen Fettanteil für die Weiterzucht verwendet (HERTRAMPF, 1998). Die
einseitige Selektion auf erhöhten Fleischanteil hatte die bekannten negativen
Auswirkungen auf Stressstabilität und Fleischbeschaffenheit sowohl in den Vaterrassen
als auch zum Teil in den Mutterrassen. Mit dem zwar intensiv durchgeführten
konventionellen und oft fatal ausgehenden Halothanbelastungstest konnte nicht
eindeutig zwischen einerseits homozygot normalen und heterozygoten Tieren bzw.
andererseits zwischen heterozygoten und homozygoten Defektallelträgern
unterschieden werden (EIKELENBOOM and MINKEMA, 1974, EIKELENBOOM, 1980).
Beispiele für die Unzulänglichkeit des Halothantestes finden sich in den Arbeiten von
REMPEL et al. (1993), SCHMITTEN (1993), KNORR et al. (1994), HAMMEL u.a. (1996),
KNYAZEV et al. (1998) wieder. Erst nachdem FUJII et al. (1991) eine signifikante
Beziehung zwischen der malignen Hyperthermie und einer Mutation an der Position
1843 der cDNA des Chromosomen 6 von Cytosin zu Thymidin feststellten und
entsprechende DNA-Testverfahren für diese Mutation entwickelt wurden, war es
möglich, die drei Genotypen eindeutig zu differenzieren (BREM und BRENIG, 1992,
BRENIG und BREM, 1992, FÖRSTER u.a., 1992). Der betroffene Genort kodiert den Ca2+-
Freisetzungskanal oder Ryanodin-Rezeptor 1 (RyR1) des sarkoplasmatischen
Retikulums der Skelettmuskulatur.
Besondere Aufmerksamkeit konnte jetzt den morphologischen und
physiologischen Zusammenhängen in den heterozygoten Tieren im Vergleich zu den

Einleitung
3
beiden homozygoten Genotypen gewidmet werden, da noch immer unterschiedliche
Meinungen und Erkenntnisse über die Belastungsanfälligkeit und züchterische
Weiterverwendung dieses Genotyps existieren (IRGANG, 2001). Ausgehend von der
phänotypischen Reaktion auf Halothanzufuhr, wie Muskelstarre, Herzrhythmus- und
Atmungsstörungen, wurde postuliert, dass Halothan zu 95 % maligne Hyperthermie in
den homozygot „stressanfälligen“ (nn) Schweinen induziert, aber keinen sichtbaren
Effekt bei den homozygot „stressstabilen“ und den heterozygoten Tieren (mit
Ausnahme von ca. 5 %) hervorruft. Diese Annahme führte zur These, dass die durch
Halothannarkose provozierte maligne Hyperthermie durch ein rezessives Gen an einem
autosomalen Genort gesteuert wird, wobei beide Allele eine unvollständige Penetranz
zeigen sollen.
Da eine erhöhte Stressreaktion vornehmlich mit einer fettarmen
Körperzusammensetzung und glykolytisch ausgerichteter Muskelstruktur
zusammenhängt, wurden die drei Genotypen als Modell für die Validierung der
Einsatzmöglichkeiten und der Genauigkeit verschiedener In-vivo-Techniken eingesetzt.
Speziell im Rahmen der Leistungsprüfung von Zuchttieren ist es erforderlich,
fortwährend nach genaueren (aber auch praktikablen) Methoden für die verschiedenen
Merkmale der Mast- und Schlachtleistung zu suchen. Daneben müssen weitere
Prüfmethoden etabliert werden, die das Wohlbefinden bzw. die „Fitness“ der Tiere
besser beschreiben. Eine alleinige Prüfung der Fleischqualität als korreliertes Merkmal
reicht nicht aus.
Magnetresonanz-Tomographie und Dualenergie-Röntgenabsorptiometrie bzw.
Magnetresonanz-Spektroskopie sowie Muskel-Schussbiopsie wurden verwendet, um
Beziehungen zwischen Merkmalen der In-vivo-Körperzusammensetzung bzw. des
Muskelstoffwechsels sowie der Muskelfaserstruktur zu analysieren.
Im Gegensatz zur sichtbaren Reaktion während des Halothantestes wurde
erwartet, dass die heterozygoten Defektallelträger mit einer intermediären
Stoffwechselveränderung auf Halothanzufuhr reagieren würden.
Zugleich sollte die Frage beantwortet werden, ob bereits die Untersuchung von
Tieren in sehr jungem Alter (4 - 12 Wochen) Rückschlüsse auf die Stressanfälligkeit
und ihre spätere Gewebezusammensetzung zulässt.

Methodenübersicht
4
2 Methodenübersicht
2.1 Muskelstoffwechseluntersuchungen in vivo mittels Magnetresonanz-Spektroskopie (MRS)
Neben der Magnetresonanz-Tomographie (MRT) gehört die (ortsaufgelöste)
Magnetresonanz-Spektroskopie (MRS) in vivo zu den spektakulärsten Entwicklungen,
die auf dem Phänomen der Magnetresonanz beruhen (BOESIGER, 1991, BAULAIN und
HENNING, 2001). Speziell seit der Entdeckung des Phänomens des kernmagnetischen
Resonanzverhaltens durch BLOCH und PURCELL im Jahre 1946 wird diese Eigenschaft
einiger Atomkerne intensiv in der Physik, Chemie und Biochemie genutzt (CERDAN und
SEELIG, 1990). Für biologische Proben ist die Magnetresonanz-Spektroskopie ohne
Vorbehalt nutzbar, da die zelluläre Biochemie in intaktem Gewebe bzw. originären
physiologischen Situationen nicht invasiv und wiederholt untersucht werden kann
(RADDA, 1992). Neben Körperflüssigkeiten, Zell- und Gewebeextrakten, perfundierten
Zellkulturen und Bioptaten (in vitro) (ZUPKE und FOY, 1995) ist diese Methodik auch
am lebenden Tier bzw. Embryo (in vivo) für die Erfassung von akuten und/oder
zeitabhängigen Stoffwechselvorgängen geeignet und „gewährt einen nicht invasiven
Einblick in die metabolische Seele der Zelle“ (AVISON et al., 1986). Die Spektren
besitzen eine hohe Spezifität und geben unmittelbar eine molekulare Information
wieder. Um z.B. die Isotope 1H, 19F, 31P und 13C (teilweise erst nach Anreicherung mit
markierten Substraten) in ihrem natürlichen Vorkommen messen zu können, benötigt
man jedoch in vivo Konzentrationen in der Größenordnung von mindestens 1 mmolar
(AVISON et al., 1986, CERDAN und SEELIG, 1990, LEIBFRITZ, 1996). Sowohl die
Sensitivität als auch die Spektralauflösung hängen im Wesentlichen von der
Magnetfeldstärke ab (Abb. 2.1.1). Ein Magnetfeld der Stärke 8,4 - 18,8 Tesla und ein
Objektdurchmesser von 5 - 10 mm wird für Extrakte, Zellen und „Bioflüssigkeiten“
verwendet, während Spektrometer von 1,5 – 4,7 (teilweise bis zu 9) Tesla für
Untersuchungen an lebenden Probanden genutzt werden. Intermediäre Feldstärken und
Objektdurchmesser sind für Organmodelle bzw. Tiere geeignet.
Stoffwechselbestandteile innerhalb lebender Gewebe oder Gewebeproben können mit
Hilfe der MRS nur bestimmt werden, wenn sie hoch mobil im Zytosol bzw. im
Zellzwischenraum vorkommen. Membrangebundene Moleküle und „feste“ Bestandteile
sind durch Linienerweiterung oder dipolare Kopplung gewöhnlich im Spektrum nicht
sichtbar (LEIBFRITZ, 1996).

Methodenübersicht
5
Varian 4,7 Tesla
Horizontalmagnet
mit 33 cm
Innendurchmesser
Isotop Kern- Resonanz- Natürliche Relative spin frequenz Häufigkeit Empfind- bei 4,7 T lichkeit
[MHZ] % %
1H* 1/2 200,11 99,985 1002H 1 30,72 0,015 0,963H 1/2 213,45 - 121,36
12C 0 - 98,89 013C 1/2 50,33 1,11 1,59
31P 1/2 80 100 6,63
* 1H - Resonanzfrequenz bei 1,5 T: 63,87 MHz
Abbildung 2.1.1: Eigenschaften wichtiger Atomkerne am Beispiel eines 4,7 TeslaMagnetfeldes
Weitere Anwendungsgebiete, die auf dem Phänomen der Magnetresonanz
beruhen, umfassen in der Physik bzw. Materialforschung die Strukturanalyse von festen
Stoffen, in der Chemie die Untersuchung der Zusammensetzung und der molekularen
Strukturen von Stoffen, während Biochemiker und Biophysiker die makromolekularen
Bestandteile und Veränderungen u.a. von Proteinen, DNA und Membranen (DAVIES
und WLODAWER, 1995) mittels hochauflösender Magnetresonanz-Techniken
analysieren (RADDA, 1992).
Die Magnetresonanz-Spektroskopie (MRS) basiert ebenso wie die
Magnetresonanz-Tomographie (MRT, Kapitel 2.2.3.2.5.2) auf dem Phänomen der
Magnetresonanz, welches die Wechselwirkung eines starken Magnetfeldes mit einem
spezifischen Typ von Atomkernen und Molekülen beinhaltet. Nur Atomkerne (Isotope),
die einen „Drehimpuls“ (Kernspin) besitzen, wie beispielsweise die biologisch
interessanten 1H (Wasserstoff) oder 2H (Deuterium schwerer Wasserstoff), 13C
(Kohlenstoff), 15N (Stickstoff), 17O (Sauerstoff), 19F (Fluor), 23Na (Natrium), 31P
(Phosphor), 39K (Kalium) und 87Rb (Rubidium) zeigen diese Wechselwirkung und
werden „MR-aktiv“ (RADDA, 1992). Jedes dieser Isotope besitzt ein kernmagnetisches
Moment - die Voraussetzung zur Interaktion mit dem Magnetfeld. Zur Veran-
schaulichung kann man annehmen, dass die Atomkerne mit einem Kernspin ≠ 0, den
nur Atomkerne mit ungerader Protonen- und Neutronenzahl aufweisen, wie kleine
Stabmagneten reagieren und sich antiparallel (entgegengesetzt) oder parallel

Methodenübersicht
6
(gleichgerichtet) entlang der Feldlinien eines stärkeren Magnetfeldes (B0) ausrichten
(Abb. 2.1.2). Kerne mit einem Kernspin = 1/2 eignen sich besonders für die MR-
Spektroskopie (-Tomographie), da sie nur exakt zwei Energieniveaus, parallel (+1/2)
oder antiparallel (-1/2), zu den Feldlinien des Magnetfeldes (B0) einnehmen können.
Dazu zählen neben Wasserstoff (1H) z.B. Phosphor (31P), Fluor (19F) und das
Kohlenstoff-Isotop 13 (13C). Werden „MR-aktive“ Kerne durch elektromagnetische
Strahlung im Radiofrequenzbereich (RF) angeregt, absorbieren sie Energie und ändern
ihre Ausrichtung im Magnetfeld B0 durch die Wechselwirkung zwischen den
kernmagnetischen Momenten, dem Magnetfeld B0 und dem durch die
Radiofrequenzstrahlung hervorgerufenen oszillierenden Magnetfeld B1 (Abb. 2.2.11).
Nach der Anregung kehrt die Kernmagnetisierung zum ursprünglichen
Gleichgewichtszustand (Grundzustand) zurück. Die aufgenommene Energie (im
Radiofrequenzbereich) wird wieder abgegeben. Sie ist für jeden Atomkern (13C oder31P) spezifisch und entspricht exakt der Resonanzfrequenz. Das gyromagnetische
Verhältnis γ (für Wasser: γ = 42.58 MHz/Tesla; HORNAK, 1997-2000) und die Stärke
des Magnetfeldes bestimmen diese kernspezifische Resonanzfrequenz. Entsprechend
der Boltzmann-Gleichung:
NhochNniedrig = e –∆ E / kT = e –hv / kT
nehmen die Differenzen zwischen den Kernen (Nuklei) und damit die Signalintensität
mit einer steigenden Magnetfeldstärke zu. Nhoch und Nniedrig repräsentieren die
Population (insbesondere die Anzahl) der Kerne im höheren bzw. niedrigeren
Energieniveau; k ist die Boltzmann-Konstante (1,3805 x 10-23 Joule/Kelvin), T
kennzeichnet die absolute Temperatur (°Kelvin), h Planck's-Konstante (6,626 x 10-34
Joule x Sekunden), während ∆E = hv das Bohr-Gleichgewicht (mit ∆E =
Energiedifferenzen und v = Frequenz der Nukleus-Veränderungen bzw. Resonanz-
oder Larmorfrequenz) wiedergibt (HAUSSER und KALBITZER, 1989, HORNAK, 1997-
2000, JAMES, 1999, BAERWALDE, 2001).
An diesem Punkt trennen sich die Wege von MR-Tomographie und MR-
Spektroskopie. In MRT-Studien werden zusätzliche Magnetfeldgradienten
(Gradientenspulen) verwendet, so dass das Magnetfeld in Abhängigkeit von der
räumlichen Position variiert. Damit hängt die Resonanzfrequenz zusätzlich von der

Methodenübersicht
7
Position im Magnetfeld ab. Diese Information wird für die Erzeugung von
ortsaufgelösten Bildern genutzt (siehe Kapitel 2.2.3.2.5.2 - MRT).
Für einfache MR-Spektroskopie-Studien werden hingegen keine Feldgradienten
zur Aufzeichnung der Resonanzsignale benötigt. Dadurch werden für einen gegebenen
Kerntyp die einzigen Differenzen in den Resonanzfrequenzen durch sehr kleine
Unterschiede im Magnetfeld, welches an den einzelnen Kernen anliegt, bedingt. Da die
Differenzen im Magnetfeld gewöhnlich durch Variationen in der chemischen Bindung
von einzelnen Kernen (Molekülen) bzw. benachbarten Molekülen verursacht werden,
nennt man sie „chemical shift“ bzw. chemische Verschiebung. Das Ziel jeder MRS-
Studie besteht darin, ein Plot oder Spektrum zu erzeugen, welches die Signalintensität
für jede einzelne Zustandsänderung der chemischen Verschiebung relativ zu einer
Standardfrequenz (ausgedrückt in parts per million - ppm) abbildet (Abb. 2.1.4). Die
Eigenschaften derartiger Spektren können dazu genutzt werden, die chemischen
Bestandteile einer Probe zu identifizieren. Gleichzeitig können unter günstigen
Bedingungen die Flächen unter den lokalen Gipfeln (Peaks) verwendet werden, um die
vorhandenen Substanzen zu quantifizieren. Die Signalintensität und damit die Fläche
unter den lokalen Gipfeln ändert sich proportional zur vorhandenen Menge der
entsprechenden Substanz. Erhöht man die Feldstärke des statischen Magnetfeldes,
verbessert sich das Signal-Rausch-Verhältnis und damit die Sensitivität der Technik
(DECANNIERE, 1993, KOHN, 1997).
Verschiedene Faktoren tragen maßgeblich zur Aufdeckung der Struktur eines
Moleküls bei. Die chemische Verschiebung bzw. die Frequenz, bei welcher der Nukleus
in Resonanzschwingung gerät, wenn ein magnetisches Feld anliegt, gibt Auskunft über
die chemische Umgebung eines Nukleus. Die sogenannte Spin-Spin-Kopplung oder J-
Kopplung klärt die Beziehungen zwischen unterschiedlichen Nuklei auf, die in einer
chemischen Verbindung vorkommen. Ein weiterer Effekt, der Nuklear-Overhauser-
Effekt bzw. die Interaktion zwischen den Dipol-Momenten zweier Nuklei in räumlicher
Nähe, wird genutzt, um die Distanz zwischen den Nuklei zu ermitteln und ist ein
Parameter, der in multidimensionalen MR-Studien analysiert wird (JAMES, 1999).
Die meisten Studien am Menschen und Nutztier erfolgen mit einer Feldstärke von
1,5 – 4,7 Tesla. Um definierte, einheitliche Volumina von Geweben (Muskelgruppen,
Organen, Fett) untersuchen zu können, werden Oberflächenspulen fest anliegend über
der interessierenden Region platziert (Abb. 3.1.1, 3.1.5). Pulswinkel und Pulsform des
Radiofrequenzsignals können so eingestellt werden, dass Resonanzsignale von anderen

Methodenübersicht
8
Geweben wie z.B. von subkutanem Fettgewebe weitestgehend unterdrückt werden
(BECKMANN et al., 1990).
Abbildung 2.1.2: Schema zur Ausrichtung der Moleküle (Atomkerne) in einem festenMagnetfeld B0 am Beispiel einer beliebigen Kohlenstoffverbindung. Die Anordnung in Richtung des Magnetfeldes bedeutet ein geringeres Energieniveau als dieentgegengesetzte Ausrichtung
Die Magnetresonanz-Signale werden erzeugt, indem die Kerne durch die
Ausstrahlung von Radiofrequenzwellen angeregt werden und sich die Netto-
Magnetisierung der Kerne - ausgehend von ihrer Gleichgewichts-Magnetisierung
(Orientierung) - zunächst um 90° ändert. Dafür wird nahe liegend ein 90°-RF-Puls
verwendet. Die resultierende Amplitude der Sinuskurve nimmt mit fortlaufender Zeit
durch das sogenannte „Spin-Dephasing“ ab (Nuklei kehren in die Ausgangsposition
zurück), wobei das Signal, als „free induction decay“ (FID) oder freier Induktionsabfall
aufgezeichnet und in ein Frequenz-Spektrum umgewandelt wird (HORNAK, 1997 -
2000).
In den meisten Fällen wird eine Kombination von verschiedenen RF-Pulsen
verwendet, um Resonanz-Signale zu produzieren. Eine Standardkombination besteht
aus einem 90°-RF-Puls, dem nach einer gewissen Zeitverzögerung ein 180°-RF-Puls
folgt. Das daraus hervorgehende Signal wird Spin-Echo genannt. Die Frequenz der
Sinuskurven im FID oder Spin-Echo entspricht genau der gesuchten Resonanzfrequenz.
Dabei müssen in komplexen Proben mathematische Verfahren herangezogen werden,
um die sich überlagernden individuellen Frequenzen identifizieren zu können. Mit Hilfe
der Fouriertransformation wird die Frequenz-Information, d.h. das gesuchte Spektrum,
aus dem zeitabhängigen (FID-) Signal extrahiert. Diesem Schritt folgt eine
Phasenkorrektur, die automatisch oder manuell durchgeführt werden kann (Abb. 2.1.3).
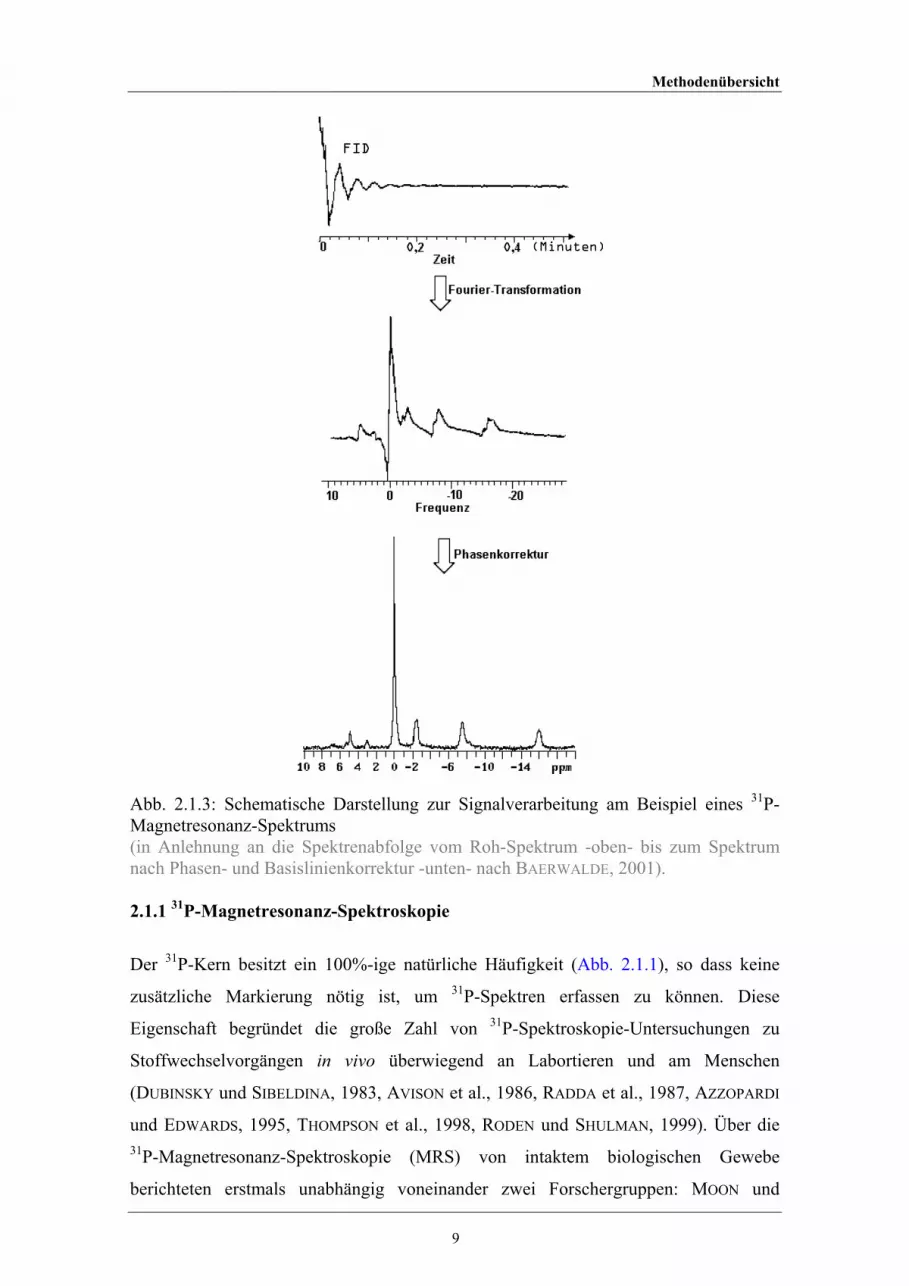
Methodenübersicht
9
FID
O, O, (Minuten)
Abb. 2.1.3: Schematische Darstellung zur Signalverarbeitung am Beispiel eines 31P-Magnetresonanz-Spektrums (in Anlehnung an die Spektrenabfolge vom Roh-Spektrum -oben- bis zum Spektrumnach Phasen- und Basislinienkorrektur -unten- nach BAERWALDE, 2001).
2.1.1 31P-Magnetresonanz-Spektroskopie
Der 31P-Kern besitzt ein 100%-ige natürliche Häufigkeit (Abb. 2.1.1), so dass keine
zusätzliche Markierung nötig ist, um 31P-Spektren erfassen zu können. Diese
Eigenschaft begründet die große Zahl von 31P-Spektroskopie-Untersuchungen zu
Stoffwechselvorgängen in vivo überwiegend an Labortieren und am Menschen
(DUBINSKY und SIBELDINA, 1983, AVISON et al., 1986, RADDA et al., 1987, AZZOPARDI
und EDWARDS, 1995, THOMPSON et al., 1998, RODEN und SHULMAN, 1999). Über die31P-Magnetresonanz-Spektroskopie (MRS) von intaktem biologischen Gewebe
berichteten erstmals unabhängig voneinander zwei Forschergruppen: MOON und

Methodenübersicht
10
RICHARDS (1973), die intakte rote Blutzellen untersuchten, sowie HOULT et al. (1974)
mit der Studie von frisch separierten Beinmuskeln der Ratte (siehe historische
Zusammenfassung bei ANDREW, 1998). Damit begann eine intensive Anwendung von31P-MRS für nahezu jedes Körperorgan, einschließlich Gehirn, Herz, Leber, Niere,
Prostata sowie Gliedmaßen. MRS ist ein nützliches Werkzeug für die Studie von
Erkrankungen/Abweichungen des Stoffwechsels, Tumoren und bestimmten
entzündlichen Vorgängen sowie Blutmangelerscheinungen. Die meisten Arbeiten beim
Menschen konzentrierten sich dabei auf die Untersuchung des Gehirns, um frühzeitig
Abweichungen im Hirnstoffwechsel erkennen zu können, die z.B. durch
Tumorerkrankungen, multiple Sklerose, Epilepsie oder Schlaganfall hervorgerufen
werden können. In der Muskulatur ergeben sich spektroskopische Veränderungen z.B.
durch Phosphofruktokinase- oder Amyloglukosidase-Defizienz, Duchenne- oder
Becker-Muskeldystrophie, Dermatomyositis, Polymyositis, Ganzkörpermyositis oder
Hypothyroidismus.
Seit der ersten 31P-MRS-Muskelstoffwechsel-Studie an Rattenmuskeln durch
HOULT et al. (1974) folgten zahlreiche Studien zum Stoffwechselverhalten der
(menschlichen) Muskulatur nach positiver oder negativer Belastung (Schwerelosigkeit)
und Erholung (CHANCE, 1984, AVISON et al., 1986, BENDAHAN et al., 1990, CERDAN
und SEELIG, 1990, BAERWALDE, 2001). Die Muskulatur (glatt oder quergestreift)
enthält, genauso wie andere interessierende Organe oder Gewebe (Herz, Leber, Gehirn),
die leicht erfassbaren Metaboliten Phosphokreatin (PCr), anorganisches Phosphat (Pi),
γ-, α- und β-ATP in relativ hoher Konzentration. Dabei überlagern sich in vivo die β-
und α-Phosphat-Resonanzen des ADP mit denen von γ- bzw. α-ATP, so dass ADP
nicht direkt ermittelt werden kann (DUBINSKY und SIBELDINA, 1983, BURT et al., 1986,
BERKOWITZ und BALABAN, 1989, DECANNIERE, 1993, JANZEN et al., 1994). Des
Weiteren ist ADP im 31P-Spektrum in vivo nicht hundertprozentig sichtbar. Allein
zytosolisches Pi ist hingegen vollständig MR-sichtbar, aber nicht in den Mitochondrien
bzw. im sarkoplasmatischem Retikulum gebundenes Pi (AVISON et al., 1986, HAUSSER
und KALBITZER, 1989, CERDAN und SEELIG, 1990). Neben den genannten Phosphor-
Komponenten können zwei weitere Signale im 31P-Spektrum beobachtet werden, die als
Phosphomonoester und Phosphodiester interpretiert werden (Abb. 2.1.4).

Methodenübersicht
11
Abb. 2.1.4: 31P Spektrum des M. biceps femoris eines Schweines in vivo(Pi - anorganisches Phosphat, PME - Phosphomonoester, PDE - Phosphodiester, PCr -Phosphokreatin, ATP - Adenosintriphosphat : γ-Phosphat von ATP, α-Phosphate vonATP, β-Phosphat von ATP)
Das im Muskel relativ schwache Signal des Phosphomonesters (PME) ist
überwiegend aus „Zucker-Phosphaten“ (Fruktose-6P, AMP, G6P, IMP)
zusammengesetzt (CERDAN und SEELIG, 1990). Außerdem kann sehr elegant über die
chemische Verschiebung des anorganischen Phosphats relativ zur Position des
Phosphokreatins der intrazelluläre pH-Wert quantifiziert werden (siehe Kapitel 3.1.1).
Da mittels 31P-MR-Spektroskopie energiereiche Phosphorverbindungen und der pH-
Verlauf im Gewebe aufgezeichnet werden können, ist diese Methodik besonders
geeignet, „metabolische Stresssituationen“, hervorgerufen durch verschiedene Ursachen
wie Hypoxie, Anoxie, Ischämie, Hypoglykämie, Verletzungen bzw. Muskelarbeit, zu
analysieren. Häufig werden Anästhetika wie Halothan verwendet, um
Stoffwechselimbalancen zu provozieren bzw. zu verstärken. In jüngster Zeit werden für
die Untersuchung von Genwirkungen künstliche Mutationen (Knockout-, transgene
bzw. transchromosomale Tiere) erzeugt, an denen im Vergleich zur normalen Kontrolle
Stoffwechselveränderungen festgestellt werden können (BOSCH et al., 1998). Einfacher
gestaltet sich die Versuchsanstellung mit bereits natürlich vorkommenden
Polymorphismen an einzelnen Genorten, die extreme phänotypische Ausprägungen
(RyR1, RN -) zeigen und mittels DNA-Analyse exakt festzustellen sind.

Methodenübersicht
12
Für MRS-Untersuchungen kommen verschiedene Techniken in Frage. KORETSKY
und WILLIAMS (1992) unterscheiden zwischen B1- und B0-Technik. Bei der lokalen B1-
Technik werden - wie in den eigenen Untersuchungen (Kapitel 3.1.1) - einfache
Oberflächenspulen über der zu untersuchenden Region platziert (Abb. 3.1.1, 3.1.5) und
auf die Resonanzfrequenz des interessierenden Kerns (z.B. 31P) bzw. der
interessierenden Kerne (z.B. 1H und 13C) abgestimmt. Im Gegensatz wird bei der B0-
Technik zunächst mittels 1H-Magnetresonanz unter Verwendung von ortsauflösenden
magnetischen Gradientenspulen ein interessierendes Volumen (VOI) definiert, von dem
anschließend ein 31P-MR-Spektrum erfasst werden kann. Verschiedene Echosequenzen
dienen dabei der Daten- bzw. Spektrenerzeugung.
Am lebenden Nutztier ist die Zahl von Muskelstoffwechseluntersuchungen mit
Hilfe der MR-Spektroskopie jedoch begrenzt (GEERS et al., 1991, 1992a,b, 1996a,
DECANNIERE, 1993, DECANNIERE et al., 1993, GAREAU et al., 1993, JANZEN et al., 1994,
KALLWEIT et al., 1997, KOHN, 1997, KOZAK-RIBBENS et al., 1997, KOHN et al., 1998,
BAULAIN und HENNING, 2001). Einerseits können notwendige Investitionen in MR-
Geräte, die auch für die Spektroskopie geeignet sind, aufgrund nicht gewährter
(vorhandener) Mittel für eine von der Gentechnologie abweichende
Grundlagenforschung besonders in den Nutztierwissenschaften nicht oder kaum
realisiert werden. Andererseits ist das Potential der Technik nicht umfassend bekannt
bzw. wird ignoriert. Gleichzeitig sind besonders in Deutschland Kooperationen mit
wissenschaftlichen humanmedizinischen Einrichtungen, die über die entsprechende
Geräteausstattung verfügen, sehr schwierig zu realisieren. Gerade diese Technik könnte
jedoch im Verbund mit konventionellen Stoffwechseluntersuchungen - anhand von
Blut-, Urin- und Speichelproben - sowie neu entwickelten Verfahren zur Aufdeckung
von Gen- und Umweltwirkungen bzw. endogenen und exogenen Effekten am lebenden
Organismus beitragen. Es ist wenig sinnvoll, nach Markergenorten oder „Quantitative
Trait Loci“ (QTL) zu suchen, wenn die meisten Methoden der Merkmalserfassung
(Leistungsprüfung) sich weiterhin an „Schlachthofstandards“ orientieren. Die gesuchten
Effekte sind häufig viel zu klein (quantitative Effekte!), um mit „ungenauen“ Methoden
nur unter höchstem Tiereinsatz aufgedeckt zu werden.
Seitdem horizontale Magneten mit Feldstärken von ≥ 1,5 T und
Innendurchmessern von ≥ 30 cm verfügbar sind, wurden erste Muskelstoffwechsel-
untersuchungen an Menschen und Labortieren durchgeführt, wobei vor allem versucht

Methodenübersicht
13
wurde, auf genetischen Defekten beruhende Muskelerkrankungen mit „normalen“
Muskeln zu vergleichen (AVISON et al., 1986).
Nachdem FUJII et al. (1991) die Beziehung zwischen maligner Hyperthermie und
einer Mutation am Ryanodin-Rezeptor 1 des sarkoplasmatischen Retikulums beim
Schwein nachweisen konnten, besteht die Möglichkeit, die Wirkungsweise des
Defektallels vor allem in den heterozygoten Tieren (Nn oder NP) gezielt zu
untersuchen. Erste In-vivo-31P-MRS-Untersuchungen an Muskeln von MHS-positiven
Schweinen bzw. Schweinen mit unterschiedlicher Stressanfälligkeit wurden durch eine
amerikanische Arbeitsgruppe (ROBERTS et al, 1983), später durch eine belgische
Arbeitsgruppe (GEERS et al., 1991, 1992a, b, 1996a, DECANNIERE, 1993 und
DECANNIERE et al., 1993) publiziert. Diesen Untersuchungen folgten neben den eigenen
Untersuchungen am Growth Biology Laboratory (GBL) des zum United States
Department of Agriculture (USDA) gehörenden Agricultural Research Service (ARS) in
Beltsville, MD, und der Howard University, Washington, DC (SCHOLZ et al., 1995,
1998) weitere Studien durch eine kanadische Arbeitsgruppe (GAREAU et al., 1993,
JANZEN et al., 1994) sowie eine deutsche Arbeitsgruppe der Bundesforschungsanstalt
für Landwirtschaft - FAL (KOHN, 1997, KOHN et al., 1998, BAULAIN und HENNING,
2001).
KOHN (1997) untersuchte mit Hilfe der 31P-MR-Spektroskopie in vivo den
Muskelstoffwechsel von MHS-homozygot positiven, heterozygoten und homozygot-
negativen Schweinen (8 - 20 Wochen alt) im Abstand von je 2 Wochen im Ruhezustand
und unter Belastung durch Halothan sowie zusätzlich post mortem die Abbauvorgänge
in Schlachthälften. Ergänzende Blut- und Serumanalysen erweiterten die Aussagekraft
der Untersuchungen. Die homozygoten Defektallelträger zeigten im Gegensatz zu den
homozygot normalen und den heterozygoten Tieren unter Halothanbelastung
dramatische Veränderungen im Muskelstoffwechsel. In Ruhe waren die Unterschiede
weitaus geringer. Die heterozygoten Tiere nahmen eine Mittelstellung zwischen den
beiden homozygoten Genotypen ein. Ihr Muskelstoffwechsel glich mehr den NN- als
den nn-Schweinen. Bei den heterozygoten Tieren wurden daher keine großen Einbußen
in Bezug auf die Fleischqualität erwartet. Ähnliche Ergebnisse erzielten bereits GEERS
et al. (1991, 1992a,b, 1996a), DECANNIERE, 1993, DECANNIERE et al. (1993), GAREAU
et al. (1993) sowie JANZEN et al. (1994). Sogar ohne eine gewollte Auslösung von
Muskelstress können Defektallelträger eine massive maligne Hyperthermie entwickeln.
KOZAK-RIBBENS et al. (1997) beobachteten bei einem von sechs „halothan-positiven“

Methodenübersicht
14
Tieren während einer 31P-MR-Studie nach Gabe von „Pentobarbitone“ eine „spontane“
Auslösung des MH-Syndroms ohne die übliche Muskelsteife beobachten zu können.
Die Auslösung des MH-Syndroms bei diesem Versuch war vorhersehbar, da zur
elektrischen Stimulation des M. semimembranosus die Haut über diesem Muskel
operativ entfernt und Elektroden im Muskel befestigt wurden. Da die Anästhesie bei
jedem Tier eine individuelle Wirkung hat, ist es denkbar, dass das betroffene Tier nicht
ausreichend sediert war. Der durch den Versuch erzeugte Stress verursachte daher die
beobachtete fatale Reaktion. Während die orale Temperatur auf 44,5 °C anstieg, sank
der intrazelluläre pH-Wert auf 6,21 und das Phosphokreatin-Niveau auf 6 % des
Startniveaus. Auf eine sehr schnelle Glykogenolyse deutend stiegen parallel die
Konzentrationen von Phosphomonoester (um das neunfache) und anorganischem
Phosphat extrem an. Das ATP-Niveau blieb jedoch unverändert. Die Ursache für das
konstante ATP-Niveau ist in der nicht vorhandenen Muskelkontraktion zu vermuten.
DECANNIERE (1993) bzw. DECANNIERE et al. (1993) beobachteten hingegen bei vier von
17 MHS-positiven Tieren parallel zur Muskelsteife einen Abfall des ATP-Niveaus.
Bei Anwesenheit des Defektallels am Ryanodin-Rezeptor-1-Genort ist die Ca2+-
Balance des sarkoplasmatischen Retikulums (SR) von Muskelzellen eingeschränkt
(MACLENNAN und PHILLIPS, 1992). Post mortem bewirkt diese Mutation nach
Stressauslösung mittels Halothan oder nach zusätzlicher Succinylcholin-Gabe einen
schnelleren Phosphokreatinabbau bei gleichzeitigem Anstieg des anorganischen
Phosphats, einen gesteigerten ATP-Umsatz sowie eine gesteigerte Glykolyserate
(KOLB, 1987, KOZAK-REISS et al., 1987, UHRIN und LIPTAJ, 1990, MOESGAARD et al.,
1993, 1995). Die erhöhte Glykolyserate ist aus dem pH-Verlauf sowie aus der
Anreicherung von Phosphomonoestern abgeleitet worden (KOZAK-RIBBENS et al.,
1997).
Neben den In-vivo-Studien zur MHS-Problematik existiert eine Anzahl von MRS-
Experimenten an Muskelbioptaten bzw. einzelnen Muskeln, die post mortem oder vom
lebenden Tier entnommen wurden (KOZAK-REISS et al., 1987, UHRIN und LIPTAJ, 1990,
AZUMA et al., 1994, BERENYI et al.,1994, BOGNER et al., 1996, KRSKA et al., 2000,
LAHUCKY et al., 2000a,b). Dabei führten BERENYI et al. (1994) sowie BOGNER et al.
(1996) keine 31P-Spektroskopie sondern eine 1H-MR-Spektroskopie durch, um T1- und
T2-Relaxationszeiten bei unterschiedlichen MHS-Genotypen festzustellen. Die
Relaxationsparameter mit einer tendenziell verlängerten T2-Zeit in MHS-positiven
Tieren unterschieden sich jedoch nur in einer von fünf Linien signifikant, so dass diese

Methodenübersicht
15
Methodik allein nicht zur Differenzierung der unterschiedlichen RyR1-Genotypen
herangezogen werden kann.
Andere Studiengruppen nutzten die 31P-MR-Spektroskopie z.B. zur
Muskelstoffwechseluntersuchung am lebenden (nicht anästhesierten) Fisch (VAN DEN
THILLART, 1989, VAN WAARDE et al., 1990) bzw. zur Untersuchung des Phosphat-
stoffwechsels in Geflügelembryonen, um anhand des ATP-zu-PDE-Verhältnisses am
18. Bruttag eine frühzeitige Selektion von fettärmeren Broilern zu ermöglichen
(TOWNER et al., 1991, LIU et al., 1992, 1994, LIRETTE et al., 1993a,b).
2.1.2 13C-Magnetresonanz-Spektroskopie
Bevor die 13C-Magnetresonanz-Spektroskopie zur Untersuchung von Kohlen-
stoffkomponenten (Glukose, Glykogen, Kreatin) im Stoffwechsel bei Tier und Mensch
in vivo eingesetzt werden konnte, dienten diesem Zweck „Tracer“-Techniken und
Gewebebiopsien (SCHÖBERLEIN, 1976, CHEAH et al., 1995, KLONT et al., 1994, KLONT
und LAMBOOY, 1995, LAHUCKY et al., 1995, RODEN und SHULMAN, 1999).
Die 13C-MR-Spektroskopie besitzt eine vierfach geringere Empfindlichkeit als die31P-MR-Spektroskopie (AVISON et al., 1986, Abb. 2.1.1). Gleichzeitig ist nur das mit
1,1 % natürlich vorkommende 13C-Isotop mittels MR-Spektroskopie detektierbar,
während das überwiegend natürlich vorkommende 12C-Isotop nicht genutzt werden
kann, da es aufgrund der geraden Protonen- und Neutronenzahl keinen Eigenspin (= 0)
aufweist. Ähnlich wie bei der 31P-MR-Spektroskopie können Oberflächenspulen zur
Erzeugung der 13C-Spektren verwendet werden (z.B. Abb. 3.1.3: 13C-Spektrum für
reines Glykogen). Speziell gestaltete Doppel-Oberflächenspulen – wie in den eigenen
Untersuchungen (Abb. 3.1.2, 3.1.5) - helfen das Protonen- bzw. Wasserstoffsignal vom
Kohlenstoffsignal zu trennen. Zusätzlich wird häufig zur Verstärkung des
Kohlenstoffsignals bei Untersuchungen des Glukose- bzw. Glykogenstoffwechsels
markierte 1-13C-Glukose intravenös verabreicht. Das verstärkte Signal kann dann
sowohl an der C1-Position als auch an der C6-Position von Glykogen bzw. Glukose
nachgewiesen werden (KORETSKY und WILLIAMS, 1992). Ein eigener Versuch am
United States Department of Agriculture in Beltsville, MD, USA, (unveröffentlichte
Ergebnisse, Scholz, 1996) an einem Schwein mit intravenös verabreichter, markierter
D-Glukose-13C6 (Sigma-Aldrich Chemical Company, Catalog Number 38,937-4, Lot
Number 085H3755) zeigte einen erheblichen Anstieg im βC1-Glukoseniveau sowie

Methodenübersicht
16
einen zweiten Peak am C1-Glykogen unmittelbar nach der Injektion einer 28 ml
Natriumchlorid-Lösung, in der 15 g Dextrose und 0,1 g D-Glukose-13C6 enthalten
waren (Abb. 2.1.5).
C1-Glykogen
Kreatin
ßC1-Glukose
Abb. 2.1.5: Verlauf der Konzentration von C1-Glykogen (100,5 ppm), Kreatin (157ppm) und βC1-Glukose (96,6 ppm) nach Verabreichung einer 28 ml Natriumchlorid-Lösung mit 15 g Dextrose und 0,1 g D-Glukose-13C6 bei einem 10,9 kg schwerenSchwein (RyR1 = Nn) mit 5/8 Duroc-Genanteilen (unveröffentlichte eigene Ergebnisse)
Wie bereits aus Abb. 2.1.5 hervorgeht, lassen „neuere“ technische Entwicklungen
in der 13C-MR-Spektroskopie eine nicht invasive Messung des Muskel-Glykogen-
gehaltes ohne zusätzliche Anreicherung mit markierter D-Glukose-13C6 zu
(DECANNIERE, 1993, PRICE et al., 1996, VAN DEN BERGH et al., 1996, SHULMAN, 1997,
PRICE und GORE, 1998, ROUSSEL et al., 1998). Deshalb erfolgten auch die eigenen

Methodenübersicht
17
Untersuchungen zum Glykogenabbau nach Auslösung von Muskelstress bei Schweinen
unterschiedlicher RyR1-Genotypen ohne Zusatz von markierter Glukose.
Die Vorteile der MR-Spektroskopie in vivo umfassen:
1. die Vermeidung von invasiven Eingriffen wie bei der Biopsie oder
Blutentnahme,
2. die Möglichkeit, den zeitlichen Verlauf und Wege von
Stoffwechselreaktionen kontinuierlich und „ohne“ Unterbrechung (on-line)
zu verfolgen,
3. ein größeres Probenvolumen als bei Bioptaten,
4. die Analyse ohne radioaktiv markierte Stoffe,
5. wenn nötig, die bildliche Darstellung der zu untersuchenden Region zur
Überprüfung der Proben-Positionierung bzw. von möglichen anatomischen
und pathologischen Variationen
Die Einschränkungen bestehen bis heute, zusätzlich zu den bereits beschriebenen
technischen Begrenzungen, die in der Natur der Methodik bzw. in den biochemischen
Gegebenheiten des zu untersuchenden Gewebes begründet sind, in:
1. der Notwendigkeit, das Untersuchungssubjekt für einen längeren Zeitraum
fixieren (sedieren) zu müssen
(Die Zeit, für ein einzelnes aussagekräftiges 31P-MR-Spektrum beträgt
0,5 - 5 Minuten, während für ein aussagekräftiges 13C-MR-Spektrum
5 - 20 Minuten veranschlagt werden müssen. Die gesamte
Untersuchungsdauer (Fixierungsdauer) richtet sich nach der Zielstellung
der Analyse bzw. nach der Geschwindigkeit der Stoffwechselabläufe.)
und
2. dem notwendigen Ausschluss von Probanden mit Metallimplantaten.

Methodenübersicht
18
2.2 Methoden zur Ermittlung der Körperzusammensetzung - in vivo2.2.1 Einflussfaktoren auf die Körperzusammensetzung und deren Messung im
Rahmen der Leistungsprüfung
Während der Wachstums- und Reifephase (Prüfperiode) oder während der Trächtigkeit
sowie der nachfolgenden Laktation variiert die Körperzusammensetzung der Individuen
in erheblichem Maße. Die sich gegenseitig beeinflussenden Faktoren für die Variation
der Körperzusammensetzung umfassen - neben der speziell den Tierzüchter
interessierenden, in Kapitel 4.2 ausführlich diskutierten genetischen Veranlagung - den
Gesundheits- und Ernährungszustand (Futteraufnahme), die Rationsgestaltung, das
Geschlecht, den Einsatz von Stoffwechsel verändernden Komponenten wie Anabolika
bzw. Katabolika sowie weitere nicht spezifizierte Umwelt- bzw. Haltungsbedingungen
(SPEER, 1988, BASTIANELLI und SAUVANT, 1997, FLACHOWSKY, 1998, LOBLEY, 1998,
NÜRNBERG et al., 1998, MITCHELL et al., 2000) (Abb. 2.2.1).
Genetik
Ernährung/Fütterung
Stoffwechsel/Hormone
Bewegungsmöglichkeit (Trainingszustand) und Aktivität[Umwelt; Haltungsbedingungen]
Körperzusammensetzung
Alter/ Körpermasse/ Geschlecht
Abb. 2.2.1: Einflussfaktoren auf die Körperzusammensetzung
Sowohl während des Wachstums als auch während der unterschiedlichen
Reproduktionszyklen verändern sich die absoluten und relativen Bestandteile von
Lipiden, Proteinen, Wasser und Mineralstoffen (Mengen- und Spurenelemente) des
Muskel- und Körperfettgewebes sowie der Knochen (ELOWSSON et al., 1998).
Andererseits zeigen SUSENBETH und KEITEL (1988), dass Schweine ab einem
Körpergewicht von ca. 60 kg einen relativ konstanten Proteingehalt von 17,6 %
innerhalb der fettfreien Fraktion (leerer Körper minus Schlachtkörper) und von 21,8 %
innerhalb der fettfreien Magermasse aufweisen.
Körperzusammensetzung, (metabolische) Körpermasse und die Verteilung der
Fettdepots (bzw. teilweise der Proteindepots) beeinflussen den Nährstoffaufwand für die

Methodenübersicht
19
grundlegenden Stoffwechselprozesse (Erhaltungsaufwand) und die Nährstoffbedürfnisse
für die unterschiedlichen Leistungsanforderungen während des Wachstums und der
Reproduktion einschließlich Laktation. Diese Aufwendungen bedingen die Effizienz der
Futteraufnahme, das heißt die Umsetzung von aufgenommener Energie in
Lebendmassezunahme (LUITING, 1993, LUITING et al., 1995). Vor allem sind die relativ
leicht messbaren Veränderungen im Fett- und Wassergehalt stark mit den
Veränderungen des Gesamtkörpergewichtes korreliert. Allein die Messung der aktuellen
Körperzusammensetzung in vivo liefert eine detaillierte Information über den
Ernährungs- und/ oder Gesundheitsstatus der Tiere, da nur auf diesem Wege genaue
Aussagen zu den unterschiedlichen „Energiespeichern“ des Körpers wie Fett, Protein,
Kohlenhydrate (Glykogen) oder zu den Komponenten der Fett- bzw. fettfreien Masse
getroffen werden können. Die Körpermasse selbst kann nur als grober Indikator für die
Körperzusammensetzung in einer Population oder Herde verwendet werden. Es ist
unmöglich, lediglich in Bezug auf die Körpermasse zu schlussfolgern, ob ein Schwein
verfettet oder gut bemuskelt ist, gesund oder krank erscheint bzw. unter- oder
überernährt wurde. Exakte In-vivo-Messmethoden müssen speziell für wertvolle
Zuchttiere angewandt oder entwickelt werden, um Veränderungen in der
Körperzusammensetzung genauestens feststellen und verfolgen zu können. Speziell im
Zusammenhang mit neu entstehenden Biotechnologien zur Modifikation der
Nahrungsbestandteile bzw. der Tiere selbst ergibt sich die Notwendigkeit zur
Verfeinerung der Messmethoden (WRAY-CAHEN et al., 1998). Die Aufdeckung von
Beziehungen zwischen genetischer Information (Genotyp) und messbarer, durch
Umwelteinflüsse modifizierter Merkmalsausprägung (Phänotyp), die im Kapitel 4.2.
insbesondere für die Körperzusammensetzung und das Wachstum beschrieben werden,
kann dadurch wesentlich erleichtert werden.
Eine Anzahl von Kriterien für eine ideale Methode zur Messung der
Körperzusammensetzung bilden Entscheidungshilfen vor Inbetriebnahme und
Anschaffung eines entsprechenden Gerätes (TOPEL und KAUFFMAN, 1988, KALLWEIT,
1994, KALLWEIT et al., 1994, ALLEN und WALSTRA, 1996, BRANSCHEID und
DOBROWOLSKI, 2000, MITCHELL und SCHOLZ, 2001):
1. hohe Messgeschwindigkeit: – abhängig vom Verwendungszweck (wenige Sekunden
für eine On-line-Schlachtkörperanalyse; <1 Minute für die Einstufung von
Verkaufstieren; bis zu < 20 Minuten für die Leistungsprüfung von Zuchtvieh),

Methodenübersicht
20
2. Zerstörungsfreiheit: – vorzugsweise nicht invasiv,
3. Mindestgenauigkeit für Leistungsprüfung und Preisbildung (Handelsklassen-
einstufung); z.B.: R2 ≥ 0,8, SEE ≤ 2,5 % für den Muskelfleischanteil (für
Wissenschaft: Wiederholbarkeit > 95 %),
4. Objektivität,
5. leichte Bedienbarkeit/ Benutzerfreundlichkeit (Praktikabilität): minimale Belastung
für Tier und Mensch (möglichst ohne Sedierung oder Anästhesie bei gleichzeitig
benutzerfreundlicher Handhabung des Gerätes und der Datenauswertung),
6. Wirtschaftlichkeit: im Hinblick auf Anschaffungskosten und laufende Kosten – die
Kosten je Messung können in Abhängigkeit vom Verwendungszweck stark
variieren; mit den niedrigsten Kosten für Vermarktungstiere und den höchsten
Kosten für Zuchttiere bzw. Forschungstiere,
7. Mess- und Analyseaufwand: möglichst Realzeitmessung mit minimaler
Datenaufbereitung und -analyse und „unmittelbarer“ Ergebnisausgabe,
8. Robustheit des Gerätes gegen mechanische und Umweltbelastungen und
9. Öffentliche Akzeptanz (Röntgenstrahlung und speziell radioaktive Strahlung sind
oft unerwünscht).
Um ein ausreichendes Genauigkeitsniveau zu erzielen, ist es in den meisten Fällen
nötig, dass die gewählte Methodik eine direkte Messung von mindestens einem oder
besser mehreren Hauptkompartimenten der Körperzusammensetzung gewährleistet.
Diese Messung kann entweder auf der Basis von Gewebezusammensetzung (Muskel,
Fett, Knochen, Bindegewebe), roh-chemischer Zusammensetzung (Protein, Lipid,
Wasser, Asche) oder elementarer Zusammensetzung (Kohlenstoff, Wasserstoff,
Stickstoff, bzw. Phosphor, Kalzium, Eisen, Kupfer, Zink – MAHAN und SHIELDS, 1998,
KIENZLE et al., 1998) beruhen. Außerdem sollte die Messung vorzugsweise den
gesamten Körper und nicht nur ausgewählte Körperteile erfassen, da die Messwerte
einzelner Bestandteile nur zum Teil sehr hoch mit der Gesamtkörperzusammensetzung
korrelieren (THOMAS et al., 1998).
In den zurückliegenden Jahren ist eine Vielzahl an Instrumenten zur Ermittlung
der Körperzusammensetzung am lebenden Tier oder Schlachtkörper eingesetzt worden.
Diese umfassen unter anderem Geräte, die auf den Prinzipien von Ultraschall,
Röntgenstrahlung, Gammastrahlung, Nahinfrarotstrahlung, magnetischer Resonanz,
elektrischer Impedanz, elektromagnetischer Konduktivität und Neutronenaktivierung

Methodenübersicht
21
basieren (Abb. 2.2.4). Viele dieser Geräte können genaue und nützliche Informationen
im Hinblick auf die Körperzusammensetzung liefern. Sie sind jedoch häufig kompliziert
zu bedienen und entsprechen nicht den kombinierten Kriterien von Genauigkeit und
Wirtschaftlichkeit.
Allgemein lassen sich die Veränderungen in der Körperzusammensetzung am
lebenden Tier oder am Schlachtkörper an einem modifizierten Fett-zu-Muskel-
Verhältnis erkennen. Um die Verbraucherwünsche nach (vermeintlich) gesünderen
Produkten zu erfüllen, standen in den zurückliegenden Jahren immer magerere
Schweine im Mittelpunkt der züchterischen Selektion. Extrem mageres Fleisch - kein
oder sehr wenig Fett enthaltend - wird von vielen Verbrauchern gewünscht. Das führte
einerseits zu extremen Rassen wie z.B. Pietrain oder Belgische Landrasse, die diese
Anforderungen erfüllten. Aufgrund der durch eine Mutation am Ryanodin-Rezeptor-1–
Genort (oder Calcium-Release-Channel-Genort) hervorgerufenen Muskelhypertrophie
verminderte sich die Fitness und gesundheitliche Stabilität dieser Tiere. Parallel
verschlechterte sich die Fleischqualität. Andererseits werden zunehmend in
überwiegend „ökologisch“ geführten Freilandbetrieben Schweinerassen wie
Schwäbisch-Hällisches Schwein (oder Angler Sattelschwein bzw. Deutsches
Sattelschwein) verwendet, die härteren Umweltbedingungen gewachsen sind,
gleichzeitig aber einen stärkeren Fettwuchs aufweisen.
Genaue Methoden zur Messung der Körperzusammensetzung sind erforderlich,
um Selektions-, Fütterungs- und/oder Vermarktungsprogramme optimieren zu können,
die die Gesundheit von Mensch und Tier berücksichtigen.
2.2.2 Physikalische und chemische Grundlagen für die Messung derKörperzusammensetzung – Modelle der Körperzusammensetzung
Lebende Systeme bestehen aus komplexen heterogenen Komponenten, die eine exakte
Messung der Körperzusammensetzung erschweren. Auf atomarem Niveau besteht der
Körper zu > 96 % aus den vier Elementen Sauerstoff, Kohlenstoff, Wasserstoff und
Stickstoff. Fügt man weitere sieben Elemente wie Natrium, Kalium, Phosphor, Chlor,
Kalzium, Magnesium und Schwefel hinzu, sind bereits mehr als 99,5 % der Gesamt-
körperzusammensetzung abgedeckt (WANG et al., 1992, KEHAYIAS und VALTUEÑA,
1999, MITCHELL und SCHOLZ, 2001). Auf molekularem Niveau bilden diese Elemente
eine Vielzahl von differenzierten Komponenten. Zur Messung der
Körperzusammensetzung erfolgt jedoch eine vereinfachte, generelle Aufteilung in Lipid,

Methodenübersicht
22
Wasser, Protein, Glykogen und Asche, die zusammen mehr als 99 % der Körper-
zusammensetzung ausmachen. Betrachtet man das Gewebe-Organ-Niveau, weist der
Körper in Abhängigkeit vom Ernährungszustand unterschiedliche Füllungsgrade auf.
Der Körper enthält neben Skelettmuskulatur (Fleisch), subkutanem, internem
(abdominalem), inter- und intramuskulärem Fett variable Mengen an Knochengewebe,
Blut, Bindegewebe, Haut, Haaren, Klauen, Organen des Herz-, Kreislauf- und
respiratorischen Systems, Nervengewebe, Reproduktions- und
Harnausscheidungsorganen sowie Organen des Gastrointestinaltraktes.
Die Techniken zur Messung der Körperzusammensetzung basieren häufig auf der
Annahme einer Vielzahl physikalischer und chemischer Konstanten. Zusätzlich bedarf
es gesicherter konstanter physiologischer Verhältnisse, um die Körperzusammensetzung
lebender Individuen oder die Schlachtkörperzusammensetzung insbesondere mit
indirekten Methoden exakt messen zu können. In jungen, wachsenden Schweinen ist das
Verhältnis zwischen den Bestandteilen aus der grobgeweblichen Zerlegung (Fettgewebe,
Mager (Muskel) - und Knochengewebe) und der chemischen Analyse (Lipid, Wasser,
Protein und Asche) jedoch nicht konstant. Während der ersten Wochen nach der Geburt
steigt mit zunehmender Körpermasse bei gleichzeitiger Abnahme des Wassergehaltes
(FUSCH et al., 1999) der Proteingehalt des fettfreien Körpers beim Schwein mit hohen
individuellen Schwankungen (ELOWSSON et al., 1998) sehr stark an (Abb. 2.2.2).
0
0.05
0.1
0.15
0.2
0.25
0.3
0 10 20 30 40 50 60 70 80 90 100 110
Körpergewicht (kg)
P:W
-Ver
häl
tnis
Abb. 2.2.2: Entwicklung des Protein-zu-Wasser (P:W)-Verhältnisses in der fettfreienMasse bzw. Magergewebemasse beim Schwein(Durchschnittliche Werte wurden für 1,3 kg [Geburt], 12,5 kg [ca. 54 Lebenstage], 40 kg [ca. 110Lebenstage] und 100 kg [ca. 195 Lebenstage] aus den Angaben in Kapiteln 2.2.2 und 3.3.3.1 in PFEIFFER
et al., 1984; für ein durchschnittliches Alter von 2,7 Wochen (3,59 ± 1,87 kg) aus den Angaben vonFUSCH et al., 1999 bzw. für ein Alter von ca. 12 Wochen (17,74 kg) aus den Angaben von ELOWSSON etal., 1998; für ein durchschnittliches Gewicht von 45 kg aus den Angaben von BIKKER et al., 1995 sowiefür ein durchschnittliches Gewicht von 85 kg aus den Angaben von BIKKER et al., 1996a berechnet.)

Methodenübersicht
23
Dieser Trend setzt sich bis zur „chemischen Reife“, die eng mit der sexuellen Reife der
Tiere gekoppelt sein soll (MALIK, 1984), im Alter zwischen 21/2 und 5 Monaten fort.
Danach geht die Veränderung der Körperzusammensetzung – bezogen auf die fettfreie
Masse – nur noch sehr langsam voran (MOULTON, 1929). In der Tendenz erhöht sich das
Protein-zu-Wasser-Verhältnis in der fettfreien Masse (Magergewebemasse) des
Schweines von ca. 0,135 bei Geburt über 0,186 im Alter von ca. 3 Wochen auf über
0,25 ab einem Alter von ca. 12 Wochen bzw. ab einem Gewicht von ca. 45 kg (Abb.
2.2.2). Allein für eine Auswahl von 7 verschiedenen Muskeln (M. triceps brachii, M.
longissimus dorsi, M. psoas major, M. glutaeus medius, M. semimembranosus, M.
biceps femoris und M. semitendinosus) bei einem durchschnittlichen Hälftengewicht
von 40,65 kg (ca. 100 - 110 kg Lebendmasse) betrug das durchschnittliche Protein-zu-
Wasser-Verhältnis = 0,281 und liegt damit in dem in der Abb. 2.2.2 gezeigten Bereich
(berechnet aus den Daten von PRUSA et al., 1989). Bei einem durchschnittlichen
Lebensalter von 147 Tagen und einem Gewicht von 91 kg lag das Protein-zu-Wasser-
Verhältnis des Fleisches aus Schinken, Kotelett und Schulter (ham, loin, picnic und butt
cuts) bei ca. 0,25 (berechnet aus den Daten von FORTIN und ELLIOT, 1985). Aus den
Literaturdaten konnte bis ca. 110 kg Lebendmasse kein maximales Protein-zu-Wasser-
Verhältnis abgeleitet werden. Eine Schwierigkeit liegt in der nicht vollständig erreichten
„absoluten Fettfreiheit“ der untersuchten „fettfreien“ bzw. Magergewebemassen. Die
Ergebnisse von LAUBE (2000) weisen ein durchschnittliches Protein-zu-Wasser-
Verhältnis von 0,31 im Kotelett (zwischen 14. und 15. Rippe) von Masttieren mit
unterschiedlicher genetischer und betrieblicher Herkunft auf, wobei der intramuskuläre
Fettgehalt (%) zwischen 1,23 (PI – nn) und 2,03 (Du – NN) schwankte.
Gleichzeitig mit dem Protein-zu-Wasser-Verhältnis in der fettfreien Masse ändern
sich Wasser- und Lipidgehalt des Fettgewebes. So enthält das Rückenfettgewebe des
neugeborenen Ferkels ~85 % Wasser und 6 % Lipid (MCMEEKAN, 1940). Bei
durchschnittlich 62,5 kg schweren Schweinen beträgt der Wassergehalt des
Rückenfettgewebes ca. 15,6 % (WARRIS et al., 1990) und sinkt im Alter von 17 Wochen
auf durchschnittlich 11,7 % bei ca. 81 % Lipid (Rohfett) bzw. im Alter von 32 Wochen
auf bis zu 5 % bei über 93 % Lipid (Rohfett) (NÜRNBERG u.a., 1988). Übereinstimmend
mit diesen Ergebnissen ermittelte MOULTAN (1929) durchschnittlich 7,5 % Wasser in
„frischem“ Schweinefett. Außerdem sind die zwischen den unterschiedlichen
Fettgewebedepots variierenden Wasser- und Lipidgehalte vom Verfettungsgrad bzw.

Methodenübersicht
24
Energieverzehr und der Rasse des Schweines abhängig (NÜRNBERG u.a., 1988,
SCHWÖRER u.a., 1990, WARRISS et al., 1990, ENSER und WOOD, 1991, ENDER u.a.,
1997a). Je magerer die Tiere bei gleichem Alter (gleichem Gewicht) sind, desto
niedriger fällt der Lipidgehalt z.B. im Rückenfettgewebe aus (66,9 % Lipidgehalt bei
60 % Muskelfleischanteil zu 71,5 % Lipidgehalt bei 53 % Muskelfleischanteil – ENDER
u.a., 1997a) bzw. desto wässriger ist das Rückenfettgewebe (WARRISS et al., 1990, siehe
Abb. 2.2.3).
0
5
10
15
20
25
Larg
e B
lack
Ber
kshi
re
Tam
wor
th
Sad
dleb
ack
Ham
pshi
re
Glo
uces
ter
Old
Spo
ts Dur
oc
Oxf
ord
San
dyan
d B
lack
Larg
e W
hite
Land
race
Pie
trai
n
Rückenspeckdicke [mm] Wassergehalt im Rückenspeck [%]
Abb. 2.2.3: Abhängigkeit des Wassergehalts im Rückenfettgewebe von Rücken-speckdicke und Rasse bei ∅ 62,5 kg Körpergewicht(Daten stammen aus WARRISS et al., 1990)
Der Lipidgehalt im Rückenfettgewebe steigt aber – wie oben beschrieben - mit
zunehmendem Alter bei gleichzeitiger Vergrößerung des Durchmessers der Adipozyten
an (NÜRNBERG et al., 1998).
In der nachfolgenden Übersicht (Abb. 2.2.4) werden physikalische und chemische
Eigenschaften aufgelistet, die zur Messung der Körperzusammensetzung verwendet
werden. Die Genauigkeit der jeweils verwendeten Methodik hängt von der
Messpräzision für die einzelnen Komponenten ab.

Methodenübersicht
25
Abb. 2.2.4: Nutzbare physikalische und chemische Eigenschaften zur Messung derKörperzusammensetzung
Für eine effiziente Verwendung der Techniken zur Analyse der
Körperzusammensetzung (in vivo) ist es sowohl erforderlich, die physikalischen und
chemischen Prinzipien der Messtechniken, als auch die grundlegenden biologischen
Modelle der Körperzusammensetzung zu kennen (PIETROBELLI et al., 1996). Während
die Ermittlung der Körperzusammensetzung oder des Fettgehaltes in vivo beim
Menschen auf verschiedene Modellannahmen angewiesen ist (PIETROBELLI et al., 1996,
BUTTE et al., 1997, GORAN et al., 1998, WANG et al., 1998a, b, 2000), sind beim
Schwein (Nutztier) Referenzwerte für den Gehalt bzw. die Masse von Fett, Protein,
Glykogen, Weichgewebemineralstoffen, Wasser und Knochenmineralstoffen (6-
Komponenten-Modell) relativ leicht mit Hilfe einer chemischen Analyse des gesamten
Schlachtkörpers zu bestimmen (WAGNER et al., 1999). Eine Voraussetzung für eine
hohe Genauigkeit bildet die getrennte Untersuchung von Weichgewebe und Knochen
nach Zerlegung und Homogenisierung (ELOWSSON et al., 1998, WAGNER et al., 1999).
Beim Menschen wird dieses „Problem“ durch Referenzstudien an Leichnamen oder
entsprechenden Körperteilen (LLOYD und MAYS, 1987, ABATE et al., 1994, JEBB, 1997,
MITSIOPOULUS et al., 1998) bzw. unter Zuhilfenahme von Tiermodellen oder
Referenzphantomen gelöst. Das hauptsächlich genutzte Tiermodell für Referenzstudien
in Bezug auf die Körperzusammensetzung des Menschen ist das Schwein (SHENG et al.,
1988, FOWLER et al., 1992, BRUNTON et al., 1993, 1997, SVENDSON et al., 1993, ELLIS et
al., 1994, FULLER et al., 1994, JEBB et al., 1995, MITCHELL et al., 1996a, PICAUD et al.,
1996, PINTAURO et al., 1996, ELOWSSON et al., 1998). Einige generelle Aspekte aus der
Zellen
Energiespektren der radioaktiven Strahlung (Gamma, Alpha) nach Neutronenaktivierung
bzw. magnetische Resonanz
Kohlenstoffgehalt
Lipidgehalt
ProtonengehaltSauerstoffgehalt
Wassergehalt
spezifische Dichte
Fett
Stickstoffgehalt
Proteingehalt
ProtonengehaltSauerstoffgehalt
Wassergehalt
spezifische Dichte
Magergewebe
KalziumgehaltPhosphorgehalt
spezifische Dichte
Knochen
Kohlenstoffgehalt
Glykogengehalt
RöntgenschwächungNahinfrarot- bzw. Ultraschallreflektion und -brechung
Elektrischer Widerstand bzw. Leitfähigkeit
Kaliumgehalt
Gewebeniveau
MolekularesNiveau
AtomaresNiveau

Methodenübersicht
26
Erfassung der Körperzusammensetzung beim Menschen werden deswegen zusätzlich
zur „reinen“ Nutztier-Körperzusammensetzung berücksichtigt.
WANG et al. (1998a) unterteilen die Methoden zur Schätzung des
Körperfettgehaltes (Körperzusammensetzung) in vivo in zwei - zu den statischen
Modellen gehörenden - Hauptgruppen. Die erste Gruppe basiert auf statistischen oder
beschreibenden Fett-Schätzmethoden. Sie hängt von einer geeigneten Referenzmethodik
und der nachfolgenden statistischen Datenaufbereitung zur Entwicklung einer
Schätzgleichung - in den meisten Fällen lineare Regressionsgleichungen - ab. Die
zweite Gruppe - modellabhängige oder mechanistische Fett-Bestimmungsmethoden -
basiert auf festgelegten (etablierten) „konstanten“ Relationen zwischen den
Komponenten der Körperzusammensetzung. Im Gegensatz zu den statischen Modellen
werden dynamische Modelle zur Vorhersage des Wachstumsverlaufs
(Wachstumsmodelle) genutzt (z.B. BALDWIN und SAINZ, 1995, MOUGHAN et al., 1995,
BASTIANELLI und SAUVANT, 1997, EMMANS und KYRIAZAKIS, 1997).
Die meisten mechanistischen Methoden zur Ermittlung der Körperzusammen-
setzung bzw. des Körperfettgehaltes in vivo messen Gesamtgewebe-Komponenten unter
Verwendung eines Zwei-Komponenten- oder eines Drei-Komponenten-Modells
(ROUBENOFF und KEHAYIAS, 1991, PIETROBELLI et al., 1996, BASTIANELLI und
SAUVANT, 1997, ZEMEL et al., 1997, HEYMSFIELD et al., 1997, TÖLLI et al., 1998,
FORBES, 1999, TRIPPO, 2000). Abbildung 2.2.5 gibt einen Überblick zu den
unterschiedlichen Bestandteilen, die zur Schätzung des Gesamtkörperfettgehaltes bei
(Nutz-)Tieren und Menschen genutzt werden. Das einfache von Adolf Magnus-Levy im
Jahre 1906 aufgestellte Zwei-Komponenten-Modell (FORBES, 1999) unterscheidet für
den Gesamtkörper unter Annahme von konstanten spezifischen Dichten für Fett
(≈0,9 g/ml) und den fettfreien Körper (≈1,1 g/ml) bei allen Individuen nur zwischen
Fettmasse und fettfreier Masse (oder Magermasse - ROUBENOFF und KEHAYIAS, 1991).
Die jetzt häufiger verwendeten Drei-Komponenten-Modelle unterteilen die
Gesamtkörpermasse (Gesamtkörpergewicht) in:
I. Fett, Magergewebe bzw. Magerweichgewebe (Wasser und Protein enthaltend)
und Knochen (Knochenminerale) oder
II. Fett, feste Gewebemasse (Protein und Mineralstoffe) und Wasser
(ROEMMICH et al., 1997).

Methodenübersicht
27
Chemisch basiert das Zwei-Komponenten-Modell auf der Unterteilung des
Säugetierkörpers in einerseits Fett sowie andererseits Protein (ursprünglich
Stickstoffbestandteile), Asche (Mineralstoffe) und Wasser (ANDERSON, 1860,
MOULTON, 1929, KEYS und BROZEK, 1953). Diese Unterteilung führt zu einem Vier-
Komponenten-Modell (HÖRNICKE, 1962, EMMANS und KYRIAZAKIS, 1997, ROEMMICH
Et al., 1997, GORAN et al., 1998). Alle physikalischen Methoden zur Ermittlung der
Körperzusammensetzung beruhen letztendlich auf diesen zwei Modellen (LUKASKI,
1987). Ein weiteres Vier-Komponenten-Modell, welches hauptsächlich die
metabolisch aktive Körperzellmasse berücksichtigt, besteht aus den Kompartimenten:
1. Fett (Fettgewebe, Gehirn, Membranen, Bindegewebe und Fettzellen umfassend),
2. Körperzellmasse (Bindegewebe, Fettzellen, feste Zellbestandteile und intrazelluläres
Wasser umfassend),
3. extrazelluläre Flüssigkeit (Plasmavolumen, interstitielles und transzelluläres Wasser
umfassend) sowie
4. Skelettgewebe (Bindegewebe und Knochen umfassend) (PIERSON et al., 1997).
Abb. 2.2.5: Körperkompartimente als Grundlage für verschiedene Modelle derKörperzusammensetzung
Mager-masse♦
Lipid
Knochen-Mineral
Protein
Kohlenhydrat (C)* Weich-Gesamtkör-permasse Mager-
gewebe♣weich-
gewebe♥
(Ca, P)
(N)
Fettfreieoder
Mineral (K, Na, Cl, Ca)
* hauptsächlich Glykogen in Muskeln und Leber (nicht berücksichtigt in 2K-, 3K-, 4K-oder 5K-Modellen – K = Kompartiment)
♣ Skelettmuskelgewebe, innere Organe, Blut und Rest (z.B. Haut)
♦ = ♣ + Knochen (Skelett)
¥ Hauptfettdepots: subkutan, inter- und intramuskulär, abdominal
♥ = ♣ + ¥
¥
(0 ~ 63 %, C ~ 20 %, H ~ 10 %, N ~ 3 % der Gesamtkörpermasse, SAJONSKI und SMOLLICH, 1981)
Wasser (1H, ²H, ³H)
intrazellulär | extrazellulär
(Fett)
Knochen-gewebe
Triglyzeride | Nicht-Triglyzeride

Methodenübersicht
28
Die theoretische Einschränkung des Zwei-Komponenten-Modells resultiert aus
der Annahme, dass die prozentualen Anteile von Mineralien, Wasser und Proteinen in
der fettfreien Masse unabhängig von Alter, Körpergewicht, Körperzusammensetzung
sowie Geschlecht konstant sind und dass Individuen bezogen auf die Körpermasse allein
in ihrem prozentualen Fettgehalt variieren – vorausgesetzt, die fettfreie Masse ergibt
sich aus einer einzigen Messung (MACKIE et al., 1989). Bezogen auf das Schwein,
speziell während der Wachstumsperiode oder in extremen Schweinerassen (sehr mager
bzw. sehr fett) kann diese Annahme nicht gelten (SHENG, 1988, TOPEL und KAUFMANN,
1988). Erst relativ neue technologische Entwicklungen wie die Neutronenaktivierungs-
Analyse (siehe Kapitel 2.2.3.2.5.11) verwenden komplexere Fünf- oder Sechs-
Komponenten-Modelle auf molekularem Niveau (RYDE et al., 1993, HEYMSFIELD et
al., 1997, WANG et al., 1998a, KEHAYIAS und VALTUEÑA, 1999).
Eine modifizierte Herangehensweise gliedert die Zusammensetzung des Körpers
in seine einzelnen atomaren Elemente auf, die in der Addition wieder den Gesamtkörper
bzw. seine Kompartimente beschreiben. Im einfachsten Fall können die zwei
Kompartimente Protein und Knochenmineral jeweils direkt über die Bestimmung eines
einzelnen Elements quantifiziert werden, da sich mehr als 98 % des Körperstickstoffs
(GKN) im Protein bzw. des Körperkalziums (GKCa) im Knochen befinden. Die
Formeln lauten dann:
Gesamtkörper-Protein (GKProtein) = Gesamtkörper-Stickstoff (GKN) • 6,25 bzw.
Knochenmineralgehalt (-asche) (GKAsche) = Gesamtkörper-Kalzium (GKCa)/0,34.
Für die Bestimmung des Körperfettgehaltes kann das Kohlenstoff-Stickstoff-
Kalzium-Modell herangezogen werden. Kohlenstoff (C) ist im Körper vor allem im
Fett (mit ca. 76 - 77 % C) und Protein (53,1 - 53,2 % C) enthalten. Knochenmineralien
bzw. Knochenasche (1,73 - 2 % C) und Glykogen (ca. 44,4 % C) tragen weniger als 3 %
zum Gesamtkohlenstoffgehalt des Körpers (GKC) bei (WANG et al., 1998b, KEHAYIAS
und VALTUEÑA, 1999). Der Körperfettgehalt bzw. zunächst der Kohlenstoffgehalt des
Fettes (CFett) wird aus dem Gesamtkohlenstoffgehalt (GKC) nach Abzug der
Kohlenstoffgehalte von Protein (CProtein), Knochenasche (CKnochen) und Glykogen
(CGlykogen) nach folgender Formel berechnet:

Methodenübersicht
29
CFett = GKC – [CKnochen + CProtein + CGlykogen]
mit
CProtein = GKN • 3,31,
CKnochen = GKN • 0,12 und
CGlykogen = GKCa • 0,05.
GKC, GKN und GKCa werden mittels „inelastischer“ Neutronenstreuung, „prompter“
Neutronenaktivierung bzw. „verzögerter“ Neutronenaktivierung gemessen (siehe
Kapitel 2.2.3.2.5.11). Schließlich ergibt sich der Fettgehalt über die Beziehung zwischen
Kohlenstoffgehalt des Fettes und dem Triglyzeridgehalt:
Gesamtkörper-Fett (GKFett) = CFett/0,77.
Ein weiteres Modell zur Ermittlung des Fettgehaltes nutzt unter der Annahme
eines fixen Wassergehaltes der fettfreien Masse von ~73 % das Verhältnis zwischen
Kohlenstoff (C) und Sauerstoff (O) innerhalb einzelner Geweberegionen bzw. für den
Gesamtkörper. Sauerstoff wird wie Kohlenstoff mit Hilfe der „inelastischen“
Neutronenstreuung gemessen, wobei Körperwasser (mit ca. 88,8 % O im Wasser),
Protein (22,6 % O), Fett (11,2 % O), Knochenasche (41,6 % O) und Glykogen (49,3 %
O) die Hauptträger des Gesamtkörpersauerstoffs (GKO) darstellen. Die Formel lautet:
Fett = [0,722 • C/O – 0,116]/[0,610 • C/O + 0,653](KEHAYIAS et al., 2000).
Neben dem durchschnittlich „konstanten“ Wassergehalt in der fettfreien Masse in neun
verschiedenen Säugetierarten einschließlich des Menschen von 73,9 ± 0,15 % (WANG et
al., 1999) existiert ein weiteres „konstantes“ Verhältnis zwischen Gesamtkörper-
Sauerstoff und kohlenstofffreier Körpermasse in der Höhe von 0,8 ± 0,009 bei Männern
(WANG et al., 1998b). Allerdings werden 58 % der Variation im Verhältnis von
Gesamtkörper-Sauerstoff und kohlenstofffreier Körpermasse (0,81 zu 0,77) durch den
Körperfettgehalt (5 - 40 %) erklärt.
In Abhängigkeit von der Messtechnik unterscheiden WANG et al. (1992) zwischen
Modellen auf:
I. atomarem Niveau (den Körper in seine atomaren Bestandteile aufteilend),
II. molekularem Niveau (den Körper in chemisch unterschiedliche Komponenten
wie Lipid, Protein, Kohlenhydrate, Wasser und Mineralien aufteilend),
III. zellulärem Niveau (Zellmasse, extrazelluläre Flüssigkeit und extrazelluläre
Feststoffe),

Methodenübersicht
30
IV. Gewebesystem-Niveau (Fettgewebe, Skelettmuskelgewebe, innere Organe,
Hautgewebe und Knochengewebe) und
V. Gesamtkörper-Niveau (Körpergewicht oder Körpervolumen).
Dabei kombinieren die meisten Modelle für die Körperzusammensetzung mindestens
zwei der vorab genannten Niveaus (Abb. 2.2.4 und 2.2.5). Auf molekularem Niveau
machen Fett, Wasser, Protein, Zellmineralien und Knochenmineralien bereits ca. 96 %
des Körpergewichts eines Schweines aus (SAJONSKI und SMOLLICH, 1981, SHULMAN,
1993). Oft wird der Kohlenhydratgehalt des Körpers (insbesondere Glykogen und
Glukose) einfach ignoriert, was zu einer leichten Überschätzung von mindestens einer
der Komponenten Fett, Protein oder Wasser - ohne Zell- bzw. Knochenmineralstoffe
einzuschließen - führt. Das höhere Modell-Niveau besteht immer aus Komponenten des
jeweils niedrigeren Niveaus. Ein zweites, mehr in Frage zu stellendes Konzept geht von
der Existenz eines „Gleichgewichts der Körperzusammensetzung“ aus, welches
konstante Verhältnisse zwischen den Komponenten der Körperzusammensetzung zur
Folge hat, solange das Körpergewicht und der Hydrierungs-Status unverändert bleiben
(FOGELHOLM und MARKEN LICHTENBELT, 1997, HEYMSFIELD et al., 1997, WANG et al.,
1998b, 1999). Ein Beispiel ist das angenommene konstante Verhältnis von 4/5 zwischen
Fettmasse auf molekularem Niveau und Fettgewebemasse auf dem Gewebesystem-
Niveau. Dieses Verhältnis berücksichtigt nicht die möglichen unterschiedlichen Anteile
der sogenannten „leeren Fettzellen“ und die signifikanten Veränderungen des Anteils
der „Nicht-Triglyzerid-Lipide“ innerhalb der Lipidfraktion des Gesamtkörpers speziell
während Situationen mit einer negativen Energiebalance, die aus absoluten bzw.
relativen Restriktionen in der Futteraufnahme und der körperlichen Aktivität resultieren
(ENDER u.a., 1997a, COMIZIO et al., 1998). Allerdings ändern sich unter diesen
Bedingungen die postulierten stabilen Körpergewichte und Hydrierungszustände.
Während des Wachstums nimmt der Wassergehalt in der fettfreien Masse ab, aber mit
steigendem Alter nach Beendigung des Wachstums oder bei stärkerer Verfettung nimmt
er tendenziell wieder zu (WANG et al., 1999).
Da Kalibrierungsdaten (Referenzwerte) für Regressionsgleichungen häufig anhand
spezifischer Populationen gewonnen und überprüft wurden, können Schätzgleichungen
nicht ohne weiteres auf Individuen anderer Populationen übertragen werden. Um genaue
Schätzgleichungen für die verschiedensten Messmethoden der Körperzusammensetzung
entwickeln und Differenzen in der Zusammensetzung des fettfreien Körpers aufgrund

Methodenübersicht
31
der Unterschiede zwischen Zuchtlinien, extremen Rationen, Geschlechts- und
Altersgruppen quantifizieren zu können, sollten Mehr-Komponenten-Modelle
angewandt werden, die die individuelle Variation im Wasser- und Mineralgehalt der
fettfreien Komponente berücksichtigen. Gleichzeitig ist es erforderlich, alle
Fehlerquellen wie z.B. den Referenzmethodefehler (z.B. chemische Analyse,
Vollzerlegung), den Modellfehler und den Messfehler weitestgehend auszuschalten
(GUO et al., 1996, ATKINSON und NEVILL, 1998, WANG et al., 1999).
2.2.3 Methoden der Leistungsprüfung
2.2.3.1 Subjektive Methoden
Visuelle und taktile Beurteilungsverfahren (Konditions- oder Exterieurbenotungen im
Hinblick auf Verfettungs- bzw. Bemuskelungsgrad) waren beginnend mit der
Domestikation vor 11000 – 7000 Jahren für Jahrhunderte (Jahrtausende) die einzigen
Methoden zur züchterischen Selektion bzw. zur Einschätzung der
Körperzusammensetzung (Abb. 2.2.6). Auch heute dienen sie noch immer zur linearen
bzw. nicht linearen Bewertung von Schweinebeständen (SCHULZE u.a., 1998).
Diese einfachen Methoden der Leistungsprüfung resultierten in einer Vielzahl von
Schweinerassen mit einer weiten Variation in ihrer Körperzusammensetzung. Dabei
stehen die extrem bemuskelten (am RyR1-Genort homozygot „defekten“) Tiere der
Rasse Pietrain auf der einen und die sehr fruchtbaren, aber gleichzeitig sehr fetten Tiere
chinesischer Herkünfte wie Meishan, Minzhu bzw. Fenjing auf der anderen Seite des
Spektrums. Die charakteristische Form von fleischreichen Schweinelinien kann als
relativ kurz und gedrungen mit gut ausgeprägten, runden Schinken und einer mehr
Ω-förmigen als n-förmigen Hinteransicht beschrieben werden (WHITTEMORE, 1987).
Eine ausgeprägte Muskelhypertrophie wirkt sich beim Schwein durch eine Mutation am
Ryanodin-Rezeptor-1-Genort (RyR1) ebenso wie die „Doppellendigkeit“ beim Rind
besonders stark auf die Keulen- bzw. Schinkenregion aus (BERG und BUTTERFIELD,
1968, FEWSON, 1987). Im Gegensatz zur Mutation am RyR1-Genort, die ausführlich im
Kapitel 4.1 diskutiert wird, ist die „Doppellendigkeit“ beim Rind nicht auf
Muskelhypertrophie sondern Muskelhyperplasie zurückzuführen. Die
Muskelhyperplasie wird durch eine auch bei der Maus nachgewiesene Mutation am
Myostatin-Genort hervorgerufen (z.B. MCPHERRON et al., 1997, VARGA et al., 1997,
SZABO G. et al., 1998, SONSTEGARD et al., 1998). Weitere Genorte, die die phänotypisch

Methodenübersicht
32
messbare Körperzusammensetzung und speziell den Bemuskelungs- und
Verfettungsgrad beeinflussen, werden ausführlich in Kapitel 4.2 erläutert.
Inzwischen sind Exterieurmerkmale, die die Größe und Struktur des Körpers
beschreiben, nicht mehr länger relevant zur Bestimmung von Wachstumsleistung und
Körperzusammensetzung (VAN STEENBERGEN, 1990), da verschiedene Techniken eine
objektivere und genauere Messung der Körperzusammensetzung am lebenden Tier
ermöglichen (Abb. 2.2.6). Häufig dienen hoch korrelierte Hilfsmerkmale am lebenden
Tier wie Rückenspeckdicke oder –fläche und Rückenmuskeldicke bzw. -fläche zur
Schätzung der Gesamtkörperzusammensetzung. Es ist jedoch zum Teil immer noch
schwierig, diese indirekten Verfahren in sehr großen Schweineproduktionsbetrieben
umzusetzen. Als Alternative zur Schätzung der Körperzusammensetzung beschreiben
CHARETTE et al. (1996) die Bewertung der Körperkondition von Sauen mit Hilfe eines
linearen Benotungsverfahrens, welches auf der Messung morphologischer
Hilfsmerkmale in vivo beruht. Dabei weist die Kombination von linearen Messwerten
(Breite über den „Hinterschinken“, Pelvishöhe) mit „halb-quantitativen“ Noten (visuelle
und taktilen [Palpations-] Noten für die THORAX-Morphologie und den SCHWANZ-
Ansatz) mit 0,91 eine höhere Wiederholbarkeit (Verhältnis von Tier- zu Gesamtvarianz)
auf als mit 0,77 die Körperkonditions-Note (PATIENCE und THACKER, 1989). Die
Differenz wurde durch den größeren BEWERTER-Effekt für die Körperkonditionsnote
verursacht, was auf eine sehr subjektive Bewertungstechnik hinweist. Beide von
CHARETTE et al. (1996) beschriebenen Verfahren sind jedoch nicht mit Hilfe objektiver
Referenzdaten wie z.B. aus der chemischen Schlachtkörperzusammensetzung überprüft
worden.
Generell kann eine subjektive Körperkonditionsbewertung keinen züchterischen
Informationsgewinn zu einer bereits „ausreichenden“ objektiven Bewertung (Schätzung)
der tatsächlichen Körperzusammensetzung von Nutztieren beitragen. So ermittelten
SCHULZE u.a. (1998) für die Rückenspeckdicke Heritabilitäten von 0,55 für Large White
und 0,37 für Landrasse, während die Heritabilitäten für das Merkmal Bemuskelung aus
der nicht linearen Exterieurbeschreibung nur bei 0,37 bzw. 0,29 lagen. Die genetischen
Korrelationen (rg) zwischen Rückenspeckdicke und Bemuskelung betrugen 0,64 bzw.
0,42.

Methodenübersicht
33
Abb. 2.2.6: Geschichtliche Entwicklung der Methoden zur Bestimmung derKörperzusammensetzung bei Nutztieren (modifiziert nach FORBES, 1999, PIERSON et al.,2000 und MITCHELL und SCHOLZ, 2001)
2.2.3.2 Physikalische und chemische Methoden
2.2.3.2.1 Körpermasse
Mit steigender Körpermasse (und steigendem Alter) nimmt nicht allein beim Schwein
(z.B. SHIELDS et al., 1983, PFEIFFER u.a., 1984, BIKKER et al., 1995, 1996a, WHITE et al.,
1995, KUHN, M. u.a., 1997, WAGNER et al., 1999), sondern auch beim Menschen
(SIERVOGEL et al., 1998, GUO et al., 1999) und seinen Haustieren (Hund und Katze,
HARPER, 1998) in der Regel der prozentuale Körperfettanteil zu bzw. sinkt der
Magergewebeanteil. Daraus resultiert eine hohe positive Korrelation zwischen
DNA-Analyse (RyR1, RN, LEP ...)
CT, MRT (MRS), DXA, TOBEC, BIA
SPA, DPA,Nahinfrarot-SpektroskopieNeutronenaktivierung, Ultraschall
Metall-„Ruler“, K-40-Zählmethode
Körperdichte
D2O-Verdünnung, Radiographie
Stoffwechsel-Kalorimetrie
Blutvolumen
Fettfreie Körpermasse
„Chemische“ AnalyseZerlegung
Visuelle und taktileExterieurbewertung
2000
1950
1900
Domestikation
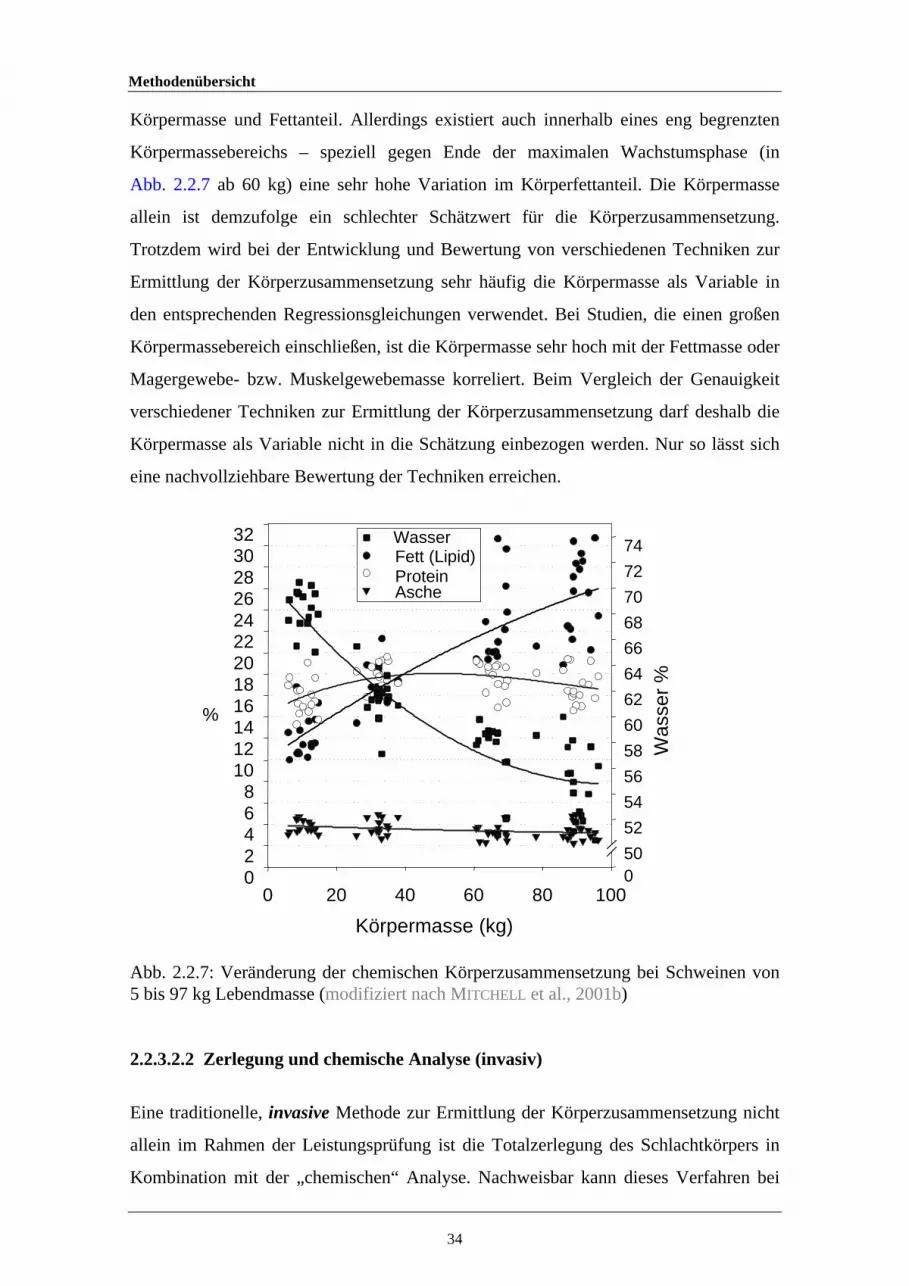
Methodenübersicht
34
Körpermasse und Fettanteil. Allerdings existiert auch innerhalb eines eng begrenzten
Körpermassebereichs – speziell gegen Ende der maximalen Wachstumsphase (in
Abb. 2.2.7 ab 60 kg) eine sehr hohe Variation im Körperfettanteil. Die Körpermasse
allein ist demzufolge ein schlechter Schätzwert für die Körperzusammensetzung.
Trotzdem wird bei der Entwicklung und Bewertung von verschiedenen Techniken zur
Ermittlung der Körperzusammensetzung sehr häufig die Körpermasse als Variable in
den entsprechenden Regressionsgleichungen verwendet. Bei Studien, die einen großen
Körpermassebereich einschließen, ist die Körpermasse sehr hoch mit der Fettmasse oder
Magergewebe- bzw. Muskelgewebemasse korreliert. Beim Vergleich der Genauigkeit
verschiedener Techniken zur Ermittlung der Körperzusammensetzung darf deshalb die
Körpermasse als Variable nicht in die Schätzung einbezogen werden. Nur so lässt sich
eine nachvollziehbare Bewertung der Techniken erreichen.
Abb. 2.2.7: Veränderung der chemischen Körperzusammensetzung bei Schweinen von5 bis 97 kg Lebendmasse (modifiziert nach MITCHELL et al., 2001b)
2.2.3.2.2 Zerlegung und chemische Analyse (invasiv)
Eine traditionelle, invasive Methode zur Ermittlung der Körperzusammensetzung nicht
allein im Rahmen der Leistungsprüfung ist die Totalzerlegung des Schlachtkörpers in
Kombination mit der „chemischen“ Analyse. Nachweisbar kann dieses Verfahren bei
Körpermasse (kg)
0 20 40 60 80 100
%
02468
101214161820222426283032
Was
ser
%
050
52
54
56
58
60
62
64
66
68
70
72
74Fett (Lipid)ProteinAsche
Wasser

Methodenübersicht
35
Schwein, Rind und Schaf (Tab. 2.1.1, Abb. 2.2.6) erstmals auf Mitte des 19.
Jahrhunderts im Rahmen der „Rothamsted Agricultural Experiments“ (England) von
John Bennet LAWES und Joseph Henry GILBERT datiert werden (ANDERSON 1860,
CAMERON, 1868, HALL, 1905).
Tab. 2.2.1: Schlachtkörperzusammensetzung von zehn Tieren in unterschiedlichen
Maststadien (nach den Angaben von ANDERSON 1860, CAMERON, 1868, HALL, 1905)
Tierart und
Verfettungsgrad
Anteil im Schlachtkörper
– einschließlich Nieren
und Nieren umgebendes
Fett (in %)
Anteil im Schlachtabfall
einschließlich Haut1),
Füße1), Kopf1), innere
Organe2), aber ohne Inhalt
des Verdauungstraktes –
Mägen und Därme)
(in %)
Anteil am Gesamtkörper bezogen
auf Lebendmasse; nüchtern (in %)
Min
eral
stof
fe
Stic
ksto
ff-
best
andt
eile
Fett
Tro
cken
-su
bsta
nz in
sg.
Was
ser
Min
eral
stof
fe
Stic
ksto
ff-
best
andt
eile
Fett
Tro
cken
-su
bsta
nz in
sg.
Was
ser
Min
eral
stof
fe
Stic
ksto
ff-
best
andt
eile
Fett
Tro
cken
-su
bsta
nz in
sg.
Mag
en u
nd
In-
nere
ien-
Inha
ltFr
isch
subs
t. 4)
Was
ser
Fettes Kalb 4,48 16,6 16,6 37,7 62,3 3,41 17,1 14,6 35,1 64,9 3,80 15,2 14,8 33,8 3,17 63,0
Halb-fetter Ochse 5,56 17,8 22,6 46,0 54,0 4,05 20,6 15,7 40,4 59,6 4,66 16,6 19,1 40,3 8,19 51,5
Fetter Ochse 4,56 15,0 34,8 54,4 45,6 3,40 17,5 26,3 47,2 52,8 3,92 14,5 30,1 48,5 5,98 45,5
Fettes Lamm 3,63 10,9 36,9 51,4 48,6 2,45 18,9 20,1 41,5 58,5 2,94 12,3 28,5 43,7 8,54 47,8
Mageres Schaf (wachsend) 4,36 14,5 23,8 42,7 57,3 2,19 18,0 16,1 36,3 63,7 3,16 14,8 18,7 36,7 6,0 57,3
Halb-fettes, altes Schaf 4,13 14,9 31,3 50,3 49,7 2,72 17,7 18,5 38,9 61,1 3,17 14,0 23,5 40,7 9,05 50,2
Fettes Schaf 3,45 11,5 45,4 60,3 39,7 2,32 16,1 26,4 44,8 55,2 2,81 12,2 35,6 50,6 6,02 43,4
Extra fettes Schaf 2,77 9,1 55,1 67,0 33,0 3,64 16,8 34,5 54,9 45,1 2,90 10,9 45,8 59,6 5,18 35,2
Mageres Schwein
(wachsend)
2,57 14,0 28,1 44,7 55,3 3,07 14,0 15,0 32,1 67,9 2,67 13,7 23,3 39,7 5,22 55,1
Fettes Schwein 1,40 10,5 49,5 61,4 38,6 2,97 14,8 22,8 40,6 59,4 1,65 10,9 42,2 54,7 3,97 41,3
∅ Rind3) 4,87 16,5 24,7 46,0 54,0 3,62 18,4 18,9 40,9 59,1 4,13 15,4 21,3 40,9 5,78 53,3
∅ Schaf3) 3,67 12,2 38,5 54,3 45,7 2,66 17,5 23,1 43,3 56,7 3,00 12,8 30,4 46,3 6,96 46,8
∅ Schwein3) 1,99 12,2 38,8 53,0 47,0 3,02 14,4 18,9 36,4 63,6 2,16 12,3 32,8 47,2 4,60 48,2
1) beim Schwein zählen Haut, Füße und Kopf nicht zum Schlachtabfall, 2) ohne Nieren und Nierenumgebendes Fett 3) eigene Berechnungen, 4) muss zum Trockensubstanz- und Wassergehalt addiertwerden, um 100 % zu erreichen
Systematische Zerlegungen im Rahmen der Leistungsprüfung beim Schwein
begannen in Deutschland unter der Leitung von Prof. J. Schmidt am Versuchsgut
Friedland der Universität Göttingen im Jahr 1926 (SCHMIDT u.a., 1931, BLENDL u.a.,
1986). Seit diesen ersten Untersuchungen bildet die Zerlegung in Kombination mit der
chemischen Analyse von Fett-, Wasser-, Protein- und Aschegehalt die Grundlage für das
vorhandene Wissen über die Körperzusammensetzung des Schweines (ANDERSON,

Methodenübersicht
36
1860, SCHMIDT u.a., 1931, MCMEEKAN, 1940, WALSTRA, 1980, WAGNER et al., 1999).
Beide Verfahren dienen nach wie vor als Standardmessverfahren zur Bewertung anderer
bzw. neuer Methoden im Bereich der (Nutz-)Tierwissenschaften (GRIEP, 1991, KIENZLE
et al., 1998). Totalzerlegung (grobgewebliche Zerlegung) und chemische Analyse sind
jedoch in Kombination und einzeln sehr zeitaufwendig und können aufgrund der
anatomischen Gegebenheiten nicht 100%-ig fehlerfrei durchgeführt werden. Eine
detaillierte quantitative Totalzerlegung, wie bei CUTHBERTSON und POMEROY (1962)
beschrieben, nimmt im Durchschnitt 110 Arbeitskraft-Stunden je Schlachthälfte in
Anspruch. Schlachtkörperzerlegungen werden mit unterschiedlichem Aufwand
betrieben. Eine „einfache“ Zerlegung umfasst die (teilweise abgespeckten) „wertvollen“
Teilstücke Kamm, Bug mit Eisbein vorn, Brustspitze, Kotelett, Filet, Bauch mit
Wamme, Schinken mit „Zu Wamme“, Eisbein hinten sowie die Abschnitte Kopf,
Spitzbeine, Schwanz, Flomen und Niere. Andere Varianten beinhalten eine Zerlegung in
Organe und Gewebe oder eine Zerlegung von Schlachtkörperhälften bzw. –teilstücken
in Fett (subkutanes Fett inklusive Haut + intermuskuläres Fett), Magergewebe (Muskeln
+ Bindegewebe + Sehnen) und Knochen bis hin zu einer detaillierten Totalzerlegung in
individuelle Muskeln, Knochen, Sehnen, Schwarten (Haut) und Fettschichten (z.B.
SCHEPER und SCHOLZ, 1985, OSTER u.a., 1987, WALSTRA und MERKUS, 1995). Die
chemische Analyse dient in den meisten Fällen zur Bestimmung des Gehalts an Rohfett
(Ether-Extraktion mit oder ohne Salzsäureaufschluss), Rohprotein (Stickstoff • 6,25),
Trockensubstanz (Wasser) und Rohasche. Unabhängig von Objekt und Art der
Probengewinnung (Gesamtkörper, leerer Schlachtkörper, Schlachthälfte, individuelle
Teilstücke mittels Zermahlung, axiale Schnitte oder „Sägemehl“ nach axialer Zerlegung
in „Scheiben“) kommt es immer darauf an, eine homogene, repräsentative und
vollständige Probe zu untersuchen. Die meisten Studien, die auf einer Zerlegung und
chemischen Analyse beruhen, beschreiben „einfach“ die Körperzusammensetzung von
Schweinen unterschiedlicher genetischer Herkünfte und Geschlechter während
verschiedener Wachstums- und Entwicklungsphasen (SCHMIDT u.a., 1931,
DOORNENBAL, 1971, 1972, KNUDSON et al., 1985, SUSENBETH und KEITEL, 1988,
SCHMIDT, 1988, GU et al., 1992, WHITE et al., 1995, QUINIOUS und NOBLET, 1995,
WAGNER et al., 1999), zum Zeitpunkt der Vermarktung (ROTHFUSS, 1981, JOHNSON et
al., 1990, RATHFELDER, 1991, KUHN u.a., 1997, 1998), nach unterschiedlicher
Ernährung bzw. Aufzuchtsintensität (NÜRNBERG u.a., 1988, KAUFMANN, 1990, FRIESEN

Methodenübersicht
37
et al., 1994, NOLD et al., 1999) bzw. nach dem Ansatz von HUXLEY (1932, S. 4 - 9:
y = bxβ/a è mit β/a = k è log y = log b + k log x ) zur Darstellung des allometrischen
Wachstums (DAVIES, 1984, TESS et al., 1986, FORTIN et al., 1987, FEWSON u.a., 1990,
Ž GUR, 1991, KOUBA et al., 1999, WAGNER et al., 1999).
Ein Großteil dieser Daten ist jedoch nur auf bestimmte Situationen /Zeitpunkte
während des Wachstums und der Entwicklung übertragbar und kann nicht zur
Abschätzung der Gesamt-Schlachtkörperqualität aus Teilinformationen herangezogen
werden (ORCUTT et al., 1990). Die separierbaren Gewebeanteile erklären jedoch vor
allem im Rückenmuskelbereich zwischen 9. und 14. Rippe einen hohen Anteil der
Variation im Magerfleischanteil der Schlachthälfte (ORCUTT et al., 1990, SWENSEN et
al., 1998).
2.2.3.2.3. Densitometrie (spezifische Dichte) - hydro- und aerostatische Methoden
Die Analyse der spezifischen Dichte, die in der Regel als Unterwasserwägung
(-volumenverdrängung) durchgeführt wird, ist vor allem im Bereich der Forschung zur
Körperzusammensetzung am Menschen eingesetzt worden (LUKASKI, 1987).
Vergleichbare Studien erfolgten an lebenden Tieren bzw. am Schlachtkörper beim
Schwein, Rind, Schaf und Ziege (Übersichten bei STREITZ, 1995, TREITL, 1998,
MITCHELL und SCHOLZ, 2001).
Die Ermittlung der Körperzusammensetzung mittels Densitometrie oder
Spezifischer Dichte basiert auf einem Zwei-Komponenten-Modell. Die Differenzen in
der spezifischen Dichte zwischen Fett und fettfreier Masse dienen als
Unterscheidungskriterium, wobei Wasser und Knochen als prozentual konstante
Bestandteile der fettfreien Masse angesehen werden. Gebräuchliche Angaben der
spezifischen Dichten sind für Fett = 0,9 g/ml, Wasser = 0,99 g/ml, Magergewebe bzw.
Muskelgewebe = 1,1 g/ml, Protein + Glykogen = 1,34 g/ml, Knochen = 1,8 g/ml und
Gesamtkörpermineral = 3,042 g/ml (SIRI, 1956, LOHMANN, 1984, SICOLNOLFI et al.,
1995, JEBB, 1997, GORAN et al., 1998).
Da Knochen- und Muskelgewebe dichter als Fettgewebe sind, wird ein fettarmer
Körper unter Wasser vergleichsweise mehr „wiegen“, ein Körper mit einem höheren
Fettanteil vergleichsweise weniger. Der Test, bei dem der Proband bis zu 30 Sekunden
„bei ausgeatmeter Luft samt Kopf“ untertauchen muss, vergleicht das

Methodenübersicht
38
„Unterwassergewicht“ mit dem „normalen“ Gewicht. Am lebenden Schwein ist dieses
Verfahren zur Errechnung des Körperfettanteils nicht realisierbar.
Nach Ermittlung der spezifischen Dichte mit Hilfe der „Unterwasserwägung“ oder
einer anderen Volumenmessung wie z.B. mit der akustischen bzw. „Luftverdrängungs-“
Plethysmographie (SHENG et al., 1988, SARDINHA et al., 1998) oder einem aus der
Textilbranche stammenden dreidimensionalen Photonen- bzw. Laser-Scanverfahrens
(WELLS et al., 2000) basiert die Schätzung der Körperzusammensetzung im
Allgemeinen auf experimentell ermittelten Regressionsgleichungen.
Verschiedene Autoren schlugen unter statischen Bedingungen folgende
empirische Beziehungen zwischen prozentualem Körperfettgehalt und spezifischer
Dichte des Gesamtkörpers (σ, in g/ml) vor:
Körperfett % = [(5,405/σ) - 4,914] • 100 KRAYBILL et al. (1953)
Körperfett % = [(4,950/σ) - 4,500] • 100 SIRI (1956)
Körperfett % = [(4,570/σ) - 4,142] • 100 BROZEK et al. (1963)
Körperfett % = [(4,971/σ) - 4,519] • 100 BROZEK (1966)
Körperfett % = [(5,300/σ) - 4,890] • 100 LOHMANN et al. (1984).
Bei jüngeren, wachsenden Individuen (Tieren) bringen die Gleichungen nur
unbefriedigende Genauigkeiten, da der Wassergehalt der fettfreien Masse und damit die
spezifische Dichte der fettfreien Masse nicht konstant ist (SHIELDS et al., 1983, ALOIA et
al., 1998, ELOWSSON et al., 1998, siehe auch Kapitel 2.2.3.2.5.4 – DXA und 2.2.3.3.1 -
Verdünnungsmethoden). Wenn nach DEURENBERG et al. (1989), ROEMMICH et al.
(1997) sowie ROGOL (1997) die Gleichung von SIRI (1956) - besonders bei Probanden
mit einem sehr hohen Fettgehalt bzw. bei wachsenden Individuen (Kindern) - zu einer
systematischen Überschätzung des Körperfettanteils um 2 - 4 % führt, gilt dieselbe
Aussage für die Gleichungen von KRAYBILL et al. (1953) bzw. von BROZEK et al. (1963)
und BROZEK (1966). Allein die Gleichung von LOHMANN et al. (1984) scheint den
Verhältnissen beim wachsenden Schwein am ehesten nahe zu kommen (ELOWSSON et
al., 1998). Da diese Gleichung speziell für wachsende Individuen (Kinder, Jugendliche,
Erwachsene < 25 Jahre) entwickelt wurde, dient sie bevorzugt humanmedizinischen
Untersuchungen, die über einen längeren Zeitraum die Veränderung der
Körperzusammensetzung (bis zu einem Alter von 65 Jahren) verfolgen (SIERVOGEL et
al., 1998).

Methodenübersicht
39
Eine spezifische Dichte für den Gesamtkörper von z.B. 1,05 g/ml führt nach der
Gleichung von SIRI (1956) zu einem Körperfettgehalt von 21,43 %, nach BROZEK et al.
(1963) zu 21,04 %, nach BROZEK (1966) zu 21,53 % und nach KRAYBILL et al. (1953)
zu 23,36 %, während die Gleichung von LOHMANN et al. (1984) den niedrigsten
rechnerischen Körperfettgehalt von 15,76 % ergibt. Alle Formeln ergeben spätestens ab
einer spezifische Dichte für den Gesamtkörper von ≥ 1,103332 g/ml negative
Körperfettgehalte (%), da ~1,1 g/ml die angenommene Dichte für die Mager-
gewebemasse darstellt.
Unter Bedingungen, die einen Gewichtsverlust bzw. eine Gewichtszunahme und
damit eine Änderung der Körperzusammensetzung zur Folge haben, müssen die
„statischen“ Formeln modifiziert werden (BROZEK, 1966, LOHMANN, 1984). Um die
nachgewiesenen Mängel der Zwei-Komponenten-Modelle zu umgehen, schlagen
LOHMANN (1984), ROEMMICH et al. (1997) sowie ELOWSSON et al. (1998) speziell für
wachsende Individuen bzw. für Individuen, deren Körperzusammensetzung sich ändert,
vor, die bereits beschriebenen Drei- bzw. vorzugsweise Vier-Komponenten-Modelle zu
verwenden oder weiter zu entwickeln.
Als erhebliche Fehlerquellen gehen in diese Methoden - neben den Variationen
des Wassergehalts in der fettfreien Masse und des Muskel-zu-Knochen-Verhältnisses
(SHIELDS et al., 1983) - die mit „Luft“ gefüllten Räume des Körpers ein. Den Gasinhalt
des Magen-Darm-Traktes kann man nicht genau bestimmen. Er wird mit 50 - 300 ml
angenommen. Mit dem Alter nimmt die Mineraldichte der Knochen wie auch das
Körperwasser ab und somit auch das spezifische Gewicht der fettfreien Masse. Bei
Tieren mit einer größeren Knochendichte ist das spezifische Gewicht der fettfreien
Masse höher (FOGELHOLM et al., 1996).
Neben der Unterwasserwägung wird zur Ermittlung der spezifischen Dichte des
Gesamtkörpers die bereits erwähnte akustische Plethysmographie (SHENG et al., 1988)
genutzt. Diese Methode nutzt das Prinzip des Helmholtzresonators, bei welchem sich
die Resonanzfrequenz einer Kammer umgekehrt proportional zur Quadratwurzel des
Luftvolumens innerhalb der Kammer verhält. Nachdem ein Objekt in der Kammer
platziert wurde, kann die Veränderung der Resonanzfrequenz herangezogen werden, um
das Volumen des Objekts zu messen. Die anhand von 13 Miniatur-Schweinen (Alter: 5 -
18 Tage, Gewicht: 1253 g - 2631 g) durchgeführten Wiederholungsmessungen des
Körpervolumens resultierten in Variationskoeffizienten zwischen 0,3 und 3,2 % (SHENG

Methodenübersicht
40
et al., 1988), wobei diese Messwerte nicht signifikant von den Körpervolumina aus der
chemischen Analyse (Summe der Volumina von Gesamtkörperwasser, Fett, Protein und
Mineralstoffen) abwichen.
Insgesamt ist für beide Methoden, Unterwasserwägung und akustische
Plethysmographie, festzustellen, dass sie für die Verwendung am lebenden Schwein
aufgrund der Verfahrensgestaltung als ungeeignet erscheinen.
2.2.3.2.4. Lineare Messungen
Der Körper oder die Schlachthälfte eines Schweines bietet selbst eine Vielzahl von
möglichen linearen Messstellen an. Körpergröße, Körperlänge, Brust- bzw.
Rumpfumfang und -tiefe, Brust- bzw. Beckenbreite, Vorder- und
Hintergliedmaßenlänge bzw. –umfang sind nur einige Beispiele. In den meisten Fällen –
insbesondere beim Schwein - gestatten diese Maße jedoch keine hinreichende
Schätzung der Körperzusammensetzung und dienen allein zur linearen bzw. nicht-
linearen Beschreibung der äußeren Körpermaße von Individuen bzw. Populationen
(LÓPEZ FERNÁNDEZ, et al., 1992, KRÄUßLICH, 1998, SCHULZE u.a., 1998). Zwei
Parameter bilden sowohl am lebenden Tier als auch am Schlachtkörper eine Ausnahme.
Rückenspeckdicke bzw. –fläche und Rückenmuskeldicke bzw. -fläche sind aufgrund der
relativ engen Korrelationen (r ≥ |±0,45|) geeignete Hilfsmerkmale zur Schätzung von
Fett- und Fleischanteil und werden seit Etablierung der Echolotmessung für die
Leistungsprüfung von Zuchttieren bzw. im Rahmen von Wachstumsversuchen
verwendet (RITTLER, 1969, HORST, 1971, FENDER u.a., 1979, PFEIFFER u.a., 1984,
SATHER et al., 1986, 1987, MOELLER et al., 1998; siehe auch Kapitel 4.2). Vor der
Einführung der Ultraschallmessungen erfolgte die Messung der Rückenspeckdicke nach
der „Ruler“-Methodik (HAZEL und KLINE, 1952). Mit einem Skalpell oder „scharfem“
Messer wurde die Haut über der Messstelle durchtrennt und der Messstab („Ruler“)
wurde bis zum Erreichen des Muskelstranges in das Rückenfettgewebe eingeführt, so
dass die Rückenspeckdicke abgelesen werden konnte (Abb. 2.2.8).

Methodenübersicht
41
Abb. 2.2.8: Messung der Rückenspeckdicke mittels „Ruler“ oder „Back Fat Probe“(Bild von William G. Luce, Spezialberater Schweine der Oklahoma State University)
Voraussetzungen für hohe Korrelationen (hohe Schätzgenauigkeiten) bilden
wiederum hohe Variationen für Rückenmuskelfläche und Rückenfettfläche bzw. –dicke
innerhalb eines engen Gewichtsbereiches innerhalb der betrachteten Populationen.
2.2.3.2.5 Radiologische und bildgebende Methoden im Überblick
Eine frühzeitige objektive und wiederholbare Bestimmung der Körperzusammensetzung
(in vivo) ist für praktische Tierzüchter, Nutztierwissenschaftler und Veterinärmediziner
gleichermaßen von Interesse, um einerseits das Generationsintervall im Rahmen der
Selektion und den Produktionsprozess optimal gestalten zu können bzw. andererseits
genetische, physiologische, ernährungs- und haltungsbedingte sowie wirtschaftliche
Fragestellungen zu beantworten. Die Entwicklung von neuen, leistungsfähigen
Methoden wurde vor allem durch die Humanmedizin vorangetrieben, wovon der
Nutztierbereich mit zeitlicher Verzögerung profitierte (FOSTER und FOWLER, 1988).
Übersichten zu genutzten Verfahren in der Humanmedizin bzw. in den
(Nutz-)Tierwissenschaften für die Bestimmung der Körperzusammensetzung finden
sich bei LUKASKI (1987), JEBB (1997) und ELLIS (2000) bzw. STREITZ (1995),
STANFORD et al. (1998a), TREITL (1998) sowie MITCHELL und SCHOLZ (2001). Auf
radiologische bzw. bildgebende Techniken beschränken sich WELLS (1991),
HEYMSFIELD et al. (1997b), PLOURDE (1997) sowie SZABO et al. (1999). Die
radiologischen Techniken umfassen die (Röntgen-)Computer-Tomographie ((R)CT)

Methodenübersicht
42
oder „Computer-Aided-Tomographie“ bzw. Computer-Axial-Tomographie (CAT),
Magnetresonanz-Tomographie (MRT), bzw. -Spektroskopie - MRS), Photonen-
Absorptiometrie (Single- SPA oder Dual- DPA), Dualenergie-Röntgenabsorptiometrie
(DXA) und Neutronenaktivierungs-Analyse (NAA).
Erste erfolgversprechende Arbeiten zur Nutzung der Röntgenmesstechnik
(Röntgen-Durchleuchtung) für die Bestimmung des „erblich-konstitutionell“ bedingten
Fettwuchses am lebenden Schwein führte HOGREVE (1938a,b) in Berlin-Dahlem an der
damaligen Friedrich-Wilhelm-Universität durch. Bereits KRONACHER und HOGREVE
(1936) versuchten die Variation der Schinkenform mit derselben Technik zu ermitteln.
Der Ursprung für die Röntgenaufnahmetechnik wurde bereits viel früher gelegt und lässt
sich nach REISNER (1996), SCHUBERT (1998) und LINDGREN (2001) in drei Abschnitte
gliedern:
1. Nach Entdeckung der X-Strahlen durch Wilhelm Conrad Röntgen (1845-
1923) am 8.11.1895 (Nobelpreis 1901) Entwicklung der
Summationsaufnahmetechnik einschließlich Spezialprojektionen (ca. 1895-
1930):
Einführung der Röntgenröhren nach dem Glühkatodenprinzip
(Coolidge-Röhre) und des Streustrahlenrasters (Bucky-Blende)
(1913-1914),
2. Entwicklungsphase der Schnittbildtechnik beginnend mit der
konventionellen Tomographie (1921-1970):
Erster „Planigraph“ nach theoretischen Vorarbeiten von
v.a. Bocage (1921) und eigentlicher Bau durch B. Ziedses des
Plantes (1931),
1930 Einführung der Drehanodenröhre durch A. Bouwers und
Einführung der Tomographie durch A. Vallebona,
1936 Einführung der Schirmbildmethode durch M. de Abreu,
Serienproduktion des Hochleistungsschichtgerätes "Polytom"
(mit hypozykloider Verwischung) durch Konstrukteur Massiot
(nach 1950),
1952 Einführung der Bildverstärker-Technik,
3. Entwicklung der Computer-Tomographie:

Methodenübersicht
43
ab 1957 Entwicklung der Computer-Tomographie durch
A. MacLeod/Cormack und
ab 1967 Einführung der Computer-Tomographie durch Godfrey
N. Hounsfield.
Neben den oben genannten Verfahren wird die Positronen-Emissions-
Tomographie als weiteres bildgebendes Verfahren vor allem für
Stoffwechseluntersuchungen genutzt (u.a. HSU et al., 1996, MÜLLER et al., 1997,
SCHWAIGER, 1997, SELBERG und BURCHERT, 1997, MÜLLER-GÄRTNER, 1998).
Zusätzlich zu den radiologischen Verfahren Magnetresonanz-Tomographie und
Computer-Tomographie wird besonders beim Schwein bereits über einen langen
Zeitraum mit Ultraschalltechniken (US) gearbeitet (KLIESCH u.a., 1957, STOUFFER et al.,
1961, 1991, HORST, 1964a, HORST u.a., 1971, RITTLER u.a., 1964, SZABO et al, 1999).
Jüngere Übersichtsreferate zur Nutzung von Ultraschalltechniken finden sich bei
FOSTER und FOWLER (1988), HORN (1991), WELLS (1991), MAYRHOFER u.a. (1995),
STREITZ (1995), TREITL (1998) sowie MITCHELL und SCHOLZ (2001). In der
Entwicklung befinden sich neben bildgebenden Verfahren, die intermediäre
Frequenzbereiche der elektromagnetischen Strahlung wie Mikrowellen- und Infrarot-
Strahlung (Abb. 2.2.9) nutzen, die Elektrische-Impedanz-Tomographie (WELLS, 1991,
FOGELHOLM und VAN MARKEN LICHTENBELT, 1997, LI, 2000) sowie auf der
Neutronenaktivierung beruhende Bildgebungsverfahren. Viele Isotope werden durch
Neutroneneinfang radioaktiv und zerfallen danach unverwechselbar unter Aussendung
individueller Gammastrahlung. Aus der Aktivitätsmessung kann äußerst empfindlich
der gleichzeitige Nachweis von mehreren (Spuren-)elementen in einer Probe geführt
werden. Bei vielen chemischen Elementen liegt die Nachweisgrenze sehr niedrig, so
dass die Neutronenaktivierungsanalyse als „Goldstandard“ zur Kalibrierung anderer In-
vivo-Methoden herangezogen wird (KEHAYIAS und VALTUEÑA, 1999).

Methodenübersicht
44
Abb. 2.2.9: Frequenzbereiche für elektromagnetische Strahlung (modifiziert nachSCHUBERT, 1998)
In der visuellen Abbildung des Körperinneren ähneln sich die beiden
dreidimensionalen Verfahren (Röntgen-)Computer-Tomographie und Magnetresonanz-
Tomographie außerordentlich, obwohl sie auf stark abweichenden Prinzipien beruhen
(WELLS, 1991, VANGEN und JOPSON, 1996). Die Magnetresonanz-Tomographie (MRT)
besitzt im Vergleich zur Computer-Tomographie (CT) Vorteile in der strukturellen
Abbildung weicher Gewebe (KALLWEIT et al., 1994), abgesehen von der nicht
vorhandenen Belastung durch Röntgenstrahlen (THOMAS et al., 1998, ROSS et al., 2000).
Nicht nur begrenzt auf Ganzkörperanwendungen in vivo (bzw. post mortem) können
diese beiden Verfahren neben klinischen (THIET und BAULAIN, 1992, KRESKEN u.a.,
1993, JANZEN und GAREAU, 1993, THIELEBEIN, 1994, MAYRHOFER u.a., 1995),
(umwelt-)toxikologischen (BLACKBAND und STOSKOPF, 1990, GASSNER et al., 1990),
anatomischen (DAVIES et al., 1987, JANZEN et al., 1989, JOPSON, 1995, MASON et al.,
1995, SCHULTE SPECHTEL u.a., 1997, WOLTER, 1997, BÖTTCHER et al., 1999) und
embryologischen Zwecken (GASSNER und LOHMAN, 1987, LIRETTE et al., 1993a)
speziell für genetische (LIU et al., 1992, 1994, SCHOLZ et al., 1995a), physiologische
(SCHOLZ et al., 1995b, 1998) sowie ernährungswissenschaftliche (MITCHELL et al.,
1991b,c, LUITING et al., 1992, KOLSTAD, 1996, KOLSTAD et al., 1996) Studien genutzt
werden. Parallel können kurzfristige oder langfristige Veränderungen in der
Körperzusammensetzung sowie in der Zusammensetzung von Gewebedepots während
des Wachstums oder zum Zeitpunkt der Vermarktung (BAULAIN, 1994a,b, BAULAIN
u.a., 1990, 1993, 1996, 1998, STREITZ, 1995, STREITZ u.a., 1995, WIEDERHOLD, 1996,
Elektromagnetische StrahlungFrequenz (Hz)
Ionisierende Strahlung (CT/ DXA)
Nichtionisierende Strahlung
(MRT/ MRS)
Röntgen-Strahlung
UV-sichtbares
Infrarot-LichtMikrowellen-Strahlung
Radio-Frequenz-Strahlung
1018
1016
1014
1012
1010
108
106

Methodenübersicht
45
KASTELIC et al., 1996, KASTELIC, 1997, KOLSTAD, 2000) auf individueller Basis
quantifiziert werden. Eine Stärke der Verfahren liegt in der Aufhebung der Variation
zwischen Tieren, die bei Stufenschlachtungen zu berücksichtigen wäre (WELLS, 1991,
BAULAIN u.a., 1996, KASTELIC, 1997), so dass sie als Referenzverfahren sehr gut
geeignet sind (BAULAIN und HENNING, 2001). Computer- und Magnetresonanz-
Tomographie sowie zum Teil Magnetresonanz-Spektroskopie werden zur Bestimmung
der Körperzusammensetzung für ein weites Spektrum von Nutztierarten eingesetzt.
In vivo (und post mortem) wurden überwiegend Schwein und Schaf untersucht,
beginnend mit den CT- bzw. MRT-Studien von SKJERVOLD et al. (1981), ALLEN und
VANGEN (1983), ETTINGER et al. (1983), GROENEVELD et al. (1983), GROENEVELD
(1985), FULLER at al. (1983) sowie SEHESTED (1983, 1986). Nach Implementierung von
CT- bzw. MRT(S)-Geräten in Australien, Deutschland, Kanada, Neuseeland, Norwegen,
Österreich, Schottland (UK), Ungarn und USA für den Nutztierbereich folgte eine Reihe
weiterer Untersuchungen zur Körperzusammensetzung an verschiedenen
Nutztierspezies gekoppelt mit den verschiedensten Fragestellungen beim Schwein
(BAULAIN, 1994a, b, BAULAIN et al., 1990, 1993, 1996, FOWLER et al., 1992, GRIEP,
1991, HENNING et al., 1991, HENNING, 1992, HORN et al., 1996, 1997, MITCHELL et al.
1991b,c,d, 1993, SCHOLZ u.a., 1993,1995a,b, 1998, KOLSTAD, 1996, KOLSTAD et al.,
1996, 2000, KOLSTAD und VANGEN, 1996, KASTELIC, 1997, SZABO et al., 1998,
VANGEN und ENTING, 1990, VANGEN, 1992, VILLÉ et al., 1991, 1997), Schaf (PÁSZTHY
et al., 1991, AFONSO, 1992, AFONSO und THOMPSON, 1996, STANDAL, 1992, VANGEN
und THOMPSON, 1992, STREITZ, 1995, STREITZ u.a., 1995, BAULAIN und STREITZ, 1997),
bei der Ziege (MASON et al., 1995), beim Geflügel (BENTSEN und SEHESTED, 1989,
MCBRIDE et al., 1991, MITCHELL et al., 1991a, LIU et al., 1992, 1994, LIRETTE et al.,
1993a,b, KALLWEIT u.a., 1994, ROMVÁRI et al., 1994, WIEDERHOLD, 1996), Kaninchen
(SZENDRÕ et al. 1992, ROMVÁRI et al., 1996, 1998) und Fisch (GJERDE, 1987a). Ein
Großteil der MR-Arbeiten zum Wachstum und der Körperzusammensetzung beim
Schwein wurde bzw. wird, neben Schaf und Geflügel, am Institut für Tierzucht und
Tierverhalten der Bundesforschungsanstalt für Landwirtschaft in Mariensee (Neustadt a.
Rübenberge) durchgeführt (BAULAIN u.a., 1990, SMIDT u.a. 1990, 1991, SMIDT, 1992,
KALLWEIT u.a., 1991, 1994, KALLWEIT, 1994, BAULAIN und HENNING, 2001).
Eine Zusammenfassung der ersten CT-Studien in vivo an Schweinen, Schafen,
laktierenden Ziegen und Broilern sowie post mortem an Regenbogenforellen und

Methodenübersicht
46
Schweineschlachtkörpern findet sich bei VANGEN (1988a, b). HORN (1991) vergleicht in
einem Übersichtsbeitrag Ultraschall und CT und gelangt zu dem Schluss, dass CT die
zwei- bis dreifache Genauigkeit zur Schätzung der Körperzusammensetzung gegenüber
Ultraschall erreicht.
Tab. 2.2.2: Bewertung „radiologischer“ Methoden zur Bestimmung der Körper-
zusammensetzung (modifiziert nach PLOURDE, 1997 und MITCHELL und SCHOLZ, 2001)
Methode Spezifität für die Bestimmungder Körperzusammensetzung
Wieder-hol-
barkeit
(%)
Messdauer
(Minuten)
Strahlen-dosis
(mrem)
Kosten Verfügbar-keit im
Veterinär-und
Nutztier-bereich
CT
QCT
Gewebeniveau,abdominales Fettgewebe undSkelettmuskelfettgehaltKnochen
> 98
> 85
10Sekunden
15 - 20
200
9
hoch ++
°
MRT Atomares Niveau,abdominales Fettgewebe undSkelettmuskelfaserzusammen-setzung
85 - 90 1 – 30 - sehrhoch
+
MRS Atomares und molekularesNiveau sowielokaler Fett- und Wassergehalt
>95 1 - 10 - sehrhoch
°
DXA Gewebeniveau,Knochenmineralgehalt und –dichte,(abdominales) Fettgewebe undMagergewebe
>88 10 – 30 0,5 moderat–hoch
+
PAA Atomares und molekularesNiveau,Gesamtkörper-C- , -N und -O-Gehalt sowieSchätzung desGesamtkörperfettgehaltes
96 45 63 cGy hoch -
US Gewebeniveau sowielokales Fett- und Muskelgewebe
75 – 80 1 - 10 - gering –moderat
+++
NAA Atomares und molekularesNiveau,Gesamtkörper-C-, N-, O-, Ca-,Gehalt sowieSchätzung desGesamtkörperfettgehaltes
97 20 0,2 hoch -sehrhoch
-
(Q)CT=(Quantitative) Röntgen-Computer-Tomographie, MRT=Magnetresonanz-Tomographie,MRS=Magnetresonanz-Spektroskopie, DXA= Dualenergie-Röntgenabsorptiometrie,PAA=Photonenaktivierungsanalyse, US=Ultraschall, NAA=Neutronenaktivierungs-Analyse (darunter Neutronen-Inelastische-Streuung, K40-Ganzkörperzählung)
Speziell in der Fleischschafzucht in Schottland, Neuseeland und neuerdings in
Österreich (http://www.schafe-ooe.at/, Stand Dezember 2001) werden verstärkt
Anstrengungen unternommen, die mittels CT bestimmte Körperzusammensetzung
direkt in die Selektion auf Nukleusebene einzubeziehen (SIMM, 1994, THOMPSON et al,

Methodenübersicht
47
1996, STANFORD et al., 1998a), während in Norwegen ein Teil der
Eigenleistungsprüfung für potentielle Zuchteber bereits seit längerer Zeit mittels CT
durchgeführt wird (BAULAIN, 1994a).
Weitere Verfahren, die zur Bestimmung der Körperzusammensetzung (in vivo)
genutzt werden, umfassen die Unterwasservolumenverdrängung (überwiegend im
Humanbereich, BUTTE et al., 1997, ELOWSSON et al., 1998), Bioimpedanz (SWANTEK et
al., 1992), Gesamtkörperwiderstandsmessung [TOBEC] (KEIM et al., 1988, HENNING et
al., 1992, FORREST, 1994, BUTTE et al., 1997, HIGBIE, 1998), Verdünnungsanalysen
(FULLER et al., 1971, HOUSEMAN et al., 1973, BUTTE et al., 1997, TREITL, 1996, 1998)
sowie Hautfaltendicke-Messungen (BUTTE et al., 1997). Größere Nutztiere, die zur
Fleischproduktion verwendet werden (z.B. Rinder), sind aufgrund der begrenzten
Geräteausmaße sowie der anatomischen Gegebenheiten für In-vivo-Untersuchungen
mittels MRT, CT, DXA oder TOBEC nur bis zu einer maximalen Körpermasse von ca.
120 kg – 130 kg geeignet (ALLEN und WALSTRA, 1996, SCHOLZ u.a., 2001 - siehe Abb.
2.2.20). Referenzuntersuchungen an Schlachtkörperteilstücken vom Rind sind jedoch
ohne weiteres durchführbar (BAULAIN u.a., 1991, ALLEN und KEANE, 1996, MITCHELL
et al., 1997b, HOLLÓ et al., 1998).
Tabelle 2.2.3: Vor- und Nachteile gebräuchlicher Verfahren zur nicht invasiven
Bildgebung für die Untersuchung der Körperzusammensetzung
Vorteile Nachteile
Ultraschall - leicht verfügbar
- keine Strahlenbelastung
- kostengünstig
- Darstellung von B-Bild in
Echtzeit
- im Vergleich zu CT und MRT
weniger genaue anatomische
Auflösung
Videobildanalyse - leicht verfügbar
- keine Strahlenbelastung
- kostengünstig
- nur Information über Körperform
und Körperproportionen am
lebenden Tier
Computer-Tomographie - sehr gute anatomische
Auflösung
- Strahlenbelastung
- relativ hohe Investitionskosten
Magnetresonanz-
Tomographie
- sehr gute
Gewebedifferenzierung
- multiplanare Darstellung
- hoher technischer Aufwand
- hohe Investitions- und laufende
Kosten
- eingeschränkte Verfügbarkeit

Methodenübersicht
48
Stellt man ergänzend zu Tab. 2.2.2 die für die Ermittlung der
Körperzusammensetzung gebräuchlichsten bildgebenden Verfahren unter Einbeziehung
der noch nicht erwähnten Videobildanalyse gegenüber, ergeben sich die in Tab. 2.2.3
zusammengestellten Vor- und Nachteile.
2.2.3.2.5.1 Computer-Tomographie (CT)
Im Zeitraum 1967 - 1971 ging mit der Einführung der Röntgen-Computer-Tomographie
(CT) ein lang gehegter Wunsch der Human- und Tierwissenschaftler in Erfüllung.
Erstmals konnten computergestützt die räumlichen Verhältnisse im Körperinneren
(Gehirn) bildlich auf nicht invasive Weise dargestellt werden (HOUNSFIELD, 1973).
RADEN (1917) legte mit seiner theoretischen Vorarbeit den Grundstein zur von
CORMACK (1980) weitergeführten Entwicklung der mathematischen Algorithmen für die
Bildrekonstruktion aus den Daten der Röntgenschwächung. Auf diesen Grundlagen baut
eine Vielzahl von Forschern auf, um die Röntgentechnik selbst und deren
mathematische Umsetzung in dreidimensionale Bildinformationen stetig zu verbessern
(z.B. SCHALLER et al., 1996a, b).
Generell liefert CT oder CAT (Computer-Axial-Tomographie) Schnittbilder des
Körperinneren, die den Bildern aus der Magnetresonanz-Tomographie (Kapitel
2.2.3.2.5.2) sehr ähneln. Die CT-Bilder verfügen zwar über einen guten Kontrast
zwischen Fett- und Magergewebe (speziell zwischen Weichgewebe und Knochen) sind
aber im Vergleich zur Magnetresonanz-Tomographie im detaillierten Kontrast zwischen
oder innerhalb verschiedener Gewebe und Organe der Magergewebekategorie
eingeschränkt. Wie der Name sagt, ist ein CT-Bild ein Querschnittsbild (Tomogramm),
das mit Hilfe eines Rechners erstellt wird.
Ein CT-Bild entsteht vereinfacht wie folgt:
Senkrecht zur Körperachse des Probanden (Kopf-Fuß-Richtung) rotiert ein festes
System aus Röntgenröhre und ihr gegenüber liegenden Detektoren. Der Proband wird in
Einzelschritten durch das Gerät geschoben. Die Röntgenröhre erzeugt dabei mit einer
Hochspannung von 120 bis 150 kV (1 kV = 1000 Volt) einen punktförmigen oder
zumeist fächerförmigen Röntgenstrahl, der den Körper in der gewünschten Ebene
durchstrahlt. In Abhängigkeit von der Dichte, der Dicke sowie der Ordnungszahl der
durchstrahlten Materie, also des Gewebes, wird der Strahl mehr oder weniger stark
geschwächt. Gegenüber der Röhre befindet sich halbkreisförmig angeordnet eine Anzahl
(z.B. 500 bis 1000) von Detektoren, die in Abhängigkeit von der auftreffenden

Methodenübersicht
49
Röntgenstrahlenintensität elektrische Signale erzeugen. Diese Signale werden weiter
verarbeitet und zur endgültigen Bilderzeugung verwendet (LEYMASTER, 1986).
Die Röhre mit den gegenüberliegenden Detektoren dreht sich schrittweise z.B. um
1° oder 0,5° weiter und sendet erneut einen Fächerstrahl aus, dessen Signal dem
Rechner zugeführt wird. Dies geschieht z.B. 360- oder 720-mal. Auf diese Weise
werden verschiedene Ansichten (Projektionen) derselben Schicht erzeugt und die für
jedes Bildelement (Pixel) gemessene Röntgenschwächung wird im Computer zu einem
Graustufen-Bild umgerechnet. Die Anzahl der Bildelemente ist geräte- und
softwareseitig einzustellen und ergab bei konventionellen CT-Geräten z.B. eine
Bildmatrix von 256 x 256 Bildelementen (BAULAIN, 1994a). „Spiral“-CAT-Scanner der
jüngsten Generation mit Bildmatrizen von ≥ 1024 x 1024 Bildpunkten lassen eine viel
höhere Auflösung zu (HORN et al., 1997). Es entsteht ein in seiner Dicke einstellbares
Querschnittsbild des Körpers. Die Schichtdicke kann für den jeweiligen
Untersuchungszweck zwischen etwa 0,5 mm und > 10 mm vorgewählt werden.
Einzelheiten der Organe bzw. Gewebe sind mit geringsten Schichtdicken am genausten
zu erkennen. Neueste Mehrzeilen-Spiral-CT-Geräte verfügen über > 8 Detektorein-
heiten, die gleichzeitig um den Probanden kreisen und Scheibendicken von > 2 x 10 mm
gewährleisten. Die Standardumdrehungszeit beträgt 0,75 Sekunden und kann bis auf 0,5
Sekunden reduziert oder bis auf 2 Sekunden erhöht werden. Aus einer variierenden
Scanlänge von 16 bis 1440 mm resultieren Bildzahlen von 41 bis 700.
Die Strahlenexposition beträgt, je nach Untersuchung, zwischen 0,5 cSv bis 1 cSv
(1 cSv = 1/100 Sievert) für den zu untersuchenden Bereich. Wie konventionelle
Röntgenstrahler lässt sich die Computer-Tomographie im Prinzip auf alle Körperteile
anwenden.
Diese Technik - wie auch die Dualenergie-Röntgenabsorptiometrie - nutzt die
unterschiedliche Schwächung der Röntgenstrahlen in differenzierten Körpergeweben
wie Muskel (Wasser und Protein), Fett oder Knochen, die durch die verschiedenen
(angenommenen) spezifischen Dichten von 0,993 g/cm3 für Wasser, 1,34 g/cm3 für
Protein, 0,9 g/cm3 für Fett und 3,075 g/cm3 für (Knochen-) Mineralstoffe bei 37 °C
verursacht werden (SUTCLIFFE, 1996). Die linearen Schwächungskoeffizienten werden
in sogenannte Hounsfield-Einheiten (HU) oder CT-Werte transformiert, die zwischen
+1023 für Knochen (bzw. theoretisch +3000 bei kompletter Absorption von extrem
dichten Materialien) und –1024 (keine Absorption) für Luft schwanken. Protein liegt bei

Methodenübersicht
50
349 HU, Wasser bei 7,5 HU, und Lipid bei –191 HU. Fettgewebe weist Werte zwischen
–30 und –190 HU auf, während Magerweichgewebe (hauptsächlich Muskel) zwischen –
29 und 150 HU klassifiziert werden kann (BAULAIN, 1994a, LUITING et al., 1995).
Ein „quantitatives“ CT-Gerät (QCT) liefert nicht nur Bilder, sondern wie
beschrieben quantitative Messwerte auf der Hounsfield-Skala. Solche digitalen CT-
Daten lassen sich ähnlich wie bei der Dualenergie-Röntgenabsorptiometrie (Kapitel
2.2.3.2.5.4) zum Beispiel bei der Überwachung der Osteoporose nutzen. Osteoporose
verringert den Kalziumgehalt und damit die Röntgenabsorption im Knochengewebe.
Deshalb weisen z.B. die Wirbelsäule oder Oberschenkelknochen erkrankter Probanden
niedrigere Werte auf als die Knochen gesunder Probanden. Verschiedene
Einflussfaktoren auf Knochenmineraldichte und Knochenmasse sind in Abb. 2.2.23
zusammengefasst.
Die ermittelten Fett- und Muskelgewebeverteilungen können sich jedoch im
Bereich von 0 HU überschneiden, obwohl Fett (Lipid) theoretisch nur im negativen HU-
Bereich und Muskel (Protein + Wasser) nur im positiven HU-Bereich liegen kann
(VANGEN, 1992). HORN et al. (1997) definierten „Dichtebereiche“ für Fett zwischen –10
und –150 HU, für Wasser zwischen -10 und +10 HU, für Muskelgewebe zwischen +10
und +150 HU und für Knochen zwischen +250 und +1000 HU. Basierend auf dieser
Einteilung quantifizieren einerseits verschiedene Computer-Programme mit
halbautomatischer oder automatischer Gewebeklassifizierung innerhalb der digitalen
CT-Bilder unabhängig von Regressionsgleichungen die Gewichte (Volumina) der
interessierenden Körperkomponenten wie subkutane Fettauflage, inter-/intramuskuläres
Fett, Gesamtkörperfett, Abdominalfett, Magergewebe, fettfreie Innereien, Knochen und
Gesamtgewebe (THOMPSON und KINGHORN, 1992, JOPSON et al., 1995). Andererseits
sind Regressionsgleichungen unter Verwendung von Referenzwerten aus der
chemischen Analyse oder der Schlachtkörperzerlegung als unabhängige Variablen und
der CT-Daten als abhängige Variablen weiterhin die Methode der Wahl zur
Genauigkeitsüberprüfung der neuen Messtechniken. Zusätzlich zu Bestimmtheitsmaß
(R2) und Reststandardabweichung (RSD) als Präzisionsparameter kann das prozentuale
Verhältnis zwischen Standardfehler der Vorhersage (SEP) und Standardabweichung der
abhängigen Variable (SD) [SEP/SD = RSD] als Genauigkeitsparameter dienen.
Messungen der Fettmenge (kg) von lebenden Schweinen ergaben RSD-Werte von 18,6
% in Jungsauen und 27,1 % in Jungebern, während die Schätzung der Proteinmenge

Methodenübersicht
51
(kg) etwas höhere Schätzfehler mit 31,8 % bei Jungsauen und 27,1 % bei Jungebern
ergab (VANGEN, 1988a,b, BAULAIN, 1994a). In der ersten CT-Studie an 23 Schweinen in
vivo veröffentlichten SKJERVOLD et al. (1981) R2-Werte von 0,89 für den Fettanteil (%)
im Schlachtkörper, 0,83 für den Proteinanteil (%) und 0,85 für den Energiegehalt (%).
Eine etwas geringere Genauigkeit resultierte aus den Messungen der „mageren“ Linie
mit R2 = 0,73 für den Energiegehalt (%) im Schlachtkörper. VANGEN und ENTING
(1990) studierten das Potential der Computer-Tomographie, um den intramuskulären
Fettgehalt in Schweinen mit unterschiedlichen Duroc-Genanteilen (0 %, 25 % und 50
%) zu ermitteln. Zu diesem Zeitpunkt indizierten ihre Ergebnisse eine niedrige
Genauigkeit zur Schätzung des intramuskulären Fettgehaltes mit einem
Bestimmtheitsmaß von R2 ≤ 0,33 und einer Reststandardabweichung (RSD) von ≥ 0,79
(n = 823). JOPSON et al. (1995) ermittelten mittels CT post mortem ein R2 von 0,93 und
eine RSD von 0,32 für die Fettmasse des Schlachtkörpers (kg), wobei sich die Referenz-
Fettmasse aus einer Kombination von Nahinfrarot-Spektroskopie und Zerlegung ergab.
Im Vergleich zeigte die Schlachtkörper-Magermasse, die mittels CT systematisch um
durchschnittlich 3,44 kg überschätzt wurde, ein R2 von 0,86 und ein RSD von 0,63. Die
Unterschiede zwischen diesen Techniken hängen von verschiedenen Faktoren ab. Die
Genauigkeit der Zerlegung, z.B. - die nicht immer zu gewährleistende - exakte
Trennung von Mager- bzw. Muskelgewebe von Knochen- und Fettgewebe, ein exakter
CT-Schwellwert zur Unterscheidung zwischen Muskel- und Knochengewebe (JOPSON et
al., 1995), die Güte der Beziehungen zwischen Gewebedichte und Hounsfield-Einheiten
speziell bei Geweben mit sehr hoher Dichte und die variable Definition der mittels CT,
Zerlegung oder Nahinfrarot-Spektroskopie gemessenen Gewebe beeinflussen in
erheblichem Umfang die geschätzte Genauigkeit und Präzision der CT-Messung. SZABO
et al. (1998) fanden mit maximalen R2-Werten von 0,51 bzw. 0,8 niedrigere
Beziehungen zwischen Magergewebe- bzw. Fettgewebevolumen aus der CT in vivo und
dem Protein- bzw. Fettgehalt aus der chemischen Analyse von 20
Schweineschlachtkörpern (10 Jungsauen und 10 männliche Kastraten) bei 105 kg
Lebendgewicht. Allein die Schlachtkörpermasse kombiniert mit dem Effekt des
Geschlechts der Tiere erklärte 44 % bzw. 37 % der Variation im Protein- bzw.
Fettgehalt.
ALLEN und LEYMASTER (1985) studierten die grundlegenden Fehlerquellen, die
sich aus der Technik der Röntgen-Computer-Tomographie ergeben. Das niedrigste

Methodenübersicht
52
„Rauschniveau“ bzw. die beste zwei- oder dreidimensionale Bildqualität resultiert aus
der Kombination einer höheren Spannung (z.B. 150 kV) mit einer größeren
Scheibendicke (z.B. 8 mm) und der höchsten Stromstärke bei der höchsten Anzahl von
Projektionen.
Während Hardware und Software für die Röntgen-Computer-Tomographie - CT
(oder Computer-Axial-Tomographie – CAT) ständige Verbesserungen erfahren, nimmt
der sogenannte „Spiral-CAT-Scanner“ die jüngste Stufe der Entwicklung in der
Röntgen-Tomographie ein. Bei der Spiral-Computer-Tomographie kreist eine
leistungsfähige, sehr hitzestabile Röntgenröhre kontinuierlich um das Objekt (Tier),
welches auf dem Untersuchungstisch mit einer bestimmten Geschwindigkeit durch die
Öffnung des Gerätes geschoben wird. Durch das Körpergewebe wird der von der Röhre
ausgesandte Röntgenstrahl (Photonen) - wie bereits beschrieben - geschwächt. Eine
Messkammer mit Röntgensensoren, die gegenüber der Strahlenquelle angebracht ist,
misst die Intensität der auf sie treffenden Strahlen. Die so gewonnenen Ergebnisse
werden kontinuierlich aufgezeichnet und vom Computer zu einer Vielzahl
hochauflösender Schnittbilder des Körpers umgerechnet (HORN et al., 1997). Aus diesen
Datensätzen können wiederum dreidimensionale Abbildungen des Körpers bzw. von
Körperregionen errechnet werden, die eine volumetrische Bildanalyse gestatten. In vivo
ergaben sich hohe Schätzgenauigkeiten: für die Schinken-Muskelmasse ein R2–Wert
von 0,92 und ein RSD (SEE/SD) von 0,30; für die Schinken- plus Knochenmasse ein
R2-Wert von 0,91 sowie ein RSD von 0,33; für die Masse des M. longissimus dorsi ein
R2-Wert von 0,96 sowie ein RSD von 0,19; für die Masse des M. longissimus dorsi plus
Knochen ein R2 -Wert von 0,91 sowie ein RSD von 0,27; für die Gesamtmuskelmasse
(M. longissimus dorsi + Psoas major + Schinken + „Bauchseitenfleisch“ [bacon side
meat]) ein R2-Wert von 0,9 sowie ein RSD von 0,34 und außerdem für das gesamte
subkutane Fett plus Haut an der Schlachtkörpermasse ein R2 -Wert von 0,92 sowie ein
RSD von 0,34. Etwas höhere Genauigkeiten (R2 und RSD) können aus (Spiral-)CAT
oder CT Messungen post mortem erwartet werden (z.B. SEHESTED und VANGEN, 1988,
JOPSON et al., 1995, HORN et al., 1997).
Zwischen volumetrischen Spiral-CAT-Messungen und den traditionellen
(planaren, zweidimensionalen) CAT-Bildparametern bestehen enge Beziehungen. HORN
et al. (1997) ermittelte R2 -Werte zwischen 0,93 und 0,99 für Fettgewebedistanz- und
-flächenmaße an verschiedenen Scan-Positionen (9., 11., 13. Rippenwirbel, 2., 4., 6.

Methodenübersicht
Allgemein hat die volumetrische Spiral-CAT-Bildgebung – trotz des höheren
Preises - eine Reihe von Vorteilen gegenüber der traditionellen (planaren) CT für
Messungen der Körperzusammensetzung in vivo:
1. Eliminierung von Bewegungsartefakten und niedrige Strahlungsbelastung durch
sehr kurze Scanzeiten (120 Sekunden für 110 Scans mit einer Scheibendicke von 10
mm),
2. (fast) keine Einschränkungen für 2D und dreidimensionale Rekonstruktionen mit
einer hohen Auflösung von (bis zu) 1024 x 1024 Bildpunkten/Scheibe im Vergleich
zur konventionellen 256 x 256 Bildmatrix (BAULAIN, 1994a),
3. Weichteile sind besser voneinander abzugrenzen und abzubilden sowie
4. Überlagerungsfreie Darstellung einzelner Körperabschnitte, einschließlich
Dichtebestimmung anatomischer Strukturen.
Abb. 2.2.10: Funktionsweise der Spiral-Röntgen-Computer-Tomographie links: Computertomograph mit „virtuellem“ Schwein, rechts: Funktion (modifiziert nach http://www.tilak.at/azwhome/rtakademie/ct.html; KNOFLACH et al., 2000) 2.2.3.2.5.2 1H-Magnetresonanz-Tomographie
Das Phänomen der Magnetresonanz (MR) ist erst seit 1946 (BLOCH et al., 1946 und
PURCELL et al., 1946) bekannt und wurde als 2D-Spektroskopie zur Analyse
53
Lendenwirbel und Femurkopf einschließlich "Bacon-Seite") und R²-Werte zwischen
0,87 und 0,97 für Muskelgewebedistanz- und flächenmaße an denselben anatomischen
Positionen.

Methodenübersicht
54
biologischer Makromoleküle erstmals von JEAN JEENER im Jahre 1971 verwendet
(HAUSSER und KALBITZER, 1989). LAUTERBUR (1973) nutzte die 1H-Magnetresonanz
erstmals zur Abbildung eines flüssigkeitsgefüllten Phantoms. Seit etwa 1980 wird das
Verfahren in der Medizin eingesetzt (LÜNING u.a., 1988, HARNAK, 1996-2000, REISNER,
1996).
Eine wesentliche Aufgabe der Tierzuchtforschung liegt in der Entwicklung und
Bereitstellung von Verfahren, die die Körperzusammensetzung von lebenden und
geschlachteten Tieren mit hoher Genauigkeit erfassen (BAULAIN und HENNING, 2001).
Die für die Humanmedizin entwickelte Magnetresonanz-Tomographie ist wie die
Röntgen-Computer-Tomographie ein Verfahren zur bildlichen Darstellung des
Körperinneren, das bei Anwendung geeigneter Bildauswertungsmethoden die Schätzung
der Gewebeanteile (Fleisch, Fett, Knochen) lebender Tiere und Schlachtkörper
ermöglicht. Für die Nutztierwissenschaften in Deutschland wird die MR-Tomographie
seit Ende 1987 im Institut für Tierzucht und Tierverhalten (FAL), Mariensee – Neustadt
am Rübenberge, genutzt (BAULAIN u.a., 1990, SMIDT u.a., 1990, 1991, BAULAIN und
HENNING, 2001). Zahlreiche Ergebnisse hinsichtlich der Schätzung der grobgeweblichen
Zusammensetzung von Schlachtkörperteilstücken, Schlachtkörpern sowie der
Gewebezusammensetzung in vivo bei den Tierarten Schwein, Schaf und Wassergeflügel
zeigten die Eignung des Verfahrens für die Nutztierwissenschaften (BAULAIN u.a., 1990,
1991, 1993, 1996, 1998a,b, BAULAIN, 1997, FOWLER et al., 1992, GRIEP, 1991,
HENNING et al., 1996, MITCHELL et al., 1991a,b,c,d, SCHOLZ u.a., 1993, 1995a, STREITZ,
1995, WIEDERHOLD, 1996, BALL et al., 1996, BAULAIN und HENNING, 2001).
Kurzbeschreibungen der Prinzipien der Magnetresonanz-Technik im Rahmen der
Nutztier- bzw. Veterinärliteratur erfolgten in diversen Publikationen (BAULAIN u.a.,
1990, SMIDT u.a., 1990, 1991, KRESKEN u.a., 1993, KALLWEIT et al., 1994, MAYRHOFER
u.a., 1995, BAULAIN, 1997, PIETSCH, 1998), während im humanmedizinischen Bereich
eine nahezu unüberschaubare Anzahl von Publikationen zu diesem Thema existiert. Nur
drei deutschsprachige Referenzen von GALLA (1988), LÜNING u.a. (1988) sowie von
HAUSSER und KALBITZER (1989) seien hier genannt. Eine sehr anschauliche Einführung
in die Magnetresonanz-Tomographie gibt SCHILD (1990).
Nachfolgend werden die Prinzipien nochmals ausführlicher dargestellt:
Materie besteht bekanntlich aus Atomen, die ihrerseits Moleküle bilden können. Das
Atom besteht aus einer Elektronenhülle und dem Atomkern. Das neutrale Atom enthält

Methodenübersicht
55
genauso viele Elektronen in seiner Hülle wie der Kern Protonen besitzt. Zusammen mit
den Protonen bilden die elektrisch neutralen Neutronen den Atomkern. Protonen sind
elektrisch positiv geladen und haben annähernd die gleiche Masse wie die Neutronen.
Jede bewegte Ladung hat ein Magnetfeld zur Folge. Daher erzeugt auch die
Eigenrotation (Spin) der Protonen ein, wenn auch sehr kleines, atomares Magnetfeld.
Aus diesem Magnetfeld resultiert ein magnetisches Moment der Protonen. Diese
Momente können sich gegenseitig verstärken oder, wenn die Protonen gegeneinander
rotieren, auslöschen. Für die Magnetresonanz-Tomographie (-Spektroskopie) werden
Atome mit einem nach außen wirkenden magnetischen Moment (Eigenspin) verwendet
wie 1H, 2H, 7Li, 11B, 13C, 14N, 15N, 17O, 19F, 31P, 23Na, 27Al, 29Si, 35Cl, 39K, 69 Ga, 71Ga,77Se, 87Rb, 109Ag, 129 Xe, 131Xe, 199Hg. Diese Liste lässt sich noch erweitern, da
verfeinerte Messmethoden immer geringere Konzentrationen nachweisen können.
Einzige Voraussetzung ist die notwendige Eigenschaft des „Kernspins“, den nur
Atomkerne mit ungerader Protonen- und Neutronenzahl aufweisen. Grundsätzlich
können alle aufgeführten chemischen Elemente für tomographische und/oder
spektroskopische Untersuchungen mit unterschiedlicher Auflösung verwendet werden.
Die zeitliche und örtliche Auflösung wird von der relativen Empfindlichkeit der Atome
bestimmt, die weitestgehend von den magnetischen Eigenschaften der Atome (gemessen
als sogenanntes gyromagnetisches Verhältnis) und ihrem natürlichen Vorkommen
abhängt (Abb. 2.1.1). Eine gezielte Anreicherung von Atomen mit geringer natürlicher
Häufigkeit führt entsprechend zu einem Anstieg ihrer Empfindlichkeit.
Die Bildgebung (Tomographie) bedarf vereinfacht nur eines zusätzlichen
Rechnerschrittes im Vergleich zur Spektroskopie, für die üblicherweise auch keine
Magnetfeldgradienten verwendet werden. Über eine Fouriertransformation kann aus den
Spektraldaten ein Abbild der entsprechenden Voxel-Signalintensitäten in Form von
sogenannten „Grauwerten“ mit unterschiedlicher Datentiefe (8 Bit - 64 Bit) berechnet
werden. Mit den Geräten und der Software der neuesten Generation lassen sich beide
Verfahren (Tomographie und Spektroskopie) zur „funktionellen Bildgebung“ erweitern
und mehrere Atome (z.B. 1H, 31P und 13C) gleichzeitig in einem Untersuchungsgang
analysieren.
Das Phänomen der Magnetresonanz wird durch die physikalische Eigenschaft der
Atomkerne (Protonen) hervorgerufen, indem sie nach Einbringen in ein stabiles
Magnetfeld mit einem spezifischen Radiofrequenzimpuls zu einer parallelen oder

Methodenübersicht
56
antiparallelen Ausrichtung und Bewegung (Präzession) um die Achse des Magnetfeldes
(B0) gebracht werden können (Abb. 2.2.11). Die Geschwindigkeit, mit der die Protonen
um die Magnetfeldachse rotieren (Präzessionsfrequenz), ist der Magnetfeldstärke direkt
proportional und folgt der Larmor-Beziehung: ωL = γ • B0 (ωL = Larmorfrequenz der
Präzession, γ = gyromagnetisches Verhältnis, B0 = magnetische Feldstärke in Tesla).
Abb. 2.2.11: Kernspin und Präzession (links: parallele Ausrichtung, rechts: antiparalleleAusrichtung), modifiziert nach eigener Grafik in MITCHELL und SCHOLZ, 2001
Abschirmung Magnet
Magnet
Gradienten-Spulen
Gradienten-Spulen
Ganzkörper-RF-Spulen
Ganzkörper-RF-SpulenHub-tisch
Gradienten-Verstärker
Gradienten-Pulsgeber
Hochfrequenz-empfänger
A-D-Wandler
RF-Pulsgeber
RF-Quelle
Radiofre-quenz (RF)-VerstärkerComputer
Daten-speicher
Abb. 2.2.12: Bau- und Funktionsprinzip eines MR-Tomographen(modifiziert nach HORNAK, 1996-2000 und MITCHELL und SCHOLZ, 2001)
In der Regel erzeugen supraleitende Magneten dieses Feld (Abb. 2.2.12). Bei
Raumtemperatur bewirkt dieses Feld einen kleinen „Überschuss“ der Spins der parallel
zur Feldrichtung ausgerichteten Protonen im Vergleich zu den antiparallel
ausgerichteten Protonen. Je stärker das Feld, desto mehr Protonenspins richten sich in

Methodenübersicht
57
dem Feld aus. Für die Bildgebung können jedoch nur die „Überschussprotonen“ genutzt
werden. Durch einen spezifischen („resonanten“) Hochfrequenzimpuls werden die sich
kreiselförmig bewegenden Kerne, den Eigenspin kennzeichnend, aus der Richtung
gekippt und gelangen in ein höheres Energieniveau. Nach Beendigung des
Hochfrequenzimpulses bzw. der -impulsfolge schwingen die Atomkerne in ihre
Ausgangslage zurück. Dabei geben sie die aufgenommene Energie wieder ab, d.h. die
Atomkerne senden selbst Radiowellen aus, die mit Antennen (Hochfrequenzempfängern
– siehe Abb. 2.2.12) empfangen werden können und von einem Computer als
Grauwertstufen in Bildinformationen umgesetzt werden (BAULAIN et al.,1990, SMIDT et
al.,1991).
Die Höhe der gewebespezifisch abgegebenen Energie, die Kern-
Resonanzfrequenz und die Resonanzdauer sind abhängig von der Stärke des
Magnetfeldes und auch von der Art des Atomkerns sowie seiner Molekülumgebung. Die
häufigste Substanz im Tierkörper ist das Wasser (über 70 %). Zur MR-Bildgebung
eignen sich daher die Wasserstoffatome (Protonen) am besten. Protonen im Magnetfeld
weisen Eigenschaften eines schräg rotierenden Magneten auf. Sie besitzen den
beschriebenen „Eigenspin“ und können nur zwei Energieniveaus, entweder das
Basisniveau oder das Anregungsniveau, annehmen (Abb. 2.2.11).
Um den Grauwert (Helligkeit) für jeden Bildpunkt (Pixel) bzw. für jedes
Volumenelement (Voxel) zu erhalten, muss die Signalintensität der entsprechenden
Punkte im Körper bestimmt werden. Neben dem Hauptmagneten ist jedes MR-Gerät mit
zusätzlichen Magnetspulen (sog. Gradientenspulen) ausgestattet, um ein
Magnetfeldgefälle in beliebiger Richtung erzeugen zu können. Durch mehrfach zeitlich
exakt programmiertes Wiederholen von Anregung und Empfangen der Resonanzwellen
(Pulssequenzen) bei jeweils unterschiedlich geschalteten Gradientenspulen erhält man
die erforderliche Information für die Errechnung von Schnittbildern.
Im Unterschied zur Röntgen-Computer-Tomographie (CT) und Dualenergie-
Röntgenabsorptiometrie (DXA), mit denen die unterschiedliche Schwächung der
Röntgenstrahlen in den verschiedenen Körpergeweben gemessen wird, ist die
Messgröße bei der Bilddarstellung durch das MRT-Verfahren die Kernmagnetisierung
in einem gegebenen Volumenelement (Voxel). Bei einer gegebenen Feldstärke (z.B. 1,5
Tesla oder 4,0 Tesla) und einer vorgegebenen Pulssequenz (z.B. ein 90° Puls gefolgt
von einem 180° Puls) ist die Signalintensität (Kernmagnetisierung) abhängig von der

Methodenübersicht
58
Protonendichte sowie von den longitudinalen (Spin-Gitter) bzw. axialen (Spin-Spin)
Relaxationszeiten (T1 bzw. T2 mit T1 > T2), die durch Aufbau und Struktur der
Gewebe (Wasser/Muskel, Fett oder Knochen) beeinflusst werden (LUTZ und SCHULTZ,
1987, LÜNING u.a., 1988, HAUSSER und KALBITZER, 1989). Mit zunehmender
Magnetfeldstärke erhöht sich das Signal-zu-Rausch-Verhältnis. Die Bildqualität
verbessert sich (TRATTNIG u.a., 1997).
Um ein Signal messen zu können, müssen die sehr schwachen Schwingungen
synchronisiert werden. Die Art der Synchronisation (z. B. als sog. Gradientenecho oder
Spinecho) und der Zeitpunkt der Synchronisation, die Zwischenechozeit (TE), sowie die
Zeit bis zur nächsten Anregung, die Repetitionszeit (TR), werden vorgegeben. Da die
Strukturen im Körper unterschiedlich lange „nachschwingen“ bzw. verschieden lange
synchronisiert bleiben, tragen diese Zeitvorgaben maßgeblich zum Bildcharakter bei
(womit eine sogenannte T1- oder T2-Wichtung erzielbar ist). Die Untersuchung muss
dem interessierenden Gewebe angepasst werden. Fettgewebe ist durch kurze T1- und
lange T2-Relaxationszeiten gekennzeichnet, die hohe Signalintensitäten (helle Pixel
oder Voxel) in T1-gewichteten Bildern verursachen. Muskelgewebe besitzt hingegen
unter denselben Vorraussetzungen niedrige Signalintensitäten (dunkle Pixel oder
Voxel). Luftgefüllte Räume werden schwarz abgebildet, da keine Signale empfangen
werden (Abb. 2.2.13).
Untersuchungen an Schlachtkörpern bzw. Schlachtkörperteilstücken von Rind und
Schwein sowie lebenden Schweinen aber auch beim Menschen zeigten, dass T1-
gewichtete MRT-Messsequenzen, beispielsweise sogenannte Multi-Slice-Multi-Echo-
Puls-Sequenzen (z.B. mit einer Pulswiederholzeit - TR von 1000 Millisekunden und
einer Echoausleseverzögerung - TE von 33 Millisekunden) oder auch Spin-Echo-Puls-
Sequenzen (z.B. mit TR = 210 Millisekunden und TE = 17 Millisekunden), besonders
für die Unterscheidung von Muskel- und Fettgewebe geeignet sind (BAULAIN et al.,
1990, 1991, GRIEP, 1991, HENNING et al., 1991, SCHOLZ et al., 1993, SCHOLZ, 1994,
ROSS et al., 2000). Hierbei ist die gleichzeitige Messung mehrerer paralleler Scheiben
sowie Echos (Signalauslesezeiten) möglich. Abbildung 2.2.13 zeigt ein Beispiel von
vier zeitlich aufeinander folgenden „Echos“ eines Thorax-Schnittbildes, welches mittels
einer Multi-Slice-Multi-Echo-Puls-Sequenz erzeugt wurde. Im Rahmen des sogenannten
freien Induktionsabfalls (FID) nach Anregung der Protonen durch die genannte
Pulssequenz nimmt die Signalintensität kontinuierlich von Echo 1 bis Echo 4 ab,

Methodenübersicht
59
während der Kontrast von Echo 1 zu Echo 2 ansteigt und danach bis zum Echo 4
abnimmt (Abb. 2.2.13 und 2.2.14).
Abb. 2.2.13: Darstellung von MR-“Echo“-Bildern, die sich aus einer Multi-Slice-Multi-Echo-Puls-Sequenz ergeben (aus SCHOLZ, 1994; Fett erscheint hell bzw. weiß undMager- bzw. Muskelgewebe grau bis dunkelgrau)
Abb. 2.2.14: Veränderung von Signalintensität und Bildkontrast während der MR-Bilderzeugung
Besonders im Thorax- und Abdominalbereich des Menschen wird im Vergleich zu
1-Fouriertransformations-Puls-Sequenzen, die im Bereich der Gliedmaßen Verwendung
finden, mit 1/2-Fouriertransformations-Puls-Sequenzen gearbeitet, um die Aufnahmezeit
und damit Bewegungsartefakte durch die Atmung zu reduzieren (ROSS et al., 2000).
Sig
nal
Ko
ntr
ast
Fett
Muskel
TE TE
|Fett – Muskel|
Methodenübersicht
60
Abb. 2.2.15: Beispiel für ein MR-Axialschnittbild im Bereich der Nieren(Sichtbare Schatten sind durch Atmungsbewegungen hervorgerufene Artefakte.)
Abb. 2.2.16: Beispiel für ein MR-Axialschnittbild im Beckenbereich
Die Magnetresonanz-Tomographie gehört im Rahmen von Untersuchungen der
Körperzusammensetzung zu den ‘deskriptiven’ Methoden, solange Schätzgleichungen
auf der Basis von Referenzwerten aus der chemischen Analyse oder geweblichen
Zerlegung verwendet werden. Basierend auf den bekannten Konstanten für die
spezifische Dichte von unterschiedlichen Körpergeweben ermöglicht die

Methodenübersicht
61
Magnetresonanz-Tomographie auch direkte Messungen der Gewichte von anatomisch
definierten Körperteilen und -geweben bzw. des Gesamtkörpers, nachdem die
entsprechenden Volumina mittels einer dreidimensionalen Bildanalyse ermittelt wurden.
Inzwischen schließen routinemäßige Anwendungen der Magnetresonanz-
Tomographie die Messung des Gesamtkörpers und regionaler Verteilungen des
subkutanen und abdominalen Fettgewebes, die Quantifizierung des Magergewebes und
speziell der Skelettmuskulatur ein. Neben den routinemäßigen, klinischen
Anwendungen in der Human- und Veterinärmedizin wird die MR-Tomographie als
Methode der Wahl für Kalibrierungen von „Feldmethoden“ bezeichnet, die für die
Messung von Körperfett und Skelettmuskulatur in vivo Verwendung finden sollen
(BAULAIN, 1996, JEBB, 1997, ROSS et al., 2000). Erst seit kurzem existieren Proton- (1H)
und Natrium- (23Na) MRT-Protokolle, die die Qualität (Lipid- und
Natriumkonzentration) des Skelettmuskelgewebes messen können (ROSS et al., 2000).
Hohe Anschaffungs- und Unterhaltskosten beschränken jedoch den Einsatz im Bereich
der Nutztiere (BAULAIN, 1996).
2.2.3.2.5.2.1 Statistische Bildanalyse zur Quantifizierung von Gewebeanteilen
Abb. 2.2.17: Schema für den Ablauf der Bildverarbeitung mittels Clusteranalyse – vomOriginal (links) über interessierende Region (ROI) mit dreidimensionalerSignalintensitätsdarstellung zum Clusterbild (rechts oben nach rechts unten)
Ebenso wie die Röntgen-Computer-Tomographie und die Ultraschall-Bildgebung
ist die Magnetresonanz-Tomographie ein Messsystem, welches zur Quantifizierung von
Komponenten der Körperzusammensetzung auf dem Gewebe-System-Niveau eingesetzt

Methodenübersicht
62
wird und die anatomische Verteilung der Körpergewebe in vivo sowohl beim Tier
(Schwein) (FOSTER und FOWLER, 1988, SMIDT u.a., 1990, 1991, FOWLER et al., 1992,
FULLER et al., 1994, BAULAIN UND SCHOLZ, 1996, SZABO et al., 1999, MITCHELL und
SCHOLZ, 2001) als auch beim Menschen (HEYMSFIELD et al., 1997, ELBERS et al., 1997)
darstellt. Die Differenzen in der Konzentration, der Mobilität und den
Relaxationseigenschaften der Wasserstoffatomkerne in verschiedenen Geweben
verursachen - wie beschrieben - eine Variation der Signalintensitäten (Grauwerte)
innerhalb der MR-Bilder (Scheiben). Verschiedene Verfahren, die für alle bildgebenden
Methoden verwendet werden können (TERADA et al., 1997), dienen der Bildbearbeitung
und der statistischen Analyse der Informationen aus den Signalintensitäten, um
Gewebevolumina bzw. Gewebeflächen quantifizieren zu können (Abb. 2.2.17, 2.2.18).
In der 3D-Abbildung der Signalintensitätsverteilung (Abb. 2.2.17) ist deutlich zu
erkennen, dass Fett höhere Signalintensitäten als Muskelgewebe (hier: Mm.l.d.)
aufweist. In den meisten Fällen beruht der erste Schritt der Bildverarbeitung auf einer
qualifizierten (z.B. auf anatomischem Vorwissen beruhenden) Definition von
sogenannten „Regions of Interest“ (interessierenden Regionen). Diese können manuell,
halbautomatisch oder im besten Fall vollautomatisch unter Zuhilfenahme einer
entsprechenden Bildbearbeitungssoftware festgelegt werden. Die anschließende
Ermittlung der Gewebevolumina kann erfolgen, indem die Anzahl der Bildpunkte, die
entweder Fett, Muskel oder Knochengewebe enthalten, gezählt wird. Aus den
Pixelausmaßen (Höhe x Breite) resultieren entsprechende Flächenmaße bzw. aus der
Kombination von Scheibendicke (Höhe x Breite x Tiefe), Scheibenabstand (möglichst =
0) und Scheibenzahl resultieren die Volumenmaße für die interessierenden Gewebe. Die
Anzahl der Bildpunkte ergibt sich aus der Flächenfestlegung (Bildsegmentierung)
entweder mittels frei definierbarer Linienziehung oder sogenannter „regionaler
Wachstumsverfahren“ bzw. aus Histogramm-, Korrelations- oder/und Clusteranalysen
(Abb. 2.2.18). Die Linienziehung bzw. Flächeneingrenzung (mit einem
Bildschirmkursor) erfolgt in Abhängigkeit von anatomischen Kenntnissen und
Gegebenheiten entweder unter vollständiger Kontrolle des Anwenders oder über eine -
auf Signalintensitätsdifferenzen beruhende - vollautomatische bzw. manuell regulierbare
Schwellwertsetzung (ONG et al., 1997, BEIER u.a., 1998a). Auf diesem Weg kann der
Anwender die exakte anatomische Definition (Eingrenzung) von interessierenden
Geweberegionen steuern. Die computergesteuerten „regionalen Wachstumsverfahren“

Methodenübersicht
63
umgrenzen eine Region, ausgehend von einem Startpunkt innerhalb des interessierenden
Gewebes, bis alle dem entsprechenden Gewebe zuzuordnenden Bildpunkte
eingeschlossen sind. Diese Technik birgt Fehlerquellen, wenn das interessierende
Gewebe an ein Gewebe mit derselben Signalintensitätsausprägung grenzt – z.B. zwei
verschiedene Muskeln, verschiedene Abdominalfettschichten – oder wenn das
interessierende Gewebe über das Bild ungleichmäßig verteilt ist, wie z.B.
intramuskuläres oder intermuskuläres Fett (ELBERS et al, 1997, TANG et al., 2000).
Die Histogrammanalyse kann zu einem aktiven konturverfolgenden Algorithmus
mittels Rauschkorrektur erweitert werden, indem die Signalintensitätshäufigkeiten eines
Bildes (Scheibe) und dynamische Schwellwertintensitäten zur Trennung der Bildpunkte
in unterschiedliche Gewebeklassen herangezogen werden (THOMAS et al., 1998, TANG
et al., 2000). Probleme ergeben sich aus Bildpunkten, die z.B. sowohl Fett- als auch
Muskelgewebe enthalten. Hier kann der Anwender bei einigen
Bildverarbeitungsprogrammen unterstützend eingreifen, indem morphologische
Bildverarbeitungsoperatoren (z.B. Dilation, Erosion) eingesetzt werden, um den
sogenannten Partialvolumeneffekt zu reduzieren (BAULAIN, 1997, BEIER u.a., 1998b).
Dabei ist jedoch zu berücksichtigen, dass sich die Ausmaße der zu segmentierenden
Gewebe verändern können, was zu falschen quantitativen Ergebnissen führt und allein
den Bildkontrast verbessert.
Im Zusammenhang mit einer Histogrammauswertung kann für die Eingrenzung
der großen Anzahl potentieller Regressionsvariablen (Signalintensitätsklassen) zur
Bestimmung der Körperzusammensetzung eine schrittweise Regressionsanalyse
durchgeführt werden. Diese Methodik ist statistisch jedoch nicht sehr robust.
Schätzgleichungen für einen Datensatz sind oftmals nur unter erheblichen
Genauigkeitseinbußen auf einen anderen Datensatz für dieselbe Population und
insbesondere für eine andere Population übertragbar (SZABO et al., 1999).
LUITING et al. (1995) beschreiben für CT-Daten ein weiteres Bildauswertungs-
verfahren unter Nutzung von Grauwerthäufigkeiten. Neben den zwei
„Normalverteilungen“ der Signalintensitäten (Hounsfield-Einheiten) für Fett- und
Muskelgewebe wird eine dritte „Normalverteilung“ für kombinierte Gewebe, die
sogenannte „Mischverteilung“ (Mixture Distribution), berechnet. Diese Information
dient der Kalkulation der Gewebeanteile und schließlich der Gewebevolumina.

Methodenübersicht
64
Eine weitere Methodik zur Auswertung der Grauwertverteilung (Signal-
intensitätsverteilung) innerhalb eines MR-Bildes bietet die Clusteranalyse. Im ersten
Schritt der Bildverarbeitung müssen die interessierenden Regionen (ROI’s) definiert
werden, um den auszuwertenden Datenumfang zu reduzieren und die Körperteile, die
nicht Fett- und Magergewebe enthalten, auszugrenzen. Ein derartig „maskiertes“ Bild
bzw. die über x- und y-Koordinaten örtlich definierten Signalintensitäten durchlaufen
eine „Disjoint-Clusteranalyse“ (mit eineindeutiger Zuordnung der Beobachtungswerte
zu einem einzigen Cluster) unter Verwendung „Euklidischer Distanzmaße“, die aus
einer oder mehreren Variablen berechnet werden. Als mögliche Variablen kommen
spezifische MR-Parameter wie Protonendichte und Relaxationszeiten oder
Signalintensitäten der Pixel, die zu unterschiedlichen Echozeiten gemessen wurden, in
Betracht. Die Beobachtungen werden in Cluster aufgeteilt, so dass jeder Bildpunkt
genau einem Cluster (Gewebeklasse oder Gewebeunterklasse) ohne Überschneidungen
angehört. Die Ergebnisse der Clusteranalyse lassen sich über die örtlich definierten
Bildpunkte (x- und y-Koordinaten) wieder als Bilder darstellen, so dass die Güte der
Clusteranalyse anhand eines Vergleichs von Originalbild und Clusterbild überprüft
werden kann (Abb. 2.2.17). Der Ablauf der Clusteranalyse unter Zuhilfenahme des
SAS-Programmpakets FASTCLUS ist im Anhang IV am Beispiel von digitalen
Videoaufnahmen des Koteletts vom Schwein detailliert dargestellt. Nach der
Multiplikation mit der Pixelgröße können die Clusterflächen (Fett- und Muskelflächen)
als Regressoren in einer multiplen Regressionsanalyse zur Schätzung der
Körperzusammensetzung oder, wenn entsprechende Schätzgleichungen bereits
vorhanden sind, direkt zur Ermittlung der Magergewebe- und Fettgehaltswerte
verwendet werden (SCHOLZ u.a., 1993, BAULAIN, 1997). Eine zusätzliche Multiplikation
mit der Scheibendicke (und Scheibenanzahl) bei MR-Aufnahmen resultiert in einem
Volumenmaß. Liegen keine direkt aufeinander folgenden Schnitte (Scheiben) vor, kann
die Formel von ENGELSON et al. (1999):
MRI-Volumen (L) = 0,001 • Abstand zwischen den Scans (cm) • (B1 + B2)/2,
mit A= Abstand zwischen den Scans (cm) und B1 bzw. B2=Gewebeflächen in
getrennten Scans (cm2) verwendet werden.
In einer anderen Methode, der Texturanalyse, versucht man die Pixel eines Bildes
mittels sogenannter Texturmaße oder „Texture Features“ einer endlichen Anzahl von

Methodenübersicht
65
Kategorien zuzuordnen. Diese Texturmaße sind zumeist statistische Funktionen, die alle
Werte in der Umgebung um einen Bildpunkt (Pixel) berücksichtigen. Beispiele hierfür
sind der Mittelwert oder die Varianz der „Grauwerte“ in dieser Umgebung. Auf diese
Weise lassen sich Aussagen darüber treffen, ob der Bildpunkt zu einer Textur gehört.
Mit Textur bezeichnet man einen kleinen Bildbereich, dessen „Pixel“ eine typische
Mikrostruktur aufweisen. Jedem „Pixel“ können auf diese Weise eine Reihe von
Texturmaßen zugeordnet werden. Diese bilden den Parametervektor, der mittels eines
Klassifikators einer Kategorie zugeordnet wird. Die Leistungsfähigkeit der
Texturanalyse hängt sehr stark von der Wahl geeigneter Texturparameter ab.
Aufwendige Forschungen befassen sich daher mit der Suche nach geeigneten
Texturmaßen für spezifische Anwendungen (KOLB, 1991).
Generell sollte für eine reproduzierbare Verwendung von Schätzgleichungen
möglichst eine Kreuzvalidierung vorgenommen werden. Insbesondere beim Vergleich
von zwei Messmethoden, die jeweils nicht als „Goldstandard“ angesehen werden, ist die
statistische Analyse nach dem Verfahren von BLAND und ALTMANN (1986) zu
empfehlen.
Unabhängig davon sind direkte Messungen der Körperzusammensetzung – wie
Gewebevolumina aus der MRT – genauer als indirekte oder geschätzte Maße (GUO et
al., 1996).
Bland-Altman-Analysen
Um einen Einblick in die Präzision der Quantifizierungen von Messwerten zubekommen, wird vor allem in der humanmedizinischen Forschung die sogenannteBland-Altman-Analyse durchgeführt. Dieses Verfahren dient dem Vergleich zwischenzwei Untersuchungsmethoden, die eine stetig verteilte biologische Variable messen,ohne dass eine der beiden Methoden (z.B. Ultraschall und MRT) als „Goldstandard“gelten kann. Zunehmend wird diese statistische Methodik in den Bereich derVersuchtierforschung Eingang finden, da Tierversuche, bei denen die Probanden amEnde des Versuchs Referenzschlachtungen zugeführt werden müssen, weiter abnehmenwerden. Da der Regressionsquotient R² extrem von der Streuung der Werte abhängt,wird zusätzlich der Unterschied zwischen den Ergebnissen der zwei Methoden gegenderen Mittelwert aufgetragen, womit sich ein „Bias“ (systematische Abweichung)erkennen lässt. Es werden die Übereinstimmungsgrenzen („limits of agreement“) alsmittlere Differenz zwischen Methode A und B ± 2 Standardabweichungen angegeben.
Die Bland-Altman-Analyse erlaubt die Beurteilung der Konkordanz zweier Messreihenin mehrerer Hinsicht: Erstens erkennt man an der vertikalen Streuweite der Punkte dieAbweichungen der einzelnen Messergebnisse voneinander (die Differenzen). NachBerechnung einer linearen Regressionsgerade und deren Darstellung im Diagramm,können zweitens Lage und Neigungswinkel der Regressionsgerade systematische Fehler

Methodenübersicht
66
darlegen. Die Lage (über oder unter Null) erklärt die Tendenz zur Über- oderUnterbewertung, während die Steigung über eine mögliche Beeinflussung derSchwankungsbreite durch die Höhe der Ergebnisse informiert.
Tab. 2.2.4: Erklärungen zur Interpretation von Bland-Altman-Plots
Beispiele zur Interpretation der Bland-Altman-PlotsLage derRegressionsgeraden
Methodik A (z.B. subkutanes Fett-Volumen aus MRT) liegt inden meisten Fällen höher (∅ = 5,5 %) als Methodik B (z.B.subkutanes Fett-Volumen aus Ultraschallmessung) - Daten ausABE et al., 1996):Die Gerade liegt über Null (y = A - B; Differenz > 0)
Steigung derRegressionsgeraden
Je höher die Messergebnisse ausfallen, desto größer werden dieUnterschiede der beiden Messverfahren: Die Gerade steigt an(Anstieg > 0).
Aspekt Die eigentliche Verteilung der Koordinaten kann Aufschlussüber Schwerpunkte im Untersuchungsmaterial geben undbesonders fehleranfällige Messniveaus anzeigen, welche nichtdurch eine Gerade dargestellt werden können.
Generell ist die Reproduzierbarkeit von subkutanen Fettflächenmessungen an
MR-Bildern unter Verwendung von Bildverarbeitungssoftware höher als für
Abdominalfettflächen (Variationskoeffizienten < 5,0 % vs. > 9 – 26 %). Das Ab-
dominalfettdepot unterliegt stärkeren Messfehlern durch Anwender- und Softwarefehl-
klassifikationen der Pixel, die vor allem durch Bewegungsartefakte und
Magnetfeldinhomogenitäten hervorgerufen werden. Beim Schwein ergibt der subkutane
Fettgehalt eine leicht höhere Genauigkeit als der Magergewebe- bzw. Fleischgehalt
(SZABO et al., 1999, BAULAIN und HENNING, 2001, MITCHELL und SCHOLZ, 2001,
MITCHELL et al., 2001b; eigene Ergebnisse siehe Kapitel 3.2.2.3). Die Verwendung
mehrerer Messpositionen, d.h. die Aufnahme mehrerer, aufeinander folgender Bilder,
erhöht – nicht allein für MR-Bildaufnahmen - die Genauigkeit (HASSEN et al., 1999,
SZABO et al., 1999, BAULAIN und HENNING, 2001, MITCHELL und SCHOLZ, 2001,
MITCHELL et al., 2001b, siehe Kapitel 3.2.2.3). Die beste Wahl für eine genaue und
reproduzierbare MR-Bildanalyse zur Ermittlung der Körperzusammensetzung besteht in
einer anwenderkontrollierten, wissensbasierten Bildverarbeitung (Definition von
interessierenden Regionen) mit Optionen für eine Korrektur automatisch festgelegter
ROI’s. Eine objektive statistische Bildanalyse beruht auf Signalintensitätsverteilungen
(Grauwertverteilungen) für jede Einzelscheibe und setzt eine „artefaktfreie“ MR-
Bilderzeugung voraus. Exakte Volumenmessungen liefern ein direktes Maß der
Körperzusammensetzung und sollten gegenüber einer Schätzung der

Methodenübersicht
67
Körperzusammensetzung mittels multipler Regressionsgleichungen unter Einbeziehung
einer Vielzahl von MR-Parametern favorisiert werden (BAULAIN, 1997, ELBERS et al.,
1997, THOMAS et al., 1998, SZABO et al., 1999, TANG et al., 2000, BAULAIN und
HENNING, 2001, MITCHELL und SCHOLZ, 2001, MITCHELL et al., 2001b).
- Längen-, Flächen- und Volumenmessungen
innerhalb manuell, semi- oder vollautomatisch definierter interessierender
Regionen bzw. Volumen (ROI’s bzw. VOI’s) unter teilweiser Verwendung
nachfolgender Techniken
- Schwellwertanalyse (mit festen oder variablen Schwellen in Abhängigkeit vom
auszuwertenden Gewebe)
Grauwert- bzw. Signalintensitätsprofile
- Histogrammanalyse (ein- und mehrdimensional)
Klassen von Grauwerthäufigkeiten oder „Mixtur-Verteilungen“
- Texturanalyse
Beziehung zwischen benachbarten Bildpunkten
- Korrelationsanalyse
- Clusteranalyse
Objektmerkmale (z.B. zeitlich aufeinander folgende Signalintensitäts-
werte) in einem Merkmalsvektor
Abb. 2.2.18: Übersicht zu statistischen Verfahren für die quantitative Bildauswertung(modifiziert nach SCHOLZ, 1994 und MITCHELL und SCHOLZ, 2001)
Die beschriebenen Verfahren stellen einen Querschnitt der möglichen statistischen
Auswertungsmethoden dar und werden durch die kombinierte Verwendung neuronaler
Netze und Fuzzy-Logik-Methoden erweitert und verbessert (BAULAIN und SCHOLZ,
1996, BAULAIN, 1997). Besonders für die Evaluierung biologischer Systeme und deren
Messung werden zunehmend diese statistischen Methoden verwendet. In Anlehnung an
das Design des visuellen Wahrnehmungsapparates von Säugetieren bilden sich unter
Hilfestellung des Anwenders (selbst organisierend) „Neuronen“ aus, die als
Klassifikatoren für bestimmte Bildparameter dienen. Es sollte also möglich sein,
Klassifikatoren zu entwickeln, die sich selbständig auf die in einem Bild vorhandenen
Texturen, Grauwerte sowie weitere Parameter einstellen; z.B. können als
Eingabesignale in das Netz die Grauwerte der Umgebung eines Pixels eingehen. In der
Trainingsphase werden dabei zufällig gewählte Pixelumgebungen oder interessierende
Regionen eines Beispielbildes als Eingabemuster verwendet. Nach dem Training ordnet

Methodenübersicht
68
das Netz jedem Pixel im Bild eindeutig eines seiner Neuronen zu. Als nächstes werden
die Neuronen z.B. durch einen Farbcode einer Gewebekategorie zugeordnet. Diese
Zuordnung kann sowohl durch direkte Eingabe interaktiv am Computer geschehen oder
auch aus einer Menge von Beispielmustern für die Kategorien automatisch gewonnen
werden.
2.2.3.2.5.3 Magnetresonanz-Spektroskopie
Das Phänomen der kernmagnetischen Resonanz (NMR oder MR) ist Basis sowohl
für die Tomographie als auch für die entsprechende Spektroskopie. Eine Beschreibung
des Prinzips erfolgte bereits im Kapitel Muskelstoffwechsel (2.1) bzw. Kapitel
Magnetresonanz-Tomographie (2.2.3.2.5.2). Spektroskopische Anwendungen
konzentrieren sich – wie die Positronen-Emissions-Tomographie (z.B. HSU et al., 1996,
MÜLLER et al., 1997) - hauptsächlich auf Komponenten des Stoffwechsels wie
Glykogen und Glukose unter Verwendung der 13C-NMR-Spektroskopie oder unter
Verwendung der 31P-NMR-Spektroskopie auf Phosphokreatin, ATP und anorganisches
Phosphat (DECANNIERE, 1993, SCHOLZ et al., 1998). Da sich die Resonanzfrequenzen
von Wasser und Fett um ca. 3,5 ppm (parts per million) unterscheiden, verwendeten
MITCHELL et al. (1991 b,c,d) die Gesamtkörper-1H-NMR-Spektroskopie für
Untersuchungen der Körperzusammensetzung von Geflügel (Putenküken und Hühnern)
post mortem, Muskelproben vom Schwein in vitro sowie von Mäuselinien in vivo. Im
Zusammenhang mit einem Vergleich des Lipid- und Wassergehaltes von normalen
(C57b6/6J) und fetten (C57b6/6J-ob) Mäuselinien ermittelten MITCHELL et al. (1991d)
in vivo eine Korrelation von 0,92 zwischen dem chemischen Körperlipidgehalt und dem
NMR-Verhältnis von Wasser-zu-Lipid-Flächen mit einem 0,5 Tesla NMR-System (21
MHz) bzw. eine Korrelation von 0,995 unter Verwendung eines 4,7 Tesla NMR-
Systems (200 MHz).
FUSCH et al. (1999) untersuchten unter Nutzung der sogenannten drei-
dimensionalen „Chemical-Shift“-Bildgebung, einer erweiterten Form der
zweidimensionalen 1H-NMR-Spektroskopie, in einem 1,5 Tesla NMR-System Ferkel
im Alter von 1 bis 37 Tagen post mortem. Sie ermittelten ein Bestimmtheitsmaß von
0,988 (n = 15) für die Beziehung zwischen dem chemischen Körperfettgehalt und dem
Fettgehalt aus der NMR-Analyse, der aus dem NMR-Fett-zu-Wasser-Verhältnis und der
Deuterium-Verdünnungsanalyse (in vivo) errechnet wurde. Die Beziehung zwischen

Methodenübersicht
69
dem chemisch ermittelten Fett-zu-Wasser-Verhältnis und dem NMR-Fett-zu-Wasser-
Verhältnis wurde mit einem Bestimmtheitsmaß von 0,984 (n = 19) angegeben. Die
mittels NMR ermittelten Messwerte fielen jeweils um einen Faktor von 0,772 für das
Fett-zu-Wasser-Verhältnis bzw. von 0,841 für den Fettgehalt niedriger als die chemisch
bzw. in Kombination mit der Verdünnungsanalyse ermittelten Werte aus.
Generell wird die Gesamtkörper-Spektroskopie und/oder –Tomographie nach dem
jetzigen Stand der Technik für Untersuchungen der Körperzusammensetzung auf relativ
kleine Tiere (z.B. Schweine < 120 kg) oder Körperteile von größeren Tieren (z.B.
Teilstücke von Rinderschlachtkörpern) begrenzt bleiben (GROENEVELD, 1985,
MITCHELL et al., 1991a).
GEERS et al. (1995) und VILLE et al. (1997) ermittelten in vivo den
intramuskulären Fettgehalt (IMF) des M. longissimus dorsi von Schweinen mittels 1H-
NMR-Spektroskopie und verglichen diese Messungen mit den in vitro Ergebnissen aus
der chemischen (gravimetrischen) und einer infrarotspektroskopischen Analyse. Beide
Untersuchungen ergaben signifikante Differenzen zwischen den IMF-Gehaltswerten in
vitro und dem NMR-IMF-Gehalt in vivo. Besonders große Unterschiede traten in einer
genetischen Linie mit dem höchsten chemischen IMF-Gehalt von 1,76 % zu 1,11 %
NMR-IMF-Gehalt auf (VILLÉ et al., 1997). Die Beziehungen zwischen gravimetrischem
IMF-Gehalt und NMR-IMF-Gehalt mit einem R2 von 0,41 deuten auf eine immer noch
relativ geringe Verlässlichkeit von Messwerten hin, die mit einem IMF-Gehalt von
< 1,5 % nur einen kleinen Anteil der gesamtem Gewebemasse repräsentieren (GEERS et
al., 1995). Eine Verbesserung der Auflösung mit den jüngsten NMR-/MRT-
Technologien und/oder der Auswahl größerer Messvolumina unter Verwendung
größerer Oberflächenspulen wird zur Erhöhung der Messgenauigkeit von anteilmäßig
kleinen Gewebekomponenten führen.
2.2.3.2.5.4 Dualenergie-Röntgenabsorptiometrie (DXA)
Die Dualenergie-Röntgenabsorptiometrie (DXA oder DEXA) wird im
Humanbereich zunehmend als Methode der Wahl oder als „Goldstandard“ für die
Ermittlung der Körperzusammensetzung in vivo angesehen (LUKASKI, 1987,
HEYMSFIELD et al., 1989, 1997, PIETROBELLI et al., 1996, FOGELHOLM und VAN
MARKEN LICHTENBELT, 1997, PRIOR et al., 1997). Zum Teil werden jedoch berechtigte,

Methodenübersicht
70
später diskutierte Kritiken an dieser Methode geäußert (ROUBENOFF et al., 1993,
LASKEY, 1996, LAPILLONNE et al., 1997, KOHRT, 1997, SPEAKMAN et al., 2001).
Für eine richtige Verwendung dieser Technik zur Ermittlung der
Körperzusammensetzung ist es notwendig, über die physikalischen Prinzipien und
grundlegenden biologischen Modelle der Körperzusammensetzung informiert zu sein
(PIETROBELLI et al., 1996, 1998a,b).
DXA basiert wie alle Röntgentechniken auf der von einer Röntgenquelle
ausgehenden Emission von Photonen, deren Intensität in Abhängigkeit von der
Körperzusammensetzung unterschiedlich stark geschwächt wird. Vorläufergeräte waren
mit einer einfachen Radionuklidquelle wie 125I oder/ und 241Am ausgestattet. Jedes
Radionuklid ist durch eine spezifische Halbwertzeit und Photonenenergie
(Photonenflussdichte) charakterisiert. Wird eine Radionuklidquelle mit einem
„Photonen-Peak“ verwendet, spricht man von „Monophotonen-Absorptiometrie“ oder
„Single Photon Absorptiometry“ (SPA). Nutzt man eine Radionuklidquelle wie z.B.153Gd mit zwei Hauptphotonen-“Peaks“ bei zwei diskreten Energieniveaus (z.B. 44 und
100 keV) ergibt sich die Dualphotonen-Absorptiometrie – DPA - (PEPPLER und
MAZESS, 1981, GOTFREDSEN et al., 1984, 1986, GRIER et al., 1996, PLOURDE, 1997).
Energiequellen, die Strahlung auf zwei Energieniveaus emittieren (wie z.B. 109Cd und153Gd), ersetzen die einfachen Energiequellen seit Beginn der achtziger Jahre
(PIETROBELLI et al., 1996).
Später wurden die Radionuklide durch Röntgenstrahlung erzeugende Quellen
abgelöst, um mit DXA eine höhere Photonenflussdichte und damit höhere
Geschwindigkeiten sowie Genauigkeiten im Vergleich zu SPA und DPA zu erreichen
(PLOURDE, 1997). Im Gegensatz zu den monoenergetischen und dienergetischen
Radionuklidquellen bei der Mono- und Dualphotonen-Absorptiometrie bestehen bei der
Dualenergie-Röntgenabsorptiometrie die erzeugten Röntgenstrahlen aus einem breiten
Spektrum von Photonenenergieniveaus im Bereich von ca. 15 keV bis 80 keV.
Verschiedene Verfahren können genutzt werden, um die zwei erforderlichen
Hauptenergieniveauspitzen zu erzeugen. Häufig werden sogenannte „Photonenfilter“
verwendet, die die besonderen energetischen Eigenschaften der Elektronenhülle von
selten vorkommenden Elementen wie Cerium (Ce) zur Grundlage haben. Treffen die
Photonen aus der Röntgenquelle auf die Elektronen der innersten Elektronenhülle (K-
Hülle) von Cerium, kommt es zu einer Interaktion zwischen Elektronen und Photonen,

Methodenübersicht
71
die die Photonenintensität schwächt. Dazu müssen die Photonen aus der Röntgenquelle
dasselbe Energieniveau wie die Elektronen in der K-Hülle bzw. ein geringfügig höheres
Energieniveau aufweisen. Die Schwächung der Photonenintensität resultiert
hauptsächlich aus Absorption und zum Teil aus einer ungerichteten Streuung
(Ablenkung) der Photonen beim Durchdringen der K-Elektronenhülle. Daraus ergibt
sich in einem polychromatischen Röntgenspektrum eine typische „Absorptionskante“,
die so genannte „K-Kante“ (K-edge). An diesem Punkt (Energieniveau) wird die
Photonenintensität schlagartig geschwächt. Für Cerium liegt die „K-Kante“ bei
40,45 keV. Polyenergetische Röntgenstrahlung kann unter Verwendung des
Ceriumphotonenfilters in zwei Hauptphotonen-Energieniveaus bei einerseits etwa 38-40
keV und andererseits bei ca. 70 keV aufgespaltet werden, so dass vereinfacht zwei
getrennte Photonenstrahlen entstehen.
Werden diese beiden Photonenstrahlen auf Gewebe (Probanden) gerichtet,
vermindert sich die transmittierte Photonenintensität (in vivo) vor allem durch
Absorption und ungerichtete Streuung. Die zwei physikalischen Phänomene „Compton
Scattering“ (Streuung) und „photoelektrischer Effekt“ sind dafür maßgeblich
verantwortlich. Ein Detektor (Photonenzählgerät) ermittelt die Anzahl der das Gewebe
durchdringenden Photonen getrennt für das niedrige (L) und das hohe (H)
Energieniveau, so dass die Anfangsintensität (I0) relativ zur Endintensität (I) gemessen
werden kann. Jedes Gewebe oder Gewebegemisch verfügt über eine spezifische
Verhältniszahl (R), die sich aus der folgenden Formel ergibt:
R= ln(I/ I0)L / ln(I/ I0)H .
Fettgewebe (Fettsäuren und Triglyzeride) weist theoretische R-Werte im Bereich
von 1,2058 - 1,2333 auf, während Protein mit einem theoretischen R-Wert von 1,2906,
Glykogen mit R = 1,3010, Skelettmuskulatur mit R = 1,352 - 1,353, Herz- und Leber-
gewebe mit R = 1,353 - 1,356, Nierengewebe mit R = 1,358 - 1,360, Wasser mit
R = 1,3572 und Knochenmineralstoffe mit einem weit höheren R-Wert von 2,8617
angegeben werden (PIETROBELLI et al., 1996, 1998a). Die R-Werte einzelner Elemente
liegen zwischen 1,0891 für Wasserstoff und 4,0162 für Eisen (PIETROBELLI et al.,
1998b).
Theoretisch können diese R-Werte aus physikalischen Zusammenhängen abgeleitet
werden, die detailliert von PIETROBELLI et al. (1996) und LASKEY (1996) beschrieben
werden. Jede Substanz besitzt einen spezifischen linearen „Schwächungskoeffizienten“

Methodenübersicht
72
(linear attenuation coefficient = µ) und ist durch eine spezifische „Dicke“
gekennzeichnet, die der Photonenstrahl als Weglänge (L) zurücklegen muss. Diese
Vorgaben führen zur Formel für die Photonenintensitätsschwächung:
-d (I/ I0) = µ • dL
bzw. nach Integration der Gleichung zu:
I = I0 e -µ • L.
Folglich hängt die transmittierte Photonenintensität (I) von der Ausgangsintensität (I0),
dem linearen Schwächungskoeffizienten (µ) und der Weglänge (L) ab. Da der lineare
Schwächungskoeffizient (µ) dichteabhängig ist, wird üblicherweise bei in der Dichte (∂)
variierenden Substanzen mit dem Masseschwächungskoeffizienten (µm) als µ/∂ (in
cm2/g) gearbeitet. Dadurch wird die Abhängigkeit zwischen physikalischer Dichte und
linearem Schwächungskoeffizienten kompensiert. Für eine spezifische Substanz
(Element) ist der Masseschwächungskoeffizient (µm) in Abhängigkeit von der
Photonenintensität eine Konstante, die sich experimentell mit Hilfe der
Neutronenaktivierungs-Analyse ermitteln lässt.
Die Formel für den R-Wert lautet allgemein:
R=Σ[fi • (µmi)L]/Σ[fi • (µmi)
H],
mit fi= Massefraktion der i-ten Komponente in einer heterogenen absorbierenden Substanz
µmi = Masseschwächungskoeffizient der i-ten Komponente
L = niedriges Photonenenergieniveau
H = hohes Photonenenergieniveau.
Speziell für Weichgewebe lautet die Formel:
RWG = (µWG)L/(µWG)H mit WG = Weichgewebe
bzw. für Knochengewebe:
RK = (µKG)L/(µKG)H, mit KG = Knochengewebe.
Wenn die RWG-Werte für reines „Magergewebe“ und reines Fett bekannt sind, kann die
Magergewebefraktion nach der folgenden Formel berechnet werden, da sich die RWG-
Werte linear mit der Magergewebefraktion ändern (GOTFREDSEN et al., 1986):
Magergewebefraktion = (RWG – RFett)/(RMagergewebe – RFett),
wobei RWG den Durchschnittswert für den gesamten Scan, RFett den niedrigsten
gemessenen R-Wert im (Weich-)Gewebe und RMagergewebe den höchsten gemessenen R-

Methodenübersicht
73
Wert im (Weich-)Gewebe darstellt. Falls ein R-Wert den RMagergewebe – Wert übersteigt,
enthält der entsprechende Bildpunkt Knochengewebe.
Generell sinkt mit steigender Photonenflussdichte der Masseschwächungskoeffizient
(µm) für ein bestimmtes Element ab, wobei Elemente mit einer höheren atomaren Masse
auch höhere Masseschwächungskoeffizienten (µm) aufweisen. Bei Elementen mit
höheren Masseschwächungskoeffizienten ist der relative Rückgang der transmittierten
Photonenintensität mit steigender Anfangsphotonenintensität stärker als bei Elementen
mit niedrigeren Masseschwächungskoeffizienten (atomaren Massen).
DXA-Messungen beruhen auf der Modellannahme, dass der (Tier-)Körper aus
drei Komponenten besteht, die durch die spezifischen Eigenschaften der
Photonenschwächung in Fett, Knochenmineralstoffe und Rest oder
„Magerweichgewebe“ getrennt werden können (Abb. 2.2.19).
Abb. 2.2.19: Drei-Komponenten-Modell der Körperzusammensetzung für DXA-Untersuchungen (Fett-/ Magerweichgewebe und Knochengewebe),modifiziert nach PIETROBELLI et al. (1996).
Obwohl im Ergebnis der Röntgendurchstrahlung ein zweidimensionales Bild
entsteht (Abb. 2.2.21), ist DXA nicht direkt von einer Bildauswertung abhängig wie z.B.
die Magnetresonanz-Tomographie oder Röntgen-Computer-Tomographie.
Theoretisch wäre es für ein Drei-Komponenten-Modell nötig, Messungen mit drei
verschiedenen Photonenenergieniveaus durchzuführen. Das DXA-Verfahren basiert
jedoch auf der Nutzung von zwei Photonenenergieniveaus, so dass DXA „direkt“ nur
* u.a. Glukose, Glykogen..
Mager-gewebe
Mager-weich-gewebe
Fette (Lipide)
Wasser
Knochen-Mineralstoffe
Proteine
Kohlenhydrate*
Mineralstoffe
Weich-gewebe
Körper-masse

Methodenübersicht
74
die Masseanteile einer Zwei-Komponenten-Mischung bestimmen kann (LASKEY, 1996,
PIETROBELLI et al., 1998b). Um mit DXA drei Komponenten schätzen zu können,
müssen die Bildpunkte (Pixel) zuerst aufgeteilt werden in Pixel, die nur Weichgewebe
(Fett und Magergewebe) enthalten und in Pixel, die sowohl Weichgewebe als auch
Knochenmineralstoffe (Rohasche oder intakter Knochen) enthalten. Dazu wird der R-
Wert benötigt. Der R-Wert für Knochen(-mineralstoffe) fällt mit > 2,8 bedeutend höher
aus als der R-Wert für Weichgewebe mit ~1,2 für Fettgewebe und ~1,35 für „reines
Magergewebe“ bzw. Muskel (bei 38 - 40 und 70 keV). Aufgrund der hohen R-Wert-
Unterschiede zwischen Knochen- und Weichgewebe kann über eine Schwellensetzung
auf Basis einer iterativen Histogramm-Analyse kombiniert mit komplexen
Bildverarbeitungs-Algorithmen zwischen Bildpunkten (Pixel), die Knochen-
mineralstoffe enthalten und Bildpunkten, die allein aus Weichgewebe bestehen,
differenziert werden. Für die Knochenpixel wird angenommen, dass sie nie einzeln
sondern immer in „Clustern“, d.h. größeren zusammenhängenden Gruppen auftreten.
Die anatomischen Kenntnisse des Untersuchers dienen als zusätzliches Hilfsmittel für
die Unterscheidung zwischen Knochen- und Weichgewebe.
Abb. 2.2.20: Positionierung eines Probanden (Kalb) für einen Ganzkörperscan (ausSCHOLZ u.a., 2001)

Methodenübersicht
75
Da nahezu jeder Weichgewebe-Bildpunkt aus fettfreiem Magergewebe und Fett
zusammengesetzt ist und damit kaum R-Werte auftreten, die eindeutig Fettgewebe bzw.
Magergewebe zugeordnet werden können, wird die Fettmasse bzw. Magergewebemasse
mit Hilfe softwareinterner Schätzgleichungen ermittelt. Diese basieren auf den
beschriebenen R-Werten, die zum Teil unter Verwendung von Kalibrier-Phantomen
bestimmt wurden, die entweder aus Polyethylenhohlzylindern (TOTHILL et al., 1997)
oder reinem Fett bzw. Knochen bestehen.
Abb. 2.2.21: Beispiel für das Ergebnis einer DXA-Studie eines Kalbes mit einemKörpergewicht von 42,43 kg (oben: Knochengewebe, unten: Fett- und Magergewebe)(aus SCHOLZ u.a., 2001)

Methodenübersicht
76
Die DXA-Methodik geht außerdem davon aus, dass der Wassergehalt der
fettfreien Masse mit ca. 73 - 74 % konstant bleibt (ROUBENOFF et al., 1993, LASKEY,
1996, PIETROBELLI et al., 1998b). Da Tierkörper ebenso wie junge, sehr alte oder kranke
Probanden des Menschen diesen Idealvorstellungen eines fixen Wassergehalts in der
fettfreien Masse nicht entsprechen (Abb. 2.2.2) und damit Diskrepanzen zwischen den
DXA-Gehalten an Fett, Magergewebe sowie Knochenmineral und den chemischen
Analysewerten verursachen, sind umfassende Referenzuntersuchungen zur Erzeugung
genauerer Schätzgleichungen erforderlich (ROUBENOFF et al., 1993, TOTHILL et al.,
1994, LAPILLONNE et al., 1997, PIETROBELLI et al., 1998b, FUSCH et al., 1999, LUKASKI
et al., 1999, SPEAKMAN et al., 2001). Beispielsweise würde nach LASKEY (1996) mittels
DXA bei einem Individuum mit einem überdurchschnittlichen Wassergehalt in der
fettfreien Masse (> 74 %) der Fettgehalt im Gesamtkörper überschätzt werden. Diese
Aussage steht im Widerspruch zu den Ergebnissen von ELOWSSON et al. (1998), die
jedoch bei ca. 12 Wochen alten Schweinen mit Körpermassen zwischen 14,51 und
21,69 kg (∅ = 17,74 ± 2,93 kg) einen durchschnittlichen Wassergehalt von 78,2 % in
der fettfreien Masse ermittelten. DXA unterschätzte hier den Fettgehalt im
Gesamtkörper im Vergleich zur chemischen Analyse signifikant. Eine Ursache liegt
möglicherweise in der Verwendung von enthaupteten Schlachtkörpern ohne innere
Organe, die durch das DXA-Scannen und Unterwasserwiegen bis zur chemischen
Analyse der rechten Schlachthälfte bereits an Substanz (Wasser) verloren haben
könnten. Noch widersprüchlicher zur Aussage von LASKEY (1996) sind die Ergebnisse
von FUSCH et al. (1999). Diese Autoren fanden zwar mit einem Bestimmtheitsmaß von
R2 = 0,961 eine hohe Beziehung zwischen chemisch ermittelter Fettmasse und DXA-
Fettmasse, aber mit einem durchschnittlichen Wassergehalt von 80 % in der fettfreien
Masse bei Ferkeln im Alter von 1 bis 37 Tagen wurden bei sehr niedrigen Fettmassen
die chemisch ermittelten Werte mittels DXA überschätzt und bei höheren Fettmassen
unterschätzt.
Generell sind die DXA-Messungen des Fettgehalts von lebenden Schweinen hoch
mit dem Fettgehalt aus dem chemisch analysierten Schlachtkörper korreliert (r = 0,912 -
0,950 für Fettgehalt in % und 0,970 - 0,989 für den Fettgehalt in kg) (MITCHELL et al.,
1996b, LUKASKI et al., 1999). Eine negative Abweichung von den Referenzwerten tritt
jedoch bei niedrigeren Fettgehalten (%) auf, die scheinbar mit der Körpermasse
korreliert sind und unkorrigiert zu einer Unterschätzung des Fettgehaltes bei kleinen

Methodenübersicht
77
Schweinen führen (MITCHELL et al., 1998e). Diese Abweichung hängt zusätzlich vom
Gerätehersteller und der Softwareversion ab, da andere Studien eine Unterschätzung,
aber in den meisten Fällen eine Überschätzung des Fettgehaltes bei kleinen Schweinen
(Probanden) feststellten (BRUNTON et al., 1993, 1997, ELLIS et al., 1994, PICAUD et al.,
1996, PINTAURO et al, 1996, ELOWSSON et al., 1998, RIGO et al., 1998, persönliche
Mitteilung John Leja, Manager Product Engineering, GE Medical Systems – Lunar,
2000, 2001). FUSCH et al. (1999) ermittelten zwischen dem DXA-Knochenmineralgehalt
und dem chemisch ermitteltem Kalziumgehalt eine Korrelation von r = 0,996 bei einem
Anteil von 44,1 ±± 4,2% Kalzium im Knochenmineralgehalt. Die DXA-Fettmasse
korrelierte bei einem Bestimmtheitsmaß von R2 = 0,961 ebenfalls hoch mit der
Gesamtfettmasse. Eine Verwendung von entsprechend angepassten Regressions-
gleichungen verbesserte die Genauigkeit der DXA-Messungen:
Gesamtfettmasse korrigiert (g) = 1,31 • DXA-Fettmasse – 68,8 bzw.
Kalziumgehalt korrigiert (g) = 0,402 • DXA-Knochenmineralgehalt + 1,7.
Für Schweine zum Zeitpunkt der Vermarktung liefert DXA hingegen genaue
Messwerte für Gesamtkörperfett-, Magergewebe- und Knochenmineralmasse
(SVENDSEN et al., 1993, MITCHELL et al. 1996a, b, 1998a). GEERS et al. (1996)
bestätigen zwar die Genauigkeit der DXA-Methodik für die Messung des
Gesamtkörpermagergewebes, bezweifeln jedoch die Verwendbarkeit von DXA für eine
genaue Messung des Gesamtkörperfettes. Sie erzielten Wiederholbarkeitswerte
innerhalb der Schweine mit einem Fehler von 4,63 ± 8 (%) für das Gesamtkörperfett
und von 0,33 ± 1,8 (%) für das Magergewebe.
LUKASKI et al. (1999) ermittelten ebenfalls bei sehr hohen Korrelationen zwischen
chemischen Referenzwerten und DXA-Messwerten (R2 = 0,927 - 0,988) eine Unter-
schätzung der Fettmasse mittels DXA um ca. 4 kg (20,5 kg zu 16,4 kg in Bauchlage
bzw. 16,0 kg in Seitenlage). Dabei spielt es keine (signifikante) Rolle, ob die Tiere, die
zwischen 52 und 113 kg wogen, auf dem DXA-Tisch in Bauch- oder Seitenlage
untersucht wurden. PIETROBELLI et al. (1998b) charakterisieren drei Ursachen für
systematische Abweichungen der DXA-Fettschätzwerte bei überdurchschnittlichen
Wassergehalten im Körper:
1. die elementare Zusammensetzung des über dem Durchschnitt liegenden
Wassergehalts,

Methodenübersicht
78
2. die Fraktion von kombinierter Flüssigkeit und Weichgewebe als
Wasserüberschuss und
3. die ursprüngliche Weichgewebezusammensetzung.
Abb. 2.2.22: Beispiel für DXA-Daten nach einem Ganzkörperscan eines totgeborenenFerkels (1,52 kg Körpermasse) mittels Kleintier-Ganzkörper-Modus – (DerGesamtkörperfettgehalt von 9,79 % [Σ Fettgewebemasse/ΣGewebemasse + ΣBMC • 100] fürdieses Tier ist offenbar überschätzt; eigene Daten, nicht veröffentlicht.)
Als weitere systematische Fehlerquelle geben LUKASKI et al. (1999) sowie SPEAKMAN et
al. (2001) die inhomogene Fett- bzw. Weichgewebeverteilung innerhalb des Tierkörpers
an. Da DXA nur ein zweidimensionales Abbild der Körperzusammensetzung liefert, gilt
die zum Teil nicht zutreffende Annahme, dass das Fettgewebe „hinter“ bzw. „unter“
dem Knochengewebe genauso verteilt ist wie „davor“ bzw. „darüber“. Außerdem ist
1à
2à
3à

Methodenübersicht
79
DXA bei großen Probanden mit kompakten Knochen nicht in der Lage, den Fettgehalt
innerhalb des Knochens – wie im Gehirn bzw. Knochenmark - zu bestimmen. Eine
Unterschätzung des Fettgehalts bei größeren Probanden ist die Folge. Hingegen kann bei
kleineren Probanden der Fettgehalt innerhalb des Knochens durchaus ermittelt werden,
da die Photonen diese noch durchdringen. Eine Überschätzung des Fettgehalts könnte
durch die mögliche Unterschätzung des Knochenmineralgehalts erklärt werden.
Besonders bei Kleintieren (z.B. Ferkeln) sind die Ergebnisse in Abhängigkeit vom
Gerätehersteller und der verwendeten Softwareversion noch sehr widersprüchlich.
Mittlerweile existieren Regressionsgleichungen zur genaueren Schätzung von Fett-,
Protein- und Wassergehalt (%) am lebenden Schwein, die jedoch nur für baugleiche
Geräte verwendet werden können. Speziell für die Kleintier-Ganzkörpersoftware
(Version 4.7c), die auf LUNAR DPX-IQ™-Scannern installiert ist, wurden von
LUNAR® Korrekturgleichungen übermittelt:
BMC, g (korrigiert) = 0,638 • DXA-BMC + 18,5
Fett, % (korrigiert) = 0,321 • DXA-Fett % + 7,15
Fett, g (korrigiert) = 0,517 • DXA-Fett + 118
Magergewebe, g (korrigiert) = 1,108 • DXA-Magergewebe – 366 und
Gewebemasse, g (korrigiert) = 0,949 • DXA-Gewebemasse – 33,9
(LEJA, 2001, persönliche Mitteilung). Aufgrund der niedrigen Referenztierzahl
(n = 4) erscheinen diese Gleichungen jedoch sehr zweifelhaft. Spezielle
Kalibrierungsarbeiten sind deswegen bei kleineren Probanden (Labortiere, junge
Schweine) weiterhin erforderlich, da mit den herstellerseitigen Hard- und
Softwarelösungen der DXA-Fettanteil (%) besonders in der niedrigen Gewichtsgruppe
ohne Korrektur (sehr stark) von den Werten aus der chemischen Analyse abweicht
(MITCHELL et al. 1996, 1998a,e, GEERS et al., 1996, ELOWSSON et al., 1998, ROSE et al,
1998, FUSCH et al., 1999). Die Einführung neuer Gerätelösungen („Fanbeam“ anstelle
von „Pencilbeam“) sowie neuer Software soll eine exaktere Bestimmung der
Körperzusammensetzung speziell für Kleintiere ermöglichen (BERTIN et al., 1998,
FUSCH et al., 1999). Gleichzeitig ist zu beachten, dass große Unterschiede in der
Schätzung der Körperzusammensetzung zwischen den verschiedenen Herstellern
bestehen (TOTHILL et al., 1994, LASKEY, 1996, KISTORP und SVENDSEN, 1997, KOHRT,
1997).

Methodenübersicht
80
Unter Berücksichtigung der oben genannten möglichen Abweichungen von den
Referenzwerten liefert DXA sowohl präzise Messwerte für Weichgewebe (Fett- und
Magergewebe) als auch Knochengewebe auf regionaler und Gesamtkörperbasis.
Deswegen gehört diese Methodik zu den umfassendsten Untersuchungsmethoden unter
den vorhandenen Techniken und ist damit neben der Anwendung im Humanbereich
(WANG et al., 1998) besonders für den (Nutz-)Tierbereich interessant (GRIER et al.,
1996, SPEAKMAN et al., 2001). Die entscheidende Schwierigkeit im Humanbereich, die
Körperzusammensetzung und insbesondere den Gesamtkörperfettgehalt allein auf der
Basis von Modellannahmen schätzen zu können (PIETROBELLI et al., 1996, BUTTE et al.,
1997, GORAN et al., 1998, WANG et al., 1998), wird im (Nutz-)Tierbereich durch die
relativ einfache Möglichkeit von Referenzschlachtungen umgangen (SPEAKMAN et al.,
2001). Im Humanbereich behilft man sich mit Referenzuntersuchungen an verstorbenen
Personen bzw. Körperteilen (LLOYD und MAYS, 1987, ABATE et al., 1994,
MITSIOPOULUS et al., 1998), der Verwendung von Modelltieren bzw. von
Phantompräparaten (MADSEN et al., 1997). Als Modelltier wird neben der Ratte (GRIER
et al., 1996) sowie teilweise Hund und Katze (SPEAKMAN et al., 2001) auch für DXA in
der Mehrzahl der Untersuchungen das Schwein in vivo oder post mortem verwendet
(BRUNTON et al., 1993, SVENDSON et al., 1993, ELLIS et al., 1994, JEBB et al., 1995,
PICAUD et al., 1996, PINTAURO et al., 1996, ELOWSSON et al., 1998, FUSCH et al., 1999).
Für die Nutztierforschung wurden Pionierarbeiten von MITCHELL und CONWAY (1993,
1994) zur Untersuchung der Körperzusammensetzung beim Schwein am ‘Growth
Biology Laboratory’ bzw. ‘Diet and Human Performance Laboratory’ des ‘Agricultural
Research Service’ (United States Department of Agriculture), Beltsville, Maryland
geleistet. Diesen Voruntersuchungen folgten weitere Studien am Schwein, um eine
routinemäßige Anwendung für Körpermasseuntersuchungen in vivo zu gewährleisten
(MITCHELL und SCHOLZ, 1995, 1997, 1998, MITCHELL et al., 1995, 1996, 1998a, b, c, d,
e, SCHOLZ und MITCHELL, 1996). Später begannen gleichgerichtete DXA-Arbeiten am
Schwein in Belgien (GEERS et al., 1996), Australien (SUSTER et al., 2000) sowie in
Deutschland (SOFFNER u.a., 2000, 2001). ROZEBOOM et al. (1998) berichteten von ersten
Gesamtkörper-DXA-Untersuchungen beim Schaf, während MITCHELL et al. (1997a)
DXA für die Untersuchung von Geflügel bzw. Drei-Rippenstücken vom Rind
(MITCHELL et al., 1997b) heranzogen. Erste In-vivo-Untersuchungen zur
Körperzusammensetzung und Knochenmineralisierung bei neugeborenen Kälbern

Methodenübersicht
81
unterschiedlicher genetischer Herkunft werden am Lehr- und Versuchsgut
Oberschleißheim der Tierärztlichen Fakultät der Ludwig-Maximilians-Universität
München durchgeführt (SCHOLZ u.a., 2001). Zusätzlich findet sich bei GRIER et al.
(1996) eine Zusammenfassung von Tiermodellen (Ratte, Katze, Kaninchen, Hund,
Minischwein, Menschenaffe, Schaf, Pferd), die speziell für die Osteoporose-Forschung,
aber auch für Körperzusammensetzungs-Studien im Zusammenhang mit der
Haustierernährung verwendet werden.
Abb. 2.2.23: Einflussfaktoren auf die Variation in der Knochenmasse undKnochendichte
Neben den In-vivo-Anwendungen kann DXA zur Untersuchung von
Schlachtkörpern verwendet werden (MITCHELL et al., 1998c). Mit Hilfe frei
definierbarer Auswahlregionen lassen sich spezifische Schlachtkörperbestandteile wie
z.B. Schulter, Schinken, Kamm, Kotelett und Bauch separat auswerten. Im Gegensatz zu
den meisten Fällen bei den kleineren Schweinen (unter Verwendung der
Kleintierganzkörpersoftware) unterschätzt DXA (unter Verwendung der
Ganzkörpersoftware – „pediatric“ oder „adult“ Versionen) unkorrigiert den Fettgehalt
(%), ist dabei aber sehr hoch mit der chemischen Analyse korreliert (r = 0,9). Eine
Unterschätzung der Fettgehalte bei der Schlachthälftenanalyse trat dabei besonders in
den Kotelett- und Bauchregionen auf.
Weitere Anwendungsmöglichkeiten bieten sich z.B. in der Analyse des
Mineralisierungsgrades einzelner Knochen bzw. von Schildkröten sowie zur Diagnose
von Knochenerkrankungen (Abb. 2.2.24, 2.2.25, 2.2.26, 2.2.28).
Genetik
Ernährung/Fütterung
HormonellerEinfluss
Bewegungsspielraum; Aktivitätsniveau[Umwelt; Haltungsbedingungen]
Knochenmineral-masse und -dichte
Alter
Körpermasse

Methodenübersicht
82
Abb. 2.2.24: Mineralisierungsgrad von Karpal- bzw. Tarsalgelenken bei Schweinen mitunterschiedlichem Gewicht (5 = 40 kg bis 8 = 90 kg), (Material stammt von HEINRITZI
u.a., 2001, nicht veröffentlichte Daten)
Abb. 2.2.25: Mineralisierungsgrad einer Landschildkröte mit 677 g Lebendmasse(Material stammt von HEUBERGER, 2001, nicht veröffentlichte Daten)

Methodenübersicht
83
Abb. 2.2.26: Gesamt-Körperzusammensetzung einer Landschildkröte mit 677 gLebendmasse (HEUBERGER, 2001, nicht veröffentlichte Daten)
Die Abbildungen zeigen eindrucksvoll, die Möglichkeit von DXA zur Knochen-
mineraldichtemessung, während die Körperzusammensetzung bei sehr kleinen und
fettarmen Tieren nur ungenau geschätzt werden kann. Der minimal messbare Fettgehalt
liegt für den LUNAR DPX-IQ-Scanner bei ca. 3,8 % (Abb. 2.2.27).
Abb. 2.2.27: Darstellung der Beziehung zwischen DXA-R-Werten für Weichgewebeund dem prozentualen DXA-Fettgehalt (SCHOLZ, 2001)

Methodenübersicht
84
Außerdem ist eine Mindestanzahl von Weichgewebe-R-Werten erforderlich, um
eine plausible Messung der Körperzusammensetzung vornehmen zu können. Gerade bei
den untersuchten Land-Schildkröten ist deshalb, neben der Gewebemasse, allein der
Mineralisierungsgrad für sinnvolle Auswertungen verwertbar. Die Summe von
Weichgewebemasse (g) und Knochenmineralgehalt (g) ergibt die Gesamtkörpermasse
(Abb. 2.2.25 und 2.2.26). Neben der Messung von Knochenmineraldichte und Fett-
bzw. Magergewebeanteil eignet sich DXA auch für klinische Fragestellungen, z.B. zur
Diagnose von Frakturen (Abb. 2.2.28) oder Wirbelsäulendeformationen.
Abb. 2.2.28: Beispiel für klinische Anwendung - Fraktur des rechten Femurs einesSchweines – Verwendung des Schenkelhals-Modus
2.2.3.2.5.5 Ultraschall
Schall ist eine longitudinale Materiewelle. Die Moleküle oder Atome der Materie
schwingen in Richtung der Schallausbreitung. Beim Übergang von Materie zu Materie
mit verschiedener Dichte ändert sich die Schallausbreitungsgeschwindigkeit, außerdem
kann es an den Übergangsflächen zu Reflektionen kommen. Als Ultraschall wird der
Schall in einem Frequenzbereich über 16 000 Hz bezeichnet (Infraschall < 20Hz), der in
der Regel mit Hilfe elektrischer Kristalle erzeugt wird, die beim Anlegen einer
entsprechenden elektrischen Wechselspannung im Rhythmus der Spannung schwingen.
Diese Kristalle werden als piezoelektrische Kristalle bezeichnet. Materialien, die
piezoelektrische Eigenschaften aufweisen, erzeugen bei Belastungswechsel (z.B.
Druckausübung) aus mechanischer Energie elektrische Energie.

Methodenübersicht
85
In der Tierwelt dient Ultraschall z.B. bei Fledermäusen zur Orientierung. Die
Tiere erzeugen dabei Schall mit einer Frequenz von ca. 40 kHz bis zu ca. 90 kHz.
Bestimmte Insekten erzeugen Ultraschall bis zu 250 kHz sowohl zur Orientierung als
auch zur Betäubung von Beutetieren oder Gegnern.
Für routinemäßige Untersuchungen der Körperzusammensetzung beim Schwein
wird Ultraschall mit einer Frequenz von 1 MHz bis 13 MHz verwendet, wobei auf die
Hautoberfläche des Rückens – unter Verwendung einer „Wasservorlaufstrecke“ zur
Ausschaltung einer toten Zone im Hautbereich - ein Oberflächenschallkopf aufgesetzt
wird. Ultraschall dringt umso weiter ins Gewebe ein, je geringer seine Frequenz ist. Mit
abnehmender Frequenz sinkt aber das räumliche Auflösungsvermögen. Eindringtiefe
und Auflösung müssen demzufolge einer Kompromisslösung folgen. Eine Hauptquelle
für Bildrauschen und andere Störsignale bildet die Körperoberfläche selbst, wobei
Fettmenge, Hautdicke und der Flüssigkeitsgehalt Ursachen für Ablenkungen und
Streuungen des Ultraschallstrahls darstellen (SCHOELGENS, 1998). Extreme Speck- und
Muskelmaße (sehr niedrig oder sehr hoch) werden dadurch offenbar mit einer
geringeren Genauigkeit als intermediäre Ausprägungen ermittelt (MOELLER und
CHRISTIAN, 1998).
Im Prinzip basiert die Technik eines Ultraschallgerätes auf der Umwandlung von
elektrischer Energie in Form eines hochfrequenten Wechselspannungspulses in
Ultraschallenergie, die in der Lage ist, den Körper mit Hilfe kurzer Pulssequenzen
(Pulsdauer ca. 1 µs) zu durchdringen. Gebündelte oder fokussierte, als kurze Impulse
durch die Haut dringende Ultraschall-Wellen werden an Haut- und Gewebe- bzw.
Organschichtgrenzen reflektiert und vom piezoelektrischen Empfänger als Echos im
Schallkopf aufgenommen. Sobald die Ultraschallwellen den Übergang von einem
Gewebe zu einem anderen Gewebe erreicht haben, die sich in ihren akustischen
Eigenschaften unterscheiden, wird ein Teil der Ultraschallwellen reflektiert und vom
Ultraschallsensor im Messkopf, der auch den Ultraschallsender enthält, erfasst. Die
reflektierte, vom Messkopf erfasste Energie (Ultraschall-Druckwelle) wird wieder in ein
elektrisches Signal umgewandelt, amplifiziert und ursprünglich über ein Oszillogramm
angezeigt (Abb. 2.2.29). Variationen in der Gewebedicke bzw. -dichte und
Kompressionsfähigkeit des jeweiligen Gewebes resultieren in zeitlichen Differenzen der
reflektierten Ultraschallwellen. Die Schallgeschwindigkeit beträgt in Luft etwa 330 m/s,

Methodenübersicht
86
in Weichteilen etwa 1540 m/s (Fett = 1440 m/s bzw. Muskel = 1580 m/s) und in
Knochen ~4000 m/s (MAYRHOFER u.a., 1995, BRØNDUM et al., 1998).
Abb. 2.2.29: Ultraschallscanner „B-Mode“ links und Prinzip des Ultraschallverfahrensnach dem „A-Mode“ zur Ermittlung von Fett- und Muskeldicke(modifiziert nach PFEIFFER, 1984 und KALLWEIT, 1991)
Im Zuge der technologischen Entwicklung der Ultraschall-Technik im
humanmedizinischen Bereich während der letzten Jahrzehnte wurden verschiedene
eindimensionale (A-Mode) und zweidimensionale (B-Mode) Geräte im Bereich der In-
vivo-Ermittlung der Körperzusammensetzung zur Standardmethode für die Selektion
von potentiellen Zuchttieren (Schweinen) bzw. Wachstumsstudien. Dabei kommt ein
weites Spektrum von Geräten zur Anwendung, die von einfachen Distanzmessungen
ohne Bildgebung (aber mit Digitalanzeige) bis zu Flächenmessungen mit Bildgebung
reichen (CLAUS, 1957, KLIESCH et al., 1957, STOUFFER et al., 1961, HORST, 1964a,b,
1971, LANGLET u.a., 1968, MERSMANN, 1982, SATHER et al., 1986, 1987, BUSK, 1989,
KAUFMANN, 1990, KRIETER u.a., 1990, BUSEMANN, 1991, KOLB, 1991, STOUFFER,
1991, ERNST, 1994, PFEIFFER und SCHRÖDER, 1994, NITTER und KOLB, 1994, TIBAU et
al., 1996, PESCHKE u.a., 1997, MOELLER und CHRISTIAN, 1998, MOELLER et al., 1998).
Modifizierte Geräte werden für die routinemäßige Klassifizierung der Schlachtkörper
verwendet (u.a. BRANSCHEID, 1994, KÜCHENMEISTER und ENDER, 1994, VAN
BETTERAY, 1994, BUSK und OLSEN, 1996, BRØNDUM et al., 1998, HÖRETH, 2000).

Methodenübersicht
87
A-Mode-Geräte basieren auf der Messung der Schallamplitude, d.h. der Amplitude des
reflektierten Impulses, die die Stärke der Schallreflexion wiedergibt. Die ein-
dimensionale Darstellung der Echos (stehendes Bild) erfolgt als intensitätsproportionale
sinusförmige Auslenkung des Elektronenstrahls nach „oben“, wobei die zeitliche
Abfolge der Echos den Tiefen der reflektierenden Grenzzonen entspricht. Hingegen
werden bei den "B-Mode" -Geräten (Brightness) die Stärken der reflektierten
Echoimpulse nicht wie im A-Mode als Amplitudenhöhe, sondern als Punkte
proportionaler Helligkeiten dargestellt. Zudem wird nicht nur ein Echoimpuls registriert,
sondern der Schallkopf besteht aus multiplen Piezokristallen, die Echoimpulse
aussenden und empfangen. So können verschiedene Ortspunkte entlang des
Schallkopfes unterschieden werden. Durch die zusätzliche Ortsauflösung längs des
Echoimpulses durch die Schalllaufzeit ergibt sich so eine zweidimensionale
Ortsauflösung mit verschiedenen Graustufen entsprechend der Helligkeitsmodulation.
B-Scan-Geräte erzeugen eine zweidimensionale, lichtmodulierte Darstellung der Echos
in Form energie- u. intensitätsproportionaler, helligkeitsvariabler Lichtpunkte (unter
Löschung der 0-Linie) als Resultat einer in 2 Richtungen erfolgenden zeilenförmigen
und schichtweisen Abtastung. Die entsprechend der Echointensität in Grautönen
wiedergegebenen Punkte fließen – soweit von gleichwertigen Strukturen stammend – zu
Flächen und Linien zusammen und geben bei apparativer Verschiebung in schneller
Folge sofort ein sichtbares Bild wieder. „Konturechos“, die an den Gewebeübergängen
zwischen z.B. Fett/Fett oder Fett/Muskel entstehen und diffuse Binnenechos, die durch
Inhomogenitäten im Gewebeinneren hervorgerufen werden, bilden die Grundlage für ein
Ultraschallbild. Die empfangenen Realzeit-Bildsequenzen können bei Bedarf einzeln
oder in Folge für eine Auswertung von Fett- und Muskelflächen bzw. -längenmaßen
gespeichert werden. Nach dem Abtastmodus bezeichnet man die B-Scan-Geräte als
Parallel-Scan [= Bp], Konvergent- u. Divergent-Scan [= Bc bzw. Bd], Misch- oder
Compound-Scan [Bcd, Bcp, Bpd). Detailliertere Beschreibungen dieser Technik finden
sich u.a. bei KOLB (1991), HOUGHTON und TURLINGTON (1992), SEE und STANISLAW
(1994), MILLER (1996) sowie SCHUBERT (1998).
Zusätzlich bietet die in der Entwicklung befindliche 3D-Sonographie
Möglichkeiten für die Volumenbestimmung von Muskel- und Fettgeweberegionen bzw.
die Volumetrie von Organen bzw. Gelenken des (Nutz-)Tieres. Mit ihrer Hilfe können
Gelenke und Organe in ihrer Gesamtheit abgebildet werden. Verwendet werden

Methodenübersicht
88
Schallköpfe mit einem Frequenzbereich von 5 bis 7,5 Megahertz. Bei kleinen Organen
bzw. Gelenken sind auch höherfrequente Schallköpfe denkbar. Die Untersuchung
erfolgt im Realzeit-Verfahren. Eine weitere Verbesserung der 3D-Sonographie im
Hinblick auf die Detailauflösung, die Verarbeitung der Daten und Größe der
Schallköpfe (Problem der Ankoppelung) wird in Zukunft eine Ausweitung der
Einsatzmöglichkeiten dieser Technik bringen. Weitere Entwicklungen wie zunehmende
Verbesserung der Bildqualität und Bildrate, Echtzeit-3D und die Möglichkeit einer
genauen automatisierten Volumetrie lassen den 3D-Ultraschall als zukunftsweisende
und erfolgsversprechende Methode erscheinen, wobei zusätzliche, bislang
sonographisch nicht erzielbare Schnittführungen mit besserer Vergleichbarkeit zu CT-
und MR-Bildern möglich sind (BAUER u.a., 1998, FRANKE u.a., 1998).
Zweidimensionale Ultraschallbilder liefern vor allem Informationen über
Fettgewebedepot- sowie Muskelquerschnitte (BUSK, 1989) und wurden versuchsweise
für die Ermittlung des intramuskulären Fettgehalts beim Schwein getestet (KOLB, 1991).
Aufgrund des sehr niedrigen Marmorierungsgrades in den „modernen“ Schweinerassen
hatte diese Untersuchungsmethodik unter den gegebenen (eingeschränkten) technischen
Bedingungen – auch nach aufwändiger Bildverarbeitung mittels Texturanalyse - keinen
Erfolg (r = 0,35, S.E. = 0,59 % für zweiten Datensatz aus der Kreuzvalidierung),
während für den Fleischanteil im Bauch ein Korrelationskoeffizient von r = 0,62
(S.E. = 8,2 %) ermittelt wurde (KOLB, 1991). STOUFFER (1991) erzielte zwischen
Ultraschallmarmorierung und Marmorierung am Schweine-Schlachtkörper ebenfalls nur
eine Korrelation von r = 0,33, wobei die Marmorierung mittels Ultraschall stark
überschätzt wurde (2,2 ± 0,9 Punkte zu 1,3 ± 0,5 Punkte auf einer 5-Punkte-Skala).
Diese Aussagen ergänzend, erzielten auch VILLÉ et al. (1997) keine signifikanten
Korrelationen (p ≥ 0,02) zwischen Ultraschallmaßen (B-Mode bzw. A-Mode) und
anhand von Muskel-Biopsieproben chemisch ermittelten intramuskulären
Fettgehaltswerten. Die Bestimmtheitsmaße schwankten zwischen 0,312 und 0,435 für
die B-Mode-Ultraschallergebnisse bei etwa 20 kg Lebendgewicht, während die A-
Mode-Ergebnisse (ohne Bildgebung) noch schlechter ausfielen und die chemisch
(gravimetrisch) ermittelten Werte bei mittleren IMF-Gehalten zwischen 1,26 % und
1,85 % (60 bzw. 100 kg Lebendgewicht) systematisch um 1 % überschätzten. Etwas
genauer als beim Schwein lässt sich mittels Ultraschall-Bildgebung die Marmorierung
beim Rind mit einer Wiederholbarkeit von 63 ± 3 % (HASSEN et al., 1999) bzw.

Methodenübersicht
89
altersabhängigen Bestimmtheitsmaßen zwischen R2 = 0,18 und R2 = 0,51 (BRETHOUR,
2000) ermitteln.
Im Allgemeinen stimmen hingegen die aus den In-vivo-Ultraschalluntersuchungen
stammenden Maße für Muskeldicke oder –fläche und insbesondere für
Rückenspeckdicke oder -fläche relativ gut mit den Referenzwerten aus der Schlacht-
körperzerlegung beim Schwein überein. Die Güte der Ergebnisse variiert in
Abhängigkeit vom verwendeten Gerät, (der Erfahrung) der Messperson, der
untersuchten Tierpopulation (Rasse, Tierart), der Messstelle sowie vom Alter und
Körpergewicht der analysierten Tiere. Da die Rückenspeckdicke mit steigendem Alter
und Körpermasse zunimmt (MOELLER et al., 1998, siehe Abb. 2.2.30), ist speziell bei
Tieren mit geringer Rückenfettauflage bzw. mit niedrigen Körpermassen (< 30 kg) die
Aussagefähigkeit der Ultraschallmaße reduziert (KAUFMANN, 1990, ERNST, 1994,
MÜLLER, U. 1994, PFEIFFER und SCHRÖDER, 1994, TIBAU et al., 1996, MOELLER et al.,
1998).
15161718192021222324252627282930
67,4 80,3 93,4 104,9
Durchschnittliche Körpermasse (kg)
Spotted Chester White Poland China Duroc
Berkshire Landrace Yorkshire Hampshire
Abb. 2.2.30: Entwicklung der „Ultraschall“-Rückenspeckdicke im Verlauf desWachstums bei verschiedenen Schweinerassen(Daten stammen aus MOELLER et al., 1998.)
Aus Untersuchungen der vergangenen 30 Jahre mit einer Vielzahl von Geräten
ergaben sich für die Beziehung zwischen Ultraschall-Kotelett-Muskelfläche bzw. –

Methodenübersicht
90
Muskeldicke und den entsprechenden Maßen aus der Schlachtkörperzerlegung
Korrelationen zwischen 0,27 und 0,94 mit einem Durchschnitt von ca. 0,65. Höhere
Korrelationen bestehen in den meisten Fällen zwischen Ultraschall-Rückenfettdicke und
den entsprechenden Schlachtkörpermaßen mit Werten zwischen 0,19 und 0,94 und
einem Durchschnitt von ca. 0,75 (SMITH et al., 1992, Übersichten bei SZABO et al., 1999
und MITCHELL und SCHOLZ, 2001). Von besonderem Interesse für die Schweinezüchter
sind jedoch neben der Messgenauigkeit die Beziehungen zwischen den
Ultraschallmaßen und der geschätzten Körperzusammensetzung in Bezug auf
Fleischanteil und Anteil Fettgewebe (KAUFMANN, 1990). Die Korrelationen zwischen
Fleischanteil (%) und Quotienten aus Ultraschall-Fett-zu-Muskelmaßen schwanken bei
einem Durchschnitt von -0,66 (±0,16) zwischen -0,32 und -0,91 (PFEIFFER u.a., 1984,
KAUFMANN, 1990). Zusätzlich zu den einfachen Beziehungen zwischen Flächen- und
Dickenmaßen bzw. deren Quotienten kommen verschiedene Schätzgleichungen unter
Einbeziehung mehrerer Ultraschallmaße für die Ermittlung des Fleischanteils bzw. der
Fleischteilstücke am lebenden Schwein zum Einsatz. Diese Gleichungen basieren
zumeist auf transversalen oder longitudinalen Scans, die eine oder mehrere Messstellen
für (Rücken)speckdicke bzw. –fläche und Muskeldicke bzw. –fläche sowie zum Teil die
Lebendmasse bzw. Schlachtkörpermasse und das Geschlecht einschließen. Die
Genauigkeiten schwanken zwischen R²-Werten von 0,36 und korrespondierenden
Reststandardabweichungen (RSD) von 3,17 bis zu R²-Werten von 0,83 und RSD von
1.67 (KAUFMANN, 1990, GRESHAM et al., 1992, 1994, CISNEROS et al., 1996,
Übersichten bei SZABO et al., 1999 und MITCHELL und SCHOLZ, 2001). Für Messungen
am Schlachtkörper (zum Teil bereits online und automatisch) sind ähnliche Gleichungen
ermittelt worden mit Genauigkeiten zwischen R² = 0,41 mit RSD = 3,05 bis zu
R² = 0,85 mit RSD = 1,58 (KÜCHENMEISTER und ENDER, 1994, BRANSCHEID u.a., 1994,
GRESHAM et al., 1992, 1994, LIU und STOUFFER, 1995, BRØNDUM et al., 1998).
Zunehmend gelangt mit dem Ziel der Schlachtkörperklassifizierung das sogenannte
AUTOFOM-Gerät, ein automatisch am Schlachtband Fett- und Muskeldicken
messender u-förmiger Ultraschall-Scanner mit 16 kombinierten 2,0 MHz Schallköpfen
(SFK Technologie, K2KG - 67080), zum Einsatz (BUSK und OLSEN, 1996, BRØNDUM et
al., 1998, BRANSCHEID und DOBROWOLSKI, 2000, HÖRETH, 2000). Für die EU-
Zulassung zum kommerziellen Einsatz muss der SEP (Standardfehler der Vorhersage
aus der Kreuzvalidierung) unter 2,5 % Fleischanteil liegen. Das AUTOFOM-Gerät

Methodenübersicht
91
erfüllt in den meisten Fällen diese Voraussetzung mit Reststandardfehlern zwischen
1,58 und 1,84 (bzw. 2,69 für den Bauchfleischanteil, BRANSCHEID und DOBROWOLSKI,
2000) bei Bestimmtheitsmaßen zwischen R2 = 0,85 und R2 = 0,74 für die Schätzung des
prozentualen Fleischanteils (BRØNDUM et al., 1998, HÖRETH, 2000).
2.2.3.2.5.6 Bioelektrische Impedanz (BIA)
In jüngerer Zeit ist, ausgehend von LUKASKI et al. (1985), eine neue Methode eingeführt
worden, die bei einer einfachen Handhabung hohe wissenschaftliche Genauigkeit
versprechen soll: die bioelektrische Impedanz-Analyse (BIA). Da die mit der BIA
erzeugten Ströme durch alle leitenden Gewebe innerhalb des Körpers fließen, können
mit Hilfe dieser Methode die Körperkompartimente (Körperzellmasse, Fettmasse,
Extrazellulärmasse, Körperwasser) differenziert werden (DE LORENZO et al, 1997,
MATTHIE et al., 1998).
Die physikalische Hypothese geht davon aus, dass der Wechselstromwiderstand
(Scheinwiderstand) des Körpers mit dem Körperwassergehalt abnimmt und mit dem
Körperfettgehalt zunimmt, so dass man aus dem Wechselstromwiderstand auf den
Körperwasser- und Körperfettgehalt schließen kann. Aus den Widerstandsmessungen
und weiteren Variablen (Körperlänge, Körpergewicht, Gliedmaßenlänge) lässt sich die
Körperfettmasse über Regressionsgleichungen bestimmen (LUKASKI et al., 1985, DE
LORENZO et al, 1997, BAUMGARTNER et al., 1998). BIA-Messungen sind dabei eher
statistisch mit Variablen wie dem Gesamtkörperwasser oder dem Körperfett verbunden
als mit bio-physikalischen Prinzipien. Obwohl die BIA-Messwerte durch eine Vielzahl
von Variablen beeinflusst werden, wie Körperhaltung (-lage), Hydrierungs-Zustand,
Zeitabstand zur letzten Nahrungsaufnahme, Luft- und Hauttemperatur, vorangegangene
körperliche Aktivität, Platzierung der Elektroden, ungenaue Messung von Körperlänge
und Körpergewicht, dem Körperfettgehalt selbst (!) sowie durch die Leitfähigkeit der
Untersuchungseinrichtung (MONTAGNANI, 1997, BAUMGARTNER et al., 1998), besitzt
die Methode bei Messwiederholungen an mehreren Tagen eine Reproduzierbarkeit von
> 95 % (LUKASKI, 1996). Voraussetzung hierfür ist eine Standardisierung und Kontrolle
der genannten Effekte. Die BIA-Geräte sollten folglich genaue elektrische Daten und
Kalibrierungsangaben aufweisen, die eine Messgenauigkeit innerhalb von 1 % über die
volle Breite des Gesamtwiderstandsbereiches von biologischen Systemen sicherstellen.
Sie sollten den Primärwiderstand und, sofern gemessen, die Reaktanz angeben sowie die

Methodenübersicht
92
Frequenz(en), bei welcher(n) die Messungen durchgeführt wurden. Berechnungen von
Parametern der Körperzusammensetzung aus den Basismessungen sollten anhand
populationsspezifischer Gleichungen/Formeln erfolgen und die Standardfehler für die
individuelle Schätzung angeben (BRÜNGEL und KLUTHE, 1997).
Eine Vielzahl von Gruppen hat versucht, die BIA besonders am Menschen, aber
auch am Tier zu validieren (LUKASKI, 1996). Die Studien resultieren aber in sehr
unterschiedlichen Formeln für die Umrechnung der Widerstandswerte auf den
Körperfettgehalt, so dass der Einsatz der verschiedenen Formeln in einer Messserie zu
sehr unterschiedlichen Ergebnissen führt. Als Beweis für die Genauigkeit der
Übereinstimmung der Bioimpedanzmethode mit der hydrodensitometrischen Methode
werden in den meisten Studien die Korrelationskoeffizienten herangezogen, die aber zu
einer solchen Aussage untauglich sind (siehe Bland-Altman-Analysen im Kapitel
2.2.3.2.5.2). Unter den meisten Bedingungen gibt die BIA eine verlässliche Angabe des
Gesamtkörperwassers. Die daraus abgeleitete Schätzung der fettfreien Masse und des
Prozentanteils des Körperfettes variieren in Abhängigkeit von der untersuchten
Population, Alter, Geschlecht und in Abhängigkeit von der Gültigkeit der verwendeten
Gleichung, mit der die Schätzung vorgenommen wurde. Speziell bei Betrachtung
einzelner Individuen können die Ergebnisse, die mit den verschiedenen Formeln erzielt
werden, sehr stark voneinander abweichen (FOGELHOLM et al., 1996, WANG und
DEURENBERG, 1996, SNIJDER et al., 1999, BUSSOLOTTO et al., 1999).
Die Bestimmung der Körperfettmasse mittels BIA beruht, wie beschrieben, auf
dem Prinzip, dass verschiedene Körpergewebe (Fettgewebe, Muskulatur, Knochen) bei
Stromdurchfluss unterschiedliche Widerstände aufbauen. Da Muskelgewebe (aus mehr
elektrolythaltigem Wasser bestehend) eine bessere elektrische Leitfähigkeit als
Fettgewebe oder Knochen aufweist, kann man anhand des Ausmaßes der Stromstärke,
welche den Körper passiert, die Körperzusammensetzung berechnen. Bei der
ursprünglichen Messmethode werden an den Extremitäten (bzw. an anderen definierten
Stellen) jeder Körperseite zwei Elektroden angebracht und ein Strom geringer Stärke
durch den Körper geleitet. Inzwischen gibt es diverse Messmöglichkeiten und eine
Vielzahl an Geräten, die in einem einfachen oder multiplen Frequenzbereich arbeiten
(SUZUKI et al., 1996). Wird eine Wechselstromquelle geringer Stärke an ein
biologisches System angeschlossen, kann man einen Spannungsabfall oder einen
zunehmenden Widerstand für den Stromfluss feststellen. Die Impedanz

Methodenübersicht
93
(Scheinwiderstand - Z) setzt sich zusammen aus der Resistenz (Ohmscher Widerstand -
Rs) und dem frequenzabhängigen kapazitiven bzw. induktiven Blindwiderstand
(Reaktanz – Xc). Die Reaktanz hängt von der geometrischen Konfiguration und dem
Volumen der stromführenden Leiter im biologischen System und der Signalstärke sowie
der Frequenz der verwendeten Wechselspannung ab. Ist die Geometrie relativ konstant,
dann entspricht die Impedanz einer Funktion des elektrischen Potentials der Masse und
kann wie folgt kalkuliert werden: Z = (Rs2+ Xc2)5.
Da die Impedanz auch vom Volumen des stromführenden Leiters abhängt, gilt:
Z (in ohm) = ρL²/V,
mit ρ= Volumenwiderstand (ohm-cm), L = Leiterlänge (cm) und V = Volumen (cm³).
Alle Leitungsparameter wie Leitungswiderstand, Isolationswiderstand, Leitungs-
kapazität und Leitungsinduktivität gehen in die Impedanz ein. Diese ist bei Kabeln -
aber nicht in biologischen Systemen - über weite Frequenzbereiche frequenzunabhängig.
Mit steigenden Frequenzen wird die Impedanz zunehmend vom Leitungswiderstand
bestimmt. Wie beschrieben, werden zwei Sets von Elektroden an definierten
Körperstellen des Probanden platziert. Danach erfolgt die Impedanz-Messung (Rs und
Xc). Zusätzlich gehen in die Schätzgleichungen das Körpergewicht, die Distanz
zwischen den Detektor-Elektroden (bzw. die Körperlänge) sowie zum Teil die
Temperatur (Körper- bzw. Probentemperatur) ein (SWANTEK et al., 1992, 1999,
MARCHELLO et al., 1999).
Beim Schwein wurde die BIA nur in begrenztem Umfang zur Schätzung der
fettfreien Masse am lebenden Tier und Schlachtkörper (SWANTEK et al., 1992,
MARCHELLO und SLANGER, 1992, MARCHELLO et al., 1999, SWANTEK et al., 1999)
verwendet. SWANTEK et al. (1992) ermittelten zwischen Rs und fettfreier Masse (FFM)
Korrelationen von r = -0,56 am lebenden Tier bzw. r = -0,63 am Schlachtkörper sowie
entsprechend zwischen Rs und Fettanteil (%) Werte von r = 0,64 bzw. 0,7. Im Vergleich
fielen die Korrelationen zwischen den Blindwiderstandsmessungen (Reaktanz) und der
fettfreien Masse (FFM) bzw. dem Fettanteil (%) bedeutend niedriger aus (r < |±0,17|).
Die ermittelten Schätzgleichungen hatten die folgende Form:
FFM (in vivo) = 0,486 Lebendmasse - 0,881 Rs + 0,48 Distanz + 0,86 Xc + 7,959
FFM (Schlachtkörper) = 0,326 Schlachthälftenmasse - 0,081 Rs + 0,239 Distanz + 0,060 Xc + 9,945.

Methodenübersicht
94
Die Bestimmtheitsmaße für die obigen Schätzgleichungen zur Ermittlung der fettfreien
Masse im Schlachtkörper betrugen mit unkorrigierter Lebendmasse R2 = 0,817 bzw.
R2 = 0,844. In einer nachfolgenden Studie führte die Benutzung der obigen Gleichung
für lebende Schweine zwischen 50 und 130 kg zu einer systematischen Unterschätzung
der fettfreien Masse um 9 kg. Die Korrelationen zwischen geschätzter und tatsächlicher
FFM betrugen durchschnittlich 0,67.
Obwohl gerade für (Nutz-)Tiere weitere Untersuchungen nötig sind, scheint die
BIA unter kontrollierten und standardisierten Bedingungen für die Erfassung der
Körperzusammensetzung sowohl für lebende Tiere als auch Schlachtkörper geeignet zu
sein. Der Informationsgewinn durch die Messung der Impedanz-Messwerte zusätzlich
zur Information aus dem Lebendgewicht ist bisher jedoch nach eigener Ansicht zu
gering und rechtfertigt den Aufwand nicht.
Generell ist die BIA nicht geeignet für die Feststellung von kurzfristigen
Veränderungen des Körperfettanteils von Individuen, kann aber zur Charakterisierung
langfristiger Veränderungen in Populationsgruppen angewandt werden.
Humanmedizinische Vergleichsuntersuchungen von BIA mit etablierten Standard-
methoden haben Fehler von ~10 % und bei zunehmendem Übergewicht bis zu 30 %
ergeben. Die BIA ist grundsätzlich nicht unter Konditionen einsetzbar, bei denen eine
ausgeprägte Verschiebung der Wasserverteilung des Körpers vorliegt (GRAY et al.,
1989, BAUMGARTNER et al., 1998).
Mess-Elektroden Quell-Elektroden
Abb. 2.2.31: Prinzip der Elektrischen Impedanztomographie (nach BRANDSTÄTTER,2000)

Methodenübersicht
95
Eine möglicherweise viel versprechende Erweiterung der bioelektrischen
Impedanzmessung ist die in der Entwicklung befindliche elektrische Impedanz-
tomographie (MENZEL, 1997, LI, 2000). Dabei wird ein Elektrodenring um den Körper
gelegt, bei dem ein Teil der Elektroden den Erregerstrom leitet und der andere Teil für
die Potentialmessung an der Oberfläche zuständig ist. Aus den Messdaten wird die
Leitfähigkeitsverteilung rekonstruiert und in Bilddaten umgesetzt (Abb. 2.2.31).
2.2.3.2.5.7 Gesamtkörperleitfähigkeit - „Total Body Electrical Conductivity“(TOBEC)
Die Prinzipien, die für die bioelektrische Impedanzanalyse gelten, treffen identisch für
die TOBEC-Messungen (Elektrische Leitfähigkeit des Gesamtkörpers) zu. Die TOBEC-
Messung beruht ebenfalls auf der unterschiedlichen Leitfähigkeit von Muskel- und
Fettgewebe. Erste Geräte liefen unter der Bezeichnung EMME (Electronic Meat
Measuring Equipment) und wurden zur Messung von Fett- und Fleischgehalt am
lebenden Schwein (Modell SA-1) bereits 1973 durch HARKER in den USA zum Patent
angemeldet (BOILEAU, 1988, SUTCLIFFE et al., 1995). Die TOBEC-Messung wird
anstelle von Messelektroden mit einer elektromagnetischen Spule unter einer
Wechselspannung von 2,5 bis 5,0 MHz betrieben, so dass sich ein magnetisches Feld
innerhalb der Spule aufbaut. Diese Spule dient gleichzeitig als Messkammer. Bewegt
man leitendes Material durch eine stromführende Spule wird eine elektrische Spannung
im leitenden Material induziert, wodurch ein irreversibler Energieverlust in Form von
Wärme entsteht. Der Energieverlust vermindert die Impedanz der Spule und dient als
Index für die leitende Masse der Probe. Die veränderte Impedanz bzw. der induzierte
Stromfluss ist eine Funktion der elektrischen Leitfähigkeit und der dielektrischen
Eigenschaften des untersuchten biologischen Systems. Hierbei beeinflusst speziell die
Kondensatorwirkung der Zellmembranen den dielektrischen Effekt. Tatsächlich
gemessen wird jedoch die Energieabsorption – nicht die Impedanz bzw. Leitfähigkeit.
Das Messergebnis (Menge absorbierter Energie = E) ist eine Funktion der Fläche (A2),
der Magnetfeldstärke (B), der Leitfähigkeit je Volumeneinheit bei einer bestimmten
Frequenz (c) sowie einer Anzahl von Konstanten (k) mit E = A2 • B • c • k, wobei die
Messkurve der Energieabsorption vor allem vom Volumen der fettfreien Masse abhängt.
Außerdem variiert die Messung je nach Instrumentenausführung. Bei stationärer
Ausführung (das Messobjekt wird in der elektromagnetischen Spule (Röhre) nicht

Methodenübersicht
96
fortbewegt) resultiert als Messergebnis die Energieabsorption aus der Differenz der
Messwerte der Spulen-Impedanz ohne und mit Probe. Wird das Objekt während des
Messvorgangs durch die Spulenanordnung bewegt, manuell oder besser mittels
Förderband, resultiert als Messergebnis der Phasendurchschnitt oder die relative
Energieabsorption (E) aus dem elektromagnetischen Feld. Geometrie, Orientierung,
Position und Temperatur der Proben beeinflussen die elektromagnetischen Messsignale
während der Passage durch die Spulenanordnung (Messkammer). Genauso wie bei der
bioelektrischen Impedanzanalyse bestimmen absolute Menge, Zusammensetzung und
Konzentration der in der Probe vorhandenen Elektrolyte durch ihre unterschiedliche
Wirkung auf die Leitfähigkeit die Signalausprägung. Deswegen ist eine Kalibrierung des
TOBEC-Gerätes mit Objekten, die eine vergleichbare Geometrie und bekannte
Zusammensetzung aufweisen, notwendig. Für eine genaue Schätzung der fettfreien
Masse ist es außerdem erforderlich, das Gewicht der Probe zu messen. Speziell bei
Lebendtiermessungen sind Bewegungs- und Positionsartefakte zu beachten, während bei
Schlachtkörpern (Teilstücken) die Temperatur einen wichtigen Faktor darstellt (KLISH et
al., 1984, BOILEAU, 1988, HENNING et al., 1993, FORREST, 1994, MITCHELL und
SCHOLZ, 2001).
Zwei frühe Experimente zur Nutzung von TOBEC an lebenden Schweinen
resultierten in Korrelationen (r) von 0,87 (n = 42) bzw. 0,68 (n = 35) zwischen TOBEC-
Messung und Schlachtkörperwasser (DOMERMUTH et al., 1976). FREDEEN et al. (1979)
und MERSMAN et al. (1984) ermittelten stark variierende Ergebnisse mit generell
niedrigen Korrelationen zwischen TOBEC- und Schlachtkörperwerten. Hingegen
erzielten KEIM et al. (1988) in einer nachfolgenden Untersuchung mit einer neuen
Gerätegeneration an sedierten Schweinen (n = 24, 48 - 137 kg und 14 - 45 % Fettgehalt)
hohe Korrelationen zwischen TOBEC-Messwerten (Phasendurchschnitt) und
Schlachtkörperdaten wie leeres Gesamtkörperwasser (r = 0,979) bzw. fettfreie Masse
des leeren Gesamtkörpers (r = 0,980) und leeres Gesamtkörper-Rohprotein (r = 0,962)
sowie leeres Gesamtkörper-Kalium (r = 0,949). Die Standardabweichungen der
Schätzungen (SEE) betrugen für leeres Gesamtkörperwasser 2,1 kg und für die fettfreie
Masse des leeren Gesamtkörpers 2,8 kg. Ähnlich gute Beziehungen ermittelten
FIOROTTO et al. (1987) an jungen Miniaturschweinen (10 - 33 Lebenstage). Das
TOBEC-Signal (korrigiert auf die Körperlänge) korrelierte hoch mit der chemisch
analysierten fettfreien Masse (r = 0,998) und dem Gesamtkörperwassergehalt

Methodenübersicht
97
(r = 0,998), während das 95 % Vorhersageintervall für individuelle Messungen ±0,16 kg
fettfreie Masse und ±0,12 Liter Wasser betrug. Die Konfidenzintervalle konnten
signifikant verbessert werden, indem auf die individuelle Variabilität im Verhältnis von
Körpermasse und Körperlänge, das heißt auf die Körpergeometrie korrigiert wurde
(±0,08 kg bzw. ±0,06 Liter). Obgleich TOBEC in der (amerikanischen)
Schlachtkörperbewertung bei Schwein bzw. Schaf und Rind eingesetzt wird (FORREST,
1994, MESECK et al., 1997, BERG et al., 1998 bzw. BERG et al., 1994 und GWARTNEY et
al., 1995), findet es in der Lebendtierbewertung aufgrund der bereits genannten
Probleme wie Bewegung der Tiere, Körpertemperatureinfluss, unterschiedliche
Geometrie der Tiere sowie Positionierung der Tiere wenig Beachtung (Übersicht bei
MITCHELL und SCHOLZ, 2001). So wurden auch bei der individuellen Analyse der
Körperzusammensetzung lebender Lämmer bzw. Broiler keine zufriedenstellenden
Genauigkeiten mit Hilfe der TOBEC-Messungen erzielt, da das Körpergewicht allein
einen größeren Anteil der Variation erklärte als die TOBEC-Messwerte (WISHMEYER et
al., 1994 bzw. DÄNICKE et al., 1997).
2.2.3.2.5.8 Infrarot-Körperfettanalyse (Infrarot-Interaktanz)
Diese Methode basiert auf dem Ausmaß der Reflektion bzw. Absorption (Transmission)
von Infrarotlicht einer bestimmten Wellenlänge (850 - 2600 nm) an geeigneten
Körperpositionen. Bei einer bestimmten Wellenlänge wird das infrarote Licht vom Fett
absorbiert und von der fettfreien Masse reflektiert. Sensoren messen die Intensität der
reflektierten Strahlung; die angeschlossene Computereinheit ermittelt die individuelle
Körperzusammensetzung.
Unter Zuhilfenahme von Merkmalen wie Körpergewicht, Geschlecht und
Körpertyp kann der Fettgehalt kalkuliert werden. Gegenüber einer einfachen, schnellen
und bequemen Anwendung steht noch immer die Ungenauigkeit dieser Methode, da die
Zusatzinformationen häufig den größten Teil der Variation erklären.
Das so genannte FUTREX-Messsystem, welches vereinzelt im Humanbereich
zum Einsatz gelangt, wurde am United States Department of Agriculture, Beltsville
Agriculture Research Center, Beltsville, MD entwickelt (RICO et al., 1994). Es besteht
aus einem Messkopf, in dem je nach Modell 4 bis 6 Elektroden integriert sind. Der
Messkopf kann an beliebigen anatomischen Positionen angesetzt werden, um
Infrarotlicht ins Gewebe zu senden. Am besten geeignet sind Körperpositionen, die eine

Methodenübersicht
98
große Variation im subkutanen Fettgehalt aufweisen. In einer ersten Studie bei Frauen
erzielten CONWAY et al. (1984) Korrelationskoeffizienten von r = 0,94 bzw. 0,90 sowie
0,89 zwischen Nah-Infrarot-Reflektions-Messwerten und D20-Verdünnungs-, bzw.
Hautdicken- sowie Ultraschall-Messwerten. Nachfolgende Studien ergaben jedoch
weniger Erfolg versprechende Ergebnisse und zweifeln den Wert dieser Methodik an
(POLITO et al., 1994, WILMORE et al., 1994, BROOKE-WAVELL et al., 1995, FLYNN et al.,
1995).
Beim Schwein weist die Rückenpartie über dem M.l.d. die höchste Variation im
subkutanen Fettgehalt innerhalb einer Population auf. MITCHELL et al. (1986)
überprüften das Verfahren sowohl an lebenden Schweinen als auch an Schlachtkörpern.
Zwischen chemischer Analyse und NIR (Nah-Infrarot-Reflektion) ergaben sich keine
Korrelationen, die größer als r = 0,75 ausfielen, wenn die NIR-Messwerte ohne
Zusatzinformation wie Lebendmasse in der Regressionsanalyse verwendet wurden.
Potentielle Probleme der Technik ergeben sich aus der Hautfarbe, Hautdicke und
Speckdicke selbst, da mit der verwendeten Technik nur eine Eindringtiefe von 1 cm
erreicht wurde. Neuere Untersuchungen zur Körperzusammensetzung am lebenden
Schwein mittels Nah-Infrarot-Interaktanz-Messung sind dem Autor nicht bekannt. Eine
potentielle Anwendung könnte in der Ermittlung der Fettqualität beim lebenden
Schwein liegen, da sich die Fettzusammensetzung im Laufe des Wachstums ändert und
eine Eindringtiefe von 1 cm für die Ermittlung der Fettqualität ausreichend sein müsste
(MITCHELL und SCHOLZ, 2001).
Als sehr gute Alternative zur chemischen Analyse eines weiten Spektrums von
Messkriterien in entsprechend aufbereiteten Proben findet die Nah-Infrarot-
Spektroskopie (Reflektion oder Transmission) inzwischen eine breite Verwendung.
Im Gegensatz zur Verwendung von Licht im infraroten Wellenlängenbereich
arbeitet das für humanmedizinische Zwecke entwickelte LIPOMETER mit Licht im
sichtbaren Wellenlängenbereich und dient zur Ermittlung der subkutanen Fettverteilung
bis zu einer Dicke von 50 mm (SUDI et al., 1998).
2.2.3.2.5.9 Videobildanalyse und Dreidimensionale Topometrie
Jüngste Entwicklungen in der Computer-Technik und digitalen Bildaufnahmetechnik
bilden die Voraussetzung für eine objektive Ermittlung der Körperproportionen und
–formen als ein Indikator für die Körperzusammensetzung von Schweinen. Diese

Methodenübersicht
99
Technik resultiert jedoch nicht in genauen Schätzwerten für die
Körperzusammensetzung lebender Tiere in Bezug auf Fett-, Protein-, Wasser- oder
Aschegehalt. Es sollte aber möglich sein, mit Hilfe der dreidimensionalen Topometrie
genaue Messungen des Körpervolumens vornehmen zu können (TSCHÜMPERLIN, 1996).
Dieses Messergebnis in Kombination mit linearen (digitalen) Messwerten wie
Brustweite, Rumpftiefe, Beckenweite und Widerristhöhe könnte die Grundlage für eine
objektive lineare Körperform- bzw. -konformationsbewertung bilden, die im
Zusammenhang mit dem Schlachtkörperwert steht, solange die Fleischqualität nicht das
Hauptkriterium darstellt (PATTERSON, 1990).
Gegenwärtig wird die Videobildanalyse - teilweise in Verbindung mit
zusätzlichen FOM-Messungen oder Ultraschallmessungen – für die Klassifikation von
Rinder-, Schweine-, Schaf- und Geflügelschlachtkörpern genutzt, um die
Körperkonformation, die Fettklasse, den Fleischanteil oder den Anteil wertvoller
(verkaufbarer) Teilstücke zu ermitteln. Die Bestimmtheitsmaße schwanken zwischen
0,3 und 0,9 mit Reststandardabweichungen zwischen 1,52 und 0,52 % (ALLEN und
WALSTRA, 1996, BRANSCHEID et al., 1995, 1998, BORGGARD et al., 1996, BRANSCHEID
und DOBROWOLSKI, 1996, HAHN u.a., 1997, STANFORD et al., 1998b, THOLEN u.a.,
1998, SÖNNICHSEN u.a., 2001). Weitere Versuche werden unternommen, um die
Videobildanalyse für die Erfassung der Fleischqualität, insbesondere der Marmorierung
bzw. des intramuskulären Fettgehalts, an definierten Schlachtkörperanschnitten zu
nutzen (ISHII et al., 1992, SCHOLZ u.a., 1995c, ENDER u.a., 1997b, KUCHIDA et al., 1998,
1999, 2000). Im Gegensatz zum Rind, scheitert speziell beim Schwein dieses Vorhaben
für routinemäßige Anwendungen bisher aufgrund der sehr geringen Variation des
Marmorierungsgrades bzw. bei extrem fettarmen Rassen aufgrund der nicht mehr
vorhandenen Marmorierung. Schwierigkeiten bei der Messung ergeben sich außerdem
durch auftretende Reflektionen an der Fleischoberfläche, die durch Wasseraustritt oder
die unebene Fleischoberfläche am Anschnitt verursacht werden. Abhilfe kann durch
spezielle Beleuchtungssysteme geschaffen werden.
2.2.3.2.5.10 Kalium40-Gammaspektrometrie - Analyse auf atomarem Niveau
Da Kalium nicht im Fettgewebe und sonst fast ausschließlich intrazellulär vorkommt,
kann die neben der Kalium39-Emission in begrenztem Ausmaß natürlich vorkommende

Methodenübersicht
100
Kalium40–Emission (0,0118 %) sowohl am lebenden Tier als auch am Schlachtkörper
mit Hilfe der K40–Flüssigkeits-Szintillation in einem Gesamtkörper-Zählgerät bestimmt
und der Fettgehalt über die Ermittlung der Gesamtkörperzellmasse bzw.
-magergewebemasse berechnet werden (SIEMENS, 1991, MITCHELL und SCHOLZ, 2001).
Kalium ist im Körper an der Steuerung verschiedener Funktionen beteiligt, wie:
• Wasser- und Elektrolythaushaltsregulierung,
• Reizübertragung an Muskeln und Nervenzellen sowie
• am Eiweißstoffwechsel.
Gleichzeitig besitzt das radioaktive Kaliumisotop K40 mit 4 500 Becquerel (Bq) den
hauptsächlichen Anteil an der natürlichen Radioaktivität des Menschen
(HTTP://WWW.NOEZSV.AT/WISSENHILFT/RADIOAKTIVITAET/WIRKUNGAUFMENSCH.HTM).
Ein Kilogramm Schweinefleisch (Frischmasse) emittiert auf der Basis des K-40-
Gehalts - bei einer Variation zwischen 87 und 130 Bq pro Tag und Person - ca. 110 Bq
pro Tag und Person. Ähnliche Werte wurden für Rindfleisch, Reh- sowie Hirschfleisch
ermittelt (HAHN, 2000). Die festgestellten Variationen sind eher auf unterschiedliche
Fett- und Wassergehalte der Fleischproben als auf die unterschiedliche regionale
Herkunft zurückzuführen.
Gesamtkörperzählgeräte fanden erstmals im Humanbereich Verwendung (FORBES,
1962, FORBES und HURSH, 1963, LLOYD und MAYS, 1987). Bereits im Jahr 1923 setzte
der Ungar GEORGE HEVESY (1943 Nobelpreis für Chemie) das erste Mal Radioaktivität
ein, um Pflanzen zu untersuchen. Er benutzte radioaktiv markiertes Blei, um die
Aufnahme dieser Substanz in die Pflanzen und die Verteilung des markierten Bleis zu
studieren. HEVESY zeigte mit seiner Arbeit, dass es mit kleinen Mengen von radioaktiv
markierten Elementen möglich ist, die Funktion von lebender Materie zu untersuchen,
ohne das Gewebe zu zerstören (SCHWAIGER, 1997).
Eine geeignete Kalibrierung, die Vermeidung von Hintergrundstrahlung und eine
Korrektur für die Körpergeometrie unter Berücksichtigung des Alters und des
Geschlechts ermöglichen eine genaue Ermittlung des Gesamtkörper-Kaliumgehaltes.
Aufgrund der sehr hohen Gerätekosten und der begrenzten Anwendbarkeit ist diese
Technik jedoch nicht weit verbreitet und wird nur in sehr wenigen Nutztierstudien zur
Ermittlung der Körperzusammensetzung herangezogen (SIEMENS et al., 1991, JIANG et
al., 2000). Im Vergleich zur chemischen Analyse fielen jedoch (wie bei den meisten
Messmethoden) die in vivo gemessenen K40-Emissions-Werte nicht so genau wie die

Methodenübersicht
101
post mortem gemessenen Werte aus, die eine Korrelation von r = 0,7 zwischen
Ganzkörper-K40-Gehalt und chemisch ermitteltem Kaliumgehalt des Schlachtkörpers
ergaben (SIEMENS et al., 1991). Bei totgeborenen Ferkeln ermittelten SPADY et al.
(1986) zwischen dem Gesamtkörper-K40-Gehalt aus der Gammaspektrometrie und dem
Gesamtkörper-Kaliumgehalt aus der Schlachtkörperanalyse eine Übereinstimmung von
durchschnittlich 97 %. JIANG et al. (2000) konnten keinen Unterschied zwischen mittels
DXA ermittelter relativer Magergewebemasse je kg Schlachtkörper und dem
Gesamtkörper-K40-Gehalt je kg fettfreier Masse beim Vergleich von zwei
Fütterungsgruppen (Plasmazusatz bzw. Kontrollfütterung) feststellen. Folglich stimmten
Zusammensetzung und Wassergehalt der Magergewebemasse für beide
Fütterungsgruppen überein. Absolut erhöhte sich im Vergleich zur Kontrolle die
Magergewebemasse in der mit Plasma gefütterten Gruppe signifikant, wobei keine
Unterschiede in den proportionalen Differenzen der beiden Fütterungsgruppen zwischen
DXA- und K40-Messung auftraten.
2.2.3.2.5.11 Neutronenaktivierung
Seit Jahrzehnten stellt die Neutronenaktivierungs-Analyse (NAA) ein Verfahren der
(Spuren-)Elementbestimmung dar, das in vielerlei Hinsicht einzigartig und
konkurrenzlos ist. Hierzu zählen die extrem hohe Genauigkeit und die „eingebaute
Selbstkontrolle". Ein chemischer Aufschluss der Probe ist prinzipiell nicht erforderlich
(KNIGHT et al., 1986, RYDE et al., 1993, SUTCLIFFE et al., 1993).
Das Prinzip der NAA beruht auf der Tatsache, dass die meisten Elemente einer
Probe (eines Individuums) bei der Bestrahlung mit (thermischen) Neutronen (n)
radioaktiv werden und eine charakteristische, durchdringende Gammastrahlung
aussenden. Bei einigen Stoßprozessen von Neutronen mit Atomkernen kommt es zur
Absorption der Neutronen (Neutroneneinfang). Bei diesem Vorgang werden radioaktive
Isotope erzeugt, die bei ihrem Zerfall eine für jedes Isotop charakteristische
Gammastrahlung (n’γ) aussenden. Auf diesem Prinzip beruht die „verzögerte“ (delayed)
Neutronenaktivierungs-Analyse. Die mit Neutronen bestrahlten Elemente wandeln sich
in instabile radioaktive Isotope um. Bei der „Rückkehr“ in einen stabilen Zustand
(Zerfall der radioaktiven instabilen Isotope bei entsprechender langer Halbwertszeit von
Sekunden, Minuten oder Stunden) lässt sich mittels geeigneter Detektoren die für jedes
Isotop spezifische Gamma-Strahlung messen und aus ihr über vergleichsweise einfache

Methodenübersicht
102
mathematische Korrelationen die in der Untersuchungsprobe vorliegende Konzentration
berechnen. Die Wahrscheinlichkeit für einen „Neutroneneinfang“ ist nur für relativ
langsame (thermische) Neutronen gegeben. Je höher der Neutronenfluss für diese
Aktivierung, desto empfindlicher ist das Verfahren und desto geringere Mengenanteile
können noch identifiziert und quantifiziert werden (KEHAYIAS und VALTUEÑA, 1999).
Zukünftig können großvolumige Proben nach der Neutronenaktivierung durch
ortsauflösende Gamma-Spektrometrie bzw. eine Kombination von Neutronen-
aktivierung und Kernresonanzabsorption tomographisch vermessen werden, so dass
analog der beschriebenen Bildgebungsverfahren die quantitativen Messergebnisse
visualisiert werden können (LIERSE, 1998, VARTSKY et al., 2000). Anders als bei der
Röntgen-Computer-Tomographie existieren bei der Neutronen-Tomographie
Wechselwirkungen mit den Atomkernen der Probe. An den Atomkernen können die
Neutronen absorbiert (eingefangen) oder gestreut werden. Die Wahrscheinlichkeit für
eine Wechselwirkung hängt empfindlich vom inneren Aufbau der Atomkerne ab. Dabei
existiert keine vergleichbar einfache Regelmäßigkeit wie für Röntgenstrahlen. Für
unterschiedliche Isotope eines Elements gibt es zum Teil stark voneinander
abweichende Wechselwirkungen. Neutronen werden besonders an Wasserstoff stark
gestreut. Hingegen treten Röntgenstrahlen in eine Wechselwirkung mit den Elektronen,
die sich in der Atomhülle befinden. Das bedeutet, wenn die zu untersuchenden
chemischen Elemente bzw. Körpergewebe elektronenreich sind, wie z.B.
Knochengewebe, werden die Röntgenstrahlen stärker geschwächt als in Muskel- oder
Fettgewebe.
Neutronenaktivierung ist die einzige Methode, die eine simultane Analyse
mehrerer Elemente des Gesamtkörpers auf atomarem Niveau zulässt. Alle wichtigen
Elemente, die im Körper vorkommen wie Wasserstoff, Kohlenstoff, Stickstoff,
Sauerstoff, Kalzium, Phosphor, Natrium und Chlor können mit dieser Methodik
quantifiziert werden. Kalium wird mit Hilfe der speziellen K40-Gammaspektroskopie
bestimmt (Kapitel 2.2.3.2.5.10). Geräte, die für die (In-vivo-)Neutronenaktivierungs-
Analyse verwendet werden, sind (noch) sehr aufwendig und teuer, benötigen ein
spezialisiertes Bedienpersonal und sind zudem relativ langsam (15 – 60 Minuten).
Gegenwärtig werden Anstrengungen unternommen, um die kommerzielle Entwicklung
eines portablen Neutronengenerators für Großtieranwendungen voranzutreiben (MITRA
et al., 1998). Zahlreiche In-vivo-Studien unter Zuhilfenahme der Neutronen-

Methodenübersicht
103
aktivierungs-Analyse existieren im humanen Bereich (SIWEK et al., 1984, COHN, 1992,
SUTCLIFFE et al., 1993, HEYMSFIELD et al., 1997a, NATTO et al., 1998, KEHAYIAS und
VALTUEÑA, 1999), während im Bereich der Nutztiere diese Technik vergleichbar wenig
zum Einsatz gelangte. Dabei wurden Gewebeproben (Homogenate) von (nicht näher
definiertem) Fleisch (MITRA et al., 1995a), Geflügel (KRUGER und GRUVERMAN, 1962),
Schaf, z.T. unter Zusatz von Rinderfett (MITRA et al., 1995b) sowie (komplette)
Schlachtkörper von Fisch (TALBOT et al., 1986) und Ratte (PRESTON et al., 1985) bzw.
(zum Teil lebende) Probanden von Miniatur-Schweinen (ABRAMS et al., 1988),
Meerschweinchen (SHENG et al., 1981, ) oder Ratte (YASUMURA et al., 1993, 1998)
untersucht. Geplant sind Untersuchungen an jungen wachsenden Schafen und
Schweinen (MITRA et al., 1998).
Für die interessierenden Elemente existieren verschiedene spezielle Messsysteme
(Abb. 2.2.32).
Abb. 2.2.32: Reaktionen der Neutronenaktivierung zur Ermittlung der elementarenKörperzusammensetzung (modifiziert nach KEHAYIAS und VALTUEÑA, 1999)
Der Gesamtkörperstickstoffgehalt kann mit der sogenannten „Prompt-Gamma-
Technik“ oder „prompten“ Neutronenaktivierung ermittelt werden (KEHAYIAS und

Methodenübersicht
104
VALTUEÑA, 1999). Die Aktivierung des Isotops 14N zum Isotop 15N erfolgt mittels
3,5 MeV Neutronenbestrahlung. Das radioaktive Isotop 15N emittiert bei der Rückkehr
zum 14N-Stadium eine charakteristische 10,83 MeV-Gammastrahlung sowie ein im
Spektrum als Standard dienendes Wasserstoffsignal bei 2,223 MeV. In diesem Fall
dienen Natriumjodid-Detektoren zur Quantifizierung der charakteristischen
Gammastrahlung mit einer Genauigkeit für die Stickstoffanalyse von 97 % (VARTSKY et
al., 1979). Im Gegensatz zur „prompten“ Neutronenaktivierung dient die „verzögerte“
Neutronenaktivierung zur Ermittlung des Gesamtkörper-Kalziumgehalts, welcher eine
exakte Aussage zur Knochenmineralmasse ermöglicht, da 99 % des Gesamt-
körperkalziums im Knochen vorkommen (COHN, 1992). Neben Kalzium werden
Gesamtkörper-Natrium, -Chlor und -Phosphor aktiviert und sind folglich ebenfalls
messbar (COHN, 1992, KEHAYIAS und VALTUEÑA, 1999).
Das Verfahren der „inelastischen“ Neutronenstreuung∗ dient der Messung des
Kohlenstoff- und Sauerstoffgehalts (KEHAYIAS et al., 2000). Die „inelastische“
Neutronenstreuung ist eine der leistungsfähigsten Methoden zur Untersuchung
dynamischer Prozesse in „kondensierter“ Materie. Sie erlaubt es, Bewegungsvorgänge
gleichzeitig sowohl zeitlich als auch räumlich aufzulösen, da (thermische) Neutronen im
Unterschied zu Elektronen oder elektromagnetischer Strahlung sowohl Wellenlängen in
der Größenordnung von Atomabständen als auch Frequenzen im Bereich typischer
Gitterschwingungs- oder Platzwechselfrequenzen besitzen. Darüber hinaus zeichnen
sich Neutronen durch eine hohe Durchdringungsfähigkeit von Materie aus, so dass
fundamentale Volumeneigenschaften innerhalb des Untersuchungsobjektes analysiert
werden können (http://www.frm2.tu-muenchen.de/d/instrumente/puma/rahmen.html).
Beim Verfahren der „inelastischen“ Neutronenstreuung wird der Proband im
Rahmen eines Gesamtkörper-Scans einer Neutronenbeschleuniger-Quelle ausgesetzt.
KEHAYIAS et al. (1991, 2000) verwendeten am Menschen einen Miniatur-Deuterium-
Tritium-Neutronengenerator mit einer Strahlenbelastung < 0,06 mSv. Die generierten
schnellen Neutronen treten durch inelastische Kollisionen in Wechselwirkung mit der
Materie, was zu einem sofortigen (prompten) Kernzerfall und der Emission von
Gammastrahlung (n→n'γ) führt. Die vom Kohlenstoff ausgehende 4,44 MeV-Strahlung
kann entweder von Bismutgermanat- oder Natriumjodidkristallen, die an jeder Seite des
Untersuchungsobjektes angeordnet sind, erfasst werden. Die Wiederholbarkeit für die
∗ Ein Dreiachsenspektrometer (PUMA) zur wissenschaftlichen Arbeit mit der „inelastischen Neutronenstreuung“ wird

Methodenübersicht
105
Kohlenstoffbestimmung beträgt ca. 97% (KEHAYIAS et al., 1991). Anschließend kann
der Körperfettgehalt berechnet werden, indem empirische Formeln für den
Kohlenstoffgehalt von Protein, Fett, Glykogen und Knochen verwendet werden (siehe
Kapitel 2.2.2). Ein Sauerstoffsignal erscheint bei 6,13 MeV im Spektrum. Das
Kohlenstoff-zu-Sauerstoff-Verhältnis dient zusätzlich zur Berechnung der Verteilung
von Fett im Körper (KEHAYIAS et al., 2000).
Eine weitere Neutronenaktivierungstechnik im Zusammenhang mit Methoden zur
Bestimmung der Körperzusammensetzung ist die „schnelle“ Neutronenaktivierung oder
sogenannte „associated particle technique“ (MITRA et al., 1995a,b). Während der
Erzeugung eines instabilen Isotops durch energiereiche, schnelle Neutronen werden
zusätzlich einzelne Fragmente bzw. Partikel, wie z.B. zwei Neutronen (n→2n), ein
Alpha-Partikel (n→α) oder ein Proton (n→p), aus dem bestrahlten Element
herausgetrennt (KEHAYIAS und VALTUEÑA, 1999). Dieses Verfahren nutzt 14 MeV
Neutronen und Natriumjodidkristalle zur Erzeugung von Gammastrahlungs-Spektren,
die Signale von Kohlenstoff, Sauerstoff und Stickstoff enthalten. Basierend auf Scans
von homogenisiertem (Schaf-)Fleisch (+ Rinderfett) ergab sich eine Präzision von 4,1 %
bzw. 5,4 % und 1,2 % (Variationskoeffizienten) innerhalb eines 15 minütigen 14 MeV-
Scans für die Messung des Protein- bzw. Fett- und Wassergehalts einer ca. 41 kg
schweren Fleischprobe (Werte aus MITRA et al, 1995a,b). Das durch die schnellen
Neutronen hervorgerufene Strahlendosisäquivalent beträgt etwa 0,03 mSv.
Verwendet man zur Messung von Gesamtkörper-Sauerstoff, -Stickstoff und
–Kohlenstoff anstelle der Neutronenquelle eine Röntgenquelle zur Aktivierung der
Positronen emittierenden Isotope 15O, 11C und 13N, dann spricht man von
Photonenaktivierung (ULIN und ZAMENHOF, 1986).
2.2.3.3 STOFFWECHSELANALYSE
2.2.3.3.1 Verdünnungsmethoden
Der Körperfettgehalt kann ebenfalls mit verschiedenen Indikatorverdünnungsmethoden
bestimmt werden: Zur Bestimmung des Körperwassers, das sich nur in der fettfreien
Masse befindet, können „Schweres Wasser“ (Deuterium-Oxid), radioaktives Tritium-
Oxid, Harnstoff, Ethanol sowie in geringerem Maße das stabile Sauerstoffisotop 18O,
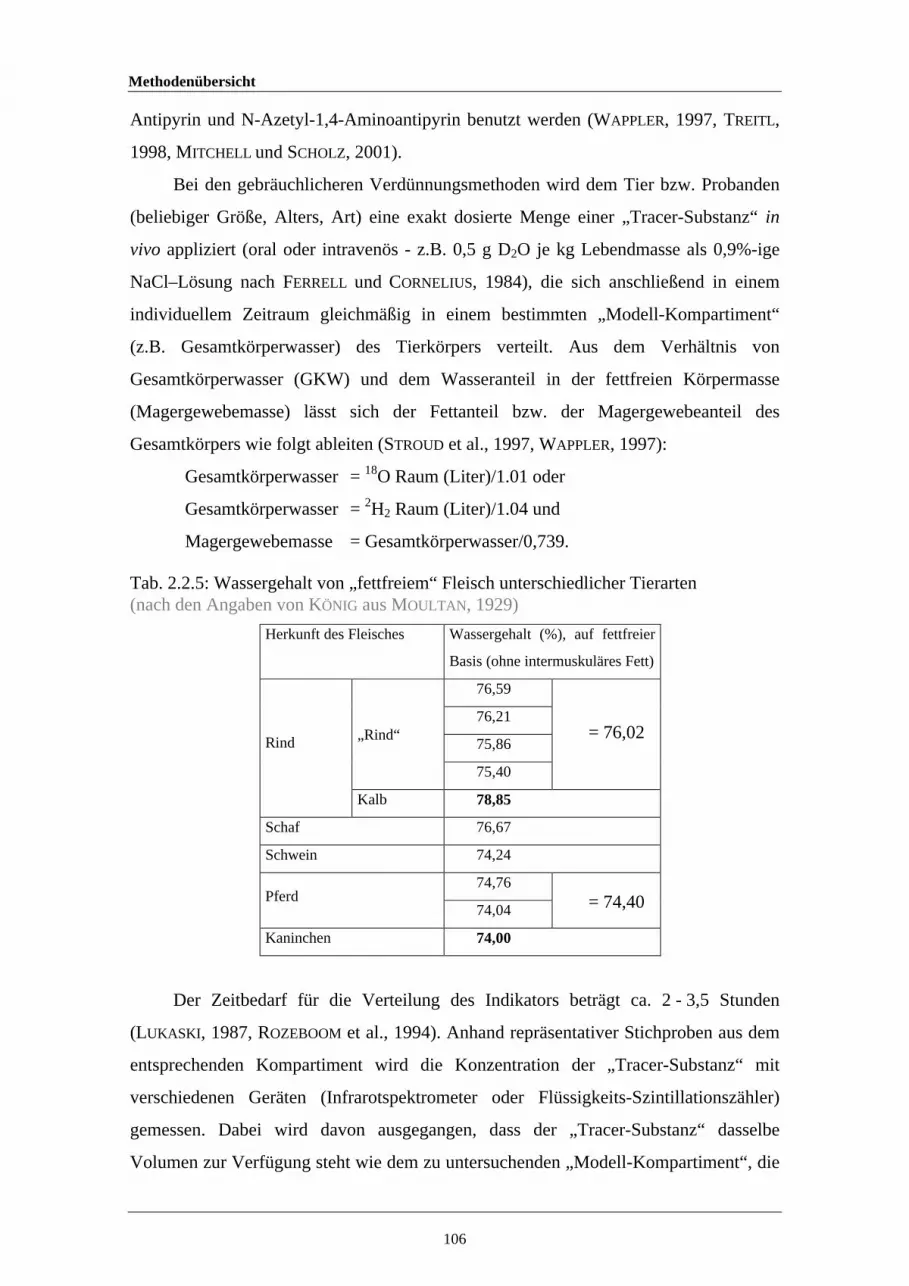
Methodenübersicht
106
Antipyrin und N-Azetyl-1,4-Aminoantipyrin benutzt werden (WAPPLER, 1997, TREITL,
1998, MITCHELL und SCHOLZ, 2001).
Bei den gebräuchlicheren Verdünnungsmethoden wird dem Tier bzw. Probanden
(beliebiger Größe, Alters, Art) eine exakt dosierte Menge einer „Tracer-Substanz“ in
vivo appliziert (oral oder intravenös - z.B. 0,5 g D2O je kg Lebendmasse als 0,9%-ige
NaCl–Lösung nach FERRELL und CORNELIUS, 1984), die sich anschließend in einem
individuellem Zeitraum gleichmäßig in einem bestimmten „Modell-Kompartiment“
(z.B. Gesamtkörperwasser) des Tierkörpers verteilt. Aus dem Verhältnis von
Gesamtkörperwasser (GKW) und dem Wasseranteil in der fettfreien Körpermasse
(Magergewebemasse) lässt sich der Fettanteil bzw. der Magergewebeanteil des
Gesamtkörpers wie folgt ableiten (STROUD et al., 1997, WAPPLER, 1997):
Gesamtkörperwasser = 18O Raum (Liter)/1.01 oder
Gesamtkörperwasser = 2H2 Raum (Liter)/1.04 und
Magergewebemasse = Gesamtkörperwasser/0,739.
Tab. 2.2.5: Wassergehalt von „fettfreiem“ Fleisch unterschiedlicher Tierarten(nach den Angaben von KÖNIG aus MOULTAN, 1929)
Herkunft des Fleisches Wassergehalt (%), auf fettfreier
Basis (ohne intermuskuläres Fett)
76,59
76,21
75,86„Rind“
75,40
∅ = 76,02Rind
Kalb 78,85
Schaf 76,67
Schwein 74,24
74,76Pferd
74,04 ∅ = 74,40
Kaninchen 74,00
Der Zeitbedarf für die Verteilung des Indikators beträgt ca. 2 - 3,5 Stunden
(LUKASKI, 1987, ROZEBOOM et al., 1994). Anhand repräsentativer Stichproben aus dem
entsprechenden Kompartiment wird die Konzentration der „Tracer-Substanz“ mit
verschiedenen Geräten (Infrarotspektrometer oder Flüssigkeits-Szintillationszähler)
gemessen. Dabei wird davon ausgegangen, dass der „Tracer-Substanz“ dasselbe
Volumen zur Verfügung steht wie dem zu untersuchenden „Modell-Kompartiment“, die

Methodenübersicht
107
„Tracer-Substanz“ nicht toxisch ist, und - im Fall von Wasser - identisch durch den
Körperstoffwechsel ausgetauscht wird (WAPPLER, 1997, TREITL, 1998).
Verdünnungsmethoden zur Analyse der Körperzusammensetzung basieren in der
Regel ebenso wie die densitometrischen Methoden auf dem bereits beschriebenen
Prinzip, dass das Gesamtkörperwasser mit ca. 74 % bei einem Variationskoeffizienten
von 2 % eine anteilmäßig fixierte Fraktion der fettfreien Masse in ausgewachsenen
Säugetieren (einschließlich Mensch) darstellt. Im Laufe des Wachstums reduziert sich
der Anteil des Gesamtkörperwassers an der fettfreien Masse von ca. 81% bei Geburt auf
die oben erwähnten 74 % [73,9 %] (WANG et al., 2000). Besonders bei Nutztieren, die
sich während der meisten Untersuchungen noch in der Wachstumsphase befinden, kann
nicht davon ausgegangen werden, dass das Gesamtkörperwasser eine fixierte Fraktion
der fettfreien Masse darstellt (SHIELDS et al., 1983). Bereits die Untersuchungen des
deutschen Forschers KÖNIG (Tab. 2.2.5) zeigten, dass der Wassergehalt in der fettfreien
Masse (Fleisch ohne intermuskuläres Fett) tierart- und altersabhängig zwischen 74,0 %
und 78,75 % variiert (MOULTAN, 1929). MOULTAN (1929) selbst bestätigte anhand einer
Vielzahl von Fleischproben diese Variation im Wassergehalt der fettfreien Masse, die
zum Teil durch die (nicht untersuchten) Abweichungen im intramuskulären Fettgehalt
verursacht werden könnte. Die Ergebnisse von LAUBE (2000) unterstreichen diese
Beobachtung nochmals. Der Wassergehalt des Koteletts zwischen 14./15. Rippe
variierte zwischen 74,10 und 75,96 % in Abhängigkeit vom intramuskulären Fettgehalt
und der genetischen Herkunft der Endstufeneber zur Erzeugung der untersuchten
Mastschweine. Der niedrigste Wassergehalt trat bei den Tieren der Rasse Duroc (NN)
bei gleichzeitig höchstem intramuskulärem Fettgehalt auf, während erwartungsgemäß in
der Tendenz die Tiere mit dem niedrigsten intramuskulärem Fettgehalt (Pietrain x
Hampshire bzw. Pietrain) den höchsten Wassergehalt aufwiesen. Insgesamt ist
hervorzuheben, dass bereits Mitte der 20er Jahre speziell für Schwein, Pferd und
Kaninchen der relativ konstante Wassergehalt in der fettfreien Masse mit ca. 74 %
ermittelt wurde. Leider gibt es bei MOULTAN (1929) keine Angaben über genaues Alter
und Gewicht der untersuchten Tiere. Allein beim Vergleich innerhalb der Tierart Rind
ist über die Unterscheidung von Rind und Kalb ein Altersvergleich nahe liegend, der in
der Tendenz zeigt, dass im wachsenden Tier (Kalb) ein höherer Wassergehalt in der
fettfreien Masse vorliegt als in den (ausgewachsenen) Rindern (Tab. 2.2.5).

Methodenübersicht
108
Die Verwendung von Deuterium-Oxid bzw. radioaktivem Tritium-Oxid führt
beim Schwein in Abhängigkeit vom Wachstumsstadium bzw. in Abhängigkeit vom
Reproduktionsstadium zu Über- bzw. Unterschätzungen der Gesamtkörper-
wassermenge, des Fettgehalts bzw. des Proteingehalts. Nur durch die Einbeziehung der
Lebendmasse bzw. der Rückenspeckdicke zusätzlich zur D2O-Konzentration können
genaue Schätzwerte für die Körperzusammensetzung innerhalb einer definierten
Population erzielt werden (FERRELL und CORNELIUS, 1984, ROZEBOOM et al., 1994).
FULLER et al. (1996) analysierten die Anwendbarkeit einer Vielzahl (> 50) von
experimentell ermittelten Regressionsgleichungen für verschiedene Techniken im
Vergleich zum mittels Deuterium- und 18O-Verdünnungsmethoden bestimmten
Gesamtkörperwasser und gelangten zu dem Schluss, dass ein Teil der Gleichungen unter
bestimmten Bedingungen – speziell bei älteren Probanden - mit „Vorsicht“ zu
behandeln ist.
Noch größer sind die Abweichungen bei Verwendung der Harnstoff-
Verdünnungsmethode, die zusätzlich zu den Effekten Wachstum, Reproduktions-
stadium, Verteilungszeit und Zeit der Probennahme durch die Rationsgestaltung sowie
die gemessene Basiskonzentration von Harnstoff im Blutplasma beeinflusst werden
kann. Die Bestimmtheitsmaße für Deuterium-Oxid bzw. radioaktives Tritium-Oxid und
Harnstoff variieren zwischen 0,29 und 0,9. (Übersicht bei MITCHELL und SCHOLZ,
2001).
Alternativ zu den „klassischen“ Verdünnungsmethoden wird bei HENEN (1991) über die
absorbierte Zyklopropangasmenge eines Individuums (Schildkröten, Ratten) bzw. bei METTAU et al.
(1977) über die absorbierte/desorbierte Xenongasmenge eines Individuums (Meerschweinchen,
Neugeborene) direkt der Fettgehalt (Lipidgehalt) ermittelt.
2.2.3.3.2 Kreatininausscheidung
Eine klassische Methode zur Schätzung der Skelettmuskelmasse des Gesamtkörpers ist
die Messung der Kreatininausscheidung mit dem Urin über einen Zeitraum von 24
Stunden. Kreatinin wird durch eine nicht-enzymatische Hydrolyse von intrazellulärem
Kreatin und Phosphokreatin erzeugt und als Stoffwechselendprodukt hauptsächlich über
die Niere ausgeschieden. Etwa 98 % des Kreatins im Körper befindet sich in der
Skelettmuskulatur – hauptsächlich als Phosphokreatin. Phosphokreatin dient im Muskel
als Energiespeicher. Somit ist die Gesamtmenge des Kreatins abhängig von der Gesamt-

Methodenübersicht
109
Muskelmasse. Untersuchungen am Menschen zeigten eine Beziehung zwischen der
Höhe der Kreatininausscheidung mit dem Urin und der Muskel- bzw. fettfreien Masse
(FORBES und BRUINING, 1976, HEYMSFIELD et al., 1983). Gleichzeitig wurde jedoch
eine sehr hohe individuelle Variation in der täglichen Kreatininausscheidung
festgestellt. Genetische und physiologische Faktoren wie Alter, Geschlecht,
Reproduktionsstadium (Trächtigkeit), Aktivitätsniveau sowie die Nierenfunktion
können die Beziehungen zwischen fettfreier Masse (Skelettmuskelmasse) bzw. den
Stoffwechselparametern Harnsäure, Blutzucker, Cholesterin oder Triglyzeride und der
Menge ausgeschiedenen Kreatinins beeinflussen (FORBES und BRUINING, 1976,
HEYMSFIELD et al., 1983, PORSCH-ÖZÇÜRÜMEZ, 1997). Ältere Individuen besitzen
niedrigere Kreatininausscheidungswerte als jüngere Individuen. Zu niedrige
Kreatininwerte können auf eine zu geringe Muskelmasse oder eine Trächtigkeit
(Schwangerschaft) hindeuten. Erhöhte Werte zeigen sich bei Niereninsuffizienz oder
Nierenschwäche sowie nach Zerstörung von Muskelmasse wie z.B. nach Quetschungen
oder bei Muskeldystrophie.
Alternativ bzw. ergänzend zur Kreatininausscheidung kann auch die 3-Methylhistidin-
Ausscheidung (über 24 Stunden) zur Ermittlung der Gesamtkörperskelettmuskelmasse
verwendet werden (ELIA et al., 1979, RIKIMARU et al., 1989, WANG et al., 1998c), wobei
Kreatinin offenbar eine höhere Beziehung zur Skelettmuskelmasse aufweist als 3-
Methylhistidin (VIRGILI et al., 1994). Sowohl die Kreatinin- als auch die 3-
Methylhistidinausscheidung können durch verschiedene Ernährungsstrategien
beeinflusst werden und variieren zusätzlich im zeitlichen Tagesablauf, so dass eine
relativ verlässliche Aussage über die Körperzusammensetzung nur unter
standardisierten und nachvollziehbaren Bedingungen erreicht werden kann.

Methodenübersicht
110
2.3 Ergänzende (invasive) Methodik zur Bestimmung von Muskelfasermerkmalen
mittels Biopsie
Für verminderte Belastbarkeit der Schweine und mangelnde Fleischbeschaffenheit wird
der Muskelstruktur große Bedeutung zugemessen. Dabei nehmen die Muskelfasern als
Ort der Proteinsynthese eine zentrale Stellung ein. Insbesondere die Qualität des
Fleisches hängt von den strukturellen und funktionellen Eigenschaften der Muskulatur
ab. Muskelfasergesamtzahl, Fasertypenanteile und Faserdurchmesser stehen in einem
ursächlichen Zusammenhang mit Fleischansatz, Fleischbeschaffenheit und
Stressempfindlichkeit. Zusätzlich können mit der Bestimmung der Glykogen- und pH-
Werte sowie der ATP-Abbauintensität die Unterschiede im Muskelenergiestoffwechsel
von Schweinen unterschiedlicher Belastungsanfälligkeit erfasst werden (VERBURG et al.,
1984, WICKE u.a., 1991, FIEDLER u.a., 1993, 2001a,b LENGERKEN et al., 1994, 1997,
KLONT et al., 1994, KLONT and LAMBOOY, 1995, KARLSSON et al., 1999).
Da die in den vorangegangenen Kapiteln beschriebenen Verfahren zur Ermittlung
der Körperzusammensetzung nicht in der Lage sind, die Muskelfaserzusammensetzung
am lebenden Tier zu bestimmen, ergänzt die invasive Muskelbiopsie diese Reihe der
Verfahren. Verschiedene Ausführungen von Biopsiegeräten dienen zur Probenentnahme
entweder von Hand, per Druckluft oder explosiv (SCHÖBERLEIN, 1976, VERBURG et al.,
1984, CHEAH et al., 1995, KLONT et al., 1994, KLONT and LAMBOOY, 1995, LAHUCKY et
al., 1995). Letzteres Verfahren wird beim bewährten Schussbiopsie-Gerät von
SCHÖBERLEIN (1976) genutzt (Abb. 2.3). Es basiert auf einem modifizierten
Bolzenschussgerät zur Betäubung von Tieren und kann erfolgreich zur Probenentnahme
in vivo eingesetzt werden (VERBURG et al., 1984, FIEDLER u.a., 1993, MAAK et al.,
1995).
Zur extrem schnellen und sauberen Probenentnahme ist das Gerät mit
Biopsienadeln unterschiedlichen Durchmessers (z.B. 8 oder 11 mm) zu bestücken. Die
Hohlnadeln enthalten im Inneren des Edelstahlzylinders kleine Metall-Flügel, die sich
beim Eindringen in das zu entnehmende Gewebe öffnen und beim Rückstoß wieder
schließen, um ein sauberes Heraustrennen der Muskelprobe zu gewährleisten. Die
Biopsie geht im Zeitraum eines Sekundenbruchteils vonstatten, wodurch die Belastung
der Tiere sehr gering gehalten werden kann.

Methodenübersicht
111
Abb. 2.3: Montiertes Schussbiopsiegerät LS1 nach SCHÖBERLEIN (1976)
Zur Weiterverarbeitung müssen die Muskelproben je nach Verwendungszweck
möglichst auf Trockeneis aufbereitet und sofort in flüssigem Stickstoff oder einem
anderen Tiefkühlmedium (Isopentan in flüssigem Stickstoff) schockgefroren werden,
um sämtliche Stoffwechselvorgänge und Strukturmerkmale zu „konservieren“.
Anschließend erfolgt eine Lagerung bis zur Weiterverarbeitung bei –70 bis –80 °C.
Sollen die Proben einer histochemischen Untersuchung dienen, besteht der
nächste Schritt in der Anfertigung von Mikro-Gefrierschnitten, die je nach Analyseziel
unterschiedlichen Färbeprozeduren (chemischen Reaktionen) unterworfen werden
(SOLOMON and DUNN, 1988, FIEDLER u.a., 1993, 2001a, KARLSSON et al., 1999,
SCHOPPMEYER u.a., 2001). Unabhängig von der verwendeten histochemischen Methode
werden drei Hauptfasertypen unterschieden. Diese werden als Typ I (im englischen
Sprachraum häufig als: „slow-twitch-oxydative - STO), Typ IIa, („fast-twitch-
oxydative“ - FTO) und Typ IIb („fast-twitch-glycolytic“ - FTG) bezeichnet. Die
Eigenschaften der Muskelfasertypen gehen aus Abb. 3.1.21 (Kapitel 3.1.5) hervor.
Zusätzlich wird in einigen Arbeiten ein Typ IIc oder Typ IIx definiert, der eine
Übergangsform zwischen Typ IIa und Typ IIb darstellt und vor allem in neonatalen
Individuen bzw. während des Heilungsprozesses nach Muskelverletzungen auftritt
(KARLSSON et al., 1999). Neben den „normalen“ Muskelfasern werden in Abhängigkeit
von ihrer Größe und Form zum Teil so genannte Riesenfasern und anguläre
(degenerierte) Fasern unterschieden, die zumindest teilweise nur post mortem

Methodenübersicht
112
nachgewiesen werden können (SWATLAND, 1994, FIEDLER u.a., 2001b, SCHOPPMEYER
u.a., 2001).
Die Anteile und die Größe der einzelnen Fasertypen variieren zwischen den
einzelnen Muskeln in Abhängigkeit von ihrer Funktion, physischer Belastung, Alter,
Geschlecht, Hormonhaushalt (pST), Rasse und Typ (fett oder mager), während die
Muskelfaseranzahl ab dem Zeitpunkt der Geburt stabil bleibt. Eine
Volumenvergrößerung der Muskeln während des postnatalen Wachstums erfolgt
demzufolge allein durch Muskelhypertrophie. Generell kann davon ausgegangen
werden, dass Muskeln, die der Bewegung dienen (speziell an Beinen und Schulter),
mehr dem roten (oxidativen) Muskelstoffwechseltyp zuzuordnen sind und Muskeln, die
das Skelett stabilisieren (speziell im Rückenbereich), mehr dem weißen
(glykolytischem) Muskelstoffwechseltyp entsprechen (SALOMON u.a., 1981, 1983,
BADER, 1982, SOSNICKI, 1987, SEIDEMANN et al., 1989, SOLOMON et al., 1990,
LEFAUCHEUR et al., 1992, ESSEN-GUSTAVSSON, 1993, 1995, LENGERKEN et al., 1994,
1997, XIONG, 1994, RUUSUNEN and POULANNE, 1997, KARLSSON et al., 1999, FIEDLER
u.a., 2001a). Eine Unterernährung der Föten im Uterus hat im Vergleich zu Föten von
ausgewogen ernährten Sauen eine verminderte Anzahl Muskelfasern zum Zeitpunkt der
Geburt zur Folge (DWYER et al., 1994). Abgesehen von Muskelerkrankungen (VAN
LOGTESTIJN und SYBESMA, 1967), könnte unter „normalen“ (physiologischen)
Wachstumsbedingungen eine Futterrestriktion über einen sehr langen Zeitraum zu einer
Verminderung der Faseranzahl führen (SALOMON u.a., 1988). Beim Menschen ist jedoch
mit zunehmendem Alter in einigen Muskeln ein signifikanter Rückgang der Faseranzahl
zu verzeichnen (CARTEE, 1995).

Eigene Untersuchungen
113
3 Eigene Untersuchungen3.1 Material und Methodik
3.1.1 Muskelstoffwechseluntersuchungen3.1.1.1 31P-Magnet-Resonanz-Spektroskopie
Die Stoffwechseluntersuchungen zu den Phosphorkomponenten mittels 31P-Magnet-
Resonanz-Spektroskopie wurden an 69 Schweinen (15 männlich, 8 männliche
Kastraten, 46 weiblich) mit Lebendmassen zwischen 6,14 und 14,96 kg (∅ 10,19 ± 2,12
kg) bei einem Alter von 28 bis 63 Tagen (∅ 44,78 ± 8,52) durchgeführt.
Der größte Teil der Tiere wurde aus der Versuchsstation der Iowa State University
bezogen (29 Duroc-Linie, 12 Hampshire-Linie und 5 Spotted Poland China). Diese
Tiere stammten aus einem Anpaarungsprogramm zur speziellen Erzeugung der drei
Ryanodin-Rezeptor-1-Genotypen. Ein kommerzieller Schweineproduzent aus Maryland
lieferte 14 reinrassige Schweine der US-Landrasse, während 9 Kreuzungstiere aus der
Verpaarung von Poland China und Landrasse in der USDA-ARS-Versuchsstation des
Livestock and Poultry Science Institute in Beltsville, Maryland, erzeugt wurden.
Für alle spektroskopischen und tomographischen Untersuchungen von Tieren bis
zu maximal 30 kg Lebendmasse stand ein horizontaler 4,7 Tesla Magnet (Varian Inc.,
Palo Alto, Cal.) mit 33 cm nutzbarem Innendurchmesser zur Verfügung (Abb. 2.1.1).
Zehn Minuten vor dem Spektroskopie-Experiment wurden die Tiere unter
Verwendung einer Mixtur von Ketamin (100 mg/10 kg Körpergewicht), Telazol and
Xylazin (50 mg + 50 mg/10 kg Körpergewicht) sediert und auf der rechten Seite liegend
in einer Versuchswiege platziert. Danach wurde auf der Haut über dem M. biceps
femoris eine im Durchmesser 5,8 cm große, doppeltgewickelte Oberflächenspule fixiert
(Abb. 3.1.1).

Eigene Untersuchungen
114
Abb. 3.1.1: Platzierung der 31P-MR-Oberflächenspule über dem M. biceps femoris einesSchweines
Bei einer Frequenz von 80,984 MHz wurden die 31P-Spektren mit einem Abstand
von 57,5 s bei einer Aufnahmedauer (acquisition time) von 0,384 s pro FID (Free
Induction Decay), einer Wiederholungszeit (TR= time of repetition) von 7,184 s und 8
Wiederholungen (transients) erfasst. Die Spektralweite wurde auf 4000 Hz festgelegt.
Für jedes Tier wurden zusätzlich zur Dauer der Halothanzufuhr (maximal 60 Minuten
bei 3 Volumen-% pro 3 Liter O2/Minute) 10 Spektren vor der Halothanzufuhr im
“Ruhezustand” und 60 Spektren nach Beendigung der Halothangabe während der
Erholungsphase aufgenommen. Für ein Tier betrug die maximale Versuchsdauer
demzufolge ca. 130 Minuten (130 Spektren). Die Halothanzufuhr wurde vorzeitig
beendet, wenn ein signifikanter Abfall der Phosphokreatin-Konzentration bzw. ein
Verhältnis Phosphokreatin zu anorganischem Phosphat (PCr/Pi) von < 7,0 beobachtet
wurde. Das durchschnittliche PCr/Pi-Verhältnis variierte zwischen 11 und 25 zu Beginn
der Spektroskopieversuche. Im Verlauf des Experiments wurden in vivo und zum Teil
anschließend post mortem die prozentualen Konzentrationsveränderungen von
Phosphokreatin (PCr), Adenosintriphosphat (ATP), anorganischem Phosphat (Pi) sowie
der pH-Wert- und der Körpertemperaturverlauf erfasst, wobei die Spektren mit Hilfe der
Varian-NMR-Software (VNMR 4.1B, 1992) unter dem Betriebssystem SunOS 4.1.2
ausgewertet wurden.

Eigene Untersuchungen
115
Nach einer “Baseline”-Korrektur wurden die Signalintensitäten (peak heights) und
die relativen Positionen im Spektrum (ppm) zur Berechnung der prozentualen
Konzentrationsveränderung von PCr, Pi, ATP sowie des pH-Niveaus verwendet. MOON
and RICHARDS (1973) berichteten erstmals von der Ermittlung des intrazellulären pH-
Wertes mittels 31P-MR-Spektroskopie bei intakten roten Blutkörperchen.
Anorganisches Phosphat (Pi) besitzt eine definierte Resonanz im 31P-MR-
Spektrum lebender Zellen und Gewebe (ELLINGTON, 1989). Unter physiologischen
Bedingungen liegt Pi bei einem pKa-Wert (mit K = Gleichgewichts- bzw. Dissoziations-
konstante) von 6,7 – 6,8 als Dihydrogenphosphat [H2PO4-] und Hydrogenphosphat
[HPO42-] vor. Nimmt der pH-Wert zu, steigt der relative Anteil von HPO4
2-, während
bei einer pH-Verminderung der Anteil des H2PO4- ansteigt. In Lösung (biologischem
Gewebe) finden schnelle chemische Übergänge zwischen H2PO4- und HPO4
2- statt. Im31P-MR-Spektrum erkennt man daher nicht zwei streng voneinander getrennte
Resonanzen, sondern lediglich einen einzigen Pi-Peak, welcher durch die jeweiligen
prozentualen Anteile der Einzelresonanzen bestimmt wird. Ändert sich der pH-Wert, so
ist im 31P-MR-Spektrum eine Verschiebung des Pi-Peaks zu erkennen.
In der eigenen Untersuchung wurden die pH-Werte unter Verwendung des
folgenden Henderson-Hasselbalch-Gleichungssystems
pH p a AHA
= + −
K log10
,
basierend auf der chemischen Verschiebung δ δ δ= +p pA A B B von Pi (A) und PCr (B)
mitpH p a= + −
−
K log10δ δδ δ
12
ermittelt (Tab 3.1.1).
Der p aK -Wert, das Gleichgewicht zwischen Protonierung und Deprotonierung,
entspricht dem negativen Logarithmus der Gleichgewichtskonstante:
[ ][ ][ ]
KaH A
HA=
+ −,
mit H+ = Wasserstoffionen-Konzentration und A- = Basenionen-Konzentration
(HAUSSER und KALBITZER, 1989, ABELES et al., 1992, GRAHAM et al., 1994).
Für die Konstanten (pKa, δ1 und δ2) werden von verschiedenen Arbeitsgruppen
(mit Ausnahme der letzten beiden Gruppen, die ex vivo arbeiteten) in einem engen
Bereich variierende Werte verwendet, die innerhalb einer Untersuchungsreihe jedoch
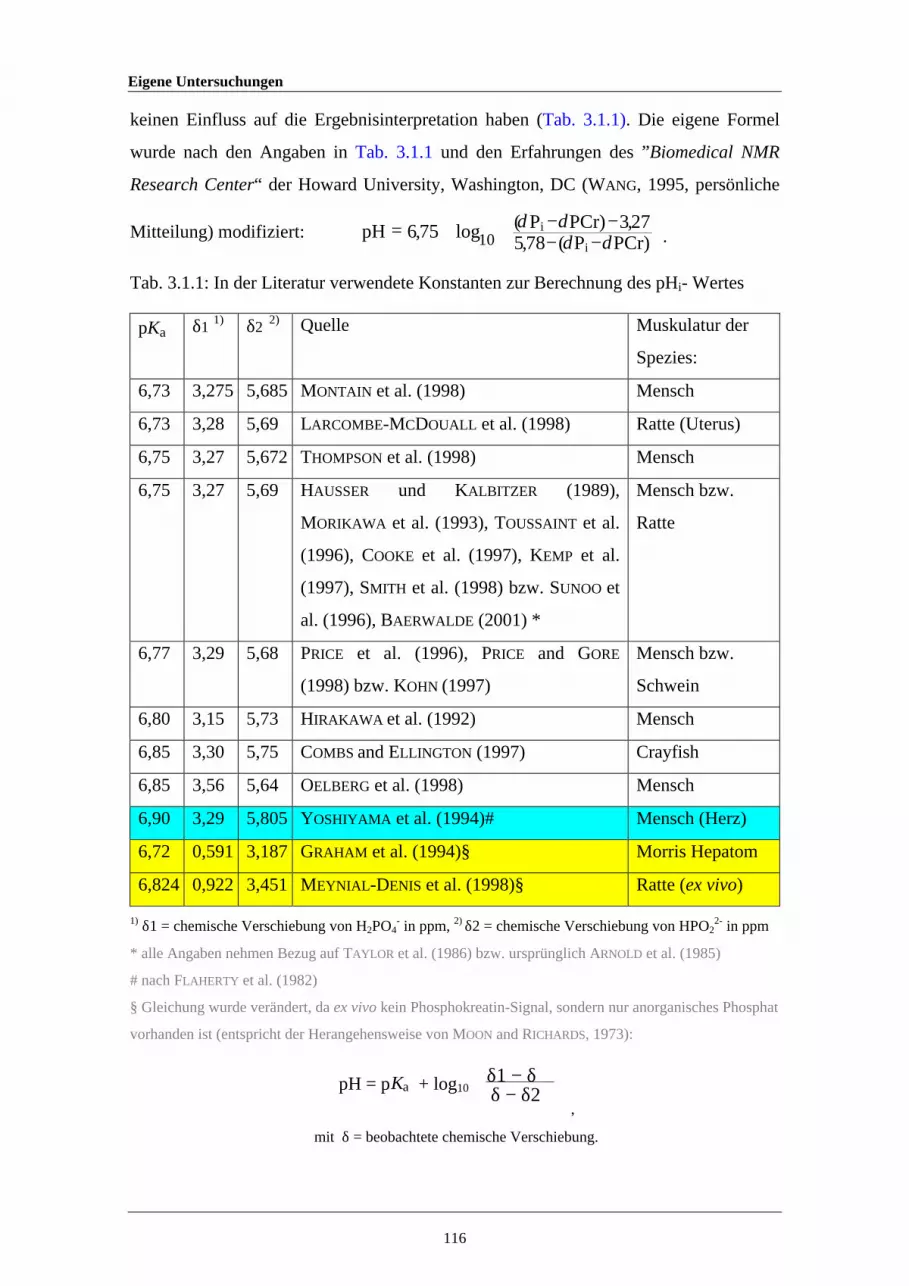
Eigene Untersuchungen
116
keinen Einfluss auf die Ergebnisinterpretation haben (Tab. 3.1.1). Die eigene Formel
wurde nach den Angaben in Tab. 3.1.1 und den Erfahrungen des ”Biomedical NMR
Research Center“ der Howard University, Washington, DC (WANG, 1995, persönliche
Mitteilung) modifiziert: pH = +6 75, log Pi PCrPi PCr
− −− −
327
578( ) ,, ( )
δ δδ δ 10 .
Tab. 3.1.1: In der Literatur verwendete Konstanten zur Berechnung des pHi- Wertes
pKa δ1 1) δ2 2) Quelle Muskulatur der
Spezies:
6,73 3,275 5,685 MONTAIN et al. (1998) Mensch
6,73 3,28 5,69 LARCOMBE-MCDOUALL et al. (1998) Ratte (Uterus)
6,75 3,27 5,672 THOMPSON et al. (1998) Mensch
6,75 3,27 5,69 HAUSSER und KALBITZER (1989),
MORIKAWA et al. (1993), TOUSSAINT et al.
(1996), COOKE et al. (1997), KEMP et al.
(1997), SMITH et al. (1998) bzw. SUNOO et
al. (1996), BAERWALDE (2001) *
Mensch bzw.
Ratte
6,77 3,29 5,68 PRICE et al. (1996), PRICE and GORE
(1998) bzw. KOHN (1997)
Mensch bzw.
Schwein
6,80 3,15 5,73 HIRAKAWA et al. (1992) Mensch
6,85 3,30 5,75 COMBS and ELLINGTON (1997) Crayfish
6,85 3,56 5,64 OELBERG et al. (1998) Mensch
6,90 3,29 5,805 YOSHIYAMA et al. (1994)# Mensch (Herz)
6,72 0,591 3,187 GRAHAM et al. (1994)§ Morris Hepatom
6,824 0,922 3,451 MEYNIAL-DENIS et al. (1998)§ Ratte (ex vivo)
1) δ1 = chemische Verschiebung von H2PO4- in ppm, 2) δ2 = chemische Verschiebung von HPO2
2- in ppm
* alle Angaben nehmen Bezug auf TAYLOR et al. (1986) bzw. ursprünglich ARNOLD et al. (1985)
# nach FLAHERTY et al. (1982)
§ Gleichung wurde verändert, da ex vivo kein Phosphokreatin-Signal, sondern nur anorganisches Phosphat
vorhanden ist (entspricht der Herangehensweise von MOON and RICHARDS, 1973):
K δ1 − δa + log10 δ − δ2pH = p
,
mit δ = beobachtete chemische Verschiebung.

Eigene Untersuchungen
117
Im Vergleich mit der pH-Formel von KOHN (1997, siehe Tab. 3.1.1), die ebenfalls
am Schwein, aber unter Verwendung eines 1,5 Tesla Magneten angewandt wurde,
würden die eigenen pH-Werte um ca. 0,06 Einheiten bei niedrigem pH-Niveau
(pH = 4,4) bzw. bis zu ca. 0,08 Einheiten bei hohem pH-Niveau (pH = 9,1) geringer
ausfallen. Die im eigenen Material am Schwein ermittelten Ruhe-pH-Werte (ca. 7,05)
stimmen jedoch sehr gut mit den unter Nutzung eines 4,7 Tesla -Magneten von PRICE et
al. (1996) am Menschen ermittelten durchschnittlichen Ruhe-pH-Werten von 7,03
überein, während KOHN (1997) Ruhe-pH-Werte von 7,10 bis 7,11 angibt. Die relativen
und absoluten Veränderungen im pH-Niveau werden durch die Wahl der Konstanten im
beschriebenen Schwankungsbereich nur minimal beeinflusst.
Ein GLM-Verfahren (SAS 6.11) mit den fixen Effekten Ryanodin-Rezeptor-
Genotyp, Zustand bei Versuchsende, Interaktion Ryanodin-Rezeptor-Genotyp mit
Zustand bei Versuchsende, Linie und Geschlecht sowie den Kovariablen Alter und
Körpermasse wurde für die Berechnung der Genotypdifferenzen verwendet:
Yijkl = µ + Ri + Lj + Sk + El + Ri • El + βAlter(Alterijkl - Alter) + βKM(KMijkl - KM) + eijkl
Yijkl = Beobachtungswert
µ = Erwartungswert von Y
Ri = Effekt des Ryanodin-Rezeptor-Genotyps (fix), i = 1 - 3
Lj = Effekt von Linie/Rasse (fix), j = 1 - 5
Sk = Effekt des Geschlechts (fix), k = 1 - 3
El = Effekt des Endzustandes (in vivo oder post mortem) (fix), l = 1 - 2
Ri • El
= Interaktion zwischen Ri und El, i • l = 1 - 6
Alterijklm = Alter (Kovariable)
KMijklm = Körpermasse (Kovariable)
eijklm = Restfehler.
Die Effekte Zustand bei Versuchsende bzw. der Interaktionseffekt fanden keine
Verwendung für die Stoffwechselparameter im “Ruhezustand”. Alter und Körpermasse
haben keinen signifikanten Einfluss (p ≥ 0,8) auf die Stoffwechselparameter nach
Stressauslösung mittels Halothan und werden deswegen bei der Endauswertung nicht im
Modell berücksichtigt.

Eigene Untersuchungen
118
3.1.1.2 13C-Magnet-Resonanz-Spektroskopie
An 27 Hybridschweinen der “Iowa State University Experimental Station” wurden
Untersuchungen zum Glykogenstoffwechsel mit Hilfe der 13C-MR-Spektroskopie unter
Verwendung eines Horizontalmagneten (“Varian© 4,7 Tesla” mit 33 cm nutzbarem
Innendurchmesser) in vivo durchgeführt. Die Genotypen NN (homozygot „normal“,
n = 7), Nn (heterozygot, n = 10) und nn (homozygote „defekt“, n = 10) wurden mit Hilfe
des beschriebenen Ryanodin-Rezeptor-1-Gentestes (Kapitel 3.1.6) bestimmt (BRENIG
and BREM, 1992, FÖRSTER u.a., 1992). Das Alter der Probanden variierte zwischen 41
und 58 Tagen bei Lebendmassen zwischen 7,3 und 19 kg. Von den 27 Tieren wurden 21
Tiere (6 NN - 8 Nn - 7 nn) in vivo untersucht, während 6 Tiere (1- 2- 3) bis zu einer
Stunde post mortem analysiert wurden. Alle Tiere stammten aus der Versuchsstation der
Iowa State University und setzten sich aus vier Zuchtlinien mit den folgenden
theoretischen Genanteilen zusammen:
I: 5/8 Duroc-, 1/4 US-Landrasse-, 1/8 Yorkshire-Genanteil
n = 9 ( 4 Nn - 5 nn) (> 50 % Duroc im Gesamtmaterial)
II: 1/2 Hampshire-, 1/4 Yorkshire-, 1/8 Pietrain-, 1/8 Spotted-Genanteil
n = 7 (4 NN - 2 Nn - 1 nn) (≥ 50 % Hampshire im Gesamtmaterial)
III: 1/4 Duroc-, 1/4 Hampshire-, 1/4 US-Landrasse-, 1/8 Spotted-, 1/8 Yorkshire-
Genanteilen
n = 9 ( 3 NN - 4 Nn - 2 nn), (DHLSY im Gesamtmaterial)
IV: 1/2 Spotted-, 1/4 Yorkshire-, 1/8 Pietrain-, 1/8 Duroc-Genanteil
n = 2 (2 nn), (≥ 50 % Spotted im Gesamtmaterial)
Davon waren 17 Tiere männlichen (4 NN - 9 Nn - 4 nn) und 10 weiblichen
Geschlechts (3 NN - 1 Nn - 6 nn).
Der intramuskuläre Glykogengehalt in vivo wurde nach einer Kalibrierung mit
einer externen Standardlösung (Phantom) bestimmt. Die Standardlösung bestand aus
reinem Glykogen (2 g), Kaliumchlorid (5 g) und destilliertem Wasser bei einem
Gesamtvolumen von 268,5 ml (Abb. 3.1.2). Die molare Masse für das “reine” Glykogen
wurde mit 179,7392 g/mol angegeben (Bestandteile: 99,5 % Glukose mit 180,15 g/mol
und 0,5 % H3PO4 mit 97,99 g/mol). Das ergab einen Glykogengehalt von
41,4422 µmol/g in der Standardlösung (0,00744879 g Glykogen/ g Lösung)
n 41,4422 = 2/268,5/179,7392 • 1000000.

Eigene Untersuchungen
119
Ein vergleichbares Vorgehen wurde von PRICE et al. (1996) beschrieben. Auf eine
bei ausgewachsenen Tieren (Probanden) notwendige “Fettkorrektur” (BUCHLI and
BOESIGER, 1993) wurde bei diesem Versuch mit sehr jungen Tieren verzichtet.
Abb. 3.1.2: Experimentelle Anordnung von 13C-MR-Doppelspule und Glykogen-standard
Das Signal des reinen Glykogens in situ ist in einem MR-Spektrum in 3 “Peaks”
aufgeteilt (Abb. 3.1.3). Allein die Signale des C1-Atoms und des C6-Atoms sind
aufgrund ihrer Anordnung im Glukosering getrennt von den anderen 4 C-Atomen
darstellbar. Die Signale der C-Atome 2 - 5 vereinigen sich zu einem Peak und waren
unter den gegebenen technischen Voraussetzungen nicht weiter aufzulösen. Da in vivo
nur das C1-Atom von Glykogen von anderen Signalen (kohlenstoffhaltigen Metaboliten
wie Glukose und Fettsäuren) unberührt bleibt, wurde das Messverfahren auf das Signal
von C1-Glykogen eingestellt, so dass das C1-Atom von Glykogen exakt die Position
100,5 ppm im Spektrum einnahm (Abb. 3.1.3).

Eigene Untersuchungen
120
C6-GlycogenC6-Glykogen
C4-, C3-, (C2-, C5)-Glycogen C4-, C3-, (C2-, C5)-Glykogen
C1-GlycogenC1-Glykogen
Abb. 3.1.3: In-situ- 13C-MR-Spektrum von reinem Glykogen, C1 auf 100,5 ppm justiert
Die Spektren wurden bei 50,295 MHz (Larmorfrequenz für Kohlenstoff in einem 4,7
Tesla MR-Tomographen) in einem Abstand von 273 Sekunden mit einer
Akquisitionszeit von 0,026 s für jedes FID (Free Induction Decay), einer
“Erholungszeit” von 0,158 s sowie 1480 Pulswiederholungen erfasst. Die Spektralweite
wurde auf 20 000 Hz festgelegt. Eine “Breitband-Wasserstoff-Rausch-Entkoppelung”,
1 ppm feldaufwärts vom Wassersignal zentriert, war nur während der Akquisitionszeit
aktiviert, um die Entwicklung des “Nuklear-Overhauser-Effekts” zu vermeiden (Abb.
3.1.4). In vivo wurden exakt dieselben Einstellungen des MR-Spektrometers sowie der
VNMR-Steuersoftware verwendet, um das Signal des C1-Glykogens auf dieselbe
Position wie in situ zu fixieren. Da sich die Signalintensität proportional zur
vorhandenen Menge Glykogen verändert, ließ sich die Glykogenkonzentration im
Muskel über den Vergleich mit der in der Standardlösung ermittelten Fläche des C1-
Glykogens messen. Zusätzlich diente während der Spektroskopie in vivo das Signal von
Kreatin an der Position 157 ppm als interner Standard.

Eigene Untersuchungen
121
Abb. 3.1.4: Aufnahmeparameter und Beispiel für ein Glykogenspektrum in vivo (C1-Glykogen bei 100,5 ppm und Kreatin bei 157 ppm)
Die Interpretation der prozentualen Veränderungen in den Metaboliten-
konzentrationen gestattet keine Aussage über die absoluten Konzentrations-
veränderungen, ohne das „wahre“ Ausgangsniveau zu kennen. Als Hilfsmittel für
Schlussfolgerungen über absolute Niveauveränderungen kann die Spektralskala nur
verwendet werden, wenn die Untersuchungen unter standardisierten Bedingungen
erfolgen. Veränderungen der Skalierung bzw. der Einstellungen für Hardware und
Software sowie der Position des Messobjektes im Magneten erschweren die
Ergebnisinterpretation erheblich.
Zehn Minuten vor dem Spektroskopie-Experiment wurden die Tiere - identisch
wie bei der Phosphorspektroskopie - unter Verwendung einer Mixtur von Ketamin (100
mg/10 kg Körpergewicht), Telazol und Rompun (50 mg + 50 mg/10 kg Körpergewicht)
anästhesiert und auf der rechten Seite liegend in einer Versuchswiege platziert. Danach

Eigene Untersuchungen
122
wurde eine im Außendurchmesser 9,6 cm (1H) und im Innendurchmesser 5,5 cm (13C)
große, kombinierte 1H-13C-Oberflächenspule auf dem M. biceps femoris fixiert (Abb.
3.1.5). Durch die Größenvariation der Probanden hervorgerufene Abweichungen der
Messregion (M. biceps femoris) vom Magnetzentrum wurden durch Schaum-
stoffunterlagen ausgeglichen, um den Bedingungen in situ zu entsprechen.
Vor der Stressauslösung mittels Halothan (3 Volumen-% per 3 Liter O2/min)
wurden für jedes Tier mindestens 4 “Ruhespektren” (18,2 Minuten) aufgenommen
(Abb. 3.1.4, 3.1.6). Mindestens 5 Spektren wurden nach Beendigung der Halothan-
zufuhr erfasst, um die Erholungsphase in vivo verfolgen zu können. Die Gesamtzahl der
Spektren in vivo hing von der Wirkung der Anästhesie ab. Nach einer “Baseline”-
Korrektur wurden die Flächen (Integrale) des C1-Glykogensignals mit Hilfe der Varian-
NMR-Software (VNMR 4.3, 1994) berechnet. Für die Kalkulation der prozentualen
Konzentrationsveränderung von Glykogen und Kreatin wurde der Mittelwert aus den
“Ruhespektren” gleich 100 % gesetzt.
Abb. 3.1.5: 1H-13C-Spulenanordnung über dem M. biceps femoris in vivo

Eigene Untersuchungen
123
C1-Glykogen
Fettsäureketten
Kreatin ßC1-Glukose
-CH=CH-CH2-CH=CH-
Abb. 3.1.6: Beispiel für ein charakteristisches 13C-Spektrum an der C1-Glykogen-Position
Zur Auswertung der Daten wurden zwei verschiedene Modelle mit Hilfe der GLM-
Prozedur (SAS 6.11) verglichen. Das Modell I deckt sich mit dem Modell für die 31P-
MR-Spektren.
Modell I:
Yijkl = µ + Ri + Lj + Sk + El + Ri • El + βAlter(Alterijkl - Alter) + βKM(KMijkl - KM) + eijkl
Yijkl = Beobachtungswert
µ = Erwartungswert von Y
Ri = Effekt des RyR1-Genotyps (fix), i = 1-3
Lj = Effekt der Rasse/Linie (fix), j = 1-4
Sk = Effekt des Geschlechts (fix), k = 1-2
El = Effekt des Endzustandes (in vivo oder post mortem) (fix), l = 1-2
Ri • El = Interaktion zwischen Ri und El
Alterijkl = Alter (Kovariable)
KMijkl = Körpermasse (Kovariable)
eijklm = Restfehler

Eigene Untersuchungen
124
Da für die untersuchten 27 Tiere die theoretische Zusammensetzung der
Kreuzungslinien bekannt war, wurden im Modell II leichte Modifikationen
vorgenommen, um den Effekt des Hampshire-Genanteils speziell auf die
Veränderungen des Glykogenniveaus überprüfen zu können.
Mit einer multiplen Regressionsanalyse (SAS 6.11) wurde vorab geprüft, ob
zwischen den Genanteilen der an den Kreuzungslinien beteiligten Reinzuchtherkünfte
(Duroc, Hampshire, Pietrain, Spotted, US-Landrasse, Yorkshire) in Kombination mit
der RyR1-Defektallelhäufigkeit (0/ 0,5/ 1) sowie der Dauer der Halothangabe und dem
interessierenden Merkmal Glykogenabbau (%) eine signifikante Beziehung besteht.
Folgende Rasse-Genanteile existieren im eigenen Material:
Duroc (Du) = 0 / 0,125 / 0,25 / 0,625,
Hampshire (Ha) = 0 / 0,25 / 0,5,
Pietrain (PI) = 0 / 0,125,
Spotted (S) = 0 / 0,125 / 0,5,
US-Landrasse (LR) = 0 / 0,25,
Yorkshire (Y) = 0 / 0,125 / 0,25.
Unter Verwendung der „Backward“-Prozedur (p = 0,05) und der Bedingung, den
Hampshire-Genanteil und der RyR1-Defektallelanteil als Gruppe zu behandeln, wurde
neben dem Hampshire-Genanteil und dem RyR1-Defektallelanteil nur noch Pietrain in
der Regressionsgleichung belassen. Die resultierenden multiplen Beziehungen zwischen
RyR1-Defektallelanteil, Hampshire- und Pietrain-Genanteil sowie dem Glykogenabbau
nach Belastung sind in Abb. 3.1.7 dargestellt. Mit steigendem (theoretischem)
Hampshire-Genanteil vermindert sich der Glykogenabbau (%), während gleichzeitig die
Anwesenheit von Pietrain-Genanteilen unabhängig vom RyR1-Genotyp einen stärkeren
Glykogenabbau zur Folge hat. Ein zunehmender RyR1-Defektallelanteil bewirkt einen
verstärkten Glykogenabbau.
In das Varianzanalyse-Modell II wurde entsprechend der Pietrain-Genanteil
zusätzlich als „Kovariable“ aufgenommen, um eine Korrektur auf den Pietrain-
Genanteil vornehmen zu können. In den vier Kreuzungslinien gab es folgende
Kombinationen von Pietrain-Genanteilen und RyR1-Genotypen: 0 % Pietrain mit 3 NN -
8 Nn - 7 nn bzw. 12,5 % Pietrain mit 4 NN - 2 Nn - 3 nn. Von den 9 Tieren mit Pietrain-
Genanteilen beendeten 5 Tiere (3 NN - 1 Nn - 1 nn) das Experiment in vivo. Das heißt, 4
von 6 Tieren, deren Untersuchung post mortem beendet wurde, besaßen Pietrain-
Genanteile (12,5 %).

Eigene Untersuchungen
125
Ha0%
Ha25%
Ha50%
PI0
%
PI12,5%
-70
-60
-50
-40
-30
-20
-10
0n 0 %
n 50 %n 100 %
n 0 %n 50 %
n 100 %
Abb. 3.1.7: Beziehungen zwischen Glykogenabbau und RyR1-Defektallelanteil (n)sowie Hampshire- bzw. Pietrain-Genanteil (Ergebnis aus der Backward-Regressions-Analyse.Unter der Restriktion, den Hampshire-Genanteil und den RYR1-Defektallelanteil als Gruppe zu behandeln,lautet die Gleichung (p = 0,05):
Glykogenabbau (%) = -31,627 + [39,092 • Ha] + [-8,476 • RyR1] + [-205,848 • PI]; mit R² = 0,35; √ MSE = 15,88.)
Modell II:
Yijkl = µ + Ri + Lj + Sk + El + Ri • El + βP (Pijkl - P) + eijkl
Yijkl = Beobachtungswert
µ = Erwartungswert von Y
Ri = Effekt des RyR1-Genotyps (fix), i = 1 - 3
Hj = Effekt des Hampshire-Genanteils (fix), j = 1 - 3 (0 %; 0,25 %; 0,5 %)
Sk = Effekt des Geschlechts (fix), k = 1 - 2 (männlich, weiblich)
El = Effekt des Endzustandes (fix), l = 1 - 2 (in vivo, post mortem)
Ri • El = Interaktion zwischen Ri und El
Pijkl = Pietrain-Genanteil (Kovariable)
eijkl = Restfehler
Identisch zur 31P-MR-Spektroskopie wurden die Effekte “Zustand bei
Versuchsende” bzw. der Interaktionseffekt für die Stoffwechselparameter im
“Ruhezustand” nicht in die Modelle I und II aufgenommen. Körpermasse und Alter
beeinflussten in dem untersuchten Tiermaterial die Merkmale aus der 13C-MR-
Spektroskopie nicht und wurden wie für die 31P-MR-Spektroskopie bei der
Endauswertung aus dem jeweiligen Modell entfernt.

Eigene Untersuchungen
126
3.1.2 Ermittlung der Körperzusammensetzung mittels MRT und DXA
3.1.2.1 1H-Magnet-Resonanz-Tomographie3.1.2.1.1 Lebendmassegruppe bis 20 kg
Im Anschluss an die Stoffwechseluntersuchungen bzw. unabhängig davon wurden
insgesamt 111 Schweine im VARIAN® 4,7 Tesla Magneten (Varian NMR Systems, Palo
Alto, CA, USA) bei einer Resonanzfrequenz von 200,23 MHz (1H-Atomkerne)
tomographiert. Die durchschnittliche Lebendmasse betrug 12,37 ± 6,18 kg mit einem
Minimum von 6,0 kg und einem Maximum von 30,08 kg bei einem durchschnittlichen
Alter von 53,69 ± 16,33 Lebenstagen (28 – 91 Lebenstage).
Wenn die Tiere vor der Bildgebung bereits einer spektroskopischen Untersuchung
dienten, wurde nur bei Bedarf nochmals die beschriebene Mixtur der Sedativa
verabreicht. Für alle anderen Tiere erfolgte die Sedierung der Tiere unter Verwendung
der Mixtur von Ketamin (100 mg/10 kg Körpergewicht), Telazol and Xylazin (50 mg +
50 mg/10 kg Körpergewicht) zehn Minuten vor der Tomographie.
Insgesamt ergab sich folgende Verteilung für die RyR1-Genotypen:
37 NN - 46 Nn - 28 nn, die aus vier „synthetischen“ Linien, zwei Kreuzungslinien und
einer Reinzuchtherkunft stammten (Tab. 3.1.2). Die „synthetischen“ Linien sowie die
Kreuzung aus Duroc und Minzhu wurden in der Versuchsstation der Iowa State
University zur Erzeugung der drei RyR1-Genotypen gezüchtet. Allein die
Einfachkreuzung „Poland China x US-Landrasse“ stammt aus der Versuchsstation des
„Livestock and Poultry Science Institute“ des USDA-ARS in Beltsville, während die
Tiere der US-Landrasse von einem Züchter aus Maryland bezogen wurden.
Tab. 3.1.2: Linienherkunft und Anzahl Probanden für 1H-MR-Tomographie bis 20 kg
Duroc-Linie (> 50% Duroc): n = 39
Hampshire-Linie (≥ 50 % Hampshire): n = 17
Kreuzungslinie mit 25 % Duroc, 25 % Hampshire, 25 % US-Landrasse,
12,5 % Yorkshire und 12,5 % Spotted Poland China (DHLYS): n = 6
Spotted Poland China (≥ 50 % Spotted): n = 6
Poland China x US-Landrasse: n = 27
US-Landrasse: n = 8
Duroc x Minzhu: n = 8
Σ: n = 111

Eigene Untersuchungen
127
Die Probanden setzten sich aus 63 weiblichen bzw. 29 intakten männlichen Tieren
und 19 männlichen Kastraten zusammen.
Für die Bildgebung (Tomographie) wurde eine sattelförmige, einmalgewickelte
Oberflächenspule zwischen der Schulter- und der Nierenregion platziert (Abb. 3.1.8).
Ein ”Multi-Slice-Spin-Echo”-Verfahren mit den folgenden Parametern:
1 Echo,
Zwischenechozeit (Time between Echoes - TE) = 20 ms (oder alternativ 24 ms),
Wiederholungszeit (Time of repetition - TR) = 400 ms (oder alternativ 200 ms),
4 Wiederholungen und
128 Phasen kodierende Schritte
diente zur Erzeugung von 5 aufeinanderfolgenden Axialschnittbildern zwischen 14. und
ca. 12. Brustwirbel mit einer Scheibendicke von 4,9 mm (Abb. 3.1.9).
Abb. 3.1.8: Versuchsanordnung zur Tomographie der Tiere bis 20 kg Lebendmassemittels 1H-MR-Oberflächenspule
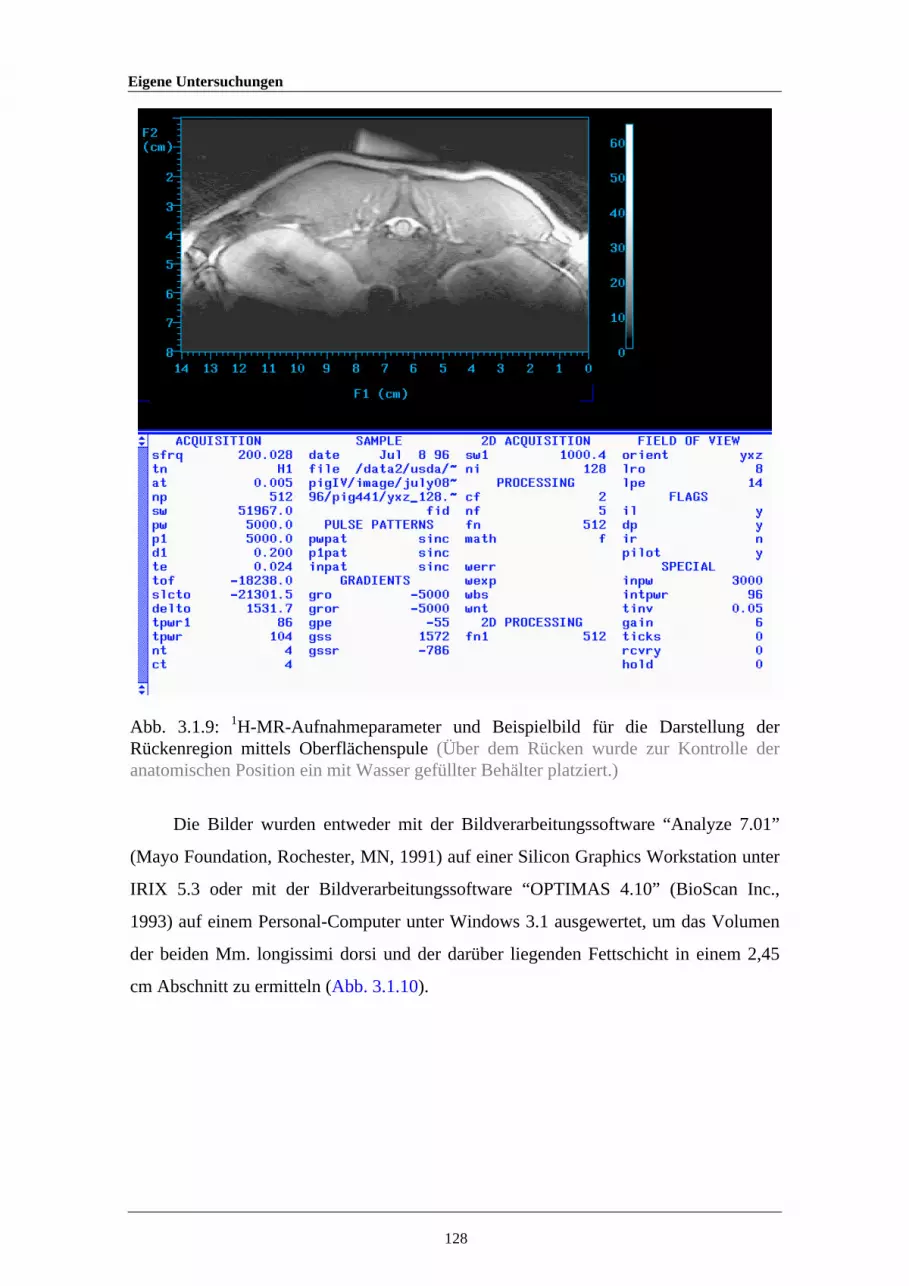
Eigene Untersuchungen
128
Abb. 3.1.9: 1H-MR-Aufnahmeparameter und Beispielbild für die Darstellung derRückenregion mittels Oberflächenspule (Über dem Rücken wurde zur Kontrolle deranatomischen Position ein mit Wasser gefüllter Behälter platziert.)
Die Bilder wurden entweder mit der Bildverarbeitungssoftware “Analyze 7.01”
(Mayo Foundation, Rochester, MN, 1991) auf einer Silicon Graphics Workstation unter
IRIX 5.3 oder mit der Bildverarbeitungssoftware “OPTIMAS 4.10” (BioScan Inc.,
1993) auf einem Personal-Computer unter Windows 3.1 ausgewertet, um das Volumen
der beiden Mm. longissimi dorsi und der darüber liegenden Fettschicht in einem 2,45
cm Abschnitt zu ermitteln (Abb. 3.1.10).

Eigene Untersuchungen
129
Musculi longissimi dorsi
Rückenspeck5 Scheiben,insgesamt 2,45 cm
Abb. 3.1.10: Darstellung der ”Regions of Interest“ (ROI’s) in den 1H-Magnet-Resonanz-Schnittbildern im Bereich 13./14. Brustwirbel für Tiere bis 20 kg Lebendmasse
Ein GLM-Verfahren (SAS 6.11) mit den fixen Effekten Ryanodin-Rezeptor-1-Genotyp,
Linie und Geschlecht sowie den Kovariablen Alter und Körpermasse wurde für die
Berechnung der Genotypdifferenzen verwendet:
Yijkl = µ + Ri + Lj + Sk + βAlter (Alterijkl - Alter) + βKM (KMijkl - KM) + eijkl
Yijkl = Beobachtungswert
µ = Erwartungswert von Y
Ri = Effekt des Ryanodin-Rezeptor-1-Genotyps (fix), i=1-3
Lj = Effekt von Linie/Rasse (fix), j=1-6
Sk = Effekt des Geschlechts (fix), k=1-3
Alterijkl = Alter (Kovariable)∗
KMijkl = Körpermasse (Kovariable)*
eijkl = Restfehler.
∗ Nicht im Modell für das Merkmal Fett-zu-Muskel-Verhältnis.

Eigene Untersuchungen
130
3.1.2.1.2 Lebendmassegruppen 30 kg, 60 kg, 90 kg
Abb. 3.1.11: Positionierung eines Probanden im Picker Vista 1,5 Tesla MR-Tomographen (63 MHz für Wasserstoff);Schweine mit bekanntem RyR1-Genotyp NN wurden zur Verminderung vonAtmungsartefakten zusätzlich mit Halothan sediert
Im Rahmen eines Wachstumsversuch sowie einer vergleichenden Studie mit
densitometrischen Untersuchungen (Dualenergie-Röntgenabsorptiometrie - DXA)
wurden insgesamt 47 Tiere, die bereits bei 10 kg Lebendmasse spektroskopiert bzw.
tomographiert wurden, erneut bei 30 kg Lebendmasse mit einem 1,5 Tesla
Ganzkörpertomographen untersucht, um die Körperzusammensetzung anhand von 1H-
MR-Bilder zu analysieren. Nach Beendigung der In-vivo-Körperzusammensetzungs-
Studien (MRT, DXA) bei 30 kg Lebendmasse wurden 15 Tiere geschlachtet und zerlegt.
Danach wurden 32 Tiere bei ca. 60 kg Lebendmasse erneut mittels DXA untersucht. Für
die MR-Tomographie in der 60 kg Gruppe konnten jedoch nur 31 Tiere genutzt werden,
da ein männlicher Kastrat der Linie „Hampshire“ nach der DXA-Untersuchung mit ca.
60 kg verendete. In der letzten Gruppe bei 90 kg Lebendmasse verblieben 17 Schweine
(6 NN - 6 Nn - 5 nn), die schließlich ebenfalls für Referenzzwecke zerlegt und chemisch
auf Rohfett-, Rohprotein-, Rohasche- und Trockensubstanzgehalt analysiert wurden.

Eigene Untersuchungen
131
Die 1H-MR-Tomographie bei den 30 - 90 kg Tieren erfolgte mit Hilfe eines Picker
Vista 1,5 Tesla Ganzkörpertomographen (Picker International, Highland Heights, OH,
USA) mit 1 Meter nutzbarem Durchmesser (Abb. 3.1.11).
Ein modifiziertes “Multi-Slice-Spin-Echo-Verfahren” (1 Echo, TR = 1,3 s, TE =
20 ms, 2 Wiederholungen) diente in zwei Schritten zur Erzeugung von insgesamt 25
Schnittbildern. Im Unterschied zu SCHOLZ u.a. (1993) wurde anstelle von 4 “Echos” nur
1 ”Echo“ aufgenommen. Im Rückenbereich wurden 10 aufeinanderfolgende
Axialschnittbilder beginnend am 14. Brustwirbel in kranialer Richtung erzeugt, um das
Volumen der Mm. longissimi dorsi und des darüber liegenden Rückenspecks zu messen
(Abb. 3.1.12, 3.1.13, 3.1.14). Weiterhin wurden im Bereich Becken/Hintergliedmaßen
15 aufeinanderfolgende Axialschnittbilder beginnend am äußeren Schwanzansatz in
kranialer Richtung ausgewählt, um das Schinkenvolumen (inklusive Knochen) und das
darüber liegende Fettvolumen zu bestimmen (Abb. 3.1.12, 3.1.15). Die Scheibendicke
betrug – ohne Abstand zwischen den Scheiben - jeweils 10 mm.
Mit Hilfe der Bildverarbeitungssoftware “Analyze 7.01” (Mayo Foundation,
Rochester, MN, USA) wurden die Volumina der Mm. longissimi dorsi und des
Rückenspecks bzw. des Schinkenmagergewebes und der Fettauflage mit Hilfe einer
halbautomatischen Gewebesegmentation bestimmt (Abb. 3.1.14, 3.1.15). Die
anatomischen Orientierungspunkte richten sich nach den Angaben von WOLTER (1997).
Dabei ist zu beachten, dass im Gegensatz zur eigenen Versuchsanordnung die 4 bis 5
Wochen alten Tiere bei WOLTER (1997) euthanasiert waren und in Rückenlage mit in
kranialer Richtung gestreckten Vordergliedmaßen tomographiert wurden. Außerdem
betrug bei einem Zwischenscheibenabstand von 0,5 mm die Scheibendicke 5 mm.
Die Verteilung der Schweine über die RYR1-Genotypen bzw. Herkünfte innerhalb
der Lebendmassegruppen zeigen die Tabellen 3.1.3 – 3.1.6.
Tab. 3.1.3: Anzahl tomographierter Tiere je RyR1-Genotyp und Lebendmassegruppe (30
– 90 kg)
NN Nn nn total
30 kg 16 22 9 47
60 kg 14 11 6 31
90 kg 6 6 5 17
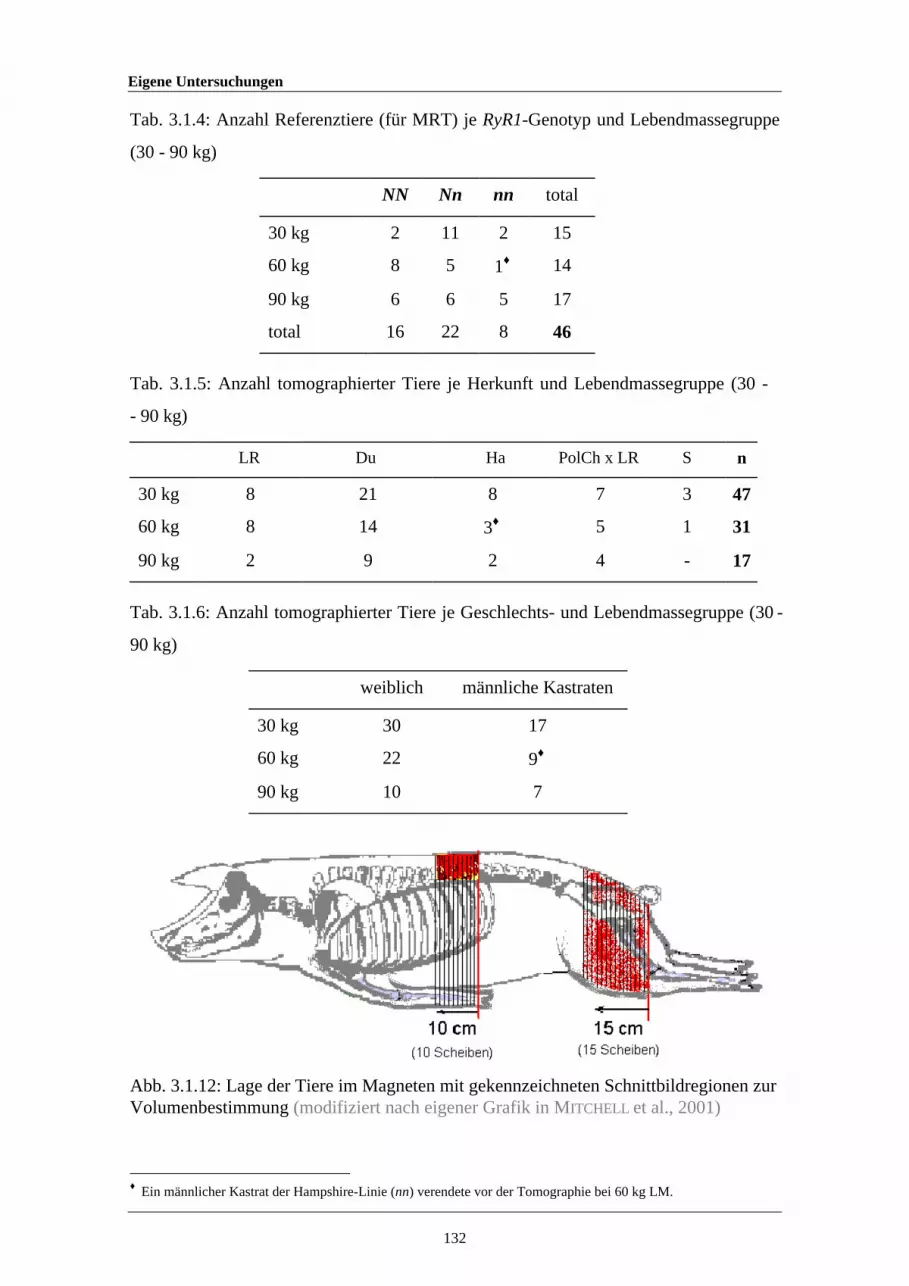
Eigene Untersuchungen
132
Tab. 3.1.4: Anzahl Referenztiere (für MRT) je RyR1-Genotyp und Lebendmassegruppe
(30 - 90 kg)
NN Nn nn total
30 kg 2 11 2 15
60 kg 8 5 1♦ 14
90 kg 6 6 5 17
total 16 22 8 46
Tab. 3.1.5: Anzahl tomographierter Tiere je Herkunft und Lebendmassegruppe (30 -
- 90 kg)
LR Du Ha PolCh x LR S n
30 kg 8 21 8 7 3 47
60 kg 8 14 3♦ 5 1 31
90 kg 2 9 2 4 - 17
Tab. 3.1.6: Anzahl tomographierter Tiere je Geschlechts- und Lebendmassegruppe (30 -
90 kg)
weiblich männliche Kastraten
30 kg 30 17
60 kg 22 9♦
90 kg 10 7
Abb. 3.1.12: Lage der Tiere im Magneten mit gekennzeichneten Schnittbildregionen zurVolumenbestimmung (modifiziert nach eigener Grafik in MITCHELL et al., 2001)
♦ Ein männlicher Kastrat der Hampshire-Linie (nn) verendete vor der Tomographie bei 60 kg LM.

Eigene Untersuchungen
133
PPaannkkrreeaass
Mm. longissimidorsi
Abb. 3.1.13: Axialschnittbild im Bereich zwischen 13./14. Brustwirbel (modifiziertnach eigener Grafik in MITCHELL et al., 2001)
Rückenspeck
Mm. longissimi dorsi
Abb. 3.1.14: Regionen für die Volumenbestimmung im Thoraxbereich (10 Scheiben, 10cm)

Eigene Untersuchungen
134
Muskel
FettOs ilium Os sacrum
Fett-ROI
Magergewebe-ROI
nicht in der ROI
Abb. 3.1.15: Anatomische Hilfspunkte und schematische Darstellung der ”Regions ofInterest“ (ROI’s) im Becken-/Hintergliedmaßenbereichoben: Anatomische Hilfspunkte und Kennzeichnung von auszuwertenden bzw.wegzulassenden Regionen für die Volumenbestimmung im BereichBecken/Hintergliedmaßenunten: Schema zur Festlegung von ”Regions of Interest” (modifiziert nach eigenerGrafik in Mitchell und Scholz, 2001)
Die anfallenden Daten zum Volumen der Mm. longissimi dorsi bzw. des
Schinkenvolumens sowie der beiden Fettvolumenmaße wurden mit der GLM-Prozedur
nach dem folgenden statistischen Modell innerhalb der Lebendmassegruppen
ausgewertet:

Eigene Untersuchungen
135
Yijkl = µ+ Ri + Lj +Sk + βAlter (Alterijkl - Alter) + βKM (KMijkl - KM) + eijkl
Yijkl = Beobachtungswert
µ = Erwartungswert von Y
Ri = Effekt des Ryanodin-Rezeptor-Genotyps (fix), i=1-3
Lj = Effekt von Linie/Rasse (fix), j=1-5
Sk = Effekt des Geschlechts (fix), k=1-2
Alterijkl = Alter (Kovariable)
KMijkl = Körpermasse (Kovariable)
eijkl = Restfehler.
Zusätzlich wurde eine gemeinsame Auswertung über alle Lebendmassegruppen (30 - 90
kg) nach dem gleichen Modell allein für die Tiere (Messung) vorgenommen, die nach
der MR-Tomographie bzw. der DXA-Untersuchung euthanasiert, zerlegt und chemisch
analysiert wurden. Das heißt, Tiere, die den Versuch bis zum Ende durchliefen, gingen
nur in der 90 kg Gruppe in die Auswertung ein. Wiederholte Messungen bei 30 und 60
kg wurden entsprechend nicht in der Varianzanalyse berücksichtigt.

Eigene Untersuchungen
136
3.1.2.2 Dualenergie-Röntgenabsorptiometrie
Insgesamt wurde die Körperzusammensetzung von 92 Schweinen in vivo mit Hilfe
eines Ganzkörper-DXA-Scanners (DPX-L©, GE LUNAR®, Waukesha, WI) analysiert.
Bei 62 Tieren wurden die DXA-Resultate mit den Ergebnissen der chemischen
Schlachtkörperanalyse verglichen (Tab. 3.1.7 - 3.1.9). Die Körperzusammensetzung für
jedes Tier wurde mit Hilfe der Software von GE LUNAR® (Version 4.6d) ermittelt. Mit
Hilfe des Ryanodin-Rezeptor-1-Gentestes wurden 29 homozygot stressstabile (NN)
Schweine, 36 heterozygote (Nn) und 26 homozygot stressanfällige Tiere (nn)
identifiziert.
Tab. 3.1.7: Aufteilung der Probanden nach RyR1-Genotyp und Lebendmassegruppe
(10 - 90 kg) für den DXA-Versuch
RyR1-Genotyp Lebendmassegruppe
10 30 60 90 Total
NN 13 (4) 16 (2) 14 (8) 6 (6) 29 (20)
Nn 14 (4) 22 (11) 11 (5) 6 (6) 36 (26)
nn 18 (7) 9 (2) 7 (2) 5 (5) 27 (16)
Total 45 (15) 47 (15) 32 (15) 17 (17) 92 (62)
*(Tiere für Referenzuntersuchung)
Tab. 3.1.8: Aufteilung der Probanden nach Rasse- bzw. Linienzugehörigkeit und
Lebendmassegruppe (10 - 90 kg) für den DXA-Versuch
Rasse/Linie Lebendmassegruppe
10 30 60 90 Total
Duroc (Du) 17 *(6) 21 (7) 14 (5) 9 (9) 38 (27)
US-Landrasse (LR) 2 (1) 8 (-) 8 (6) 2 (2) 10 (9)
Poland China * LR 2 (2) 7 (2) 5 (1) 4 (4) 9 (9)
Hampshire (Ha) 11 (4) 8 (4) 4 (2) 2 (2) 19 (12)
Spotted (S) 4 (2) 3 (2) 1 (1) - 7 (5)
DHLYS 9 (-) - - - 9 (-)
Total 45 (15) 47 (15) 32 (15) 17 (17) 91 (62)
*(Tiere für Referenzuntersuchung)
Von den 45 Tieren, die in der 10 kg Gruppe (durchschnittliche Lebendmasse =
12,55 kg; Durchschnittsalter = 53 Tage) mittels DXA untersucht wurden, dienten 15 als
Eigene Untersuchungen
137
Referenztiere für die Zerlegung und chemische Analyse. Bei den 47 Tieren ab 30 kg
erfolgte eine Stufenschlachtung (30 kg: n = 15; 60 kg: n = 15 Tiere und 90 kg: n = 17).
Tab. 3.1.9: Aufteilung der Probanden nach Geschlecht und Lebendmassegruppe (10 –
90 kg) für den DXA-Versuch
Geschlecht Lebendmassegruppe
10 30 60 90 Total
Männlich (intakt) 18 *(1) - - - 18 (1)
Männliche Kastraten 3 (2) 17 (7) 10 (3) 7 (7) 20 (19)
Weiblich 24 (12) 30 (8) 22 (12) 10 (10) 54 (42)
Total 45 (15) 47 (15) 32 (15) 17 (17) 92 (62)
*(Tiere für Referenzuntersuchung)
Die DXA-Untersuchung erfolgte für die meisten Tiere drei Tage nach der MR-
Tomographie. In Ausnahmefällen fand die DXA-Untersuchung fünf Tage vor der
geplanten MR-Tomographie bzw. für einen Großteil der Tiere aus der 10 kg Gruppe
unabhängig von der MR-Tomographie statt.
Unmittelbar vor dem Scannen wurden die Tiere elektronisch gewogen (± 0,2 kg).
Alle Tiere wurden anästhesiert (500 mg Ketamin, 80 mg Tiletamin, 80 mg Zolazepam
und 333 mg Xylazin je 100 kg Lebendmasse, i.m., gefolgt von 290 mg Ketamin, 37 mg
Tiletamin, 37 mg Zolazepam, 150 mg Xylazin, 3 mg Butorphanoltartrat und 0,16 mg
Atropinsulfat je 100 kg Lebendmasse, i.v.), um Bewegungen während des Scan-
Verfahrens zu vermeiden. Die Schweine lagen mit ausgestreckten Hintergliedmaßen auf
dem Bauch, während die Vordergliedmaßen seitlich entlang des Rumpfes gestreckt
waren und mit Schaumstoffkissen vom Körper auf Distanz gehalten wurden. Damit
wurden Überlagerungen von Rumpf und Vordergliedmaßen verhindert (Abb. 3.1.16).

Eigene Untersuchungen
138
Abb. 3.1.16: Lage des Probanden auf dem DXA-Scanner (GE LUNAR DPX-LTM)
Drei verschiedene Scan-Einstellungen wurden in Abhängigkeit von der
Körpergröße der Schweine eingesetzt: 5-29 kg “pediatric-medium”, 30-69 kg
“adult-medium” und > 69 kg “adult-slow”. Diese Einstellungen regulieren die
Scangeschwindigkeit und/ oder die Kollimation (Ausrichtung der „optischen“ und
mechanischen Achsen für eine modifizierte Fokussierung bzw. Strahldivergenz der
Photonenstrahlen). Sie beeinflussen jedoch nicht das Energieniveau der
Röntgenstrahlung.
Nach individueller Anpassung der voreingestellten Körperregionen auf die
anatomischen Verhältnisse im Schwein erscheinen Gesamtkörperfettmasse,
Magergewebe- (Muskel-)masse und Knochenmineralgehalt (BMC) bzw.
Knochenmineraldichte (BMD) als Hauptbestandteile im Ergebnisausdruck der DXA-
Analyse (Abb. 3.1.17).
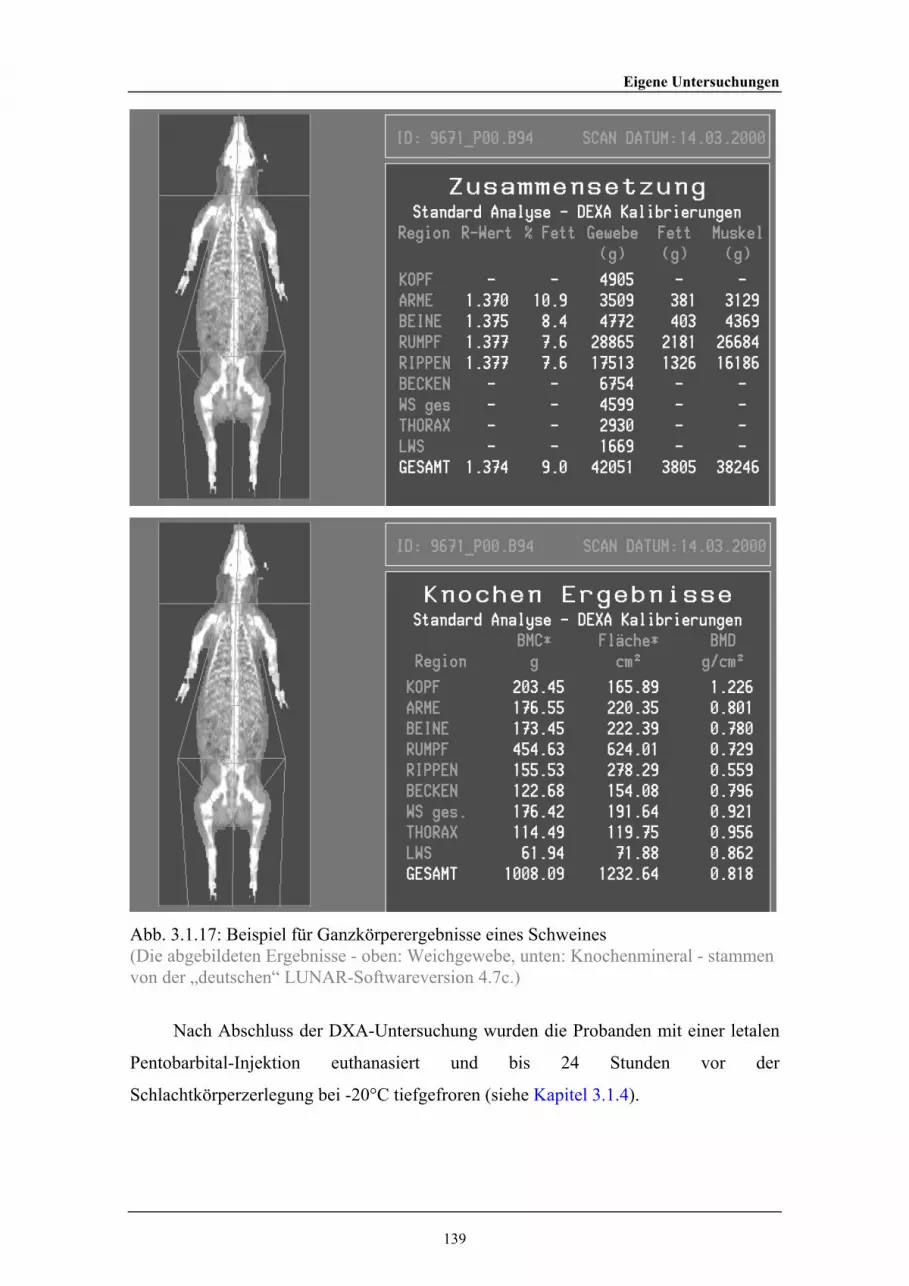
Eigene Untersuchungen
139
Abb. 3.1.17: Beispiel für Ganzkörperergebnisse eines Schweines (Die abgebildeten Ergebnisse - oben: Weichgewebe, unten: Knochenmineral - stammenvon der „deutschen“ LUNAR-Softwareversion 4.7c.)
Nach Abschluss der DXA-Untersuchung wurden die Probanden mit einer letalen
Pentobarbital-Injektion euthanasiert und bis 24 Stunden vor der
Schlachtkörperzerlegung bei -20°C tiefgefroren (siehe Kapitel 3.1.4).

Eigene Untersuchungen
140
DXA liefert keine direkten Messwerte für Muskelmasse bzw. Proteingehalt,
jedoch einen Messwert für Magergewebe mit allen Komponenten außer Fett und
Knochenmineral.
Der Gesamtproteingehalt sowie der Wassergehalt als Hauptbestandteil des
Magergewebes kann aus der DXA-Magergewebemasse geschätzt werden - nach den
Gleichungen:
Protein (g) = -1,062 + 0,22 • [DXA-Magergewebe] (MITCHELL and CONWAY, 1994)
und
Wasser (g) = 508 + 0,74 • [DXA-Magergewebe] (MITCHELL et al., 1996 a).
Die statistische Auswertung für die DXA-Merkmale erfolgte wiederum unter
Benutzung der GLM-Prozedur (SAS 6.11) innerhalb Lebendmassegruppen und über
alle Lebendmassegruppen nach folgendem statistischen Modell:
Yijkl = Beobachtungswert des m-ten Tieresµ = Erwartungswert von YRi = fixer Effekt des Ryanodin-Rezeptor-Genotyps
Lj = fixer Effekt von Linie/Rasse
Sk = fixer Effekt des Geschlechts
= Alter (Kovariable)
= Körpermasse (Kovariable)
Aijkl
KMijkl
eijkl = Restfehler.
Yijkl = µ + Ri + Lj + Sk + ßAlter (Aijkl -A) + ßLM (KMijkl -KM )+ eijkl
Für die Wachstumsanalyse in Kapitel 3.2.3 (Merkmale Lebendmassezunahme,
Magergewebezunahme, Fettgewebezunahme, Knochenmineralzunahme, Magergewebe-
futterverwertung und Futteraufnahme erfolgte keine Korrektur auf Lebendmasse und
Alter, um die vorhandene Variation nicht zu vermindern.

Eigene Untersuchungen
141
3.1.3 Haltung, Fütterung und Transport
Mit Versuchsbeginn wurden die Tiere (ab minimal 5 kg) bis zu einer Lebendmasse von
30 kg in Gruppen von 6 - 8 Tieren gehalten. Sie erhielten einheitliches Futter ad libitum
nach den Vorgaben des „National Research Council“ (NRC, 1988). Mit Ausnahme der
in Beltsville geborenen „Poland China x Landrasse“-Kreuzungstiere und den aus
Maryland bezogenen reinrassigen US-Landrasse-Tieren stammten alle Versuchstiere aus
der Versuchsfarm der Iowa State University.
Alle Schweine ab 30 kg bis 90 kg Lebendmasse waren in Einzelbuchten auf
Teilspaltenboden untergebracht. Die Fütterung erfolgte mit einer einheitlichen Ration,
die 17,62 % Rohprotein und eine verdauliche Energie (DE) von 14,78 MJ/kg enthielt. In
Abhängigkeit von den wöchentlich gemessenen Lebendmassen erhielten die Tiere eine
Ration, die 95 % der geschätzten freiwilligen Futteraufnahme deckte. Unter
Berücksichtigung der bekannten Unzulänglichkeiten (WHITTEMORE et al., 1995) bildete
die Formel des „National Research Council“ (NRC, 1988) die Grundlage für die
Futtergabe: DEIntake (MJ/Tag) = 55 • (1 - e -0,0176 • BW).
Für ein 30 kg Schwein ergibt sich entsprechend eine erwartete Futteraufnahme von 1,53
kg/Tag [60 kg à 2,43 kg/d und 90 kg à 2,96 kg/d]. Die tatsächliche individuelle
Futteraufnahme wurde täglich erfasst (gewogen), indem die Differenz aus erfolgter
Futterzuteilung (kg) und Restfutter (kg) am Folgetag gebildet wurde. Der
durchschnittliche tägliche Futterverzehr in den Versuchsabschnitten [30à60 kg und
60à90 kg] ergibt sich aus dem Quotienten:
Gesamtfutterverzehr im Versuchsabschnitt / korrespondierende Anzahl Prüftage.
Vor jedem MR- bzw. DXA-Experiment1 wurde den Tieren das Futter für etwa 18
Stunden entzogen.
Für alle MR-Experimente wurden die Tiere von den Stallgebäuden des
Agriculture Research Service (ARS) in Beltsville, MD, zum Howard University
Hospital (Department of Radiology) nach Washington, DC, gefahren. Die Tiere bis 30
kg Lebendmasse waren in entsprechenden Tiertransportbehältern untergebracht,
während für die Tiere ab 30 kg Lebendmasse ein spezieller Tiertransporter zur
Verfügung stand. Die DXA-Untersuchungen erfolgten im Spezialcontainer in
unmittelbarer Stallnähe.
1 Die Experimente wurden in Übereinstimmung mit den durch das “USDA-ARS, Beltsville Animal Care and Usecommittee“ bestätigten Versuchsprotokollen durchgeführt.

Eigene Untersuchungen
142
3.1.4 Referenzschlachtung
Nach Abschluss der MR- bzw. DXA-Untersuchungen wurden die Probanden mit einer
letalen Pentobarbital-Injektion euthanasiert und bis 24 Stunden vor der Schlacht-
körperzerlegung bei –20 °C tiefgefroren.
Für Referenzzwecke wurden in jeder Gewichtsgruppe 15 bzw. 17 Schweine
geschlachtet. Vom Schlachtkörper wurden Kopf und Innereien separiert. Danach wurde
der Schlachtkörper entlang der Wirbelsäule in zwei Hälften geteilt. Die linke
Schlachthälfte wurde immer von derselben Person in die USDA-Teilstücke Blatt
(blade), Schulter (picnic shoulder), Rücken (loin), Seite (side) und Schinken (ham)
zerlegt (Abb. 3.1.18). Die Vorder- und Hinterfüße verblieben an der Schulter bzw. dem
Schinken. Die rechte Schlachthälfte inklusive Kopf, Gastrointestinal- und Urogenital-
Trakt (alle Innereien) wurde nach der Wägung zweimal komplett in einem Ganzkörper-
Wolf (Modell 810, GH, Astoria, OR, USA) unter Verwendung eines 0,32 cm „Siebes“
homogenisiert. Repräsentative Proben aus dem Homogenat dienten der chemischen
Bestimmung von Rohfett [Lipid] (Chloroform-Methanol-Extraktion nach der Methode
von FOLCH et al., 1957), Rohprotein (Kjeldahl-Stickstoff nach AOAC, 1984:
[N]1 • 6.25) und Rohasche (Restgewicht nach Erhitzung im Muffelofen für 24 Stunden
bei 520 °C). Etwa 500 g des homogenisierten Materials wurden verwendet, um die
Protein- und Fettanalyse durchzuführen. Die Proteinbestimmung erfolgte anhand von
drei jeweils 2 bis 3 g wiegenden Zufallsstichproben, während für die Lipidbestimmung
5 bis 7 g wiegende Proben (3) entnommen wurden. Der Wassergehalt wurde aus dem
Gewichtsverlust nach 10 Tagen Gefriertrocknung anhand einer zweiten 500 g Probe des
homogenisierten Materials ermittelt. Alle chemischen Untersuchungen wurden am
Growth Biology Laboratory des ARS durchgeführt.
Gleichzeitig wurden die Massen von Herz, Leber, Nieren und verbleibenden
Innereien erfasst. Zusätzlich dienten auf Transparentpapier abgezeichnete Aussenkanten
des M. longissimus dorsi und der darüber liegenden Fettschicht am Anschnitt zwischen
13. und 14. Rippe der linken Schlachthälfte zur Messung der entsprechenden Flächen
mittels eines Sigma-Plot-Digitalisierungstabletts (Jandel, Corte Madera, CA).
1 Kjeldahl-Stickstoff = Summe von Norganisch. und NH4-N

Eigene Untersuchungen
143
Abb. 3.1.18: Schema für Teilzerlegung (Vorder- bzw. Hinterfüße verblieben amSchlachtkörper)
In die Referenzuntersuchung gingen insgesamt 63 Tiere ein. Dabei lagen jeweils
für zwei Tiere der 10 kg Gruppe Werte entweder allein aus der MR-Tomographie oder
der DXA-Analyse vor.
Nach Ermittlung der Körperzusammensetzung für die Tiere der 10 kg Gruppe im
4,7 Tesla Magneten und im Lunar DPX-L-Scanner© wurden insgesamt 16 Tiere für
Referenzzwecke geschlachtet. Nur 14 Referenztiere der 10 kg Gruppe wurden sowohl
tomographiert als auch mittels DXA analysiert. Nach der Untersuchung von 47 Tieren
im 1,5 Tesla Ganzkörpertomographen und mit dem Lunar DPX-L Scanner dienten 15
Tiere in der 30 kg Gruppe, 15 Tiere in der 60 kg Gruppe und die verbleibenden 17 Tiere
in der 90 kg Gruppe als Referenztiere. Die Aufteilung nach Rasse-, Linienzugehörigkeit
und Geschlecht innerhalb der Lebendmassegruppen geht aus den Tabellen 3.1.10 bis
3.1.13 hervor.
Tab. 3.1.10: Anzahl Referenztiere je RyR1-Genotyp und Lebendmassegruppe
NN Nn nn total
10 kg 4 5 7 16DXA 4 4 7 15MRT 4 5 6 15
30 kg 2 11 2 15
60 kg 8 5 2*) 15
90 kg 6 6 5 17
total 20 27 16 63* Ein männlicher Kastrat der Linie „Hampshire“ (nn) wurde zwar bei 60 kg mittels DXA, aber nur bei 30kg mittels MRT untersucht, da dieses Tier vor der geplanten MR-Tomographie bei 60 kg verendete.

Eigene Untersuchungen
144
Tab. 3.1.11: Linien- und Geschlechterverteilung innerhalb RyR1-Genotyp für
Referenztiere (10 kg)
RyR1-
Genotyp
Rasse/Linie Anzahl männlich
(kastriert)
weiblich
Duroc 1 - 1
US-Landrasse 1 - 1
Poland China x Landrasse 2 - 2NN
Σ 4 - 4
Duroc 2 1 1
US-Landrasse 1 - 1#)
Spotted 2 1 1Nn
Σ 5 2 3
Duroc 3 1 (intakt) 2
Hampshire 4 - 4+)nn
Σ 7 1 6
total 16 3 13
+) Ein (1) Tier ohne Daten aus dem 4,7 Tesla MR-Tomographen#) Tier ohne Daten aus dem Lunar-DPX-L- Scanner

Eigene Untersuchungen
145
Tab. 3.1.12: Linien- und Geschlechterverteilung innerhalb RyR1-Genotyp für
Referenztiere (30 - 90 kg)
RyR1-
Genotyp
Rasse/Linie Anzahl männlich
(kastriert)
weiblich
Duroc 6 2 4
US-Landrasse 7 - 7
Poland China x Landrasse 3 - 3NN
Σ 16 2 14
Duroc 8 6 2
Hampshire 6 1 5
US-Landrasse 1 - 1
Poland China x Landrasse 4 - 4
Spotted 3 3 -
Nn
Σ 22 10 12
Duroc 7 4 3
Hampshire 2 1*) 1nn
Σ 8 5 4
total 47 17 30
*Ein männlicher Kastrat der Hampshire-Linie, RyR1-Genotyp = nn, verendete vor der MR-Tomographie(nach der DXA-Untersuchung) in der 60 kg Gruppe, so dass für dieses Tier nur MR-Messwerte bei ca. 30kg Lebendmasse vorliegen.
Tab. 3.1.13: Tieranzahl in Abhängigkeit von Rasse- bzw. Linienzugehörigkeit und
Lebendmassegruppe im Wachstumsversuch (DXA)
US-
Landrasse
Duroc Hampshire Poland China x
US-Landrasse
Spotted
10 kg 8 29 10 9 5
30 kg 8 21 8 7 3
60 kg 8 14 4 5 1
90 kg 2 9 2 4 -

Eigene Untersuchungen
146
3.1.5 Bestimmung der Muskelfaserstruktur mittels Schussbiopsie
An insgesamt 22 Schweinen (7 NN - 10 Nn - 5 nn) wurden parallel zu den DXA- und
MRT-Untersuchungen Muskelproben mittels Schussbiopsie in vivo gewonnen
(SCHÖBERLEIN, 1976). Die Biopsie erfolgte aus dem rechten M. longissimus dorsi auf
Höhe des 13./14. Brustwirbel ca. 3 – 5 cm lateral der Wirbelsäule.
Die durchschnittliche Lebendmasse der Tiere betrug 83 kg bei einem
Durchschnittsalter von 150 Tagen, wobei 6 Tiere aus der 60 kg Gruppe und 16 Tiere aus
der 90 kg Gruppe stammten. Diese Tiere wurden bereits bei ca. 10 kg Lebendmasse zur
Ermittlung der 31P-MR-Stoffwechselreaktion spektroskopiert und unmittelbar vor der
Biopsie zur Bestimmung der Körperzusammensetzung tomographiert (1H). Zum
Zeitpunkt der Biopsie waren die Tiere deswegen noch sediert, auch wenn eine vorherige
Fixierung und Sedierung der Tiere für die Biopsie nicht nötig gewesen wäre. Aufgrund
der Gerätekonstruktion dauert die Probenahme (inklusive Haut und subkutaner
Fettschicht) nur ein Sekundenbruchteil (< 1/100 Sekunde). Nach der Biopsie wurde die
Probeentnahmestelle lokal mit Wundspray behandelt.
Abb. 3.1.19: Verwendetes Schussbiopsiegerät LS1 mit demontiertem Aufsatz und inAktion (rechts im Bild)
Sofort nach der Biopsie wurden die Proben mit Nylonfaden auf Holzspateln fixiert, in
flüssigem Stickstoff schockgefroren und bei -70 °C bis zur Weiterverarbeitung am Meat
Science Laboratory des ARS/USDA in Beltsville, MD, USA gelagert.

Eigene Untersuchungen
147
1 cm
Abb. 3.1.20: Beispiel für ein frisch entnommenes Bioptat aus dem M. longissimus dorsieines SchweinsDeutlich zu erkennen sind Haut (mit Borsten), Fett- und Muskelgewebe - von links nachrechts.
Zur Weiterverarbeitung wurden die tiefgekühlten Proben gleichmäßig auf eine
Temperatur von -20 °C „erwärmt“. Mit einem Microtom (Cryostat 2800 Frigocut-E,
Reichert-Jung, Cambridge Instruments, New York) wurden 10 µm dicke Gefrierschnitte
angefertigt. Histochemisch wurden die Gefrierschnitte zur Fasertyp- und
Faserquerschnittsflächenanalyse mit Hilfe eines simultanen Kombinations-
färbeverfahrens nach SOLOMON und DUNN (1988) aufbereitet. Der (sauren)
Präinkubation bei pH = 4,30 folgte eine Inkubation für Succinatdehydrogenase-Aktivität
mit einer anschließenden Anfärbung für myofibrilläre Adenosintriphosphatase-Aktivität
und einer abschließenden Gegenfärbung mit Hämatoxylin. Die mit einem „Zeiss
Standard #16 Photomikroskop“ (Carl Zeiss, New York) erzeugten Fotografien der
histochemisch aufbereiteten Proben wurden mit einem handelsüblichen Scanner
digitalisiert und anschließend mit Hilfe der Bildverarbeitungssoftware OPTIMAS 4.10
ausgewertet. Für jedes Foto existieren Kalibrierungsaufnahmen, so dass die
Faserflächen exakt bestimmt werden konnten. Die Muskelfasertypen (Abb. 3.1.21) und
Muskelfaserquerschnittsflächen wurden an mindestens 300 Muskelfasern/Probe (Tier)
bzw. einer Mindestfläche von 2 400 000 µm2 ermittelt.
Unabhängig von der verwendeten histochemischen Methode werden die drei im
Versuch unterschiedenen Hauptfasertypen als Typ I (im englischen Sprachraum häufig
als: „slow-twitch-oxydative - STO), Typ IIa, („fast-twitch-oxydative“ - FTO) und Typ
IIb („fast-twitch-glycolytic“ - FTG) bezeichnet. In Abb. 3.1.21 sind die wichtigsten
Eigenschaften der drei unterschiedenen Muskelfasertypen zusammengefasst.

Eigene Untersuchungen
148
T y p I (S T O ): - la n g s a m e K o n trak tio n“ ro t ”„ β r ed “M H C - 1 *)
- s e h r h o h e o x ida t ive K a p a z i t ä t- m o d e r a t e r G l y k o g e n g e h a lt u n d
m o d e ra te g l y k o ly t i sche K a p a z itä t- s e h r h o h e r T rig lyze r idgeha l t- s e h r h o h e K a p illa rd i ch t e
T y p I Ia (F T O ): - s c h n e lle K o n tra k tio n
“ in te rm e d iä r”„ α r ed “M H C -IIA
- h o h e o x ida t ive K a p a z i t ä t- h o h e r G l y k o g e n g e h a lt u n d h o h e
g lyko ly t i s che K a p a z itä t- m o d e r a t e r T r i g l y z e r i d g e h a l t- h o h e K a p illa rd i ch t e
T y p IIb ( F T G ): - s c h n e lle K o n tra k tio n
“ w e i ß ”„ α w h ite “M H C -IIB
- g e r i n g e o x ida t ive K a p a z i t ä t- h o h e r G l y k o g e n g e h a lt u n d h o h e
g lyko ly t i s che K a p a z itä t- ge r inge r T rig lyze r idgeha l t- m o d e r a t e K a p illa rd i ch t e
*) M H C è M y o s i n H e a v y C h a i n I s o f o r m
Typ ITyp IIa
Typ IIb
Abb. 3.1.21: Im Versuch unterschiedene Muskelfasertypen
Die Auswertung der Daten für die Merkmale Muskelfaserquerschnittsfläche der
verschiedenen Muskelfasertypen und prozentuale Muskelfasertypanteile (bezogen auf
die analysierte Muskelfaserfläche bzw. auf die Anzahl analysierter Muskelfasern)
erfolgte nach dem folgenden statistischen Modell:

Eigene Untersuchungen
149
Yijkl = Beobachtungswert des m-ten Tieres
µ = Erwartungswert von Y
Ri = fixer Effekt des Ryanodin-Rezeptor-Genotyps
Lj = fixer Effekt von Linie/Rasse
Sk = fixer Effekt des Geschlechts
A ijkl = Regression auf Alter (Kovariable)
KM ijkl = Regression auf Körpermasse (Kovariable)
eijkl = Restfehler.
Yijklm= µ + R i + Lj + Sk+ ßAlter (Aijkl -A) + ßKM(KMijkl -KM)+ eijkl
Alter und Körpermasse befinden sich nicht im Modell für das Merkmal
Muskelfaseranzahl, da sich die Muskelfaseranzahl mit steigendem Alter und steigender
Körpermasse nicht verändert.

Eigene Untersuchungen
150
3.1.6 DNA-Analyse für den Ryanodin-Rezeptor-1-Gentest
Polymerasekettenreaktion
Die Polymerasekettenreaktion (PCR) ist eine universell einsetzbare, enzymatische
Methode zur spezifischen Vervielfältigung eines bestimmten DNA-Segments.
Eine Matrizen-DNA ("template") wird mit einem „Mastermix“ aus dNTPs, einem
spezifischen Oligonukleotid-Primerpaar und einer thermostabilen DNA-Polymerase
(z.B. Taq + Coenzym MgCl2 + pH-stabilisierenden Puffer) einem dreistufigen
Temperaturprofil ausgesetzt. Durch Denaturierung, Primer-Hybridisierung und
Elongation wird dabei die Zielsequenz der DNA mit jedem Zyklus verdoppelt. Ein
definiertes DNA-Segment wird dabei so angereichert, dass eine weitergehende Analyse
oder methodische Verwendung des PCR-Produkts leicht möglich ist, z.B. für DNA-
Sequenzierung, Restriktionsverdau, Sonden für Hybridisierungen, Mutationsanalyse
oder Klonierung (NEWTON und GRAHAM, 1994, WOLZ, 1999).
Extraktion der DNA für die PCR aus Blut
Die aus einer Ohrvene gewonnene Blutprobe wird mit EDTA konserviert. Das EDTA-
Blut (250 µl) wird in einem 1,5 ml Mikrozentrifugenröhrchen mit 250 µl Lysis-Puffer
durch Invertieren gemischt. Nach 20 sec Zentrifugation bei 13 000XG wird der
Überstand abpipettiert, das gewonnene Pellet in 1 ml Lysis-Puffer durch Schütteln auf
dem Vortex resuspendiert, erneut zentrifugiert und der Überstand erneut abpipettiert.
Dieser Vorgang muss mindestens zweimal oder bis zur Gewinnung eines farblosen
Pellets wiederholt werden. Das so gewonnene Pellet wird in 250 µl PCR-Puffer mit
nichtionischen Detergenzien und Proteinase K auf dem Vortex geschüttelt und im
Wasserbad oder Heizblock bei 50 - 60 °C für 1 Stunde inkubiert. Nach der Proteolyse
muss zur Inaktivierung der Proteinase K das Röhrchen mit der in Lösung gegangenen
DNA bei 95 ºC für 10 min inkubiert werden. In 25 µl der entstandenen DNA-Lösung
sind ca.1 µg genomische DNA enthalten.
Polymerasekettenreaktion für die Ryanodin-Rezeptor-1-DNA
Für die PCR wird ein Mastermix von 34 µl pro Probe angesetzt (für komplettes
Laborprotokoll siehe Anhang III). Die bei –20 ºC gelagerten Bestandteile des
Mastermixes werden auf Eis aufgetaut und kurz auf dem Vortex geschüttelt (außer Taq-
Polymerase). In zügiger Reihenfolge wird der Ansatz (als letztes immer die Taq-
Polymerase - sehr reaktionsfreudig!) auf Eis pipettiert und kurz „gevortext“.

Eigene Untersuchungen
151
Folgende Komponenten ergeben den Mastermix (40 µl Gesamtvolumen =
34 µl Mastermix + 6 µl DNA):
22 µl H2O
+ 4 µl 10 x Puffer II (PCR 103, GeneAmp® - PERKIN ELMER)
+ 4 µl (2 mM) dNTPs
+ 1 µl 15 mM MgCl2
+ 1 µl Primer I Oligo Name: 17824 (BioServe):
5'> GTG CTG GAT GTC CTG TGT TCC CT <3';
+ 1 µl Primer II Oligo Name:17825 (BioServe):
5'> CTG GTG ACA TAG TTG ATG AGG TTT G <3'
+1 µl Taq-Polymerase (AmpliTaq® - PERKIN ELMER).
Der Mastermix wird in PCR-Röhrchen auf Eis pipettiert und 6 µl DNA-Suspension
hinzugefügt. Ein Überschichten der Proben mit 2 Tropfen Mineralöl kann ein
eventuelles Verdunsten verhindern (bei Heizblöcken ohne beheizten Deckel nötig).
Für die Amplifikation eines 134bp-Fragments der Ryanodin-Rezeptor-1-DNA
wird der nach BRENIG and BREM (1992) bzw. FÖRSTER et al. (1992) modifizierte Ansatz
für ca. 4 ½ Stunden im Thermo-Cycler (Perkin Elmer 480) inkubiert (komplettes
Protokoll siehe Anhang III). Der verwendete Thermozyklus besteht aus:
1) Anfangsdenaturierung der DNA-Doppelstränge bei 94 ºC für 3 min;
2) 35 Zyklen mit:
⇒ Denaturierung der DNA-Stränge zur Erzeugung der Matrix für Primer
und DNA-Polymerase bei 94 ºC (1 min)
⇒ Anlagerung der Primer an die komplimentären Einzelstränge bei 55 ºC
(1 min 15 sec)
⇒ Verlängerung der Primer in 5'-3'-Richtung durch Taq-DNA-Polymerase
bei 72 ºC (1 min 15 sec);
3) Abschlusszyklus bei 72 ºC (4 min);
4) 4 ºC bis zur Herausnahme der PCR-Proben.
Typisierung
Die PCR-Suspension (20 µl) wird unter Verwendung von 2 µl Restriktions-enzym Cfo1
(GIBCO BRL) und 2 µl "Storage"-Puffer bei 37 °C mindestens 2 Stunden inkubiert.
Während der Inkubation werden die DNA-Fragmente geschnitten. Anschließend erfolgt
die Trennung der DNA-Fragmente in einem 3%-igen Agarose-Ethidiumbromid-Gel.
Unter UV-Licht können die RyR1-Genotypen typisiert werden (Abb. 3.1.22).

Eigene Untersuchungen
152
- heterozygoter Defektallelträger Nn:
⇒ 3 Banden: 134 bp, 84 bp, 50 bp (links)
- homozygoter Defektallelträger nn:
⇒ 1 Bande: 134 bp (Mitte)
- homozygot normal NN:
⇒ 2 Banden: 84 bp, 50 bp (rechts).
(links außen DNA-Längenstandard)
Nn nn NN
Abb. 3.1.22: PCR-Ergebnis für die RyR1-Genotypen
Links außen (Abb. 3.1.22) läuft zur Kontrolle ein DNA-Längenstandard VIII
(Boehringer Mannheim Biochemica) mit. Insgesamt lassen sich 13 DNA-Segmente
unterscheiden:
1114 bp, 900 bp, 692, [501 bis 489] bp, 404 bp, 320 bp, 242 bp, 190 bp, 147 bp, 124 bp,
110 bp, 67 bp und [37 bis 19] bp.
Die Abbildung 3.1.23 fasst den Ablauf der PCR-Analyse schematisch zusammen.
Abb. 3.1.23: Prinzip des verwendeten RyR1-Gentestes (modifiziert nach FÖRSTER u.a.,1992 und GAISSER und HÜSING, 2000)

Ergebnisse
153
3.2 Ergebnisse
3.2.1 Muskelstoffwechseluntersuchungen3.2.1.1 Phosphorkomponenten - 31P-Magnetresonanz-Spektroskopie
Eine dramatische Veränderung der Muskelstoffwechselprozesse setzte erwartungsgemäß
im nn-Genotyp nach durchschnittlich nur 8 Minuten Halothangabe ein. Phosphokreatin,
ein wichtiger Indikator für energieverbrauchende Prozesse des Muskelstoffwechsels,
zeigte einen sehr schnellen und drastischen Konzentrationsabfall um 45 %, begleitet von
einem ebenso rasanten Absinken im pH-Niveau (Abb. 3.2.1, 3.2.2, Tab. 3.2.1).
Gleichzeitig stiegen die Konzentration von anorganischem Phosphat um 166 % sowie
die Körpertemperatur um 0,81 °C an, während sich die ATP-Konzentration verminderte.
Im Gegensatz zum nn-Genotyp veränderte sich die Stoffwechsellage der NN-Tiere sehr
langsam. Nur minimale Veränderungen konnten in der Phosphokreatin- und ATP-
Konzentration nach durchschnittlich 51 Minuten Halothangabe beobachtet werden
(Abb. 3.2.1, 3.2.2, Tab. 3.2.1). Das Niveau von anorganischem Phosphat stieg um
signifikant 86 %, während der pH-Wert auf 6,95 und die Körpertemperatur um 0,71 °C
im homozygot normalen Genotyp absank. Der heterozygote Genotyp reagierte in vivo
insgesamt intermediär zwischen den beiden homozygoten Genotypen auf die
Halothanbelastung. Phosphokreatin fiel auf 84,06 % des Ruheniveaus ab - ohne eine
signifikante Veränderung in der ATP-Konzentration oder der Körpertemperatur
hervorzurufen. Der minimale pH-Wert endete bei 6,91. Berücksichtigt man die Dauer
der Halothangabe, tendierte der heterozygote Genotyp in vivo jedoch mehr zu NN.
Zu Beginn des Versuches wurden zwischen den drei Ryanodin-Rezeptor-
Genotypen keine signifikanten Unterschiede im Muskel-pH-Wert festgestellt (Tab.
3.2.1). Im Versuchsverlauf verminderten sich pH-Wert und Phosphokreatin zumeist
gleichzeitig. Der pH-Wert erreichte seinen Tiefpunkt jedoch in den häufigsten Fällen 5-
10 Minuten später als Phosphokreatin im homozygot normalen bzw. heterozygoten
Genotyp. Bereits vor einer Konzentrationsveränderung im Phosphokreatin kann jedoch
eine pH-Absenkung vor allem im NN-Genotyp und zum Teil in den Nn-Probanden
beobachtet werden (Abb. 3.2.2). Demzufolge könnte die Energiebereitstellung zunächst
über Glukose oder/und Glykogen mit anschließender Laktatanhäufung erfolgen.

Ergebnisse
154
Tab. 3.2.1: Vergleich von Stoffwechselparametern aus der 31P-MR-Spektroskopie (PCr,
pH, ATP, Pi) sowie für die Körpertemperatur nach Halothanbelastung in Schweinen mit
unterschiedlichen RyR1-Genotypen (LSM ± SEE#)
Ryanodin-Rezeptor-1-Genotypen
Merkmal Status beiVersuchsende
NN Nn nn
in vivo 3,76 ± 4,94 a 15,94 ± 3,76 b 45,62 ± 5,82 cPCr-Abbau %
post mortem 61,76 ± 9,92 a 74,85 ± 8,79 ab 93,26 ± 10,61 b
in vivo 7,05 ± 0,01 7,05 ± 0,01 7,06 ± 0,02 pH-Start
post mortem 7,05 ± 0,03 7,05 ± 0,03 7,08 ± 0,03
in vivo 6,95 ± 0,03 6,91 ± 0,03 6,88 ± 0,04 pH-Minimum post mortem 6,86 ± 0,07a 6,64 ± 0,06 b 6,25 ± 0,07 c
in vivo +4,91 ± 3,59 a +0,39 ± 2,74 a -19,61 ± 4,23 b ATP-Verände-rung (%) post mortem +4,56 ± 7,22 a -24,07 ± 6,39 b -55,80 ± 7,72 c
in vivo 85,85 ± 32,50 ab 80,43 ± 24,75a 166,47 ± 38,31 bPi-Anstieg(%) post mortem 695,56 ± 65,29 a 643,91 ± 57,83a 415,48 ± 69,83 b
in vivo -0,71 ± 0,31 a -0,00 ± 0,24 b +0,81 ± 0,34 cKörper-temperatur (°C) post mortem -0,24 ± 0,71 a +2,75 ± 0,58 b +5,17 ± 0,62 c
in vivo 50,71 ± 3,45 a 49,16 ± 3,16 a 8,08 ± 4,37 bHalothangabe(Minuten) post mortem 41,00 ± 9,12 a 40,50 ± 7,89 a 10,00 ± 9,12 b
in vivo 21 25 13n
post mortem 3 4 3
#) Unterschiedliche Buchstaben innerhalb einer Zeile kennzeichnen Signifikanz (p ≤ 0,05).
In den (homozygoten) Defektallelträgern sind hingegen keine zeitlichen
Verschiebungen zwischen pH-Absenkung und Phosphokreatinabbau zu verzeichnen.
Der durch die Halothanbelastung hervorgerufene Energiesog führt zu einem nahezu
gleichzeitigen Verbrauch aller verfügbaren Ressourcen wie Glykogen, Glukose und

Ergebnisse
155
Phosphokreatin. Wenn diese Ressourcen stark abgenommen haben und nicht mehr
regeneriert werden können, sinkt auch die ATP-Konzentration.
- NN-- NN- - Nn -- Nn - - nn -- nn -
Abb. 3.2.1: Beispiel für den zeitlichen Verlauf der 31P-MR-Spektroskopie inunterschiedlichen RyR1-Genotypen (modifiziert nach Scholz et al., 1995)
Zusätzlich zum Experiment in vivo (Abb. 3.2.1, 3.2.2) wurde der
Muskelstoffwechsel bei 10 Probanden (3 NN - 4 Nn - 3 nn) nach einer Überdosierung
von Halothan post mortem (nach dem Einstellen der Atmung) weiterbeobachtet (Abb.
3.2.3). Spätestens eine Stunde nach Beendigung der Halothanadministration oder nach
dem völligen Abbau von Phosphokreatin wurde das Experiment beendet.
Das Phosphokreatinniveau verminderte sich im NN-Genotyp langsamer als in den
beiden anderen Genotypen. Eine Stunde nach Beendigung der Halothanzufuhr war
Phosphokreatin (fast) vollständig im homozygot „defekten“ Genotyp aufgebraucht.
Anorganisches Phosphat (Pi) erreichte ein Plateau. Überraschenderweise stieg post
mortem die Konzentration des anorganischen Phosphats im homozygot normalen
Genotyp jedoch stärker als in den Defektallelträger-Genotypen an (Tab. 3.2.1, Abb.
3.2.3). Diese Reaktion könnte eine Erklärung für die höhere Kapazität in der Ca2+-
Kompensation des „normalen“ Genotyps sein. Anorganisches Phosphat ist ein
“Feedback”-Regulator für die Ca2+-ATPase-Pumpe und hilft post mortem die
Speicherung von Ca2+ im sarkoplasmatischen Retikulum anzuregen bzw.
aufrechtzuerhalten (NEWBOLD and TUME, 1981).

Ergebnisse
156
01020
30405060
7080
10Halothangabe: Start
20 30 40 50 60 70Ende
80 90 100 110 120 Minuten
6,94
6,98
7,02
7,06
7,10
7,14
pH
Pi PCr Temperatur °C ATP pH-NN-INTERNE
EINHEITEN
0
10
20
30
40
50
60
0 10Halothangabe: Start
20 30 40 50 60 70 80 Minuten6,90
6,94
6,98
7,02
7,06
7,10
pH
Ende
-Nn-INTERNE
EINHEITEN
0
10
20
30
40
50
60
70
10Halothan: Start Ende
20 30 40 50 60 Minuten6,70
6,756,80
6,856,90
6,957,00
7,057,10
7,15
pH
-nn-INTERNE
EINHEITEN
Abb. 3.2.2: Beispiel für die Veränderung der Stoffwechselsituation in vivo nachHalothanbelastung in unterschiedlichen RyR1-Genotypen (modifiziert nach SCHOLZ etal., 1995)

Ergebnisse
157
0
10
20
30
40
50
60
10Halothangabe: Start
20 30 40 50 60 70Ende
80 90 100 110 120 Minuten6,75
6,80
6,85
6,90
6,95
7,00
7,05
pH
Pi PCr ATP Temperatur °C pH-NN-INTERNE
EINHEITEN
0
10
20
30
40
50
60
Halothangabe: Start20 30 40 50 60 70 80 90 100 110 Minuten
6,356,456,556,656,756,856,957,057,15
pH
Ende10
-Nn-INTERNE
EINHEITEN
0
10
20
30
40
50
60
6,20
6,40
6,60
6,80
7,00
7,20
pH
10 20 30 40 50 60 70 Minuten
Halothangabe: Start Ende
-nn-
INTERNE
EINHEITEN
Abb. 3.2.3: Beispiel für Stoffwechselveränderungen post mortem im Vergleich der dreiRyR1-Genotypen (modifiziert nach SCHOLZ et al., 1995)

Ergebnisse
158
Die heterozygoten Schweine reagierten intermediär auf die Halothanbelastung für
alle Merkmale, die post mortem analysiert wurden. Die Differenzen zwischen Nn und
NN waren signifikant für das pH-Minimum, die ATP-Konzentration- und die
Körpertemperaturveränderung - aber nicht signifikant für den Phosphokreatinabbau, den
Pi-Anstieg sowie für die Dauer der Halothangabe. Generell waren die Unterschiede
zwischen den drei Ryanodin-Rezeptor-1-Genoytpen post mortem größer als in vivo. Das
pH-Minimum, die ATP- und die Körpertemperaturveränderung sind dabei
hervorzuheben (Tab. 3.2.1).
Obwohl zwischen den RyR1-Genotypen für den ”Ruhestoffwechsel” (pH-Wert zu
Beginn des Experiments à pH-Start) keine Unterschiede festgestellt werden konnten
(Tab. 3.2.1), waren beim Vergleich der Linien Duroc, Hampshire, Landrasse, Poland
China x Landrasse und Spotted pH-Start-Differenzen mit dem höchsten Wert für
Spotted (alle Nn) zu verzeichnen. Die Linie Spotted reagierte auf die Halothangabe am
deutlichsten mit einem starken Absinken von PCr, ATP und pH bei einem parallelen
Anstieg von Pi sowie der Körpertemperatur. Im Gegensatz wurde der geringste
Rückgang im Phosphokreatin (~39 %) gekoppelt mit dem niedrigsten pH-Minimum
(6,696 ± 0,044) für Hampshire gemessen. Die Differenz zwischen pH-Start und pH-
Minimum war jedoch bei Spotted mit ∆pH = 0,405 am größten (Tab. 3.2.2).
Die extreme Reaktion von Spotted überraschte, da diese Linie - bei nicht
signifikanten Unterschieden im Volumen der Mm. longissimi dorsi – nach „Duroc x
Minzhu“ die stärkste Fettauflage besaß (Kapitel 3.2.2.1: Tab. 3.2.6). Da alle
untersuchten Probanden der Linie Spotted heterozygot im RyR1-Genotyp waren, liegt
hier eine dominante Wirkung des Defektallels oder eine Kopplung mit einem weiteren
Stressanfälligkeit und Körperzusammensetzung beeinflussenden Genort im Bereich des
Möglichen. Die im statistischen Modell enthaltenen Linieneffekte berücksichtigen
jedoch bereits den Einfluss von RyR1-Genotyp, Geschlecht, Zustand bei Versuchsende
sowie die Interaktion zwischen Zustand bei Versuchsende und RyR1-Genotyp. Weitere,
speziell die Körperzusammensetzung beeinflussende Genorte und Mechanismen werden
im Kapitel 4.2 diskutiert, während im Kapitel 4.1 näher auf die Wirkungsweise der
verschiedenen Ryanodin-Rezeptor-1-Genvarianten eingegangen wird.

Ergebnisse
159
Tab. 3.2.2: Vergleich von Stoffwechselparametern aus der 31P-MR-Spektroskopie (PCr,
pH, ATP, Pi) und der Körpertemperatur nach Halothanbelastung in Schweinen mit
unterschiedlicher Rasse- bzw. Linienzugehörigkeit (LSM ± SEE#)
Duroc Hampshire Landrasse Poland China
x Landrasse
Spotted
PCr-Abfall(%)
50,34 ± 4,48 ab 39,03 ± 6,56 a 45,43 ± 6,91 ab 44,43 ± 7,67 ab 66,68 ± 9,16 b
pH- Start
7,039 ± 0,011ab 7,038 ± 0,017 ab 7,030 ± 0,018 a 7,071 ± 0,019 bc 7,109 ± 0,023 c
pH-Minimum
6,737 ± 0,030 a 6,696 ± 0,044 a 6,759 ± 0,047ab 6,845 ± 0,052 b 6,704 ± 0,062 ab
ATP-Verän-derung (%)
-11,89 ± 3,19 a -11,02 ± 4,67 a -6,94 ± 4,92 a -8,09 ± 5,46 a -35,69 ± 6,52 b
Pi-Anstieg(%)
333,02 ± 29,11 a 357,74 ± 42,65 a 264,98 ± 44,91 a 264,00 ± 49,87 a 511,38 ± 59,55 b
Körpertemp-eratur (°C)
+0,69 ± 0,23 a +0,71 ± 0,37 a +1,16 ± 0,52 a +0,49 ± 0,43 a +3,41 ± 0,53 b
Halothan(Minuten)
37,48 ± 3,41 a 40,73 ± 5,07 a 29,77 ± 8,05 ab 20,57 ± 6,36 b 22,87 ± 7,99 ab
n 29 12 14 9 5
#) Unterschiedliche Superskripts innerhalb einer Zeile kennzeichnen Signifikanz (p ≤ 0,05).

Ergebnisse
160
3.2.1.2 Kohlenstoffkomponenten - 13C-Magnetresonanz-Spektroskopie
Da in Bezug auf den RyR1-Genotypeffekt nur minimale Unterschiede in den
Ergebnissen zwischen den beiden Varianzanalyse-Modellen I (Anhang II, Tab. A13)
und II (Tab. 3.2.3) bestehen, das Modell II jedoch eine zusätzliche Berücksichtigung des
Hampshireeffekts beinhaltet, wird – wenn nicht anders gekennzeichnet - in der
nachfolgenden Ergebnisinterpretation immer das Modell II betrachtet. Speziell für das
Glykogenniveau resultierte das Modell II bei identischem Bestimmtheitsmaß bzw.
Reststandardabweichung für das Gesamtmodell (R2 = 0,84, se = 9.10) in geringeren
Standardschätzfehlern (SEE) der „Kleinsten Quadrate Mittelwerte“ (LSM).
Neben dem signifikanten Effekt des RyR1-Genotyps (p = 0,007) hatte das
Geschlecht (p = 0,0057) einen maßgeblichen Einfluss auf das „Ruhe”-Glykogenniveau.
Allein durch diese beiden Effekte wurden 41,5 % der Variation im „Ruheniveau” des
Glykogens erklärt (p = 0,0056). Der Hampshire-Genanteil zeigte keinen signifikanten
Effekt auf das „Ruhe“-Glykogenniveau. Nur in der Tendenz wiesen die Tiere mit dem
Hampshire-Genanteil (50 %) ein höheres „Ruhe“-Glykogenniveau auf als die
Tiere ohne Hampshire-Genanteil bzw. mit theoretisch 25 % Hampshire-Genanteil (Tab.
3.2.4).
Weibliche Tiere verfügten zu Beginn des Versuchs über signifikant mehr
Glykogen (74,87 ± 3,95 µmol/g) als männliche Tiere (59,04 ± 2,99 µmol/g), zeigten
nach Halothangabe jedoch nur in der Tendenz einen geringeren prozentualen
Glykogenabbau als die männlichen Tiere (-35,73 ± 4,71 % zu -44,87 ± 3,37 %). Durch
das stark voneinander abweichende Startniveau ergaben sich jedoch Differenzen im
absoluten Glykogenniveau am Ende des Versuchs mit 48,12 µmol/g für die weiblichen
Tiere und 32,54 µmol/g für die männlichen Tiere, da beide Geschlechtsgruppen fast
identisch 26,7 bzw. 26,5 µmol Glykogen je Gramm Muskelgewebe über einen Zeitraum
von ca. 130 Minuten abbauten. Die durchschnittliche Abbaurate nach ca. 25 Minuten
Halothanbelastung entsprach 0,20 µmol/g • min-1.
Drastische Veränderungen der Muskelstoffwechselprozesse nach durchschnittlich
nur 8 Minuten Halothangabe (in vivo und post mortem) traten erwartungsgemäß im
Genotyp nn auf. Die Glykogenkonzentration als ein wichtiger Indikator für
energieverbrauchende Prozesse des Muskelstoffwechsels sank schnell ab und war mit
einem leichten Anstieg von Kreatin gekoppelt, während der heterozygote Genotyp (Nn)

Ergebnisse
161
erst nach längerer Halothangabe einen starken Abfall im Glykogenniveau zeigte (Tab.
3.2.3, Abb. 3.2.4).
Tab. 3.2.3: Vergleich der Ergebnisse aus der 13C-MR-Spektroskopie für die RyR1-
Genotypen (LSM ± SEE#)
NN Nn nn
„Ruhe”-Glykogen (µmol/g)+ 72,21 ± 4,41a 73,36 ± 4,22 a 55,31 ± 3,71 b
n 7 10 10
Glykogen in vivo (%) -19,10 ± 4,36 a -35,03 ± 3,37 b -33,05 ± 3,80 b
Kreatin in vivo (%) -7,86 ± 12,74 -1,47 ± 9,83 +10,99 ± 11,09
Temperatur in vivo (°C) +0,28 ± 0,54 +1,36 ± 0,41 +1,59 ± 0,47
Halothan in vivo (min) 34,85 ± 5,28 a 21,19 ± 4,07 a 6,53 ± 4,60 b
n 6 8 7
Glykogen post mortem (%) -17,53 ± 11,56 a -73,42 ± 6,59 b -49,71 ± 10,29 ab
Kreatin post mortem (%) +24,94 ± 33,72 -41,49 ± 19,21 +18,89 ± 30,01
Temperatur post mortem (°C) -1,47 ± 1,42 a +5,00 ± 0,81b +5,85 ± 1,26 b
Halothan post mortem (min) 54,67 ± 13,98 25,98 ± 7,96 11,13 ± 12,44
n 1 2 3# Unterschiedliche Superskripts kennzeichnen Signifikanz bei p ≤ 0,05.+ Nur RYR1 und Geschlecht verblieben im Modell, bei allen anderen Merkmalen wurde derGeschlechtseffekt aufgrund fehlender Signifikanz nicht berücksichtigt.
-90
-80
-70
-60
-50
-40
-30
-20
-10
0
Veränderung von Glykogen (%)
NN
Nn
nn
in vivo post mortem (1h nach Halothan)
Abb. 3.2.4: Veränderung des prozentualen Glykogenniveaus nach Halothanzufuhr beiSchweinen unterschiedlicher RyR1-Genotypen (LSM, SEE)

Ergebnisse
162
Der Reaktionsbeginn scheint folglich von der Dauer der Halothangabe (Dosis) und
von der Anzahl der Defektallele abzuhängen. Im Gegensatz zu den Defektallelträgern
(Nn und nn) veränderte sich die Stoffwechsellage im NN-Genotyp nur sehr langsam und
ohne drastische Reaktionen in Bezug auf das Glykogenniveau.
Vorwiegend im NN-Genotyp und zum Teil bei Nn sank das Glykogenniveau auch
unabhängig von einer Veränderung im Kreatin ab (Abb. 3.2.4, 3.2.5, Tab. 3.2.3). Die
Energiebereitstellung erfolgte in diesen Fällen offenbar zuerst über Glukose bzw.
Glykogen und zeitlich leicht verzögert über Phosphokreatin. Diese Beobachtungen
bestätigen die Ergebnisse aus der 31P-Magnetresonanz-Spektroskopie (SCHOLZ et al.,
1995, Kapitel 3.2.1.1), in deren Verlauf bei einigen Tieren ein Absinken des pH-Wertes
- als direkter Indikator für den Verlauf der Glykolyse - unabhängig von einer
Konzentrationsveränderung im Phosphokreatin beobachtet werden konnte. Insgesamt
waren die Veränderungen im Kreatinniveau (Position: 157 ppm im 13C-MR-Spektrum)
in vivo nicht signifikant verschieden von Null (Tab. 3.2.3, Abb. 3.2.5) und unterlagen
großen Schwankungen. Unmittelbar nach Beginn der Halothanzufuhr stieg die
Kreatinkonzentration speziell bei einigen Tieren des Genotyps nn kurzzeitig schnell an,
was eine unverzügliche Beendigung der Halothanzufuhr nach sich zog, um den
Stoffwechsel nicht zum Entgleisen zu bringen. Dabei wurde davon ausgegangen, dass
der Kreatinanstieg einen Verbrauch von Phosphokreatin signalisierte. Unabhängig vom
zeitweiligen Anstieg der Kreatinkonzentration wird zunehmend Glykogen abgebaut
(Abb. 3.2.4, 3.2.5, 3.2.6, 3.2.7).
-80
-60
-40
-20
0
20
40
60
Veränderung von Kreatin (%)
NN
Nn
nn
in vivo post mortem (1h nach Halothan)
Abb. 3.2.5: Veränderung des prozentualen Kreatinniveaus nach Halothanzufuhr beiSchweinen unterschiedlicher RyR1-Genotypen (LSM, SEE)

Ergebnisse
163
Abbildung 3.2.6 stellt den durchschnittlichen zeitlichen Verlauf des
Glykogenabbaus für jeden RyR1-Genotyp korrigiert auf die anderen Einflussfaktoren
wie Linie (Hampshire- bzw. Pietrain-Genanteil) und Geschlecht dar. Alter und
Lebendmasse beeinflussten in den untersuchten Tieren den Glykogenabbau nicht
signifikant. Vom durchschnittlichen Verlauf können die individuellen Verläufe
abweichen, da einzelne Probanden in vivo nach Belastungsreaktion und Beendigung der
Halothangabe wieder das Ausgangsniveau (Ruhezustand) in den beobachteten
Merkmalen erreichten. Insbesondere im Genotyp nn ist nicht auszuschließen, dass diese
Tiere bereits während der „Ruhespektren”-Aufnahme (ohne Halothangabe) Anzeichen
einer Stressreaktion zeigten, da der „Ruhe”-Glykogengehalt (µmol/g) bereits signifikant
unter dem der beiden anderen Genotypen lag (Tab. 3.2.3).
Die Ergebnisse der Untersuchungen, die post mortem weiter geführt wurden,
lassen aufgrund der geringen Tierzahl keine eindeutigen Schlussfolgerungen zu
(Anhang II, Tab. A14), bestätigen in der Tendenz aber die Ergebnisse aus der Analyse in
vivo und decken sich für die Körpertemperaturveränderung mit den Ergebnissen aus der31P-MR-Spektroskopie (Tab. 3.2.3).
Bei der Gegenüberstellung der Stoffwechselreaktionen der verschiedenen
Hamphire-Genanteile (Tab. 3.2.4) wird deswegen auf einen Vergleich von in vivo und post
mortem erzielten Ergebnissen verzichtet. Die dargestellten Ergebnisse sind die Kleinsten
Quadrate Mittelwerte unter Berücksichtigung aller im Modell II (Kapitel 3.1.1.2)
dargestellten Effekte und Kovariablen.
Da in der Gruppe mit dem höchsten Hampshire-Genanteil (50 %) die
Wahrscheinlichkeit für das Vorhandensein des RN --Allels am größten ist, verzeichnete
diese Gruppe erwartungsgemäß den geringsten Glykogenabbau, obwohl tendenziell die
Körpertemperatur am stärksten anstieg. Beim Vergleich der drei Hampshire-
Genanteilgruppen lässt sich jedoch - im Gegensatz zur in Abbildung 3.1.7 dargestellten
linearen Abhängigkeit - keine lineare Beziehung zwischen Glykogenabbau und
Hampshire-Genanteil feststellen. Die Gruppen mit 0 % bzw. 25 % Hampshire-
Genanteilen unterschieden sich nicht im Glykogenabbau. Dieses Ergebnis lässt auf die
Abwesenheit des RN--Allels in den Gruppen mit 0 % bzw. 25 % Hampshire-
Genanteilen schließen (Tab 3.2.4).
Im Vergleich zum Modell II wurden im Modell I nicht die Hampshire- und
Pietrain-Genanteile, sondern die Effekte der vier Kreuzungslinien in das Modell

Ergebnisse
164
einbezogen. In Übereinstimmung mit dem Modell II verläuft der Glykogenabbau auch
im Modell I bei der Kreuzungsherkunft mit 50 % Hampshire-Genanteil am langsamsten,
während die beiden anderen Herkünfte mit jeweils 25 % Hampshire-Genanteilen (50 %
Duroc bzw. DHLSY) einen schnelleren Glykogenabbau aufweisen. Am schnellsten wird
Glykogen jedoch in der Linie Spotted (ohne Hampshire-Genanteil) abgebaut (Abb.
3.2.8, siehe auch Tab. 4.1.2).
Beginn der Halothangabe
Abb. 3.2.6: Zeitlicher Verlauf des absoluten Glykogenabbaus (µmol/g) nachHalothanbelastung in verschiedenen RyR1-Genotypen in vivoNach dem Beginn der Halothanzufuhr ab der 20. Minute betrug die durchschnittliche Dauer derHalothangabe für NN = 34,85, Nn = 21,19 und nn = 6,53 Minuten (Tab. 3.2.3).
Tab. 3.2.4: Vergleich der Ergebnisse aus der 13C-MR-Spektroskopie in Abhängigkeit
vom Hampshire-Genanteil (LSM ± SEE#)
Ha 0 % Ha 25 % Ha 50 %(4 Nn/ 7 nn) (3NN/ 4 Nn/ 2 nn) (4 NN/ 2 Nn/ 1nn)
„Ruhe”-Glykogen (µmol/g) 66,70 ± 4,36 65,44 ± 4,25 68,90 ± 4,85
Glykogen (%) -44,74 ± 4,45 ab° -48,50 ± 4,79 b -20,68 ± 9,47 a*
Kreatin (%) +15,93 ± 13,01 -6,16 ± 28,55 -16,36 ± 14,03
Temperatur (°C) +1,29 ± 0,55 +1,79 ± 0,59 +2,90 ± 1,21
Halothan (min) 23,02 ± 4,29 26,63 ± 4,19 22,53 ± 4,79
End-Glykogen (µmol/g) +) 36,86 33,70 54,65
n 11 9 7#) Unterschiedliche Superkripts kennzeichnen Signifikanz bei p ≤ 0,05Werte mit ° und * überschreiten das Signifikanzniveau minimal (p ≤ 0,07).+) berechnet aus „Ruhe”-Glykogengehalt und Glykogenabbau %

Ergebnisse
165
Abb
. 3.2
.7: B
eisp
iel f
ür d
en z
eitli
chen
Ver
lauf
der
13C
-MR
-Spe
ktre
n vo
n Sc
hwei
nen
unte
rsch
iedl
iche
r RyR
1-G
enot
ypen
(je
wei
ls 2
Spe
ktre
n ad
dier
t)
C1-
Gly
koge
nK
reat
in

Ergebnisse
166
70
60
50
40
30
20
10
0
62,5 % Duroc ≥50 % Hampshire DHLSY ≥50 % Spotted
Abb. 3.2.8: Glykogenabbau in unterschiedlichen Kreuzungsherkünften (Modell I)

Ergebnisse
167
3.2.2 Körperzusammensetzung
3.2.2.1 MRT-Lebendmassegruppe bis 20 kg
Bereits bei den jungen Versuchstieren mit durchschnittlich 53,69 Lebenstagen bei einem
durchschnittlichen Gewicht von 12,37 kg bestehen Unterschiede im Volumen der Mm.
longissimi dorsi zwischen den drei RyR1-Genotypen. Der nn-Genotyp verfügt bei
gleichzeitig niedrigstem Fett-zu-Muskel-Verhältnis in dem 2,45 cm MR-Abschnitt mit
50,96 cm³ über signifikant mehr Muskelvolumen als die beiden anderen Genotypen NN
und Nn. Keine signifikanten Unterschiede existieren im Volumen der Fettauflage über
der analysierten Mm.l.d.-Region, wobei auch hier überraschenderweise der nn-Genotyp
in der Tendenz die stärkste Fettauflage aufweist (Tab. 3.2.5).
Tab. 3.2.5: Volumen der Mm. longissimi dorsi und des darüber liegenden Fettes sowie
Fett-zu- Muskel-Verhältnis in einem 2,45 cm MRT-Abschnitt von Schweinen mit
unterschiedlichen RyR1-Genotypen
(LSM ± SEE#); korrigiert auf einheitliche Lebendmasse von 12,42 kg bei einem Alter von 53,78
Lebenstagen)
NN Nn nn
Volumen Mm.l.d. (cm³) 41,62 ± 1,57a 42,89 ± 1,32a 50,96 ± 2,06 b
Fett-Volumen (cm³) 16,19 ± 0,58 16,68 ± 0,48 17,39 ± 0,76
Fett-zu-Muskel-Verhältnis 0,397 ± 0,017a 0,367 ± 0,014ab* 0,327 ± 0,022 b°
n 37 46 28#) Werte mit unterschiedlichen Buchstaben innerhalb einer Zeile sind signifikant verschieden (p ≤ 0,05).
Differenzen zwischen Werten mit * und ° überschreiten die Signifikanzschwelle minimal (p ≤ 0,0874)
Zwischen den Geschlechtergruppen bestehen geringfügige Unterschiede in der
Körperzusammensetzung. Die intakten männlichen Tiere (n = 29) haben eine geringere
Fettauflage (16,58 ± 0,84 cm3) als die weiblichen Tiere (n = 63; 17,22 ± 0,54 cm3) bzw.
die männlichen Kastraten (n = 19; 18,05 ± 0,91 cm3). Im Muskelvolumen liegen
minimal die weiblichen Probanden mit 45,86 ± 1,27 cm3 vor den männlichen Tieren
(44,95 ± 1,97 cm3) bzw. den männlichen Kastraten (44,65 ± 2,16 cm3). Tendenziell
weisen jedoch die männlichen intakten Tiere ein geringeres Fett-zu-Muskel-Verhältnis
(0,345 ± 0,020) im Vergleich zu den weiblichen Schweinen (0,366 ± 0,014) und den
männlichen Kastraten (0,380 ± 0,023) auf.
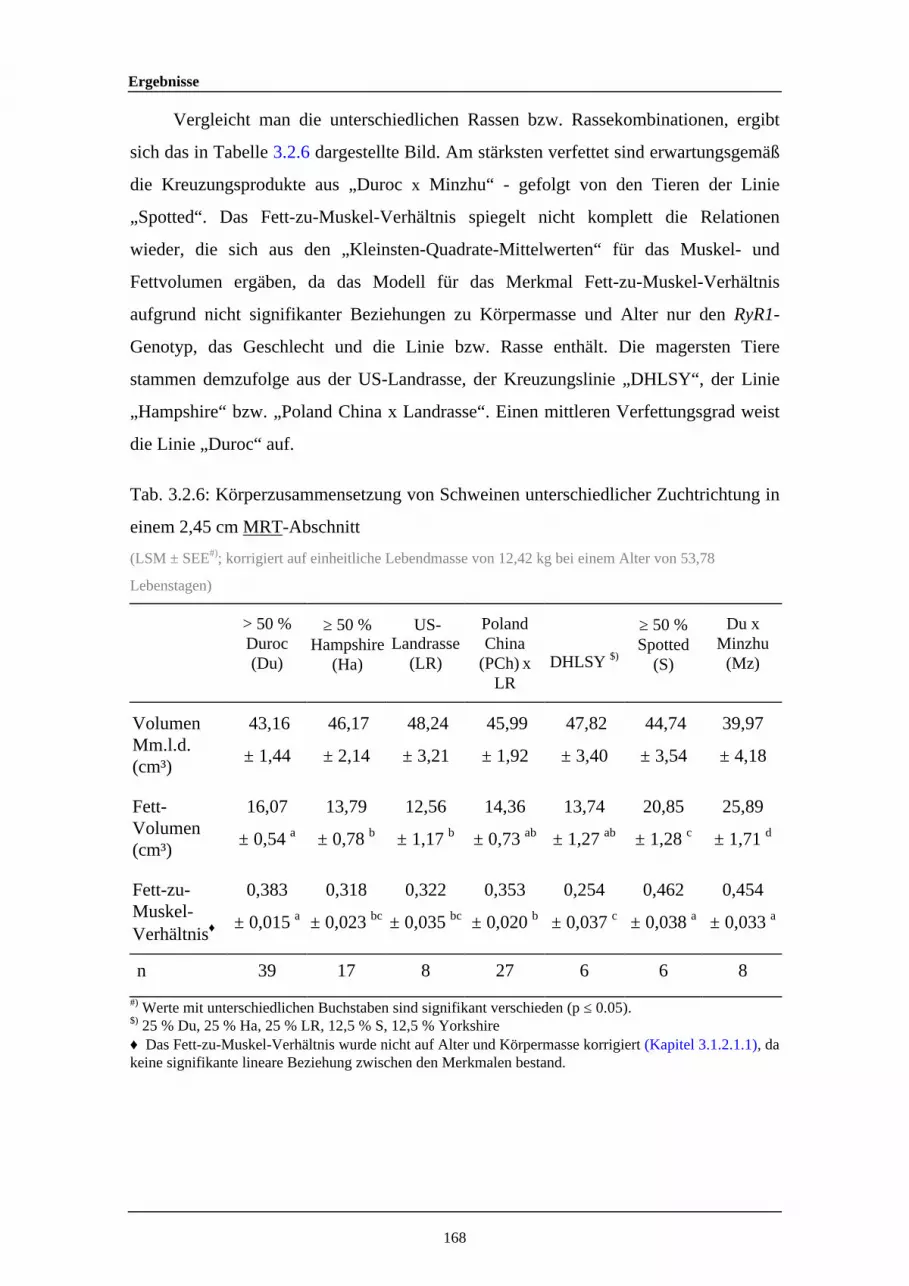
Ergebnisse
168
Vergleicht man die unterschiedlichen Rassen bzw. Rassekombinationen, ergibt
sich das in Tabelle 3.2.6 dargestellte Bild. Am stärksten verfettet sind erwartungsgemäß
die Kreuzungsprodukte aus „Duroc x Minzhu“ - gefolgt von den Tieren der Linie
„Spotted“. Das Fett-zu-Muskel-Verhältnis spiegelt nicht komplett die Relationen
wieder, die sich aus den „Kleinsten-Quadrate-Mittelwerten“ für das Muskel- und
Fettvolumen ergäben, da das Modell für das Merkmal Fett-zu-Muskel-Verhältnis
aufgrund nicht signifikanter Beziehungen zu Körpermasse und Alter nur den RyR1-
Genotyp, das Geschlecht und die Linie bzw. Rasse enthält. Die magersten Tiere
stammen demzufolge aus der US-Landrasse, der Kreuzungslinie „DHLSY“, der Linie
„Hampshire“ bzw. „Poland China x Landrasse“. Einen mittleren Verfettungsgrad weist
die Linie „Duroc“ auf.
Tab. 3.2.6: Körperzusammensetzung von Schweinen unterschiedlicher Zuchtrichtung in
einem 2,45 cm MRT-Abschnitt
(LSM ± SEE#); korrigiert auf einheitliche Lebendmasse von 12,42 kg bei einem Alter von 53,78
Lebenstagen)
> 50 %Duroc(Du)
≥ 50 % US-Hampshire
(Ha)Landrasse
(LR)
PolandChina
(PCh) xLR
DHLSY $)
≥ 50 %Spotted
(S)
Du xMinzhu
(Mz)
VolumenMm.l.d.(cm³)
43,16
± 1,44
46,17
± 2,14
48,24
± 3,21
45,99
± 1,92
47,82
± 3,40
44,74
± 3,54
39,97
± 4,18
Fett-Volumen(cm³)
16,07
± 0,54 a13,79
± 0,78 b12,56
± 1,17 b14,36
± 0,73 ab
13,74
± 1,27 ab
20,85
± 1,28 c25,89
± 1,71 d
Fett-zu-Muskel-Verhältnis♦
0,383
± 0,015 a0,318
± 0,023 bc
0,322
± 0,035 bc
0,353
± 0,020 b0,254
± 0,037 c0,462
± 0,038 a0,454
± 0,033 a
n 39 17 8 27 6 6 8
#) Werte mit unterschiedlichen Buchstaben sind signifikant verschieden (p ≤ 0.05).$) 25 % Du, 25 % Ha, 25 % LR, 12,5 % S, 12,5 % Yorkshire♦ Das Fett-zu-Muskel-Verhältnis wurde nicht auf Alter und Körpermasse korrigiert (Kapitel 3.1.2.1.1), dakeine signifikante lineare Beziehung zwischen den Merkmalen bestand.

Ergebnisse
169
3.2.2.2 MRT-Lebendmassegruppen 30 kg, 60 kg, 90 kg
Verschiedene Volumenmaße am Schinken und im Thoraxbereich des Schweins dienten dem
Vergleich der Bemuskelung bzw. des Verfettungsgrades der drei RyR1-Genotypen bei 30, 60
und 90 kg Lebendmasse. Die Volumenmaße von Schinkenmagergewebe (SMGV) und
korrespondierender Fettauflage (SFETTV) sowie das Gesamtvolumen (SGV) im Bereich der
Hintergliedmaßen/Becken resultieren aus der Aufnahme von 15 aufeinanderfolgenden
Schnittbildern mit einer Schnittdicke von 1 cm (ohne Zwischenraum) beginnend am äußeren
Schwanzansatz in kranialer Richtung. Die Messwerte für das Volumen der beiden Mm.
longissimi dorsi und des darüber liegenden Fettes ergeben sich aus der Aufnahme von 10
aufeinanderfolgenden Schnittbildern beginnend am 14. Brustwirbel in kranialer Richtung
(Abb. 3.1.12).
Erwartungsgemäß verfügen die homozygoten Defektallelträger für den RyR1-Genort
über ein größeres Muskelvolumen und eine reduzierte Fettauflage sowohl an der
Schinkenmessstelle als auch im Thoraxbereich (Mm.l.d.), woraus jeweils das niedrigste
(erwünschte) Fett-zu-Magergewebe-Verhältnis folgt. In allen Gewichtsgruppen - mit
Ausnahme der vergleichsweise niedrigsten Werte für das Fettauflagevolumen am Schinken
und das Schinkengesamtvolumen in der 90 kg Gruppe bzw. des vergleichsweise höchsten
Wertes für das Schinkengesamtvolumen in der 60 kg Gruppe - liegt der heterozygote Genotyp
zwischen den beiden homozygoten Genotypen.
Da die Tierzahl und die Zusammensetzung der Tiergruppen in jeder Gewichtsgruppe
aufgrund der Stufenschlachtung variiert, ergeben sich unterschiedlich hohe Differenzen
zwischen den Genotypen. Am aussagekräftigsten für den Vergleich der RyR1 -Genotypen sind
die Werte aus der 30 kg Gruppe bzw. der „Referenzgruppe“, die alle Tiere (n = 47) umfassen.
In der Referenzgruppe ist darauf hinzuweisen, dass ein männlicher Kastrat der Hampshire-
Linie (nn) während des Versuchszeitraumes (nach der DXA-Analyse bei 60 kg) verendete
und nicht mehr bei 60 kg (lebend) tomographiert werden konnte. Für dieses Tier liegen
folglich MRT-Ergebnisse nur in der 30 kg Gruppe vor.

Ergebnisse
170
30 kg Gruppe
Tab. 3.2.7: Volumen des Schinkenmagergewebes (einschließlich Knochen) (SMGV) und des
darüber liegenden Fettes (SFETTV) sowie des gesamten Schinkens (SGV) und das Fett-zu-
Muskel-Verhältnis (SFMV) in einer 15 cm MRT-Region für Schweine mit unterschiedlichen
RyR1-Genotypen bei ca. 30 kg Lebendmasse (LSM ± SEE #)
n SMGV (cm3) SFETTV (cm3) SFMV SGV (cm³)
NN 16 3560,82 ± 66,84 a* 801,52 ± 27,25 a 0,224 ± 0,008 a 4362,34 ± 67,36 a
Nn 22 3710,38 ± 47,15 ab° 772,52 ± 19,23 ab* 0,209 ± 0,006 a 4482,89 ± 47,52 ab
nn 9 3881,09 ± 80,97 b 704,32 ± 33,02 b° 0,184 ± 0,010 b 4585,40 ± 81,60 b
#) Werte mit unterschiedlichen Buchstaben sind signifikant verschieden (p ≤ 0,05). Differenzen zwischen den Werten mit * und °überschreiten das Signifikanzniveau nur minimal mit p ≤ 0,075.
Tab. 3.2.8: Volumen der Mm. longissimi dorsi (MLDV) und des darüber liegenden Fettes
(FETTV) sowie das Fett-zu-Muskel-Verhältnis (FMV) in einer 10 cm MRT-Region für
Schweine mit unterschiedlichen RyR1-Genotypen bei ca. 30 kg Lebendmasse (LSM ± SEE #)
n MLDV (cm3) FETTV (cm3) FMV
NN 16 285,12 ± 13,95 a 117,96 ± 6,79 a 0,44 ± 0,03 a
Nn 22 291,72 ± 9,84 a 95,36 ± 4,79 b 0,34 ± 0,02 b
nn 9 332,98 ± 16,90 b 91,21 ± 8,22 b 0,28 ± 0,04 b
#) Werte mit unterschiedlichen Buchstaben sind signifikant verschieden (p ≤ 0,05).

Ergebnisse
171
60 kg Gruppe
Tab. 3.2.9: Volumen des Schinkenmagergewebes (einschließlich Knochen) (SMGV) und des
darüber liegenden Fettes (SFETTV) sowie des gesamten Schinkens (SGV) und das Fett-zu-
Muskel-Verhältnis (SFMV) in einer 15 cm MRT-Region für Schweine mit unterschiedlichen
RyR1-Genotypen bei ca. 60 kg Lebendmasse (LSM ± SEE #)
n SMV (cm3) SFETTV (cm3) SFMV SGV (cm³)
NN 14 6668,91 ±137,79 * 1587,72 ± 58,78 a 0,239 ± 0,010 a 8256,63 ±141,09
Nn 11 6949,17 ±112,30 1519,33 ± 47,90 ab 0,222 ± 0,008 a 8468,50 ±114,98
nn 6 7029,44 ±192,42 ° 1372,27 ± 82,08 b 0,199 ± 0,014 b 8401,70 ±197,03
#) Werte mit unterschiedlichen Buchstaben sind signifikant verschieden (p ≤ 0,05). Differenzen zwischen den Werten mit * und ° überschreiten das Signifikanzniveau nur minimal mit p < 0,078.
Tab. 3.2.10: Volumen der Mm. longissimi dorsi (MLDV) und des darüber liegenden Fettes
(FETTV) sowie das Fett-zu-Muskel-Verhältnis (FMV) in einer 10 cm MRT-Region für
Schweine mit unterschiedlichen RyR1-Genotypen bei ca. 60 kg Lebendmasse (LSM ± SEE #)
n MLDV (cm3) FETTV (cm3) FMV
NN 14 549,68 ± 32,43 * 324,77 ± 21,49 a 0,62 ± 0,05 a
Nn 11 626,28 ± 26,43 ° 310,36 ± 17,51 a 0,53 ± 0,04 a
nn 6 636,90 ± 45,28 ° 220,10 ± 30,01 b 0,37 ± 0,06 b
#) Werte mit unterschiedlichen Buchstaben sind signifikant verschieden (p ≤ 0,05).Differenzen zwischen den Werten mit * und ° überschreiten das Signifikanzniveau nur minimal mit p ≤ 0,071.

Ergebnisse
172
90 kg Gruppe
Tab. 3.2.11: Volumen des Schinkenmagergewebes (einschließlich Knochen) (SMV) und des
darüber liegenden Fettes (SFETTV) sowie des gesamten Schinkens (SGV) und das Fett-zu-
Muskel-Verhältnis (SFMV) in einer 15 cm MRT-Region für Schweine mit unterschiedlichen
RyR1-Genotypen bei ca. 90 kg Lebendmasse (LSM ± SEE #)
n SMV (cm3) SFETTV (cm3) SFMV SGV (cm³)
NN 6 8479,59 ±147,55 a 1950,26 ±131,78 0,229 ± 0,015 10429,87 ±185,83 a
Nn 6 8529,31 ±148,38 a 1754,20 ±132.53 0,207 ± 0,015 10283,50 ±186,88 a
nn 5 9693,47 ±274,62 b 1829,41 ±245,28 0,188 ± 0,028 11522,88 ±345,87 b
#) Werte mit unterschiedlichen Buchstaben sind signifikant verschieden (p ≤ 0,05).
Tab. 3.2.12: Volumen der Mm. longissimi dorsi (MLDV) und des darüber liegenden Fettes
(FETTV) sowie das Fett-zu-Muskel-Verhältnis (FMV) in einer 10 cm MRT-Region für
Schweine mit unterschiedlichen RyR1-Genotypen bei ca. 90 kg Lebendmasse (LSM ± SEE #)
n MLDV (cm3) FETTV (cm3) FMV
NN 6 772,58 ± 26,77 a 416,43 ± 43,85 a 0,55 ± 0,06 a
Nn 6 775,38 ± 26,92 a 372,26 ± 44,10 ab 0,49 ± 0,06 a
nn 5 941,39 ± 49,83 b 209,61 ± 81,62 b 0,15 ± 0,11 b
#) Werte mit unterschiedlichen Buchstaben sind signifikant verschieden (p ≤ 0,05).

Ergebnisse
173
Referenztiere (26,4 kg bis 97,2 kg)
Analysiert man die Messwerte der 47* Tiere (30 kg: 2 x NN, 11 x Nn, 2 x nn; 60 kg: 8 x NN,
5 x Nn, 2 x nn*; 90 kg: 6 x NN, 6 x Nn und 5 x nn), die unmittelbar nach MR-Tomographie
und DXA-Untersuchung für die Referenzzerlegung und chemische Analyse verwendet
wurden, ergibt sich das gleiche Bild wie bei den Auswertungen getrennt nach
Lebendmassegruppen. Der heterozygote Genotyp liegt für alle Merkmale zwischen den
beiden homozygoten Genotypen. Bei einem durchschnittlichen Lebensalter von 122,06 Tagen
betrug die durchschnittliche Lebendmasse zum Zeitpunkt der MR-Tomographie 62,8 kg,
während das durchschnittliche Schlachtalter bei 125,23 Lebenstagen und 63,72 kg
Lebendmasse lag.
Tab. 3.2.13: Volumen des Schinkenmagergewebes (einschließlich Knochen) (SMV) und des
darüber liegenden Fettes (SFETTV) sowie des gesamten Schinkens (SGV) und das Fett-zu-
Muskel-Verhältnis (SFMV) in einer 15 cm MRT-Region für Schweine mit unterschiedlichen
RyR1-Genotypen zwischen 26,4 bis 97,2 kg Lebendmasse (LSM ± SEE #)
(korrigiert auf eine durchschnittliche Lebendmasse von 62,8 kg und 122,06 Lebenstage)
n SMV (cm3) SFETTV (cm3) SFMV SGV (cm3)
NN 16 6292,57 ±147,04 a 1490,79 ± 63,63 a 0,232 ± 0,010 a 7783,36 ±158,81
Nn 22 6498,39 ±103,92 ab 1384,81 ± 44,97 ab 0,218 ± 0,007 a 7883,19 ±112,23
nn 9 6796,14 ±175,28 b 1266,41 ± 75,86 b 0,190 ± 0,011 b 8062,55 ±189,30
#) Werte mit unterschiedlichen Buchstaben sind signifikant verschieden (p ≤ 0,05).
* 1 männlicher Kastrat der Hampshire-Linie (nn) verendete vor der MR-Tomographie bei 60 kg, wurde aber bei30 kg tomographiert und bei 60 kg mittels DXA untersucht und anschließend zerlegt sowie chemisch analysiert.

Ergebnisse
174
Tab. 3.2.14: Volumen der Mm. longissimi dorsi (MLDV) und des darüber liegenden Fettes
(FETTV) sowie das Fett-zu-Muskel-Verhältnis (FMV) in einer 10 cm MRT-Region für
Schweine mit unterschiedlichen RyR1-Genotypen zwischen 26,4 bis 97,2 kg Lebendmasse
(LSM ± SEE #)
(korrigiert auf eine durchschnittliche Lebendmasse von 62,8 kg und 122,06 Lebenstage)
n MLDV (cm3) FETTV (cm3) FMV
NN 16 524,08 ± 26,50 a 295,41 ± 21,43 a 0,548 ± 0,044 a*
Nn 22 591,98 ± 18,73 b 261,48 ± 15,15 a 0,446 ± 0,031 a*
nn 9 640,17 ± 31,59 b 198,21 ± 25,55 b 0,314 ± 0,053 b
#) Werte mit unterschiedlichen Buchstaben sind signifikant verschieden (p ≤ 0,05). Differenzen zwischen den Werten mit * und ° überschreiten das Signifikanzniveau nur minimal mit p ≤ 0,072.
Schlachtkörperanalyse - Chemische Körperzusammensetzung und Schlachtkörpermaße
(30 - 90 kg Gruppen)
Die Ergebnisse der chemischen Analyse bzw. der Schlachtkörperzerlegung decken sich mit
den Resultaten aus der MR-Tomographie. Der heterozygote Genotyp liegt sowohl für die
Parameter aus der chemischen Analyse (Tab. 3.2.15) als auch für die Parameter aus der
Schlachtkörperanalyse (Tab. 3.2.16 – 3.2.19) zwischen den beiden homozygoten Genotypen.
Tab. 3.2.15: Chemische Körperzusammensetzung für Schweine mit unterschiedlichen RyR1-
Genotypen zwischen 26,4 bis 97,2 kg Lebendmasse (LSM ± SEE #)
(korrigiert auf eine durchschnittliche Lebendmasse von 63,72 kg und 125,23 Lebenstage)
n Fett (%) Magergewebe (%)*) Protein (%)
NN 16 23,96 ± 0,88 a 72,51 ± 0,78 a 16,71 ± 0,44
Nn 22 22,18 ± 0,63 a 73,81 ± 0,55 a 17,35 ± 0,31
nn 9 19,60 ± 1,06 b 76,79 ± 0,93 b 17,72 ± 0,52
#) Kleinste Quadrate Mittelwerte mit unterschiedlichen Buchstaben sind signifikant verschieden (p ≤ 0,05).*) Magergewebe (%) = Wasser (%) + Protein (%)

Ergebnisse
175
Tab. 3.2.16: DXA-Körperzusammensetzung für Schweine mit unterschiedlichen RyR1-
Genotypen zwischen 26,4 bis 97,2 kg Lebendmasse, bei einer durchschnittlicher DXA-
Lebendmasse von 63,06 kg (124,85 Lebenstage) (LSM ± SEE #)
n DXA-Fett DXA-Fett DXA-Magergewebe DXA-Magergewebe (%) (kg) (%) (kg)
NN 16 16,54 ± 0,87 a 11,76 ± 0,72 a 81,93 ± 0,86 49,91 ± 0,71 a
Nn 22 14,98 ± 0,61 ab 10,27 ± 0,51 ab 82,54 ± 0,61 51,35 ± 0,50 ab
nn 9 13,42 ± 1,03 b 8,92 ± 0,85 b 83,98 ± 1,03 52,72 ± 0,85 b
#) Kleinste Quadrate Mittelwerte mit unterschiedlichen Buchstaben sind signifikant verschieden (p ≤ 0,05).
Tab. 3.2.17: Kotelettfläche, Rückenspeckfläche und Speck-zu-Muskelverhältnis (FMR) für
Schweine mit unterschiedlichen RyR1-Genotypen zwischen 26,4 bis 97,2 kg Lebendmasse
(LSM ± SEE #)
(korrigiert auf eine durchschnittliche Lebendmasse von 63,72 kg und 125,23 Lebenstage)
n Kotelett (cm2) Rückenspeck (cm2) FMR
NN 16 32,75 ± 1,42 a 19,90 ± 1,08 a 0,616 ± 0,040 a*
Nn 22 33,23 ± 1,01 a 16,59 ± 0,77 b 0,516 ± 0,028 ab*°
nn 9 37,08 ± 1,69 b 14,31 ± 1,29 b 0,418 ± 0,048 b°
#) Kleinste Quadrate Mittelwerte mit unterschiedlichen Buchstaben sind signifikant verschieden (p ≤ 0,05).
*Differenzen zwischen Werten mit * und ° überschreiten das Signifikanzniveau nur minimal mit p ≤ 0,065.
In den Teilstückgewichten liegt der heterozygote Genotyp zwar ebenfalls zwischen den
beiden homozygoten Genotypen, allerdings gibt es teilweise eine Verschiebung in der
Rangierung zwischen den homozygoten Genotypen. Während die homozygoten
Defektallelträger das höchste Schinken- und Schultergewicht aufweisen, verfügt der
homozygot normale Genotyp bedingt durch den höheren Fettanteil über die höchsten Blatt-,
Rücken- und Bauchgewichte (Tab. 3.2.18).

Ergebnisse
176
Tab. 3.2.18: Schinken-, Schulter-, Blatt- und Rückengewicht für Schweine mit
unterschiedlichen RyR1-Genotypen zwischen 26,4 bis 97,2 kg Lebendmasse (LSM ± SEE #)
(korrigiert auf eine durchschnittliche Lebendmasse von 63,72 kg und 125,23 Lebenstage)
n Schinken (kg) Schulter (kg) Blatt (kg) Rücken (kg) Bauch (kg)
NN 16 6,47 ± 0,11 a* 4,48 ± 0,11 a 3,46 ± 0,13 a 5,11 ± 0,11 * 4,15 ± 0,10 a
Nn 22 6,73 ± 0,08 a° 4,66 ± 0,07 ab* 3,36 ± 0,09 a 4,89 ± 0,08 3,89 ± 0,07 b
nn 9 7,25 ± 0,14 b 4,91 ± 0,13 b° 2,93 ± 0,15 b 4,79 ± 0,13 ° 3,77 ± 0,11 b
#) Werte mit unterschiedlichen Buchstaben sind signifikant verschieden (p ≤ 0,05). Differenzen zwischen Werten mit * und ° überschreiten das Signifikanzniveau nur minimal mit p ≤ 0,075.
Zusätzlich zu den Teilstückgewichten wurden die Gewichte einzelner Körperorgane erfasst
(Tab. 3.2.19). In allen Fällen liegen die Organgewichte des homozygoten Defektallel-
Genotyps unter den Gewichten der beiden anderen Genotypen. Das trifft auch für die nicht in
der Tabelle aufgeführten Gewichte des Gastrointestinaltraktes einschließlich Lunge mit
6,22 ± 0,30 kg für nn, 6,89 ± 0,18 kg für Nn und 7,32 ± 0,25 kg für NN zu.
Tab. 3.2.19: Leber-, Herz- und Nierengewichte für Schweine mit unterschiedlichen RyR1-
Genotypen zwischen 26,4 bis 97,2 kg Lebendmasse (LSM ± SEE #)
(korrigiert auf eine durchschnittliche Lebendmasse von 63,72 kg und 125,23 Lebenstage)
n Leber (kg) Herz (kg) Niere (kg)
links rechts
NN 16 1,77 ± 0,06 a 0,296 ± 0,012 0,172 ± 0,007 a 0,163 ± 0,007 a
Nn 22 1,78 ± 0,04 a 0,308 ± 0,009 0,165 ± 0,005 a 0,158 ± 0,005 a
nn 9 1,47 ± 0,07 b 0,287 ± 0,015 0,146 ± 0,008 b 0,136 ± 0,008 b
#) Werte mit unterschiedlichen Buchstaben sind signifikant verschieden (p ≤ 0,05).

Ergebnisse
177
3.2.2.3 Genauigkeit der MR-Tomographie
Zur Überprüfung der Genauigkeit der Ergebnisse aus beiden MRT-Versuchen (4,7 Tesla1H-„Oberflächen“-Tomographie der Thoraxregion bei ca. 10 kg Lebendmasse – Versuch
1 und 1,5 Tesla 1H-„Ganzkörper“-Tomographie bei 30 - 90 kg Lebendmasse – Versuch
2) dienten die korrespondierenden Referenzwerte aus der Stufenschlachtung.
Regressionskoeffizienten zwischen den Einzelmerkmalen und die entsprechenden
Standardschätzfehler beschreiben die gemeinsame Variation der beiden Messmethoden.
Zusätzlich zu den gemessenen MR-Volumina im Schinken- bzw. Thoraxbereich wurde
der Volumenanteil bezogen auf die aktuelle Lebendmasse in cm³/kg berechnet (Tab.
3.2.20). Im Versuch 2 stehen sowohl der Schlachtkörperfettanteil (%) als auch der
Schlachtkörpermagergewebeanteil (%) in engster Beziehung zum MR-Fettauflage-
volumen im Thoraxbereich, gefolgt vom Fettauflagevolumen im Schinkenbereich. Im
Versuch 1 korreliert hingegen das MR-Fett-zu-Muskelverhältnis am höchsten mit dem
Schlachtkörperfettanteil bzw. –magergewebeanteil (jeweils in %). Außerdem weist das
MR-Schinkenmagervolumen (cm³) gefolgt vom MR-Mm.l.d.-Volumen die engste
Beziehung zur Schlachtkörpermager- und Proteinmasse (g) auf, während die
entsprechenden Fettauflagevolumina am stärksten mit der Schlachtkörperfettmasse (g)
korrelieren. Kein MR-Volumenmesswert ist hoch mit dem Schlachtkörper-Proteinanteil
(%) korreliert. Allein das Fett-zu-Muskel-Verhältnis im Thoraxbereich weist eine etwas
höhere Beziehung als alle anderen Merkmale zum Proteinanteil (%) auf. Kombiniert
man beide Experimente, indem nur die gemeinsamen Merkmale aus der Thoraxregion
(Mm.l.d.) berücksichtigt werden, ergeben sich im Vergleich zu den Einzelexperimenten
leicht erhöhte Regressionskoeffizienten zwischen dem MR-Mm.l.d.-Anteil (cm³/kg) und
dem Schlachtkörperfett- bzw. –magergewebeanteil (jeweils in %). Die Ursache liegt in
der geänderten Variation der Merkmale über den gesamten Lebendmassebereich im
Vergleich zu den getrennten Lebendmasseabschnitten. Für die Kombination beider MR-
Experimente wurden die Messwerte für das Volumen der Mm. longissimi dorsi und des
Auflagefettes der 10 kg Gruppe nach der folgenden Formel an die Messwerte ab der 30
kg Gruppe angepasst:
Volumen (korrigiert) = gemessenes Volumen (Versuch 1) • 10/2,45.
Da die MR-Bilder im Versuch 1 (10 kg Gruppe) in einer auf 2,45 cm begrenzten Region
unter Einsatz einer Oberflächenspule entstanden, während im Versuch 2 eine auf 10 cm
ausgedehnte Region analysiert wurde, war diese Anpassung erforderlich. Es wurden

Ergebnisse
178
keine Korrekturfaktoren für individuelle Variationen im Muskel- und Fettvolumen
eingesetzt, da die relative schmale 2,45 cm MR-Region keine gerichtete Zunahme oder
Abnahme der Volumenmaße von Axialschnittebene 1 (14. Brustwirbel) zu
Axialschnittebene 5 in kranialer Richtung zeigte (Beispiel in Abb. 3.2.9). Die
anatomischen Gegebenheiten und das Design der Oberflächenspule verhinderten den
sinnvollen Einsatz über der Becken-/ Hintergliedmaßenregion.
y = -0,1441x + 6,0623
R2 = 0,2692
y = -0,0353x + 1,9902
R2 = 0,4818
0
1
2
3
4
5
6
7
1 2 3 4 5
Schnittebene
cm³
Mm.l.d.-Volumen Fett-Volumen
y = -0,0078x + 5,1501
R2 = 0,0634
y = 0,0035x + 2,0312
R2 = 0,0024
0
1
2
3
4
5
6
1 2 3 4 5
Schnittebene
cm³
Mm.l.d.-Volumen Fett-Volumen
Abb. 3.2.9: Beispiele für die Variation der MRT-Axialschnitt-Messwerte (1-5) innerhalbeines Tieres
Ergänzend wurden die einfachen Regressionskoeffizienten zwischen den MR-
Volumenmaßen am Rücken bzw. Schinken und den korrespondierenden
Teilstückgewichten aus der Schlachtkörperzerlegung sowie den korrespondierenden
Maßen für Kotelett- und Fettfläche am Anschnitt zwischen 13./14. Brustwirbel
berechnet. Erwartungsgemäß besteht zwischen Schinkengewicht und MR-
Schinkengesamtvolumen (n = 46, Versuch 2) mit R2 = 0,976 (√MSE = 0,424) ebenso
wie zwischen dem Rückengewicht und dem „MR-Mm.l.d.+Fettauflagevolumen“
(n = 61, Versuch „1+2“) mit R2 = 0,961 (√MSE = 0,500) eine sehr enge Beziehung. Wird
die Regressionskurve durch den Nullpunkt (x, y) geführt, ergeben sich für die
betrachteten Merkmale im Vergleich zur Regressionsanalyse ohne Restriktion bei
gleichzeitig erhöhten Standardschätzfehlern zwar leicht erhöhte Bestimmtheitsmaße,
systematische Abweichungen werden aber bei Betrachtung der Einzelmesswerte im
Vergleich zur Regressionsgeraden deutlicher. Da der definierte 15 cm „MR-
Schinkenabschnitt“ bei den 30 kg Tieren mehr Gewebe als das Schlachtkörperteilstück
„Schinken“ umfasst, liegen alle Messwerte dieser Gruppe unterhalb der

Ergebnisse
179
Regressionsgeraden, während bei den 90 kg Tieren die umgekehrte Situation eintritt
(Abb. 3.2.10). Für den „Rücken“ existiert eine ähnliche Situation (Abb. 3.2.11). Speziell
in der 10 kg Gruppe, die den Messwerten der Gruppen von 30, 60 und 90 kg angepasst
wurde, ist zu beobachten, dass die multiplikativ korrigierten Messwerte unterhalb der
Regressionsgeraden angeordnet sind. Offensichtlich führt die lineare Anpassung von
einem 2,45 cm auf einen 10 cm MR-Abschnitt zu einer Überschätzung des
Rückenanteils, während bei den 30 - 90 kg Tieren keine systematischen Abweichungen
auftreten. Die Kombination beider Versuche ist deshalb nur unter Berücksichtigung
dieser Beziehung zu interpretieren. Generell ist es schwierig, zwei voneinander
abweichende Messverfahren sinnvoll zu vereinheitlichen. Beide Verfahren beruhen
zwar auf der Magnetresonanz-Bildgebung, umfassen jedoch voneinander abweichende
Messbereiche. Außerdem diente in der 10 kg Gruppe ein Oberflächenspulensystem zur
Bilderzeugung, während für die Tiere ab 30 kg ein Ganzkörperspulensystem verwendet
wurde.
Abb. 3.2.10: Beziehung zwischen Schinkenmasse der linken Schlachthälfte (kg) unddem „Schinken“-Gesamtvolumen (cm³) aus der MR-Tomographie[R2 = 0,994, √MSE = 0,590, mit Restriktion: Interzept = 0, links (rechts ohne Restriktion:R2 = 0,976; √MSE = 0,424)]
Da mit zunehmender Körpermasse der Anteil des mittels MRT gemessenen
Rückenvolumens (Mm.l.d. + Auflagefett) am vorhandenen, gesamten Rückenvolumen
aufgrund der auf 10 cm begrenzten MR-Messregion abnimmt, resultiert die mit höheren
Teilstücksgewichten des Rückens steigende Variation zwangsläufig (Abb. 3.2.11). Für
eine exakte Bestimmung des Rückenvolumens oder anderer Teilstücke ist demnach die
MR-Messung der gesamten Teilstücke oder Organe nötig (MITCHELL et al., 1993,

Ergebnisse
180
MITCHELL et al., 2001b). Neue, auf schnelleren Computern basierende
Auswertungsverfahren werden diese sehr aufwendige Methode erleichtern.
Interzept=0 ohne Restriktion
Abb. 3.2.11: Beziehung zwischen Rückenmasse (kg) der linken Schlachthälfte und dem„Rücken“-Volumen (Mm.l.d + Auflagefett, cm³) aus der MR-Tomographie[R2 = 0.987, √MSE = 0.524; mit Restriktion: Interzept = 0, links (rechts ohneRestriktion: R2 = 0,961, √MSE = 0,500)]
Abb. 3.2.12: Beziehung zwischen Kotelettfläche (cm2) der linken Schlachthälfte (13./14.Brustwirbel) und dem „Kotelett“-Volumen (cm³) aus der MR-Tomographie (14.Brustwirbel + 10 cm in kraniale Richtung)[R2 = 0,982, √MSE = 4,149; mit Restriktion: Interzept = 0 (ohne Restriktion:R2 = 0,929, √MSE = 4,143)]

Ergebnisse
181
Analysiert man die Beziehungen zwischen MR-Kotelett- bzw. MR-Rückenspeck-
Volumen und Kotelett- bzw. Rückenspeckfläche am Schlachtkörper getrennt, ergeben
sich erwartungsgemäß ebenfalls hohe Regressionskoeffizienten für diese
Merkmalspaare (Abb. 3.2.12). Mit steigenden Körpermassen bzw. Volumen- und
Flächenmaßen nehmen die Abweichungen von der Regressionsgeraden jedoch zu, da
Kotelett- und Rückenspeckfläche am Schlachtkörperanschnitt zwischen 13./14.
Brustwirbel die zunehmende Variation der Muskel- und
Rückenspeckquerschnittsflächen in dem 10 cm MR-Volumenabschnitt nicht
wiedergeben können. (Abb. 3.2.12, 3.2.13).
Abb. 3.2.13: Beziehung zwischen Rückenspeckfläche (cm2) der linken Hälfte (13./14.Brustwirbel) und dem „Rückenspeck“-Volumen (cm³) aus der MR-Tomographie (14.Brustwirbel + 10 cm in kraniale Richtung)[R2 = 0,961; √MSE = 3,062; Restriktion: Interzept = 0; (ohne Restriktion: R2 = 0,889;√MSE = 2,704)]
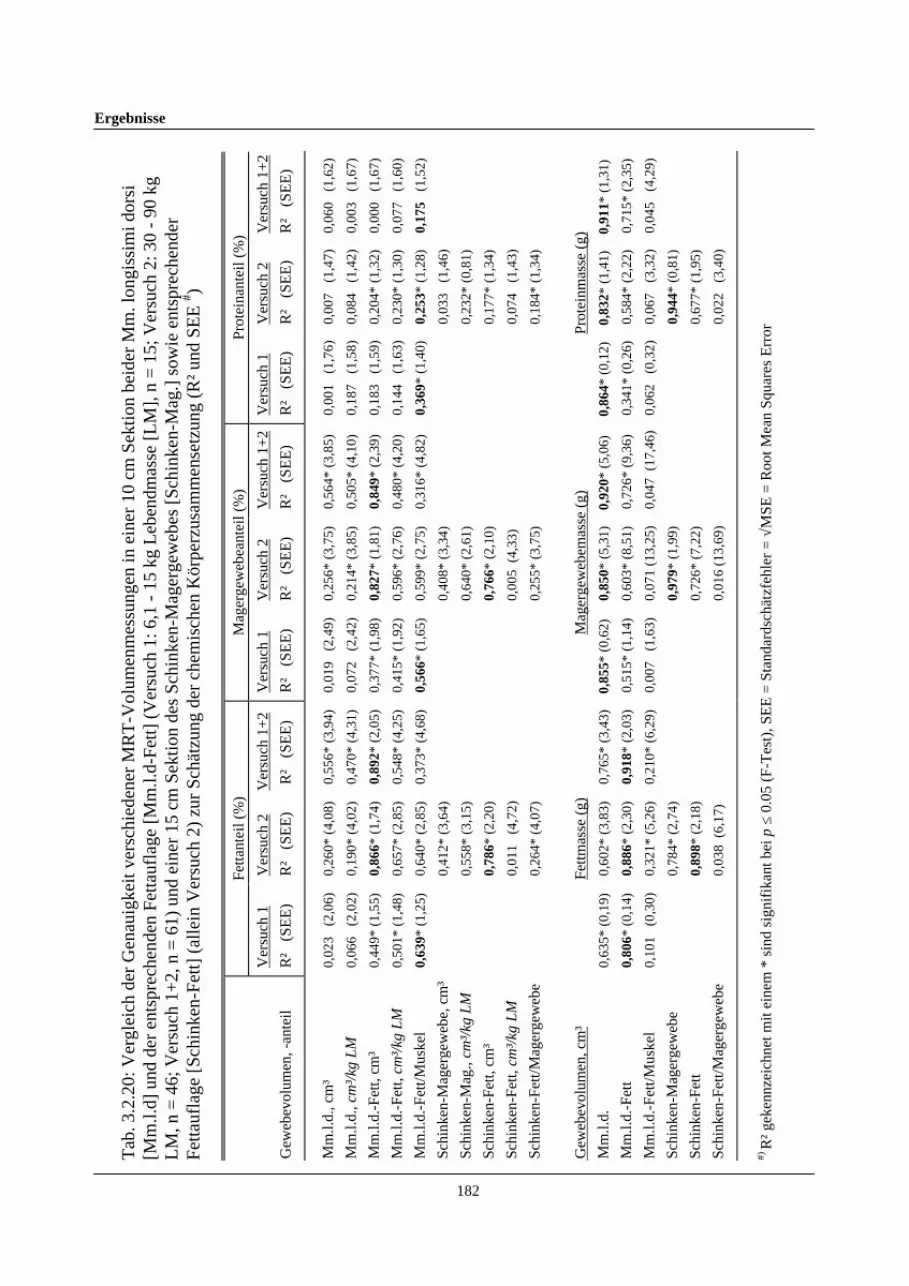
Tab
. 3.2
.20:
Ver
glei
ch d
er G
enau
igke
it ve
rsch
iede
ner
MR
T-V
olum
enm
essu
ngen
in e
iner
10
cm S
ektio
n be
ider
Mm
. lon
giss
imi d
orsi
[Mm
.l.d]
und
der
ent
spre
chen
den
Fetta
ufla
ge [
Mm
.l.d-
Fett]
(V
ersu
ch 1
: 6,1
- 1
5 kg
Leb
endm
asse
[L
M],
n =
15;
Ver
such
2: 3
0 -
90 k
gL
M, n
= 4
6; V
ersu
ch 1
+2,
n =
61)
und
ein
er 1
5 cm
Sek
tion
des
Schi
nken
-Mag
erge
web
es [
Schi
nken
-Mag
.] s
owie
ent
spre
chen
der
Fetta
ufla
ge [
Schi
nken
-Fet
t] (
alle
in V
ersu
ch 2
) zu
r Sc
hätz
ung
der
chem
isch
en K
örpe
rzus
amm
ense
tzun
g (R
² und
SE
E # )
Fetta
ntei
l (%
)M
ager
gew
ebea
ntei
l (%
)Pr
otei
nant
eil (
%)
Gew
ebev
olum
en, -
ante
il
Ver
such
1
R²
(S
EE
)
Ver
such
2
R²
(S
EE
)
Ver
such
1+
2
R²
(S
EE
)
Ver
such
1
R²
(S
EE
)
Ver
such
2
R²
(S
EE
)
Ver
such
1+
2
R²
(S
EE
)
Ver
such
1
R²
(S
EE
)
Ver
such
2
R²
(S
EE
)
Ver
such
1+
2
R²
(S
EE
)
Mm
.l.d.
, cm
³0,
023
(2,
06)
0,26
0* (
4,08
)0,
556*
(3,
94)
0,01
9 (
2,49
)0,
256*
(3,
75)
0,56
4* (
3,85
)0,
001
(1,
76)
0,00
7 (
1,47
)0,
060
(1,
62)
Mm
.l.d.
, cm
³/kg
LM
0,06
6 (
2,02
)0,
190*
(4,
02)
0,47
0* (
4,31
)0,
072
(2,
42)
0,21
4* (
3,85
)0,
505*
(4,
10)
0,18
7 (
1,58
)0,
084
(1,
42)
0,00
3 (
1,67
)
Mm
.l.d.
-Fet
t, cm
³0,
449*
(1,
55)
0,86
6* (
1,74
)0,
892*
(2,
05)
0,37
7* (
1,98
)0,
827*
(1,
81)
0,84
9* (
2,39
)0,
183
(1,
59)
0,20
4* (
1,32
)0,
000
(1,
67)
Mm
.l.d.
-Fet
t, cm
³/kg
LM
0,50
1* (
1,48
)0,
657*
(2,
85)
0,54
8* (
4,25
)0,
415*
(1,
92)
0,59
6* (
2,76
)0,
480*
(4,
20)
0,14
4 (
1,63
)0,
230*
(1,
30)
0,07
7 (
1,60
)
Mm
.l.d.
-Fet
t/Mus
kel
0,63
9* (
1,25
)0,
640*
(2,
85)
0,37
3* (
4,68
)0,
566*
(1,
65)
0,59
9* (
2,75
)0,
316*
(4,
82)
0,36
9* (
1,40
)0,
253*
(1,
28)
0,17
5 (
1,52
)
Sch
inke
n-M
ager
gew
ebe,
cm
³0,
412*
(3,
64)
0,40
8* (
3,34
)0,
033
(1,
46)
Sch
inke
n-M
ag.,
cm³/
kg L
M0,
558*
(3,
15)
0,64
0* (
2,61
)0,
232*
(0,
81)
Sch
inke
n-Fe
tt, c
m³
0,78
6* (
2,20
)0,
766*
(2,
10)
0,17
7* (
1,34
)
Sch
inke
n-Fe
tt, c
m³/
kg L
M0,
011
(4,
72)
0,00
5 (
4,33
)0,
074
(1,
43)
Sch
inke
n-Fe
tt/M
ager
gew
ebe
0,26
4* (
4,07
)0,
255*
(3,
75)
0,18
4* (
1,34
)
Gew
ebev
olum
en, c
m³
Fettm
asse
(g)
Mag
erge
web
emas
se (
g)Pr
otei
nmas
se (
g)
Mm
.l.d.
0,63
5* (
0,19
)0,
602*
(3,
83)
0,76
5* (
3,43
)0,
855*
(0,
62)
0,85
0* (
5,31
)0,
920*
(5,
06)
0,86
4* (
0,12
)0,
832*
(1,
41)
0,91
1* (
1,31
)
Mm
.l.d.
-Fet
t0,
806*
(0,
14)
0,88
6* (
2,30
)0,
918*
(2,
03)
0,51
5* (
1,14
)0,
603*
(8,
51)
0,72
6* (
9,36
)0,
341*
(0,
26)
0,58
4* (
2,22
)0,
715*
(2,
35)
Mm
.l.d.
-Fet
t/Mus
kel
0,10
1 (
0,30
)0,
321*
(5,
26)
0,21
0* (
6,29
)0,
007
(1,
63)
0,07
1 (1
3,25
)0,
047
(17
,46)
0,06
2 (
0,32
)0,
067
(3,
32)
0,04
5 (
4,29
)
Sch
inke
n-M
ager
gew
ebe
0,78
4* (
2,74
)0,
979*
(1,
99)
0,94
4* (
0,81
)
Sch
inke
n-Fe
tt0,
898*
(2,
18)
0,72
6* (
7,22
)0,
677*
(1,
95)
Sch
inke
n-Fe
tt/M
ager
gew
ebe
0,03
8 (
6,17
)0,
016
(13,
69)
0,02
2 (
3,40
)
#) R
² gek
ennz
eich
net m
it ei
nem
* s
ind
sign
ifik
ant b
ei p
≤ 0
.05
(F-T
est)
, SE
E =
Sta
ndar
dsch
ätzf
ehle
r =
√M
SE =
Roo
t Mea
n S
quar
es E
rror
182
Ergebnisse

Ergebnisse
183
Zusätzlich zur linearen Regressionsanalyse der Einzelmerkmale wurde eine Stepwise-
Regressionsanalyse (für mehrere Merkmale) durchgeführt, um das beste Modell für die
Vorhersage von Fett-, Protein- und Magergewebegehalt der rechten Schlachthälfte (jeweils in
%) auswählen zu können (Tab. 3.2.21). Sowohl für den Schlachtkörperfettanteil (%) als auch
für den Schlachtkörpermagergewebeanteil (%) ergab die Kombination des MR-Fettvolumens
(cm³) mit dem korrespondierenden MR-Mm.l.d.-Volumenanteil (cm³/kg) für die
zusammengefassten Versuche „1+2“ das beste Modell. Für den Versuch 2 resultierte die
Kombination des Fettvolumens über der Mm.l.d.-Region (cm³) und des Schinken-Fett-zu-
Muskel-Verhältnisses im besten Modell für die Vorhersage des Fettgewebeanteils (%),
während für den Magergewebeanteil (%) zusätzlich der Schinken-Magergewebe-
Volumenanteil (cm³/kg LM) in das Modell aufgenommen wurde. Für den Versuch 1 verbleibt
allein das Mm.l.d.-Fett-zu-Muskel-Verhältnis für die Vorhersage aller drei
Schlachtkörpermerkmale im Modell. Der Proteingehalt (%) wird in allen Fällen mit einer
geringen Genauigkeit geschätzt.
Tab. 3.2.21: Modelle aus der Stepwise-Regressionsanalyse für Merkmale aus der MR-
Tomographie, die am besten die chemische Schlachtkörperzusammensetzung vorhersagen
(p ≤ 0,05)
__________________________________________________________________________________
Variable R² SEEa DFb Parameter in der Gleichungc
Chemischer Fettgehalt im Schlachtkörper (%)Versuch 1 0,64 1,253 1 Mm.l.d.-Fett:Muskel-VerhältnisVersuch 2 0,90 1,500 2 Mm.l.d.-Fettvol., Schinken-Fett-zu-Muskel-VerhältnisVersuch 1+2 0,91 1,761 2 Mm.l.d.-Fettvol., Mm.l.d.-Volumenanteil (cm³/kg LM)
Chemischer Magergewebegehalt (Wasser + Protein) im Schlachtkörper (%)Versuch 1 0,57 1,654 1 Mm.l.d.-Fett:Muskel-VerhältnisVersuch 2 0,88 1,539 3 Mm.l.d.-Fettvol., Schinken-Fett-zu-Muskel-Verhältnis,
Schinken-Magergewebe-Volumenanteil (cm³/kg LM)Versuch 1+2 0,89 1,984 2 Mm.l.d.-Fettvol., Mm.l.d.-Volumenanteil (cm³/kg LM)
Chemischer Proteingehalt im Schlachtkörper (%)Versuch 1 0,37 1,396 1 Mm.l.d.-Fett:Muskel-VerhältnisVersuch 2 0,25 1,280 1 Mm.l.d.-Fett:Muskel-VerhältnisVersuch 1+2 0,27 1,442 2 Mm.l.d.-Fettvol., Mm.l.d.-Fett-zu-Muskel-Verhältnis
__________________________________________________________________________________________aStandardschätzfehler, b Freiheitsgrade,cAbkürzungen: Mm.l.d = Mm. longissimi dorsi (linke und rechte Seite, vol. = Volumen (cm³),
Vergleicht man die Schätzgenauigkeit unterschiedlicher Informationsquellen, dass heißt
unterschiedlicher Körperregionen, die mittels MRT gemessen wurden, wird deutlich, dass die

Ergebnisse
184
Kombination mehrerer Messpositionen (hier: Thorax + Schinken) die höchste Genauigkeit
ergibt. Die alleinige Verwendung der Ergebnisse aus der Thoraxregion bzw. der
Schinkenregion führt bereits zu relativ hohen Genauigkeiten (R2 > 0,82, √MSE < 2,01), liegt
jedoch sowohl für die Schätzung des Schlachtkörperfettgehaltes (%) als auch für die
Schätzung des Schlachtkörpermagergewebegehaltes (%) jeweils unter den kombinierten
Ergebnissen (R2 ≥ 0,88; √MSE < 1,54; Abb. 3.2.14).
0,7
0,74
0,78
0,82
0,86
0,9
R²
1
1,2
1,4
1,6
1,8
2
MSE
MLDMm.l.d..
DF=2DF = 2
HAM
DF=2DF = 2
MLD+HAMMm.l.d.+Schinken
DF=2DF = 2
Fett % (21,78 ± 4,58) Magergewebe % (74,31 ± 4,16)
√
Schlachtkörper-Schlachtkörper-
MLD Mm.l.d.
DF=2DF = 2
HAM
DF=2DF = 3
MLD+HAMMm.l.d.+Schinken
DF=2DF = 3
Schinken Schinken
Abb. 3.2.14: Bestimmtheitsmaße und Standardschätzfehler (√MSE) für die Schätzung deschemischen Fettgehalts im Schlachtkörper (%) und des chemischen Magergewebegehalts(Wasser + Protein) im Schlachtkörper (%) beim Vergleich verschiedener Informationsquellenaus der In-vivo-MR-Tomographie zwischen 30 und 90 kg Lebendmasse (p ≤ 0,05)
Tabelle 3.2.22 gibt einen Überblick zu den - dem Autoren bekannten - beim Schwein
durchgeführten In-vivo-MR-Tomographie-Experimenten zur Ermittlung der Körper-
zusammensetzung. In Abhängigkeit von Referenzmethodik und Bildauswertungsverfahren
werden die Schätzgenauigkeiten verglichen. Generell bestätigen die Ergebnisse aus der
Literatur die eigenen Resultate. Mit steigender Lebendmasse (Variation) und höherer Anzahl
von Messpositionen verbessert sich die Schätzgenauigkeit für die Körperzusammensetzung.
Der Fettanteil (oder Lipidanteil) lässt sich in der Regel besser mit der MR-Tomographie als
der Magerfleisch- bzw. Magergewebeanteil ermitteln. Die Clusteranalyse oder eine manuelle
bzw. semiautomatische Eingrenzung der „Regions of Interest“ zur Flächen- bzw.
Volumenbestimmung ist der Histogrammanalyse überlegen.

Tab
3.2
.22:
Gen
auig
keit
der
Schä
tzun
g de
r K
örpe
rzus
amm
ense
tzun
g be
im S
chw
ein
mitt
els
MR
T in
viv
o(m
odif
izie
rt n
ach
MIT
CH
EL
L u
nd S
CH
OL
Z, 2
001
und
SZ
AB
O e
t al.,
199
9)
Ras
se/
Popu
lati
onG
ewic
hts-
grup
pe(-
bere
ich)
nR
efer
enzm
erkm
alR
2
RSD
= √
MSE
SD
Anz
ahl v
onSc
anpo
sitio
nen
(Sch
eibe
n)
Met
hodi
k de
r B
ildau
swer
tung
(nac
h je
wei
liger
Fes
tlegu
ng d
erR
egio
ns o
f In
tere
st -
RO
I)
Ref
eren
z
Deu
tsch
eL
andr
asse
90 k
g75
Schl
acht
körp
erfe
tt %
Schl
acht
körp
erm
uske
lfle
isch
% (Zer
legu
ng)
0,66
0,64
0,60
0,63
2 (5
)2
(7)
(Sch
ulte
r un
dSc
hink
en)
His
togr
amm
anal
yse
Hen
ning
et a
l.(1
991)
Deu
tsch
eL
andr
asse
90 k
g75
Schl
acht
körp
erfe
tt %
Schl
acht
körp
erm
uske
lfle
isch
% (Zer
legu
ng)
0,74
0,76
0,52
0,50
1 (3
)1
(3)
(13.
/14.
Bru
stw
irbe
l)
Clu
ster
anal
yse
Scho
lz u
.a.
(199
3)
Kre
uzun
gen
mit
75 %
Lar
geW
hite
und
25
%B
ritis
che
Lan
dras
se
76-7
9 kg
(mag
erun
d fe
tt)
12Sc
hlac
htkö
rper
lipid
%(c
hem
isch
)Fe
ttgew
ebe
%(Z
erle
gung
)
0,96
0,96
0,24
0,22
13 13 (Sch
ulte
r bi
sSc
hink
en)
Sem
iaut
omat
isch
eSc
hwel
lwer
tset
zung
(Reg
ione
nwac
hstu
m)
kom
bini
ert
mit
man
uelle
r K
orre
ktur
Fow
ler
et a
l.(1
992)
Deu
tsch
eL
andr
asse
20 k
g
50 k
g
90 k
g
43 40 60
Fettg
eweb
e %
Mag
erfl
eisc
h %
Fettg
eweb
e %
Mag
erfl
eisc
h %
Fettg
eweb
e %
Mag
erfl
eisc
h %
(alle
s au
s Z
erle
gung
)
0,68
0,55
0,80
0,83
0,89
0,87
0,59
0,73
0,46
0,46
0,36
0,38
≤ 5
≤ 5
≤ 5
≤ 5
≤ 5
≤ 5
(Sch
ulte
r bi
sSc
hink
en)
Clu
ster
anal
yse
Bau
lain
u.a
.(1
996)
185

Ergebnisse
186
3.2.2.4 DXA-Lebendmassegruppen 10 kg, 30kg, 60 kg und 90 kg
Parallel zu den MR-Studien erfolgte eine Analyse der Gesamtkörperzusammensetzung
(Fettmasse, Magergewebemasse und Knochenmineralmasse) mittels Dualenergie-
Röntgenabsorptiometrie unter Verwendung eines LUNAR DPX-L© Scanners innerhalb
eines Abstands von ±3 Tagen zur MR-Studie. Neben dem Vergleich mit der
Magnetresonanz-Tomographie diente die DXA-Studie zur Ermittlung des
Magergewebewachstums und der Magergewebefutterverwertung. Dazu wurde zwischen 30
und 90 kg Lebendmasse der individuelle Futterverzehr der Tiere täglich erfasst. Aufgrund
der durchgeführten Stufenschlachtung absolvierten nur 17 Tiere (6 NN - 6 Nn - 5 nn) den
gesamten Versuch bis 90 kg Lebendmasse. Bis 30 kg Lebendmasse wurden die Probanden
in Gruppen zu 6 - 8 Tieren gehalten. Eine Erfassung des individuellen Futterverzehrs war
bis 30 kg Lebendmasse nicht möglich. Einen Anhaltspunkt über die DXA-
Körperzusammensetzung der RyR1-Genotypen zu Beginn des Versuchs bei ca. 10 kg
Lebendmasse lieferte eine stichprobenartige Analyse an 45 Tieren (13 NN - 14 Nn - 18 nn).
Die relative niedrige Zahl von Probanden in der 10 kg Gruppe (im Vergleich zur MR-
Tomographie) liegt an der erst später erfolgten Inbetriebnahme des LUNAR Scanners am
Growth Biology Laboratory. In der 10 kg Gruppe wurden mit dem DXA-Scanner im
Gegensatz zur MR-Tomographie signifikante Unterschiede zwischen den heterozygoten
und homozygoten Defektallelträgern ermittelt. Die homozygoten Defektallelträger nn
wiesen aber überraschend – bei sehr geringfügigen Unterschieden - die niedrigste
Magergewebemasse (MGM) bei gleichzeitig höchster Fettgewebemasse (FGM) auf,
während die heterozygoten Tiere am magersten waren (Tab. 3.3.23).

Ergebnisse
187
Tab. 3.2.23: Magergewebemasse (MGM), Fettgewebemasse (FGM) und
Knochenmineralmasse (KMM) aus der DXA-Analyse in Schweinen mit unterschiedlichen
RyR1-Genotypen bei durchschnittlich 12,55 kg Lebendmasse und 53,29 Lebenstagen (LSM
± SEE#)
n MGM (kg) FGM (kg) KMM (kg)
NN 13 11,431 ± 0,059 ab 0,774 ± 0,056ab 0,348 ± 0,010
Nn 14 11,517 ± 0,067 a 0,695 ± 0,063a 0,340 ± 0,011
nn 18 11,346 ± 0,089 b 0,857 ± 0,084b 0,350 ± 0,015
#) Werte mit unterschiedlichen Buchstaben sind signifikant verschieden (p ≤ 0,05).
Ab der 30 kg Gruppe decken sich die DXA-Ergebnisse wieder mit denen aus der MR-
Tomographie (Tab. 3.2.24 – Tab. 3.2.26). Die homozygot normalen Tiere verfügen bei
niedrigster Magergewebemasse über die höchste Fettgewebe- und Knochenmineralmasse,
während der heterozygote Genotyp in allen Messwerten zwischen den beiden homozygoten
Genotypen liegt.
Tab. 3.2.24: Magergewebemasse (MGM), Fettgewebemasse (FGM) und
Knochenmineralmasse (KMM) aus der DXA-Analyse in Schweinen mit unterschiedlichen
RyR1-Genotypen bei durchschnittlich 32,31 kg Lebendmasse und 84,11 Lebenstagen (LSM
± SEE)
n MGM (kg) FGM (kg) KMM (kg)
NN 16 28,365 ± 0,172 3,156 ± 0,172 0,783 ± 0,015
Nn 22 28,489 ± 0,122 3,055 ± 0,122 0,761 ± 0,011
nn 9 28,791 ± 0,209 2,751 ± 0,209 0,763 ± 0,019
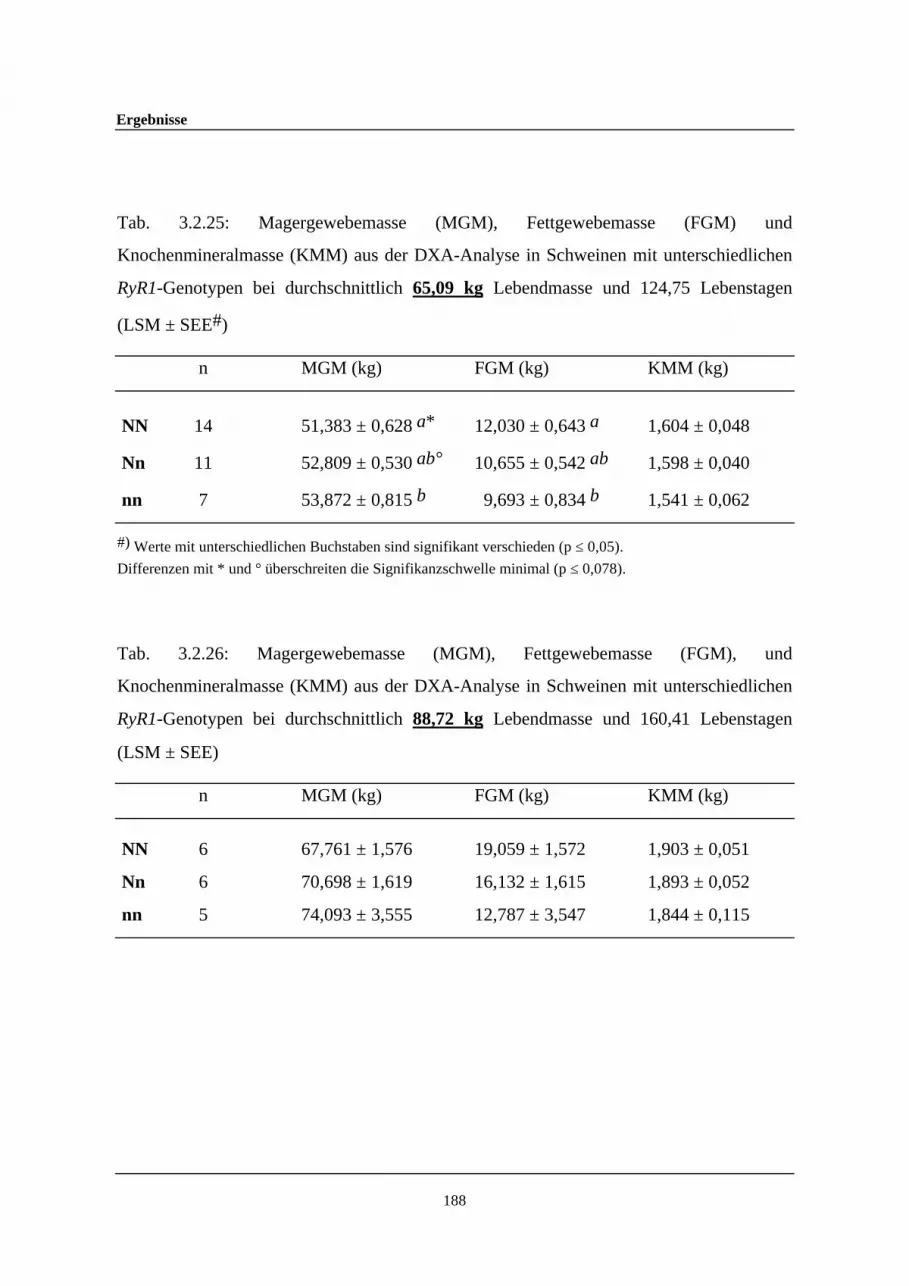
Ergebnisse
188
Tab. 3.2.25: Magergewebemasse (MGM), Fettgewebemasse (FGM) und
Knochenmineralmasse (KMM) aus der DXA-Analyse in Schweinen mit unterschiedlichen
RyR1-Genotypen bei durchschnittlich 65,09 kg Lebendmasse und 124,75 Lebenstagen
(LSM ± SEE#)
n MGM (kg) FGM (kg) KMM (kg)
NN 14 51,383 ± 0,628 a* 12,030 ± 0,643 a 1,604 ± 0,048
Nn 11 52,809 ± 0,530 ab° 10,655 ± 0,542 ab 1,598 ± 0,040
nn 7 53,872 ± 0,815 b 9,693 ± 0,834 b 1,541 ± 0,062
#) Werte mit unterschiedlichen Buchstaben sind signifikant verschieden (p ≤ 0,05).
Differenzen mit * und ° überschreiten die Signifikanzschwelle minimal (p ≤ 0,078).
Tab. 3.2.26: Magergewebemasse (MGM), Fettgewebemasse (FGM), und
Knochenmineralmasse (KMM) aus der DXA-Analyse in Schweinen mit unterschiedlichen
RyR1-Genotypen bei durchschnittlich 88,72 kg Lebendmasse und 160,41 Lebenstagen
(LSM ± SEE)
n MGM (kg) FGM (kg) KMM (kg)
NN 6 67,761 ± 1,576 19,059 ± 1,572 1,903 ± 0,051
Nn 6 70,698 ± 1,619 16,132 ± 1,615 1,893 ± 0,052
nn 5 74,093 ± 3,555 12,787 ± 3,547 1,844 ± 0,115

Ergebnisse
189
3.2.2.5 Genauigkeit der Dualenergie-Röntgenabsorptiometrie
10 kg Gruppe(Durchschnittliche DXA-Masse: 9,91 kg, durchschnittliches Schlachtalter/DXA-Alter:46,87 Tage, n = 15)
In der 10 kg Gruppe wurde ein Großteil der Tiere komplett zermahlen und
homogenisiert (Schlachthälfte + Innereien + Kopf). Im Gegensatz zu den Tieren ab 30
kg werden deshalb die Gehaltswerte für Fett, Magergewebe und Knochenmineral auf
den Gesamtkörper und nicht auf die rechte Schlachthälfte bezogen. Aufgrund der
geringen Tierzahl und der insgesamt geringen Variation innerhalb der untersuchten
Merkmale sind die Differenzen zwischen den RyR1-Genotypen in der 10 kg Gruppe
nicht signifikant. Mit Ausnahme des Knochenmineralgehaltes deckt sich jedoch die
Rangierung der chemischen Gehaltswerte mit denen der DXA-Gehaltswerte für Fett (%)
und Magergewebe (%) innerhalb der RyR1-Genotypen. Gleichzeitig wird deutlich, dass
die Original-DXA-Werte im Vergleich zur chemischen Analyse den Fett- und den
Knochenmineralgehalt systematisch unter- sowie den Magergewebegehalt
überschätzen. Nach Verwendung der von MITCHELL und CONWAY (1994)
vorgeschlagenen Regressionsgleichung zur Berechnung des DXA-Fettgehaltes aus dem
Röntgenschwächungskoeffizienten für Weichgewebe
[DXA-Fett (%) korrigiert = 493,4 – (348,8 • R-Wert)]
ist keine systematische Abweichung mehr zwischen den beiden Fettgehaltsmaßen
nachweisbar (Tab. 3.2.27). Der korrigierte Magergewebegehalt aus der DXA-Messung
liegt aufgrund des fixen – niedrigeren - DXA-Knochenmineralgehalts mit Ausnahme
des nn-Genotyps weiterhin leicht über dem chemisch ermitteltem Magergewebegehalt
(Tab. 3.2.28). Der Knochenmineralgehalt aus der DXA-Messung lässt sich nicht über
den R-Wert ableiten oder korrigieren, da LUNAR-DXA-Scanner allein den
Weichgewebe-R-Wert angeben. Ab einem bestimmten Schwellenwert für den
Röntgenschwächungskoeffizienten (R-Wert) wird davon ausgegangen, dass in der
durchstrahlten Pixel-Ebene Knochenmineral überwiegt. Diese Bildpunkte (Pixel)
werden generell dem Knochenmineral zugeordnet (Tab. 3.2.29).

Ergebnisse
190
Tab. 3.2.27: Vergleich von chemischem und DXA-Fettgehalt (%) in unterschiedlichen
RyR1-Genotypen innerhalb der 10 kg Gruppe (LSM ± SEE)
RyR1-Genotyp
n Chemischer Fettgehalt(%) [gesamter
Schlachtkörper]
DXA-Gesamt-fettgehalt (%)
DXA-Gesamt-fettgehalt (%)
(berechnet aus R-Wert$)
NN 4 10,560 ± 1,146 4,715 ± 1,124 9,124 ± 1,038
Nn 4 8,836 ± 1,340 4,000 ± 1,274 8,179 ± 1,176
nn 7 11,750 ± 1,830 7,523 ± 1,797 13,213 ± 1,659
$) Gleichung nach MITCHELL und CONWAY, 1994: DXA-Fett (%) korrigiert = 493,4 – [348,8 • R-Wert]
Tab. 3.2.28: Vergleich von chemischem und DXA-Magergewebegehalt (%) in
unterschiedlichen RyR1-Genotypen innerhalb der 10 kg Gruppe (LSM ± SEE#)
RyR1-Genotyp
n Magergewebe (%)(chem. Wasser +Protein) [rechteSchlachthälfte]
DXA-Gesamt-magergewebe (%)
DXA-Gesamt-magergewebe (%)(berechnet aus R-
Wert$)
NN 4 85,962 ± 0,381 a 92,761 ± 0,944 88,353 ± 0,867
Nn 4 87,711 ± 0,446 a 93,594 ± 1,071 89,419 ± 0,982
nn 7 84,702 ± 0,608 b 89,770 ± 1,510 84,079 ± 1,385
#Werte mit unterschiedlichen Buchstaben sind signifikant verschieden (p ≤ 0,05).$) DXA-Magergewebe % korrigiert = 100 - DXA-Fett % korrigiert - DXA-Knochenmineralgehalt %
Tab. 3.2.29: Vergleich von chemischem Rohaschegehalt (%) und DXA-
Knochenmineralgehalt (%) in unterschiedlichen RyR1-Genotypen innerhalb der 10 kg
Gruppe (LSM ± SEE)
RyR1-Genotyp
n Chem. Aschegehalt (%)[Gesamtschlachtkörper]
DXA-Gesamt-knochenmineralgehalt (%)
NN 4 3,396 ± 0,435 2,523 ± 0,184
Nn 4 3,512 ± 0,508 2,403 ± 0,208
nn 7 4,739 ± 0,694 2,707 ± 0,294

Ergebnisse
191
30 - 90 kgdurchschnittliche DXA-Masse: 63,06 kg (124,85 Lebenstage); durchschnittlicheLebendmasse zur Schlachtung: 63,72 kg (125,23 Lebenstage); n = 47
Für die Tiere ab 30 kg Lebendmasse, werden den DXA-Messwerten die chemischen
Analysewerte der rechten Schlachthälfte (ohne Kopf) gegenübergestellt (Tab. 3.2.30 –
Tab. 3.2.32). Die Ergebnisse decken sich mit denen aus der MR-Tomographie. Sowohl
nach den Ergebnissen aus der DXA-Analyse, als auch aus der chemischen Analyse sind
die homozygoten Defektallelträger am magersten, gefolgt von den heterozygoten
Defektallelträgern. Chemische und DXA-Werte stimmen - trotz hoher Beziehungen für
den Fett- und Magergewebegehalt (%) - in der absoluten Größenordnung nicht überein,
da die DXA-Werte den gesamten Körper (lebend) umfassen, während die chemischen
Analysewerte lediglich die rechte Schlachthälfte betreffen (Tab. 3.2.20, Abb. 3.2.15).
Der DXA-Fettgehalt basiert auf den DXA-Weichgewebe-R-Werte. Mit steigendem R-
Wert nimmt der Fettgehalt ab und der Magergewebegehalt zu (Tab. 3.2.30, 3.2.31).
Tab. 3.2.30: Vergleich von chemischem und DXA-Fettgehalt (%) in unterschiedlichen
RyR1-Genotypen im Lebendmassebereich von 30 - 90 kg (LSM ± SEE#)
RyR1-Genotyp
n Chem. Fettgehalt (%)[rechte Schlachthälfte]
DXA-Gesamt-fettgehalt (%)
DXA-R-Wert
NN 16 23,967 ± 0,886 a* 16,544 ± 0,867 a 1,355 ± 0,002 a*
Nn 22 22,178 ± 0,627 a° 14,984 ± 0,614 ab 1,359 ± 0,001 ab°
nn 9 19,604 ± 1,057 b 13,420 ± 1,033 b 1,362 ± 0,002 b
#)Werte mit unterschiedlichen Buchstaben sind signifikant verschieden (p ≤ 0,05).Differenzen zwischen Werten mit * und ° überschreiten das Signifikanzniveau minimal (p ≤ 0,089).
Tab. 3.2.31: Vergleich von chemischem und DXA-Magergewebegehalt (%) in
unterschiedlichen RyR1-Genotypen im Lebendmassebereich von 30 - 90 kg (LSM ±SEE#)
RyR1-Genotyp
n Magergewebe (%)(chem. Wasser + Protein)[rechte Schlachthälfte]
DXA-Gesamt-magergewebegehalt (%)
NN 16 72,513 ± 0,776 a 81,932 ± 0,864
Nn 22 73,815 ± 0,549 a 82,542 ± 0,612
nn 9 76,790 ± 0,925 b 83,975 ± 1,030
#) Werte mit unterschiedlichen Buchstaben sind signifikant verschieden (p ≤ 0,05).

Ergebnisse
192
Tab. 3.2.32: Vergleich von chemischem Rohaschegehalt und DXA-
Knochenmineralgehalt in unterschiedlichen RyR1-Genotypen im Lebendmassebereich
von 30 – 90 kg (LSM ± SEE#)
RyR1-Genotyp
n Chem. Aschegehalt (%)[rechte Schlachthälfte]
DXA-Gesamt-knochenmineralgehalt (%)
NN 16 3,742 ± 0,249 2,327 ± 0,067
Nn 22 3,367 ± 0,176 2,277 ± 0,048
nn 9 3,369 ± 0,296 2,324 ± 0,080
In Tabelle 3.2.33 sind die Schätzgenauigkeiten der In-vivo-DXA-Messwerte (R2, SEE)
für die chemische Zusammensetzung der rechten Schlachthälfte aufgeführt. Versuch 1
umfasst allein die Tiere der 10 kg Gruppe, während Versuch 2 die Tiere im
Lebendmassebereich von 30 - 90 kg einschließt. Insgesamt bestehen für den Fettgehalt
(%) und den Magergewebegehalt (%) hohe Beziehungen zwischen den
korrespondierenden DXA-Messwerten und den Werten aus der chemischen Analyse,
wobei der Fettgehalt noch genauer geschätzt wird als der Magergewebegehalt. Der
DXA-Fettgehalt (%) und das DXA-Fett-zu-Magergewebeverhältnis stehen in engerer
Beziehung zu den chemischen Analysewerten - einschließlich des chemischen
Magergewebegehalts - als der DXA-Magergewebegehalt (%). Im Gegensatz zum Fett-
und Magergewebegehalt (%) sind die Beziehungen zwischen DXA-
Knochenmineralgehalt (%) und chemischem Rohaschegehalt (%) nicht signifikant von
Null verschieden.
Erwartungsgemäß erklären die absoluten Gewebemassen (kg) aus der DXA-
Analyse einen sehr hohen Anteil der chemisch ermittelten Gewebemassen (R2 ≥ 0,89 für
die korrespondierenden Merkmale). Allein die Beziehung zwischen DXA-
Knochenmineralmasse (kg) und Rohasche (kg) fällt bei getrennter Betrachtung von
Versuch 1 und 2 mit einem Bestimmtheitsmaß von R2 = 0,61 geringer aus.
Fasst man beide Versuche zusammen, erhöhen sich generell alle
Bestimmtheitsmaße, da die messbare Variation zunimmt. Eine Ausnahme bildet der
Rohascheanteil (%), für den die Bestimmtheitsmaße auch nach Zusammenfassung von
Versuch 1 und 2 nicht signifikant verschieden von Null sind.
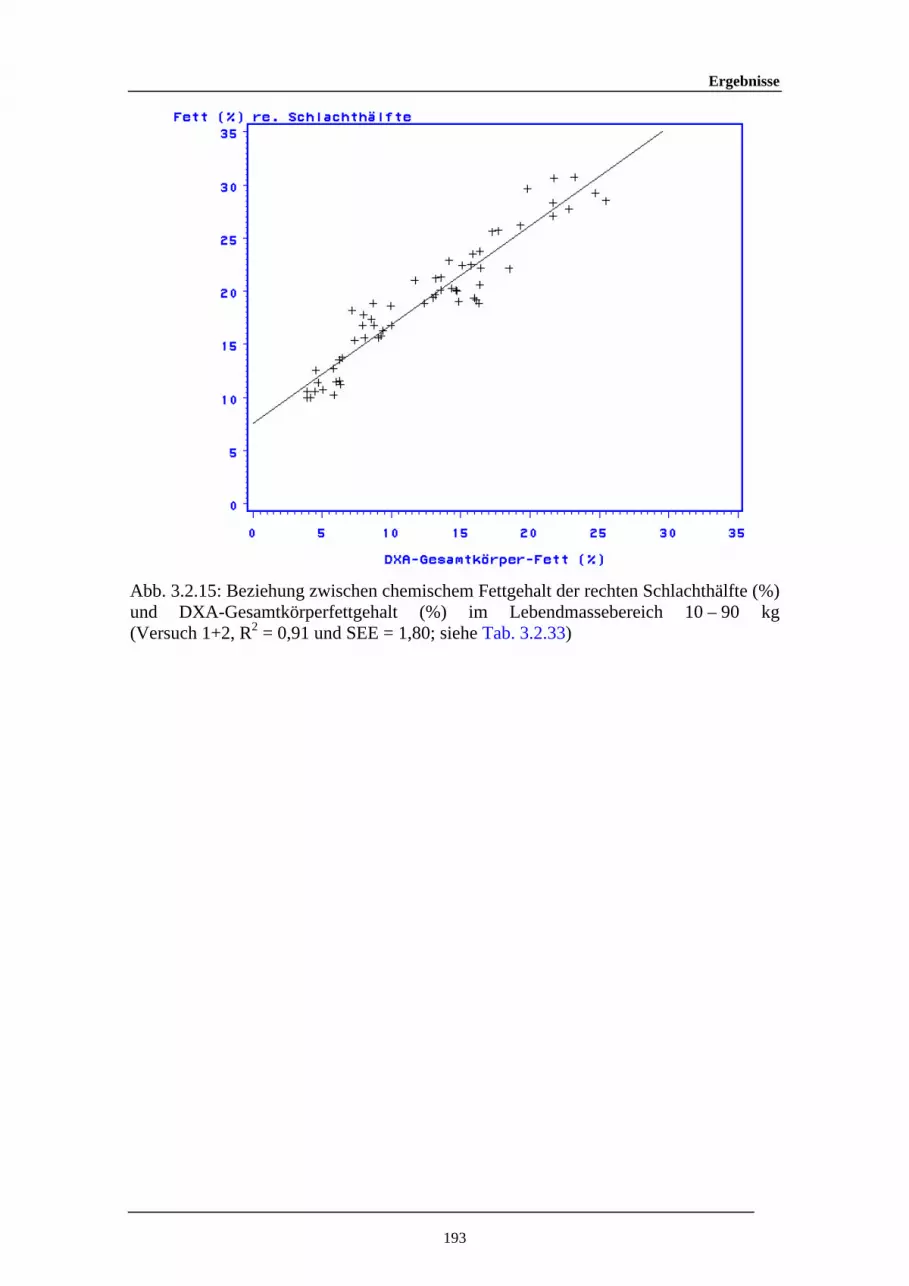
Ergebnisse
193
Abb. 3.2.15: Beziehung zwischen chemischem Fettgehalt der rechten Schlachthälfte (%)und DXA-Gesamtkörperfettgehalt (%) im Lebendmassebereich 10 – 90 kg(Versuch 1+2, R2 = 0,91 und SEE = 1,80; siehe Tab. 3.2.33)

Tab
. 3.2
.33:
Gen
auig
keit
der
DX
A-M
essu
ng (
in v
ivo)
zur
Sch
ätzu
ng d
er c
hem
isch
en Z
usam
men
setz
ung
der
rech
ten
Schl
acht
hälf
te, n
= 4
7 (R
² und
SEE
# )
Fetta
ntei
l (%
)M
ager
gew
ebea
ntei
l (%
)A
sche
ante
il (%
)
Gew
ebea
ntei
l, %
Ver
such
1
R²
(SE
E)
Ver
such
2
R²
(SE
E)
Ver
such
1+
2
R²
(SE
E)
Ver
such
1
R²
(SE
E)
Ver
such
2
R²
(SE
E)
Ver
such
1+
2
R²
(SE
E)
Ver
such
1
R²
(SE
E)
Ver
such
2
R²
(SE
E)
Ver
such
1+
2
R²
(SE
E)
DX
A-F
ett
0,70
8* (
1,03
)0,
846*
(1,
84)
0,91
0* (
1,80
)0,
630*
(1,
13)
0,81
5* (
1,86
)0,
880*
(2,
08)
0,13
3 (
0,92
)0,
032
(0,
74)
0,02
7 (
0,80
)
DX
A-M
ager
gew
ebe
0,68
5* (
1,07
)0,
765*
(2,
28)
0,86
4* (
2,21
)0,
579*
(1,
20)
0,74
6* (
2,17
)0,
841*
(2,
40)
0,14
0 (
0,92
)0,
024
(0,
74)
0,02
1 (
0,80
)
DX
A-K
noch
enm
iner
al0,
081
(1,
83)
0,12
0 (
4,40
)0,
223
(5,
29)
0,01
7 (
1,84
)0,
126
(4,
03)
0,24
6 (
5,22
)0,
051
(0,
96)
0,01
4 (
0,74
)0,
032
(0,
80)
DX
A-F
ett-
zu-M
ager
gew
ebe
0,71
0* (
1,03
)0,
851*
(1,
81)
0,90
0* (
1,90
)0,
627*
(1,
13)
0,81
2* (
1,87
)0,
859*
(2,
26)
0,12
8 (
0,92
)0,
028
(0,
74)
0,02
7 (
0,80
)
Gew
ebem
asse
, kg
Fett
(kg)
Mag
erge
web
e (k
g)A
sche
(kg
)
DX
A-F
ett
0,89
0* (
0,08
)0,
966*
(1,
14)
0,97
7* (
1,08
)0,
665*
(1,
26)
0,77
8* (
7,69
)0,
845*
(8,
71)
0,49
6* (
0,10
)0,
628*
(0,
44)
0,76
1* (
0,42
)
DX
A-M
ager
gew
ebe
0,74
4* (
0,13
)0,
825*
(2,
58)
0,88
8* (
2,38
)0,
991*
(0,
21)
0,99
2* (
1,50
)0,
997*
(1,
30)
0,58
1* (
0,09
)0,
685*
(0,
40)
0,83
0* (
0,36
)
DX
A-K
noch
enm
iner
al0,
764*
(0,
12)
0,79
7* (
2,77
)0,
875*
(2,
52)
0,94
3* (
0,52
)0,
900*
(5,
15)
0,96
0* (
4,45
)0,
613*
(0,
09)
0,61
5* (
0,44
)0,
802*
(0,
38)
#) B
estim
mth
eits
maß
e m
it *
sind
sig
nifi
kant
ver
schi
eden
von
Nul
l (p
≤ 0,
05).
Ergebnisse
194

Ergebnisse
195
3.2.2.6 Vergleich der Genauigkeit von Magnetresonanz-Tomographie undDualenergie-Röntgenabsorptiometrie zur Ermittlung der Körperzusam-mensetzung
Da von 46 Schweinen ab einer Lebendmasse von 30 kg sowohl Werte aus der MR-
Tomographie als auch aus der DXA-Analyse und parallel deren chemische
Referenzwerte vorliegen, ist ein Vergleich der beiden Methoden MRT und DXA nahe
liegend. Für den Vergleich wurde jeweils eine Stepwise-Regressionsanalyse unter
Verwendung der chemischen Referenzwerte als unabhängige Variablen und der MR-
bzw. DXA-Ergebnisse als abhängige Variablen durchgeführt. Die Signifikanzschwelle
für Aufnahme und Verbleib im Modell beträgt jeweils p ≤ 0,05. Vorab ist festzustellen,
dass ein derartiger Vergleich nur einen begrenzten Aussagewert besitzt, da MR-
Tomographie und Dualenergie-Röntgenabsorptiometrie verschiedene Komponenten der
Körperzusammensetzung messen. Während mittels MR-Tomographie anatomisch
definierbare, dreidimensionale Volumenmaße (cm³) erhoben werden, können mittels
DXA allein aufgrund interner Schätzgleichungen Fett-, Magergewebe- und
Knochengewebemassen (g) bzw. –anteile (%) für zweidimensional festlegbare
Körperregionen oder für den gesamtem Körper erfasst werden. Im vorliegenden
Vergleich sind mittels MR-Tomographie nur zwei Körperregionen (Thorax und
Schinken) in einem 25 cm Abschnitt des gesamten Körpers analysiert worden. Hingegen
umfasste die DXA-Analyse den gesamten Körper. Die chemische Analyse (Fettgehalt
mittels Chloroform/Methanolextraktion nach FOLCH et al., 1957) wiederum wurde nur
zum Teil für die kompletten Schlachtkörper (n = 40) durchgeführt. Von allen
Schlachtkörpern (n = 46) liegen die chemischen Analysewerte der rechten
Schlachthälfte vor. Deshalb erfolgt der Vergleich getrennt nach diesen beiden Kriterien.
Insgesamt existieren nur minimale Unterschiede in der Genauigkeit der beiden
Messverfahren. Mit leichtem Vorteil für die MR-Tomographie resultieren beide
Verfahren in sehr hohen Bestimmtheitsmaßen und relativ niedrigen
Standardschätzfehlern (√MSE) zur Ermittlung des Fettgehalts der rechten
Schlachthälfte (Tab. 3.2.34). Für beide Messverfahren verbleiben zwei Variablen in der
Schätzgleichung, so dass eine Vergleichbarkeit der beiden Methoden gegeben ist. Im
reduzierten Material für die gesamten Schlachtkörper (rechte Schlachthälfte + Kopf +
vollständige Innereien) liegen die Bestimmtheitsmaße (bei niedrigeren
Standardschätzfehlern) sogar geringfügig höher als allein für die rechte Schlachthälfte
(Tab. 3.2.35). Sowohl für den Fettgehalt der rechten Schlachthälfte als auch für den
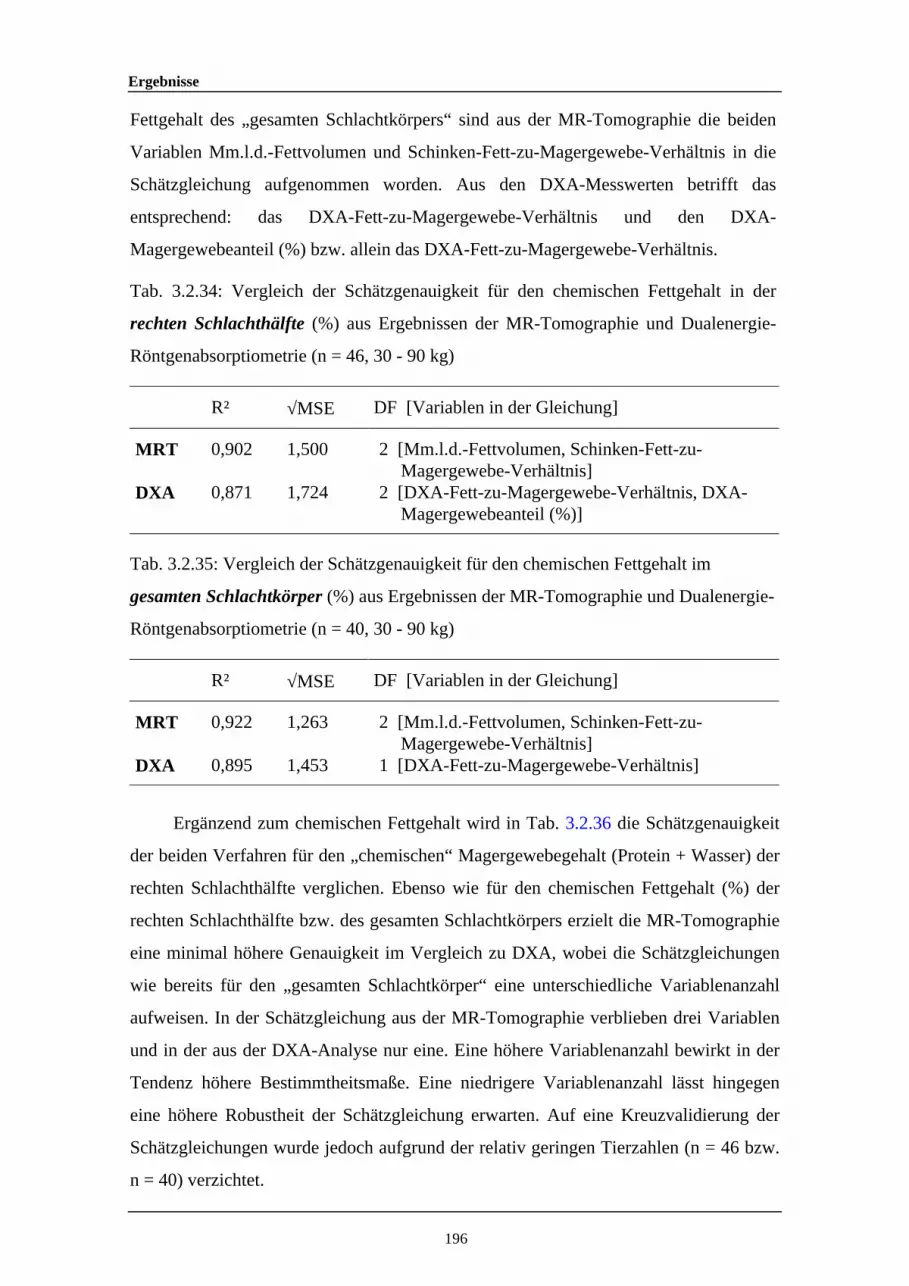
Ergebnisse
196
Fettgehalt des „gesamten Schlachtkörpers“ sind aus der MR-Tomographie die beiden
Variablen Mm.l.d.-Fettvolumen und Schinken-Fett-zu-Magergewebe-Verhältnis in die
Schätzgleichung aufgenommen worden. Aus den DXA-Messwerten betrifft das
entsprechend: das DXA-Fett-zu-Magergewebe-Verhältnis und den DXA-
Magergewebeanteil (%) bzw. allein das DXA-Fett-zu-Magergewebe-Verhältnis.
Tab. 3.2.34: Vergleich der Schätzgenauigkeit für den chemischen Fettgehalt in der
rechten Schlachthälfte (%) aus Ergebnissen der MR-Tomographie und Dualenergie-
Röntgenabsorptiometrie (n = 46, 30 - 90 kg)
R² √MSE DF [Variablen in der Gleichung]
MRT 0,902 1,500 2 [Mm.l.d.-Fettvolumen, Schinken-Fett-zu-Magergewebe-Verhältnis]
DXA 0,871 1,724 2 [DXA-Fett-zu-Magergewebe-Verhältnis, DXA-Magergewebeanteil (%)]
Tab. 3.2.35: Vergleich der Schätzgenauigkeit für den chemischen Fettgehalt im
gesamten Schlachtkörper (%) aus Ergebnissen der MR-Tomographie und Dualenergie-
Röntgenabsorptiometrie (n = 40, 30 - 90 kg)
R² √MSE DF [Variablen in der Gleichung]
MRT 0,922 1,263 2 [Mm.l.d.-Fettvolumen, Schinken-Fett-zu-Magergewebe-Verhältnis]
DXA 0,895 1,453 1 [DXA-Fett-zu-Magergewebe-Verhältnis]
Ergänzend zum chemischen Fettgehalt wird in Tab. 3.2.36 die Schätzgenauigkeit
der beiden Verfahren für den „chemischen“ Magergewebegehalt (Protein + Wasser) der
rechten Schlachthälfte verglichen. Ebenso wie für den chemischen Fettgehalt (%) der
rechten Schlachthälfte bzw. des gesamten Schlachtkörpers erzielt die MR-Tomographie
eine minimal höhere Genauigkeit im Vergleich zu DXA, wobei die Schätzgleichungen
wie bereits für den „gesamten Schlachtkörper“ eine unterschiedliche Variablenanzahl
aufweisen. In der Schätzgleichung aus der MR-Tomographie verblieben drei Variablen
und in der aus der DXA-Analyse nur eine. Eine höhere Variablenanzahl bewirkt in der
Tendenz höhere Bestimmtheitsmaße. Eine niedrigere Variablenanzahl lässt hingegen
eine höhere Robustheit der Schätzgleichung erwarten. Auf eine Kreuzvalidierung der
Schätzgleichungen wurde jedoch aufgrund der relativ geringen Tierzahlen (n = 46 bzw.
n = 40) verzichtet.

Ergebnisse
197
Tab. 3.2.36: Vergleich der Schätzgenauigkeit für den chemischen Magergewebegehalt
(Wasser + Protein) der rechten Schlachthälfte (%) aus Ergebnissen der MR-
Tomographie und Dualenergie-Röntgenabsorptiometrie (n = 46, 30 - 90 kg)
R² √MSE d.f. (Variablen in der Gleichung)
MRI 0,880 1,539 3 [Mm.l.d.-Fettvolumen, Schinken-Fett-zu-Muskel-Verhältnis, Schinken-Magergewebe-Volumenanteil(cm³/kg LM)]
DXA 0,813 1,877 1 (DXA-Fett %)
Zusätzlich ist in Abb. 3.2.16 die Regression zwischen zwei repräsentativen
Messwerten aus der MR-Tomographie und DXA dargestellt (Versuch 1 + 2). Über den
gesamten Lebendmassebereich von 10 – 90 kg besteht erwartungsgemäß bei einem
Bestimmtheitsmaß von R2 = 0,906 (RMSE = √MSE = 1,901) eine sehr enge Beziehung
zwischen dem Rückenfettvolumen über den Mm. longissimi dorsi (cm³) und der DXA-
Fettmasse (kg). Mit zunehmender Lebendmasse erhöht sich jedoch gleichzeitig die
Variation zwischen den beiden Messverfahren. Der Abstand der Merkmalspaare von der
Regressionsgeraden wird größer.
Abb. 3.2.16: Beziehung zwischen DXA-Gesamtkörperfettmasse (kg) und MR-Fettvolumen (cm³) in einem 10 cm MR-Abschnitt über den Mm. longissimi dorsi

Ergebnisse
198
3.2.3 Magergewebewachstum und Magergewebefutterverwertung
Eine Wachstumsstudie an Schweinen mit unterschiedlichen RyR1-Genotypen
(homozygot negativ = NN, heterozygot = Nn und homozygot positiv = nn) diente der
Verlaufskontrolle des Magergewebe- und Fettansatzes von 10 - 90 kg Lebendmasse
(LM) mittels Dualenergie-Röntgenabsorptiometrie (DXA).
Für diesen Teilversuch (Versuch 1) wurden bei 10 kg Lebendmasse 45 Schweine
(13 NN - 14 Nn - 18 nn) mit einem LUNARDPX-L© Scanner untersucht. Ab 30 kg
Lebendmasse wurden 47 Tiere einzeln gehalten und der Futterverzehr wurde individuell
erfasst. Aufgrund der durchgeführten Stufenschlachtung verblieben für die 60 kg
Gruppe 32 Tiere und für die 90 kg Gruppe 17 Tiere im Versuch.
Da aus den DXA-Messungen unmittelbar Magergewebemasse, Fettgewebemasse
und Knochenmineralgehalt des gesamten Tieres resultieren (Kapitel 3.2.2.4), lassen sich
aus diesen Werten in Kombination mit dem Futterverzehr und dem Lebensalter
individuell die Magergewebewachstumsrate (g/d) und die Magergewebe-
futterverwertung (kg/kg) in den einzelnen Versuchsabschnitten berechnen. Zusätzlich
wurden die Fettgewebewachstumsrate und die Knochenmineralzunahme aus den
Differenzen der Fettgewebe- bzw. Knochenmineralmessungen zwischen den vier
Messzeitpunkten bei ca. 10 kg, 30 kg, 60 kg und 90 kg ermittelt. Die
Magergewebewachstumsrate für den Abschnitt von Geburt bis zu 10 kg Lebendmasse
wurde unter der Annahme ermittelt, dass sich der prozentuale Magergewebegehalt der
Individuen zum Zeitpunkt der Geburt (1,2 kg Lebendmasse) proportional zum
Magergewebegehalt zum Zeitpunkt der Messung bei 10 kg verhält. Dafür wurde die
lineare Regressionsgleichung zwischen Magergewebegehalt und Lebendmasse zum
Zeitpunkt der DXA-Messung für alle untersuchten Tiere berechnet:
DXA-Magergewebegehalt (%) = 93,6386 0,1697 • Lebendmasse.
Die Gleichung für die Berechnung der Magergewebewachstumsrate vom Zeitpunkt der
Geburt (1,2 kg) bis zur ersten DXA-Messung bei ca. 10 kg Lebendmasse (MGWR10)
lautet:
MGWR10 (g/d) = (Magergewebemasse10 - (1,2 • ((Magergewebegehalt10/100) + (0,1697/100) • 1,2 ))) / Alter10 • 1000
mit Magergewebemasse10 = Magergewebemasse bei 10 kg Messung (kg),
Magergewebegehalt10 = Magergewebegehalt bei 10 kg Messung (%) und
Alter10 = Alter bei 10 kg Messung (Lebenstage).

Ergebnisse
199
Da die Tiere, die bei 10 kg mittels DXA untersucht wurden, bei 30 kg nicht mehr zur
Verfügung standen, wurde für den Abschnitt von ca. 10 bis 30 kg Lebendmasse die
Magergewebewachstumsrate unter Benutzung desselben Regressionskoeffizienten wie
oben (b = 0,1697) berechnet. Alle Tiere, die bei 30 kg mittels DXA untersucht wurden,
dienten vorher bei ca. 10 kg zur Untersuchung mittels MR-Tomographie bzw. MR-
Spektroskopie, so dass das Alter und die Lebendmasse zum Zeitpunkt der MR-
Untersuchung als Anhaltspunkt für den 10 kg Bereich zur Verfügung standen. Die
Formel lautet:
MGWR1030 (g/d) = (Magergewebemasse30 - (Lebendmasse10 • ((Magergewebegehalt30/100) + (0,1697/100) • Lebendmasse10)))/(Alter30 - Alter10) • 1000.
Die Magergewebewachstumsrate (MGWR) und Magergewebefutterverwertung
(MFUV) für die Gewichtsabschnitte 30 60 kg bzw. 60 90 kg wurden in Anlehnung
an die Formel von FOWLER et al. (1976) berechnet:
MGWR (g/d) = ∆ Magergewebemasse (g) / ∆ Alter (d)
MFUV (kg/kg) = Futterverzehr (kg/d) / (MGWR (g/d) /1000)∆ = Differenz zwischen den Gewichtsabschnitten
Tab. 3.2.37: Tageszunahme (TZ10) und Magergewebewachstumsrate (MGWR10) in
Schweinen mit unterschiedlichen RyR1-Genotypen von Geburt (1,2 kg) bis zur DXA-
Messung bei 10 kg Lebendmasse (LSM ± SEE#)
n TZ10 (g) MGWR10 (g/d)
NN 13 212 ± 16 a 194 ± 14 a
Nn 14 196 ± 18 a 181 ± 16 a
nn 18 152 ± 22 b 140 ± 19 b
#) Werte mit unterschiedlichen Buchstaben sind signifikant verschieden (p ≤ 0,05).
Tab. 3.2.38: Tageszunahme (TZ1030) und Magergewebewachstumsrate (MGWR1030)
in Schweinen mit unterschiedlichen RyR1-Genotypen im Lebendmasseabschnitt von 10
bis 30 kg (LSM ± SEE#)
n TZ1030 (g) MGWR1030 (g/d)
NN 16 545 ± 21 475 ± 19 *
Nn 22 565 ± 15 492 ± 13
nn 9 599 ± 25 527 ± 22 °
#) Differenzen zwischen Werten mit * und ° überschreiten die Signifikanzgrenze nur minimal (p ≤0,067).
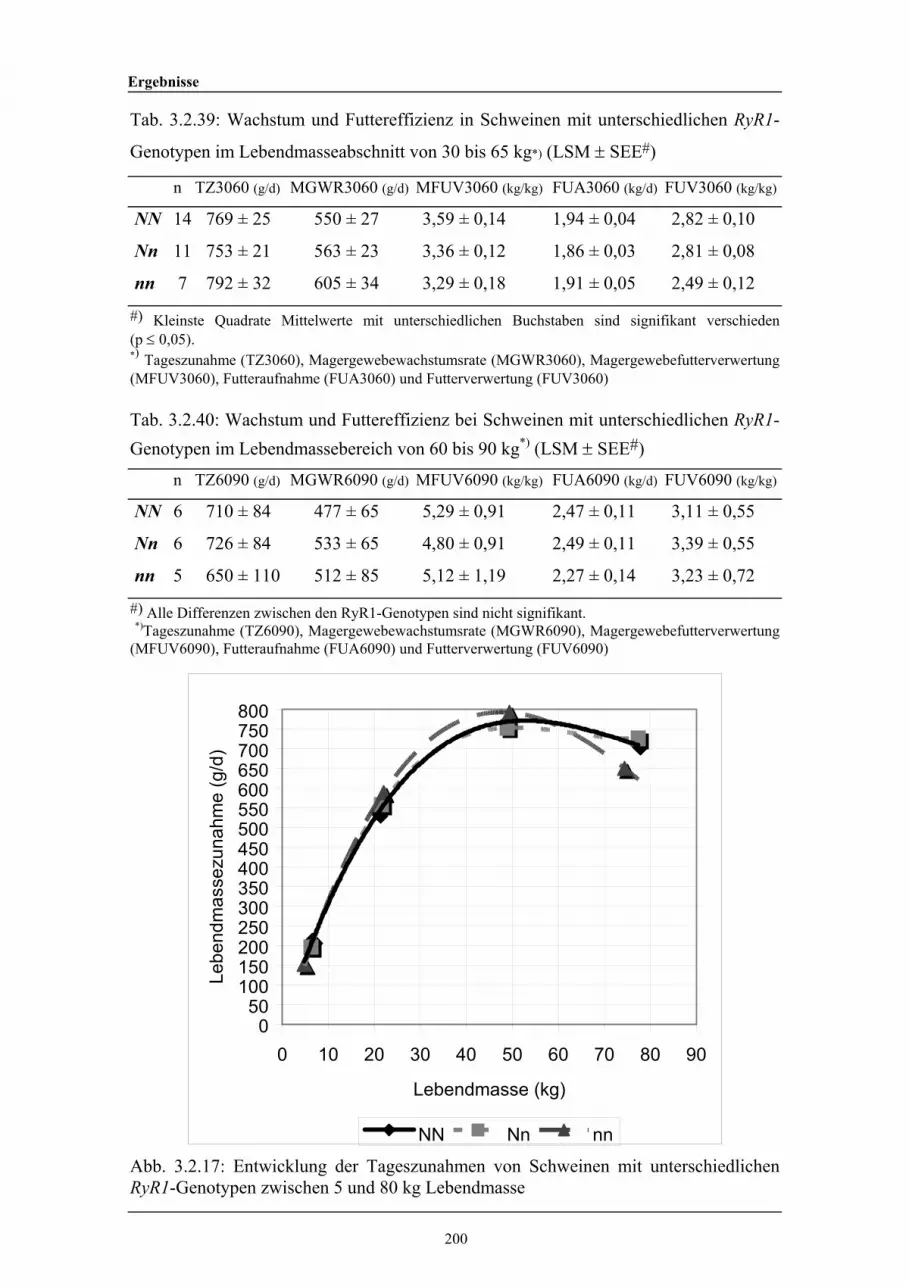
Ergebnisse
200
Tab. 3.2.39: Wachstum und Futtereffizienz in Schweinen mit unterschiedlichen RyR1-
Genotypen im Lebendmasseabschnitt von 30 bis 65 kg*) (LSM ± SEE#)
n TZ3060 (g/d) MGWR3060 (g/d) MFUV3060 (kg/kg) FUA3060 (kg/d) FUV3060 (kg/kg)
NN 14 769 ± 25 550 ± 27 3,59 ± 0,14 1,94 ± 0,04 2,82 ± 0,10
Nn 11 753 ± 21 563 ± 23 3,36 ± 0,12 1,86 ± 0,03 2,81 ± 0,08
nn 7 792 ± 32 605 ± 34 3,29 ± 0,18 1,91 ± 0,05 2,49 ± 0,12
#) Kleinste Quadrate Mittelwerte mit unterschiedlichen Buchstaben sind signifikant verschieden(p ≤ 0,05).*) Tageszunahme (TZ3060), Magergewebewachstumsrate (MGWR3060), Magergewebefutterverwertung(MFUV3060), Futteraufnahme (FUA3060) und Futterverwertung (FUV3060)
Tab. 3.2.40: Wachstum und Futtereffizienz bei Schweinen mit unterschiedlichen RyR1-Genotypen im Lebendmassebereich von 60 bis 90 kg*) (LSM ± SEE#)
n TZ6090 (g/d) MGWR6090 (g/d) MFUV6090 (kg/kg) FUA6090 (kg/d) FUV6090 (kg/kg)
NN 6 710 ± 84 477 ± 65 5,29 ± 0,91 2,47 ± 0,11 3,11 ± 0,55
Nn 6 726 ± 84 533 ± 65 4,80 ± 0,91 2,49 ± 0,11 3,39 ± 0,55
nn 5 650 ± 110 512 ± 85 5,12 ± 1,19 2,27 ± 0,14 3,23 ± 0,72
#) Alle Differenzen zwischen den RyR1-Genotypen sind nicht signifikant. *)Tageszunahme (TZ6090), Magergewebewachstumsrate (MGWR6090), Magergewebefutterverwertung(MFUV6090), Futteraufnahme (FUA6090) und Futterverwertung (FUV6090)
050
100150200250300350400450500550600650700750800
0 10 20 30 40 50 60 70 80 90
Lebendmasse (kg)
Lebe
ndm
asse
zuna
hme
(g/d
)
NN Nn nnAbb. 3.2.17: Entwicklung der Tageszunahmen von Schweinen mit unterschiedlichenRyR1-Genotypen zwischen 5 und 80 kg Lebendmasse

Ergebnisse
201
050
100150200250300350400450500550600650
0 10 20 30 40 50 60 70 80 90
Lebendmasse (kg)
Mag
erge
web
ewac
hstu
m (g
/d)
NN Nn nn
Abb. 3.2.18: Entwicklung der Magergewebewachstumsrate in Schweinen mitunterschiedlichen RyR1-Genotypen zwischen 5 und 80 kg Lebendmasse
400
450
500
550
600
650
1,75 1,85 1,95 2,05 2,15 2,25 2,35 2,45 2,55
Futterverzehr (kg/d)
Mag
erge
web
ewac
hstu
m (g
/d)
NNNnnn
30 - 60 kg LM 60 - 90 kg LM
Abb. 3.2.19: Durchschnittliche Futteraufnahme (kg/d) und Magergewebe-wachstumsrate (g/d) in Abhängigkeit vom Prüfabschnitt bei Schweinen mitunterschiedlichen RyR1-Genotypen
Bei minimal unterschiedlichen Tageszunahmen steigt die Magergewebe-
wachstumsrate (g/d) mit zunehmendem Alter bzw. bis zu einer Lebendmasse von ca. 40

Ergebnisse
202
kg bei allen drei Genotypen stetig an. Aufgrund der mittels DXA ermittelten
fettärmeren Körperzusammensetzung und gleichzeitig annähernd identischen
Lebendmasse-zunahmen (Abb. 3.2.17) liegt ab ca. 15 - 20 kg die
Magergewebewachstumsrate (g/d) der homozygoten Defektallelträger nn über den
Magergewebewachstumsraten der beiden anderen Genotypen. Erst gegen Ende des
Versuchs nähern sich die Magergewebewachstumsraten der beiden
Defektallelgenotypen wieder an. Aufgrund der im zweiten Prüfabschnitt (60 - 90 kg
Lebendmasse) nur geringfügig gesteigerten Futteraufnahme - im Vergleich zum ersten
Prüfabschnitt (30 60 kg Lebendmasse) - nehmen das Lebendmassewachstum und das
Magergewebewachstum im homozygoten Defektallelgenotyp stärker ab als im
heterozygoten Genotyp. Der homozygot normale Genotyp nimmt genauso schnell wie
der heterozygote Genotyp an Lebendmasse zu, liegt aber aufgrund des höheren
Fettanteils unter den Magergewebewachstumsraten der beiden Defektallelgenotypen.
Da das Magergewebe hauptsächlich aus den zwei Komponenten Protein und
Wasser (+ minimale Anteile von Kohlenhydraten [Glykogen] und Mineralstoffen)
besteht, lässt sich aus der Magergewebewachstumsrate die Proteinansatzrate ableiten.
Unter Verwendung einer modifizierten Formel von MITCHELL and CONWAY (1994) zur
Berechnung des Proteingehaltes aus der DXA-Magergewebemasse
[Proteinansatz (g/d) = -1,062 + 0,22 • DXA-Magergewebewachstumsrate (g/d)]
ergeben sich die in Tabelle 3.2.24 dargestellten Proteinansatzraten in den untersuchten
Gewichtsabschnitten. Die Fettansatzraten resultieren aus der Differenz von
Lebendmassezunahme und Magergewebewachstumsrate abzüglich der Knochen-
mineralzunahme. Der unbekannte Knochenmineralgehalt zum Zeitpunkt der Geburt mit
einer angenommenen Lebendmasse von 1,2 kg wurde aus der am eigenen Material
berechneten Regressionsgleichung abgeleitet:
[DXA-Knochenmineralgehalt (%) = 2,5674 (0,0041 • Lebendmasse)].
Aus dieser Regressionsgleichung ergibt sich die Knochenmineralwachstumsrate
zwischen Geburt (1,2 kg) und 10 kg DXA-Messung mit:
Knochenmineralzunahme (g/d) =
(((DXABMPC10/100 • DXALM10) - (1,2 • ((DXABMPC10/100) + (0,0041/100) • 1,2))))/DXAAlter10 • 1000
mit
DXABMPC10 = Knochenmineralgehalt (%) aus DXA-Messung in der 10 kg Gruppe,
DXALM10 = Lebendmasse zum Zeitpunkt der DXA-Messung in der 10 kg Gruppe (kg) und
DXAAlter10 = Alter zum Zeitpunkt der DXA-Messung in der 10 kg Gruppe (Tage).

Ergebnisse
203
Tab. 3.2.41: Protein-, Fett- und Knochenmineralansatzraten (g/d) in Schweinen mit
unterschiedlichen RyR1-Genotypen bei einer Fütterung mit 95% der geschätzten
freiwilligen Futteraufnahme
RyR1-Genotyp
NN Nn nn
Wachs-
tumsab-
schnitt Zunahme (g/d)
Protein Fett Knochen Protein Fett Knochen Protein Fett Knochen
6,56 kg 6,27 kg 4,81 kgI*)
42 ± 3 13 ± 2 6 ± 1 39 ± 4 10 ± 2 5 ± 1 30 ± 4 8 ± 3 4 ± 1
20,79 kg 21,50 kg 21,50 kgII
104 ± 4 57 ± 4 13 ± 1 107 ± 3 59 ± 3 13 ± 0 115 ± 5 58 ± 5 14 ± 1
48,82 kg 48,91 kg 49,38 kgIII
120 ± 6 199 ± 13 19 ± 1 123 ± 5 172 ± 11 19 ± 1 132 ± 7 168 ± 16 19 ± 1
77,16 kg 77,37 kg 74,33 kgIV
104 ± 14 225 ± 45 9 ± 1 116 ± 14 183 ± 45 9 ± 1 112 ± 19 125 ± 58 12 ± 1
*) Durchschnittliche Lebendmassen in den VersuchsabschnittenI: 1,2 kg ca. 10 kg, II: 10 kg 30 kg, III: 30 kg 60 kg, IV: 60 90 kg.
Da bei allen drei Genotypen die Magergewebewachstumsrate gegen Ende des
Versuchs bei steigendem täglichen Futterverzehr leicht abnimmt, kann das häufig
angenommene Linear-Plateau-Modell für den Proteinansatz in Bezug auf die
Energieaufnahme (WHITTEMORE, 1985, KANIS, 1988, BIKKER, 1994) in diesem Versuch
nicht bestätigt werden (Abb. 3.2.18, 3.2.19). Das Linear-Plateau-Modell unterstellt
einen steilen Anstieg der Proteinansatzrate bei steigender Energieaufnahme bis zum
Erreichen des maximalen Proteinansatzpotentials. Nach Erreichen der maximalen
Proteinansatzrate bleibt der Proteinansatz auch bei weiter steigender Energiezufuhr
konstant. Jedoch verweisen BIKKER et al. (1996a,b) darauf, dass bei einem relativ
niedrigen Futteraufnahmeniveau wie im eigenen Versuch zwischen 30 und 60 kg
Lebendmasse - anteilmäßig mehr Magergewebe (Muskelgewebe bzw. Protein)
angesetzt wird als bei einem hohen Futteraufnahmeniveau, welches den Fettansatz
fördert.

Ergebnisse
204
0102030405060708090
100110120130140150160170180190200210220230240250
0 10 20 30 40 50 60 70 80 90
Lebendmasse (kg)
Zuna
hme
(g/d
)
NN_Protein Nn_Protein nn_ProteinNN_Fett Nn_Fett nn_FettNN_Knochen Nn_Knochen nn_Knochen
Abb. 3.2.20: Wachstumsraten für Protein, Fett und Knochenmineral bei Schweinen mitunterschiedlichen RyR1-Genotypen
Nach einem schnellen Anstieg zwischen Geburt und 20 kg Lebendmasse und
einem Maximum zwischen 45 und 60 kg sinkt die Proteinansatzrate bis zum Ende des
Versuchs bei ca. 90 kg Lebendmasse wieder minimal. Die Ergebnisse von SCHINCKEL
et al. (1996) decken sich mit den eigenen Ergebnissen für das Ende des Versuchs (90
kg) und zeigen in verschiedenen Genotypen einen deutlichen Rückgang der
Proteinansatzraten ab 60 kg Lebendmasse, während WHITTEMORE et al. (1988) einen
erheblichen Rückgang in der Proteinansatzrate erst ab ca. 100 kg Lebendmasse
verzeichnen. Ähnlich wie in den eigenen Ergebnissen wenn man den RyR1-Genotyp
NN ausnimmt - wich die Proteinansatzrate im Bereich zwischen 45 und 110 kg
Lebendmasse maximal um 10 g von der maximalen Proteinansatzrate ab. Im eigenen
Versuch liegen die maximalen Proteinansatzraten (PDmax) theoretisch um 5 % über
den ermittelten Werten, da die Tiere im Versuch nur mit 95 % der geschätzten Ad-
libitum-Futteraufnahme versorgt wurden (Kapitel 3.1.3). Die durchschnittliche
maximale Proteinansatzrate für den RyR1-Genotyp nn beträgt unter den gegebenen

Ergebnisse
205
Bedingungen (Rasse, Fütterung und Haltung) ca. 139 g/d, während für den
heterozygoten Genotyp eine durchschnittliche maximale Proteinansatzrate von ca.
129 g/d bzw. für den homozygot normalen Genotyp von ca. 126 g/d zu veranschlagen
ist. Ein Kastrat der Linie Duroc (nn) erreichte mit 164 g/d (PDmax = 172 g/d) die
höchste Proteinansatzrate im Gewichtsabschnitt zwischen 30 und 60 kg.
Tiere mit höchstem Fleischanteil (Magergewebeanteil) müssen nicht zwingend die
höchsten Protein- bzw. Magergewebeansatzraten aufweisen. Sobald die fleischreicheren
Tiere (wie z.B. Pietrain oder homozygote Defektallelträger) aufgrund einer
vergleichsweise niedrigeren Futteraufnahme stark reduzierte Tageszunahmen erzielen,
sinken die Proteinansatzraten unter die der Tiere mit einem mittleren Verfettungsgrad -
hier heterozygoter Genotyp Nn ab (Tab. 3.2.41, Abb. 3.2.20). Dabei ist vorausgesetzt,
dass die Tiere mit einem mittleren Verfettungsgrad mehr Futter für die Realisierung
höchster Tageszunahmen aufnehmen als die magersten Tiere (WÄFLER u.a., 1984,
KRIETER, 1986, KANIS, 1988, WEBB, 1989, CAMERON and CURRAN, 1994).
In der eigenen Untersuchung zeigt der fleischreiche Genotyp nn im letzten
Prüfabschnitt (60 - 90 kg) im Vergleich zum vorangegangenen Prüfabschnitt (30 - 60
kg) neben einer verminderten Tageszunahme eine verminderte
Magergewebewachstumsrate und folglich eine verminderte Proteinansatzrate.
Korrespondierend ergibt sich im Vergleich zu den beiden anderen Genotypen NN und
Nn eine geringere Futteraufnahme. Insgesamt resultiert aus den verminderten
Lebendmassezunahmen und der niedrigsten Futteraufnahme eine mittlere
Futterverwertung für den Genotyp nn. Bezogen auf die Magergewebemasse ergibt sich
aufgrund der höheren Magergewebewachstumsraten die beste Futterverwertung für den
heterozygoten Genotyp (Abb. 3.2.19, Tab 3.2.33).

Ergebnisse
206
3.2.4 Muskelfaserstruktur und ihre Beziehung zu Parametern aus der 31P-MR-Spektroskopie
Muskelfaserstrukturmerkmale werden als wichtiger Indikator für die Stressstabilität und
mit der Belastungsstabilität einhergehender Fleischqualität angesehen.
Besonders die Muskelfaserquerschnittsfläche zeigte erhebliche Unterschiede beim
Vergleich des normalen Genotyps (NN) mit den beiden Genotypen, die das Defektallel
tragen (Nn und nn).
T
TT
Abb. 3.2.21: Vergleich der Muskelfaserquerschnitte in Abhängigkeit vom RyR1-Genotyp: oben: kleine Faserquerschnitte - NN -
unten: große Faserquerschnitte - Nn und nn -

Ergebnisse
207
Im normalen Genotyp ist die Muskelfaserquerschnittsfläche je nach Typ signifikant
um 35 % bis 57 % kleiner als in den beiden anderen Genotypen, während sich die
Genotypen mit mindestens einer Kopie des Defektallels in der
Muskelfaserquerschnittsfläche nicht signifikant unterscheiden (Abb. 3.2.21, Tab.
3.2.42).
Tab. 3.2.42: Muskelfaserquerschnittsflächen der Muskelfasertypen I, IIa und IIb sowie
Kotelettflächen von Schweinen mit unterschiedlichen RyR1-Genotypen bei einem
Durchschnittsgewicht von 83,48 kg und 152,77 Lebenstagen (LSM ± SEE#)
a
b
b
b b
b
a
n n N n N N
a a b 49 ,6 4 ± 1 ,46 4 0 ,93 ± 0 ,9 1 42 ,1 4 ± 1 ,09ØM .l.d . (cm ²)
b6 41 9 ± 38 2
5 17 9 ± 56 54 43 7 ± 35 0
5 22 0 ± 71 75 08 8 ± 44 4
1 94 6 ± 4 2 0
T yp IIb 41 2 3 ± 4 6 0
T yp I
T yp IIa
2 58 5 ± 5 3 4 a
6 83 7 ± 61 7
F läche (µ m ²)
10 n 7 5
a bb 6 60 1 ± 60 36 01 6 ± 37 3 3 75 8 ± 4 4 9X
#) Werte mit unterschiedlichen Buchstaben sind signifikant verschieden (p ≤ 0,05).
Außerdem bestehen zwischen dem normalen und dem heterozygoten Genotyp keine
signifikanten Unterschiede in der Fläche des M. longissimus dorsi (Kotelettfläche).
Zwischen 60 und 90 kg Lebendmasse nehmen die Muskelfaser-
querschnittsflächen in der Tendenz um ca. 39,3 µm² je kg Lebendmasse zu (r = 0,27,
n.s., Abb. 3.2.22). Die Anzahl der Muskelfasern ändert sich erwartungsgemäß nicht.
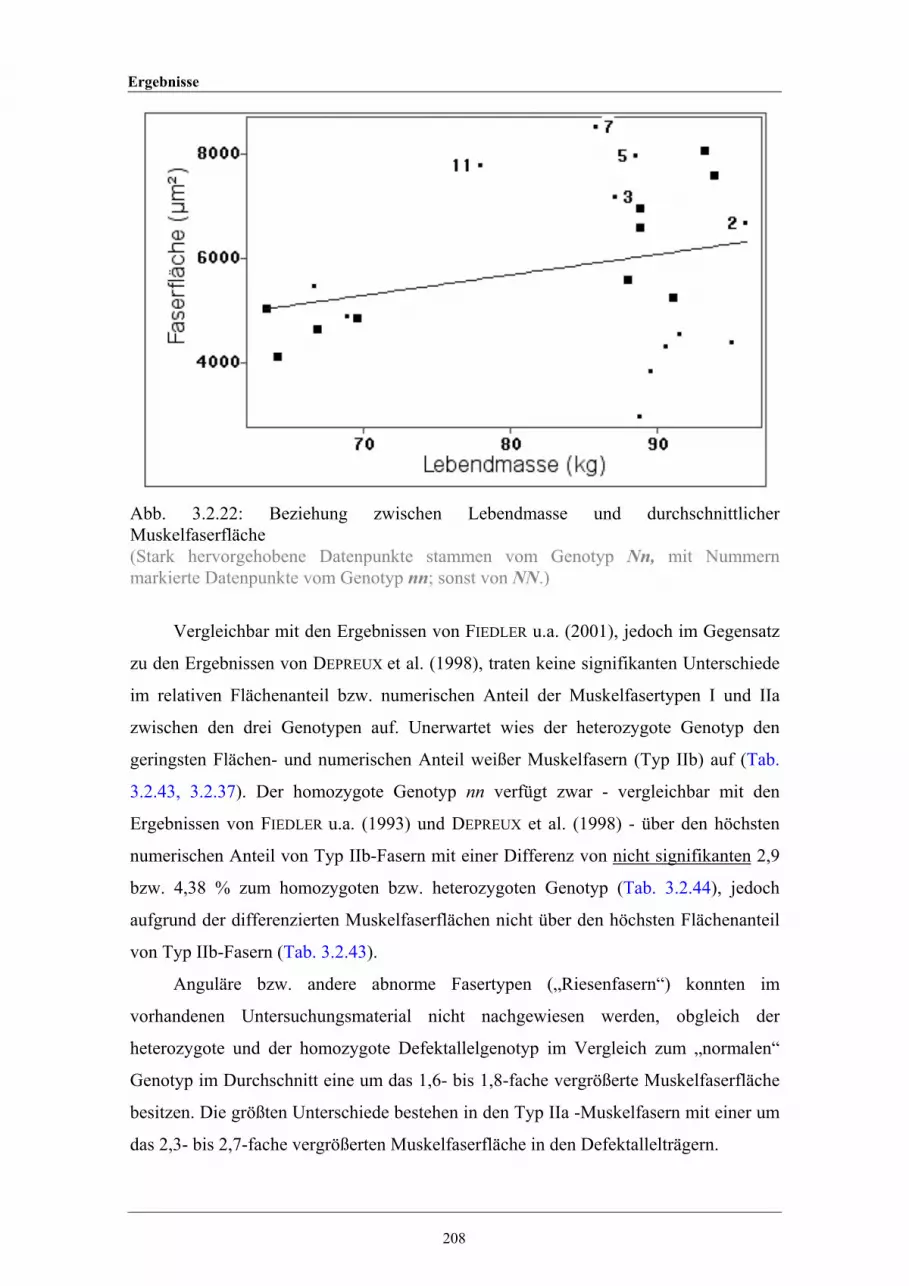
Ergebnisse
208
Abb. 3.2.22: Beziehung zwischen Lebendmasse und durchschnittlicherMuskelfaserfläche (Stark hervorgehobene Datenpunkte stammen vom Genotyp Nn, mit Nummernmarkierte Datenpunkte vom Genotyp nn; sonst von NN.)
Vergleichbar mit den Ergebnissen von FIEDLER u.a. (2001), jedoch im Gegensatz
zu den Ergebnissen von DEPREUX et al. (1998), traten keine signifikanten Unterschiede
im relativen Flächenanteil bzw. numerischen Anteil der Muskelfasertypen I und IIa
zwischen den drei Genotypen auf. Unerwartet wies der heterozygote Genotyp den
geringsten Flächen- und numerischen Anteil weißer Muskelfasern (Typ IIb) auf (Tab.
3.2.43, 3.2.37). Der homozygote Genotyp nn verfügt zwar - vergleichbar mit den
Ergebnissen von FIEDLER u.a. (1993) und DEPREUX et al. (1998) - über den höchsten
numerischen Anteil von Typ IIb-Fasern mit einer Differenz von nicht signifikanten 2,9
bzw. 4,38 % zum homozygoten bzw. heterozygoten Genotyp (Tab. 3.2.44), jedoch
aufgrund der differenzierten Muskelfaserflächen nicht über den höchsten Flächenanteil
von Typ IIb-Fasern (Tab. 3.2.43).
Anguläre bzw. andere abnorme Fasertypen (Riesenfasern) konnten im
vorhandenen Untersuchungsmaterial nicht nachgewiesen werden, obgleich der
heterozygote und der homozygote Defektallelgenotyp im Vergleich zum normalen
Genotyp im Durchschnitt eine um das 1,6- bis 1,8-fache vergrößerte Muskelfaserfläche
besitzen. Die größten Unterschiede bestehen in den Typ IIa -Muskelfasern mit einer um
das 2,3- bis 2,7-fache vergrößerten Muskelfaserfläche in den Defektallelträgern.

Ergebnisse
209
Tab. 3.2.43: Flächenanteile der Muskelfasertypen I, IIa und IIb in unterschiedlichen
RyR1-Genotypen (LSM ± SEE#)
n n N n N N
5 ,4 8 ± 1 ,2 6 7 ,4 5 ± 1 ,0 5 7 ,8 6 ± 1 ,6 9
8 5 ,8 7 ± 1 ,7 08 3 ,7 0 ± 1 ,0 5
6 ,8 8 ± 1 ,2 6 8 ,8 5 ± 1 ,0 5 6 ,2 7 ± 1 ,7 0
T y p IIb 8 7 ,6 4 ± 1 ,2 6
T y p IIa
T y p I
a b ab
F läch en an te il (% )
#) Werte mit unterschiedlichen Buchstaben sind signifikant verschieden (p ≤ 0,05).
Tab. 3.2.44: Prozentuale Verteilung der Anzahl der Muskelfasertypen I, IIa und IIb in
unterschiedlichen RyR1-Genotypen (LSM ± SEE#)
T y p IIa
T y p I
N N N n n n
8 ,0 5 ± 2 ,1 3 9 ,3 5 ± 1 ,7 7 9 ,3 9 ± 2 ,8 6
1 2 ,0 3 ± 1 ,5 9 1 2 ,2 1 ± 1 ,3 2 7 ,7 9 ± 2 ,1 3
T y p IIb 7 9 ,9 2 ± 1 ,9 2 7 8 ,4 4 ± 1 ,6 0 8 2 ,8 2 ± 2 ,5 7
A n zah l (% )
#) Werte mit unterschiedlichen Buchstaben sind signifikant verschieden (p ≤ 0,05).
In der Tendenz - bei einer Korrelation von r = -0,56 - nahm der numerische Anteil
von Typ-IIbFasern mit steigender Lebendmasse von 63,4 bis 96,2 kg ab, während der
numerische Anteil von Typ I Fasern leicht anstieg (r = 0,42). Der Anteil von Typ-IIa
Fasern zeigte einen minimalen nicht signifikanten Anstieg mit zunehmenden Alter bzw.
Lebendmasse. Bezogen auf die Fläche resultieren Korrelationen von r = - 0,66 zwischen
Typ-IIb-Faseranteil und Lebendmasse bzw. von r = 0,57 zwischen Typ-I-Faseranteil
und Lebendmasse. Nur minimal geringere Korrelationskoeffizienten ergeben sich in
Bezug auf das Alter im Bereich von 120 bis 177 Lebenstagen. Diese Ergebnisse stehen
einerseits im Gegensatz zu den von KARLSSON et al. (1999) zusammengefassten
Erkenntnissen, die von einer Zunahme der glykolytischen Fasern (Typ IIa und IIb ) und
einer Abnahme der oxidativen Fasern (Typ I) im Laufe des Wachstums speziell für den
untersuchten M. longissimus dorsi ausgehen. Eine Ursache in den kontroversen
Ergebnissen könnte darin bestehen, dass im eigenen Material keine fortlaufenden
Untersuchungen an einzelnen Individuen vorliegen, sondern Tiere mit
unterschiedlichem Alter und Gewicht in die Untersuchung eingingen. Andererseits

Ergebnisse
210
weist ESSÉN-GUSTAVSSON (1993) die eigenen Ergebnisse bestätigend - darauf hin,
dass der Anteil der Typ-I-Fasern bis zur sexuellen Reife zunimmt und erst in älteren
Schweinen wieder zurückgeht.
Aus der planimetrierten Fläche des M. longissimus dorsi (Tab. 3.2.42) und der
durchschnittlichen Muskelfaserquerschnittsfläche ergibt sich die Gesamtanzahl
Muskelfasern:
[Muskelfaseranzahl = M.l.d. (cm²) x 100000 / durchschnittliche Faserfläche (µm²)].
Die Muskelfaseranzahl ist bereits pränatal festgelegt und lässt sich durch äußere
Einflüsse wie extreme Futterrestriktion nur minimal ändern (SWATLAND, 1993,
REHFELDT et al., 1999). Zwischen Muskelfaseranzahl und Muskelfaserfläche existiert
übereinstimmend mit den Ergebnissen von REHFELDT et al. (1999) eine hohe negative
Korrelation von r = -0,833 (Abb. 3.2.23, n = 22).
Abb. 3.2.23: Beziehung zwischen Muskelfaseranzahl und Muskelfaserfläche
(r = -0,833)
Der homozygot normale Genotyp NN verfügt infolge der kleinsten
Faserquerschnittsflächen über die höchste Anzahl von Muskelfasern innerhalb des M.
longissimus dorsi (M.l.d.). Aufgrund der größten Fläche des M.l.d. (Koteletts) bei
gleichzeitig größten Faserquerschnittsflächen folgt der homozygote Defektgenotyp nn

Ergebnisse
211
an nächster Stelle (p = 0,0566). Beruhend auf der großen Muskelfaserquerschnittsfläche
bei gleichzeitig kleiner Fläche des M. longissimus dorsi weist Nn die niedrigste Anzahl
Muskelfasern auf, die sich jedoch nicht signifikant von nn unterscheidet (Tab. 3.2.45).
Ähnlich wie in den vorliegenden Ergebnissen fanden FIEDLER u.a. (2001) keine
statistisch gesicherten Unterschiede in der Fasergesamtzahl zwischen den homozygoten
RyR1-Genotypen (NN und nn), wobei die Probenahme post mortem und nicht in vivo
erfolgte. Eine Muskel(faser)hypertrophie muss demzufolge nicht notwendigerweise mit
einer reduzierten Muskelfaseranzahl gekoppelt sein, obwohl eine enge negative
Korrelation zwischen den beiden Merkmalen besteht. Generell besitzen jedoch die
Defektallelträger (Nn und nn) im Vergleich zum normalen Genotyp aufgrund der
größeren Muskelfaserflächen eine verminderte Anzahl an Muskelfasern. Im eigenen
Material besteht keine signifikante Beziehung zwischen Muskelfaserfläche und
Muskelfläche (Kotelett) (r = 0,24).
Tab. 3.2.45: Gesamtanzahl Muskelfasern im M. longissimus dorsi in Abhängigkeit vom
RyR1-Genotyp (LSM ± SEE#)
b a°b
nn N n N N
840 ,7 ± 113 ,9708 ,6 ± 70 ,5A nzahl F asern 1094 ,1 ± 85 ,4 a*
(T ausend)
#) Werte mit unterschiedlichen Buchstaben unterscheiden sich signifikant mit p ≤ 0,05. Differenzen mit * und ° überschreiten die Signifikanzgrenze minimal mit p = 0,0566.(M.l.d.-Fläche zwischen 13./14. Brustwirbel direkt nach Schlachtung planimetrisch gemessen)
Da die mittels Biopsie untersuchten Tiere bereits bei ca. 10 kg Lebendmasse einer
Stoffwechselanalyse mit Hilfe der 31P-MR-Spektroskopie unterzogen wurden, konnten
die Beziehungen zwischen Stoffwechselreaktion (Phosphokreatinabbau) und
durchschnittlicher Muskelfaserfläche berechnet werden (Abb. 3.2.24). Für den
Aussagewert dieser Berechnung ist jedoch der zeitliche Abstand zwischen den beiden
Untersuchungen sowie die Verwendung unterschiedlicher Muskelgruppen (M.l.d. für
Biopsie bzw. M. biceps femoris für MR-Spektroskopie) zu berücksichtigen. Zum
Zeitpunkt der 31P-MR-Untersuchung (10 kg) konnten noch keine Biopsieproben
entnommen werden, da das Schussbiopsiegerät erst später verfügbar war. Insgesamt
nimmt die Phosphokreatinabbaurate und damit die Stressanfälligkeit mit steigender
Muskelfaserfläche zu (r = 0,533). Zwischen der Muskelfaseranzahl und der
Stressanfälligkeit (Phosphokreatinabbau, %) konnte hingegen mit r = -0,30 (n = 22)

Ergebnisse
212
keine signifikante Beziehung ermittelt werden (Abb. 3.2.25). Die Höhe der Korrelation
zwischen Muskelfaserfläche bzw. Muskelfaseranzahl und Phosphokreatinabbau hängt
stark von der untersuchten RyR1-Genotypzusammensetzung ab, denn innerhalb der
RyR1-Genotypgruppen bestehen nicht allein wegen der geringen Beobachtungszahl -
keine signifikanten Beziehungen zwischen den Merkmalen (Abb. 3.2.26).
Abb. 3.2.24: Beziehung zwischen der auf einheitliche 90 kg Lebendmasse korrigiertenMuskelfaserfläche am M. longissimus dorsi*) und dem Phosphokreatinabbau nachStressauslösung mittels Halothan am M. biceps femoris (bei ca. 10 kg Lebendmasse)Regressionsgleichung zur Korrektur der Muskelfaserfläche auf 90 kg Lebendmasse:*)Muskelfaserfläche90 = gemessene Muskelfaserfläche + [39,3757 • (90 - Lebendmasse)]Zwischen Phosphokreatinabbau und Lebendmasse innerhalb des untersuchten Lebendmassebereiches(6,14 14,96 kg) besteht kein signifikanter Zusammenhang.

Ergebnisse
213
Abb. 3.2.25: Beziehung zwischen Muskelfaseranzahl am M. longissimus dorsi und demPhosphokreatinabbau nach Stressauslösung mittels Halothan am M. biceps femoris(r = 0,367, p = 0,093)
Abb. 3.2.26: Beziehung zwischen Muskelfaseranzahl am M. longissimus dorsi und demPhosphokreatinabbau nach Stressauslösung mittels Halothan am M. biceps femorisinnerhalb der RyR1-Genotypen

Ergebnisse
214
Übereinstimmend mit den Erkenntnissen von SCHMITTEN u.a. (1986),
RATHFELDER (1991), ESSÉN-GUSTAVSSON (1995), V. LENGERKEN u.a. (1997), MARTENS
(1997, 1998), KARLSSON et al. (1999) und FIEDLER u.a. (2001) ist generell davon
auszugehen, dass Tiere mit einem erhöhten Anteil weißer (glykolytischer) Muskelfasern
und vor allem mit größeren, hypertrophierten Muskelfasern belastungsanfälliger sind
und eine schlechtere Fleischqualität aufweisen als Tiere mit einem normalen
Muskelfaserdurchmesser bzw. Muskelfaserquerschnittsfläche.

Diskussion
215
4 Diskussion
4.1 Ursachen für die Variation in Muskelstoffwechselvorgängen
Glykogen, Glukose, Phosphokreatin (synonym Kreatinphosphat) und
Adenosintriphosphat (ATP) sind wesentliche Energielieferanten des Muskel-
stoffwechsels. Zum Zeitpunkt der Schlachtung beeinflussen das Glykogenniveau sowie
die Geschwindigkeit der Glykolyse post mortem wesentlich die Fleischqualität.
Ergebnisse mehrerer In-vitro-Studien zeigen, dass niedrige Glykogenreserven zum
Zeitpunkt der Schlachtung in schlechter Fleischqualität resultieren (KOLB, 1987,
MÜLLER, 1994, STALDER et al., 1998). In vivo konnten bisher keine signifikanten
Differenzen zwischen verschiedenen Rassen bzw. Linien gefunden werden (LINDNER,
1991).
In Kombination mit neuen DNA-Analyseverfahren eröffnet die Magnetresonanz-
Spektroskopie die Möglichkeit, die Reaktion der Phosphor- und Kohlenstoffbestandteile
des Muskelstoffwechsels auf Stressoren (z.B. Halothan) in vivo und nicht invasiv an
definierten Genotypen zu untersuchen (GAREAU et al, 1993, GEERS et al., 1992 a, b,
JANZEN et al., 1994, MOESGAARD et al., 1995).
Bei der malignen Hyperthermie des Schweins ist seit langem bekannt, dass es sich
um eine genetische Krankheitsdisposition handelt, die eng mit der Ausbildung von PSE-
Fleisch verbunden ist (KOLB, 1987). Der Defekt wird von einem einzigen Genort, dem
Ryanodin-Rezeptor-1-Gen (RyR1) verursacht, welches auf dem Chromosom 6p12-q22
lokalisiert wurde. Synonyme Bezeichnungen für den RyR1-Genort sind „Halothan“-
Genort [HAL1843 TM], „Malignes Hyperthermie Syndrom“-Genort [MHS], „Calcium
Release Channel“-Genort [CRC] bzw. „Porcine Stress Syndrome“-Genort [PSS]
(LISTER, 1987, FUJII et al., 1991, MACLENNAN und PHILLIPS, 1992). Bei der Mutation
des RyR1-Gens handelt es sich um eine Punktmutation am Nukleotid 1843 der porcinen
DNA [ HAL1843 TM]. Dadurch erscheint anstatt einer Cytosin- eine Thymin- Base in
der Nukleotidsequenz. Der Basenaustausch bewirkt bei der Translation, dass anstatt von
Arginin Cystein in dem als „Foot-Region“ bezeichneten Abschnitt des Ryanodin-
Rezeptors eingesetzt wird (FUJI et al, 1991). Bei Anwesenheit des Defektallels am
Ryanodin-Rezeptor-1-Genort ist die Ca2+-Balance des Sarkoplasmatischen Retikulums
von Muskelzellen – vor allem bei Stresssituationen - eingeschränkt (KIM et al., 1984,
NELSON et al., 1991, BRENIG und BREM, 1992, MACLENNAN und PHILLIPS, 1992,
MICKELSON und LOUIS, 1993, 1996, FIEDLER u.a., 2001a).

Diskussion
216
Die Ryanodin-Rezeptoren (RyR 1-3) wurden ursprünglich aufgrund ihrer
Interaktion mit dem Pflanzenalkaloid Ryanodin gefunden und erforscht. Ryanodin ist
ein insektizides Pflanzenalkaloid, das aus den Wurzeln und dem Stamm des
südafrikanischen Strauchgewächses Ryania speciosa gewonnen wird. Es hat zwei aktive
Hauptkomponenten: Ryanodin und 9,21-Didehydroryanodin. Das Alkaloid hält den
Rezeptor permanent offen (KREMPLER, 2000).
Nach Auslösung einer Stressreaktion (z. B. mit Halothan) resultiert die Mutation
am Ryanodin-Rezeptor post mortem in einem schnelleren Phosphokreatinabbau bei
gleichzeitigem Anstieg des anorganischen Phosphats, einem gesteigerten ATP-Umsatz,
einem schneller abfallenden pH-Wert sowie in einer gesteigerten Glykolyserate (KOLB,
1987). Die erhöhte Glykolyserate kann aus dem pH-Verlauf während der 31P-
Spektroskopie abgeleitet werden (MOESGAARD et al., 1995, HENNING et al., 2000).
Halothan bewirkt bei Vorliegen des Defektallels n eine längere und häufigere Öffnung
des RyR1-Kanals im Vergleich zum normalen Genotyp NN (NELSON und LIN, 1995,
MICKELSON und LOUIS, 1996). Die Freisetzungsrate für sarkoplasmatisches Kalzium
(Ca2+) durch den defekten Ryanodin-Rezeptor ist erhöht. In der Folge wird das
Zellinnere mit Ca2+-Ionen überschwemmt. Der erhöhte Kalziumeinstrom in die
Muskelzelle (und der verminderte Kalziumausstrom, FIEDLER u.a., 2001a) regt eine
maximale, ununterbrochene Muskelkontraktion an, die eine allgemeine
Muskelverkrampfung und -steifheit herbeiführt. Kalzium verstärkt durch die
Aktivierung der Phosphorylase-Kinase die Glykogenolyse, die wiederum die katabolen
Aktivitäten wie ATP- und Wärmeproduktion ansteigen lässt. In einer normalen
Muskelzelle wird das Kalzium nach der Kontraktion unter Energieverbrauch in das
sarkoplasmatische Retikulum zurücktransportiert und die Muskelzelle kann auf den
nächsten elektrischen Impuls erneut mit einer Kontraktion reagieren (Abb. 4.1.10). Die
mitochondrialen Enzyme der Atmungskette, Pyruvat-Dehydrogenase, Isozitrat-
Dehydrogenase und 2-Oxoglutarat-Dehydrogenase werden ebenfalls durch Kalzium
aktiviert und tragen zur Erhöhung der Stoffwechselaktivitäten bei (BABCOCK und
HILLE, 1998). Außerdem ist in vitro (post mortem) eine Inaktivierung der Ca2+-ATPase
für die verminderte Aufnahme von Ca2+-Ionen durch das sarkoplasmatische Retikulum
bei den Defektallelträgern verantwortlich (KÜCHENMEISTER et al., 1999, FIEDLER u.a.,
2001a). In vivo scheint die Ca2+-ATPase für die Stoffwechselentgleisungen der
Defektallelträger keine Rolle zu spielen, da die Aktivität der Ca2+-ATPase unmittelbar
nach der Schlachtung in den homozygoten Defektallelträgern höher als in den

Diskussion
217
homozygot „normalen“ Tieren ausfällt (FIEDLER u.a., 2001a). Da der defekte
Kalziumfreigabekanal (calcium release channel - CRC) einen verstärkten Einstrom von
Ca2+-Ionen in die Muskelzelle bedingt, ist die höhere Aktivität der Ca2+-ATPase in den
Defektallelträgern nötig, um die Kalzium-Homöostase so lange wie möglich aufrecht zu
erhalten. Während bei den homozygoten Defektallelträgern der Kalziumfreigabekanal
durchgängig fast vollständig geöffnet wird, ist bei den homozygot „normalen“ Tieren
der CRC nur partiell geöffnet (KÜCHENMEISTER et al., 1999)
Unter extremen Belastungssituationen in vivo übersteigt in den Defektallelträgern
der ATP-Bedarf die Synthese von ATP – die zuerst über Phosphokreatin und aerobe
Glykolyse erfolgt – erheblich, und der Stoffwechsel schaltet um auf die ineffizientere
anaerobe ATP-Gewinnung durch Glykolyse. Als Folgeerscheinungen treten
metabolische Azidose, Anstieg des exspiratorischen CO2 und Sauerstoffdefizit im Blut
auf (MACLENNAN und PHILLIPS, 1992, Abb. 4.1.10). Der Hypermetabolismus, der
generell mit Herz-/Kreislaufsymptomen einhergeht, führt zu Blutdruckanstieg,
Herzrhythmusstörungen und Hyperventilation (KOLB, 1987, HARTUNG, 1995,
MARTENS, 1997, 1998). Im fortgeschrittenen Stadium kommt es zur Schädigung der
Zellmembran, verbunden mit einer zunehmenden Permeabilität für Ionen und Enzyme
sowie anderer Moleküle, die zu Hyperkaliämie, Hyperkalziämie, einem Anstieg der
Kreatinphosphokinase (CK) und Myoglobin im Blut sowie einer schwerwiegenden
Schädigung der Skelett- und Herzmuskulatur führen. Ohne Gegenmaßnahmen
resultieren aus diesen Vorgängen irreversible und letale Zellschäden.
In vivo wurden diese Beobachtungen nach Halothanbelastung bisher nur für die
homozygoten Defektallelträger bestätigt (GEERS et al., 1991, 1992a, DECANNIERE, 1993,
DECANNIERE et al., 1993, GAREAU et al., 1993, JANZEN et al., 1994, KOZAK-RIBBENS et
al., 1997, HENNING et al., 2000, BAULAIN und HENNING, 2001). Muskelsteife als
klinische Beobachtung während des klassischen Halothantestes stand nach GEERS et al.
(1992a) in Beziehung zum Grad des Phosphokreatinabbaus. Während der eigenen
Untersuchungen ließ die Positionierung der Tiere im geschlossenen Magneten keine
derartigen Beobachtungen zu. Die homozygot halothansensitiven Schweine zeigten
auch in vivo einen signifikanten Rückgang in der Phosphokreatinkonzentration,
während das Signal des anorganischen Phosphats breiter wurde, in der Amplitude
zunahm und sich die Position im Spektrum verschob (GEERS et al., 1992a).
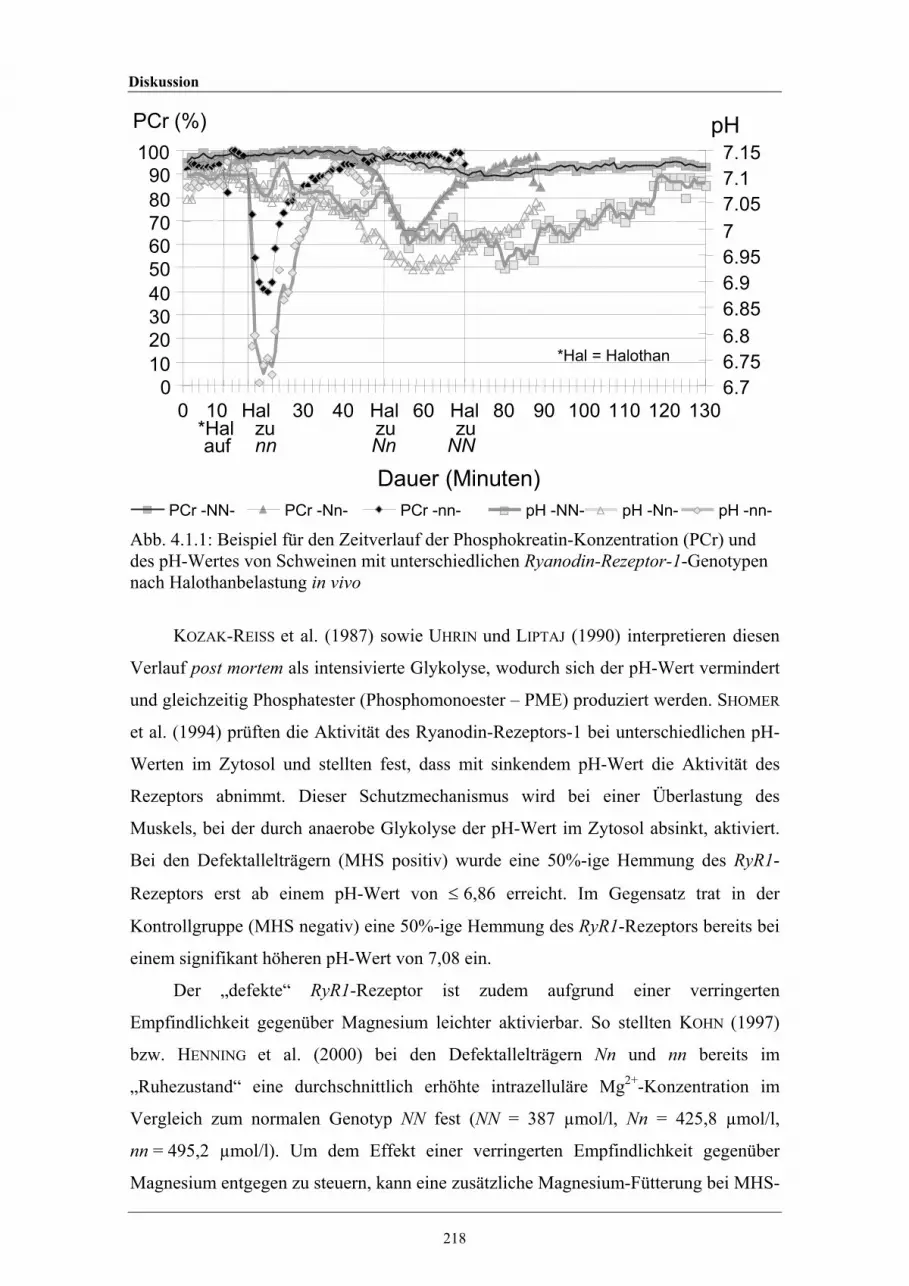
Diskussion
218
Abb. 4.1.1: Beispiel für den Zeitverlauf der Phosphokreatin-Konzentration (PCr) unddes pH-Wertes von Schweinen mit unterschiedlichen Ryanodin-Rezeptor-1-Genotypennach Halothanbelastung in vivo
KOZAK-REISS et al. (1987) sowie UHRIN und LIPTAJ (1990) interpretieren diesen
Verlauf post mortem als intensivierte Glykolyse, wodurch sich der pH-Wert vermindert
und gleichzeitig Phosphatester (Phosphomonoester – PME) produziert werden. SHOMER
et al. (1994) prüften die Aktivität des Ryanodin-Rezeptors-1 bei unterschiedlichen pH-
Werten im Zytosol und stellten fest, dass mit sinkendem pH-Wert die Aktivität des
Rezeptors abnimmt. Dieser Schutzmechanismus wird bei einer Überlastung des
Muskels, bei der durch anaerobe Glykolyse der pH-Wert im Zytosol absinkt, aktiviert.
Bei den Defektallelträgern (MHS positiv) wurde eine 50%-ige Hemmung des RyR1-
Rezeptors erst ab einem pH-Wert von ≤ 6,86 erreicht. Im Gegensatz trat in der
Kontrollgruppe (MHS negativ) eine 50%-ige Hemmung des RyR1-Rezeptors bereits bei
einem signifikant höheren pH-Wert von 7,08 ein.
Der „defekte“ RyR1-Rezeptor ist zudem aufgrund einer verringerten
Empfindlichkeit gegenüber Magnesium leichter aktivierbar. So stellten KOHN (1997)
bzw. HENNING et al. (2000) bei den Defektallelträgern Nn und nn bereits im
„Ruhezustand“ eine durchschnittlich erhöhte intrazelluläre Mg2+-Konzentration im
Vergleich zum normalen Genotyp NN fest (NN = 387 µmol/l, Nn = 425,8 µmol/l,
nn = 495,2 µmol/l). Um dem Effekt einer verringerten Empfindlichkeit gegenüber
Magnesium entgegen zu steuern, kann eine zusätzliche Magnesium-Fütterung bei MHS-
Dauer (Minuten)
PCr (%)
*Hal = Halothan
0102030405060708090
100
0 10*Halauf
Halzunn
30 40 HalzuNn
60 Halzu
NN
80 90 100 110 120 1306.76.756.86.856.96.9577.057.17.15
pH
PCr -NN- PCr -Nn- PCr -nn- pH -NN- pH -Nn- pH -nn-

Diskussion
219
heterozygoten Schweinen im Vergleich zu nicht zusätzlich mit Magnesium versorgten
Tieren einen verlangsamten PCr-Abbau bzw. pH-Abfall und folglich eine verminderte
ATP-Umsatzrate bewirken (MOESGAARD et al., 1993).
Speziell wenn das Defektallel doppelt vorliegt, führen die hohen intrazellulären
Kalziumkonzentrationen nur unzureichend oder gar nicht zu einer Inaktivierung des
RyR1-Rezeptors. Dadurch entstehen vermehrt unkontrollierte Muskelkontraktionen, die
durch die (indirekt) kalziumabhängige Interaktion von Myosin und Aktin unter
Verbrauch von ATP zu erklären sind. ATP wird zusätzlich von der Ca-ATPase des
Sarkoplasmatischen Retikulums verbraucht. Dabei wird es zunächst durch
Phosphokreatin regeneriert, muss aber dann durch aerobe Glykolyse und bei
anhaltender Belastung durch anaerobe Glykolyse neu synthetisiert werden. Bei
übermäßiger Belastung erschöpfen sich die Energiereserven sehr schnell. Laktat wird
angehäuft und infolge des ATP-Mangels kann die Ionenverteilung zwischen intra- und
extrazellulärem Raum (Na+ und K+) nicht mehr aufrechterhalten werden, wodurch
osmotische Effekte ausgelöst werden (ABELES et al., 1992, SHOMER et al., 1994, KOHN,
1997, MARTENS, 1997).
In keiner aus der Literatur bekannten 31P-NMR-Untersuchungen konnte
nachgewiesen werden, dass der heterozygote Genotyp in vivo einen intermediären
Phosphokreatinabbau aufweist (GEERS et al., 1992b, KOHN et al., 1994, 1998, KOHN,
1997, HENNING et al., 2000). Diese Ergebnisse waren hauptsächlich durch die
Versuchsanstellung bedingt, denn die Halothanbelastung wurde über einen kürzeren
Maximalzeitraum (20 min) durchgeführt und/oder zusätzlich wurden geringere
Halothankonzentrationen (2 Volumen-%) verwendet als im eigenen Versuch (60 min,
bzw. 3 Volumen-%).
Die Ergebnisse des eigenen 31P-MR-Spektroskopie-Experimentes zeigen in vivo
erwartungsgemäß, dass die Präsenz beider Defektallele durchgängig eine drastische
Reaktion nach Halothanbelastung hervorruft, während das Vorhandensein nur eines
Defektallels am Ryanodin-Rezeptor-1-Genort nicht zwingenderweise eine extreme
Belastungsreaktion nach sich ziehen muss, aber ebenfalls erwartet werden kann (Abb.
4.1.1, 4.1.10). Insgesamt konnte kein Beweis dafür gefunden werden, dass das mit dem
Ryanodin-Rezeptor-1-Gen übereinstimmende Halothan-Gen in Bezug auf die
Stressreaktion eine homozygot rezessive Genwirkung mit unvollständiger Penetranz
besitzt. Die These der homozygot rezessiven Genwirkung mit unvollständiger Penetranz
trifft allein auf den ungenauen konventionellen Halothanmaskentest zu. Die 31P-MR-

Diskussion
220
Spektroskopie-Untersuchungen von GEERS et al. (1992a) besagen ebenfalls, dass die
Stärke des Phosphokreatinabbaus in vivo innerhalb des M. biceps femoris beim
Vergleich von halothansensitiven und „halothanresistenten“ Tieren keine falsch-
negativen Ergebnisse erbrachte und mit 100%-iger Zuverlässigkeit die homozygot
halothansensitiven Tiere identifiziert. Die eigenen Untersuchungen können diese
Aussage nicht bestätigen, denn es können auch heterozygote Tiere in vivo einen
Phosphokreatinabbau aufweisen, der in der Stärke dem der homozygoten
Defektallelträger gleicht. Abgesehen von den Kosten, wäre die 31P-MR-Spektroskopie
in dieser Hinsicht nicht für die Identifizierung der RyR1-Genotypen geeignet.
Während des konventionellen Halothanbelastungstests werden die Schweine
maximal 5-8 Minuten einer Halothanzufuhr ausgesetzt. Diese Zeit ist nicht zwingend
ausreichend, um in den homozygot „halothansensitiven“ Tieren eine sichtbare Reaktion
zu provozieren, während bei den heterozygoten Tieren dafür in der Regel eine
bedeutend längere Halothanbelastung notwendig war. In der eigenen Untersuchung
dauert es übereinstimmend mit KOLB (1987) zwischen 3 und 19 Minuten nach Beginn
der Halothangabe, um bei den homozygot halothansensitiven Tieren einen signifikanten
Rückgang in der Phosphokreatin- bzw. Glykogenkonzentration als Indikator für
Stoffwechselimbalancen im Muskel feststellen zu können. Vergleichsweise zeigen die
heterozygoten Defektallelträger die größte Variation in ihrer Stoffwechselreaktion auf
eine Halothanbelastung. Drei heterozygote Schweine (von 29) reagierten mit einem
signifikanten Rückgang in der Phosphokreatinkonzentration bereits nach 5 bis 14
Minuten Halothanadministration, während 24 % (7) der heterozygoten Schweine 20 bis
39 Minuten Halothanzufuhr benötigten. In 65 % (19) der untersuchten heterozygoten
Schweine betrug die Dauer der Halothangabe zwischen 52 und 60 Minuten. Insgesamt
zeigten 24 % (7) der heterozygoten Schweine in Bezug auf Phosphokreatin keinen
signifikant veränderten Muskelstoffwechsel nach 60 Minuten Halothangabe genauso
wie 76 % (16) der homozygot „normalen“ Schweine.
Offensichtlich führt die einfache Anwesenheit des Defektallels nicht zwingend zu
nachweisbaren Stoffwechselveränderungen im Vergleich zu den „Normaltieren“, was
die Wechselwirkung mit anderen Genen sehr nahe liegend erscheinen lässt. LE ROY et
al. (2000) unterstützen diese These mit dem Hinweis auf eine Wechselwirkung von
RyR1- und RN-Gen (Hampshireeffekt) auf Fleischqualitätsmerkmale. Träger des RyR1-
Defektallels (n) und des RN --Allels (gekennzeichnet durch einen erhöhten Muskel-
Glykogengehalt) zeigten einen stärker verminderten pH1-Wert (35 Minuten post

Diskussion
221
mortem) oder Dripverlust als Träger des RyR1-Defektallels (n) in Kombination mit dem
„normalen“ rn+-Allel (rn+/rn+, gekennzeichnet durch einen „normalen“ Muskel-
Glykogengehalt). Hingegen ergab sich ein niedrigerer Kochverlust bei Trägern des RN -
-Allels in Kombination mit dem homozygot „defekten“ RyR1-Genotyp (nn) im
Vergleich zur Kombination mit dem homozygot „normalen“ RyR1-Genotyp (NN).
Da kein Beweis für einen homozygot rezessiven Effekt des Defektallels gefunden
werden konnte, ist ein additiver Effekt des Defektallels und die Interaktion mit anderen
Genorten sowohl für den Muskelstoffwechsel als auch für die Körperzusammensetzung
(Kapitel 4.2) wahrscheinlicher. Zusätzlich könnten die monogen und polygen bedingten
Effekte auf den Muskelstoffwechsel durch die Struktur des Ryanodin-Rezeptors
moduliert werden. Speziell wenn nur ein Defektallel (Nn) vorhanden ist, sind
entsprechend der eigenen Ergebnisse vielfältige Reaktionen des Muskelstoffwechsels zu
erwarten. Von einer extrem starken Belastungsreaktion mit einem beschleunigten
Phosphokreatin- und Glykogenabbau bis zu einer sehr geringen Belastungsreaktion
ohne messbare Abbauraten für Phosphokreatin und Glykogen sind alle Zwischenstufen
möglich. FRANZINI-ARMSTRONG und JORGENSEN (1994), ORLOVA et al. (1996) sowie
WAGENKNECHT und RADERMACHER (1997) belegen unter kombinierter Verwendung
von „Immunolabeling“, (Tiefgefrier-)Elektronenmikroskopie und Bildverarbeitung,
dass der Ryanodin-Rezeptor-Kanal die Form eines quadratisch-prismenähnlichen
Tetramers aufweist, welche von MEISSNER (1994) in Form eines vierblättrigen
Kleeblattes mit einer Dimension von 27 x 27 x 14 nm beschrieben wurde. MA und
ZHAO (1994) fanden vier getrennte Leitfähigkeits-Zustände am RyR-Kanal und
bestätigen damit die Tetramerstruktur. Der Rezeptor besteht aus vier Untereinheiten, die
symmetrisch um ein „Fußstück“ („Foot Region“) oder Verbindungsstück (plug)
angeordnet sind (Abb. 4.1.2, 4.1.3) und besitzt eine aus cDNA-Sequenzen geschätzte
Masse von 4 x ~565 kDa = ~2,3 Millionen Dalton (FURUICHI, et al. 1994, MEISSNER,
1994, EHRLICH, 1995, SORRENTINO et al., 2000). Entgegen der ursprünglichen
Homotetramer-Theorie (MARTENS, 1997, 1998) - bestehend aus Verbindungsstück
(„Foot Region“) und vier kanalformenden Untereinheiten - enthält die
Proteingrundstruktur des Ryanodin-Rezeptors-1 als zusätzlichen festen Bestandteil an
jeder Untereinheit ein 12 kDa Protein, das „FK506-Binding Protein“ (FKBP12). Es ist
in die Kommunikation zwischen potentialabhängigem Dihydropyridin-Rezeptor und
Ryanodin-Rezeptor während der Muskelarbeit (excitation-contraction coupling)
eingebunden (EHRLICH, 1995, MICKELSON und LOUIS, 1996, WOLZ, 1999, KREMPLER,

Diskussion
222
2000). Entfernt man FKBP12 vom Ryanodin Rezeptor, wird der Ca2+-Kanal
durchlässig. Offenbar ist FKBP12 an der Koordinierung des RyR1-Tetramers bzw. an
der Stabilisierung der Funktion des RyR1-Kanals beteiligt (MAYRLEITNER et al., 1994,
EHRLICH, 1995).
Nach KREMPLER (2000) entwickelt sich der (elektromechanische) EC-
Kopplungsmechanismus (excitation-contraction coupling) im Skelettmuskel in den
ersten Wochen nach der Geburt. Im Zuge der postnatalen Entwicklung werden Protein-
Protein-Interaktionen innerhalb des Ca2+-Regulationssystems immer komplexer. Die
Bildung von Oligomeren scheint eine wichtige Voraussetzung für die physiologische
Funktion der RyR-Proteine zu sein.
Die elektromechanische Koppelung ist der Prozess, der ein Aktionspotential eines
motorischen Nervs in mechanische Energie in Form einer Gleitbewegung von Aktin-
und Myosinfilamenten "übersetzt". Ein an der motorischen Endplatte einer Muskelfaser
ankommendes Aktionspotential führt dabei zu einer Depolarisierung des Sarkolems, das
sich über die Einstülpungen der „transversalen Tuben“ rasch bis ins Innere der
Muskelfaser ausbreitet. Nach Erfassung der Membrandepolarisierung durch einen
Spannungssensor wird das Signal an das sarkoplasmatische Retikulum (SR)
weitergeleitet, was in der Freisetzung von Ca2+ aus dessen terminalen Zisternen
resultiert. Im Sarkoplasma bindet das Ca2+ an die Troponin-C-Moleküle eines dünnen
Aktinfilaments und löst das eigentliche Filamentgleiten aus (WOLZ, 1999).
Die im sarkoplasmatischen Retikulum verankerte „Foot Region“ dient nicht nur
als Basis für die vier kanalformenden Untereinheiten, die den zytoplasmatischen Teil
des Rezeptors bilden, sondern stellt auch die Verbindung zwischen Lumen des SR und
dem Zytoplasma dar. Die kanalformenden Untereinheiten zeigen in den
elektronenmikroskopischen Tomogrammen eine stark zerklüftete Struktur mit jeweils
drei porenartigen Öffnungen. Für die Tetramerstruktur spricht ebenso, dass die mit dem
Ryanodin-Rezeptor direkt zusammenwirkenden Dihydropyridin-Rezeptoren in
Vierergruppen angeordnet sind (Abb. 4.1.2, 4.1.3), die mit ihrer zweireihigen
Netzstruktur genau der durch die vier Untereinheiten des Ryanodin-Rezeptors
gebildeten Struktur angepasst sind (BRENIG u.a., 1991, ORLOVA et al., 1996,
WAGENKNECHT und RADERMACHER, 1997, RÍOS und STERN, 1997, KREMPLER, 2000,
SORRENTINO et al., 2000). Die phylogenetisch verwandten Inositol-Trisphosphat-
Rezeptoren, die die Inositol-1,4,5-Triphosphat und zum Teil - ebenso wie RyR1-

Diskussion
223
Rezeptoren - die Ca2+-induzierte Ca2+-Freisetzung regulieren, bestehen hingegen aus
sechs „Transmembran“-Untereinheiten (MIKOSHIBA, 1997, SORRENTINO et al., 2000).
Wie beschrieben, kodiert der betroffene RyR1-Genort den Ca2+-Freigabekanal
oder den Ryanodin-Rezeptor-1-Kanal des Sarkoplasmatischen Retikulums der
Skelettmuskulatur. Speziell im Dihydropyridin-Rezeptor zugewandten Teil der „Fuß-
Region“ des Ryanodin-Rezeptor-1-Kanals findet sich bei den Defektallelträgern in der
Position 615 anstelle der normalerweise vorhandenen Aminosäure Arginin die „falsche“
Aminosäure Cystein (MARTENS, 1997). SORRENTINO et al. (2000) beschreiben drei
Haupt-Mutationsbereiche („main hot spots of mutation“) für das RyR1-Gen (des
Menschen). Der erste Bereich liegt zwischen den Aminosäuren 35 und 614, die die
sogenannten MIR-Domänen (Mannosyltransferase, Inositol-(1,4,5)-Trisphosphat-
Rezeptor und Ryanodin-Rezeptor) kodieren. Der zweite Bereich befindet sich zwischen
den Aminosäuren 2162 und 2458 der zweiten RIH-Domäne (Ryanodin-Rezeptor- und
Inositol-(1,4,5)-Trisphosphat-Rezeptor-Homologien) und der dritte Bereich zwischen
den Residuen 4796 und 4898 der Region für die Ionenkanalformation. Insgesamt
wurden beim Menschen inzwischen mehr als 20 Mutationen am RyR1-Gen beschrieben,
die mit dem malignen Hyperthermie-Syndrom in Verbindung gebracht werden (PAUL,
1998, WOLZ, 1999).
Da im heterozygoten Genotyp offenbar beide beim Schwein bekannten Allele des
Ryanodin-Rezeptors-1 zur Ausprägung gelangen, könnte diese Tetramerstruktur aus
allen denkbaren Kombinationen der durch die beiden Allel-Varianten (normal und
defekt) kodierten Ryanodin-Rezeptor-Typen bestehen: alle vier Untereinheiten normal,
alle vier Untereinheiten abnormal (defekt), und eine Mixtur aus normalem und
abnormalem Typ (MICKELSON und LOUIS, 1993, MARTENS, 1997, 1998).
Übereinstimmend mit den eigenen Ergebnissen würde diese Formation in einer großen
Variation von Stressreaktionen zum Ausdruck kommen. Eine genaue Kenntnis der
Struktur des Ryanodin-Rezeptors ist erforderlich, um die Stoffwechselreaktion der
heterozygoten Tiere voraussagen zu können.
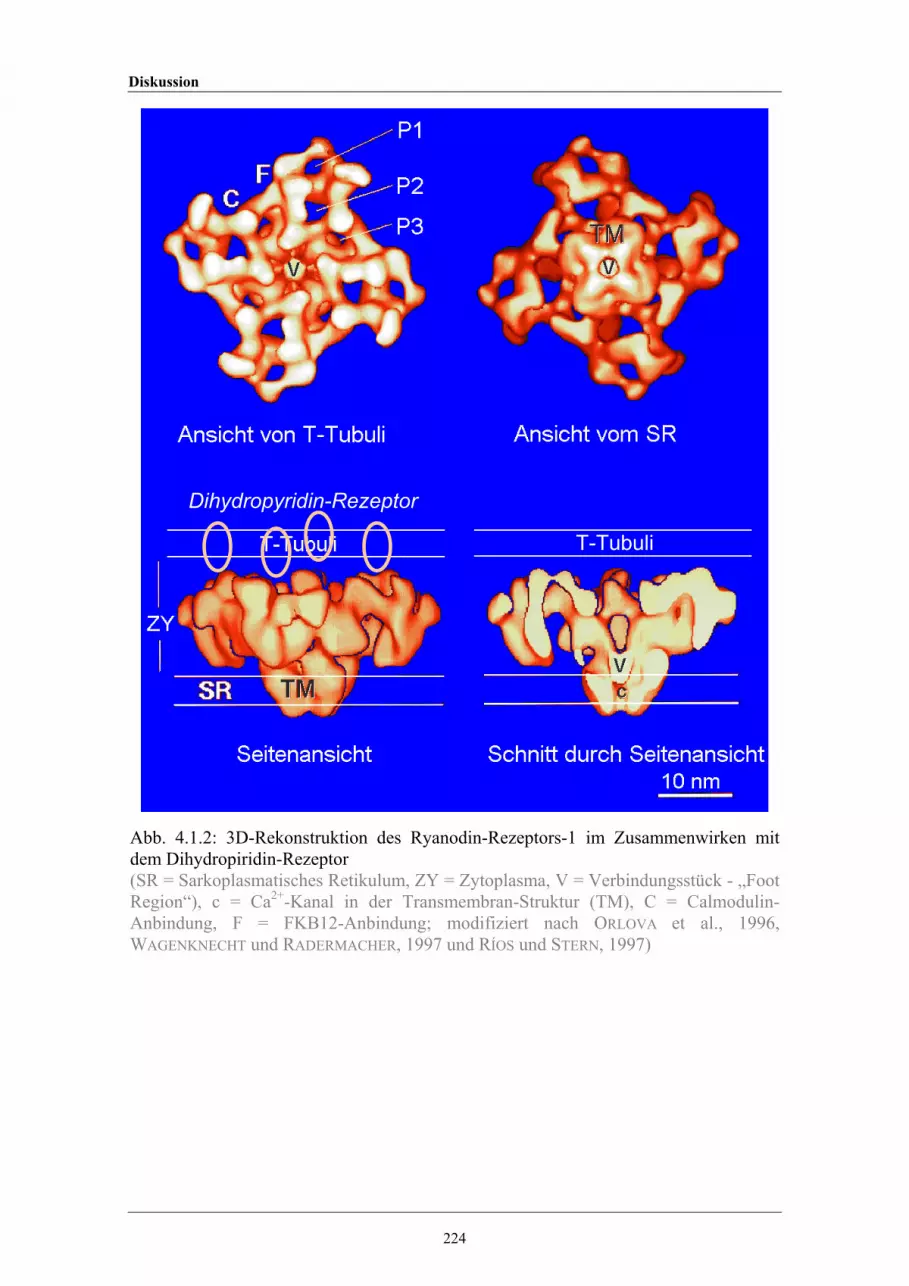
Diskussion
224
Dihydropyridin-Rezeptor
T-Tubuli
Abb. 4.1.2: 3D-Rekonstruktion des Ryanodin-Rezeptors-1 im Zusammenwirken mitdem Dihydropiridin-Rezeptor(SR = Sarkoplasmatisches Retikulum, ZY = Zytoplasma, V = Verbindungsstück - „FootRegion“), c = Ca2+-Kanal in der Transmembran-Struktur (TM), C = Calmodulin-Anbindung, F = FKB12-Anbindung; modifiziert nach ORLOVA et al., 1996,WAGENKNECHT und RADERMACHER, 1997 und RÍOS und STERN, 1997)
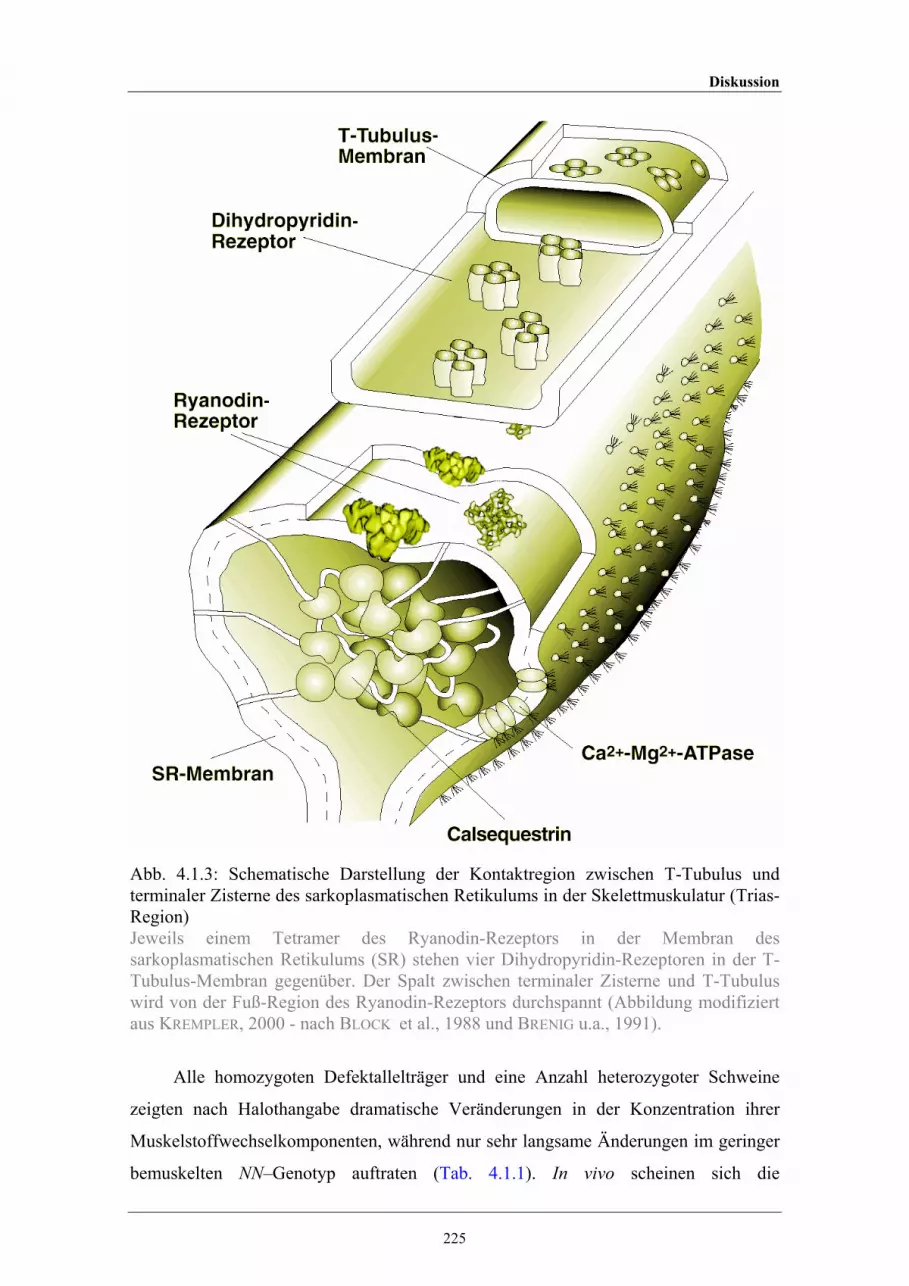
Diskussion
225
Abb. 4.1.3: Schematische Darstellung der Kontaktregion zwischen T-Tubulus undterminaler Zisterne des sarkoplasmatischen Retikulums in der Skelettmuskulatur (Trias-Region)Jeweils einem Tetramer des Ryanodin-Rezeptors in der Membran dessarkoplasmatischen Retikulums (SR) stehen vier Dihydropyridin-Rezeptoren in der T-Tubulus-Membran gegenüber. Der Spalt zwischen terminaler Zisterne und T-Tubuluswird von der Fuß-Region des Ryanodin-Rezeptors durchspannt (Abbildung modifiziertaus KREMPLER, 2000 - nach BLOCK et al., 1988 und BRENIG u.a., 1991).
Alle homozygoten Defektallelträger und eine Anzahl heterozygoter Schweine
zeigten nach Halothangabe dramatische Veränderungen in der Konzentration ihrer
Muskelstoffwechselkomponenten, während nur sehr langsame Änderungen im geringer
bemuskelten NN–Genotyp auftraten (Tab. 4.1.1). In vivo scheinen sich die

Diskussion
226
Muskelstoffwechselprozesse des NN- und des Nn-Genotyps mehr zu gleichen als post
mortem (SCHOLZ et al., 1995).
Generell sind während der 31P-Spektroskopie die Differenzen zwischen allen drei
Genotypen post mortem deutlicher als in vivo, wobei Nn eine intermediäre Stellung
einnimmt. Dieselbe Tendenz ergibt sich für den Glykogenabbau (%) während der 13C-
MR-Studie, obgleich der Nn-Genotyp post mortem sogar eine höhere Abbaurate
aufweist als der nn-Genotyp. Die Kleinsten Quadrate Mittelwerte
(± Standardschätzfehler) betragen: 20,50 ± 12,71 im NN Genotyp, 77,68 ± 7,20 in Nn
und 53,22 ± 7,52 in nn (Modell I).
Ein stärkerer Effekt post mortem im Vergleich zur Reaktion in vivo liegt
möglicherweise neben der myoglobinabhängigen Sauerstoffzufuhr (WITTENBERG und
WITTENBERG, 1989) ursächlich in der über den Blutstrom nicht mehr gegebenen
Nachlieferung von Glukose als insulinabhängigem Energielieferanten der Muskelzelle
(BARON und CLARK, 1997) sowie der Inaktivierung der Ca2+-ATPase (FIEDLER u.a.,
2001a). Die Unterschiede in der Körpertemperaturentwicklung in vivo zu post mortem
fallen entsprechend sehr drastisch aus (Abb. 4.1.5).
Unter In-vivo-„Ruhebedingungen“ unterscheiden sich die RyR1-Genotypen nicht
im Muskel-pH-Niveau (~7,06), jedoch begannen die Versuche beim Nn-Genotyp mit
einer niedrigeren Körpertemperatur als bei den beiden homozygoten Genotypen NN
bzw. nn. Im Gegensatz zu den Ergebnissen von KOHN (1997) weisen die nn-Tiere
bereits zu Versuchsbeginn ein geringeres Phosphorilierungspotential (PCr/Pi-
Verhältnis) auf als NN- bzw. Nn–Tiere, hervorgerufen durch ein geringfügig erhöhtes
Pi-Startniveau. Insgesamt korrespondiert der Verlauf des 13C Experiments mit den
Ergebnissen des 31P Experiments. So verfügen NN- und Nn-Schweine über ein
signifikant höheres „Start“-Glykogenniveau als nn-Schweine (Tab. 4.1.1, Abb. 4.1.4).
In Biopsie-Proben fand LINDNER (1991) hingegen keine signifikanten Unterschiede im
„Ruhe“-Glykogenniveau (in vivo) zwischen stressstabilen und stressanfälligen
Schweinen. Ursache für die unterschiedliche Start-Konzentration von anorganischem
Phosphat und Glykogen könnten die teilweise während der Sedierung aufgetretenen
verhaltensbedingten Stressreaktionen gewesen sein.

Diskussion
227
Tab. 4.1.1: Stoffwechselreaktion und Körperzusammensetzung in Schweinen
unterschiedlicher RyR1-Genoytpen mit einer Lebendmasse von ca. 10 kg (LSM ± SEE
nach Modell I#)
NN Nn nn
PCr/Pi-Verhältnis Start 15,53 ± 0,69 a 15,21 ± 0,52 a 13,35 ± 0,79 b
PCr-Abbau (%) 32,11 ± 5,27 a 44,01 ± 4,17 b 72,12 ± 5,63 c
Pi-Anstieg (%) 394 ± 39 a 355 ± 33 ab 291 ± 43 b
pH-Minimum 6,908 ± 0,041 a 6,769 ± 0,035 b
6,563 ± 0,046 c
ATP-Änderung (%) +4,02 ± 4,24 a -10,53 ± 3,63 b -37,74 ± 4,73 c
N (n - post mortem) 24 (3) 29 (4) 16 (3)
Glykogen Start (µmol/g) 74,12 ± 5,91 a 76,11 ± 5,15 a 57,12 ± 4,35 b
Glykogenabbau (%) 20,88 ± 8,45 a 56,71 ± 5,11 b 46,76 ± 3,51 b
N (n - post mortem) 7 (1) 10 (2) 10 (3)
Körpertemperatur Start (°C) 39,38 ± 0,14 a 38,96 ± 0,13 b 39,25 ± 0,15 ab
Körpertemperatur-Änderung (°C) -0,71 ± 0,34 a +1,70 ± 0,29 b +2,82 ± 0,33 c
Halothangabe (Minuten) 44,16 ± 2,90 a 38,43 ± 2,59 a 6,57 ± 3,02 b
N 13C+31P (n - post mortem) 31 (4) 39 (6) 26 (6)
Volumen Mm.l.d. (cm³) 40,78 ± 1,64 a 41,24 ± 1,53a 46,53 ± 1,91c
Fett-Volumen (cm³) 13,19 ± 0,69 12,08 ± 0,65 13,58 ± 0,82
N 31 42 23
#) Werte mit unterschiedlichen Buchstaben sind signifikant verschieden (p ≤ 0,05).
Berücksichtigt man sowohl die Untersuchungen in vivo als auch post mortem, dann
ergeben sich für die drei Ryanodin-Rezeptor-1-Genotypen folgende Glykogen-
abbauraten:
15,47 µmol/g in durchschnittlich 135 Minuten bei NN, entspricht 0,11 µmol/g • min-1;
43,16 µmol/g in durchschnittlich 130 Minuten bei Nn, entspricht 0,33 µmol/g • min-1
und
26,71 µmol/g in durchschnittlich 100 Minuten bei nn, entspricht 0,27 µmol/g • min-1.
Da die Halothanbelastung von nn mit 6,57 Minuten zu 38,43 Minuten für Nn signifikant
geringer ausfiel, sind die Glykogenabbauraten der beiden Genotypen nur bedingt
vergleichbar (Abb. 4.1.4).
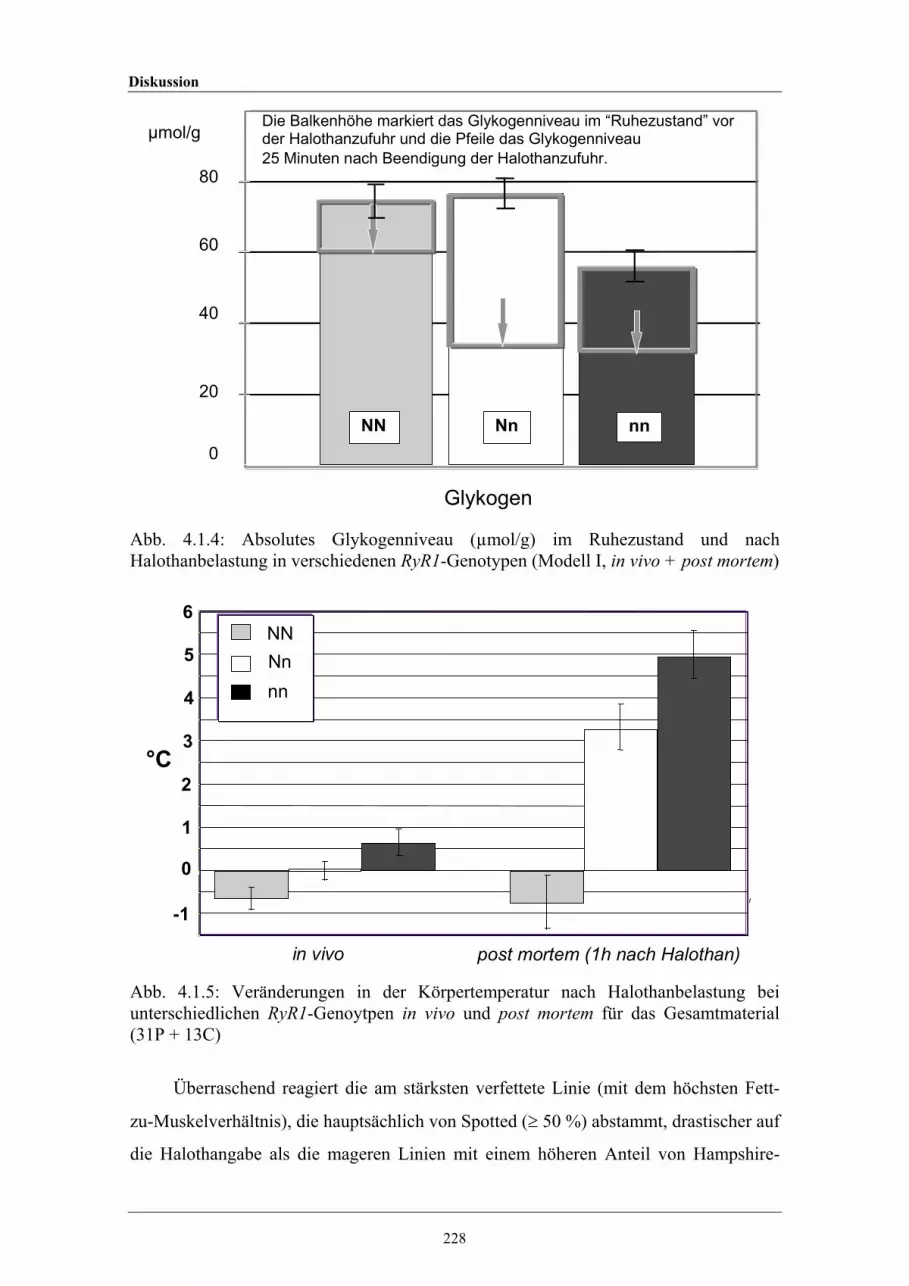
Diskussion
228
µmol/g
0
20
40
60
80
Die Balkenhöhe markiert das Glykogenniveau im “Ruhezustand” vorder Halothanzufuhr und die Pfeile das Glykogenniveau25 Minuten nach Beendigung der Halothanzufuhr.
NN
Glykogen
Nn nn
Abb. 4.1.4: Absolutes Glykogenniveau (µmol/g) im Ruhezustand und nachHalothanbelastung in verschiedenen RyR1-Genotypen (Modell I, in vivo + post mortem)
-1-1
00112233445566
°C°C
in vivoin vivo post mortem (1h nach Halothan)post mortem (1h nach Halothan)
NNNN
NnNn
nnnn
Abb. 4.1.5: Veränderungen in der Körpertemperatur nach Halothanbelastung beiunterschiedlichen RyR1-Genoytpen in vivo und post mortem für das Gesamtmaterial(31P + 13C)
Überraschend reagiert die am stärksten verfettete Linie (mit dem höchsten Fett-
zu-Muskelverhältnis), die hauptsächlich von Spotted (≥ 50 %) abstammt, drastischer auf
die Halothangabe als die mageren Linien mit einem höheren Anteil von Hampshire-

Diskussion
229
(Effekt des RN--Allels !), Poland China-, Landrasse- oder Duroc- Genanteilen (Tab.
4.1.2, Abb. 4.1.6).
Tab. 4.1.2: Stoffwechselreaktion und Körperzusammensetzung in verschiedenen
Zuchtlinien (Modell I, LSM ± SEE#)
> 50 %
Duroc
(Du)
≥ 50 %
Hampshire
(Ha)
≥ 50 %
Spotted
(S)
US-
Landrasse
(LR)
Poland
China
(PCh) x LR
DHLSY♦ Du x
Minzhu
PCr-zu-Pi-
Verhältnis♥15,50
± 0,49 a15,53
± 0,87 a16,71
± 1,24 a10,78
± 0,90 b14,95
± 1,00 a- -
PCr-
Abbau (%)
51,07
± 3,96 ac
38,94
± 6,28 b66,90
± 8,91 c45,74
± 6,35 ab
44,41
± 7,18 ab
- -
Pi-
Anstieg (%)
323
± 26 a352
± 42 a510
± 59 b275
± 43 a273
± 48 a- -
pH-
Start♥7,05
± 0,01 a7,05
± 0,01 a7,09
± 0,02 b7,04
± 0,01 a7,08
± 0,02 ab
- -
pH-
Minimum
6,76
± 0,03 ab
6,71
± 0,04 a6,71
± 0,06 ab
6,74
± 0,04 a6,83
± 0,05 b- -
ATP-
Abbau (%)
11,59
± 2,86 a10,86
± 4,58 a35,64
± 6,46 b7,28
± 4,65 a8,39
± 5,26 a- -
n (31P-MRS) 29 12 5 14 9 - -
Fett-zu-Mus-
kel-Verhältnis
0,383
± 0,015 a0,318
± 0,023 bc
0,462
± 0,038 a0,322
± 0,035 bc
0,353
± 0,020 b0,254
± 0,037 c0,454
± 0,033 a
n (1H-MRT) 39 17 6 8 27 6 8
Glykogen♥
(µmol/g)
66,48
± 5,05
69,72
± 5,15
72,63
± 10,43
- - 67,63
± 5,57
Glykogen-
abbau (%)
40,31
± 4,73 b27,28
± 8,87 a55,74
± 12,58 b- - 42,48
± 4,56 b
n (13C-MRS) 9 7 2 - - 9
#) Werte mit unterschiedlichen Buchstaben sind signifikant verschieden (p ≤ 0,05).♥ ) im Ruhezustand♦) 25 % Du, 25 % Ha, 25 % LR, 12,5 % S, 12,5 % Yorkshire
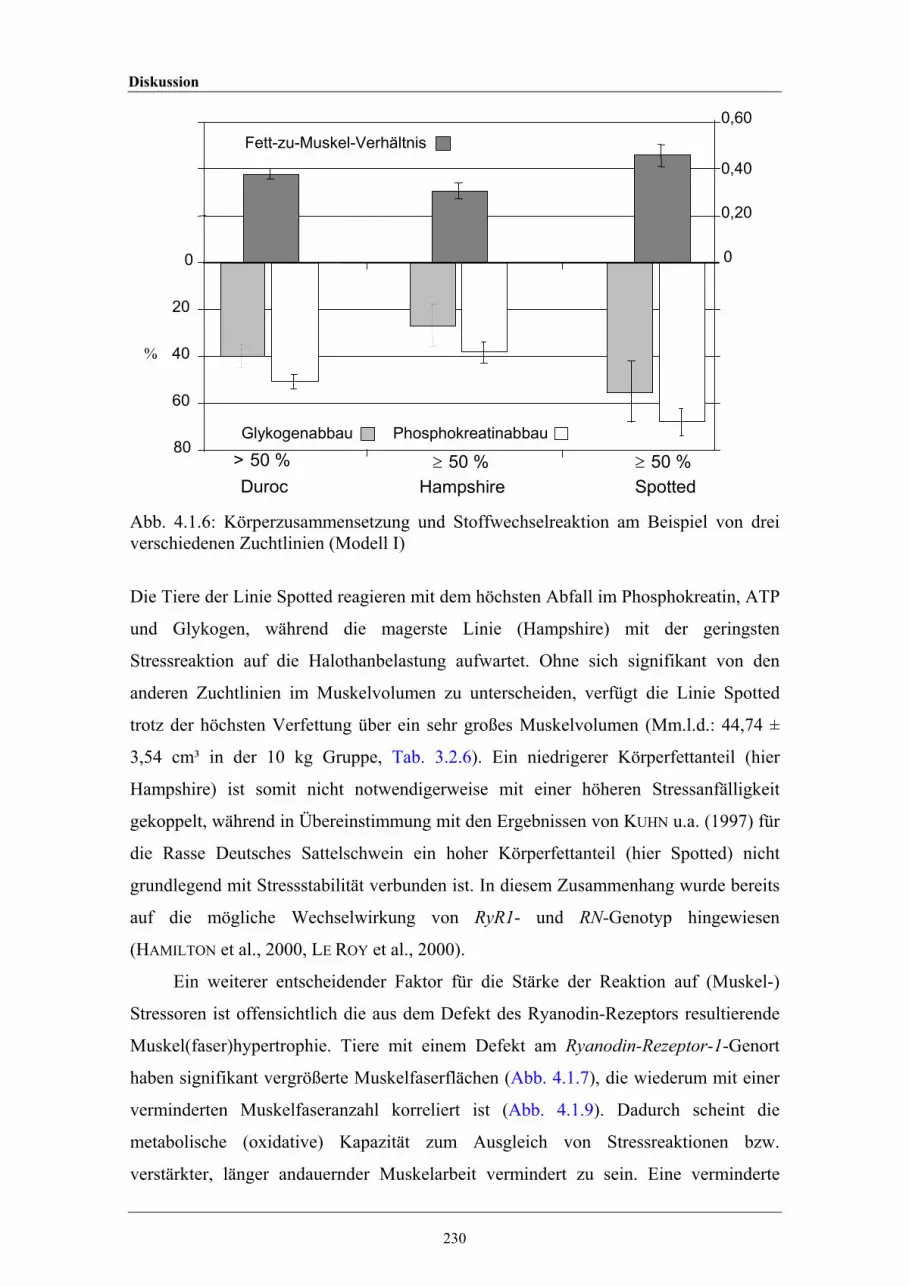
Diskussion
230
80
60
40
20
0
0,20
0,40
0,60
0
Fett-zu-Muskel-Verhältnis
Glykogenabbau Phosphokreatinabbau
%
> 50 %Duroc
≥ 50 %Hampshire
≥ 50 %Spotted
Abb. 4.1.6: Körperzusammensetzung und Stoffwechselreaktion am Beispiel von dreiverschiedenen Zuchtlinien (Modell I)
Die Tiere der Linie Spotted reagieren mit dem höchsten Abfall im Phosphokreatin, ATP
und Glykogen, während die magerste Linie (Hampshire) mit der geringsten
Stressreaktion auf die Halothanbelastung aufwartet. Ohne sich signifikant von den
anderen Zuchtlinien im Muskelvolumen zu unterscheiden, verfügt die Linie Spotted
trotz der höchsten Verfettung über ein sehr großes Muskelvolumen (Mm.l.d.: 44,74 ±
3,54 cm³ in der 10 kg Gruppe, Tab. 3.2.6). Ein niedrigerer Körperfettanteil (hier
Hampshire) ist somit nicht notwendigerweise mit einer höheren Stressanfälligkeit
gekoppelt, während in Übereinstimmung mit den Ergebnissen von KUHN u.a. (1997) für
die Rasse Deutsches Sattelschwein ein hoher Körperfettanteil (hier Spotted) nicht
grundlegend mit Stressstabilität verbunden ist. In diesem Zusammenhang wurde bereits
auf die mögliche Wechselwirkung von RyR1- und RN-Genotyp hingewiesen
(HAMILTON et al., 2000, LE ROY et al., 2000).
Ein weiterer entscheidender Faktor für die Stärke der Reaktion auf (Muskel-)
Stressoren ist offensichtlich die aus dem Defekt des Ryanodin-Rezeptors resultierende
Muskel(faser)hypertrophie. Tiere mit einem Defekt am Ryanodin-Rezeptor-1-Genort
haben signifikant vergrößerte Muskelfaserflächen (Abb. 4.1.7), die wiederum mit einer
verminderten Muskelfaseranzahl korreliert ist (Abb. 4.1.9). Dadurch scheint die
metabolische (oxidative) Kapazität zum Ausgleich von Stressreaktionen bzw.
verstärkter, länger andauernder Muskelarbeit vermindert zu sein. Eine verminderte

Diskussion
231
Anzahl Muskelfasern im Muskel bewirkt - neben einer reduzierten
Gesamtkapillarisierung - automatisch eine verminderte Zahl an Mitochondrien, die
gehäuft in den „langsamen“ Typ-I–Fasern vorkommen. Parallel verfügen speziell die
homozygoten, zum Teil aber auch die heterozygoten Defektallelträger in der Tendenz
über einen höheren Anteil glykolytischer, „schneller“ Fasern vom Typ IIb (ESSÉN-
GUSTAVSSON, 1993, 1995, VON LENGERKEN et al., 1994, 1997, KUHN u.a., 1998,
KARLSSON et al., 1999, FIEDLER u.a., 2001a). Die Genotypen (Nn und nn) mit einer
verminderten Muskelfaseranzahl bei gleichzeitig größeren Muskelfasern weisen zum
Zeitpunkt der Muskelstoffwechseluntersuchung einen stärkeren Abfall im
Phosphokreatin auf (Abb. 4.1.7, 4.1.8, 4.1.9). Da die Muskelfaseruntersuchungen im
eigenen Material nicht parallel mit der Muskelspektroskopie, sondern erst zu einem viel
späteren Zeitpunkt erfolgte, kann nicht exakt geklärt werden, ob bereits zum Zeitpunkt
der Muskelstoffwechseluntersuchung bei ca. 10 kg Lebendmasse Unterschiede in den
Muskelfaserflächen zwischen den drei RyR1-Genotypen bestanden. Diese sind jedoch
anzunehmen, da die Muskelfaseranzahl ab der Geburt konstant bleibt und bereits bei
den ca. 10 kg schweren Schweinen ein vergrößertes Muskelvolumen in den
Defektallelgenotypen auftritt (Tab. 4.1.1). Der heterozygote Genotyp weist zum
Zeitpunkt der Muskelfaseruntersuchung zwar eine minimal kleinere Kotelettfläche als
der homozygot „normale“ Genotyp auf, ist aber mit signifikant vergrößerten
Muskelfaserflächen ausgestattet (Abb. 4.1.4). Außerdem ist bei der Interpretation der
Ergebnisse – speziell für die Beziehung zwischen Muskelfaserstruktur- und
Muskelstoffwechselparametern - zu berücksichtigen, dass die 31P-MR-Spektroskopie
am (äußeren) M. biceps femoris erfolgte, die Biopsie jedoch am M. longissimus dorsi
durchgeführt wurde. Beide Muskeln (äußerer M. biceps femoris und der komplette M.
longissimus dorsi) gehören jedoch - im Gegensatz zum oxidativen Muskelstoff-
wechseltyp - mehr dem glykolytischen Muskelstoffwechseltyp an. Dabei weist der M.
biceps femoris insgesamt (äußerer + innerer M. biceps femoris) einen etwas höheren
Anteil Typ I-Muskelfasern auf als der M. longissimus dorsi (SALOMON u.a., 1981, 1983,
ESSEN-GUSTAVSSON, 1993, 1995; KARLSSON et al., 1993, RUUSUNEN und POULANNE,
1997, KARLSSON et al., 1999, BROCKS et al., 2000, FIEDLER u.a., 2001a).
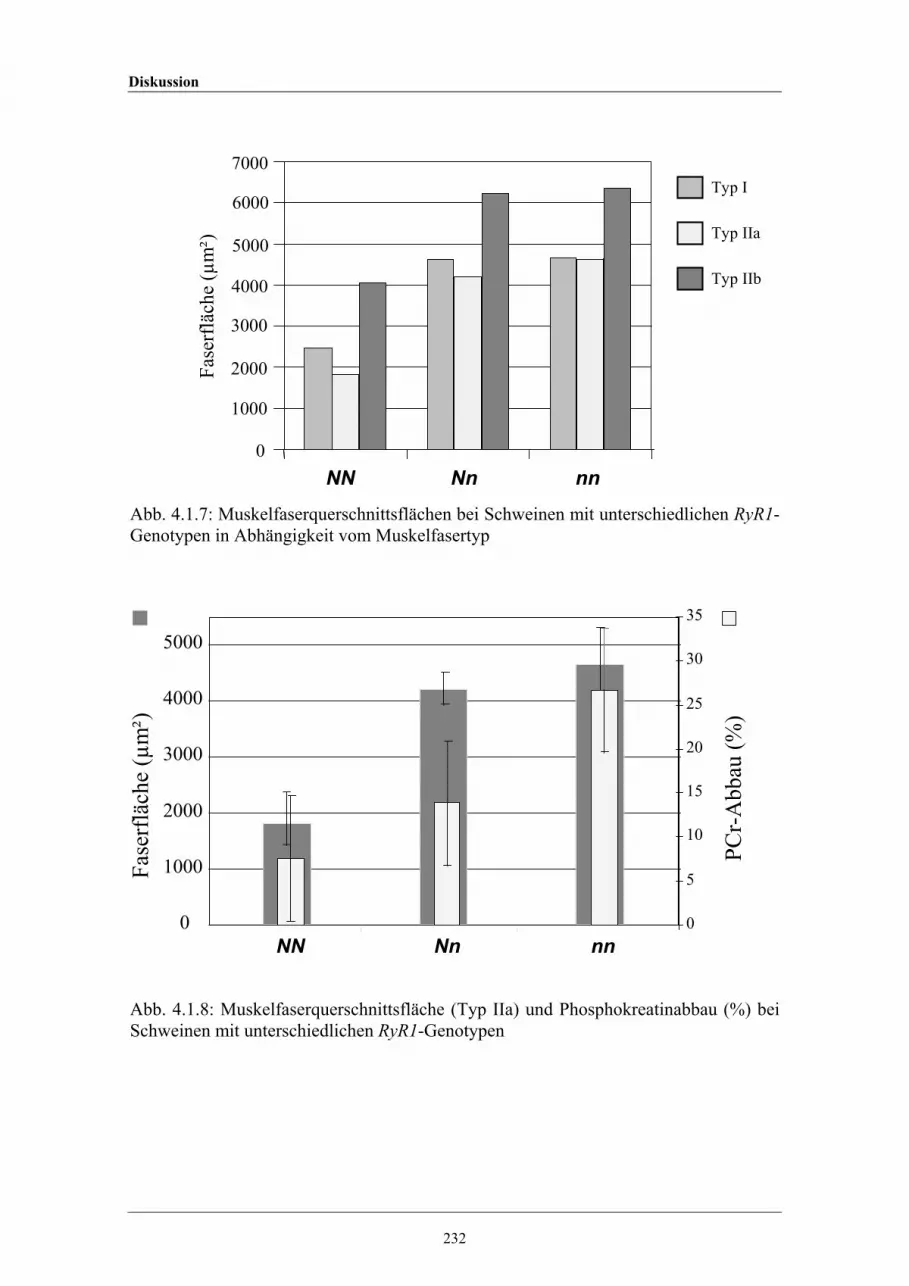
Diskussion
232
Abb. 4.1.7: Muskelfaserquerschnittsflächen bei Schweinen mit unterschiedlichen RyR1-Genotypen in Abhängigkeit vom Muskelfasertyp
Abb. 4.1.8: Muskelfaserquerschnittsfläche (Typ IIa) und Phosphokreatinabbau (%) beiSchweinen mit unterschiedlichen RyR1-Genotypen
0
1000
2000
3000
4000
5000
6000
7000
Fase
rflä
che
(µm
²)Typ I
Typ IIa
Typ IIb
NN Nn nn
0
1000
2000
3000
4000
5000
NN Nn nn0
5
10
15
20
25
30
35
Fase
rflä
che
(µm
²)
PCr-
Abb
au (%
)

Diskussion
233
Abb. 4.1.9: Muskelfaseranzahl und Phosphokreatinabbau (%) bei Schweinen mitunterschiedlichen RyR1-Genotypen
Der RyR1-Genotyp ist nach eigenen Berechnungen zu etwa 20 % für die Variation
in der Stressreaktion (PCr-Abbaurate) bei einer Lebendmasse von ca. 10 kg
verantwortlich. Er erklärt jedoch mit 3 - 17 % (10 - 90 kg Lebendmasse) nur einen
relativ geringen Anteil der Variation in den Merkmalen der Körperzusammensetzung,
so dass ausreichend Variation für weitere Selektionsmaßnahmen innerhalb des
homozygot stressstabilen Genotyps sowie Umweltverbesserungen verbleibt (WEBB et
al., 1994). Die signifikanten Unterschiede in der Körperzusammensetzung (aus der MR-
Tomographie) bei 10 kg Lebendmasse zwischen nn und Nn bzw. NN ergaben sich
unerwartet, da davon ausgegangen wurde, dass sich die stärkere Bemuskelung der
Defektallelträger erst später während des Wachstums entwickeln würde. Obgleich keine
Differenzen in der subkutanen Fettauflage auftraten, hatten die homozygot
stressanfälligen Tiere ein größeres Volumen der Mm. longissimi dorsi (Tab. 4.1.1).
Offenbar reicht das durchschnittliche Alter von 53 Tagen bei einem durchschnittlichen
Gewicht von 12,5 kg bereits aus, um ein vergrößertes Muskelwachstum infolge der
Mutation am RyR1-Genort mit Hilfe der MR-Bildgebung feststellen zu können. Auf
weitere genetische Faktoren, die die Körperzusammensetzung regulieren, wird
umfassend im nächsten Kapitel 4.2 eingegangen.
Es ist sehr plausibel, dass die Wirkung des Ca2+-steuernden /gesteuerten
Ryanodin-Rezeptor-1-Gens neben dem RN-Gen (LE ROY et al., 2000) durch weitere
0100000200000300000400000500000600000700000800000900000
10000001100000
NN Nn nn051015202530354045
Fase
ranz
ahl
PCr-
Abb
au (%
)

Diskussion
234
Gene, die den Energiestoffwechsel bzw. die Zellzyklus-Regulation beeinflussen,
abgeschwächt oder auch verstärkt werden kann, wie z.B. durch einen
zellproliferationsabhängigen Transkriptionsfaktor, das Protoonkogen c-Myc (REINER et
al., 1999). Die Expression des Protoonkogens c-Myc ermöglicht der Zelle, die
Zellteilung (Mitose) einzuleiten und aus der G0-Phase („Gap“-Phase 0 oder Ruhephase
0) in die S-Phase (durch DNA-Synthese charakterisierte Intermitosephase) der
Zellteilung zu gelangen, um diese anschließend zu durchlaufen. In der ruhenden Zelle
ist dieser Vorgang stark eingeschränkt (JENTZSCH, 1983, SCHMOETZER, 1998).
Außerdem wird die Expression von c-Myc durch Peroxisomen-Proliferatoren angeregt,
die wiederum eine wichtige Rolle in der Aufrechterhaltung der Lipid-Homöostase
spielen, da Fettsäuren als Peroxisomen-Proliferatoren wirken. Peroxisomen sind
vorzugsweise auf oxidative Reaktionen mit molekularem Sauerstoff spezialisiert, das
heißt, diese Organellen beherbergen insbesondere Oxidasen des Cytochrom-P450-
Systems. Darüber hinaus kommen in ihnen u.a. Dehydrogenasen, Katalasen sowie
Acyltransferasen vor. Diese Enzyme spielen eine Rolle im Stoffwechsel zahlreicher
körpereigener und körperfremder Substanzen (FELDMANN u.a., 2000).
Parallel ist eine indirekte Beteiligung der mitochondrialen DNA (LIMPER u.a.,
1991, BABCOCK und HILLE, 1998) sowie eine Fehlfunktion der Ca2+-ATPase (FIEDLER
u.a., 2001a) an den Auswirkungen der RyR1-Mutation anzunehmen.
Die vielfältigen Möglichkeiten von Interaktionen zwischen mehreren
Genen/Allelen und deren Aufklärung beschreiben BRENT und FINLEY (1997). Darunter
fallen die Interaktionen zwischen Ryanodin-Rezeptor-1 und FKBP12 sowie FHL1, die
ebenfalls zur Variation der Stoffwechselreaktionen beitragen können. FHL1 („Four und
a half LIM Domain Protein“) wird als ein Promotorfragment des RyR1 beschrieben und
ist - neben der MyoD-Genfamilie sowie einer Reihe von sogenannten Zinkfinger-
Protein-Genen - ein Regulator der Muskelzellfunktionen (KREMPLER, 2000, KREMPLER
et al., 2000). Ein weiterer Kandidat ist Triadin, welches wahrscheinlich in Kontakt zum
Ryanodin-Rezeptor-1 und Calsequestrin steht und an der Initiierung der
Muskelkontraktion mitwirkt, indem Kalzium aus dem sarkoplasmatischen Retikulum in
das Sarkoplasma freigesetzt wird. Calsequestrin dient als Speicher für Kalzium
(MICKELSON und LOUIS, 1996, WOLZ, 1999, KREMPLER, 2000).
HOFFMAN (1995) beschreibt eine Reihe von Mutationen an Genorten, die Defekte
an den potentialgesteuerten Ionenkanälen für Na+/ K+, Cl- und Ca2+ in der
Muskelfasermembran verursachen können und zu verschiedenen Krankheitsbildern

Diskussion
235
führen. Die hyperkalemische periodische Paralyse sowohl beim „Quarter Horse“ als
auch beim Menschen wird durch eine Punktmutation des Skelettmuskel-Na+-Kanal-
Gens hervorgerufen, welches sich auf Chromosom 17 des Menschen befindet.
Für den gesamten Komplex Stressreaktion sind Effekte verantwortlich, die neben
der genetischen Komponente endokrinologisch zu erklären sind und zum Teil mit einem
erhöhten Adrenalinausstoß einhergehen können, gefolgt von einem verstärkten
Gesamtenergieverbrauch. Zur besseren begrifflichen Differenzierung zwischen
belastender (z.B. emotionsauslösender) Reizkonstellation und Belastungsreaktion
bezeichnen MANTEUFFEL und PUPPE (2000) sowie VON BORELL (2000) die
entsprechende Reizkonstellation als Stress und die im Einzelnen analysierbaren
belastenden Reizaspekte als „Stressoren“. Stress bezieht sich also auf objektive
Reizeigenschaften einer Situation, die in einem Individuum eine entsprechende
Reaktion (Angst, Flucht, Angriff) auslöst. Nach MANTEUFFEL und PUPPE (2000),
TUCHSCHERER und MANTEUFFEL (2000) sowie VON BORELL (2000) tritt eine
Stresssituation ein, wenn Abweichungen von einem angestrebten biologischen bzw.
„psycho-physiologischen“ Zustand (Wert) eliminiert werden müssen, um den optimalen
Funktionsspielraum des Organismus beizubehalten (Prinzip der Homöostase =
Aufrechterhaltung der Körperfunktionen).
In diesem Zusammenhang verfügten am RyR1-Genort heterozygote Eber im
Vergleich zu homozygot „normalen“ Ebern zwar über eine signifikant niedrigere Basal-
Plasmakonzentration für ACTH und Kortisol, zeigten aber keine Unterschiede in der
Reaktion nach Stressinduzierung durch Nasenschlingenfixierung über einen Zeitraum
von 5 Minuten bzw. durch die Gabe von Dexamethason (0,04 mg/kg Körpergewicht)
(WEAVER et al., 2000). Auch diese Beobachtungen bestätigen die eigenen Resultate.
Eine relativ kurzzeitige oder niedrig dosierte Exposition zu Stressoren bewirkt bei
heterozygoten Tieren keine nachweisbare Stressreaktion. Langfristig (über eine
komplette Mastperiode hinweg) zeigen die heterozygoten Tiere jedoch im Durchschnitt
ein negativen Effekt auf die Fleischqualität (z.B. WOLF-SCHWERIN und KALLWEIT,
1991, SCHMITTEN, 1993, SCHOLZ und HARDGE, 1994, LOUIS et al. 1994, GOODWIN et al.
1994, HANSET et al., 1995, DE SMET et al., 1996, KUHN u.a., 1997, 1998, ESTANY et al.,
1998, BEČKOVÁ und DAVID, 2000, HENNING et al., 2000, KORTZ et al., 2000, WEAVER
et al., 2000, THALLER et al., 2000). In Übereinstimmung mit VON LENGERKEN u.a.
(1997), MARTENS (1997, 1998), WITTMANN u.a. (1998), BIEDERMANN u.a. (2000) sowie
KRIETER und THOLEN (2001) sollte folglich zur Vermeidung unnötiger Belastungen für

Diskussion
236
das Tier das Defektallel (n) am RyR1-Genort komplett aus allen Zuchtpopulationen -
sowohl aus den Mutterrassen als auch aus den Vaterrassen - eliminiert werden. Die
Stressstabilität und die mit ihr korrelierte Fleischqualität verbessern sich durch die
Elimination des Defektallels (n) „automatisch“. Bei der weiteren Zuchtarbeit ist jedoch
zu berücksichtigen, dass sich durch die Elimination des Defektallels (n) der Marktwert
bestimmende Fleischanteil am Schlachtkörper erheblich (um 1 - 7 %) reduzieren kann
(GÖDEKE u.a., 1998, BIEDERMANN u.a., 2000, IRGANG, 2001, siehe Kapitel 4.2).
Unabhängig von der Anwesenheit des Defektallels (n) am RyR1-Genort existieren
außerdem individuell stark variierende Stressreaktionen auf diverse multifaktorielle
Umweltreize wie Temperatur, Bewegungsspielraum, Liegefläche und soziale
Umgebung (Gruppengröße, Umstallungen, Gruppenneuzusammenstellung), die sich
additiv auf das Schwein auswirken können (CLAUS, 1996, HYUN et al., 1998,
TUCHSCHERER und MANTEUFFEL, 2000, VON BORELL, 2000). Pränatal erfahrener Stress
spielt bei der Stressbewältigung im Laufe des Lebens (Wachstums) eine prägende Rolle
und wirkt sich störend auf die Ontogenese der neuroendokrinen Systeme aus (OTTEN et
al., 2000).

„def
ekte
r“ M
uske
l (R
yR1=
Nn/nn
)„n
orm
aler
“ M
uske
l (R
yR1=NN/ N
n)
Mus
kelfa
ser
Sark
opla
smat
isch
es R
etik
ulum
Gly
koge
n
Glu
kose
-1-P
Lakt
at
Wär
me
Mito
chon
drie
n
Sark
opla
smat
isch
es R
etik
ulum
Mus
kelfa
ser
Gly
koge
n
Glu
kose
-1-P
Lakt
at
Mito
chon
drie
n
AT
P
Abb
. 4.1
.10:
Sch
emat
isch
er V
ergl
eich
der
Mus
kelk
ontra
ktio
nszy
klen
von
Sch
wei
nen
mit
„nor
mal
er“
Mus
kula
tur (
links
) und
„de
fekt
er“
Mus
kula
tur
(rec
hts)
; mod
ifizi
ert n
ach
MA
CLE
NN
AN
und
PH
ILLI
PS (1
992)
237
Diskussion

Diskussion
238
4.2 Genetische Steuerung des Körperfettgehaltes und des Wachstums beimSchwein - Konsequenzen für die Zuchtarbeit
In Übereinstimmung mit den eigenen Ergebnissen (Abb. 4.2.1, Kapitel 3.2.2,
Kapitel 4.1) vermindert bereits eine einfache Mutation am Ryanodin-Rezeptor-1-Genort
(RyR1) neben der Belastungsstabilität und Fleischqualität in den meisten Populationen
den Fettansatz und erhöht das Muskelvolumen bzw. die Muskelfläche (WOLF-
SCHWERIN und KALLWEIT, 1991, SCHMITTEN, 1993, SCHOLZ und HARDGE, 1994,
SCHOLZ et al. 1995, LOUIS et al. 1994, GOODWIN et al. 1994, HANSET et al., 1995, DE
SMET et al., 1996, MITCHELL und SCHOLZ, 1997 a,b, ESTANY et al., 1998, WEAVER et
al., 2000, THALLER et al., 2000, KRIETER und THOLEN, 2001). In einigen Populationen
wurde jedoch keine eindeutige Verminderung des Fettansatzes durch die Wirkung des
Defektallels festgestellt (PARK et al., 1998). Generell können Genotyp-Umwelt-
Interaktionen bzw. Genotyp-Genotyp-Interaktionen (BOUCHARD und TREMBLAY, 1997,
DE VRIES und PLASTOW, 1998, SPURLOCK et al., 1998) oder Genotyp-Geschlecht-
Interaktionen (PARK et al., 1998) die Genwirkung bzw. Genexpression beeinflussen
(MACKAY, 2001). Da die Körperzusammensetzung des Schweines (neben
Belastungsstabilität und damit korrelierter Fleischqualität, siehe Kapitel 4.1) nicht allein
vom Majorgen RyR1, sondern polygen gesteuert wird, ist es für den Tierzüchter von
besonderem Interesse die genetischen Zusammenhänge umfassend aufzudecken.
0
100
200
300
400
500
600
700
800
900
1000
1100
Volu
men
(cm
3 )
Mm.l.d.10 kg
Fett10 kg
Mm.l.d.30 kg
Mm.l.d.60 kg
Mm.l.d.90 kg
Fett30 kg
Fett60 kg
Fett90 kg
NN Nn nn
Abb. 4.2.1: Entwicklung von Muskel- und Fettauflagevolumen in einem 10 cm MRT-Bereich des Rückens bei Schweinen mit unterschiedlichen RyR1-GenotypenDie Werte für die 10 kg Gruppe wurden auf einen 10 cm MRT-Bereich korrigiert:
[Volumen (korrigiert) = gemessenes Volumen • 10 /2,45].

Diskussion
239
Beim Nutztier (Schwein) beeinflusst der Körperfettgehalt vor allem die
Produktqualität in Form des Schlachtkörpers sowie die Effizienz des Wachstums bzw.
des Fleischansatzes. Der Körperfettgehalt - und der Muskelgewebe- bzw. Proteingehalt
- sind die wichtigsten Nährstoffbestandteile im Schlachtkörper. Diese Merkmals-
komplexe sind primär in den Zuchtzielen der Schweineproduktion verankert. Abhängig
von der mit den beschriebenen Methoden (MRT, DXA, US, CT) messbaren
phänotypischen Variation in der Körperzusammensetzung interessiert den Tierzüchter
im Rahmen der Leistungsprüfung und Zuchtwertschätzung - neben der
Einzelgenwirkung - vor allem der additiv (aber auch nicht additiv) genetisch bedingte
Anteil an der Gesamtvariation der zu verändernden Merkmale. Es existieren zahlreiche
Arbeiten zur Heritabilität dieser Merkmale (Anteil der additiv genetischen Varianz an
der phänotypischen Varianz eines polygen gesteuerten Merkmals). Anstelle der
Ganzkörperanalyse mittels CT (Kapitel 2.2.3.2.5.1), MRT (Kapitel 2.2.3.2.5.2) oder
DXA (Kapitel 2.2.3.2.5.4) wird die Körperzusammensetzung hauptsächlich als
subkutane Fettauflage (Dicken- oder Flächenmaße) bzw. als subkutanes Fett-zu-
Muskel-Verhältnis an verschiedenen Messstellen des Rückens und der Seite des
Schweinekörpers entweder mittels Ultraschall (Kapitel 2.2.3.2.5.5) oder optischen
Verfahren (Kapitel 2.2.3.2.5.9) im Rahmen der Leistungsprüfung gemessen. Die
Heritabilitätsschätzwerte (h2) für Merkmale wie Rückenspeckdicke, Rückenmuskel-
fläche oder dem aus den vorangegangenen Maßen berechneten (Mager-)Fleischanteil
variieren zwischen 0,2 und 0,8. In Abhängigkeit von der Population (Rasse, Zuchtlinie),
Prüfmethodik (Einzel- oder Gruppenfütterung, ad libitum oder restriktiv), Prüfdauer
(zeit- oder masseabhängige Prüfung), Prüfabschnitt, Schätzmodell, geprüftem
Geschlecht (LANGHOLZ, 1966, PUFF, 1975, VALLE-ZARATE, 1981, LAMPO et al., 1985,
BUSSE und GROENEVELD, 1986, HUAROTO u.a., 1986, BRANDT, 1987, BERESKIN und
STEELE, 1988, SCHMIDT, 1988, HOFER, 1990, SCHOLZ, 1990, SCHOLZ und TRIEBLER,
1992, CAMERON und CURRAN, 1994, CAMERON et al., 1994, LI und KENNEDY, 1994,
BIDANEL et al., 1994, SEE, 1994, STERN et al., 1994, VON FELDE u.a., 1996,
ENGELLANDT u.a., 1997, GROENEVELD u.a., 1998, SCHULZE et al., 1998, SONESSON et
al., 1998, RUTEN u.a., 1999, HERMESCH et al., 2000, http://www.gparm.csiro.au/) oder
vom Schätzverfahren (REISER, 1984, DUCOS et al., 1992, HOFER, 1998) ergeben sich
große Abweichungen zwischen den Literaturwerten. Generell kann man davon
ausgehen, dass die additive genetische Varianzkomponente etwa 45 % bis 60 % der
Gesamtvariation in den Merkmalen der Körperzusammensetzung bzw.

Diskussion
240
Schlachtkörperqualität erklärt. Allein der RyR1-Genotyp erklärt in Abhängigkeit von
Population, Alter bzw. Lebendmasse ca. 3 – 17 % der Variation in den Merkmalen der
Körperzusammensetzung (Kapitel 4.1). Die vollständige Elimination des Defektallels
am RyR1-Genort, besonders in Populationen mit mittleren bis hohen Frequenzen für
dieses Allel, führt demzufolge zu einer Verminderung der genetischen Variation
(KRIETER und THOLEN, 2001). Solange die phänotypische Variation proportional
vermindert würde, blieben die Heritabilitätsschätzwerte (für Merkmale der
Körperzusammensetzung wie Rückenmuskelfläche oder Rückenspeckdicke)
unbeeinflusst. In Populationen mit einer annähernd 100%-igen Frequenz des
Defektallels (n) ist die genetische Variation für Merkmale der Körperzusammensetzung,
wie z.B. den Muskelfleischanteil, vermindert (BUSK et al., 1999). Die phänotypische
Variation ist offenbar nicht proportional reduziert, so dass die Heritabilitätsschätzwerte
und damit die nutzbare genetische Variation geringer ausfallen.
Generell liegen die h2-Schätzwerte für die Rückenmuskelfläche (-dicke) meist
über den Werten für die Rückenspeckdicke (-fläche), da die Rückenspeckdicke noch
stärker durch Umwelteinflüsse, z.B. durch restriktive Fütterung, verändert werden kann
als die Rückenmuskelfläche bzw. -dicke (DICKERSON und TEAGUE, 1987). Nach GERI et
al. (1990a) ist die Heritabilität für das Volumen der Adipozyten (Zellularität) ebenfalls
höher als die Heritabilität für die Rückenspeckdicke, wobei eine engere genetische
Korrelation zwischen Rückenspeck und Körpermasse bzw. Körperlänge als zur
Fettgewebezellularität besteht. Außerdem korreliert die Fettgewebezellularität zum
Zeitpunkt der Geburt mit dem Zellvolumen, dem Wassergehalt und dem Gehalt an
vollständig ungesättigten Fettsäuren C18:0 und C18:2 im Rückenspeck zum Zeitpunkt
des Schlachtens (GERI et al. 1990b). Fettere Schweinelinien besitzen dabei größere
Fettzellen (-durchmesser bzw. –volumina) als magere Schweinelinien (KALBITZ und
MUELLER, 1988).
Für Wachstumsmerkmale liegen die Heritabilitätsschätzwerte im Bereich von
0,10 bis 0,60 zumeist etwas niedriger als für Merkmale der Körperzusammensetzung
bzw. Schlachtkörperqualität (BUSSE und GROENEVELD, 1986, BERESKIN und STEELE,
1988, STERN et al., 1994, ENGELLANDT u.a., 1997, HERMESCH et al., 2000). Wachstum
wird überwiegend als durchschnittliche Tageszunahme während der Prüfperiode (Mast)
oder von Geburt bis zur Schlachtung erfasst. Eine differenziertere Herangehensweise
unterscheidet zwischen Magergewebe- und Fettgewebe- bzw. so genanntem
Restwachstum (FOWLER et al., 1976, WÄFLER u.a., 1984, WEBB und CURRAN, 1986,

Diskussion
241
KANIS, 1988, SCHOLZ, 1990, SCHOLZ und TRIEBLER, 1992). Hierfür müssen jedoch
geeignete Messmethoden wie Ultraschall, Dualenergie-Röntgenabsorptiometrie,
Computer- oder Magnetresonanz-Tomographie zur Ermittlung des Magergewebe- bzw.
Fettanteils im Laufe des Wachstums eines Individuums zur Verfügung stehen. Ohne die
exakte Messung der Körperzusammensetzung zu Beginn und am Ende eines
Wachstums- bzw. Prüfabschnitts, kann z.B. die Magergewebe- oder
Fettgewebewachstumsrate nur geschätzt und nicht genau ermittelt werden. Wenn die
phänotypische Variation für ein Merkmal in einer Population nur ungenau ermittelt
werden kann, ist sowohl die Heritabilitätsschätzung als auch die Identifikation der
Wirkung von Einzelgenen erheblich erschwert. In den meisten Prüfprogrammen wird
vor allem aus Kostengründen allein die Lebendmassezunahme geprüft und erst am Ende
der Prüfung die Körperzusammensetzung der Tiere ermittelt.
0
50
100
150
200
250
300
350
400
450
500
0 10 20 30 40 50 60 70 80 90Lebendmasse (kg)
Mag
erge
web
ewac
hstu
m (g
/d)
NN Nn nn
0
100
200
300
400
500
600
700
0 10 20 30 40 50 60 70 80 90Lebendmasse (kg)
Lebe
ndm
asse
zuna
hme
(g/d
)
NN Nn nn
Abb. 4.2.2: Entwicklung des Magergewebewachstums (links) und derLebendmassezunahme (rechts) bei Schweinen mit unterschiedlichen RyR1-Genotypen(Daten aus SCHOLZ, 2001)
Ähnlich wie im Ergebnisteil zu den Versuchen am ”Agricultural Research
Center“ des USDA in Beltsville (Maryland, USA) dargestellt (Kapitel 3.2.3), zeigen
noch „unveröffentlichte“ Ergebnisse von DXA-Untersuchungen am Lehr- und
Versuchsgut Oberschleißheim der Ludwig-Maximilians-Universität München deutliche
Unterschiede im Magergewebewachstum zwischen den drei untersuchten RyR1-
Genotypen (SCHOLZ, 2001). Zu Beginn des Wachstums bis zu etwa 40 kg Lebendmasse
zeigt der homozygote Defektallelgenotyp (nn) im Vergleich zum homozygot normalen
(NN) sowie zum heterozygoten Genotyp (Nn) bei gleichzeitig etwas geringeren
Lebendmassezunahmen (Abb. 4.2.2) noch ein langsameres Magergewebewachstum. Im
Laufe der Wachstumsphase bis zu ca. 65 kg nimmt die Magergewebewachstumsrate

Diskussion
242
schnell zu und übersteigt die Geschwindigkeit der Magergewebezunahme der beiden
anderen Genotypen bis zum Ende der Untersuchung bei etwa 90 kg Lebendmasse um
50 – 100 g/d. Nach dem Maximum bei 40 bzw. 65 kg sinkt die
Magergewebewachstumsrate in allen drei Genotypen wieder. Allein im heterozygoten
Genotyp nimmt die Magergewebewachstumsrate sehr langsam ab. Abbildung 4.2.2
zeigt in Bezug auf die durchschnittlichen Lebendmassen während der Messzeiträume
leicht voneinander abweichende Datenpunkte, da die homozygoten RyR1-
Defektallelträger bei gleichem (nicht aufgeführtem) Alter immer etwas geringere
Lebendmassen als die beiden anderen Genotypen aufweisen.
Insgesamt kann das Wachstum (Mastleistung) leichter durch
Umweltveränderungen modifiziert werden als die Schlachtkörperzusammensetzung.
Generell erklärt die additive genetische Varianzkomponente etwa 30 % bis 40 % der
Gesamtvariation in den Wachstumsmerkmalen (Lebenstagszunahme, Prüftagszunahme,
Magergewebezunahme).
Da die „modernen“ Zuchtprogramme den größten Selektionsdruck auf die
Züchtung magerer Schweine gerichtet haben, wurden - im Gegensatz zur zunehmenden
„Fettleibigkeit“ der in den hoch industrialisierten Ländern lebenden Menschen (BARTH
et al., 1997, KROMEYER-HAUSCHILD und JAEGER, 1998, LUKE et al., 1998, TRIPPO,
2000) und seiner Haustiere (HARPER, 1998) - sehr kleine, aber in der
Fettsäurezusammensetzung stärker ungesättigte, subkutane Fettdepots in Kombination
mit einer verminderten lipogenen Enzymkapazität erreicht (STEELE und FROBISH 1976,
METZ, 1985, KALBITZ und MUELLER, 1988, MAILÄNDER 1988, SCHLINDWEIN, 1990,
MÜLLER u.a., 1996). Die Ergebnisse einer über 9 Generationen andauernden Selektion
auf verminderte lipogene Enzymaktivität (NADPH-lieferende Enzyme), die zu einer
verminderten Rückenspeckdicke führte (ROGDAKIS u.a., 1996, MÜLLER u.a., 1996),
bestätigen diese Beziehung.
Nach MERKS et al. (1997) ging die Ultraschall-Rückenspeckdicke im Herdbuch
der niederländischen Yorkshire-Vaterlinien von etwa 20 mm an der P2-Messstelle im
Jahr 1960 auf weniger als 10 mm im Jahr 1997 zurück. Das mittlere Heritabilitätsniveau
verminderte sich dabei nicht, da die phänotypische und genetische Variation
proportional sanken. Die Autoren weisen zugleich darauf hin, dass sehr magere
Schweine weniger genetisch variieren als fettere Schweine, genauso wie Schweine mit
wenig Rückenspeck (< 10 mm) und hohen Tageszunahmen (> 650 g) bzw. Schweine
mit einer starken Speckauflage (> 10 mm) und niedrigen Tageszunahmen (< 650 g).

Diskussion
243
Sinkt unter Verwendung der herkömmlichen Methoden die messbare
phänotypische und genetische Variation für ein Merkmal wie z.B. die
Rückenspeckdicke, wird es für den Tierzüchter erforderlich, entweder verfeinerte
Messmethoden zu entwickeln oder mit modifizierten Methoden (z.B. MRT,
Videobildanalyse) Merkmale an anderen Messorten (z.B. Bauchfettanteil) für die
weitere Selektion zu etablieren (BAULAIN u.a., 1998a,b, THOLEN u.a., 1998a,b, BAULAIN
und HENNE, 1999).
In den meisten Fällen verfügen magere Schweine über eine höhere
Proteinansatzrate als fette Schweine (SIEBRITS et al., 1985), wobei eine verminderte
Rückenspeckdicke mit einem erhöhten Proteingehalt im Schlachtkörper und
insbesondere im Fleisch korreliert ist. Jeder Millimeter Reduktion in der
Rückenspeckdicke führt zu einer Erhöhung des Proteingehalts im gesamten
Muskelfleisch um 19,5 g/kg (FORTIN und ELLIOT, 1985). Sobald jedoch die
fleischreicheren (magereren) Tiere aufgrund einer vergleichsweise niedrigeren
Futteraufnahme stark reduzierte Tageszunahmen erzielen, können die
Proteinansatzraten - wie in den eigenen Ergebnissen zwischen 60 und 90 kg
Lebendmasse (Tab. 3.2.41, Abb. 3.2.20) - unter die der Tiere mit einem mittleren
Verfettungsgrad sinken. Dabei ist vorauszusetzen, dass die Tiere mit einem mittleren
Verfettungsgrad – im eigenen Versuch Nn - mehr Futter für die Realisierung höchster
Tageszunahmen aufnehmen als die magersten Tiere (WÄFLER u.a., 1984, KRIETER,
1986, KANIS, 1988, WEBB, 1989, CAMERON und CURRAN, 1994).
Als eine Konsequenz des verminderten Verhältnisses zwischen Fett- und
Magergewebeansatz wird die Fettqualität negativ verändert. Der Anteil von Rohfett
innerhalb des Fettgewebes ist vermindert (FORTIN und ELLIOT, 1985); und die
Fettsäuren des Fettgewebes sind infolge eines relativ geringeren Beitrages der De-novo-
Fettsäuresynthese zur Gesamtsynthese weniger gesättigt (METZ, 1985).
Korrespondierend erlangen andere (sichtbare) Fettdepots wie abdominales und
intermuskuläres Fett eine größere Bedeutung (DE VRIES und KANIS, 1994), wobei das
subkutane Fettdepot dominiert und rasseunabhängig im Verlauf des Wachstums vor den
beiden anderen Fettdepots angelegt wird. Der Anteil des abdominalen (internen) und
intermuskulären/intramuskulären Fettes am Gesamtfett nimmt jedoch mit steigender
Lebendmasse zu (KOLSTAD, 2000).
Korrespondierend demonstriert der Fett-Verteilungs-Index nach LISTER (1976)
eine große Variation zwischen mehreren Schweine-Genotypen mit dem Wildschwein

Diskussion
244
am unteren Ende und Hampshire an der Spitze, während Pietrain, Landrasse sowie
Large White (Edelschwein) dazwischen liegen. Der Fett-Verteilungs-Index ist als
Verhältnis der Masse des messbaren subkutanen Fettes zur Summe der Massen von
intermuskulärem Fett, Nieren umlagerndes Fett und „Inguinalfett“ (in der
Leistengegend vorhandenes Fett) in einem Schlachtkörper definiert.
Diese Ergebnisse werden u.a. von TESS et al. (1986) und KOLSTAD et al. (1996)
bestätigt. Nach KOLSTAD et al. (1996) besitzt die Norwegische Landrasse mehr
Innereienfett und weniger inter/intramuskuläres Fett als Duroc. Es konnten jedoch keine
Unterschiede im subkutanen Fettgewicht zu Beginn einer Erhaltungsfütterungsperiode
festgestellt werden. Als Ergebnis der restriktiven Fütterung auf Erhaltungsniveau kam
es in den Landrasse-Schweinen durch einen höheren Prozentsatz an Fettmobilisierung
aus allen Fettdepots zu einem höheren Grad der Fettdepotumverteilung als bei Duroc.
In Übereinstimmung mit den eigenen MRT-Ergebnissen für die Kreuzung [Duroc
x Minzhu] im Vergleich zu den anderen Linien (Kapitel 3.2.2.1, Tab. 3.2.6) zeigen sich
die größten Unterschiede im Körperfettgehalt von kommerziell genutzten
Schweinerassen zwischen einer Vielzahl von Chinesischen Schweinerassen (und deren
Kreuzungsprodukten) wie Meishan, Minzhu, Fengjing, Jiaxing, Jinhua und den
mageren „modernen“ Schweinerassen mit europäischem oder amerikanischem
‘Ursprung’ wie Pietrain, diversen Landrassen, Yorkshire oder Hampshire (MOLENAT,
1987, MOLENAT und CARITEZ, 1987, HERPIN et al., 1993, QUINIOU und NOBLET, 1995,
WHITE et al., 1995, CLAUS, 1996). Bereits YOUATT und MARTIN (1865) zitierten Hurtrel
D‘Arboval, dass insbesondere die chinesischen und siamesischen Schweine für einen
extremen Fettwuchs prädestiniert sind („There is, however no animal so liable to
become over-fat, as the pig, and especially Chinese and Siamese swine.“). Duroc bzw.
Linien oder Kreuzungen mit hohen Duroc-Genanteilen besitzen innerhalb der mageren
„modernen“ Schweinelinien den höchsten intramuskulären Fettgehalt (SCHWÖRER und
REBSAMEN, 1990a, VANGEN und ENTING, 1990, AFFENTRANGER et al., 1991, ALTMANN
u.a., 1992, MAASSEN-FRANCKE u.a., 1992, DE VRIES et al., 1993, KALLWEIT et al., 1993,
MÜLLER, S. u.a., 1996, BAULAIN et al., 2000).
Diese Differenzen können zum Teil durch genetisch bedingte
Konzentrationsunterschiede zwischen Wachstumshormon bzw. „Insulin ähnlichem
Wachstumsfaktor“ (insulin like growth factor - IGF-1) mit anaboler Wirkung und
Kortisol mit kataboler Wirkung bedingt sein (CLAUS und WEILER, 1994, LOUVEAU et
al., 1995). Auf Fettgewebe wirkt das Wachstumshormon jedoch katabol, d.h. lipolytisch

Diskussion
245
(KARG, 1989, CLAUS, 1996, ELSASSER und PARVIZI, 1996, ETHERTON und BAUMAN,
1998, MÜLLER et al., 1999). In diesem Zusammenhang weist die Rasse Meishan
geringere IGF-1- und höhere Kortisolkonzentrationen als zum Beispiel die Rasse
Edelschwein (bzw. Large White) auf (LOUVEAU et al., 1995, CLAUS, 1996). Da IGF-1
unter der Mitwirkung von Wachstumshormon gebildet wird und über die Besetzung der
IGF-1-Rezeptoren das Wachstum von Muskel- und Skelettgewebe stimuliert (ELSASSER
und PARVIZI, 1996, MÜLLER et al., 1999), ist der sehr hohe Verfettungsgrad der Rasse
Meishan zumindest teilweise durch die vergleichsweise niedrige IGF-1-Konzentration
erklärbar. WINDISCH u.a. (2000) fanden jedoch im Verlaufe eines Proteinstoffwechsel-
versuches unter restriktiven Fütterungsbedingungen bei gleichem Gehalt an
Wachstumshormon (3,8 bzw. 4,0 ng/ml) in Tieren der Rasse Pietrain (MHS-Status Nn
oder nn) mit 157 ng/ml einen niedrigeren IGF-1-Blutplasma-Gehalt als in den etwas
fetteren Tieren der Deutschen Landrasse (MHS-Status NN) mit 298 ng/ml. Erklärt
wurden diese Unterschiede mit einer stärkeren hormonellen Drosselung der
Proteinsynthesekapazität bei der Rasse Pietrain, die durch die mengenmäßig identische
Restriktion der Nährstoffzufuhr möglicherweise stärker in ihrem Leistungspotential
eingeschränkt wurde als die Tiere der Deutschen Landrasse. Aufgrund dieser zum Teil
widersprüchlichen Ergebnisse ist es nicht verwunderlich, dass KNORR et al. (1997) zwar
Assoziationen zwischen Wachstumshormon-Genotypen und dem Körperfettgehalt in
einer Meishan x Pietrain Familie fanden, aber keine signifikanten Assoziationen in einer
Wildschwein x Pietrain Familie nachweisen konnten. CASAS-CARRILLO et al. (1997)
berichteten über eine mögliche Kopplung zwischen dem IGF-1-Genotyp und der
täglichen Zunahme nach dem Absetzen in einer Eber-Familie, aber nicht zwischen dem
Wachstumshormon-Genotyp und dem Körperfettgehalt oder Wachstumsmerkmalen.
Diese Ergebnisse beruhen möglicherweise auf der Aktion der IGF-Bindungsproteine
(IGFBP 1-6), die die Wirkung von IGF I bzw. II in Muskel- und Lebergewebe
modulieren (LOUVEAU et al., 1995, SIMMEN et al., 1998, GERRARD et al., 1999, MÜLLER
et al., 1999, CLAPPER et al., 2000, SCHNEIDER et al., 2000). Transgene Mäuse, die
IGFBP2 (mRNA und Protein) im Überschuss produzieren, weisen im Vergleich zur
gleichaltrigen nicht transgenen Kontrollgruppe ab dem 25. Lebenstag ein vermindertes
Körpergewicht auf. Ein erhöhtes Niveau von IGFBP2 kann folglich die Wirkung von
IGF-I einschränken (HOEFLICH et al., 1999, SCHNEIDER et al., 2000, WOLF et al., 2000).
CLAPPER et al. (2000) vermuten, dass altersabhängige Veränderungen in der Serum-
Konzentration von IGF-I und relativen Mengen von IGFBP, die auf der

Diskussion
246
Konzentrationsveränderung von Estradiol-17ß und Testosteron beruhen, zu den
Wachstumsdifferenzen zwischen unterschiedlichen Geschlechtern beim Schwein
beitragen. Eber zeigen dabei durchgängig die höchsten Serum-Konzentrationen an
IGF-I, womit deren höhere Wachstumsleistung im Vergleich zu männlichen Kastraten
und Sauen partiell erklärt werden kann.
1.4
1.6
1.8
2
2.2
2.4
2.6
2.8
3
30 - 40 40 - 60 60 - 80 80 - 100
Gewichtsabschnitt (kg)
durc
hsch
nittl
iche
rFu
tterv
erze
hr (k
g/d)
DL DE LC (46% DU)PI x F1 LB x F1 HAPI x F1
Abb. 4.2.3: Durchschnittlicher Futterverzehr verschiedener Rassen bzw. Kreuzungenwährend der Leistungsprüfung von ca. 30 – 100 kg Lebendmasse (mit DL=Deutsche Landrasse, DE=Deutsches Edelschwein, LC=Leicoma, DU=Duroc, PI x F1=Pietrainx F1-Sau, LB x F1= Belgische Landrasse x F1-Sau, HAPI x F1=[Hampshire x Pietrain] x F1-Sau; Datenstammen aus PAULKE und SCHOLZ, 1999)
In Übereinstimmung mit den vorliegenden Ergebnissen (Abb. 3.2.19, Kapitel
3.2.3) und den in Abbildung 4.2.3 dargestellten Ergebnissen von PAULKE und SCHOLZ
(1999) sind die magersten, stressanfälligen Schweinerassen (Vaterrassen) - mit einer
hohen Allelfrequenz für das Defektallel (n) am RyR1-Gen - wie Pietrain und Belgische
Landrasse sowie deren Kreuzungsprodukte im Vergleich zu den belastbareren
Mutterrassen - mit einer sehr niedrigen Allelfrequenz für das RyR1-Defektallel - durch
ein vermindertes Futteraufnahmevermögen in ihrem Fettansatzpotential begrenzt
(LISTER, 1976, HONG, 1985, KRIETER und KALM, 1986, STIEWE, 1986, BRANDT, 1987,
KANIS, 1988, WHITTEMORE et al., 1995). Untersuchungen am Lehr- und Versuchsgut
Oberschleißheim zeigen, dass homozygot „normale“ Tiere innerhalb der Rasse Pietrain
(weiß) signifikant mehr Fett ansetzen als ihre heterozygoten bzw. homozygoten
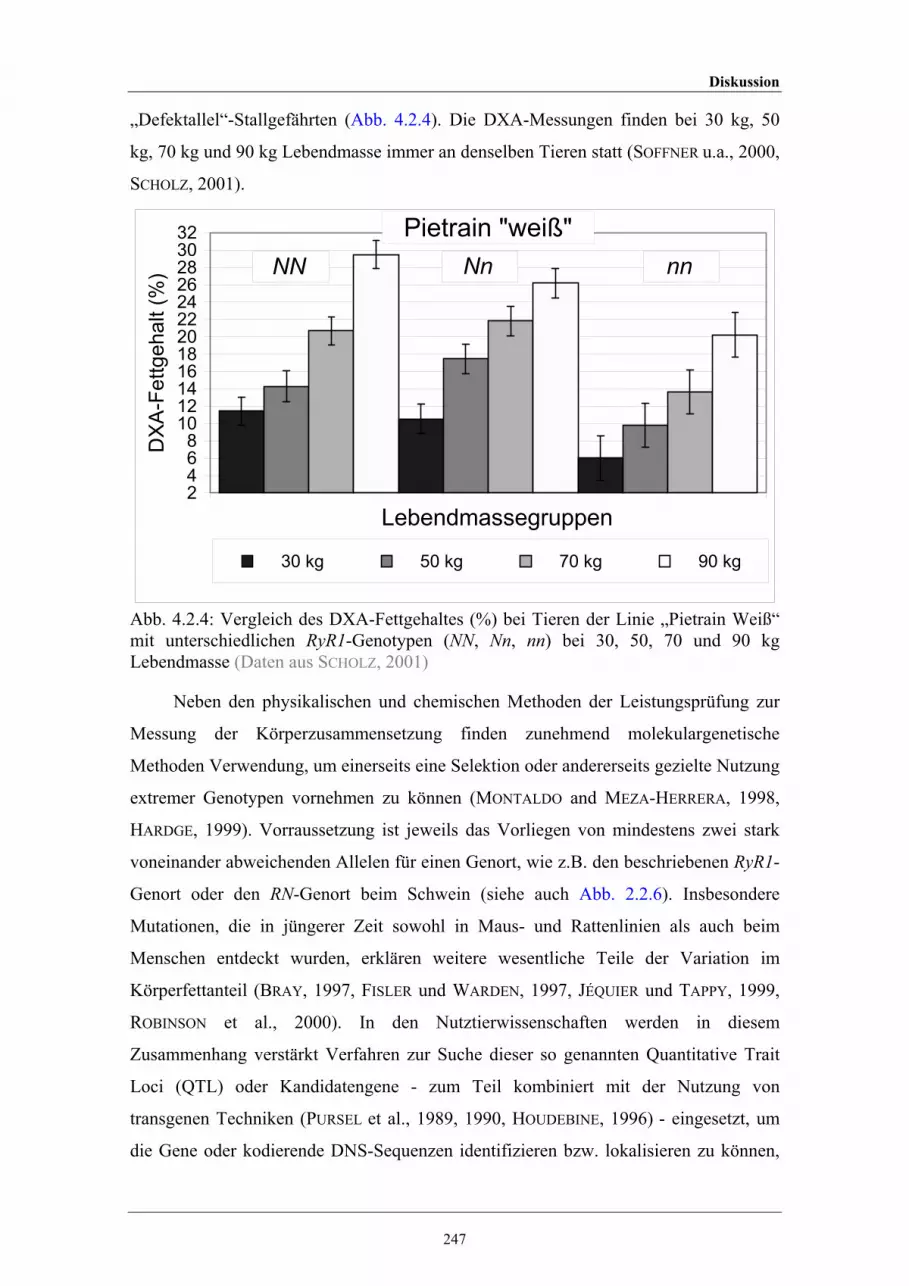
Diskussion
247
„Defektallel“-Stallgefährten (Abb. 4.2.4). Die DXA-Messungen finden bei 30 kg, 50
kg, 70 kg und 90 kg Lebendmasse immer an denselben Tieren statt (SOFFNER u.a., 2000,
SCHOLZ, 2001).
Abb. 4.2.4: Vergleich des DXA-Fettgehaltes (%) bei Tieren der Linie „Pietrain Weiß“mit unterschiedlichen RyR1-Genotypen (NN, Nn, nn) bei 30, 50, 70 und 90 kgLebendmasse (Daten aus SCHOLZ, 2001)
Neben den physikalischen und chemischen Methoden der Leistungsprüfung zur
Messung der Körperzusammensetzung finden zunehmend molekulargenetische
Methoden Verwendung, um einerseits eine Selektion oder andererseits gezielte Nutzung
extremer Genotypen vornehmen zu können (MONTALDO and MEZA-HERRERA, 1998,
HARDGE, 1999). Vorraussetzung ist jeweils das Vorliegen von mindestens zwei stark
voneinander abweichenden Allelen für einen Genort, wie z.B. den beschriebenen RyR1-
Genort oder den RN-Genort beim Schwein (siehe auch Abb. 2.2.6). Insbesondere
Mutationen, die in jüngerer Zeit sowohl in Maus- und Rattenlinien als auch beim
Menschen entdeckt wurden, erklären weitere wesentliche Teile der Variation im
Körperfettanteil (BRAY, 1997, FISLER und WARDEN, 1997, JÉQUIER und TAPPY, 1999,
ROBINSON et al., 2000). In den Nutztierwissenschaften werden in diesem
Zusammenhang verstärkt Verfahren zur Suche dieser so genannten Quantitative Trait
Loci (QTL) oder Kandidatengene - zum Teil kombiniert mit der Nutzung von
transgenen Techniken (PURSEL et al., 1989, 1990, HOUDEBINE, 1996) - eingesetzt, um
die Gene oder kodierende DNS-Sequenzen identifizieren bzw. lokalisieren zu können,
2468
101214161820222426283032
DXA
-Fet
tgeh
alt (
%)
30 kg 50 kg 70 kg 90 kg
NN Nn nn
Lebendmassegruppen
Pietrain "weiß"

Diskussion
248
die einen komplexen multifaktoriellen Phänotyp beschreiben helfen (MONTALDO and
MEZA-HERRERA, 1998, HARDGE, 1999).
Zu den jüngsten molekulargenetischen Errungenschaften gehört die Entdeckung
von einzelnen regulatorischen Genen (Mutationen) des Fettstoffwechsels wie das
LEPTIN- (LEP) /OBESE- (OBS) Gen auf dem Chromosom 6 der Maus (FRIEDMANN, 1991,
ZHANG et al., 1994). Das „normale“ Gen produziert ein den Appetit steuerndes Hormon
Leptin, welches vom Fettgewebe des Schweines proportional zur Menge angesetzten
Fettes sekretiert wird (SPURLOCK et al., 1998, Abb. 4.2.5). Diese Beziehung deckt sich
mit den von ROSENBAUM et al. (1996) ermittelten Beziehungen beim Menschen (zitiert
bei LUKE et al., 1998), während LUKE et al. (1998) eine exponentielle Beziehung
zwischen Plasma-Leptin-Konzentration (µg/L) und Fettmasse bzw. Fettanteil (%)
berechneten.
0
20
40
60
80
100
120
140
160
0 10 20 30 40 50
Fettmasse (kg)
obese
mRN
A
Abb. 4.2.5: Anstieg des Gehalts an obese mRNA (in Analyse internen Einheiten) inAbhängigkeit von der Fettmasse beim Schwein, Gewichte der Schweine betrugen in derReihenfolge: 23, 55, 107 und 162 kg (nach Daten von SPURLOCK et al., 1998)
Generell signalisieren hohe Leptinkonzentrationen dem Gehirn (Hypothalamus),
den Appetit oder das Hungergefühl zu vermindern. Individuen mit einem Defekt im
OBESE-Gen sind nicht in der Lage Leptin zu produzieren. Im Wachstum befindliche
Individuen können bei gesteigerten Lipogeneseraten der Leber ihren Appetit nicht
steuern, was eine ungezügelte Futteraufnahme zur Folge hat. Neben negativen Effekten
auf das Immunsystem und vor allem auf die Sexualentwicklung sind eine verminderte
Körpertemperatur und eingeschränkte Basalstoffwechselraten zu beobachten. Leptin
leitet sich vom griechischen leptos (= dünn) ab. Es handelt sich um ein 16 kD großes

Diskussion
249
Protein, das nur in Fettzellen gebildet wird. Das aus der Fettzelle in die Blutbahn
freigesetzte Leptin signalisiert dem ZNS die Menge des gespeicherten Fetts. Das heißt,
je größer die Fettzelle ist, desto mehr Leptin wird von der einzelnen Fettzelle
freigesetzt. Dieser Vorgang wird durch eine Reihe humoraler Faktoren moduliert.
Insulin und Glucokortikoide führen zu einer vermehrten Leptinsynthese, während
Geschlechtshormone, Katecholamine und Schilddrüsenhormone die Leptinsynthese
vermindern. Leptin gelangt durch die Blut-Hirn-Schranke zum ZNS und bindet dort
insbesondere an Leptinrezeptoren im Hypothalamus. Dies führt zu einer verminderten
Nahrungsaufnahme und zu einem erhöhten Energieverbrauch. Die Bedeutung der
Leptinrezeptoren in anderen Anteilen des ZNS, aber auch verschiedenen peripheren
Geweben ist noch unklar. Das Neuropeptid Y ist möglicherweise an der weiteren
Vermittlung des Signals beteiligt, da Leptinrezeptoren auf den Neuropeptid-Y-haltigen
Neuronen nachweisbar sind und die Zufuhr von Leptin zu einer Verminderung der
Neuropeptid-Y-Bildung und Ausschüttung im Hypothalamus führt. Individuen mit zu
hohen Leptinkonzentrationen vermindern - neben einem erhöhten Energieverbrauch -
ihre Futteraufnahme und lehnen Futter sogar ab, woraus signifikante
Körpergewichtsverluste resultieren. Bei der übergewichtigen, fettwüchsigen ob/ob-
Maus liegt eine Mutation vor, die zu einem falschen Stop-Codon führt, so dass nur ein
defektes, zu kurzes "Leptin" synthetisiert wird. Die Gabe von Leptin führt bei diesen
Mäusen durch verminderte Nahrungsaufnahme und vermehrten Energieverbrauch zur
Gewichtsabnahme (HWANG et al, 1997, HOUSEKNECHT et al., 1998, NORTH, 1999,
STROSBERG und ISSAD, 1999, XIE u.a., 1999, AHIMA und FLIER, 2000).
In engem Wechselspiel zum OBESE-Gen steht das DIABETIC-Gen (db-, Lepr- oder
Ob-R-Gen), welches für die Kodierung des Leptinrezeptors zuständig ist. Der Defekt im
db-Gen - vor allem in seiner „Langform“ Ob-Rb aber auch in der kürzesten „Spleiß“-
Variante OB-Re führt zu einer starken Verfettung bereits in frühem Alter sowie
ineffizienter Appetitsregulation (NORTH, 1999, STROSBERG und ISSAD, 1999, REICHART
et al., 2000a, b).
ERNST et al. (1997) lokalisierten das Leptinrezeptor-Gen des Schweines (PORCINE
LEPTIN RECEPTOR gene - LEPR) physisch auf dem Chromsom 6 (6q3.3-q3.5) mit 100 %
Konkordanz zu den korrespondierenden Ergebnissen von VINCENT et al. (1997) in einer
Kopplungsanalyse zur Chromosomenkartierung.
Nachdem zwei Gruppen (NEUENSCHWANDER et al., 1996 und SASAKI et al., 1996)
das „Verfettungs“-Gen des Schweines (PORCINE OBESITY oder LEPTIN-Gen) dem

Diskussion
250
Chromosom 18 zugewiesen hatten, wurde es von mehreren Forschungsgruppen
(BIDWELL et al., 1997, DAI et al., 1998, RAMSAY et al., 1998) kloniert. Dadurch
eröffnen sich in Kombination mit den beschriebenen Messmethoden (CT, MRT, DXA)
viel versprechende Möglichkeiten, den Effekt dieses Gens auf die Regulation der
Fettdepots und der Körpermasse zu untersuchen. Das homologe humane LEPTIN-Gen
mit nachgewiesenen rezessiv pathogenen Mutationen liegt auf dem Chromosom 7
(7q31-7q32; WEIGLE und KUIJPER, 1996, STROSBERG und ISSAD, 1999).
Erste Ergebnisse einer Assoziationsstudie zwischen Polymorphismen des LEPTIN-
Gens und Leistungsmerkmalen offenbarten nur einen signifikanten Effekt auf den
Körperfettgehalt innerhalb einer stark verfetteten F2-Resource-Familie, die aus
reziproken Kreuzungen von Schweinen der Rasse Duroc und des Berliner Miniatur-
Schweines entstand. Andere kommerzielle Schweinerassen wie Deutsche Landrasse,
Edelschwein und Pietrain zeigten keine signifikanten Assoziationen zwischen Varianten
des LEPTIN-Gens und dem Körperfettgehalt (HARDGE et al., 1998), während JIANG und
GIBSON (1999) für Hampshire, Large White, Landrasse und Duroc bzw. KULIG et al.
(2001) für Tiere der Polnischen Landrasse entsprechende Assoziationen fanden. Tiere
mit dem Genotyp CC sollen einen geringeren Fettgehalt (höheren Fleischgehalt)
aufweisen als Tiere mit dem Genotyp TC oder TT (KULIG et al., 2001).
Die gewichtsreduzierende Wirkung von Leptin ist offenbar nicht allein über seine
durch die Hypothalamus-Leptinrezeptoren vermittelte Wirkung auf die
Appetitregulation zu erklären. Leptin übt ebenfalls direkte Effekte auf (braunes und)
weißes Fettgewebe aus, da sich auch im Fettgewebe mRNA der Leptinrezeptoren
nachweisen ließ. Physiologische Konzentrationen von Leptin steigern die Lipolyse im
weißen Fettgewebe, während dieser Effekt bei genetisch adipösen Ratten (Fa/fa), die
infolge einer Mutation eine Inaktivierung ihres Leptinrezeptors aufweisen, nicht
gefunden werden konnte (SIEGRIST-KAISER et al., 1997).
Im Gegensatz zu den rezessiven OBESE- und DIABETIC-Mutationen (in Mäusen) mit
einer bereits im Jugendstadium beginnenden Verfettung, scheinen ausgewachsene
Träger der dominanten AGOUTI YELLOW-Mutation effizienter in der Speicherung von
übermäßig konsumierten Nährstoffen in der Form von Fett zu sein. Überschüssige
Energie wird folglich nur zu einem relativ geringen Grad für physische Aktivitäten bzw.
für die Aufrechterhaltung der Körpertemperatur umgesetzt. Die AGOUTI- und AGOUTI
verwandten Genprodukte unterdrücken im Zusammenspiel mit PROOPIOMELANOCORTIN-
abhängigen Peptiden die Bindung des α-Melanozyten stimulierenden Hormons an den

Diskussion
251
Melanocortin-4-Rezeptor sowie den Melanocortin-1-Rezeptor (WILSON et al., 1999,
KRUDE und GRÜTERS, 2000), welcher durch das EXTENSION-Gen – eine weitere, die
Fellfarbe bestimmende Allelserie - kodiert ist. Mutationen am EXTENSION-Genort – auf
dem Schweine-Chromosom 6 befindlich (KIJAS et al., 1998) - agieren epistatisch
gegenüber AGOUTI-Mutationen. Ferner ist die durch AGOUTI induzierte Verfettung
wahrscheinlich durch antagonistische Wirkungen der Melanocortin-4- und
Melanocortin-3-Rezeptoren im Gehirn und/oder peripheren Geweben beeinflusst.
Während Neuropeptid Y den Appetit stimuliert und den Energieverbrauch einschränkt,
wird entgegengesetzt der Melanocortin-4-Rezeptor (MC4-R) aktiviert, wenn das
Melanozyten stimulierende Hormon sich an ihn bindet. Das geschieht, wenn der
Leptinspiegel bei einer Gewichtszunahme steigt. MC4-R dämpft den Appetit und
steigert den Energieverbrauch (MILTENBERGER et al., 1997, OLLMANN et al., 1998,
WILSON et al., 1999).
Innerhalb der obigen Kaskade von Genwirkungen unterdrückt das MAHOGANY-Gen
(mg) auf dem Chromosom 2 der Maus die genetisch induzierte Verfettung von AGOUTI-
YELLOW Mäusen, aber nicht die „verfetteten“ Phänotypen des Melanocortin-4-Rezeptor-
Null-Allel-Modells oder der monogenetisch bedingten „Verfettungs-Modelle“ (OB, DB,
TUB, oder FAT). Zusätzlich kann MAHOGANY eine nahrungsinduzierte Verfettung
unterdrücken. Mäuse mit einer Mutation im MAHOGANY-Gen halten unabhängig vom
Fettgehalt (42 % zu 9 %) bei konstantem Energiegehalt in der Ration ein ‘normales’
Gewicht aufrecht, während Mäuse ohne Mutation nach Verzehr der Hoch-Fett-Ration
sehr stark zunehmen (NAGLE et al., 1999). Das MAHOGANY-Gen agiert der Transkription
von AGOUTI nachgeschaltet, aber der Transkription des Melanocortin-4-Rezeptors
vorgeschaltet. Da Menschen eng verwandte Homologien zum Maus-AGOUTI-Gen
(KWON et al., 1994) und zum Maus-MAHOGANY-Gen (HUMAN ATTRACTIN) aufweisen
(EDEAL et al., 2000), könnte es möglich sein, auch beim Schwein Kopplungen zwischen
Genen aufzudecken, die die Fellfarbe oder die Immunzell-Interaktionen kodieren und
Leistungsmerkmale beeinflussen. Gegenwärtig ist es jedoch völlig offen, ob Varianten
in der Fellfarbe des Schweines bzw. die Immunantwort kodierende Gene signifikant mit
der Körperzusammensetzung oder dem Wachstum assoziiert sind. Das MAHOGANY-Gen
der Maus wurde von NAGLE et al. (1999) und GUNN et al. (1999) kloniert. Beim
Schwein ist das MAHOGANY-Gen (PORCINE ATTRACTIN) auf Chromosom 17 lokalisiert
(EDEAL et al., 2000).

Diskussion
252
Ein weiterer möglicher Hauptgenort für den Fettansatz (Rückenspeck und
Abdominalfett) bzw. für den Anteil von „Muskelfleisch + Knochen“ wurde von
ANDERSSON et al. (1994, 1996, 1998) bzw. ANDERSSON-EKLUND (1998) am proximalen
Teil des Chromosoms 4 vom Schwein mittels QTL-Kartierung identifiziert, wofür eine
Kreuzung aus Wildschwein mit Large White genutzt wurde. WALLING et al. (1998),
KNOTT et al. (1998) sowie PEREZ-ENCISO et al. (2000) fanden korrespondierende
Effekte für die durchschnittliche Rückenspeckdicke bzw. die Fettsäure-
zusammensetzung in der gleichen Region des Chromosoms 4. Der QTL-Effekt auf den
subkutanen Fettansatz betrug signifikant 15 – 18 % der gesamten phänotypischen
Varianz, d.h. die absolute Differenz zwischen den homozygoten Genotypen entspricht
etwa 5 mm durchschnittlicher Rückenspeckdicke. Außerdem ergaben erste Ergebnisse
von LIU et al. (1995), die durch WANG, L. et al. (1997) gestützt werden, Hinweise auf
eine Assoziation der Region um den porcinen TUMOR NECROSE FACTOR alpha (TNFα)
auf dem Chromosom 7 mit Differenzen im Niveau des Körperfettgehaltes, der
Bemuskelung und des Wachstums. Jedoch zeigen nur die Speckdicken an der letzten
Rippe, der 10. Rippe sowie die durchschnittliche Speckdicke signifikante Marker-QTL-
Beziehungen, wobei die Allele der chinesischen Rassen (Meishan und Minzhu) über
einen negativen additiven Geneffekt die „mageren“ Allele darstellten. MOSER et al.
(1998) und MILAN et al. (1998) lieferten den Nachweis für ein (kryptisches) Allel für
das Merkmal Rückenspeckdicke auf dem Chromosom 7 des Schweines, welches
ROHRER und KEELE (1998) und ROHRER (2000) ebenfalls fanden.
Genomuntersuchungen beim Menschen und Nagetieren stützen im Einklang mit den
Assoziationen beim Schwein die These eines Effekts von (Markern in der Nähe von)
TNFα und der Lipoprotein-Lipase auf den Fettansatz (NORMAN et al., 1995, KERN,
1997, MOLLER, 2000). Das PORCINE COLIPASE–Gen (CLPS) als ein erforderlicher
Kofaktor für den durch die Lipase regulierten Nahrungslipid-Stoffwechsel ist ein
weiteres Kandidatengen für einen potentiellen Effekt auf die Körperzusammensetzung
innerhalb des Haupthistokompatibilitäts-Komplexes auf Chromosom 7 (BASKIN und
POMP, 1998).
YU et al. (1997) kartieren den PIT-1-Genort auf dem Chromosom 13 des
Schweines. PIT-1 ist ein transkriptionaler regulatorischer Wachstumsfaktor für das
WACHSTUMS-HORMON-Gen sowie PROLAKTIN- und THYROTROPIN-B-Gen-
Untereinheiten. Die Autoren berichten, dass sich PIT-1 (unter Verwendung des
Restriktionsenzyms MspI) in enger Nachbarschaft der Region auf Chromosom 13

Diskussion
253
befindet, welche QTL für Geburtsgewicht und frühes Wachstum enthält, die zuerst von
ANDERSSON et al. (1994) gefunden wurden.
Wie vorab beschrieben, ist es nahe liegend, dass Teile der natürlichen Variation
des Körperfettgehaltes mit Modifikationen in der Effizienz der Leptinregulation
zusammenhängen (XIE et al., 1999, AHIMA und FLIER, 2000). Eine Vielzahl anderer
Faktoren ist gleichermaßen in die Steuerung des Körperfettgehaltes eingebunden (Abb.
4.2.6) - zum Beispiel die Anwesenheit von energieverbrauchendem braunem
Fettgewebe im Zusammenspiel mit dem sogenannten mitochondrialen UNCOUPLING
PROTEIN 1 (SAXTON und EISEN, 1984, LOWELL und FLIER, 1997, NORTH, 1999).
LOWELL et al. (1993) beschreiben, wie genetisch veränderte Mäuse, die ihr eigenes
braunes Fettgewebe zerstörten, danach sehr stark verfetteten. Braunes Fettgewebe ist
beim Schwein jedoch histologisch nicht nachweisbar. Aus diesem Grund ist
anzunehmen, dass die Effekte von UNCOUPLING PROTEIN 2- oder 3- und ß3-
ADRENERGISCHEN REZEPTOR-Gen-Varianten auf die Körperzusammensetzung und
Energiebalance, die bei Nagetieren und teilweise beim Menschen nachgewiesen
wurden, beim Schwein keine signifikante Auswirkung haben.
Interessanter, speziell für Schweine(züchter), ist die Entschlüsselung des
Gens, welches für die Doppellendigkeit beim Rind zuständig ist. Mindestens zwei
„Funktionsverlust“-Mutationen innerhalb des “GROWTH/DIFFERENTIATION FACTOR-8”-
(GDF8) oder MYOSTATIN-Gens – ein Mitglied der “TRANSFORMING GROWTH FACTOR-ß”-
(TGF-ß) Superfamilie – sind für die extreme Muskelzellhyperplasie und daraus
resultierenden hervorgehobenen Schulter- und Hüftbereichen speziell bei den
Rinderrassen Blau-Weiße Belgier und Piemonteser sowie bei (homozygot) mutierten
Mäusen zuständig. Der hohe Grad der Sequenzkonservierung für GDF8 in Tieren
einschließlich des Schweines (Chromosom 15) stellt ein potentielles Ziel zur
Vergrößerung der Muskelmasse ohne negative Seiteneffekte auf die Fleischqualität dar
(z.B. MCPHERRON et al., 1997, VARGA et al., 1997, SZABO G. et al., 1998, SONSTEGARD
et al., 1998). Die Muskelfaseranzahl und Muskelmasse wird neben dem MYOSTATIN-Gen
durch die MYOGENIN-Gen-Familie reguliert (TE PAS et al., 1999).
Innerhalb des Muskelgewebes wird zwischen intermuskulärem und
intramuskulärem Fett differenziert. Dabei wird dem intramuskulären Fettgehalt (IMF)
ein wesentlicher Beitrag für den Geschmack und die Zartheit von Schweinefleisch
zugemessen. Mit durchschnittlichen Werten zwischen 0,3 und 0,5 verfügt der
intramuskuläre Fettgehalt über eine hohe Heritabilität und ist in Kombination mit einer

Diskussion
254
negativen genetischen Beziehung zum Fleischanteil im Schlachtkörper (rg ~ -0,2)
positiv mit der Rückenspeckdicke (rg ~ 0,3) korreliert. Eine weitere züchterische
Reduktion der Rückenspeckdicke zieht folglich eine unerwünschte Verminderung des
intramuskulären Fettgehaltes nach sich (SCHWÖRER et al, 1987, SCHWÖRER und
REBSAMEN, 1990, HOVENIER et al., 1993, DE VRIES UND VAN DER WAL, 1993, DE
VRIES et al., 1994, HERMESCH et al., 2000a,b).
BRASCAMP et al. (1995) erwähnen jedoch, dass das von JANSS et al. (1994) in
Meishan statistisch identifizierte „Hauptgen“ für das intramuskuläre Fett den
Gesamtkörperfettgehalt nicht kontrolliert. Folglich wäre es möglich, den
intramuskulären Fettgehalt „unabhängig“ vom Rückenspeck in Abhängigkeit vom
Leistungsniveau zu erhöhen, worauf bereits SCHWÖRER u. a. (1987) bzw. WARRISS et al.
(1990) hinwiesen. In einer späteren Untersuchung bestimmte dieselbe Arbeitsgruppe
getrennt für intramuskuläres Fett bzw. Rückenspeckdicke den Effekt eines einzelnen
rezessiven Allels, welches wahrscheinlich von der Rasse Meishan stammt und in den
„westlichen“ Gründerfamilien (founder lines) nicht vorhanden ist (JANSS ET al., 1997).
Dieses Beispiel kennzeichnet die Schwierigkeiten, die mit der Suche von so genannten
“Economic Trait Loci - ETL“ oder “Quantitative Trait Loci - QTL“ zur Nutzung in
markergestützten Selektionsprogrammen verbunden sind. Polymorphismen, die in einer
Population (Familie) gefunden werden, sind in anderen Populationen nicht unbedingt
informativ (SCHWERIN, 1994, MACKAY, 2001). Weitere begrenzende Faktoren ergeben
sich aus dem Rekombinationsverlust und der genetischen Drift über Generationen, der
bekannten Komplexität der Genwirkungen, den benötigten statistischen
Computerverfahren sowie aus der gleichzeitigen Selektion auf mehrere ökonomisch
wichtige Merkmale, die biologisch konkurrieren (SMITH und SMITH, 1993, MONTALDO
and MEZA-HERRERA, 1998, FRIES, 2000).
Ein weiteres Kandidatengen für die Variation im intramuskulären Fett ist das in
Fettzellen auftretende und für den intrazellulären Fettsäuretransport mitverantwortliche
Fettsäure-Bindungs-Protein-Gen (ADIPOCYTE FATTY ACID-BINDING PROTEIN: A-FABP) auf
dem Chromosom 4 des Schweines (GERBENS et al., 1998). Die Autoren berichten über
einen signifikanten Kontrast von ca. 1% intramuskulärem Fettgehalt zwischen den
unterschiedlichen Genotypklassen, ohne jedoch einen Effekt auf Rückenspeckdicke und
Dripverlust (Wasserbindungsvermögen) in einer Duroc-Population feststellen zu
können. In nachfolgenden Untersuchungen innerhalb verschiedener Kreuzungslinien
ohne Duroc-Genanteile konnte jedoch keine eindeutige Assoziation zwischen A-FABP-

Diskussion
255
Genotypen und dem intramuskulären Fettgehalt nachgewiesen werden (GERBENS et al.,
2000, 2001). Das A-FABP- (FABP4) oder ADIPOCYTE LIPID-BINDING PROTEIN-Gen (AP2,
ALBP) wird hauptsächlich im Verlauf der Fettgewebe-Konversion reguliert, wenn sich
die Zellen von fibroblastischen Vorläuferzellen zu ausgereiften Adipozyten
differenzieren. Verschiedene Proteine wie Glucokortikoide, CCAAT/ENHANCER-BINDING
PROTEIN α (C/EBPα), ADIPOCYTE DETERMINATION UND DIFFERENTIATION-DEPENDENT
FACTOR 1 (ADD1), PEROXISOME PROLIFERATOR-ACTIVATED RECEPTOR α (PPARα) und
PEROXISOME PROLIFERATOR-ACTIVATED RECEPTOR γ1 und 2 (PPARγ1, PPARγ2) sowie
HORMONSENSITIVE LIPASE (HSL) sind an den Differenzierungsprozessen beteiligt
(BERNLOHR et al., 1997, DING et al., 2000, ROBINSON et al., 2000, VOGEL HERTZEL und
BERNLOHR, 2000). Speziell für die Gene, die ADD1, PPARα und
PPARγ1 bzw. PPARγ2 sowie ACYL-COA-OXIDASE (ACO) regulieren, konnten keine
Genotypunterschiede beim Vergleich von Schweinen mit Duroc- bzw. Newsham-
Abstammung nachgewiesen werden. Allein in der mRNA-Konzentration von FAS
(FATTY ACID SYNTHASE) im Fettgewebe bestehen tendenzielle Unterschiede zwischen
genetisch „fetten“ Schweinen mit Duroc-Abstammung und genetisch „mageren“
Schweinen mit Newsham-Abstammung. Schweine mit Duroc-Abstammung weisen eine
höhere mRNA-Konzentration für FAS auf (DING et al., 2000).
Das HEARTY FATTY ACID-BINDING PROTEIN-Gen (h-FABP oder FABP3) ist ebenfalls
von Interesse für die Steuerung der Körperzusammensetzung und speziell des
intramuskulären Fettgehaltes, da es neben der Herzmuskulatur auch im Skelettmuskel
vorkommt (VOGEL HERTZEL und BERNLOHR, 2000). Bislang konnte jedoch beim
Schwein nur ein signifikanter Effekt von h-FABP auf die Tageszunahme gefunden
werden (GERBENS et al., 1997, VAN ERP et al., 1997), der möglicherweise durch die
unterschiedliche Testosteron-Konzentration in männlichen und weiblichen Tieren
moduliert wird (VOGEL HERTZEL und BERNLOHR, 2000, GERBENS et al., 2001). Ein
Effekt auf den intramuskulären Fettgehalt wird jedoch nicht ausgeschlossen (GERBENS
et al., 2000, 2001).
Eine besondere Rolle an der Oberfläche der Muskel- und Fettzellen spielt das
INSULIN-SENSITIVE GLUKOSETRANSPORT-PROTEIN 4 (GLUT4) bzw. sein kodierendes Gen.
Homozygote GLUT4-knockout Mäuse zeigten vielfache Abnormalitäten, die von stark
vermindertem Wachstum über extrem verminderten Fettansatz bis zu einer durch eine
Herzhypertrophie verursachten kürzeren Lebensdauer reichten (OLSON und PESSIN,
1996). Neben diesen Wirkungen unterstützt Glukose über den Weg zu Glukose-6-

Diskussion
256
Phosphat die Kontrolle der Transkription von Genen für lipogene und glykolytische
Enzyme in Fettgewebe, Leber und Bauchspeicheldrüse (GIRARD et al., 1997).
Neben dem bereits beschriebenen LEPTIN-GEN und dem LEPTIN-REZEPTOR-Gen
scheint eine wachsende Liste von Genen, die diverse Peptid-Hormone bzw. ihre
Rezeptoren regulieren (z.B. CHOLECYSTOKININ-A, GLUKAGON-ÄHNLICHE PEPTIDE 1,
NEUROPEPTID Y, α-MELANOZYTEN-STIMULIERENDES HORMON, CORTICOTROPIN-RELEASING
HORMON, PRO-OPIOMELANOCORTIN, COCAINE- UND AMPHETAMINE-REGULATED
TRANSCRIPT, OREXIN A, OREXIN B), die kurzzeitige Futteraufnahme zu modulieren
(NORTH, 1999, WILSON et al., 1999, AHIMA und FLIER, 2000, Abb. 4.2.6). Sie sind
jedoch nicht essentiell für die Aufrechterhaltung des Körpergewichts (KOPIN et al.,
1999). Das „Futter“-Rezeptor-Gen des Schweines: NEUROPEPTID Y REZEPTOR 5 (NPY5)
befindet sich auf dem Chromosom 8 (8p11 - TORNSTEN et al., 1998). Auf diesem
porcinen Chromosom häufen sich mit dem CHOLECYSTOKININ-A-REZEPTOR-GEN
(CLUTTER et al., 1998) und dem CARBOXYPETIDASE-E-GEN (CPE, CARGILL et al., 1998)
weitere Gene, die an der Regulation der Futteraufnahme und der
Körperzusammensetzung beteiligt sind. Im Gegensatz befindet sich das PRO-
OPIOMELANOCORTIN-Gen auf dem Chromosom 3 des Schweines (MESSER et al., 2001).
Mutationen im TUBBY-Gen (TUB) könnten einen weiteren Faktor darstellen
(NOBEN-TRAUTH et al., 1996, KLEYN et al., 1996). Homozygote Mäuse mit einer G-zu-
T-Transversion an einer Donor-Spleiß-Position, die das Spleißen des Carboxyl-
Terminal-Introns verhindert, haben ein längeres Transkript als „normale“ Mäuse. Diese
Mäuse sind mager und aktiv nach der Geburt, verfetten jedoch mit zunehmendem Alter.
Die Alterung ist mit Diabetes sowie zunehmender Seh- und Gehörschwäche gekoppelt -
medizinischen Zuständen, die in engem Zusammenhang zur Fettleibigkeit stehen.

Diskussion
257
Abb. 4.2.6: Beziehungen zwischen Fettgewebe, genetischer Steuerung von Zentren imHypothalamus und der Futteraufnahmeregulation (eigene Abbildung modifiziert ausMITCHELL et al., 2001a)
Andere genetische Defekte, die mit Diabetes und Fettwüchsigkeit bei der Maus
verbunden sind, erzeugen weit dramatischere Effekte wie extreme Verfettung in sehr
frühen Altersstadien. Das Hauptinteresse bei der TUBBY-Maus liegt jedoch in der
zunehmenden Blind- und Taubheit und weniger in ihrer Diabetes bzw.
Gewichtszunahme. Die Zellen der Retina sterben bei diesen Mäusen nach dem so
genannten Prozess des „programmierten Zelltodes“ ab. Ein noch unbekannter Auslöser
aktiviert in den Zellen einen eingebauten „Suizidschalter“. Der Auslöser könnte eine
abnormale Zunahme von zyklischem GMP sein, welches normalerweise nur in sehr
geringen Konzentrationen vorkommt. Die Konzentration wird durch das Enzym
zyklische GMP-Phosphodiesterase niedrig gehalten. Nach NOBEN-TRAUTH et al. (1996)
enthält das TUBBY-Gen die Instruktionen für die Produktion einer Phosphodiesterase.
Zwischen dem Genom von Schwein bzw. Mensch und Maus existieren
weitreichende Kopplungskonservationen mit einer variierenden Neuanordnung der
Genreihenfolge innerhalb homologer Chromosomensegmente (Tab. A15 im Anhang V).
Die durchschnittliche Länge der konservierten Chromosomenabschnitte zwischen
Schwein und Mensch wurden mit 37 cM und zwischen Schwein und Maus mit 21 cM
geschätzt (JOHANSSON et al., 1995).
Da die Merkmale der Körperzusammensetzung und des Wachstums beim
Schwein polygen gesteuert werden, ist es nicht überraschend, eine derartig große
Fettgewebe
Leptin
ZNS (Hypothalamus)Leptin-Rezeptor
Neuropeptid Y'Glukagon-like' Peptid 1
GLP-1-Rezeptor
'Tubby'-Mutation
Cholecystokinin-Peptidase
Cholecystokinin
Gastrointestinaltrakt
Füllungszustand
Futteraufnahme
+ -
Energieverbrauch+ -
+ -
(Energiespeicher)
-
- α-Melanozyten-stimulierendes Hormon
BombesinApolipoprotein B, E4, A4
Nervus vagus
Vegetatives
Serotonin
Serotonin-Rezeptor 2c
Nervensystem
?
(Thermoregulation)
(kein Rezeptor in db/db Mäusen)
(gesteigertes Niveau in ob/ob Mäusen)
Wachstumshormon
+ -
(Hunger) +
( 'yellow'-Allel am 'agouti'- Locus)
Melanocortin-4-Rezeptor-
Proopiomelanocortin

Diskussion
258
Anzahl von Genen oder Markern zu finden, die mit Variationen im Körperfett- bzw.
Magergewebegehalt und Veränderungen im Körpergewicht assoziiert sind. Bis zum
Ende des Jahres 1996 entdeckten verschiedene Forschungsgruppen 13 Genorte, die 10
„Fettwuchs“-Syndrome beim Menschen beschrieben, fünf Einzelgen-Mutationen bei
Nagetieren (LEPR, LEP, AGOUT, TUBBY, FAT) und 25 QTL in Beziehung zu Wachstum
oder Körperzusammensetzung bei Maus, Ratte und Schwein. Weitere
Kandidatengenorte schließen die Gene ein, die das Wachstumshormon, die Insulin-
ähnlichen Wachstumsfaktoren, ihre Rezeptoren und Bindungsproteine kodieren
(BROCKMANN et al., 1998). Die Anzahl der lokalisierten Gene auf den Chromosomen
von Mensch, Schwein und Maus wächst ständig (MACKAY, 2001). Nachdem FÖRSTER
(1991 a,b) noch feststellte, dass die Zahl der mit gentechnischen Methoden
identifizierten Gene (für allgemeine und spezielle Produktionsleistungen sowie für
Produktqualität – GLODEK, 1991) beim Schwein weit unter 100 lag (1989: 40 Gene),
berichtet ROTHSCHILD (2001) von über 4000 Marker- und Kandidatengenorten auf der
„Genkarte“ des Schweines. Allein die Arbeitsgruppe um BROCKMANN et al. (1998)
identifizierte bei der Untersuchung von 13 Maus-Chromosomen 31 QTL mit Effekten
auf Körpergewicht, abdominales Fettgewicht und auf die Gewichte von Leber, Niere
und Milz, die mindestens signifikant auf dem Vorschlagsniveau (p < 0,05) waren.
Verschiedene dieser QTL befinden sich in denselben chromosomalen Regionen, die
durch andere Gruppen identifiziert wurden und könnten tatsächlich identische Gene
betreffen (FISLER und WARDEN, 1997, MACKAY, 2001). POMP (1999) weist bereits auf
50 chromosomale Regionen hin, die Gene zur potentiellen Steuerung der
Körperzusammensetzung beherbergen. Dabei können diese Gene in Abhängigkeit vom
Geschlecht und multiplen Genotyp (Rasse, Linie) unterschiedliche Effekte (dominant
bzw. rezessiv, additiv, epistatisch oder pleiotrop) unter variierenden
Umweltbedingungen (z.B. unterschiedlicher Ernährung oder Haltung) hervorrufen
(SPURLOCK et al., 1998, MACKAY, 2001). Die Identifikation dieser Gene und
genetischer Polymorphismen wird in Kombination mit modernen In-vivo-
Analyseverfahren, wie z.B. MRS, MRT, CT, DXA und NAA zur Aufklärung der
komplexen Biologie des Schweines (bzw. der landwirtschaftlicher Nutztiere sowie der
Haus- und Heimtiere) beitragen. Genauere Methoden zur Messung der phänotypischen
Variation im Einklang mit verfeinerten statistischen Methoden zur Zuchtwertschätzung
und markergestützten Selektion können schließlich helfen, den züchterischen Fortschritt
im Sinne von Mensch und Tier voran zu bringen. Obwohl die komplette Elimination

Diskussion
259
des Defektallels am RyR1-Genort neben der verbesserten Fleischqualität und
Belastungsstabilität eine erhebliche Verminderung des Fleischanteils (~ 1 – 7 %) und
damit eine zum Teil unerwünschte Erhöhung des Fettanteils im Schlachtkörper zur
Folge hat (siehe eigene Ergebnisse, SCHOLZ und HARDGE, 1994, WITTMANN u.a., 1998,
BIEDERMANN u.a., 2000, THALLER u.a., 2000, IRGANG, 2001), sollte es möglich sein -
unter Verwendung der umfassenden Informationen über die genetische Steuerung der
Körperzusammensetzung und des Wachstums - weiterhin Tiere zu züchten, die den
vielfältigen Wünschen der Verbraucher gerecht werden. Erste Selektionsergebnisse
innerhalb einer homozygot normalen Pietrain-Eberpopulation zeigen, dass stressstabile
Tiere bei verminderten Nachteilen im Fleischanteil auch überlegene Mastleistungen
erzielen können (WITTMANN u.a., 1999).
Moderne In-vivo-Messverfahren (MRT, MRS, CT, DXA, US) sind nötig, um die
exakte Wirkungsweise neu entschlüsselter genetischer Polymorphismen in den
verschiedensten Zuchtpopulationen aufzuklären. Außerdem werden auch die zur Zeit
noch (sehr) teueren Verfahren wie CT, DXA und möglicherweise MRT zunehmend für
die Leistungsprüfung von Zuchttieren Verwendung finden, um einerseits die regional
sehr variierende Gewebezusammensetzung verstärkt in die Züchtung einbeziehen zu
können und um andererseits durch eine frühere Leistungsinformation das
Generationsintervall in der Selektion zu verkürzen.

Zusammenfassung
260
5 Zusammenfassung
Schweine mit unterschiedlichen Ryanodin-Rezeptor-1-Genotypen (RyR1) dienen als
Modell für die Evaluierung verschiedener Methoden zur In-vivo-Messung von
Muskelenergiestoffwechsel und Körperzusammensetzung. Im Mittelpunkt der
Untersuchungen stehen 31P- und 13C-Magnetresonanz-Spektroskopie bzw.
Magnetresonanz–Tomographie sowie Dualenergie-Röntgenabsorptiometrie. Zusätzlich
zur Referenzschlachtung und chemischen Analyse werden diese Methoden im
Vergleich zu anderen Verfahren einer ausführlichen Bewertung in der
Methodenübersicht unterzogen. Die Muskel-Schussbiopsie ergänzt diese
Untersuchungen zur Ermittlung der Muskelfaserzusammensetzung. Insgesamt gehen
111 Versuchstiere in die Auswertung ein (Tab. 5.1).
Tab. 5.1: Anzahl Tiere innerhalb der RyR1-Genotypen – Gesamtmaterial
NN Nn nn31P-MR-Spektroskopie *
n (n - post mortem) 24 (3) 29 (4) 16 (3)13C-MR-Spektroskopie *
n (n - post mortem) 7 (1) 10 (2) 10 (3)1H-MR-Tomographie #
n 37 46 28
Muskelbiopsie +
n 7 10 5
Herkünfte: [US-Landrasse - LR], [Poland China x LR], [≥50 % Duroc - Du], [≥50 % Hampshire – Ha],[≥50 % Spotted –S-], [25 % Du, 25 % Ha, 25 % LR, 12,5 % S, 12,5 % Yorkshire] und [Du x Minzhu]§
* Lebendmasse (LM): 6 - 20 kg; #LM-Gruppen: 10-30-60-90 kg; +LM: 60-90 kg.
Dualenergie-Röntgenabsorptiometrie #
n 29 36 27
Referenzschlachtung und chemische Analyse#
n 20 27 16
§ Du x Minzhu wurden zusätzlich allein für die 1H-MR-Tomographie verwendet.
13C- bzw. 31P-MR-Spektroskopie ermöglichen in sehr geeigneter Weise in vivo bzw.
post mortem nicht invasiv und kontinuierlich Veränderungen von Glykogen, Kreatin
bzw. Phosphokreatin (PCr), anorganischem Phosphat (Pi), Adenosintriphosphat (ATP)
und pH-Wert direkt in relativen oder absoluten Konzentrationsmengen über eine
“unbegrenzte” Zeit zu messen. Die 31P-MR-Spektroskopie besitzt dabei im Vergleich
zur 13C-MR-Spektroskopie Vorteile in der Sensitivität und zeitlichen Auflösung für die

Zusammenfassung
261
Messung von Konzentrationsveränderungen der Muskelstoffwechselkomponenten.
Während sich 31P-MR-Spektren zur Bestimmung der Konzentrationsveränderungen von
PCr, ATP, Pi und pH-Wert in einem zeitlichen Abstand von < 1 Minute zuverlässig in
vivo erfassen lassen, ist für eine sinnvolle Interpretation von 13C-MR-Spektren mit dem
Ziel der Glykogen- und Kreatinbestimmung ein Zeitraum von > 5 Minuten erforderlich.
Fortschritte in den Techniken zur Analyse der Körperzusammensetzung basieren
in der Regel auf elektronischen, computergesteuerten Methoden, um eine möglichst
zerstörungsfreie, schnelle und objektive Untersuchung gewährleisten zu können. Die
Auswahl der Methodik richtet sich nach Zielstellung (Forschung, Leistungsprüfung,
Produktion oder Diagnose) und finanziellen Mitteln für die jeweilige Studie bzw.
klinische Anwendung. Technische Details in Bezug auf Genauigkeit, Präzision sowie
Parameterspektrum und maximale/minimale Probandengröße spielen dabei ebenso eine
Rolle wie eine einfache, robuste und umwelt- bzw. probandenschonende Handhabung.
Höchste Genauigkeiten für Ganzkörperuntersuchungen können innerhalb der
bildgebenden Verfahren mit Magnetresonanz- und Computer-Tomographie gefolgt von
Dualenergie-Röntgenabsorptiometrie (DXA) erzielt werden. Die DXA-Methodik
besticht in der einfachen Handhabung und Datenanalyse sowohl für
Ganzkörperuntersuchungen als auch für ausgewählte Körperregionen. Zusätzlich ist
DXA neben der quantitativen Computer-Tomographie die einzige nicht invasive
Methode, die direkte Maße zur Knochenmineraldichte liefert. Entsprechend der
Literaturauswertung ist im Rahmen von Gerätekalibrierungen die
Neutronenaktivierungs-Analyse als „nicht invasive“ Methode einer chemischen
Analyse bzw. geweblichen Totalzerlegung vorzuziehen. Aufgrund der sehr begrenzten
Verfügbarkeit dieser Methodik sind die chemische Analyse oder die grobgewebliche
Zerlegung bis auf weiteres als Standardreferenzmethoden erforderlich. Die
Fehlerquellen der nicht invasiven (bildgebenden) Methoden sind jedoch geringfügiger
als die möglichen Fehler bei der grobgeweblichen Zerlegung.
Die Hauptvorteile der nicht invasiven Bildgebungs- oder/und spektroskopischen
Verfahren in vivo liegen:
1. in leicht standardisierbaren und wiederholbaren Messungen (Wiederholbarkeit
> 75 %),
2. in der Möglichkeit von separaten Analysen großer interessierender Volumina des
gesamten Körpers, von Körpergeweben und Körperteilen,

Zusammenfassung
262
3. in der Möglichkeit kontinuierliche Messungen über einen „unbegrenzten“ Zeitraum
vorzunehmen sowie
4. in der Zerstörungs- bzw. Verletzungsfreiheit der Tiere und hoher hygienischer
Zuverlässigkeit.
Magnetresonanz, Ultraschall und digitale Bildgebung besitzen einen Vorteil gegenüber
den Röntgentechniken, da sie ohne ionisierende Strahlung auskommen, während sich
speziell die Spiral-Computer-Tomographie durch sehr hohe Untersuchungs-
geschwindigkeiten auszeichnet.
Der Muskelstoffwechsel der Defektallelträger am RyR1-Genort Nn bzw. nn zeigt
bereits in vivo stressbedingte Abweichungen im Vergleich zum normalen Genotyp NN.
Nach Stressauslösung mittels Halothan reagieren die Defektallelgenotypen mit einem
starken Abfall im Glykogen-, Phosphokreatin-, ATP- und pH-Niveau unter
gleichzeitigem Anstieg im Niveau von anorganischem Phosphat und der
Körpertemperatur. Die homozygoten Defektallelträger weisen im Durchschnitt eine
stärkere Stoffwechselentgleisung als die heterozygoten Defektallelträger auf. Post
mortem sind die Differenzen zwischen den Genotypen noch stärker als in vivo. Die
Stoffwechselentgleisungen gehen einher mit einer Muskelfaserhypertrophie bei beiden
Defektallelgenotypen. Im Laufe des Wachstums setzen die homozygoten
Defektallelträger mehr Muskelmasse (-volumen) und weniger Fett als die heterozygoten
Defektallelträger an. Bereits bei ca. 10 kg Lebendmasse kann im homozygoten
Defektallelgenotyp ein größeres Muskelvolumen mittels 1H-Magnetresonanz-
Tomographie nachgewiesen werden als in den beiden anderen Genotypen. Neben dem
untersuchten Effekt des RyR1-Genotyps (und des vor allem in der Rasse Hampshire
vorkommenden RN—-Allels) beeinflussen erwartungsgemäß eine Vielzahl von
genetischen, morphologischen und Umweltfaktoren den Muskelstoffwechsel und die
Körperzusammensetzung beim Schwein, die ausführlich im Diskussionsteil beschrieben
werden. Das RN—-Allel bewirkt einen verlangsamten Glykogenabbau in vivo (und post
mortem). Parallel weist die untersuchte Hampshire-Line (≥ 50 % Hampshire-Genanteil)
neben der US-Landrasse und der synthetischen Linie „Duroc x Hampshire x US-
Landrasse x Spotted x Yorkshire“ im Vergleich zu Duroc, „Poland China x Landrasse“,
Spotted und „Duroc x Minzhu“ einen niedrigen Körperfettgehalt auf. Spotted und
„Duroc x Minzhu“ sind am stärksten verfettet. Unerwartet reagiert Spotted (alle Nn) am
stärksten auf die Stressauslösung mittels Halothan.

Summary
263
6 Summary
In vivo techniques for the analysis of muscle metabolism and body composition inpigs of different genotypes
Pigs of different ryanodine receptor 1 (RyR1) genotypes serve as a model for the
evaluation of various non-invasive in vivo methods to measure muscle energy
metabolism and body composition. The main focus is set on one hand on 31P and 13C
nuclear magnetic resonance spectroscopy and on the other hand on nuclear magnetic
resonance imaging and dual energy X-ray absorptiometry. In addition to a reference
dissection and chemical analysis, the above mentioned methods are being extensively
compared to other methods in the methodology part. An invasive muscle shot biopsy
serves as an additional method in order to determine the muscle fiber composition of the
longissimus dorsi muscle. Totally, 111 animals have been considered in the data
analysis (Tab. 5.1a).
Tab. 5.1a: Number of animals within ryanodine receptor 1 (RyR1) genotype – total
material
NN Nn nn31P-MR-Spectroscopy *
n (n - post mortem) 24 (3) 29 (4) 16 (3)13C-MR-Spectroscopy *
n (n - post mortem) 7 (1) 10 (2) 10 (3)1H-MR-Imaging #
n 37 46 28
Muscle Biopsy+
n 7 10 5
breeds: [US-Landrace - LR], [Poland China x LR, [≥50 % Duroc – Du], [≥50 % Hampshire - Ha],[≥50 % Spotted – S], [25 % Du, 25 % Ha, 25 % LR, 12,5 % S, 12,5 % Yorkshire] and [Du x Minzhu]§
* live body weight (BW): 6 - 20 kg; #BW groups: 10-30-60-90 kg; +BW: 60-90 kg
Dual Energy X-Ray Absorptiometry #
n 29 36 27
Reference Dissection and Chemical Analysis#
n 20 27 16
§ Du x Minzhu were used alone in addition to all other probands for MR Imaging
13C- and 31P NMR spectroscopy are very appropriate methods to measure in vivo or post
mortem, non-invasively and continuously changes in the concentration of glycogen and
creatine or phosphocreatine (PCr), inorganic phosphate (Pi), adenosintriphosphate
(ATP), and pH-value directly in relative or absolute amounts over an „unlimited“ time

Summary
264
period. 31P NMR spectroscopy compared to 13C NMR spectroscopy has advantages in
the sensitivity and timely resolution for measuring changes in the concentration within
the components of muscle metabolism. While 31P NMR spectra in vivo yield sufficient
results for measuring changes of PCr, ATP, Pi and pH value within a time interval < 1
minute, it takes > 5 minutes to acquire reasonable 13C NMR spectra in order to measure
changes in the concentration of glycogen and creatine.
Progress in the techniques of body composition analysis is mainly based on
electronic and computer driven methods in order to provide non-invasive, fast and
objective measurements. The selection of the appropriate method depends on the
objective (research, performance testing, production, or diagnosis) and financial budget
for each single study and clinical application. Technical details considering accuracy,
precision, parameter selection and maximum/minimum size of an individual affect the
decision as well as does a simple, robust, environment-safe, patient-comfortable and
risk-free handling of the technique.
The highest accuracy for whole body studies within the imaging methods provide
magnetic resonance imaging (MRI) and computer tomography (CT) followed by dual
energy x-ray absorptiometry (DXA). DXA, however, outperforms MRI and CT due to
its simple handling and easy data analysis for whole body or regional body composition
studies. In addition, DXA is beside the Quantitative CT the only non-invasive method
which provides direct bone mineral density measurements.
According to the literature survey, the neutron activation analysis as a non-
invasive method should be preferred for the calibration of body composition
measurement devices instead of chemical analysis or dissection. Chemical analysis and
total dissection will remain standard reference methods as long as neutron activation
facilities are available only on a very limited basis. However, the non-invasive
(imaging) methods are less prone to error sources than are the diverse dissection or
chemical analysis procedures.
The main advantages of the non-invasive imaging and/or spectroscopic methods
in vivo are:
1. the easy way to standardize the methods with a high repeatability (>75 %),
2. the opportunity of analyzing separately large volumes of interest of the whole
body, body tissues and/or body parts,
3. the opportunity of performing continuous measurements over an “unlimited”
period of time, and

Summary
265
4. the harm “free” animal/patient precautions with a high degree of hygienic
safety.
An advantage for magnetic resonance, ultrasound and digital imaging is provided
by the function without radiation, while especially the “spiral computer tomography” is
characterized by a very high speed of analysis.
The muscle metabolism of the defect allele carriers at the RyR1 locus Nn and nn
shows already in vivo stress associated deviations in comparison to the “normal”
genotype NN. After generating muscle stress by halothane, defect genotypes react with
a decline in the glycogen, phosphocreatine, ATP, and pH level. Parallel inorganic
phosphate and body temperature increase significantly. In the average show the
homozygous defect allele carriers a stronger metabolic distress than do the
heterozygous carriers. Differences among genotypes are higher post mortem compared
to the metabolism in vivo. The muscle metabolic distress is connected with a muscle
fiber hypertrophy in both defect allele genotypes. During growth, the homozygous
defect allele genotype deposits more muscle mass (volume) and less fat than does the
heterozygous genotype followed by the homozygous normal genotype.
Already at a live body weight of about 10 kg, magnetic resonance imaging
provides evidence for a larger muscle volume in the homozygous defect genotype in
comparison to the two other genotypes. Beside the studied RyR1 genotypes (and the
RN—-allel especially present in Hampshire), a large number of genetic, morphologic
and environmental factors influence muscle metabolism and body composition in Swine
-- as described in the discussion part. The RN—-allel causes a reduced glycogen
depletion in vivo (and post mortem). Corresponding, the Hampshire line (≥ 50 %
Hampshire genes) is beside the US-Landrace and the synthetic line “Duroc x Hampshire
x US-Landrace x Spotted x Yorkshire” less obese than are the lines Duroc, “Poland
China x Landrace”, Spotted, and “Duroc x Minzhu”. Spotted and “Duroc x Minzhu” are
the most obese lines. In addition comparing all lines, Spotted (all Nn) responses --
unexpectedly -- most severely to (muscle) stress caused by halothane administration.

Literaturverzeichnis
7 Literaturverzeichnis Abate, N., D. Burns, R.M. Peshock, A. Garg, S.M. Grundy (1994): Estimation of adipose tissue mass by magnetic resonance imaging: validation against dissection in human cadavers. J. Lipid Res. 35: 1490-1496.
Abe, T., F. Tanaka, Y. Kawakami, K. Yoshikawa, T. Fukunaga (1996): Total and segmental subcutaneous adipose tissue volume measured by ultrasound. Med. Sci. Sports Exerc. 28: 908-912.
Abeles, R.H., P.A. Frey, W.P. Jenks (1992): Biochemestry. Jones and Bartlett Publishers, Boston, London.
Abrams, S.A., R.J. Schanler, H.P. Sheng, H.J. Evans, A.D. Leblanc, and C. Garza (1988): Bone mineral content reflects total body calcium in neonatal miniature piglets. Pediatr. Res. 24: 693-695.
Affentranger, P., C. Gerwig, G. Seewer, N. Künzi, D. Schwörer, A.L. Prabucki (1991): Comparison of fattening and carcass performance as well as meat and fat quality of crossbred genotypes fed with different feeding intensities. 42nd Annual Meeting of the EAAP, Berlin, 9-12 September, Paper Copy, pp. 8.
Afonso, J.J.M. (1992): Changes in body composition in sheep during growth, pregnancy and lactation. PhD Thesis, University of New England, Australia.
Afonso, J. and J.M. Thompson (1996): Fat distribution in sheep selected for/ against backfat depth, during growth on ad libitum feeding. Livest. Prod. Sci. 46: 97-106.
Ahima, R.S. and J.S. Flier (2000): Adipose Tissue as an Endocrine Organ. Trends in Endocrinology and Metabolism 11: 327-332.
Allen, P. and O. Vangen (1983): X-ray Tomography of pigs some preliminary results. CEC Meeting: In Vivo Measurement of Body Composition in Meat Animals. Bristol, UK, November 30-December 1.
Allen, P. and K.A. Leymaster (1985): Machine error in x-ray computer tomography and its relevance to prediction of in vivo body composition. Livest. Prod. Sci. 13: 383-398.
Allen, P. and M.G. Keane (1996): Prediction of beef pistolsa hind composition by TOBEC. 47th Annual Meeting of EAAP, Lillehammer, Norway, Book of Abstracts No. 2: 280.
Allen, P. and P. Walstra (1996): Non-invasive techniques for carcass assessment - grading applications. Paper, 47th Annual Meeting of EAAP, Lillehammer, Norway, pp. 18.
Aloia, J.F., A. Vaswani, E. Flaster, and R. Ma (1998): Relationship of body water compartments to age, race, and fat-free mass. J. Lab. Clin. Med. 132: 483-490.
266

Literaturverzeichnis
Altmann, M., G.von Lengerken, H. Pfeiffer (1992): Intramuskulärer Fettgehalt - Bedeutung, Ermittlung und Selektionsmaßnahmen in der Schweinezucht. Schweinezucht und Schweinemast 40: 252 – 254.
ANALYZE 7.01 (1991): Biomedical Imaging Resource, Mayo Foundation, 200 First Street SW, Rochester, MN 55905.
Anderson, Th. (1860): Elements of agricultural chemistry. Adam and Charles Black, Edinburgh, GB. Albert R. Mann Library. 2000. Core Historical Literature of Agriculture (CHLA). Ithaca, NY: Albert R. Mann Library, Cornell University. http://chla.library.cornell.edu (Version October 2000). http://cdl.library.cornell.edu/cgi-bin/chla/chla-cgi?notisid=AAM3671
Andersson, L., C.S. Haley, H. Ellegren, S.A. Knott, M. Johansson, K. Andersson, L. Andersson-Eklund, I. Edfors-Lilja, M. Fredholm, I. Hansson, J. Håkansson and K. Lundström (1994): Genetic mapping of quantitative trait loci for growth and fatness in pigs. Science 263: 1771-1774.
Andersson, L., L: Marklund, P.-E. Nyström and S. Stern (1996): Identification of a major QTL for fatness on pig chromosome 4. 47th Annual Meeting of EAAP, Lillehammer, Norway, p. 37.
Andersson-Eklund, L., L. Marklund, K. Lundström, C.S. Haley, K. Andersson, I. Hansson, M. Moller, and L. Andersson (1998): Mapping Quantitative Trait Loci for Carcass and Meat Quality traits in a Wild Boar x Large White Intercross. J. Anim. Sci. 76: 694-700.
Andrew, R. (1998): The Impact of NMR and MRI. In: Making the human body transparent: The impact of nuclear magnetic resonance and magnetic resonance imaging. The transcript of a Witness Seminar held at the Wellcome Institute for the History of Medicine, London, on 2 July 1996, (Edited by D.A. Christie and E.M. Tansey). In: Making the human body transparent: The impact of nuclear magnetic resonance and magnetic resonance imaging. – Research in general practice. – Drugs in psychiatric practice. – The MRC common cold unit. Witness Seminar Transcripts (Edited by: E.M. Tansey, D.A. Christie, L.A. Reynolds), Volume 2: 4-13.
AOAC (1984): Methods of Analysis. 14th ed. Association of Official Analytical Chemists, Washington, DC.
Arnold, D.L., D.J. Taylor, and G.K. Radda (1985): Investigations of human mitochondrial myopathies by phosphorous magnetic resonance spectroscopy. Ann. Neurol. 18: 189-196.
Atkinson, G. and A.M. Nevill (1998): Statistical methods for assessing measurement error (reliability) in variables relevant to sports medicine. Sports Med. 26: 217-238.
Avison, M.J., H.P. Hetherington, R.G. Shulman (1986): Applications of NMR to studies of tissue metabolism. Ann. Rev. Biophys. Chem. 15: 377-402.
267

Literaturverzeichnis
Azzopardi, D. and D. Edwards (1995): Magnetic resonance spectroscopy in neonates. Current Opinion in Neurology 8: 145-149.
Azuma, Y., N. Manabe, F. Kawai, M. Kanamori, and H. Miyamato (1994): Phosphorous-31 nuclear magnetic resonance study of postmortem changes in the intact tissue of goat muscles. Anim. Sci. Technol. (Jpn.) 65: 416-422.
Babcock, D.F. and B. Hille (1998): Mitochondrial oversight of cellular Ca2+ signaling. Current Opinion in Neurobiology 8: 398-404.
Baerwalde, S.O. (2001): Der Einfluss einer simulierten Orthostase auf den Energiestoffwechsel des menschlichen Skelettmuskels. Dissertation, Universität Köln.
Bader, R. (1982): Enzymhistochemische und histometrische Untersuchungen an Skelettmusklen von ausgemästeten, gesunden Schweinen der deutschen Landrasse. Zbl. Vet. Med. A 29: 443-457.
Baldwin, R.L. and R.D. Sainz (1995): Energy Partitioning and Modeling in Animal Nutrition. Annu. Rev. Nutr. 15:191-211.
Ball, A.J., J.M. Thompson, and G.N. Hinch (1996): Seasonal oscillations in the mass of body components of mature ewes fed at a constant intake. Proc. Aust. Soc. Anim. Prod. Vol. 21: 479.
Baron, A.D. and M.G. Clark (1997): Role of blood flow in the regulation of muscle glucose uptake. Annu. Rev. Nutr. 17: 487-499.
Barth, N., A. Ziegler, G.W. Himmelmann, H. Coners, M. Wabitsch, K. Hennighausen, H. Mayer, H. Remschmidt, H. Schäfer, J. Hebebrand (1997): Significant weight gains in a clinical sample of obese children and adolescents between 1985 and 1995. Intl. J. Obesity 21: 122-126.
Baskin, L.C. and D. Pomp (1998): Rapid Communication: Mapping of the Porcine Colipase Gene to Chromosome 7 Using Linkage Analysis. J. Anim. Sci. 76: 1241-1242.
Bastianelli, D. and D. Sauvant (1997): Modelling the mechanism of pig growth. Livest. Prod. Sci. 51: 97-107.
Bauer, A., U. Bogdahn, A. Haase, R. Schlief (1998): Dreidimensionale echosignalverstärkte transkranielle Doppler-Ultraschalldiagnostik. Der Radiologe 38: 394-398.
Baulain, U. (1994a): Bestimmung der Körperzusammensetzung landwirtschaftlicher Nutztiere mit Hilfe der Röntgen-CT. Landbauforschung Völkenrode, Sonderheft 145: 110-118.
Baulain, U. (1994b): MR-Imaging zur Erfassung der Körperzusammensetzung in vivo. Landbauforschung Völkenrode, Sonderheft 145: 119-126.
268

Literaturverzeichnis
Baulain, U. (1996): Das Schwein in Scheiben - Kernspintomographie macht's möglich! Schweinezucht und Schweinemast 44/1: 42-44.
Baulain, U. (1997): Magnetic resonance imaging for the in vivo determination of body composition in animal science. Computers and Electronics in Agriculture 17: 189-203.
Baulain, U. and A. Scholz (1996): Image evaluation for different acquisition techniques in animal production. 47th Annual Meeting of the EAAP, Lillehammer, Norway, 25-29 August, Book of Abstracts No. 2, Wageningen Pers, p. 269 (PS5.5) and paper copy (4 pages).
Baulain, U. und E. Streitz (1997): Moderne Messmethode. Fleischanteil und Verfettung am lebenden Lamm messen. Deutsche Schafzucht 89: 451-453.
Baulain, U. und H. Henne (1999): Variation des Fleischanteils im Bauch von Schweinen aus Mutterlinien. Arch. Tierzucht (Dummerstorf) 42: 593-600.
Baulain, U. und M. Henning (2001): Untersuchungen zur Schlachtkörper- und Fleischqualität mit Hilfe von MR-Tomographie und MR-Spektroskopie. Arch. Tierz. (Dummerstorf) 44: 181-192.
Baulain,U., W. Griep, M. Henning, E. Hüster, E. Kallweit (1990): Schätzung der geweblichen Körperzusammensetzung mittels Magnet-Resonanz-Tomographie (MRT). Landbauforschung Völkenrode, 40. Jahrgang, Heft 3: 218 – 227.
Baulain, U., R.Schwerdtfeger, E.Kalm (1991): Composition of carcass cuts assessed by Magnetic Resonance Tomography (MRT). 42nd Annual Meeting of the EAAP, Berlin, September 8 - 12.
Baulain, U., A. Scholz, M. Henning, E. Hüster, E. Kallweit (1993): Composition of live pigs of different weight assessed by Magnetic Resonance Imaging (MRI). Proc. 44th Annual Meeting of EAAP, Aarhus, Denmark, 16-19 August.
Baulain, U., M. Henning, E. Kallweit (1996): Bestimmung der Körperzusammensetzung von Landrasse-Schweinen unterschiedlichen Alters mittels MRI. Arch. Tierzucht (Dummerstorf) 4: 431-440.
Baulain, U., M. Henning, E. Tholen, und W. Peschke (1998a): Der Schweinebauch in Scheiben. Schweinezucht und Schweinemast 46/4: 16-17.
Baulain, U., M. Henning, E. Tholen, W. Wittmann und W. Peschke (1998b): Objektive Erfassung des Fleischanteils im Schweinebauch. 2. Mitteilung: Verwendung von Bildinformationen aus dem MR-Imaging. Züchtungskunde 70: 205-212.
Baulain, U., P. Köhler, E. Kallweit, W. Brade (2000): Intramuscular fat content in some native German pig breeds. In: Quality of meat and fat in pigs as affected by genetics and nutrition (Eds.: C. Wenk, J.A. Fernández, and M. Dupuis). EAAP publication No. 100: 181-184.
Baumgartner, R.N, R. Ross, and St.B. Heymsfield (1998): Does adipose tissue influence bioelectric impedance in obese men and women? J. Appl. Physiol. 84: 257-262.
269

Literaturverzeichnis
Beckmann, N., J. Seelig, and H. Wick (1990): Analysis of glycogen storage disease by in vivo 13C NMR: comparison of normal volunteers with a patient. Magn. Reson. Med. 16: 150–160.
Bečková, R. and P. David (2000): Effect of the RYR 1 gene on meat quality in pigs of Large White, Landrace and Czech Meat Pig Breeds. In: Quality of meat and fat in pigs as affected by genetics and nutrition (Eds.: C. Wenk, J.A. Fernández, and M. Dupuis). EAAP publication No. 100: 151-155.
Beier, J., T. Büge, C. Stroszczynski, H. Oellinger, E. Fleck, R. Felix (1998a): 2D- und 3D-Parameterbilder zur Analyse der Kontrastmittelverteilung bei dynamischen CT- und MR-Untersuchungen. Der Radiologe 38: 832–840.
Beier, J., R.C. Bittner, N. Hosten, J. Tröger, und R. Felix (1998b): Morphologische Bildverarbeitungsoperatoren: Reduktion von Partialvolumenartefakten für eine verbesserte dreidimensionale Visualisierung von CT-Untersuchungen. Der Radiologe 38: 860-866.
Bejerholm, C. and P. Barton-Gade (1986): Effect of intramuscular fat level on eating quality of pig meat. Proceedings of the 32nd European Meeting of Meat Research Workers: 389-391.
Bendahan, D., Sylviane Confort-Gouny, Geneviève Kozak-Reiss, and P.J. Cozzone (1990): Pi trapping in glycogenolytic pathway can explain transient Pi disappearance during recovery from muscular exercise. FEBS Letters 269: 402-405.
Bentsen, H.B. and E. Sehested (1989): Computerised Tomography of Chickens. British Poultry Science 30: 575-589.
Berenyi, E., P. Bogner, A. Miseta, G. Köver, P. Horn, and D.N. Wheatley (1994): Pre-slaughter characteristics of proton NMR relaxation parameters in malignant hyperthermia susceptible pigs. Sciences des Aliments 14: 459-467.
Bereskin, B and N.C. Steele (1988): Estimates of genetic parameters for carcass measures of body composition and growth in swine. J. Anim. Sci. 66: 2498-2507.
Berg, R.T. and R.M. Butterfield (1968): Growth patterns of bovine muscle, fat and bone of young bulls. J. Anim. Sci. 27: 611-619.
Berg, E.P., J.C. Forrest, D.L. Thomas, N. Nusbaum, R.G. Kauffman (1994): Electromagnetic scanning to predict lamb carcass composition. J. Anim. Sci. 72: 1728-1736.
Berg, E.P., B.A. Engel, and J.C. Forrest (1998): Pork Carcass Composition Derived from Neural Network Model of Electromagnetic Scans. J. Anim. Sci. 76: 18-22.
Berkowitz, B.A. and R.S. Balaban (1989): Improvement in 31P NMR Signal-to-Noise for ATP in Vivo Using Homonuclear Decoupling. Magnetic Resonance in Medicine 12: 249-252.
Bernlohr, D.A., M.A. Simpson, A. Vogel Hertzel, and L.J. Banaszak (1997): Intracellular lipid-binding proteins and their genes. Annu. Rev. Nutr. 17: 277-303.
270

Literaturverzeichnis
Bertin, E., J.-C. Ruiz, J. Mourot, P. Peiniau, B. Portha (1998): Evaluation of dual-energy x-ray absorptiometry for body-composition assessment in rats. J. Nutr. 128: 1550-1554
Biedermann, G., Christiane Jatsch, W. Peschke, J.-P. Lindner und W. Wittmann (2000): Mast- und Schlachtleistung sowie Fleisch- und Fettqualität von Pietrain-Schweinen unterschiedlichen MHS-Genotyps und Geschlechts. 1. Mitteilung: Mast- und Schlachtleistung sowie Fleischqualität. Arch. Tierzucht (Dummerstorf) 43: 151-164.
Bikker, P. (1994): Protein and lipid accretion in body components of growing pigs. Thesis, Wageningen.
Bikker P., V. Karabinas, M.W.A. Verstegen, and R. G. Campbell (1995): Protein and Lipid Accretion in Body Components of Growing Gilts (20 to 45 Kilograms) as Affected by Energy Intake. J. Anim. Sci. 73: 2355-2363.
Bikker P., M.W.A. Verstegen, B. Kemp and M.W. Bosch (1996a): Performance and Body Composition of Finishing Gilts (45 to 85 Kilograms) as Affected by Energy Intake and Nutrition in Earlier Life: I. Growth of the Body and Body Composition. J. Anim. Sci. 74: 806–816.
Bikker P., M.W.A. Verstegen, and R. G. Campbell (1996b): Performance and Body Composition of Finishing Gilts (45 to 85 Kilograms) as Affected by Energy Intake and Nutrition in Earlier Life: II. Protein and Lipid Accretion in Body Components. J. Anim. Sci. 74: 817–826.
Bidanel, J.P., A. Ducos, R. Guéblez and F. Labroue (1994): Genetic parameters of backfat thickness, age at 100 kg and ultimate pH in on-farm tested French Landrace and Large White pigs. Livest. Prod. Sci. 40: 291-301.
Bidwell, C.A., S. Ji, G.R. Franks, S.G. Cornelius, G.M. Willis, and M.E. Spurlock (1997): Cloning and expression of the porcine obese gene. Anim. Biotechnol. 8: 191-206.
Biochemestry (1992): Editors Robert H. Abeles, Perry A. Frey, and William P. Jencks. Jones and Bartlett Publishers, Boston.
BioScan OPTIMAS (1991): User Guide. BioScan Inc., 170 West Dayton, Edmonds, WA 98020.
Billington, C.J. and A.S. Levine (1996): Appetite regulation: Shedding new light on obesity. Current Biology 6: 920-923.
Blackband, S.J. and M.K. Stoskopf (1990): In vivo nuclear magnetic resonance imaging and spectroscopy of aquatic organisms. Magnetic Resonance Imaging 8: 191-198.
Bland, J.M. and D.G. Altman (1986): Statistical methods for assessing agreement between two methods of clinical measurement. Lancet (February 8): 307-310.
Blendl, H.M., W. Knott, A. Baumann, E. Goretzki und H. Männer (1986): 30 Jahre Mast- und Schlachtleistungsprüfung beim Schwein in Grub. Bayer. Landw. Jahrbuch 63/6: 683-710.
Bloch, F, W.W. Hansen, M. Packard (1946): Nuclear induction. Phys. Rev. 69: 127.
271

Literaturverzeichnis
Block, B.A., T. Imagawa, A. Leung, P. Campbell, C. Franzini-Armstrong (1988): Structural evidence for direct interaction between the molecular components of the transverse tubules/sarcoplasmic reticulum junction in skeletal muscle. J. Cell Biol. 107: 2587-2600.
Boesiger, P. (1991): Localized NMR spectroscopy in-vivo. Proc. 12th Symposium Energy Metabolism of Farm Animals, Kartause Ittingen, Switzerland, 1-7 September: 210-217.
Bogner, P., E. Berényi, A. Miseta, P. Horn, M. Kellermeyer, D.N. Wheatley, and F.A. Jolesz (1996): Nuclear Magnetic Resonance Relaxation Parameters of Muscle in Malignant Hyperthermia-Susceptible Swine. Acad. Radiol. 3: 26-30.
Boileau, R.A. (1988): Utilization of Total Body Electrical Conductivity in Determining Body Composition. In: “Designing Foods: Animal Product Options in the Marketplace”, National Academy Press, Washington, DC, pp. 251-257. http://books.nap.edu/books/0309037956/html/R1.html Boling C.A., W. Taylor, C. Entemor, A.R. Behnke (1962): Total exchangeable potassium and chloride and total body water in healthy men of varying fat content. J. Clin. Invest. 31: 1840-1849.
Bosch, F., A. Pujol, A. Valera (1998): Transgenic mice in the analysis of metabolic regulation. Annu. Rev. Nutr. 18: 207-232.
Böttcher, P., J. Maierl, Th. Schiemann, Ch. Glaser, R. Weller, K.-H. Hoehne, M. Reiser, H.-G. Liebich (1999): The visible animal project: a three-dimensional, digital database for high quality three-dimensional reconstructions. Veterinary Radiology & Ultrasound 40: 611-616.
Bouchard, C. and A. Tremblay (1997): Genetic Influences on the Response of Body Fat and Fat Distribution to Positive and Negative Energy Balances in Human Identical Twins. J. Nutr. 127: 943S–947S.
Borggard, C., N.T. Madsen & H.H. Thodberg (1996): In-line Image Analysis in the Slaughter Industry, Illlustrated by Beef Carcass Classification. Meat Science 43 (No. S): S151-S163.
Brandt, H. (1987): Development and genetic aspects of feed intake in three breeds of pigs at German test stations and measures to prevent further deterioration. Pig News and Information Vol. 8 No. 1: 29-33.
Brandstätter, B. (2000): (Institut für Grundlagen und Theorie der Elektrotechnik), TU-Graz, Österreich, http://www-igte.tu-graz.ac.at/services/da/eit.htm
Branscheid, W. (1994): Ultraschall als Referenz für die Zulassung von Klassifizierungsgeräten. Landbauforschung Völkenrode, Sonderheft 145: 5-14.
Branscheid, W. und A. Dobrowolski (1996): Zur Genauigkeit der Video-Image-Analyse: Erfassung des Teilstückwertes und der Fleischhelligkeit von Schweineschlachtkörpern. Fleischwirtschaft 76: 1228-1238.
272

Literaturverzeichnis
Branscheid, W. and A. Dobrowolski (2000): Evalaution of market value: comparison between different techniques applied on pork carcasses (short communication). Arch. Tierzucht (Dummerstorf) 43: 131-137.
Branscheid, W., A. Dobrowolski und R. Höreth (1994): Untersuchungen zur Zuverlässigkeit der Bestimmung des Muskelfleischanteils von Schweinehälften nach der Kühlung. Arch. Tierzucht (Dummerstorf) 37: 121-131.
Branscheid, W., A. Dobrowolski, R. Höreth (1995): Die Video-Image-Analyse: Methode zur On-line-Erfassung des Teilstückwertes von Schweineschlachtkörpern. Fleischwirtschaft 75: 636-642.
Branscheid, W., A. Dobrowolski, M. Spindler, Ch. Augustini (1998): Apparative Klassifizierung und Erfassung des Schlachtwertes beim Rind mit Hilfe der Videbildanalyse. Fleischwirtschaft 78: 1046-1050.
Brascamp, E.W., C.S. Haley, M.A.M. Groenen and L.L.G. Janss (1995): PiGMaP: gene mapping and its contribution to meat production and meat quality parameters. Pig News and Information 16: 41N-46N.
Bray, G.A. (1997): Progress in Understanding the Genetics of Obesity. J. Nutr. 127: 940S-942S.
Brem, G. und B. Brenig (1992): Molekulare Genotyp-Diagnostik des Malignen Hyperthermie-Syndroms zur effizienten Zucht streßresistenter Schweine. Wien. Tierärztl. Mschr. 79: 301-305.
Brenig, B and G. Brem (1992): Molecular cloning and analysis of the porcine “halothane” gene. Arch. Tierzucht, (Dummerstorf) 35: 129-135.
Brenig, B., E.-L. Winnacker, G. Brem (1991): Molekulargenetische Untersuchung der malignen Hyperthermie des Schweines. In: Fortschritte in der Tierzüchtung, Ulmer, Stuttgart: 315-326.
Brent, R. and R.L. Finley Jr. (1997): Understanding gene and allele function with two-hybrid methods. Annu. Rev. Genet. 31: 663-704.
Brethour, J.R. (2000): Using serial ultrasound measures to generate models of marbling and backfat thickness changes in feedlot cattle. J. Anim. Sci. 78: 2055-2061.
Brockmann, G.A., Ch.S. Haley, U. Renne, S.A. Knott, and M. Schwerin (1998): Quantitative Trait Loci Affecting Body Weight and Fatness From a Mouse Line Selected for Extreme High Growth. Genetics 150: 369-381.
Brocks, L., R.E. Klont, W. Buist, K. de Greef, M. Tieman, and B. Engel (2000): The effects of selection of pigs on growth rate vs. leanness on histochemical characteristics of different muscles. J. Anim. Sci. 78: 1247–1254.
Brooke-Wavell, K., P.R.M. Jones, N.G. Norgan, and A.E. Hardman (1995): Evaluation of near infra-red interactance for assessment of subcutaneous and total body fat. European Journal of Clinical Nutrition 49: 57-65.
Brozek, J. (1966): Body Composition: Models and Estimation Equations. Am. J. Phsy. Anthrop. 24: 239-246.
273

Literaturverzeichnis
Brozek, J., F. Grande, J. T. Anderson and A. Keys (1963): Densitometric analysis of body composition: revision of some quantitative assumptions. Ann. N. Y. Acad. Sci. 110: 113-140.
Brøndum, J., M. Egebo, C. Agerskov, and H. Busk (1998): On-line pork carcass grading with the Autofom ultrasound system. J. Anim. Sci. 76: 1859-1868.
Brunton, J.A., H.S. Baley, St.A. Atkinson (1993): Validation and application of dual-energy x-ray absorptiometry to measure bone mass and body composition in small infants. Am. J. Clin. Nutr. 58: 839-845.
Brunton, J.A., H.A. Weiler, and St.A. Atkinson (1997): Improvement in the Accuracy of Dual Energy X-ray Absorptiometry for Whole Body and Regional Analysis of Body Composition: Validation Using Piglets and Methodologic Considerations in Infants. Pediatric Research 41: 590-596.
Brüngel, M. und R. Kluthe (1997): Die bioelektrische Impedanzanalyse als Methode zur Erfassung der Körperzusammensetzung. DGE-info Mai 1997, Review, [Bioelectrical impedance analysis in body composition measurement: National Institutes of Health Technology Assessment Conference Statement. Am J Clin Nutr 64 (suppl. 3): 524S–532S]. http://www.dge.de/Pages/navigation/fach_infos/rev0597.htm
Buchli, R. and P. Boesiger (1993): Comparison of Methods for the Determination of Absolute Metabolite Concentrations in Human Muscles by 31P MRS. Magnetic Resonance in Medicine 30: 552-558.
Burt, C.T., J. Koutcher, J.T. Roberts, R.E. London, B. Chance (1986). Magnetic resonance spectroscopy of the musculoskeletal system. Radiol. Clin. North Am. 24: 321-331.
Busemann, E. (1991): Möglichkeiten der Schlachtkörperbewertung am lebenden Schwein mit Hilfe von Ultraschallmessungen. Schriftenreihe des Instituts für Tierzucht und Tierhaltung der Christian-Albrechts-Universität zu Kiel, Heft 64.
Busk, H. (1989): Applications of Ultrasound Imaging in Animal Science. In: Application of NMR Techniques on the Body Composition of Live Animals (Eds.: Kallweit, E., M. Henning, E. Groeneveld), Elsevier Applied Science: 75-90.
Busk, H. and E. Olsen (1996): Determination of percent meat in pig carcasses with the AUTOFOM equipment. 47th Annual Meeting of the EAAP, Lillehammer, Norway, 25-29 August, (paper copy, 9 pages).
Busk, H., A. Karlsson and S.H. Hertel (2000): Halothane gene effect on carcass and meat quality by use of Duroc x Pietrain Boars. In: Quality of meat and fat in pigs as affected by genetics and nutrition. EAAP publiction No. 100: 129-133.
Busse, W. und E. Groeneveld (1986): Schätzung von Populationsparametern bei Schweinen der Deutschen Landrasse an Daten aus dem Marienseer-Herdbuch-Informationssystem. 1. Mitteilung: Schätzung der Populationsparameter für die Merkmale der Geschwisterleistung auf Station. Züchtungskunde 58: 175-183.
274

Literaturverzeichnis
Busse, W. und E. Groeneveld (1986): Schätzung von Populationsparametern bei Schweinen der Deutschen Landrasse an Daten aus dem Marienseer-Herdbuch-Informationssystem. 2. Mitteilung: Schätzung von Populationsparametern für Merkmale der Eigenleistungsprüfung von Jungebern auf Auktion und Schätzung von genetischen Korrelationen zwischen Merkmalen der Eigenleistungsprüfung auf Auktion und Merkmalen der Geschwisterleistungsprüfung auf Station unter Berücksichtigung von Genotyp-Umwelt-Interaktionen. Züchtungskunde 58: 184-195.
Bussolotto, M., A. Ceccon, G. Sergi, V. Giantin, P. Beninca, G. Enzi (1999): Assessment of body composition in elderly: accuracy of bioelectrical impedance analysis. Gerontology 45: 39-43.
Butte, N.F., J.M. Hopkinson, K.J. Ellis, W.W. Wong, E.O. Smith (1997): Changes in fat-free mass and fat mass in postpartum woman: a comparison of body composition models. Intl. J. Obesity 21: 874-880.
Cameron, Charles Alexander (1868): The stock-feeder's manual. The chemistry of food in relation to the breeding and feeding of live stock. Cassell, Petter, and Galpin, London, | New York, Albert R. Mann Library. 2000. Core Historical Literature of Agriculture (CHLA). Ithaca, NY: Albert R. Mann Library, Cornell University. http://chla.library.cornell.edu (Version October 2000). http://cdl.library.cornell.edu/cgi-bin/chla/chla-cgi?notisid=AFB3188
Cameron, N.D. and Curran, M.K. (1994): Responses in performance test traits to selection on components of efficient lean growth in pigs. Proc. 5th World Congr. Genet. Appl. Live. Prod., Guelph, Vol. 19: 83–86.
Cameron, N.D., Curran, M.K. and Kerr, J.C. (1994): Selection for lean growth in pigs with a restricted feeding regime. Proc. 5th World Congr. Genet. Appl. Live. Prod., Guelph, Vol. 19: 87–90.
Cargill, E.J., L.C. Baskin, and D.Pomp (1998): Rapid Communication: Localization of the Porcine Carboxypeptidase-E Gene by Linkage Analysis Further Extends the region of Synteny Between Human Chromosome 4 and Porcine Chromosome 8. J. Anim. Sci. 76: 2211-2212.
Cartee, G.D. (1995): What Insights Into Age-Related Changes in Skeletal Muscle Are Provided by Animal Models. The Journal of Gerontology 50A (Special Issue): 137-141.
Cerdan, S. and J. Seelig (1990): NMR Studies of Metabolism. Annu. Rev. Biophys. Chem. 19: 43-67.
Cisneros F., M. Ellis, K.D. Miller, J. Novakofski, E.R. Wilson, F.K. McKeith (1996): Comparison of transverse and longitudinal real-time ultrasound scans for prediction of lean cut yields and fat-free lean content in live pigs. J. Anim. Sci. 74: 2566-2576.
Chance, B. (1984): Applications of 31P NMR to Clinical Biochemestry. Ann. N. Y. Acad. Sci. 428: 318-332.
Charette, R., M. Bigras-Poulin, G.-P. Martineau (1996): Body condition evaluation in sows. Livest. Prod. Sci. 46: 107-115.
275

Literaturverzeichnis
Cheah, K.S., A.M. Cheah, and D.I. Krausgrill (1995): Variations in Meat Quality in Live Halothane Heterozygotes Identified by Biopsy Samples of M. longissimus dorsi. Meat Sci. 39: 293-300.
Clapper, J.A., T.M. Clark, and L.A. Rempel (2000): Serum concentrations of IGF-I, estradiol-17ß, testosterone, and relative amounts of IGF binding proteins (IGFBP) in growing boars, barrows, and gilts. J. Anim. Sci. 78: 2581-2588.
Claus, A. (1957): Die Messung natürlicher Grenzflächen im Schweinekörper mit Ultraschschall. Fleischwirtschaft 9: 552-554.
Claus, R. (1996): Physiologische Grenzen der Beeinflußbarkeit von Leistungen beim Schwein. Züchtungskunde 68: 493-505.
Claus, R. and U. Weiler (1994): Endocrine regulation of growth and metabolism in the pig: a review. Livest. Prod. Sci. 37: 245-260.
Clutter, A.C., S. Sasaki, and D. Pomp (1998): Rapid Communication: The Cholecystokinin Type-A Receptor (CCKAR) Gene Maps to Porcine Chromosome 8. J. Anim. Sci. 76: 1983-1984.
Cohn, S.H. (1992): In vivo neutron activation analysis; a new technique in nutritional research. The journal of nutritional biochemistry 3: 378-386.
Comizio, R., A. Pietrobelli, Y.X. Tan, Z.M. Wang, R.T. Withers, S.B. Heymsfield, C.N. Boozer (1998): Total body lipid and triglyceride response to energy deficit: relevance to body composition models. Am. J. Physiol. 274: E860-E866.
Comuzzie, A.G., J.E. Hixson, L. Almasy, B.D. Mitchell, M.C. Mahaney, T.D. Dyer, M.P. Stern, J.W. MacCluer and J. Blangero (1997): A major quantitative trait locus determining serum leptin levels and fat mass is located on human chromosome 2. Nat. Genet. 15: 273-276.
Conway, J.M., K.H. Norris, and C.E. Bodwell (1984): A new approach for the estimation of body composition: infrared interactance. Am. J. Clin. Nutr. 40: 1123-1130.
Cooke, S.R., St.R. Peterson, H. A. Quinney (1997): The influence of maximal aerobic power on recovery of skeletal muscle following anerobic exercise. Eur. J. Appl. Physiol. 75: 512-519.
Cormack, M.M. (1980): Early two-dimensional reconstruction and recent topics stemming from it. Med. Phys. 7: 277-281.
Cuthbertson, A. and R.W. Pomeroy (1962): Quantitative anatomical studies of the composition of the pig at 50, 68 and 92 kg carcass weight. I: Experimental material and methods. J. Agric. Sci. 59: 207-214.
Dai, R., N. Li, Ch. Wu (1998): Molecular cloning of “ob” cDNA and polymorphic analysing on “ob” locus in swine.
276

Literaturverzeichnis
Proc. 6th World Congress on Genetics Applied to Livestock Production, Armidale, Australia, Vol. 23: 612-615. Davies, A.S. (1984): Wachstumsverlauf von Muskeln und Knochen bei Schweinen unterschiedlicher Endgröße. Dissertation, Tierärztliche Hochschule Hannover.
Davies, A.S., K.L. Garden, M.J. Young and C.S.W. Reid (1987): An atlas of X-ray tomographical anatomy of the sheep. Science Information Publishing Centre, DSIR, Wellington, New Zealand.
Davies, D.R. and A. Wlodawer (1995): Cytokines and their receptor complexes. FASEB J. 9: 50-56.
Dänicke S., Ingrid Halle, H. Jeroch (1997): Evaluation of the non invasive TOBEC (total body electrical conductivity) procedure for prediction of chemical components of male broilers with special consideration of dietary protein level. Arch. Anim. Nutr. 50: 137-153.
Decanniere, Caroline (1993): In-Vivo Localized NMR Spectroscopy at High Field Strength. Thesis, Katholieke Universiteit Leuven, Belgium, pp. 149.
Decanniere, Caroline, P. van Hecke, F. Vanstapel, H. Villé, R. Geers (1993): Metabolic response to halothane in piglets susceptible to malignant hyperthermia: an in vivo 31P-NMR study. J. Appl. Physiol. 75: 955-962.
De Koning, D.J., A.P. Rattink, B. Harlizius, M.A.M. Groenen, E.W. Brascamp, J.A.M. van Arendonk (2001): Detection and characterization of quantitative trait loci for growth and reproduction traits in pigs. Livest. Prod. Sci. 72: 185-198.
De Lorenzo, A., A. Andreoli, J. Matthie, and P. Withers (1997): Predicting body cell mass with bioimpedance by using theoretical methods: A technological review. J. Appl. Physiol. 82: 1542–1558.
Depreux, F.F.S., A.L. Grant, J.C. Forrest, and D.E. Gerrard (1998): Effect of halothane gene status on myosin heavy chain content and meat quality attributes. J. Anim. Sci. 76 (Suppl. 1): 151.
Deurenberg, P., R. Leenen, K. van der Kooy, J.G. Hautvast (1989): In obese subjects the body fat percentage calculated with Siri's formula is an overestimation. Eur. J. Clin. Nutr. 43: 569-575.
De Smet, S.M., H. Pauwels, S. De Bie, D.I. Demeyer, J. Callewier, and W. Eeckhout (1996): Effect of Halothane Genotype, Breed, Feed Withdrawal, and Lairage on Pork Quality of Belgian Slaughter Pigs. J. Anim. Sci. 74: 1854-1863.
De Vries, A.G. and P.G. van der Wal (1993): Breeding for Pork Quality. In: Pork Quality: Genetic and Metabolic factors (Editors: E. Poulanne and D.I. Demeyer), CAB International: 58-75.
De Vries, A.G. and E. Kanis (1994): Swine breeding goals, short- and long-term considerations. Proc. 5th World Congress on Genetics applied to Livestock Production. Guelph, Canada, Vol. 17, 390-397.
277

Literaturverzeichnis
De Vries, A.G. and G.S. Plastow (1998): Major genes for quality. Pig Progress 14: 43-48. De Vries, A.G., R. Hovenier, E.W. Brascamp, J.W.M. Brascamp (1993): Breeding for Pork Quality in the Netherlands. 44th Annual Meeting of the EAAP, Denmark, 16-19 August, paper copy, pp. 7.
De Vries, A.G., P.G. van der Wal, T. Long, G. Eikelenboom and J.W.M. Merks (1994): Genetic parameters of pork quality and production traits in Yorkshire populations. Livest. Prod. Sci. 40: 277-289.
Dickerson, G.E. and H.S. Teague (1987): Energy intake restriction and lean growth efficiency in swine. J. Anim. Breed. Genet. 104: 82-95.
Diehl, B.W.K. und W. Ockels (1995): Ermittlung der Fettsäureverteilung durch 13C-NMR-Spektroskopie. Fett 97(3): 115-118.
Ding, S.-T., A.P. Schinckel, T.E. Weber, and H.J. Mersmann (2000): Expression of porcine transcription factors and genes related to fatty acid metabolism in different tissues and genetic populations. J. Anim. Sci. 78: 2127-2134.
Domermuth, W., T.L. Veum, M.A. Alexander, H.B. Hedrick, J. Clark, and D. Eklund. (1976): Prediction of lean body composition of live market weight swine by indirect methods. J. Anim. Sci. 43: 966-976.
Doornenbal, H. (1971): Growth, development and chemical composition of the pig. I. Lean tissue and protein. Growth 35: 281-295.
Doornenbal, H. (1972): Growth, development and chemical composition of the pig. II. Fatty tissue and chemical fat. Growth 36: 185-194.
Drudik, D.K., D. Pomp, Z-B. Zeng and E.J. Eisen (1995): Identification of major genes controlling body weight and fat percentage on mouse chromosome 2. J. Anim. Sci. 73 (Suppl. 1): 110.
Dubinsky, V.Z. and L.A. Sibeldina (1983): [31P-NMR Study of P-Compounds Metabolism in Mouse Liver under Stress.] in Russisch, Biophysika XXVIII: 92-95.
Ducos, A., J.P. Bidanel, and J. Naveau (1992): Estimation of genetic parameters and genetic trends for production traits in the Sino-European Tiameslan composite line. J. Anim. Breed. Genet. 109: 108-118.
Dwyer, Catherine M., N.C. Stickland, and J.M. Fletcher (1994): The influence of maternal nutrition on muscle fiber number development in the porcine fetus and on subsequent postnatal growth. J. Anim. Sci. 72: 911-917.
Edeal, B., M. Rumph, R. Mass, K. Killinger, N. Jerez, S. Elnagar, T. McDaneld, R. Aljumaah, U. Pithpongsiriporn, K. Nephawe, G. Martinez, C.D. Gladney, M.F. Allan, and D. Pomp (2000):
278

Literaturverzeichnis
Rapid communication: Linkage mapping of the Mahogany (attractin) locus in cattle and pigs. J. Anim. Sci. 78: 2479-2480.
Eikelenboom, G. (1980): New Ways To Achieve An Optimum Combination Of Carcass Quality; Meat Quality And Stress Resistance In Pigs. Hohenheimer Arbeiten, Reihe Tierische Produktion 107: 60-67. Eikelenboom, G. and D. Minkema (1974): Prediction of pale, soft, exudative muscle with a non-lethal test for the halothane -induced porcine malignant hyperthermia syndrome. Neth. J. Vet. Sci. 99: 421-426.
Ehricke, H.-H. and G. Laub (1990): Tissue Discrimination in Three-dimensional Imaging by Texture Analysis. In: Tissue Characterization in MR Imaging. (Eds.: H.P. Higer, G. Bielke), Springer Verlag, Berlin, Heidelberg: 149-153.
Ehrlich, Barbara E. (1995): Functional properties of intracellular calcium-release channels. Current Opinion in Neurobiology 5: 304-309.
Elia, M., A. Caner, R. Smith (1979): The 3-methylhistidine content of human tissues. Br. J. Nutr. 42: 567-570. Elbers, J.M.H., G. Haumann, H. Asscheman, J.C. Seidell, L.J.G. Gooren (1997): Reproducibility of fat area measurements in young, non-obese subjects by computerized analysis of magnetic resonance images. Intl. J. Obesity 21: 1121-1129.
Ellington, W.R. (1989): Phosphocreatine represents a thermodynamic and functional improvement over other muscle phosphagens. J. Exp. Biol. 143: 177-194.
Ellis, K.J. (2000): Human Body Composition: In Vivo Methods. Physiological Reviews 80: 649-680.
Ellis, K.J., R.J. Shypailo, J.A. Pratt, W.G. Pond (1994): Accuracy of dual-energy x-ray absorptiometry for body-composition measurements in children. Am. J. Clin. Nutr. 60: 660-665.
Elowsson, P., A.H. Forslund, H. Mallmin, U. Feuk, I. Hansson, J. Carlsten (1998): An evaluation of dual-energy x-ray absorptiometry and underwater weighing to estimate body composition by means of carcass analysis in piglets. J. Nutr. 128: 1543-1549. Elsaesser, F. und Nahid Parvizi (1996): Zwanzig Jahre Forschungsbereich Endokrinologie Mariensee: Neuere Erkenntnisse zur hormonalen und molekularen Steuerung des Wachstums beim Schwein. Züchtungskunde 68: 280-296.
Emmans, G.C., and I. Kyriazakis (1997): Models of pig growth: problems and proposed solutions. Livest. Prod. Sci. 51: 119-129.
279

Literaturverzeichnis
Ender, K., K. Nürnberg und H.-J. Papstein (1997a): Die zwei Seiten des Fettes – eine Standortbestimmung. Arch. Tierz. (Dummerstorf) 40: 35-45.
Ender, K., Elke Albrecht und J. Wegner (1997b): Die Video-Bildanalyse zur Messung qualitätsrelevanter Merkmale des Rindfleisches. Arch. Tierz. (Dummerstorf) 40 (Sonderheft): 148-154.
Engellandt, Th., N. Reinsch, S. Reinicke und E. Kalm (1997): Schätzung genetischer Parameter für die Vaterlinien Pietrain und Belgische Landrasse der Schweineherdbuchzucht Schleswig-Holstein. Züchtungskunde 69: 39-53.
Engelson, Ellen S., D.P. Kotler, Y.X. Tan, Denise Agin, J. Wang, R.N. Pierson, and St.B. Heymsfield (1999): Fat distribution in HIV-infected patients reporting truncal enlargement quantified by whole-body magnetic resonance imaging. Am. J. Clin. Nutr. 69: 1162-1169.
Enser, M. and J.D. Wood (1991): Factors controlling fat quality in pigs. 42nd Annual Meeting of the EAAP, Berlin, 9.-12. September, (paper copy, pp. 9).
Ernst, E. (1994): Ultraschall zur Ermittlung der Körperzusammensetzung bei Schwein und Rind. Landbauforschung Völkenrode, Sonderheft 145: 84-89.
Ernst, C.W., P. Kapke, M.Yerle and M.F. Rothschild (1997): The leptin receptor gene (LEPR) maps to porcine Chromosome 6. Mammalian Genome 8: 226.
Essèn-Gustavsson, Birgitta (1993): Muscle-fiber Characteristics in Pigs and Relationships to Meat-quality Parameters - Review. In: Pork Quality: Genetic and Metabolic Factors (Eds.: E. Poulanne, D.J. Demeyer, M. Ruusunen and S. Ellis), CAB International, Wallingford, UK, 140-159.
Essèn-Gustavsson, Birgitta (1995): Effects of physical stress on muscle fibre characteristics in pigs – relation to meat quality. Proc. 2nd Dummerstorf Muscle-Workshop Muscle Growth and Meat Quality. FBN Schriftenreihe 6: 65-76.
Estany, J., J. Tibau, J. Soler, J.L. Noguera (1998): Responses in production, carcass and meat quality traits after an hyperprolific cycle of selection in pigs. Proc. 6th World Congress on Genetics Applied to Livestock Production, Armidale, Australia, Vol. 23: 628-631.
Etherton, T.D. and D.E. Bauman (1998): Biology of Somatotropin in Growth and Lactation of Domestic Animals. Physiol. Rev. 78: 745-761.
Ettinger, K.V., M.A. Foster and U.J. Miola (1983): Future developments in the in vivo measurement of body composition in pigs. CEC Meeting: In Vivo Measurement of Body Composition in Meat Animals. Bristol, UK, November 30 - December 1.
Feddern, Elke, J. Wegner, K. Ender und E. Kalm (1995): Untersuchungen von Muskelstrukturmerkmalen bei Hampshire-Reinzuchttieren und verscheidenen Kreuzungskombinationen. Arch. Tierzucht (Dummerstorf) 38: 43-56.
280

Literaturverzeichnis
Feldmann, H.U., G. Göretzlehner, A. Heufelder, F. Husmann, F. Jockenhövel, R. Körfer, H. Koertke, H. Kuhl, B. Leiber, G. Ludwig, J.F. Schindler, A.T. Teichmann (2000): gynehormonweb - Intrazelluläre Rezeptoren: Waisen-Rezeptoren: P, PP und PPAR. (Online Editor: J.F. Schindler) http://www.gynehormonweb.de/WebSite/A-Intrazell_Rezeptoren/C-Waisen-Rezeptoren/Ca-PPAR.html - in Zusammenarbeit mit gyne (Zeitschrift für den Frauenarzt — den Hausarzt der Frauen) und aner (Zeitschrift für Endokrinologie in der Männerheilkunde).
Fender, Maria, S. Kühnle und D. Fewson (1979): Selektionsversuch beim Schwein zur Verbesserung der Schlachtkörper-zusammensetzung. Z. Tierzüchtg. Züchtgsbiol. 96: 86-95.
Ferrell C.L. and S.G. Cornelius (1984): Estimation of body composition of pigs. J. Anim. Sci. 58: 903-912.
Fewson, D. (1987): Muskelproportionen und Typfragen in der Schweinezucht. Züchtungskunde 59: 416-429.
Fewson, D., W. Branscheid und E. Sack (1990): Untersuchungen über das relative Wachstum der Teilstücke und Gewebe von männlichen und weiblichen Mastschweinen verschiedener Herkünfte. Züchtungskunde 62: 304-316.
Fiedler, Ilse, K. Ender, M. Wicke und G. von Lengerken (1993): Zusammenhänge zwischen Mikrostruktur des Muskelgewebes bei Schweinen der Landrasse und ihrer Streßempfindlichkeit (Halothanreaktion). Arch. Tierz., (Dummerstorf) 36: 525-538.
Fiedler, Ilse, Gerda Kuhn, Margitta Hartung, U. Küchenmeister, Karin Nürnberg, Charlotte Rehfeldt, Korinna Huber und Danuta Klosowska (2001a): Auswirkungen des Malignen Hyperthermie-Syndroms (MHS) auf Fleischqualität, Muskelfasereigenschaften und Stoffwechselkriterien des M. longissimus von Pietrain-Schweinen. Arch. Tierzucht (Dummerstorf) 44: 203-217.
Fiedler, I., M. Hartung, G. Dietl, H. Schewe (2001b): Muskelstrukturanalyse liefert Aussagen zu Fleischansatz und Fleischqualität von Rassen. Schweinezucht und Schweinemast 49/2: 35-36.
Fiorotto, M.L., W.J. Cochran, R.C. Funk, H.P. Sheng, W.J. Klish (1987): Total body electrical conductivity measurements: effects of body composition and geometry. Am. J. Physiol. 252(4 Pt 2): R794-800.
Fisler, J.S and C.H. Warden (1997): Mapping of Mouse Obesity Genes: A Generic Approach to a Complex Trait. J. Nutr. 127: 1909S-1916S.
Flachowsky, G. (1998): Einflußmöglichkeiten der Tierernährung auf Inhaltsstoffe und Qualität von Lebensmitteln tierischer Herkunft. Verbraucherdienst 43-3: 388-392.
Flynn, M.A., G.B. Nolph, and G. Krause (1995): Comparison of body composition measured by total body potassium and infrared interactance. Journal of the American College of Nutrition 14: 652-655.
281

Literaturverzeichnis
Fogelholm, M. and W. van Marken Lichtenbelt (1997): Comparison of body composition methods: a literature analysis. Eur. J. Clin. Nutr. 51: 495-503.
Fogelholm, M., T.K. Kukkonen-Harjula, H.T. Sievänen, P. Oja and I.M. Vuori (1996): Body composition assessment in lean and normal-weight young women. Brit. J. Nutr. 75: 793-802.
Folch, J.M., M. Lees, and G.H. Sloan-Stanley (1957): A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 226: 497-509.
Forbes, G.B. (1962): Methods for determining composition of the human body. Pediatrics 29: 477-494.
Forbes, G.B. (1999): Body Composition: Overview. J. Nutr. 129: 270S-272S.
Forbes, G.B., and J.B. Hursch (1963): Age and sex trends in LBM calculated from K40 measurements: With a note on the theoretical basis for the procedure. Ann. N.Y. Acad. Sci. 110: 255-263.
Forbes G.B. and G.J. Bruining (1976): Urinary creatinine excretion and lean body mass. Am. J. Clin. Nutr. 29: 1359-1366.
Forrest, J.C. (1994): Klassifizierung von Schlachtkörpern und Teilstücken mit dem TOBEC-Gerät. Landbauforschung Völkenrode, Sonderheft 145: 49-58.
Förster, M. (1991a): Genomanalyse bei landwirtschaftlichen Nutztieren. Züchtungskunde 63: 174-182.
Förster, M. (1991b): Zur Bedeutung der Genomanalyse und Gendiagnostik in der Tierzüchtung. In: Fortschritte in der Tierzüchtung, Ulmer, Stuttgart, 295-313.
Förster, M. (1992): Gendiagnostische Erbfehleranalyse zur Verbesserung der Tiergesundheit. Züchtungskunde 64: 405-410.
Förster, M., B. Brenig, G. Brem, W. Wittmann, B. Littmann, W. Peschke (1992): Der MHS-Gentest für die sichere Streßresistenzzüchtung beim Schwein. Schweinezucht und Schweinemast 40: 4-6.
Fortin, A. and J.I. Elliot (1985): Relationships between backfat thickness and chemical composition of the body and components of swine. J. Anim. Sci. 61: 158-164.
Fortin, A., J.D. Wood, and O.P. Whelehan (1987a): Breed and sex effects on the development and proportions of muscle, fat and bone in pigs. J. Agric. Sci. (Camb.) 108: 39-45.
Fortin, A., J.D. Wood, and O.P. Whelehan (1987b): Breed and sex effects on the development, distribution of muscle, fat and bone, and the partition of fat in pigs. J. Agric. Sci. (Camb.) 108: 141-153.
Foster, M.A. and P.A. Fowler (1988): Non - invasive methods for assessment of body composition. Proc. Nutr. Soc. 47: 375 - 385.
282

Literaturverzeichnis
Fowler, V.R. (1986): Biological advances towards genetic improvement in pigs. 3 rd World Congress on Genetics applied to Livestock Production, Lincoln, Nebraska, USA, Vol. XI: 345-354.
Fowler, V.R., M. Bichard, A. Pease (1976): Objectives in pig breeding. Anim. Prod. 23: 365 - 387.
Fowler, P.A., M.F. Fuller, Ch.A. Glasbey, G.G. Cameron, M.A. Foster (1992): Validation of the in vivo measurement of adipose tissue by magnetic resonance imaging of lean and obese pigs. Am. J. Clin. Nutr. 56: 7-13.
Franke, A., H. P. Kühl, und P. Hanrath (1998): Dreidimensionale Rekonstruktion echokardiographischer Schnittbilder. Methodik, klinischer Stellenwert und Zukunftsperspektiven. Dtsch. med. Wschr. 123: 1245-1249.
Franzini-Armstrong, C. and A.O. Jorgensen (1994): Structure and development of E-C coupling units in skeletal muscle. Annu. Rev. Physiol. 56: 509-534.
Fredeen, H.T., A.H. Martin, and A.P. Sather (1979): Evaluation of an electronic technique for measuring lean content of the live pig. J. Anim. Sci. 48: 536-540.
Friedman, J.M. (1991): Molecular mapping of the mouse ob mutation. Genomics 11: 1054-1062.
Fries, R. (2000): DNA-Variation – Polymorphismus – Genetische Variation. Hülsenberger Gespräche: 35-43.
Friesen, K.G., J.L. Nelssen, J.A. Unruh, R.D. Goodband, M.D. Tokach (1994): Effects of the interrelationship between genotype, sex, and dietary lysine on growth performance and carcass composition in finishing pigs fed to either 104 or 127 kilograms. J. Anim. Sci. 72: 946-954.
Fujii, J., K. Otsu, F. Zorzato, S. de Leon, V. K. Khanna, J. Weiler, P.J. O'Brien, D.H. MacLennan (1991): Identification of a mutation in the porcine ryanodine receptor that is associated with malignant hyperthermia. Science 253: 448-451.
Fuller, M.F., R.A. Houseman, A. Cadenhead (1971): The measurement of exchangeable potassium in living pigs and its relation to body composition. Br. J. Nutr. 26: 203-214.
Fuller, M.F., P.A. Fowler, G. McNeill, and M.A. Foster (1994): Imaging Techniques for the Assessment of Body Composition. J. Nutr. 124: 1546S-1550S.
Fuller, N.J., M.B. Sawyer, M.A. Laskey, P. Paxtons, and M. Elia (1996): Prediction of body composition in elderly men over 75 years of age. Annales of Human Biology 23: 127-147.
Fusch, Ch., J. Slotboom, U. Fuehrer, R. Schumacher, A. Keisker, W. Zimmermann, A. Moessinger, Ch. Boesch, and J. Blum (1999): Neonatal Body Composition: Dual-Energy X-Ray Absorptiometry, Magnetic Resonance Imaging, and Three-Dimensional Chemical Shift Imaging versus Chemical Analysis in Piglets. Pediatric Research 46: 465-473.
283

Literaturverzeichnis
Furuichi, T., K. Kohda, A. Miyawaki, and K. Mikoshiba (1994): Intracellular channels. Current Opinion in Neurobiology 4: 294-303.
Gaisser, Sybille und Bärbel Hüsing (2000): "Reproduktions- und Gentechnik bei Tieren" Dokumentation der Lehrerfortbildung, 22./23. November 1999 in Tübingen, 101 Seiten. http://www.leu.bw.schule.de/allg/gentechnik/dokuberarbeitet.pdf
Galla, H.-J. (1988): Spektroskopische Methoden in der Biochemie, Thieme Verlag, Stuttgart, New York.
Gareau, P.J., E.G. Janzen, R.A. Towner, and W.A. Stewart (1993): In vivo 31P NMR Spectroscopy studies of halothane induced Porcine Stress Syndrome. No effect of C-Phenyl N-Tert-Butyl Nitrone (PBN). Free Rad. Res. Comms. 19: 43-50.
Gassner, G. and J.A.B. Lohman (1987): Combined proton NMR imaging and spectral analysis of locust embryonic development. Proc. Natl. Acad. Sci. USA 84: 5297-5300.
Gassner, G., M.J. Line, M.M. Millard, J.R. Plimmer, W.K. Vogelbein, P. van Veld (1990): Feasibility Study: Magnetic Resonance Imaging of Hepatic Neoplasm in Mummichog Fundulus heteroclitus from a Creosote-Contaminated Environment. Cheasapeake Research Consortium Publication No. 137: 469-475. Geers, R.; Decanniere, C.; Ville, H.; van Hecke, P.; Bosschaerts, L; Goedseels, V.; Deley, J.; Janssens, S. (1991): In Vivo 31P-NMR-Spectroscopy on Muscle Tissue of Piglets: Dynamics of Intracellular Phosphocreatine and pH. Proc. 12th Symposium: Energy Metabolism of Farm Animals, Kartause Ittingen, Switzerland, 1-7 September, 246-248.
Geers, R., C. Decanniere, H. Villé, P. van Hecke, V. Goedseels, F. Vanstapel, L. Bosschaerts, J. De Ley, W. Zhang, St. Janssens (1992a): In vivo muscle 31P nuclear magnetic resonance spectroscopy during treatment of halothane-sensitive and halothane-nonsensitive pigs. Am. J. Vet. Res. 53: 613-616.
Geers, R., C. Decanniere, H. Villé, P. van Hecke, V. Goedseels, L. Bosschaerts, J. De Ley, St. Janssens, W. Nierynck (1992b): Identification of halothane gene carriers by use of in vivo 31P nuclear magnetic resonance spectroscopy in pigs. Am. J. Vet. Res. 53: 1711-1714. Geers, R.C. Decanniere, H.Villé, P.van Hecke, and L. Bosschaerts (1995): Variability within intramuscular fat content of pigs as measured by gravimetry, FTIR and NMR spectroscopy. Meat Sci. 40: 373-378. Geers, R., C. Decanniere, A. Rosier, H. Villé, P. van Hecke, F. Vandesande, J. Jourquin (1996a): Variability of energy metabolism and nuclear T3-receptors within the skeletal muscle tissue of pigs with respect to the halothane gene. J. Anim. Sci. 74: 717-722.
284

Literaturverzeichnis
Geers, R., H. Villé, G. Spincemaille, J. Vits and W. Duchateau (1996b): In vivo measurement of body composition of pigs with DEXA. 47th Meeting EAAP, Lillehammer, 25 - 29 August, 278.
Geldermann, H. (1996): Analyse von Genwirkungen auf Leistungsmerkmale. Dtsch. tierärztl. Wschr. 103: 378-383.
Gerbens, F., J.W.M. Merks, J.H. Veerkamp, and M.F.W. te Pas (1997): Effects of heart fatty acid-binding protein (h-FABP) on intramuscular fat and production traits in pigs. J. Anim. Sci. 75 (Suppl. 1): 150.
Gerbens, F., A. Jansen, A.J.M. van Erp, F.L. Harders, T.H.E. Meuwissen, G. Rettenberger, J.H. Veerkamp, M.F.W. te Pas (1998): The adipocyte fatty acid-binding protein locus: characterization and association with intramuscular fat content in pigs. Mammalian Genome 9: 1022-1026.
Gerbens, F., D.J. de Koning, F.L. Harders, T.H.E. Meuwissen, L.L.G. Janss, M.A.M. Groenen, J.H. Veerkamp, J.A.M. van Arendonk, and M.F.W. te Pas (2000): The effect of adipocyte and heart fatty acid-binding protein genes on intramuscular fat and backfat content in Meishan crossbred pigs. J. Anim. Sci. 78: 552–559.
Gerbens, F., F.J. Verburg, H.T.B. van Moerkerk, B. Engel, W. Buist, J.H. Veerkamp, and M.F.W. te Pas (2001): Associations of heart and adipocyte fatty acid-binding protein gene expression with intramuscular fat content in pigs. J. Anim. Sci. 79: 347–354.
Geri, G., A. Zappa, G. Campadoni, O. Franci and B.M. Poli (1990a): Relationships between adipose tissue characteristics of newborn pigs and subsequent performance: I. Characteristics at 8 days of age and growth rate until slaughter. J. Anim. Sci. 68: 1922-1928.
Geri, G., B.M. Poli, A. Zappa, G. Campadoni and O. Franci (1990b): Relationships between adipose tissue characteristics of newborn pigs and subsequent performance: III. Histological and chemical characteristics of backfat. J. Anim. Sci. 68: 1936-1943.
Gerrard, D.E., C.S. Okamura, and A.L. Grant (1999): Expression and Location of IGF Binding Proteins-2, -4, and –5 in Developing Fetal Tissues. J. Anim. Sci. 77: 1431-1441.
Girard, J., P. Ferré and F. Foufelle (1997): Mechanisms by which carbohydrates regulate expression of genes for glycolytic and lipogenic enzymes. Annu. Rev. Nutr. 17: 325-352.
Gjerde, B. (1987): Predicting carcass composition of rainbow trout by computerized tomography. Z. Tierzüchtung und Züchtungbiol. 104: 121-136.
Glodek, P. (1991): Einflüsse der Biotechnik auf dieTierzüchtung und –produktion. Züchtungskunde 63: 201-210.
Glodek, P. (1996): Die Wahl der Vaterlinie bestimmt die Qualität der Endprodukte in der Schweinezucht. Züchtungskunde 68: 483-492.
285

Literaturverzeichnis
Goodwin, R.N., L.L. Christian, C.F. Louis, P.J. Berger, K.J. Prusa, D.C. Beitz (1994): Effect of the HAL gene on pork carcass composition and eating quality traits. J. Anim. Sci. 72 (Suppl. 1): 249.
Goran, M.I., M.J. Toth, E.T. Poehlman (1998): Assessment of research-based body composition techniques in healthy elderly men and women using the 4-compartment model as a criterion method. Intl. J. Obesity 22: 135-142.
Gotfredsen, A., J. Borg, C. Christiansen, R. B. Mazess (1984): Total body bone mineral in vivo by dual photon absorptiometry. I. Measurement procedures. Clin. Physiol. 4: 343-355.
Gotfredsen, A., J. Jensen, J. Borg, and C. Christiansen (1986): Measurement of Lean Body Mass and Total Body Fat Using Dual Photon Absorptiometry. Metabolism 35: 88-93.
Gödeke, Katja, P. Glodek, J.N. Meyer und H. Brandt (1998): Der Einfluß des MHS-Streßgens auf die Fleischleistung und Fleischbeschaffenheit der Nachkommen von Pietrain- und (Pietrain x Hampshire)-Kreuzungsebern. Arch. Tierzucht (Dummerstorf) 41: 433-446.
Graham, R.A., Anne H. Taylor, and T.R. Brown (1994): A method for calculating the distribution of pH in tissues and a new source of pH error from the 31P-NMR spectrum. Am. J. Physiol. 266: R638-R645.
Gray D.S., G.A. Bray, N. Gemayel, K. Kaplan (1989): Effect of obesity on bioelectrical impedance. Am. J. Clin. Nutr. 50: 255-260.
Gregor,G. und T. Hardge (1995): Zum Einfluß von Ryanodin-Rezeptor-Genvarianten auf Spermaqualitätsmerkmale bei KB-Ebern. Arch. Tierzucht (Dummerstorf) 38: 527-538.
Gresham, J.D., S.R. McPeake, J.K. Bernard, and H.H. Henderson (1992): Commercial adaptation of ultrasonography to predict pork carcass composition from live animal and carcass measurements. J. Anim. Sci. 70: 631-639.
Gresham J.D., S.R. McPeake, J.K. Bernard, M.J. Riemann, R.W. Wyatt, H.H. Henderson (1994): Prediction of live and carcass characteristics of market hogs by use of a single longitudinal ultrasonic scan. J. Anim. Sci. 72: 1409-1416.
Griep, W (1991): Schätzung der grobgeweblichen Schlachtkörperzusammensetzung von Schweinen unterschiedlichen Alters mit der Magnet-Resonanz-Tomographie. Diss., Göttingen. 118 S.
Griep, W., E. Hüster, U. Baulain, M. Henning, E. Kallweit (1990): MR (Magnetic - Resonance) - Tomography as a tool for the evaluation of porcine carcasses in relation to growth. 41st Annual Meeting of the EAAP, Toulouse - France, July 8 – 12.
Grier, S.J., A.S. Turner, and M.R. Alvis (1996): The use of Dual-Energy X-ray Absorptiometry in Animals. Investigative Radiology 31: 50-62.
286

Literaturverzeichnis
Groeneveld, E. (1985): Schätzung der Schlachtkörperzusammensetzung lebender Tiere mit neueren Verfahren. Der Tierzüchter 37: 305-306.
Groeneveld, E., E. Kallweit, M. Henning and A. Pfau (1983): Evaluation of Body Composition of Live Animals by X-Ray and NMR Computed Tomography. CEC Meeting: In Vivo Measurement of Body Composition in Meat Animals. Bristol, UK, November 30 - December 1, 84-89.
Groeneveld, E., J. Wolf, M. Wolfová, V. Jelínková und D. Večerová (1998): Schätzung genetischer Parameter für tschechische Schweinerassen mit einem Mehrmerkmals-Tiermodell. Züchtungskunde 70: 96-107.
Gu, Y., A.P. Schinckel, T.G. Martin (1992): Growth, development, and carcass composition in five genotypes of swine. J. Anim. Sci. 70: 1719-1729.
Gunn, T.M., K.A. Miller, L. He, R.W. Hyman, R.W. Davis, A. Azarani, S.F. Schlossmann, J.S. Duke-Cohan, and G.S. Barsh (1999): The mouse mahogany locus encodes a transmembrane form of human attractin. Nature 398: 152-156.
Guo, Sh.S., W. Cameron Chumlea, and D.B. Cockram (1996): Use of statistical methods to estimate body composition. Am. J. Clin. Nutr. 64 (suppl): 428S-435S.
Guo, Sh.S., W. Cameron Chumlea, and R.M. Siervogel (1999): Aging, body composition, and lifestyle: the Fels Longitudinal Study. Am. J. Clin. Nutr. 70: 405-411.
Gwartney, B.L., N.L. Meseck, and C.R. Calkins (1995): The Effects of Orientation and Storage Time on the Prediction of Beef Rib Composition Using Electromagnetic Scanning. J. Anim. Sci. 73: 387-392.
Hahn, Gisela, A. Dobrowolski, R. Dünkel und W. Branscheid (1997): Handelswertbestimmung von Puten: Eignung der Videobildauswertung zur Erfassung der Schlachtkörperzusammensetzung. Jahresbericht Bundesanstalt für Fleisch-forschung (BAFF), Kulmbach: 13-14. http://www.dainet.de:8080/JBBAFF97FORM/DDD/JAHRESBERICHTBAFF1997JB97FGES.PDF.pdf
Hahn, M. (2000): Strahlenschutz – Umweltdaten, Aktuelle Messdaten: Dezember 2000. http://www.brandenburg.de/land/mlur/s/gamma03.htm
Hall, A.D. (1905): The Book of the Rothamsted Experiments. E.P. Dutton and Company, New York http://cdl.library.cornell.edu/CDL/neh_H.html
Hamilton, D.N., M. Ellis, K.D. Miller, F.K. McKeith, and D.F. Parrett (2000): The effect of the Halothane and Rendement Napole genes on carcass and meat quality characteristics of pigs. J. Anim. Sci. 78: 2862–2867.
Hammel, M., E. Müller und D. Fewson (1996): Zuchtexperiment mit DL-Schweinen zur Untersuchung von genetischen Antagonismen zwischen Fleischanteil, Fleischbeschaffenheit und Streßresistenz. Züchtungskunde 68: 384-397.
287

Literaturverzeichnis
Hanset, R., C. Dasnois, S. Scalais, C. Michaux and L. Grobet (1995): Génotypes au locus de sensibilité á l’halothane et caractéres de croissance et de carcasse dans une F2 Piétrain x Large White. Genet. Sel. Evol. 27: 63–76.
Hardge, T., K. Köpke, G. Leuthold, K. Wimmers, and Th. Paulke (1997): Differences in allele frequencies of candidate genes for growth, carcass value and meat quality between extreme phenotypes of commercial pig breeds. 48th Annual Meeting of the EAAP, August 25-28, Vienna (Paper copy, pp. 6).
Hardge, T, K. Köpke, K. Wimmers and G. Leuthold (1998): Association between Polymorphisms of the Leptin-Gene (LEP) and Performance Traits in a porcine ressource family and in commercial outbred populations. Proc. XXVI International Conference on Animal Genetics, 9-14 August, Auckland, NZ, p. 106.
Hardge, T. (1999): Untersuchungen zu den genetischen Ursachen von quantitativen Leistungsmerkmalen mittels Kandidatengenanalyse beim Schwein. Habilitationsschrift, Humboldt-Universität zu Berlin.
Harper, E.J. (1998): Changing Perspectives on Aging and Energy Requirements: Aging, Body Weight and Body Composition in Humans, Dogs and Cats. J. Nutr. 128: 2627S-2631S.
Hartung, E. (1995): Die Bedeutung des Pflanzenalkaloids Ryanodin für die Diagnostik der Malignen Hyperthermie. Habilitationsschrift, Inst. für Anästhesiologie der Universität Würzburg.
Hassen, A., D.E. Wilson, V.R. Amin, and G.H. Rouse (1999): Repeatability of Ultrasound-Predicted Percentage of Intramuscular Fat in Feedlot Cattle. J. Anim. Sci. 77: 1335-1340.
Hausser, K.H. und H.R. Kalbitzer (1989): NMR für Mediziner und Biologen: Strukturbestimmung, Bildgebung, In-vivo-Spektroskopie. Springer-Verlag; Berlin, Heidelberg, New York, London, Paris, Tokyo.
Hazel, L.N. and E.A. Kline (1952): Mechanical measurement of fatness and carcass value on live hogs. J. Anim. Sci. 11: 313-318.
Hegele, R.A. , J.H. Brunt and P.W. Connelly (1995): Genetic variation on chromosome 1 associated with variation in body fat distribution in men. Circulation 92: 1089-1093.
Henen, B.T. (1991): Measuring the lipid content of live animals using cyclopropane gas. Am. J. Physiol. 261: R752-R759.
Henning, M. (1992): Magnetic resonance imaging for the assessment of body composition in pigs. Pig News and Information 13: 163N-166N.
Henning, M., E. Hüster, U. Baulain, E. Kallweit (1991): Evaluation of porcine body composition during growth by means of Magnetic Resonance Tomography (MRT). 42nd Annual Meeting of the EAAP, Berlin, September 8 - 12.
Henning, M., U. Baulain, E. Kallweit, J.C. Forrest (1993): Determination of lean and fat in carcass cuts with TOBEC (Total Body Electrical
288

Literaturverzeichnis
Conductivity). 44th Annual Meeting of the EAAP, Aarhus, Denmark, 16-19 August, 453 (P5.24) + paper copy, p. 5.
Henning, M., U. Baulain, W. Wittmann (1996): Investigation of pork bellies by means of Magnetic Resonance Imaging. 47th Annual Meeting of the EAAP, Lillehammer, Norway, 25-29 August, Book of Abstracts No. 2: 279.
Henning, M., U. Baulain, G. Kohn, and R. Lahucky (2000): In vivo and post mortem changes of muscle phosphorous metabolites in pigs of different Malignant Hyperthermia Genotype. In: Quality of meat and fat in pigs as affected by genetics and nutrition (Eds.: C. Wenk, J.A. Fernández, and M. Dupuis). EAAP publication No. 100: 135-138.
Hermesch, S., B.G. Luxford, H.-U. Graser (2000a): Genetic parameters for lean meat yield, meat quality, reproduction and feed efficiency traits for Australian pigs. 1. Description of traits and heritability estimates. Livest. Prod. Sci. 65: 239-248.
Hermesch, S., B.G. Luxford, H.-U. Graser (2000b): Genetic parameters for lean meat yield, meat quality, reproduction and feed efficiency traits for Australian pigs. 2. Genetic relationships between production, carcase and meat quality traits. Livest. Prod. Sci. 65: 249-259.
Herpin, P., J. Le Dividich and N. Amaral (1993): Effect of selection for lean tissue growth on body composition and physiological state of the pig at birth. J. Anim. Sci. 71: 2645-2653.
Hertrampf, J.W. (1998): Das marktgerechte Schwein vor 200 Jahren – Ein Exkurs in die Vergangenheit. Fleischwirtschaft 78: 862-864.
Heymsfield, S.B., C. Arteaga, C. McManus, J. Smith, S. Moffitt (1983): Measurement of muscle mass in humans: validity of the 24-hour urinary creatinine method. Am. J. Clin. Nutr. 37: 478-494.
Heymsfield, S.B., J. Wang, J. Kehayias, S.Heshka, S. Lichtman, and R.N. Pierson Jr. (1989): Chemical determination of human body density in vivo: relevance to hydrodensitometry. Am. J. Clin. Nutr. 50: 1282-1289.
Heymsfield, S. B., J. Wang, S. Hesjka, J. J. Kehayias, R. N. Pierson (1989): Dual-photon absorptiometry: Comparison of bone mineral and soft tissue mass measurements in vivo with established methods. Am. J. Clin. Nutr. 49: 1283-1289.
Heymsfield, S.B., Z. Wang, R.N. Baumgartner, R.Ross (1997a): Human Body Composition: Advances in Models and Methods. Annu. Rev. Nutr. 17: 527-558.
Heymsfield, S.B., R.Ross, Z. Wang, D. Frager (1997b): Imaging Techniques of Body Composition: Advantages of Measurement and New Uses. In: Emerging Technologies for Nutrition Research: Potential for Assessing Military Performance Capability (Eds.: S.J. Carlson-Newberry and Rebecca B. Costello), National Academy Press, Washington, DC, pp 127-150.
Hiegbi, A.D., T.D. Bidner, J.O. Matthews, L.L. Southern, T.G. Page, M.A. Persica, M.B. Sanders, C.J. Monlezun (1998):
289

Literaturverzeichnis
Prediction of swine body composition by total body electrical conductivity (TOBEC). J. Anim. Sci. 76 (Suppl. 1): 150.
Hirakawa, K., K. Yoshizaki, T. Morimoto and H. Watari (1992): The transition from aerobic to anerobic metabolism during exercise in human skeletal muscle studied by 31P-NMR and electromyography. 11th Annual Scientific Meeting of the Society of Magnetic Resonance in Medicine, Berlin, August 8-14, Book of Abstracts V2: 2724.
Hoeflich, A., M. Wu, S. Mohan, J. Föll, R. Wanke, T. Froehlich, G.J. Arnold, H. Lahm, H.J. Kolb, and E. Wolf (1999): Overexpression of Insulin-Like Growth Factor-Binding Protein-2 in Transgenic Mice Reduces Postnatal Body Weight Gain. Endocrinology 140: 5488-5496.
Hofer, A. (1990): Schätzung von Zuchtwerten feldgeprüfter Schweine mit einem Mehrmerkmals-Tiermodell. Dissertation, ETH Zürich.
Hofer, A. (1998): Variance component estimation in animal breeding: a review. J. Anim. Breed. Genet. 115: 247-265.
Hoffman, E.P. (1995): Voltage-Gated Ion Channelopathies: Inherited Disorders Caused by Abnormal Sodium, Chloride, and Calcium Regulation in Skeletal Muscle. Annu. Rev. Med. 46: 431-441.
Hogreve, F. (1938a): Ausbau eines neuen Forschungsweges zur Bestimmung der Fettwüchsigkeit und Fettleistung in verschiedenen Mastabschnitten beim lebenden Schwein verschiedener Rassenzugehörigkeit. Z. Züchtg.: B Tierzüchtg. u. Züchtungsbiol. 40: 377-395.
Hogreve, F. (1938b): Untersuchungen über die Fettbildung wachsender Mastschweine mittels der Röntgendurchleuchtung. Züchtungskunde 13: 178-191.
Holló, G., I. Repa, J. Tözsér, I Holló, E. Szücs (1998): Estimation of carcass lean meat yield and percentage based on adipocity morphometry and using CT from rib samples. J. Anim. Sci. 76 (Suppl. 1): 143.
Hong, K. Ch. (1985): Der Antagonismus zwischen Fleischleistung und Futteraufnahme in der deutschen Schweinezucht und seine züchterischen Konsequenzen. Diss. Göttingen.
Horn, P. (1991): New methods of in vivo body composition evaluations in the selection of species bred for meat production with special reference to x-ray computerized tomography (RCT). Review article, (Hungarian) Magyar Állatorvosok Lapja 46: 133-137.
Horn, P., Gy. Kövér, I. Repa, E. Berényi, G. Kovách (1996): The use of spiral CAT for estimating in vivo body composition of pigs. 47th Annual Meeting of the EAAP, Lillehammer, Norway, 25-29 August, Paper copy, pp 4.
Horn, P., Gy. Kövér, I. Repa, E. Berényi, G. Kovách (1997): The use of spiral CAT for volumetric estimation of body composition of pigs. Arch. Tierzucht (Dummerstorf) 40: 445-450.
290

Literaturverzeichnis
Hornak, J.P. (1996-2000): “The Basics of MRI” Online Textbook, http://www.cis.rit.edu/htbooks/mri/inside.htm (Stand 22.10.2001).
Hornak, J.P. (1997-2000): "The Basics of NMR" Online Textbook, http://www.cis.rit.edu/htbooks/nmr/inside.htm (Stand 22.10.2001).
Horst, P. (1964a): Entwicklung eines Verfahrens zur Durchführung von Ultraschallmessungen beim lebenden Schwein. Z. Tierz. Züchtungsbiol. 80: 341-364.
Horst, P. (1964b): Kritische Bewertung der Methoden zur Bestimmung des Schlachtkörperwertes beim lebenden Schwein. Z. Tierz. Züchtungsbiol. 80: 365-401.
Horst, P. (1971): Erste Untersuchungsergebnisse über den Einsatz des ”Vidoson”-Schnittbildgerätes beim Schwein. Züchtungskunde 43: 208-218.
Höreth, B.R. (2000): Autofom: Sicherheitskonzept und Überwachung. http://www.isn-ev.de/doku/do8.htm; Copyright 2000 by ISN e.V. bzw. Originalmanuskript aus: Autofom: Neutrale Kontrolle gibt Sicherheit. Landwirtschaftsblatt Weser-Ems, Nr. 5 vom 4. Feb. 2000 sowie Bayerisches Wochenblatt Heft 5 vom 5. Feb. 2000: Bis ins Teilstück vermessen. Nach Stand 01.08.2000 ergänzt und verändert, pdf-Datei, 14 Seiten. http://www.dainet.de/baff/ZulassungAFOM_1_.pdf
Hörnicke, H. (1962): Methoden zur Bestimmung der Körperzusammensetzung lebender Tiere unter besonderer Berücksichtigung des Schweines. V- Die Körperzusammensetzung normal ernährter Schweine im Verlauf des Wachstums. Z. Tierernährung u. Futtermittelkunde 17: 28-60.
Houdebine, L.-M. (1996): Animal Biotechnolgy. STI Review 19: 75-92.
Houghton, P.L. and L.M. Turlington (1992): Application of ultrasound for feeding and finishing animals: a review. J. Anim. Sci. 70: 930-941.
Hoult, D.I., S.J.W. Busby, D.G. Gadian, G.K. Radda, R.E. Richards, and P.J. Seeley (1974): Observation of tissue metabolites using 31P nuclear magnetic resonance. Nature 252: 285-287.
Hounsfield, G.N. (1973): Computerized transverse axial scanning (tomography). Br. J. Radiol. 46: 1016-1021.
Houseman, R.A., I. McDonald, K. Pennie (1973): The measurement of total body water in living pigs by deuterium oxide dilution and its relation to body composition. Br. J. Nutr. 30: 149-156.
Hsu, H., M. Yu Yong, J. W. Babich, J.F. Burke, E. Livni, R.G. Tompkins, V.R. Young, N.M. Alpert, and A.J. Fischman (1996): Measurement of muscle protein synthesis by positron emission tomography with L-[methyl-11C] methionine. Proc. Natl. Acad. Sci. USA 93: 1841-1846.
291

Literaturverzeichnis
Huaroto, J.R., W. Peschke, D. Sprengel, H.M. Blendl, J.H. Weniger und G. Averdunk (1986): Genetische Parameter für Merkmale der Eigenleistungsprüfung von Jungebern in Station. Bayer. Landw. Jahrbuch 63/6: 711-720.
Huxley, Julian S. (1932): Problems of Relative Growth. Lincoln MacVeagh, The Dial press, New York. Albert R. Mann Library. 2000. Core Historical Literature of Agriculture (CHLA). Ithaca, NY: Cornell University. http://chla.library.cornell.edu (Version October 2000). http://chla.library.cornell.edu/cgi-bin/chla/chla-idx?notisid=ADX7659
Hwang, C.-S., Th. M. Loftus, S. Mandrup, M.D. Lane (1997): Adipocyte differentiation and leptin expression. Annu. Rev. Cell. Dev. Biol. 13: 231-259.
Hyun, Y., M. Ellis, G. Riskowski, and R.W. Johnson (1998): Growth Performance of Pigs Subjected to Multiple Concurrent Environmental Stressors. J. Anim. Sci. 76: 721-727.
Irgang, R. (2001): Retrospective and perspective of breeding for carcass and pork quality. Second International Virtual Conference on Pork Quality. November 05 to December 06. http://www.conferencia.uncnet.br/pork/seg/pal/anais01p2_irgang_en.pdf
Ishii, T., R.G. Cassens, K.K. Scheller, S.C. Arp and D.M. Schaefer (1992): Image analysis to determine intramuscular fat in muscle. Food Structure 11: 55-60.
James, T.L. (1999): "Fundamentals of NMR," In: Nuclear Magnetic Resonance (NMR), D. Gorenstein, ed., Online Biophysics Textbook, Biophysical Society. http://biosci.umn.edu/biophys/OLTB/NMR.html
Jamison, W. (2000): Who eats meat: factors effecting pork consumption in Europe and the United States. In: Quality of meat and fat in pigs as affected by genetics and nutrition (Eds.: C. Wenk, J.A. Fernández, and M. Dupuis). EAAP publication No. 100: 101-107.
Janss, L.L.G., J.A.M. van Arendonk and E.W. Brascamp (1994): Identification of a single gene affecting intramuscular fat in Meishan crossbreds using Gibbs sampling. 5th World Congress on Genetics applied to Livestock Production. Guelph, Canada, Vol. 18: 361-364.
Janss, L.L.G., J.A.M. van Arendonk and E.W. Brascamp (1997): Bayesian statistical analyses for presence of single genes affecting meat quality traits in a crossed pig population. Genetics 145: 395-408.
Janzen, E.G. and P.J. Gareau (1993): In vivo proton magnetic resonance imaging (MRI) of liver in young pigs exposed to halothane. Comparison between normal and stress susceptible pigs. Genet. (Life Sci. Adv.) 12: 93-98.
Janzen, E.G., St. Leeson, G.D. Partlow, U.M. Oehler, R.A. Towner (1989): Cross-Sectional Anatomy of the Chicken: An Atlas for Magnetic Resonance Imaging. University of Guelph, MRI Facility.
Janzen, E.G., P.J. Gareau, W.A. Stewart, R.A. Towner (1994): The use of in vivo magnetic resonance imaging and spectroscopy to study porcine
292

Literaturverzeichnis
stress syndrome in young, halothane-susceptible pigs: Preliminary results. Can. J. Anim. Sci. 74: 37-43.
Jebb, S.A. (1997): From chemical analysis of the body to metabolic insights provided by the new methodology. British Journal of Nutrition 78 (Suppl. 2): S101-S112.
Jebb, S.A., G.R. Goldberg, G. Jennings, M. Elia (1995): Dual-energy X-ray absorptiometry measurements of body composition: effects of depth and tissue thickness, including comparisons with direct analysis. Clinical Science 88: 319-324.
Jentzsch, K.-D. (1983): Regulation des Wachstums und der Zellvermehrung. Wissensstand und Probleme. Wissenschaftliche Taschenbücher, Band 283, Akademie Verlag, Berlin.
Jéquier, E. and L. Tappy (1999): Regulation of Body Weight in Humans. Physiological Reviews 79: 451-480.
Jiang, Z. and J.P. Gibson (1999): Genetic polymorphisms in the leptin gene and their association with fatness in four pig breed. Mamm Genome. 10: 191-193.
Jiang, R., X. Chang, B. Stoll, K.J. Ellis, R.J. Shypailo, E. Weaver, J. Campbell, and D.G. Burrin (2000): Dietary Plasma Protein Is Used More Efficiently than Extruded Soy Protein for Lean Tissue Growth in Early-Weaned Pigs. J. Nutr. 130: 2016-2019.
Johansson, M., H. Ellegren and L. Andersson (1995): Comparative mapping reveals extensive linkage conservation -- but with gene order rearrangements -- between the pig and the human genomes. Genomics 25: 682-690.
Johnson L.P., M.F. Miller, K.D. Haydon, J.O. Reagan (1990): The prediction of percentage of fat in pork carcasses. J. Anim. Sci. 68: 4185-4192.
Jopson, N. (1995): Aries, the digital sheep: A Computed Tomography (CT) Atlas of the sheep. AgResearch, Invermay Agricultural Centre, Mosgiel, New Zealand (Software).
Jopson, N.B., K. Kolstad, E. Sehested, and O. Vangen (1995): Computed Tomography as an accurate and cost effective alternative to carcass dissection. Proc. Aust. Assoc. Anim. Breed. Genet. 11: 635-638.
Kalbitz, S. and E. Mueller (1988): Selection for the activity of NADPH-generating enzymes in backfat of pigs. VI. Insulin receptors and glucose metabolism in isolated fat cells. J. Anim. Breed. Genet. 105: 306-316.
Kallweit, E. (1991): Methodische Entwicklung der Wachstumsanalyse bis zur Magnet-Resonanz-Tomographie. Landbauforschung Völkenrode, 41. Jahrgang Heft 4: 208-216.
Kallweit, E. (1992): Tierhaltung und Produktqualität. Züchtungskunde 64: 283-291.
Kallweit, E. (1993): Methodical Development of Growth Analysis up to Magnetic Resonance Imaging. Animal Research and Development 37: 79-93.
293

Literaturverzeichnis
Kallweit, E. (1994): Bedeutung "nicht-invasiver Methoden" zur Messung der Körperzusammensetzung in der Tierproduktion. Landbauforschung Völkenrode, Sonderheft 145: 1-4.
Kallweit, E. (1997): Sicherung einer marktgerechten Fleischerzeugung. Arch. Tierzucht (Dummerstorf) 40 (Sonderheft): 142-147.
Kallweit, E., U. Baulain, K.H. Hoppenbrock (1993): Intramuscular fat in some German pig breeds and crosses. 44th Annual Meeting of the EAAP, Århus, Denmark, September 16 -19, P1.5, 302 (paper copy, pp. 5).
Kallweit, E., H.H. Wesemeier, D. Smidt, U. Baulain (1994): Einsatz der Magnet-Resonanz-Messung in der Tierzuchtforschung. Arch. Tierzucht (Dummerstorf) 37: 105-120.
Kallweit, E., U. Baulain, M. Henning und G :Kohn (1997): In-vivo-Untersuchungen zur Streßanfälligkeit beim Schwein mit Hilfe der Magnet-Resonanz-Spektroskopie. Dtsch. tierärztl. Wschr. 104: 380-383.
Kalm, E. (1998): Qualitätskriterien Fleisch – welche Aspekte nutzt die Zucht- und Produktionsstufe. Lohmann Information 4/98: 9-14.
Kanis, E. (1988): Food intake capacity in relation to breeding and feeding of growing pigs. Thesis, Wageningen Agricultural University.
Karg, H. (1989): Hormone in der Tierproduktion. Teil 1: Einsatzmöglichkeiten und Wirkungsweise von Wachstumshormonen und Lipidmodulatoren. Der Tierzüchter 41: 14-17.
Karlsson, A., Ann-Charlotte Enfält, Birgitta Essén-Gustavsson, Kerstin Lundström, Lotta Rhydmer, and Susan Stern (1993): Muscle Histochemical and Biochemical Properties in Relation to Meat Quality During Slection for Increased Lean Tissue Growth Rate in Pigs. J. Anim. Sci. 71: 930-938.
Karlsson, A.H., R.E. Klont and X. Fernandez (1999): Skeletal muscle fibres as factors for pork quality. In: Quality of meat and fat in pigs as affected by genetics and nutrition (Eds.: C. Wenk, J.A. Fernandez and Monique Dupuis) Wageningen Pers, 2000, EAAP publication No. 100: 47-67.
Kastelic, M. (1997): Bestimmung der Wachstumsallometrie von Muskel- und Fettgewebe aus In-vivo-Messungen beim Schwein. Landbauforschung Völkenrode. Sonderheft 172: 88 S.
Kastelic, M., U. Baulain, E. Kallweit (1996): Allometric growth of muscle and fat areas in German Landrace pigs. 47th Annual Meeting of EAAP, Lillehammer, Norway, Book of Abstracts No. 2: 279.
Kaufmann, A. (1990): Einfluss der Aufzuchtintensität auf die Prüfmast und Vergleich von Prüfverfahren beim Schwein. Dissertation, ETH Zürich.
Kehayias, J.J. and Silvia Valtueña (1999): Neutron activation analysis determination of body composition. Current Opinion in Clinical Nutrition and Metabolic Care 2: 453-463.
294

Literaturverzeichnis
Kehayias, J.J., S.B. Heymsfield, A.F. LoMonte, J. Wang, and R.N. Pierson Jr. (1991): In vivo determination of body fat by measuring total body carbon. Am. J. Clinic. Nutr. 53: 1339-1344.
Kehayias, J.J., Silvia Valtueña, Anathea B. Waitekus, Ch. A. Sheahan, and M. O’Neill (2000): In Vivo Elemental Partition Analysis Using Fast Neutrons – A Tool for Testing the Efficacy of New Clinical Interventions. Ann. N. Y. Acad. Sci. 904: 140-147.
Keightley, P.D., T. Hardge, L. May, G. Bulfield (1996): A Genetic Map of Quantitative Trait Loci for Body Weight in the Mouse. Genetics 142: 227-235
Keim, N.L., P.L. Mayclin, S.J. Taylor, D.L. Brown (1988): Total-body electrical conductivity method for estimating body composition: validation by direct carcass analysis of pigs. Am. J. Clin. Nutr. 47: 180-185.
Kemp, G.J., C.H. Thompson, D.J. Taylor, G.K. Radda (1997): Proton efflux in human skeletal muscle during recovery from exercise. Eur. J. Appl. Physiol. 76: 462-471.
Kern, P.A. (1997): Potential Role of TNFa and Lipoprotein Lipase as Candidate Genes for Obesity. J. Nutr. 127: 1917S-1922S.
Keys, A. and J. Brozek (1953): Body fat in adult man. Physiol. Rev. 33: 245-325.
Khurana, B. (2000): Characterization of DLIM1, a novel cytoskeleton-associated LIM domain containing protein of Dictyostelium discoideum. Diss., Math.-Nat. Fakultät, Universität zu Köln.
Kienzle, Ellen, J. Zentek, and H. Meyer (1998): Body Composition of Puppies and Young Dogs. J. Nutr. 128: 2680S-2683S.
Kim D.H., F.A. SrÈter, S.T. Onishi, J.F. Ryan, J. Roberts, P.D. Allen, L.G. Meazaros, B. Antoniu, and N. Ikemoto (1984): Kinetic studies of Ca2+ release from sarcoplasmic reticulum of normal and malignant hyperthermia susceptible pig muscles. Biochim. Biophys. Acta. 775: 320-327.
Kistorp, C.N. and O.L. Svendsen (1997): Body composition analysis by dual energy X-ray absorptiometry in female diabetics differ between manufacturers. European Journal of Clinical Nutrition 51: 449-454.
Kijas, J., R. Wales, A. Tornsten, P. Chardon, M. Moller, L. Andersson (1998): Melanocortin Receptor 1 (MC1R) mutations and coat colour in pigs. Genetics 150:117-118.
Kleyn, P.W., W. Fan, S.G. Kovats, J.J Lee, J.C. Pulido, Y. Wu, L.R. Berkemeier, D.J. Misumi, L. Holmgren, O. Charlat, E.A. Woolf, O. Taybe, T. Brody, P. Shu, F. Hawkins, B. Kennedy, L. Baldini, C. Ebeling, G.D. Alperin, J. Deeds, N.D. Lakey, J. Culpepper, H. Chen, M.A. Glücksmann-Kuis, G. A. Carlson, G.M. Duyk, and K.J. Moore (1996): Identification and characterization of the mouse obesity gene tubby: a member of a novel gene family. Cell 85:281-290.
295

Literaturverzeichnis
Kliesch, J., U. Neuhaus, E. Silber, and H. Kostzewske (1957): Versuche zur Messung der Speckdicke am lebenden Tier mit Hilfe des Ultraschalls. Zeitschrift für Tierzüchtung und Züchtungsbiologie 70: 29-32.
Klish, W.J., G.B. Forbes, A. Gordon, and W.J. Cochran (1984): New Method for the Estimation of Lean Body Mass in Infants (EMME Instrument): Validation in Monhuman Models. J. Pediatr. Gastroenterol. Nutr. 3: 199-204.
Klont, R.E. and E. Lambooy (1995): Effects of Preslaughter Muscle Exercise on Muscle Metabolism and Meat Quality Studied in Anesthetized Pigs of Different Halothane Genotype. J. Anim. Sci. 73: 108-117.
Klont, R.E., E, Lambooy, and J.G. Logtestijn (1994): Effect of Dantrolene Treatment on Muscle Metabolism and Meat Quality of Anesthetized Pigs of Different Halothane Genotypes. J. Anim. Sci. 72: 2008-2016.
Knapp, P., A. Willam and J. Sölkner (1996): Genetic parameters for lean meat content and meat quality traits in different types of pig breeds. 47th Annual Meeting of EAAP, Lillehammer, Norway, p. 254.
Knight, G.S., A.H. Beddoe, S.J. Streat, and G.L. Hill (1986): Body composition of two human cadavers by neutron activation and chemical analysis. Am. J. Physiol. 250 (Endocrinol. Metab. 13): E179-E185.
Knoflach M., M Kuprian, und G. Walk (2000): RT-Akademie. http://www.tilak.at/azwhome/rtakademie/ct.html.
Knorr, C., M. Schwille, G. Moser, E. Müller, H. Bartenschlager and H. Geldermann (1994): Calcium-release-channel genotypes in several pig populations - associations with halothane and CK reactions. J. Anim. Breed. Genet. 111: 243-252.
Knorr, C., G. Moser, M. Müller, H. Geldermann (1997): Association of GH gene variants with performance traits in F2 generations of European wild boar, Piétrain and Meishan pigs. Animal Genetics 28: 124-128.
Knott, S.A., L. Marklund, C.S. Haley, K. Andersson, W. Davies, H. Ellegreen, M. Fredholm, I. Hansson, B. Hoyheim, K. Lundström, M. Moller, and L. Andersson (1998): Multiple marker mapping of quantitative trait loci in a cross between outbred wild boar and Large White pigs. Genetics 149: 1069-1080.
Knudson B.K., M.G. Hogberg, R.A. Merkel, R.E. Allen, W.T. Magee (1985): Developmental comparisons of boars and barrows: II. Body composition and bone development. J. Anim. Sci. 61: 797-801.
Knyazev, S., K. Zhuchaev, V. Hart, T. Hardge, S. Papshev (1998): About the discordance of halothane reactivity and ryr1 mutation among Siberian swine populations. Proc. 6th World Congress on Genetics Applied to Livestock Production, Armidale, Australia, Vol. 23: 644-647.
Kohn, G. (1997): In-vivo-Untersuchungen zur Muskelphysiologie von Schweinen mit Hilfe der Magnet-Resonanz-Spektroskopie. Landbauforschung Völkenrode, Sonderheft 171: 133 S.
Kohn, G., U. Baulain, M. Henning, R. Lahucky, D. Leibfritz, E. Kallweit (1998): In vivo und post mortem Untersuchungen des Muskelstoffwechsels von Schweinen mit
296

Literaturverzeichnis
unterschiedlichem MHS-Genotyp mit Hilfe der 31P-NMR-Spektroskopie. Arch. Tierz., (Dummerstorf) 41: 299-310.
Kohrt, Wendy M. (1997): Dual-Energy X-Ray Absorptiometry: Research Issues and Equipment. In: Emerging Technologies for Nutrition Research: Potential for Assessing Military Performance Capability (Eds.: S.J. Carlson-Newberry and Rebecca B. Costello), National Academy Press, Washington, DC, pp 151-167.
Kolb, E. (1987): Neue biochemische Erkenntnisse zur Entstehung der Streßanfälligkeit und zu den Veränderungen bei Schweinen mit blasser, weicher und wäßriger Fleischqualität (PSE-Fleisch). Tierzucht (Berlin) 41: 557-559.
Kolb, R. (1991): Analyse digitalisierter Ultraschallbilder an lebenden Schweinen zur Abschätzung des Fleischanteils im Bauch und des intramuskulären Fettgehaltes im Rückenmuskel. Diss., Univ. Hohenheim.
Kolstad, K. (1996): Genetic differences in fat mobilisation of growing pigs fed at maintenance, and the use of blood components as indicators of changes in body composition. 47th Annual Meeting of the EAAP, Lillehammer, Norway, 25-29 August, Book of Abstracts No. 2: 276.
Kolstad, K. (2000): Fat deposition and distribution in three genetic lines of pigs from 10 to 105 kilograms liveweight. In: Quality of meat and fat in pigs as affected by genetics and nutrition (Eds.: C. Wenk, J.A. Fernández, and M. Dupuis), Wageningen Pers, EAAP publication No. 100: 199-202.
Kolstad, N. and O. Vangen (1996): Breed differences in maintenance requirements of growing pigs when accounting for changes in body composition. Livest. Prod. Sci. 47: 23-32.
Kolstad, N., N.B. Jopson and O. Vangen (1996): Breed and sex differences in fat distribution and mobilization in growing pigs fed at maintenance. Livest. Prod. Sci. 47: 33-41.
Koretsky, A.P. and D.S. Williams (1992): Application of localized in vivo NMR to whole organ physiology in the animal. Annu. Rev. Physiol. 54: 799-826.
Kortz, J., W. Kapelanski, S. Grajewskaja, J. Kuryl, M. Bocian, and A. Rybarczyk (2000): Meat quantity to meat quality relationships when the RYR1 gene effect is eliminated. In: Quality of meat and fat in pigs as affected by genetics and nutrition (Eds.: C. Wenk, J.A. Fernández, and M. Dupuis). EAAP publication No. 100: 143-146.
Kopin, A.S., W. Foulds Mathes, E.M. McBride, M. Nguyen, W. Al-Haider, F. Schmitz, S. Bonner-Weir, R. Kanarek, and M. Beinborn (1999): The cholecystokinin-A receptor mediates inhibition of food intake yet is not essential for the maintenance of body weight. J. Clin. Invest. 103: 383-391.
Kouba, M., M. Bonneau, J. Noblet (1999): Relative development of subcutaneous, intermuscular, and kidney fat in growing pigs with different body compositions. J. Anim. Sci. 77: 622-629.
297

Literaturverzeichnis
Kozak-Reiss, G., F. Desmoulin, P. Canioni, P. Cozzone, J.P. Gascard, G. Monin, J.M. Pusel, J.P. Renou, A. Talmant (1987): Contraction and metabolism traits in skeletal muscle biopsies from halothane-positive pigs as studied by mechanical measurements and 31P NMR. In: "Evaluation and Control of meat quality in pigs" (Ed. P. V. Tarrant, G. Eikelenboom and G. Monin), Martinus Nijhoff Publishers, The Hague: 27-38.
Kozak-Ribbens, G., D. Bendahan, L. Rodet, S. Confort-Gouny, A. Miri, A. Talmant, G. Monin, and P.J. Cozzone (1997): Metabolic events with spontaneous malignant hyperthermia crisis in an anaesthetized pig. Can. J. Anaesth. 44: 757-764.
Kraybill, H.F., E.R. Goode, R.S.B. Robertson, and H.S. Sloane (1953): In vivo measurement of body fat and body water in swine. J. Appl. Physiol. 6: 27-32.
Kräußlich, H. (1998): Geschichte der Tierbeurteilung. In: Exterieurbeurteilung landwirtschaftlicher Nutztiere (Hrsg.: G. Brem), Verlag Eugen Ulmer.
Krempler, Andrea (2000): Analyse der RyREF2-Bindungsstelle im Promotor des porcinen RyR1 Gens und Charakterisierung des porcinen FHL1-Gens. Dissertation, Georg-August-Universität zu Göttingen, Göttingen. http://webdoc.gwdg.de/diss/2000/krempler/
Krempler, A., S. Kollers, R. Fries and B. Brenig (2000): Isolation and characterization of a new FHL1 variant (FHL1C) from porcine skeletal muscle. Cytogenet Cell Genet 90: 106–114.
Kresken, J.-G., U. Baulain und R.G. Köstlin (1993): Kernspintomographie, Magnet-Resonanz-Tomographie (MRT), Nuclear Magnetic Resonance Imaging (NMR): Technische und methodische Voraussetzungen zur Anwendung der Kernspintomographie am Hüftgelenk des Hundes. Tierärztl. Prax. 21: 559-565.
Krieter, J. (1986): Entwicklung von Selektionsmethoden für das Wachstum und Futteraufnahmevermögen beim Schwein. - Diss. Kiel. - Schriftenreihe des Instituts für Tierzucht und Tierhaltung der Christian-Albrechts-Universität zu Kiel, Heft 31.
Krieter, J. und E. Kalm (1986): Futteraufnahme aus der Sicht der Tierzucht. Betriebswirtschaftliche Mitteilungen der Landwirtschaftskammer Schleswig-Holstein Nr. 370: 3-12.
Krieter, J. und E. Tholen (2001): Berücksichtigung der Fleischqualität bei der Selektion innerhalb Linien beim Schwein – eine Studie. Arch. Tierz. (Dummerstorf) 44: 531-546.
Krieter, J., T. Hölscher, P. Hartjen, E. Kalm und E. Ernst (1990): Vergleich von Ultraschallverfahren zur Abschätzung der Schlachtkörper-zusammensetzung am Schwein. Züchtungskunde 62: 29-37.
Krska, P., R. Lahucky, U. Küchenmeister, Karin Nürnberg and Gerda Kuhn (2000): Post-mortem muscle metabolism of phosporous compounds assessed by 31P NMR spectroscopy in relation to vitamin E administration of pigs genotyped on malignant hyperthermia and stress susceptibility. Arch. Tierz. (Dummerstorf) 43 (Sonderheft): 229.
298

Literaturverzeichnis
Kromeyer-Hauschild, Katrin und U. Jaeger (1998): Zunahme der Häufigkeit von Übergewicht und Adipositas bei Jenaer Kindern. Monatsschr. Kinderheilkd. 146: 1192–1196.
Kronacher, C. und F. Hogreve (1936): Beiträge zur Kenntnis der Grundlagen der Beckenformen bei verschiedenen Schweinerassen, gewonnen an Hand röntgenologischer Studien. Z. Zücht.: B Tierzüchtg. u. Züchtungsbiol. 35: 161-167.
Krude, H. and A. Grüters (2000): Implications of Proopiomelanocortin (POMC) Mutations in Humans: The POMC Deficiency Syndrome. Trends in Endocrinology and Metabolism 11: 15-22.
Kruger, P. and I.J. Gruverman (1962): Neutron activation analysis of homogenized chicken. Appl. Radiat. Isot. 13: 106-110.
Kuchida, K., K. Konishi, M. Suzuki and S. Miyoshi (1998): Prediction of the Crude Fat Contents in Rib-Eye Muscle of Beef Using the Fat Area Ratio Calculated by Computer Image Analysis. Anim. Sci. Technol. (Jpn) 69: 585-588.
Kuchida, K., S. Tsurata, L.D. van Vleck, M. Suzuki and S. Miyoshi (1999): Prediction Method of Beef Marbling Standard Number Using Parameters Obtained from Image Analysis for Beef Ribeye. Animal Science Journal 70: 107-112.
Kuchida, K., S. Kono, K. Konishi, L.D. van Vleck, M. Suzuki, and S. Miyoshi (2000): Prediction of crude fat content of longissimus muscle of beef using the ratio of fat area calculated from computer image analysis: Comparison of regression equations for prediction using different input devices at different stations. J. Anim. Sci. 78: 799-803.
Kuhn, G., M. Hartung, H. Falkenberg, G. Nürnberg, M. Langhammer, M. Schwerin, K. Ender (1997): Wachstum, Körperzusammensetzung und Fleischbeschaffenheit von im Fettansatz genetisch differenten Schweinen. Arch. Tierzucht (Dummerstorf) 40: 345-355.
Kuhn, G., M. Hartung, K. Nürnberg, I. Fiedler, H. Falkenberg, G. Nürnberg, K. Ender (1998): Körperzusammensetzung und Muskelstruktur von genetisch differenten Schweinen in Abhängigkeit vom MHS-Status. Arch. Tierzucht (Dummerstorf) 41: 589-596.
Kuhn, M., L. Beesten, C. Jatsch (1997): Zum Einfluß der Fütterungsintensität und des Mastendgewichts auf die Mast- und Schlachtleistung von Schweinen sowie das Fettsäuremuster der Gesamt- und Phospholipide des M. long. dorsi – 1. Mitteilung: Parameter der Mast- und Schlachtleistung, der Fleischbeschaffenheit sowie der Trockensubstanz – und Aschegehalte der Depotfette. Züchtungskunde 69: 294-306.
Kuhlig, Hanna, W. Grezesiak and Iwona Szatkowska (2001): Effect of leptin gene polymorphism on growth and carcass traits in pigs (short communication). Arch. Tierz. (Dummerstorf) 44: 291-296.
Küchenmeister, U. und K. Ender (1994): Klassifizierung mit Ultraschall vor dem Enthäuten. Landbauforschung Völkenrode, Sonderheft 145: 15-24.
Küchenmeister U., Gerda Kuhn, J. Wegner, G. Nürnberg, and K. Ender (1999): Post mortem changes in Ca2+ transporting proteins of sarcoplasmic reticulum in dependence on malignant hyperthermia status in pigs. Mol. Cell Biochem. 195: 37-46.
299

Literaturverzeichnis
Kwon, H.Y., S.J. Bultman, C. Loffler, W.J. Chen, P.J. Furdon, J.G. Powell, A.L. Usala, W. Wilkison, I. Hansmann, and R.P. Woychik (1994): Molecular structure and chromosomal mapping of the human homolog of the agouti gene. Proc. Natl. Acad. Sci. USA 91: 9760-9764.
Lahucky, R., V. Madajová, O. Mharapara, J. Marák, D. Kaniansky, J. Poltarsky, and L. Kovac (2000a): A Comparison of Some Metabolites in Skeletal Muscle of Different Halothane Sensitivity with Meat Quality Traits of Pigs Using a Capillary Isotachophoresis Technique. Meat Sci. 39: 403-414.
Lahucky, R., P. Krska, U. Küchenmeister, Karin Nürnberg, T. Liptaj, G. Nürnberg, I. Bahelka, P. Demo, Gerda Kuhn, and K. Ender (2000b): Effect of Vitamin E on changes in Phosphorous Compounds assessed by 31P NMR Spectroscopy and ATPase from postmortem Muscle samples and Meat quality of Pigs. Arch. Tierzucht (Dummerstorf) 43: 487-497.
Lampo, Ph, A. Calus, W. Nauwynck and Y. Bouqet (1985): De fokwaarde- beoordeling van varkens op grond van selectiemesterijresultaten. II. De genetische en phenotypische parameters voor fokkerij - versnijdingsresultaten. Landbouwtijdschrift 38: 403-411.
Langlet, J.F., E. Ernst, H.D. Glaner (1968): Die Möglichkeit einer Selektion bei Schweinen mit Hilfe von Echolotmessungen unter den Verhältnissen der Praxis. Zeitschrift f. Tierzüchtung und Züchtungsbiologie 85: 27-35.
Langholz, H.-J. (1966): Das züchterische Hilfsmittel der stationären Nachkommenprüfung beim Schwein. III. Heritabilitäten und genetische Korrelationen beim norwegischen Landschwein. Acta Agricultrae Scandinavica 16: 97-114.
Lapillonne, A. P.M. Braillon, P.D. Delmas, B.L. Salle (1997): Dual-Energy X-Ray Absorptiometry in Early Life. Horm. Res. 48 (suppl. 1): 43-49.
Larcombe-McDouall, J.B., N. Harrison, S. Wray (1998): The in vivo relationship between blood flow, contractions, pH and metabolites in the rat uterus. Plügers Archiv - Eur. J. Physiol. 435: 810-817.
Laskey, M.A. (1996): Dual-Energy X-Ray Absorptiometry and Body Composition. Nutrition 12: 45-51.
Lauterbur, P.C. (1973): Image formation by induced local interactions: Examples employing nuclear magnetic resonance. Nature 242: 190-191.
Laube, Sigurd (2000): Die Eignung spezieller Schweinekreuzungen zur Qualitätsverbesserung von Markenschweinefleisch unter besonderer Berücksichtigung von MHS-Status, Hampshirefaktor und intramuskulärem Fettgehalt. Dissertation, TiHo Hannover, 78 S.
Lawes, J.B. and J.H. Gilbert (1858): Pamphlets by Lawes and Gilbert: Experimental inquiry into the composition of some of the animals fed and slaughtered as human food. Abstract from Proceedings of the Royal Society, June 17, 1858, London, Taylor and Francis, 16 p.
300

Literaturverzeichnis
Lawes, J.B. and J.H. Gilbert (1861): Rothamsted Agricultural Experiments: Lawes and Gilbert: On the composition of oxen, sheep and pigs and of their increase whilst fattening. Reprinted from Journal of the Royal Agricultural Society of England, Vol. 21, Part 2, London, W. Clowes and sons, 58 p.
Lawes, J.B. and J.H. Gilbert (1877): Rothamsted Agricultural Experiments: Lawes and Gilbert: On the formation of fat in the animal body. Reprinted from Journal of Anatomy and Physiology, Vol.11, Part 4., Cambridge, CUP, 11 p.
Lawes, J.B. and J.H. Gilbert (1883): Rothamsted Agricultural Experiments: Lawes and Gilbert: Supplement to former paper entitled "Experimental inquiry into the composition of some of the animals fed and slaughtered as human food" - composition of the ash of the entire animals and of certain separated parts. Reprinted from Proceedings of the Royal Society No. 226, 3 p.
Lefaucheur, L., P. Buche, P. Ecolan, and M. Lemoing (1992): Classification of Pig Myofibres and Assessment of Post-Mortem Glycogen Depletion According to Fibre Type by Compterized Image Analysis. Meat Sci. 32: 267-278.
Leibfritz, D. (1996): An Introduction to the Potential of 1H-, 31P- and 13C-NMR-Spectroscopy. Anticancer Research 16: 1317-1324.
Le Roy, P., C. Moreno, J.M. Elsen, J.C. Caritez, Y. Billon, H. Lagant, A. Talmant, P. Vernin, Y. Amigues, P. Sellier, and G. Monin (2000): Interactive effects of the HAL and RN major genes on carcass quality traits in pigs: preliminary results. In: Quality of meat and fat in pigs as affected by genetics and nutrition (Eds.: C. Wenk, J.A. Fernández, and M. Dupuis). EAAP publication No. 100: 139-142.
Leymaster, K.A. (1986): Tomography to estimate changes in body tissues. J. Anim. Sci. 63(Suppl. 2): 89-95.
Li, Jianhua (2000): Multifrequente Impedanztomographie zur Darstellung der elektrischen Impedanzverteilung im menschlichen Thorax. Dissertation, Universität Stuttgart.
Li, X. and B.W. Kennedy (1994): Comparison of genetic parameter estimates for growth rate and backfat from single and multiple trait models with and without genetic groups. 5th World Congress on Genetics applied to Livestock Production. Guelph, Canada, Vol. 18: 418-421.
Lierse, Ch. (1998): Spurenanalyse für Technik und Umwelt. FRM-II-Symposium: Neutronen für Industrie und Medizin, München, 9. Juli 1998. http://www.frm2.tu-muenchen.de/d/presse/resi98/resi98-01.html
Limper, J., G. Reiner, W. Hecht und V. Dzapo (1991): Mitochondrialer (mt) Energiestoffwechsel im Diaphragma von HAL+ und HAL--Schweinen und die vermutete Beteiligung der mt-DNA an der Halothansensitivität. J. Anim. Breed. Genet. 108: 389-398.
Lindgren, Uta (2001): Experimente mit den X-Strahlen. Bayerns innovative Köpfe: Wilhelm Conrad Röntgen. Unser Bayern 50: 39-41.
301

Literaturverzeichnis
Lindner, R. (1991): Untersuchungen über die Beziehungen zwischen Glykolyse und Fleischbeschaffenheit beim Schwein - zugleich Beschreibung der Basis eines Selektionsexperimentes. Dissertation Universität Hohenheim. 179 S.
Lirette, A., R.A. Towner, Z. Liu, U.M. Oehler, E.G. Janzen, R.J. Etches, J.R. Chambers, R.W. Fairfull, S. Chu, A. Rath, J. Millam, B. Cutler, D.C. Crober (1993a): Magnetic resonance imaging (1H) and spectroscopy (1H, 31P) of growing chicken embryos. Can. J. Anim. Sci. 73: 953-965.
Lirette, A., R.A. Towner, Z. Liu, E.G. Janzen, J.R. Chambers, R.W. Fairfull, L.P. Milligan, D.C. Crober (1993b): In Vivo Nuclear Magnetic Resonance Spectroscopy of Chicken Embryos from Two Broiler Strains of Varying Fat Content. Poultry Science 72: 1411-1420.
Lister, D. (1976): Effects of nutrition and genetics on the composition of the body. Proc. Nutr. Soc. 35: 351-356.
Lister, D. (1987): The physiology and biochemistry of the porcine stress syndrome. In: Evaluation and Control of Meat Quality in Pigs. (Eds.: P.V. Tarrant, G. Eikelenboom, and G. Monin), Martinus Nijhoff Publishers, Boston, MS, USA: 3-16.
Liu, H.C., M.F. Rothschild and C.K. Tuggle (1995): Analysis of pig chromosome 7 genetic markers for growth and carcass performance traits. J. Anim. Sci. 73 (Suppl. 1): 41.
Liu, Y. and J.R. Stouffer (1995): Pork carcass evaluation with an automated and computerized ultrasonic system. J. Anim. Sci. 73: 29-38
Liu, Z., A. Lirette, R.A. Towner, R.W. Fairfull, J.R. Cahambers, E.G. Janzen, L.P. Milligan, D.C. Crober (1992): Nuclear magnetic resonance spectroscopy (Phosphorous-31) of chicken embryos of two broiler strains with different carcass fat content. Proc. XIX World’s Poultry Congress, Amsterdam, Netherlands, 20-24 September, Volume 3: 471- 475.
Liu, Z., A. Lirette, R.W. Fairfull, B.W. McBride (1994): Embryonic Adenosine Triphosphate: Phosphodiesteres Ratios Obtained with In Vivo Nuclear Magnetic Resonance Spectroscopy (Phosphorous-31): A New Technique for Selecting Leaner Broiler Chickens. Poultry Science 73: 1633-1641.
Lloyd, R.D. and Ch.W. Mays (1987): A Model for Human Body Composition by Total Body Counting. Hum. Biol. 59:7-30.
Lobley, G.E. (1998): Nutritional and hormonal control of muscle and peripheral tissue metabolism in farm species. Livest. Prod. Sci. 56: 91-114.
Lohmann, T.G. (1984): Research Progress in validation of laboratory methods of assessing body composition. Med. Sci. Sports Exerc. 16: 596-603.
302

Literaturverzeichnis
Lohmann, T.G., M.H. Slaughter, R.A. Boileau, J. Bunt and L. Lussier (1984): Bone mineral measurements and their relation to body density in children, youth and adults. Hum. Biol. 56: 667-679.
López Fernández, J.L., A. Arguello Henríquez, J. Capote Alvarez, N. Darmanin Garrido (1992): Contribution to the study of Black Canary Pigs. Arch. Zootec. 41 (extra): 531- 536.
Louis, C.F., J.R. Mickelson and W.E. Rempel (1994): The effect of skeletal muscle ryanodine receptor (ryr1) genotype on swine performance and carcass traits. J. Anim. Sci. 72 (Suppl. 1): 248.
Louveau, I., S. Schnoebelen and M. Bonneau (1995): Regulation of growth and body composition by growth hormone and related homones in pigs. Proc. 2nd Dummerstorf Muscle-Workshop Muscle Growth and Meat Quality. FBN Schriftenreihe 6: 207-214.
Lowell, B.B. and J.S. Flier (1997): Brown Adipose Tissue, ß3-Adrenergic Receptors, and Obesity. Annu. Rev. Med. 48: 307-316.
Luce, W.G.: Measuring Carcass Traits in Live Hogs. Cooperative Extension Service • Division of Agriculture • Oklahoma State University, No. 3662 (AC-321 0491 RO Revised).
Luiting, P. (1993): The biological nature of genetic variation in net feed efficiency. 44th Annual Meeting of the EAAP, Aarhus (Denmark), 16-19 August, Paper, 9 p.
Luiting, P., H. Enting, O. Vangen (1992): Breed differences in maintenance requirements in pigs measured by computer tomography. 43rd Annual Meeting of the EAAP, Madrid, Spain, 13.-17. September, Volume 1, 164.
Luiting, P., K. Kolstad, H. Enting, O. Vangen (1995): Pig breed comparison for body composition at maintenance: analysis of computerized tomography data by mixture distributions. Livest. Prod. Sci. 43: 225-234.
Lukaski, H.C. (1987): Methods for the assessment of human body composition: Traditional and new. Am. J. Clin. Nutr. 46: 537-556.
Lukaski, H.C. (1996): Biological indexes considered in the derivation of the bioelectrical impedance analysis. Am. J. Clin. Nutr. 64 (suppl.): 397S-404S.
Lukaski, H.C., Ph.E. Johnson, W.W. Bolonchuk, and I.G. Lykken (1985): Assessment of fat-free mass using bioelectrical impedance measurements of the human body. Am. J. Clin. Nutr. 41: 810-817.
Lukaski, H.C., M.J. Marchello, C.B. Hall, D.M. Schafer, and W.A. Siders (1999): Soft Tissue Composition of Pigs Measured With Dual X-Ray Absorptiometry: Comparison With Chemical Analyses and Effects of Carcass Thicknesses. Nutrition 15: 697-703.
Luke, Amy H., Ch.N. Rotimi, R.S. Cooper, A.E.Long, T.E. Forrester, R. Wilks, F.I. Bennett, O. Ogunbiyi, J.A. Compton, and R.R. Bowsher (1998):
303

Literaturverzeichnis
Leptin and body composition of Nigerians, Jamaicans, and US blacks. Am. J. Clin. Nutr. 67: 391-396.
Lutz, N.W. und E. Schultz (1987): Beschreibung des MR-Relaxationsverhaltens von Gewebe mit Hilfe einer geeigneten Modellvorstellung. Digit. Bilddiagn. 7: 56-60.
Lüning, M., J. Planitzer, K.-H. Sandring, B. Schnackenburg, C. Dewey (1988): Magnetresonanz-Tomographie - ein neues bildgebendes Verfahren. Medizin aktuell 14: 266-269.
Ma, J. and J. Zhao (1994): Highly cooperative and hysteretic response of the skeletal muscle ryanodine receptor to changes in proton concentrations. Biophysical Journal 67: 626-633.
Maak, S., M. Wicke, D.C. Beitz, G. von Lengerken (1995): Suitability of a shot biopsy test for prediction of meat quality in live pigs at different ages. J. Anim . Sci. 73 (Suppl. 1): 170.
Maassen-Francke, B., J. Krieter, E. Kalm (1992): Untersuchungen zur Mastleistung und Fleischbeschaffenheit fast vergessener Schweinerassen. Schweinezucht und Schweinemast 40: 44-48.
Mackay, Trudy F.C. (2001): The genetic architecture of quantitative traits. Annu. Rev. Genet. 35: 303-339.
Mackie, A., W.J. Hannan, and P. Tothill (1989): An introduction to body composition models used in nutritional studies. Clin. Phys. Physiol. Meas. 10: 297-310.
MacLennan, D.H. and M.S. Phillips (1992): Malignant Hyperthermia. Science (Washington, DC) 256: 789-794.
Madsen, O.R., J.-E.B. Jensen, O.H. Sørensen (1997): Validation of a dual energy X-ray absorptiometer: measurement of bone mass and soft tissue composition. Eur. J. Appl. Physiol. 75: 554-558.
Mahan, D.C. and R.G. Shields, Jr. (1998): Macro- and Micromineral Composition of Pigs from Birth to 145 Kilograms of Body Weight. J. Anim. Sci. 76: 506-512.
Mailänder, Ch. (1988): Ergebnisse eines Zuchtexperimentes zur Ermittlung der Effektivität der Selektion nach biochemischen Parametern oder Ultraschallmassen zur Verbesserung der Schlachtkörperqualität beim Schwein. Dissertation, Univ. Hohenheim.
Malik, R.C. (1984): Genetic and physiological aspects of growth, body composition and feed efficiency in mice: A Review. J. Anim. Sci. 58: 577-590.
Manteuffel, G and B. Puppe (2000): Animal well-being in husbandry and coping with stress: Introductory remarks. Arch. Tierz. (Dummerstorf) 43 (Sonderheft): 140-143.
Marchello M.J. and W.D. Slanger (1992): Use of bioelectrical impedance to predict leanness of Boston butts. J. Anim. Sci. 70: 3443-3450.
304

Literaturverzeichnis
Marchello, M.J., W.D. Slanger, and J.K. Carlson (1999): Bioelectrical impedance: fat content of beef and pork from different size grinds. J. Anim. Sci. 77: 2464-2468.
Martens, H. (1997): Physiologie und Pathophysiologie des Ryanodin-Rezeptors beim Schwein - Bedeutung für die Streßempfindlichkeit, Belastungsmyopathien, maligne Hyperthermie und die Qualität des Fleisches. Tierärztl. Prax. 25: 41-51.
Martens, H. (1998): Physiologie der Muskulatur und das MHS-Gen des Schweines: Zur Diskussion um eine Eliminierung des mutierten Ryanodin-Rezeptors aus der deutschen Schweinezucht. Arch. Tierzucht (Dummerstorf) 41: 179-192.
Mason, P.A., T.J. Walters, J.W. Fanton, D.N. Erwin, J.H. Gao, J.W. Roby, J.L. Kane, K.A. Lott, L.E. Lott, R.V. Blystone (1995): Database created from magnetic resonance images of a Sprague-Dawley rat, rhesus monkey, and pigmy goat. FASEB J. 9: 434-440.
Matthie, J., B. Zarowitz, A. de Lorenzo, A. Andreoli, K. Katzarski, G. Pan, and P. Withers (1998): Analytic assessment of the various bioimpedance methods used to estimate body water. J. Appl. Physiol. 84: 1801–1816.
Mayrleitner, M., A.P. Timerman, G. Wiederrecht und S. Fleischer (1994): The calcium release channel of sarcoplasmic reticulum is modulated by FK-506 binding protein: effect of FKBP-12 on single channel activity of the skeletal muscle ryanodine receptor. Cell Calcium 15: 99-108.
Mayrhofer, E., W. Henninger, K. Hittmair und S. Kneissl (1995): Röntgenstrahlen, Ultraschall, Magnetresonanz - Möglichkeiten ihres Einsatzes in der Veterinärmedizin. Wien. Tierärztl. Mschr. 82: 276-288.
Mazess, R. B., H. S. Barden, J. P. Bisek, J. Hanson (1990): Dual energy X-ray absorptiometry for total-body and regional bone-mineral and soft-tissue composition. Am. J. Clin. Nutr. 51: 1106.
McBride, B.W., S. Leeson, R.A. Towner, E.G. Janzen (1991): Application of magnetic resonance imaging and 31P spectroscopy to the study of body composition and energy metabolism in poultry. Proc. 12th Symposium Energy Metabolism of Farm Animals, Kartause Ittingen, Switzerland, 1-7 September, 234-237.
McLaren, D.G. and Buchanan, D.S. (1991): Impact of metabolism modifiers on animal breeding considerations. J. Anim. Sci. 69 (Suppl. 2): 100-115.
McMeekan, C.P. (1940): Growth and development in the pig, with special reference to carcass quality characters. Part I. J. Agric. Sci. 30: 266-343.
McPherron, A.C., A.M. Lawler, and S.-J. Lee (1997): Regulation of skeletal muscle mass in mice by a new TGF-ß superfamily member. Nature 387: 83-90.
Meissner, G. (1994): Ryanodine receptor/Ca2+ release channels and their regulation by endogenous effects. Annu. Rev. Physiol. 56: 485-508.
305

Literaturverzeichnis
Menzel, M. (1997): Ströme in Gebieten mit variabler Zusammenhangszahl. Diss., FB Physik, Universität Kaiserslautern, 110 Seiten.
Merks, J.W.M., E.H.T. Hanenberg and A.J.M. van Erp (1997): Effects of selection for leanness in pigs on genetic parameters and ultrasonic backfat thickness. 48th Annual Meeting of EAAP, Vienna, Austria. 25-28 August 1997, paper copy, 8 pages, author address; J.W.M. Merks, Institute for Pig Genetics B.V., P.O. Box 43, 6640 AA Beuningen, The Netherlands.
Mersmann, H.J. (1982): Ultrasonic determination of backfat depth and loin area in swine. J. Anim. Sci. 54: 268-275.
Mersmann, H.J., L.J. Brown, E.Y. Chai, and T.J. Fogg (1984): Use of electronic meat measuring equipment to estimate body composition in swine. J. Anim. Sci. 58: 85-93.
Meseck, N.L., B.L. Gwartney, C.R. Calkins, and P.S. Müller (1997): Influence of Sample Orientation on Prediction of Fresh Ham Lean Content by Electromagnetic Scanning. J. Anim. Sci. 75: 3169-3173.
Messer, L.A., E.J. Cargill, and D. Pomp (2001): Rapid communication: Mapping of the pro-opiomelanocortin (POMC) gene to porcine chromosome by linkage analysis using a PCR-RFLP. J. Anim. Sci. 79: 2241–2242.
Mettau, J.W., H.J. Degenhardt, H.K.A. Visser, and W.P.S. Holland (1977): Measurement of Total Body Fat in Newborns and Infants by Absorption and Desorption of Nonradioactive Xenon. Pediatric Res. 11: 1097-1101.
Metz, S.H.M (1985): Genetic effects on fat deposition and fat quality in the growing pig. Pig News and Information 6: 291-294.
Meynial-Denis, D., M. Mignon, L. Foucat, G. Bielecki, A. Ouali, C. Tassy, J.-P. Renou, J. Grizard, M. Arnal (1998): PH is regulated differently by glucose in skeletal muscle from fed and starved rats:A study using 31P-NMR spectroscopy. J. Nutr. 128: 28-34.
Mickelson, J.R. and C.F. Louis (1993): Calcium (Ca2+) Regulation in Porcine Skeletal Muscle - Review. In: ”Pork Quality: Genetic and Metabolic Factors” (Ed. E. Puolanne, D.I. Demeyer, M. Ruusunen; S. Ellis), CAB International, Wallingford, p. 160-184.
Mickelson, J.R. and C.F. Louis (1996): Malignant hyperthermia: Excitation-Contraction Coupling, Ca 2+ Release Channel, and Cell Ca 2+ Regulation Defects. Physiol. Rev. 76: 537-592.
Mikoshiba, K. (1997): The InsP3 receptor and intracellular Ca2+ signaling. Current Opinion in Neurobiology 7: 339-345.
Milan, D. J.P. Bidanel, P.Le Roy, C. Chevalet, N. Woloszyn, J.C. Caritez, J. Gruand, H.Lagant, M. Bonneau, L. Lefaucheur, C. Renard, M. Vaiman, P. Mormède, C. Désautés, Y. Amigues, F. Bourgeois, J. Gellin, L.Ollivier (1998): Current status of QTL detection in Large White x Meishan Crosses in France.
306

Literaturverzeichnis
Proceeding of the 6th World Congress on Genetics Applied to Livestock Production, Armidale, Australia, January 11-16, Volume 26: 414-417.
Miller, D.C. (1996): Accuracy and Application of Real-Time Ultrasound for Evaluation of Carcass Merit in Live Animals. In: Animal Science Facts. Publication Number: ANS 96-010B. http://mark.asci.ncsu.edu/Publications/factsheets/010b.htm.
Miltenberger, R.J., R.L. Mynatt, J. Erby Wilkinson, and R.P. Woychik (1997): The Role of the agouti Gene in the Yellow Obese Syndrome. J. Nutr. 127:1902S-1907S.
Mitchell, A.D. and J.M. Conway (1993): Body composition analysis of pigs by dual energy x-ray absorptiometry (DEXA). J. Anim. Sci. 71 (Suppl. 1): 146.
Mitchell, A.D. and J.M. Conway (1994): Measurement of Energy Deposition and Body Composition of Pigs by Dual-Energy X-Ray Absorptiometry (DEXA). Proc. 13th Symposium on Energy Metabolism of Farm Animals, September 18-24, Mojácar, Spain, 31-34.
Mitchell, A.D. and A.M. Scholz (1995): Analysis of total body and carcass composition of pigs of different ryanodine receptor genotypes by dual energy x-ray absorptiometry (DEXA). J. Animal Sci. 73, (Suppl. 1), 162.
Mitchell, A.D. and A.M. Scholz (1997): Dual-energy X-ray absorptiometry (DXA) analysis of growth and composition of pigs of different ryanodine receptor genotypes. Arch. Tierzucht (Dummerstorf) 40: 47-56.
Mitchell, A.D. and A.M. Scholz (1998): Energy Deposition and Body Composition Measurement of Pigs of Different Ryanodine Receptor Genotypes by Dual Energy X-Ray Absorptiometry. In: Energy Metabolism of Farm Animals (McCracken, Unsworth and Wylie, Eds.). Proc. of the 14th Symposium on Energy Metabolism, September 14 - 20, 1997, Newcastle, Northern Ireland, CAB International, pp. 249-252.
Mitchell, A.D. and A.M. Scholz (2001): Techniques for Measuring Body Composition of Swine (Chapter 41). In: Swine Nutrition, 2nd Edition (Ed.: A.J. Lewis and L. Lee Southern), CRC Press, Boca Raton, London, New York, Washington, D.C.: 917-960.
Mitchell, A.D., K.H. Norris, H.H. Klueter, N.C. Steele, and M.B. Solomon (1986): Estimation of live body and carcass composition of pigs by near-infrared reflectance. J. Anim. Sci. 63 (Suppl. 1): 234.
Mitchell, A.D., P.C. Wang, R.W. Rosebrough, T.H. Elsasser, W.F. Schmidt (1991a): Assessment of Body Composition of Poultry by Nuclear Magnetic Resonance Imaging and Spectroscopy. Poultry Sci. 70: 2494-2500.
Mitchell, A.D., P.C. Wang, C.M. Evock (1991b): Body composition analysis of control and pGH treated pigs by NMR imaging. J. Anim. Sci. 69 (Suppl. 1): 308.
Mitchell, A.D., P.C. Wang, T.H. Elsasser, W.F. Schmitt (1991c): Application of NMR Spectroscopy and Imaging for body composition analysis as
307

Literaturverzeichnis
related to sequential measurement of energy deposition. Proc. 12th Symposium Kartause Ittingen, Switzerland, 1-7 September, 222-225.
Mitchell, A.D., P.C. Wang, T.H. Elsasser (1991d): Determination of Fat and Water Content in vitro and in vivo by Proton Nuclear Magnetic Resonance. J. Sci. Food Agric. 56: 265-276.
Mitchell, A.D., P.C. Wang, H.F. Song, W.F. Schmidt (1993): Body Composition Analysis of the Pig by Magnetic Resonance Imaging. In: Human Body Composition, Eds.: K.J. Ellis, J.D. Eastman, Plenum Press, New York, 105-108.
Mitchell, A., K. Mitchell, J. Conway and A. Scholz (1995): Measurement of compositional changes of growing pigs by dual energy x-ray absorptiometry. International Symposium Swine in Biomedical Research, University of Maryland at College Park, MD, USA, October 22-25, p. 75.
Mitchell, A.D., J.M. Conway, and W.J.E. Potts (1996a): Body composition analysis of pigs by dual-energy x-ray absorptiometry. J. Anim. Sci. 74: 2663-2671.
Mitchell, A.D., J.M. Conway, A.M. Scholz (1996b): Incremental Changes in Total and Regional Body Composition of Growing Pigs Measured by Dual-energy X-ray Absorptiometry. Growth, Development & Aging 60: 113-123.
Mitchell, A.D., R.W. Rosebrough, and J.M. Conway (1997a): Body composition analysis of chickens by dual-energy x-ray absorptiometry. Poult. Sci. 76: 1746-1752.
Mitchell, A.D., M.B. Solomon, and T.S. Rumsey (1997b): Compositional analysis of beef 9,10,11-rib sections by dual-energy x-ray absorptiometry. Meat Sci. 47: 115-124.
Mitchell, A.D., A.M. Scholz and J.M. Conway. (1998a): Body composition analysis of pigs from 5 to 97 kg by dual energy x-ray absorptiometry. Appl. Radiat. Isot. 49: 521-523.
Mitchell, A.D., P.C. Wang, H.F. Song, J.M. Conway and A.M. Scholz (1998b): Application of magnetic resonance imaging and dual-energy x-ray absorptiometry for the measurement of body composition of pigs. Proc. VIII World Conference on Animal Production, Seoul, Korea, June 22-26.
Mitchell, A.D., A.M. Scholz, V.G. Pursel, C.M. Evock-Clover (1998c): Composition Analysis of Pork Carcasses by Dual-Energy X-Ray Absorptiometry. J. Anim. Sci. 76: 2104-2114.
Mitchell, A.D., V.G. Pursel, G. Bee (1998d): Evaluation of body composition of transgenic pigs using dual-energy x-ray absorptiometry (DXA). J. Anim. Sci. 76 (Suppl. 1): 115.
Mitchell, A.D., A.M. Scholz, J.M. Conway (1998e): Body Composition Analysis of Small Pigs by Dual-Energy X-Ray Absorptiometry. J. Anim. Sci. 76: 2392-2398.
Mitchell, A.D., A.M. Scholz, and Harry J. Mersmann (2001a): Growth and Body Composition (Chapter 6). In: Biology of the Domestic Pig, 2nd edition (Eds.: W. Pond and H. Mersmann) Cornell University Press, Ithaca, NY, USA, pg. 225-308.
308

Literaturverzeichnis
Mitchell, A.D, A.M. Scholz, P.C. Wang and H. Song (2001b): Body composition analysis of the pig by magnetic resonance imaging. J. Anim. Sci. 79: 1800-1813.
Mitra, S., J.E. Wolff, R. Garrett, C.W. Peters (1995a): Whole body measurement of C, N and O using 14 MeV neutrons and the associated particle time-of-flight technique. Asia Pacific J. Clin. Nutr. 4: 187-189.
Mitra, S., J.E. Wolff, R. Garrett, C.W. Peters (1995b): Application of the associated particle technique for the whole-body measurement of protein, fat and water by 14 MeV neutron activation analysis--a feasibility study. Phys. Med. Biol. 40: 1045-1055.
Mitra, S., J.E. Wolff, R. Garrett (1998): Calibration of a prototype in vivo total body composition analyser using 14 MeV neutron activation and the associated particle technique. Appl. Radiat. Isot. 49(5-6): 537-539.
Mitsiopoulos, N., R.N. Baumgartner, S.B. Heymsfield, W. Lyons, D. Gallagher, R. Ross (1998): Cadaver validation of skeletal muscle measurement by magnetic resonance imaging and computerized tomography. J. Appl. Physiol. 85: 115-122.
Moeller, S.J. and L.L. Christian (1998): Evaluation of the Accuracy of Real-Time Ultrasonic Measurments of Backfat and Loin Muscle Area in Swine Using Multiple Statistical Analysis Procedures. J. Anim. Sci. 76: 2503-2514.
Moeller, S.J., L.L. Christian, and R.N. Goodwin (1998): Development of Adjustment Factors for Backfat and Loin Muscle Area from Serial Real-Time Ultrasonic Measurements on Purebred Lines of Swine. J. Anim. Sci. 76: 2008–2016.
Moesgaard, B, I.E. Larsen, B. Quistorff, I. Therkelsen, V. GrØsfield Christeensen, and P.F. Jørgensen (1993): Effect of Dietary Magnesium on Post Mortem Phosphocreatine Utilization in Skeletal Muscle of Swine: A Non-Invasive Study Using 31P-NMR Spectroscopy. Acta Vet. Scand. 34: 397-404.
Moesgaard, B., B. Quistorff, V. Grøsfield Christeensen, T. Jørgensen and P.F. Jørgensen (1995): Differences of Post-Mortem ATP Turnover in Skeletal Muscle of Normal and Heterozygote Malignant-Hyperthermia Pigs: Comparison of 31P-NMR and Analytical Biochemical Measurements. Meat Science 39: 43-57.
Molenat, M. (1987): Chinesische Schweine für die europäische Zucht? Teil 1: Die chinesischen Schweinerassen, die in Frankreich wirtschaftlich genutzt werden. Der Tierzüchter 39: 331-334.
Molenat, M. und J.C. Caritez (1987): Chinesische Schweine für die europäische Zucht? Teil 2: Forschungsansätze. Der Tierzüchter 39: 389-391.
Moller, D.E. (2000): Potential Role of TNF-α in the Pathogenesis of Insulin Resistance and Type 2 Diabetes. Trends in Endocrinology and Metabolism 11: 212-217.
309

Literaturverzeichnis
Montagnani, M., M. Montomoli, M. Mulinari, G. Guzzo, N. Scopetani, C. Gennari (1998): Relevance of hydration state of the fat free mass in estimating fat mass by body impedance analysis. Appl. Radiat. Isot. 49: 499-500.
Montain, S.J., S.A. Smith, R.P. Mattot, G.P. Zientara, F.A. Jolesz, and M.N. Sawka (1998): Hypohydration effects on skeletal muscle performance and metabolism: a 31P-MRS study. J. Appl. Physiol. 84: 1889-1894.
Montaldo, H.H. and C.A. Meza-Herrera (1998): Use of moleculars markers and major genes in the genetic improvement of livestock. EJB - Electronic Journal of Biotechnology 1: 7 pages (http://www.ejb.org).
Moon, R.B. and J.H. Richards (1973): Determination of intracellular pH by 31P magnetic resonance. J. Biol. Chem. 248: 7276-7278.
Morikawa, S., T. Inubushi, K. Kito, C. Kido (1993): pH Mapping in Living Tissues: An Application of in Vivo 31P NMR Chemical Shift Imaging. Magnetic Resonance in Medicine 29: 249-251.
Moser, G., E. Mueller, P. Beekmann, G. Yue, H. Geldermann (1998): Mapping of QTL‘s in F2 Generations of Wild Boar, Pietrain and Meishan pigs. Proceeding of the 6th World Congress on Genetics Applied to Livestock Production, Armidale, Australia, January 11-16, Volume 26: 478-481.
Moughan, P.J., M.W.A. Verstegen and M.I. Visser-Reyneveld (Eds.) (1995): Modelling growth in the pig. EAAP Publication No. 78, Wageningen Pers, Wageningen, NL, pp. 238.
Moulton, C.R. (1929): Meat through the microscope; applications of chemistry and the biological sciences to some problems of the meat packing industry. The University of Chicago press, Chicago, Ill., USA. http://cdl.library.cornell.edu/cgi-bin/chla/chla-cgi?notisid=AFP9939
Mullis, K.B. and F. Faloona (1987): Specific Synthesis of DNA in vitro via a Polymerase-Catalyzed Chain Reaction. Methods Enzymol. 155: 335-350.
Müller, E. (1994): Untersuchungen über die Beziehungen von Fleischanteil, Fleischbeschaffenheit und Streßresistenz bei verschiedenen Schweineherkünften 2. Mitteilung: Vergleich von Enzymaktivitäten und Metabolitkonzentrationen des Glykogenabbaus im M. longissimus dorsi. Züchtungskunde 66: 349-358.
Müller, E., Ch. Mailänder, E. Rogdakis und D. Fewson (1996): Selektionsexperiment beim Schwein zur Verbesserung der Schlachtkörper-zusammensetzung durch Zuchtwahl nach biochemischen Parametern oder Ultraschallmaßen. 2. Mitteilung: Vergleich der Zuchtlinien in der letzten Generation. Züchtungskunde 68: 32-47.
Müller, E.E., V. Locatelli, and Daniela Cocchi (1999): Neuroendocrine Control of Growth Hormone Secretion. Physiological Reviews 79: 511-607.
310

Literaturverzeichnis
Müller, M.J., O. Selberg, W. Burchert (1997): Use of positron emission tomography (PET) in the assessment of skeletal muscle glucose metabolism. Z. Ernährungswiss. 36: 359-363.
Müller, S., E. Gernand, M. Mußlick (1996): Untersuchungen zu Möglichkeiten der Einbeziehung des intramuskulären Fettgehalts (IMF) bei der Qualitätsbeurteilung von Schweinefleisch. Abschlußbericht der Thüringer Landesanstalt für Landwirtschaft, Themenblatt-Nr.: 22.02.253/1996, 101 Seiten.
Müller, U. (1994): Ergebnisse von Ultraschallmessungen am lebenden Tier und deren Beziehungen zu Merkmalen der Schlachtleistung bei Voll- und Teilzerlegung. Landbauforschung Völkenrode. Sonderheft 145: 99-105.
Müller-Gärtner, H.-W. (1998): Imaging techniques in the analysis of brain function and behaviour. TIBTECH 16: 122-130.
Naggert, J., T. Harris and M. North (1997): The genetics of obesity. Current Opinion in Genetics & Development 7: 398-404.
Nagle, D.L, S.H. McGrail, J. Vitale, E.A. Woolf, B.J. Dussault Jr., L. Dirocco, L. Holmgren, J. Montagno, P. Bork, D. Huszar, V. Fairchild-Huntress, P. Ge, J. Keilty, C. Ebeling, L. Baldini, J. Gilchrist, P. Burn, G.A. Carlson, and K.J. Moore (1999): The mahogany protein is a receptor involved in suppression of obesity. Nature 398: 148-152.
Natto, S.A., D.G. Lewis, and S.J.S. Ryde (1998): Benchmarking the MCNP Code for Monte Carlo Modelling of an In Vivo Neutron Actiavtion Analysis System. Appl. Radiat. Isot. 49: 545-547.
Nelson, Th.E. and Marina Lin (1995): Abnormal function of porcine malignant hyperthermia calcium release channel in the absence and presence of halothane. Cell Physiol. Biochem. 5: 10-22.
Nelson, Th.E., Marina Lin, and P. Volpe (1991): Evidence for intraluminal Ca++ regulatory site defect in sarcoplasmic reticulum from malignant hyperthermia pig muscle. The Journal of Pharmacology and Experimental Therapeutics 256: 645-649.
Neuenschwander, S., G. Rettenberger, E. Meijerink, H. Jörg and G. Stranzinger (1996): Partial characterization of porcine obesity gene (OBS) and its localization to chromosome 18 by somatic cell hybrids. Animal Genetics 27: 275-278.
Newbold, R.P. and R.K. Tume (1981): Comparison of the effects of added orthophosphate on calcium uptake and release by bovine and rabbit muscle sarcoplasmic reticulum. J. Food Sci. 46: 1327-1332.
Newton, C.R. und A. Graham (1994): PCR. 2. Aufl., Spektrum Akademischer Verlag, Heidelberg, Berlin, Oxford.
Nitter, G. und R. Kolb (1994): Abschätzung des Fleischanteils im Bauch mit Hilfe von digitalisierten Ultraschallbildern an lebenden Schweinen. Landbauforschung Völkenrode, Sonderheft 145: 106-109.
311

Literaturverzeichnis
Nold, R.A., J.R. Romans, W.J. Costello, G.W. Libal (1999): Characterization of muscles from boars, barrows, and gilts slaughtered at 100 or 110 kilograms: differences in fat, moisture, color, water-holding capacity, and collagen. J. Anim. Sci. 77: 1746-1754.
Norman, R.A., C. Bogardus, E. Ravussin (1995): Linkage between obesity and a marker near the tumor necrosis factor-alpha locus in Pima Indians. J. Clin. Invest. 96: 158-162.
North, M.A. (1999): Advances in the molecular genetics of obesity. Current Opinion in Genetics and Development 9: 283-288.
North, M.A., J.K. Naggert, Y. Yan, K. Noben-Trauth, P.M. Nishina (1997): Molecular characterization of TUB, TULP1, and TULP2, members of the novel tubby gene family and their possible relation to ocular diseases. Proc. Natl. Acad. Sci. USA 94: 3128-3133.
NRC (1988). Nutrient requirements of swine (9th Ed.). National Academy Press, Washington, D.C.
Nürnberg, K., G. Kuhn, I.Fiedler, K. Ender (1988): Wachstum, Zusammensetzung und qualitative Eigenschaften des Fettgewebes beim Schwein. In: Genetische Probleme der Tierzucht 19 (Editoren G. Herrendörfer und J. Wolf), Akademie der Landwirtschaftswissenschaften der DDR, Dummerstorf-Rostock, 97 Seiten.
Nürnberg, K., J. Wegner, K. Ender (1998): Factors influencing fat composition in muscle and adipose tissue of farm animals. Livest. Prod. Sci. 56: 145-156.
Oelberg, D.A., A.B. Evans, M.I. Hrovat, P.P Pappagianopoulus, S. Patz, D.M. Systrom (1998): Skeletal muscle chemoreflex and pHi in exercise ventilatory control. J. Appl. Physiol. 84: 676-682.
Ollivier, L.; Sellier, P.; Monin, P. (1975): Determinisme genetique du syndrome d'hyperthermie maligne chez le porc de Pietrain. Ann. Genet. Sel. Anim. 7: 159-166.
Ollmann, M.M., M. Lynn Lamoreux, B.D. Wilson, and G.S. Barsh (1998): Interaction of Agouti protein with the melanocortin 1 receptor in vitro and in vivo. Genes and Development 12: 316-330.
Olson, Ann Louise and J.E. Pessin (1996): Structure, function, and regulation of the mammalian facilitative glucose transporter gene family. Annu. Rev. Nutr. 16: 235-256.
Ong, H.T., J. Tieman, M. Albert, F. Jolesz, T. Sandor (1997): Semi-automated extraction of brain contours from MRI. Neuroradiology 39: 797-803.
Orcutt, M.W., J.C. Forrest, M.D. Judge, A.P. Schinckel, C.H. Kuei (1990): Practical means for estimating pork carcass composition. J. Anim. Sci. 68: 3987-3997.
Orlova, E.V., I.I. Serysheva, M. van Heel, S.L. Hamilton, W Chiu (1996): Two structural configurations of the skeletal muscle calcium release channel. Nat. Struct. Biol. 3: 547-552.
312

Literaturverzeichnis
Oster, A., D. Fewson, P. Komender, W. Branscheid, und E. Sack (1987): Schätzung des Muskelgewebeanteiles beim Schwein aufgrund der Forchheimer Teilstückzerlegung sowie üblicher Schlachtkörpermaße. Züchtungskunde 59: 281-295.
Otsu, K.; Khanna V. K.; Archibald, A.L.; MacLennan D. H. (1991): Cosegregation of Porcine Malignant Hyperthermia and a Probable Causual Mutation in the Skeletal Muscle Ryanodine Receptor Gene in Backcross Families. Genomics 11: 744-750.
Otten, W., Ellen Kanitz and Margret Tuchscherer (2000): Prenatal stress in pigs: effects on growth, physiological stress reactions and immune function. Arch. Tierz. (Dummerstorf) 43 (Sonderheft): 159-164.
Park, Y.I., J.Y. Han, T.S. Park, Y.S. Shin, S.C. Lee, I.J. Lee, H.K. Lee, H.K. Kim, H.S. Oh, C.J. Son (1998): Effect of porcine stress syndrome genotype on performance traits in swine. Proc. 6th World Congress on Genetics Applied to Livestock Production, Armidale, Australia, Vol. 23: 668-671.
Pászthy, G., A. Lengyel, P. Horn (1991): Comparing breeds and fattening methods by computer tomography on live sheep. 42nd Annual Meeting of the EAAP, Berlin, Germany, 9 -12 September, 254.
Patience, J.F., and P.A. Thacker (1989): Swine Nutrition Guide. Prairie Swine Centre. 260 pp.
Patterson, D.L. (1990): Obtaining objective measurements of animal conformation by video image analysis. Proc. 4th World Congress on Genetics applied to Livestock Production. Edinburgh, 23-27 July, Volume XV: 295-298.
Paul, Sabine (1998): Molekulargenetische Untersuchungen in der alpha 1-Untereinheit des Dihydropyridinrezeptors bei der Malignen Hyperthermie des Menschen. Dissertation, Ludwig-Maximilians-Universität München, München.
Paulke, Th. und A. Scholz (1999): Die Charakteristik von Merkmalen der Futteraufnahme unter ad libitum Bedingungen bei verschiedenen Schweinerassen und Rassenkombinationen. Züchtungskunde 71: 229-243.
Peschke, W, W. Wittmann, und E. Littmann (1997): Auswirkungen unterschiedlicher Meßskalen des Ultraschallgerätes piglog 105 auf die Beziehungen der Meßergebnisse zu Schlachtkörpermaßen. Gruber Info Ausgabe 1/1997.
Peppler, W.W. and R.B. Mazess (1981): Total body bone mineral and lean body mass by dual-photon absorptiometry. I. Theory and measurement procedure. Calcif. Tissue Int. 33: 353-359.
Pérez-Enciso, M., A. Clop, J.L. Noguera, C. Óvilo, A. Coll, J.M. Folch, D. Babot, J. Estany, M.A. Olivers, I. Díaz, and A. Sánchez (2000): A QTL on pig chromosome 4 affects fatty acid metabolism: Evidence from an Iberian by Landrace intercross. J. Anim. Sci. 78: 2525-2531.
313

Literaturverzeichnis
Pfeiffer, H. (1984): Schweinezucht. 3. überarbeitete Auflage, VEB Deutscher Landwirtschaftsverlag, Berlin.
Pfeiffer, H., G. von Lengerken, G. Gebhardt (1984): Wachstum und Schlachtkörperqualität bei landwirtschaftlichen Nutztieren - Schweine. VEB Deutscher Landwirtschaftsverlag Berlin.
Pfeiffer, H. und Chr. Schröder (1994): Die Ermittlung des Muskel-Speck-Verhältnisses mit Hilfe der Ultraschall-Meßtechnik zur Erfassung des Fleischanteils am lebenden Schwein.Landbauforschung Völkenrode, Sonderheft 145: 90-98.
Picaud, J.-Ch., J. Rigo, K. Nyamugabo, J. Milet, J. Senterre (1996): Evaluation of dual-energy X-ray absorptiometry for body-composition assessment and term human neonates. Am. J. Clin. Nutr. 63: 157-163.
Pierson, R.N., J. Wang, and J.C. Thornton (1997): Measurement of Body Composition: Applications in Hormone Research. Horm. Res. 48 (suppl. 1):56-62.
Pierson, R.N., J. Wang, and J.C. Thornton (2000): Body Composition Comes of Age: A Modest Proposal for the Next Generation. In: In Vivo Body Composition Studies, Editors: S. Yasumuru, J. Wang, and R.N. Pierson Jr., Ann. N. Y. Acad. Sci. 904: 1-11.
Pietrobelli, A., C. Formica, Z. Wang, and St. B. Heymsfield (1996): Dual-energy X-ray absorptiometry body composition model: review of physical concepts. Am. J. Physiol. 271: E941-E951.
Pietrobelli, A., D. Gallagher, R. Baumgartner, R. Ross, and St.B. Heymsfield (1998a): Lean R Value for DXA Two-component Soft-tissue Model: Influence of Age and Tissue or Organ Type. Appl. Radiat. Isot. 49: 743-744.
Pietrobelli, A., Z. Wang, C. Formica, and St.B. Heymsfield (1998b): Dual-energy X-ray absorptiometry: fat estimation errors due to variation in soft tissue hydration. Am. J. Phys. 274: E808-E816.
Pietsch, H. (1998): Kernspintomographische Untersuchungen der Bauchhöhle der Katze. Diss., Freie Univ. Berlin, Journal-Nr. 2217.
Pintauro, St.J., T.R. Nagy, Ch.M. Duthie, M.I. Goran (1996): Cross-calibration of fat and lean measurements by dual-energy X-ray absorptiometry to pig carcass analysis in the pediatric body weight range. Am. J. Clin. Nutr. 63: 293-298.
Plourde, G. (1997): The Role of Radiologic Methods in Assessing Body Composition and Related Metabolic Parameters. Nutrition Review 55: 289-296.
Polito, A., J. Parizkova, E. Toti, and A. Ferro-Luzzo (1994): The measurement of body composition by infrared interactance: an evaluation. Nutrition research (N.Y.) 14: 1165-1177.
Pomp, D. (1999): Animal models of obesity. Molecular Medicine Today 5: 459-460.
314

Literaturverzeichnis
Porsch-Özçürümez, M.K. (1997): Vergleichende Untersuchung der Lipidstoffwechselparameter zwischen 35-64 jährigen deutschen und in Deutschland lebenden türkischen Teilnehmern einer Gesundheitsvorsorgeuntersuchung (Check-up 35). Dissertation, Justus-Liebig-Universität Gießen.
Preston T, P.J. Reeds, B.W. East, and P.H. Holmes (1985): A comparison of body protein determination in rats by in vivo neutron activation and carcass analysis. Clin. Sci. (Colch) 68: 349-355.
Price, Th.B. and J.C. Gore (1998): Effect of muscle glycogen content on exercise-induced changes in muscle T2 times. J. Appl. Physiol. 84: 1178-1184.
Price, Th.B., G. Perseghin, A. Duleba, W. Chen, J. Chase, D.L. Rothman, R.G. Shulman, and G.I. Shulman (1996): NMR studies of muscle glycogen synthesis in insulin-resistant offspring of parents with non-insulin-dependent diabetes mellitus immediately after glycogen-depleting exercise. Proc. Natl. Acad. Sci. (USA) 93: 5329-5334.
Prior, B.M., K.J.Cureton, Ch.M. Modlesky, E.M. Evans, M.A. Sloniger, M. Saunders, R.D. Lewis (1997): In vivo validation of whole body composition estimates from dual-energy X-ray absorptiometry. J. Appl. Physiol. 83: 623-630.
Prusa, K.J., Jane A. Love, and L.L. Christian (1989): Fat Content and Sensory Analysis of Selected Pork Muscles Taken from Carcasses with Various Backfat Levels. Journal of Food Quality 12: 135-143.
Puff, H. (1975): Genetische Parameter wichtiger Mast- und Schlachtleistungskriterien bei der Zuchtpopulation der Deutschen Landrasse in Niederbayern. Bayer. Landw. Jahrbuch 52/8: 956-975.
Purcell, E.M., H.C. Torrey, R.V. Pound (1946): Resonance absorption by nuclear magnetic moments in a solid. Phys. Rev. 69: 37-38.
Pursel, V.G., C.A. Pinkert, K.F. Miller, D.J. Bolt, R.G. Campell, R.D. Palmiter, R.L. Brinster and R.E. Hammer (1989): Genetic engineering of livestock. Science 244: 1281-1288.
Pursel, V.G., R.E. Hammer, D.J. Bolt, R.D. Palmiter and R.L. Brinster (1990): Integration, expression and germ-line transmission of growth-related genes in pigs. J. Reprod. Fert. Suppl. 41: 77-87.
Quiniou, N. and J. Noblet (1995): Prediction of Tissular Body Composition from Protein and Lipid Deposition in Growing Pigs. J. Anim. Sci. 73: 1567-1575.
Rathfelder, A. (1991): Beziehungen zwischen Muskelmorphologie, Fleischanteil, Fleischbeschaffenheit, und Stressresistenz bei verschiedenen Schweineherkünften. Dissertation, Hohenheim, 128 S.
Radda, G.K. (1992): Control, bioenergetics, and adaptation in health and disease: noninvasive biochemistry from nuclear magnetic resonance. FASEB J. 6: 3032-3038.
315

Literaturverzeichnis
Radda, G.K., R.D. Oberhaensli, and Doris J. Taylor (1987): The Biochemsitry of Human Diseases as Studied by 31P NMR in Man and Animal Models. Ann. N. Y. Acad. Sci. 508: 300-308.
Raden, J. (1917): On the determination of functions from their integrals along certain mainfolds. Math. Phys. Klasse. 69: 262-277.
Radeff, T. (1930): Über den Calcium-, Phosphor- und Stickstoffansatz junger Tiere in der Säugezeit (nach Versuchen an Hund, Kaninchen, Schwein und Ziege) Arch. Tierernähr. Tierz. 3: 659-662.
Ramsay, T.G., X.Yan, and C. Morrison (1998): The Obesity Gene in Swine: Sequence and Expression of Porcine Leptin. J. Anim. Sci. 76: 484-490.
Rathfelder, A. (1991): Beziehungen zwischen Muskelmorphologie, Fleischanteil, Fleischbeschaffenheit und Sressresistenz bei verschiedenen Schweineherkünften. Diss., Univ. Hohenheim, 128 S.
Rehfeldt, Charlotte, N.C. Stickland, Ilse Fiedler and J. Wegner (1999): Environmental and Genetic Factors as Sources of Variation in Skeletal Muscle Fibre Number. Basic Appl. Myol. 9: 237-255.
Reichart, Ursula, R. Kappler, H. Scherthan, E. Wolf, M. Müller, G. Brem, and B. Aigner (2000a): Partial Leptin Receptor Gene Deletion in Transgenic Mice Prevents Expression of the Membrane-Bound Isoforms for Ob-Rc. Biochemical and Biophysical Research Communications 269: 496-501.
Reichart, Ursula, Ingrid Renner-Müller, A. Höflich, O. J. Müller, W.M. Franz, E. Wolf, M. Müller, G. Brem, and B. Aigner (2000b): Contrasting Obesity Phenotypes Uncovered by Partial Leptin Receptor Gene Deletion in Transgenic Mice. Biochemical and Biophysical Research Communications 269: 502-507.
Reiner, G., G. Moser, H. Geldermann, V. Dzapo (1999): Associations between c-myc protooncogene and carcass quality traits in the pig – evidence for epistasis with the ryr1-gene. J. Anim. Breed. Genet. 116: 253-261.
Reiser, Gabriele (1984): Erprobung verschiedener Methoden zur Schätzung von Varianzkomponenten aus Daten der Stationsprüfung beim Schwein. Diss., Univ. Hohenheim.
Reisner, K. (1996): Geschichte der bildgebenden Verfahren im Kopf-Hals-Bereich. Der Radiologe 36: 175-180.
Rempel, W.E., M.Y. Lu, S.E. Kandelgy, C.F.H. Kennedy, L.R. Irvin, J.R. Mickelson, and C.F. Loius (1993): Relative Accuracy of the Halothane Challenge test and a Molecular Genetic Test in Detecting the Gene for Porcine Stress Syndrome. J. Anim. Sci. 71: 1395-1399.
Rempel, W.E., M.Y. Lu, J.R. Mickelson, and C.F. Loius (1995): The effect of skeletal muscle ryanodine receptor genotype on pig performance and carcass quality traits. Animal Science 60: 249-257.
316

Literaturverzeichnis
Rico, H., M. Revilla, L.F. Villa, D. Ruiz-Contreras, E.R. Hernandez, and M. A. de Buergo (1994): The Four-Compartment Models in Body Composition: Data From a Study With Dual-Energy X-Ray Absorptiometry and Near-Infrared Interactance on 815 Normal Subjects. Metabolism 43: 417-422.
Rieu, M. and J.P. van Ferneij (2001): Situation and perspectives of swine production in the European Union. Second International Virtual Conference on Pork Quality November, 05 to December, 06. http://www.conferencia.uncnet.br/pork/seg/pal/anais01p2_rieu_en.pdf
Rigo J., K. Nyamugabo, J.C. Picaud, P. Gerard, C. Pieltain, and M. De Curtis (1998): Reference values of body composition obtained by dual energy X-ray absorptiometry in preterm and term neonates. J. Pediatr. Gastroenterol. Nutr. 27: 184-190.
Rikimaru, T., T. Oozeki, M. Ichikawa, H. Ebisawa, and Y. Fujita (1989): Comparisons of urinary creatinine, skeletal muscle mass, and indices of muscle protein catabolism in rats fed ad libitum, with restricted food intake, and deprived of food. J. Nutr. Sci. Vitaminol. (Tokyo) 35: 199-209.
Ríos, E. and M.D. Stern (1997): Calcium in Close Quarters: Microdomain Feedback in Excitation-Contraction Coupling and Other Cell Biological Phenomena. Annu. Rev. Biophys. Biomol. Struct. 26: 47-82.
Rittler, A. (1969): Echolotmessungen als Hilfsmittel für die Zuchtwertschätzung beim Schwein. 3. Mitt.: Abschätzung des Fleisch- und Fettanteils aufgrund von Echolotmessungen am lebenden Schwein. Z. Tierzüchtg. Züchtgsbiol. 85: 78-89.
Rittler, A., P. Schoen, E. Schelper, D. Fewson (1964): Zur Frage der Genauigkeit von Echolot-Messungen am lebenden Schwein in verschiedenen Gewichtsabschnitten. Züchtungskunde 36: 159-168
Roberts, J.T., T. Burt, L. Gouylai, B. Chance, F. SrÈter, and J. Ryan (1983): Immediate uncoupling of high energy oxidative phosphorylation in muscle of malignant hyperthermic swine determined non-invasively by whole body 31P nuclear magnetic resonance. Anesthesiology 59: A230.
Roberts, S.A., A.L. Schaefer, A.C. Murray, L. Thibault (1998): Fall and winter hormone concentrations related to stress in pigs identified as normal and carrier for stress susceptibility. Chronobiol. Int. 15: 275-281.
Robinson, Susan W., Daniela M. Dinulescu, and R.D. Cone (2000): Genetic Models of Obesity and Energy Balance in the Mouse. Annu. Rev. Genet. 34: 687-745.
Roden, M and G.I. Shulman (1999): Applications of NMR Spectroscopy to study muscle glycogen metabolism in man. Annu. Rev. Med. 50: 277–290.
Roemmich, J.N., P.A. Clark, A. Weltman, and A.D. Rogol (1997): Alterations in growth and body composition during puberty. I. Comparing multicompartment body composition models. J. Appl. Physiol. 83: 927-935.
Rogdakis, E., E. Müller, Ch. Mailänder, und D. Fewson (1996): Selektionsexperiment beim Schwein zur Verbesserung der Schlachtkörper-zusammensetzung durch Zuchtwahl nach biochemischen Parametern oder
317

Literaturverzeichnis
Ultraschallmaßen. 1. Mitteilung: Versuchsanlage, direkte und indirekte Selektionseffekte. Züchtungskunde 68: 20-31.
Rogol, A.D. (1997): Body composition analysis during growth in children and adolescents. Current Opinion in Endocrinology and Diabetes 4: 77-79.
Rohrer, G.A. (2000): Identification of quantitative trait loci affecting birth characters and accumulation of backfat and weight in a Meishan-White Composite resource population. J. Anim. Sci. 78: 2547-2553.
Rohrer, G.A., and J.W. Keele (1998): Identification of Quantitative Trait Loci Affecting Carcass Composition in Swine: I. Fat Deposition Traits. J. Anim. Sci. 76: 2247-2254.
Rooyackers, O.E. and K.S. Nair (1997): Hormonal Regulation of Human Muscle Protein Metabolism. Annu. Rev. Nutr. 17: 457-485.
Romvári, R., M. Perényi, P. Horn (1994): In vivo measurement of total body fat content of broiler chickens by X-ray computerised tomography. Znan. Prak. Poljopr. Tehnol. 24: 215-220.
Romvári, R., G. Milisits, Zs. Szendrõ, P. Sørensen (1996): Non invasive method to study the body composition of rabbits by X-ray computerised tomography. World Rabbit Science 4: 219-224.
Romvári, R., Zs. Szendrõ, J.F. Jensen, P. Sørensen, G. Milisits, P. Bogner, P. Horn, J. Csapó (1998): Noninvasive measurement of body composition of two rabbit populations between 6 and 16 weeks of age by computer tomography. J. Anim. Breed. Genet. 115: 383-395.
Rose, B.S., W.P. Flatt, R.J. Martin, R.D. Lewis (1998): Whole body composition of rats determined by dual energy x-ray absorptiometry is correlated with chemical analysis. J. Nutr. 128: 246-250.
Ross, R., B. Goodpaster, D. Kelley, F. Boada (2000): Magnetic resonance imaging in human body composition research. From quantitative to qualitative tissue measurement. Ann. N. Y. Acad. Sci. 904: 12-17.
Roussel, R., P.G: Carlier, J.-J. Robert, G. Velho, and G. Bloch (1998): 13C/31P NMR studies of glucose transport in human skeletal muscle. Proc. Natl. Acad. Sci. (USA) 95: 1313-1318.
Rothfuss, Ursula (1981): Lipogene und lipolytische Parameter im Fettgewebe von Large White- und Pietrain-Schweinen in Abhängigkeit vom Mastgewicht. Dissertation, Universität Hohenheim.
Rothschild, M. (2001): Dissecting the genetic control of carcass merit and meat quality in the pig. J. Anim. Sci. 79 (Suppl. 1): 99.
Roubenoff, R., and J.J. Kehayias (1991): The meaning and measurement of lean body mass. Nutrition Reviews 49: 163-175.
Roubenoff, R., J.J Kehayias, B. Dawson-Hughes, and St.B. Heymsfield (1993): Use of dual-energy x-ray absorptiometry in body-composition studies: not yet a “gold standard”. Am. J. Clin. Nutr. 58: 589-591.
318

Literaturverzeichnis
Rozeboom D.W., J.E. Pettigrew, R.L. Moser, S.G. Cornelius, S.M. el Kandelgy (1994): In vivo estimation of body composition of mature gilts using live weight, backfat thickness, and deuterium oxide. J. Anim. Sci. 72: 355-366.
Rozeboom, K.J., M.G. Thomas, L. Hillman, F. Chanetsa, R.J. Lipsey, D.H. Keisler (1998): Use of dual-energy x-ray absorptiometry in estimating the carcass composition of wether lambs. J. Anim. Sci. 76 (Suppl. 1): 149.
Ruusunen, M. and E. Poulanne (1997): Comparison of histochemical properties of different pig breeds. Meat Sci. 45: 119-125.
Ruten, W., E. Tholen, M. Rodehutscord, und W. Trappmann (1999): Beurteilung des Proteinaufwandes als mögliches Selektionskriterium in der Schweinezucht. Züchtungskunde 41: 277-292.
Ryde, S.J.S., J.L. Birks, W.D. Morgan, C.J. Evans, and J. Dutton (1993): A five-compartment model of body composition of healthy subjects assessed using in vivo neutron activation analysis. European Journal of Clinical Nutrition. 47:863-874.
Sajonski, H. und A. Smollich (1981): Zelle und Gewebe. S. Hirzel Verlag, Leipzig, pp. 278.
Salomon, F.V., G. Michel, B. Salomon, F. Gruschwitz (1981): Zur Fasertypisierung an Skelettmuskeln. Mh. Vet.-med. (Jena) 36: 349-359.
Salomon, F.V., G. Michel, F. Gruschwitz (1983): Zur Entwicklung von Fasertypenkomposition und Faserdurchmesser im M. longissimus des Hausschweines (Sus scrofa domesticus). Anat. Anz. (Jena) 154: 69-79.
Salomon, F.V., D. Bittorf, B. Wulf, F. Liebert, G. Gebhardt, G. von Lengerken und H. Pfeiffer (1988): Zum Einfluß unterschiedlicher Ernährungsintensität auf die Skelettmuskulatur des Schweines. Wiss. Z. Karl-Marx-Univ. Leipzig, Math.-Naturwiss. R. 37: 44-49.
Sardinha, L.B., T.G. Lohman, P.J. Teixeira, D.P. Guedes, and S.B. Going (1998): Comparison of air displacement plethysmography with dual-energy x-ray absorptiometry and 3 field methods for estimating body composition in middle-aged men. Am. J. Clin. Nutr. 68: 786-793.
SAS (1990): SAS / STATTM Guide for Personal Computers, Version 6, Fourth Edition, by SAS Institute Inc., Cary, NC, USA.
Sasaki, S., A.C. Clutter and D. Pomp (1996): Assignment of the porcine obese (leptin) gene to Chromosome 18 by linkage analysis of a PCR-based product. Mammalian Genome 7: 471-472.
Sather, A.P., A.K.W. Tong, and D.S. Harbison (1986): A Study of Ultrasonic Probing Techniques for Swine. I: The Effect of Operator, Machine and Site. Can. J. Anim. Sci. 66: 591-598.
Sather, A.P., A.K.W. Tong, and D.S. Harbison (1987): A Study of Ultrasonic Probing Techniques for Swine. II: Prediction of Carcass Yield from the Live Pig. Can. J. Anim. Sci. 67: 381-389.
Saxton, A.M. and E.J. Eisen (1984): Genetic Analysis of Brown Adipose Tissue, Obesity and Growth in Mice. Genetics 106: 705-718.
319

Literaturverzeichnis
Schaller S., T. Flohr, P. Steffen (1996a): An Efficient Fourier Method for 3D Radon Inversion in Exact Cone-Beam CT Reconstruction. IEEE Medical Imaging Conference, Anaheim (CA), Nov. 7-9. http://www.nt.e-technik.uni-erlangen.de/~schaller/DFI_poster_html/title.html.
Schaller S., T. Flohr, P. Steffen (1996b): A new Approximate Algorithm for Image Reconstruction in Cone-Beam Spiral CT at small Cone-Angles. IEEE Medical Imaging Conference, Anaheim (CA), Nov. 7-9. http://www.nt.e-technik.uni-erlangen.de/~schaller/CFBP_poster_html/title.html.
Scheper, J und W. Scholz (1985): DLG-Schnittführung für die Zerlegung der Schlachtkörper von Rind, Kalb, Schwein und Schaf. Arbeitsunterlagen DLG, 31 Seiten.
Schild, H.H. (1990): MRI made easy. Schering AG, Berlin/ Bergkamen.
Schinckel, A.P., P.V. Preckel and M.E. Einstein (1996): Prediction of Daily Protein Accreation Rates of Pigs from Estimates of Fat-Free Lean Gain Between 20 and 120 Kilograms Live Weight. J. Anim. Sci. 74: 498-503.
Schinckel, A.P. and C.F.M. de Lange (1996): Characterization of Growth Parameters Needed as Inputs for Pig Growth Models. J. Anim. Sci. 74: 2021-2036.
Schlindwein, B. (1990): Untersuchungen über den Einfluss unterschiedlicher Selektionsmaßnahmen auf Lipolyse, Lipogenese und Fettzellparameter beim Schwein. Dissertation Hohenheim.
Schmidt, E. (1988): Analyse des Wachstums und seine Beziehungen zum Schlachtkörperwert beim Schwein. Diss. Kiel, Schriftenreihe des Institutes für Tierzucht und Tierhaltung der Christian-Albrechts-Universität zu Kiel, Heft 48: 118 S.
Schmidt, J., M. Freiin v. Schleinitz, E. Lagenau und C. Zimmermann (1931): Über die Zusammensetzung des Schweinekörpers. Archiv für Tierernährung und Tierzucht 5: 237-375.
Schmitten, F. (1993): Schweinefleischqualität in der Zuchtpraxis. Züchtungskunde 65: 455-467. Schmitten, F., A. Klingelhöller, K.H. Schepers, A. Festerling, H. Juengst (1986): Einfluss des Mastendgewichtes auf die Fleischbeschaffenheit beim Schwein. Züchtungskunde 58: 282-290.
Schmoetzer, G. (1998): Molekulare Onkologie - Grundbegriffe – Online-Textbook. http://home.t-online.de/home/schmoetzer-gruenstadt/onko_k3.htm
Schneider, M.R., H. Lahm, M. Wu, A. Hoeflich, and E. Wolf (2000): Transgenic mouse models for studying the functions of insulin-like growth factor-binding proteins. FASEB J. 14: 629-640. Schoelgens, C. (1998): NativeTM Tissue Harmonic Imaging. Der Radiologe 38: 420-423. Scholz, A. (1990): Züchterische Bewertung neuer Selektionskriterien beim Schwein: Magerfleisch-
320

Literaturverzeichnis
wachstumsrate, Magerfleischfutteraufwand und Restwachstumsrate. Diss., Humboldt-Universität zu Berlin.
Scholz, A. (1994): Quantitative Analyse von Magnet-Resonanz-Tomographie-Bildern zur Bestimmung der Körperzusammensetzung am lebenden Schwein. Landbauforschung Völkenrode; Sonderheft 145: 38-48.
Scholz, A. (1997): Videobildanalyse zur Bestimmung des Marmorierungsgrades im Kotelett mit Hilfe verschiedener SAS-Prozeduren. 1. Konferenz der SAS-Benutzer in Forschung und Entwicklung (KSFE), Humboldt-Universität zu Berlin, 20./21. Februar, 295-308. http://www.hu-berlin.de/rz/ksfe/inhalt.html
Scholz, A. (2001): Anwendungspotential der Dualenergie-Röntgenabsorptiometrie bei den Tierarten Schwein, Rind und Schildkröte. Vortragsunterlagen zum Kolloquium: “Entwicklungsstand nicht invasiver Messtechniken zur Erfassung der Körperzusammensetzung”, Institut für Tierzucht und Tierverhalten der FAL, Mariensee, 29.05.2001.
Scholz, A. und G. Triebler (1992): Zur Selektionswirkung der “biologischen” Indizes Magerfleischwachstumsrate und Magerfleischfutterverwertung bei zwei Schweinerassen. Züchtungskunde 64: 346-355.
Scholz, A. und T. Hardge (1994): Zum Einfluß von MHS-Genotyp und Genkonstruktion auf komplexe quantitative Merkmale der Mast- und Schlachtleistung sowie Fleischqualität beim Schwein. Arch. Tierzucht (Dummerstorf) 37: 245-256.
Scholz, A.M. and A.D. Mitchell (1996): Accuracy of in vivo body composition measurements comparing magnetic resonance imaging and dual energy x-ray absorptiometry. J. Animal Sci. 74 (Suppl. 1): 152
Scholz, A., U. Baulain, E. Kallweit (1993): Quantitative Analyse von Schnittbildern lebender Schweine aus der Magnet-Resonanz-Tomographie. Züchtungskunde 65: 206-215.
Scholz, A.M., A.D. Mitchell, P.C. Wang, H. Song and Z. Yan (1995a): Muscle and fat deposition of pigs with different ryanodine receptor genotypes from 10 to 90 kg body weight studied by means of Magnetic Resonance Imaging. J. Anim. Sci. 37 (Suppl. 1): 137.
Scholz, A. M., A. D. Mitchell, P. C. Wang, H. Song and Z. Yan (1995b): Muscle metabolism and body composition of pigs with different ryanodine receptor genotypes studied by means of 31P nuclear magnetic resonance spectroscopy and 1H magnetic resonance imaging. Arch. Tierzucht (Dummerstorf) 5: 539-552.
Scholz, A., Th. Paulke, H. Eger (1995c): Bestimmung des Marmorierungsgrades beim Schwein - Einsatz der computergestützten Videobildanalyse. Fleischwirtschaft 75: 320-322.
Scholz, A.M., A.D. Mitchell, H. Song and P.C. Wang (1998): In vivo glycogen and phosphate metabolism in relation to body composition in young pigs of different genotypes studied by nuclear magnetic resonance techniques. Proceedings of the 6th World Congress on Genetics Applied to Livestock Production. January 11 - 16, Armidale, NSW, Australia; V. 25: 125-128.
321

Literaturverzeichnis
Scholz, A., S. Nüske, P. Soffner und M. Förster (2001): Dualenergie-Röntgenabsorptiometrie-Untersuchungen zur Körperzusammensetzung und Knochenmineralisierung bei neugeborenen Kälbern unterschiedlicher genetischer Herkunft. Kurzfassungen Vortragstagung der DGfZ und GFT am 12./13.09.2001 in Weihenstephan, A22.
Schoppmeyer, A., I. Fiedler, M. Hartung, G. Dietl, K. Ender (2001): Untersuchungen zur Mikrostruktur des M. longissimus bei fünf Schweinerassen unter Berücksichtigung histopathologischer Fasern. Kurzfassungen Vortragstagung der DGfZ und GFT am 12./13.09.2001 in Weihenstephan, D24.
Schöberlein, L. (1976): Die Schußbiopsie – eine neue Methode zur Entnahme von Muskelproben. Mh. Vet.-Med., Jena 31: 457-460.
Schönmuth, G. (1987): Leistungszucht und Tiergesundheit. Arch. Tierzucht (Dummerstorf) 30: 7-14.
Schönmuth, G. (1988): Kombination von Fleischansatz und Belastbarkeit. In: Biologische Grundlagen der Fleischerzeugung und –bewertung, Tagungsbericht der Akademie der Landwirtschaftswissenschaften der DDR, Berlin, 268: 17-26.
Schubert, H. (1998): Radiologische Diagnoseverfahren. http://www.klinikum.rwth-aachen.de/cbt/radiologie/skript/index.html
Schulte Spechtel, M.-T., H.-H. Wesemeier, U. Baulain, M. Henning, F. Ellendorf (1997): A Magnetic Resonance Tomographic Atlas of the Turkey (Meleagris gallopavo). Landbauforschung Völkenrode, Sonderheft 179: 79 S.
Schulze, V., R. Röhe, H. Looft und E. Kalm (1998): Möglichkeiten der züchterischen Verbesserung des Exterieurs beim Schwein unter besonderer Berücksichtigung des Fundaments. Züchtungskunde 70: 43-60.
Schwaiger, M. (1997): Radioaktivität - medizinische Anwendungen. In: Forschung mit Neutronen - Nutzen für die Medizin in Diagnose und Therapie. Dokumentation der Technischen Universität München zur Neuen Forschungs-Neutronenquelle Garching - FRM-II. http://www.frm2.tu-muenchen.de/d/medizin/medizin.htm
Schwerin, M. (1994): Molekulare Genomanalyse in der Tierzucht. Arch. Tierzucht (Dummerstorf) 37 (Sonderheft): 107-112.
Schwörer, D., A. Rebsamen (1990a): Zucht auf genussreiches Schweinefleisch. Der Kleinviehzüchter 38: 1469-1478.
Schwörer, D., A. Rebsamen (1990b): Zucht auf gute Fleischbeschaffenheit durch Berücksichtigung des Gehaltes an intramuskulärem Fett. Schweinezucht und Schweinemast 38: 173 - 176.
Schwörer, D., P. Morel, A. Rebsamen (1987): Selektion auf intramuskuläres Fett beim Schwein. Tierzüchter 39: 392 - 394.
322

Literaturverzeichnis
Schwörer, D., P. Morel, A. Rebsamen (1990): Bedeutung der chemischen Zusammensetzung der Fettgewebe für die Schweinezucht. Der Kleinviehzüchter 38: 297 - 309.
See, M.T. (1994): Heterogeneity of variance amongst swine herds for backfat. Proc. 5th World Congress on Genetics Applied to Livestoc Production. Vol. 17: 409-412.
See, M.T. and C.M. Stanislaw (1994): Estimating carcass lean in the live animal. In: Animal Science Facts, ANS 94-803S. http://mark.asci.ncsu.edu/breeding/803s.htm oder http://mark.asci.ncsu.edu/Publications/factsheets/803s.htm
Sehested, E. (1983): Computerized Tomography of Sheep. CEC Meeting: In Vivo Measurement of Body Composition in Meat Animals, Bristol, UK, November 30 - December 1.
Sehested, E. (1986): In-Vivo prediction of lamb carcass composition by computerized tomography. PhD Thesis, Ås-NLH, Norway, pp 81.
Sehested, E. and O. Vangen (1988): Computer Tomography, A Non-Destructive Method of Carcass Evaluation. VI World Conference on Animal Production, Helsinki, Finland, 27th June -1st July, 5 pp.
Seidemann, S.C., J.D. Crouse and H.J. Mersmann (1989): Carcass, muscle and meat characteristics of lean and obese pigs. J. Anim. Sci. 67: 2950-2955.
Selberg, O. und W. Burchert (1997): Die Positronen-Emissions-Tomographie in der Untersuchung des Muskelstoffwechsels. Akt. Ernähr.-Med. 22: 260-265.
Sheng, H.-P. (1988): Methodologies for Measuring Body Composition in Humans. In: “Designing Foods: Animal Product Options in the Marketplace”, National Academy Press, Washington, DC, 242-250. http://books.nap.edu/books/0309037956/html/R1.html
Sheng, H.P., R.A. Huggins, C. Garza, H.J. Evans, A.D. LeBlanc, B.L. Nichols, P.C. Johnson (1981): Total body sodium, calcium, and chloride measured chemically and by neutron activation in guinea pigs. Am. J. Physiol. 241: R419-422.
Sheng, H.-P., A.L. Adolph, E. O’Brien Smith, and C. Garza (1988): Body Volume and Fat-Free Mass Determinations by Acoustic Plethysmography. Pediatr. Res. 24: 85-89.
Shields, R.G. Jr., D.C. Mahan, P.L. Graham (1983): Changes in swine body composition from birth to 145 kg. J. Anim. Sci. 57: 43-54.
Shomer, N.H., C.F. Louis, and M. Fill (1993): Reconstitution of abnormalities in the malignant hyperthermia-susceptible pig ryanodine receptor. Am. J. Physiol. 264: C125-C135.
Shulman, G.I. (1997): Nuclear Magnetic Resonance Studies of Liver and Muscle Glycogen Metabolism in Humans. In: Emerging Technologies for Nutrition Research: Potential for Assessing
323

Literaturverzeichnis
Military Performance Capability (Eds.: S.J. Carlson-Newberry and Rebecca B. Costello), National Academy Press, Washington, DC, 259-272.
Shulman, R.J. (1993): The piglet can be used to study the effects of parenteral and enteral nutrition on body composition. J. Nutr. 123: 395-398.
Siconolfi, St.F., R.J. Gretebeck, and W.W. Wong (1995): Assessing total body protein, mineral, and bone mineral content from total body water and body density. J. Appl. Physiol. 79: 1837-1843.
Siebrits, F.K., E.H. Kemm, M.N. Ras and P.M. Barnes (1986): Protein deposition in pigs as influenced by sex, type and livemass. 1. The pattern and composition of protein deposition. S. Afr. J. Anim. Sci. 16: 23-27.
Siegrist-Kaiser, C.A., V. Pauli, C.E. Juge-Aubry, O. Boss, A. Pernin, W.W. Chin, I. Cusin, F. Rohner-Jeanrenaud, A.G. Burger, J. Zapf, and C.A. Meier (1997): Direct effects of leptin on brown and white adipose tissue. J. Clin. Invest. 100: 2858-2864.
Siemens A.L., R.J. Lipsey, W.M. Martin, M.G. Siemens, and H.B. Hedrick (1991): Composition of pork carcasses by potassium-40 liquid scintillation detection: estimation and validation. J. Anim. Sci. 69: 47-53.
Siervogel, R.M., W. Wisemandle, L.Michele Maynard, Sh.S. Guo, A.F. Roche, W.C. Chumlea, B. Towne (1998): Serial Changes in Body Composition Throughout Adulthood and Their Relationships to Changes in Lipid and Lipoprotein Levels - The Fels Longitudinal Study. Arterioscler. Thromb. Vasc. Biol. 18: 1759-1764.
Simm, G. (1994): Developments in improvement of meat sheep. Proc. 5th World Congress on Genetics Applied to Livestock Production, Guelph, Canada, Vol. 18: 3-10.
Simmen, F.A., L. Badinga, M.L. Green, I. Kwak, S. Song, and R.C.M. Simmen (1998): The Porcine Insulin-Like Growth Factor System: At the Interface of Nutrition, Growth and Reproduction. J. Nutr. 128: 315S-320S.
Siri, W.E. (1956): The gross composition of the body. In: Advances in Biological and Medical Physics. (C.A. Tobias and J.H. Lawrence eds.) 4: 239-280, Academic Press New York, NY, USA.
Siwek, Rosemarie A., L. Burkinshaw, C.B. Oxby, and P.A.J. Robinson (1984): Multi-element analysis of the obes subject by in vivo neutron activation analysis. Phys. Med. Biol. 29: 687-701.
Skjervold, H., K. GrØnseth, O. Vangen and A. Evensen (1981): In vivo estimation of body composition by computerized tomography. Z. Tierzüchtg. Züchtgsbiol. 98: 77-79.
Smidt, D. (1992): Schwerpunkte und Entwicklungstendenzen in der Nutztierforschung. Arch. Tierzucht (Dummerstorf) 35: 143-159.
Smidt, D., U. Baulain, M. Henning, E. Kallweit (1990): Anwendungsperspektiven der MR-Tomographie in der Tierzuchtforschung. Tag.-Ber., Akad. Landwirtsch.-Wiss. DDR, 287: 191-195.
324

Literaturverzeichnis
Smidt, D., U. Baulain, M. Henning, E. Kallweit (1991): Magnetische Resonanztomographie ( MRT) in der morphologisch orientierten Nutztierforschung. Wien. Tierärztl. Mschr. 78: 97-104.
Smith B.S., W.R. Jones, J.D. Hough, D.L. Huffman, W.B. Mikel, D.R. Mulvaney (1992): Prediction of carcass characteristics by real-time ultrasound in barrows and gilts slaughtered at three weights. J. Anim. Sci. 70: 2304-2308.
Smith, C. and P.R. Bampton (1977): Inheritance of reaction to halothane anaesthesia in pigs. Genet. Res. 29: 287-292.
Smith, C. and D.B. Smith (1993): The need for close linkage in marker-assisted selection for economic merit in livestock. Animal Breeding Abstracts 61: 197-204.
Smith, S.A., S.J. Montain, R.P. Matott, G.P. Zientara, F.A. Jolesz, R.A. Fielding (1998): Creatine supplementation and age influence muscle metabolism during exercise. J. Appl. Physiol. 85: 1349-1356.
Snijder, M.B., B.E. Kuyf, and P. Deurenberg (1999): Effect of body build on the validity of predicted body fat from body mass index and bioelectrical impedance. Ann. Nutr. Metab. 43: 277-285.
Solomon, M.B. and M.C. Dunn (1988): Simultaneous Histochemical Determination of Three Fiber Types in Single Sections of Ovine, Bovine, and Porcine Skeletal Muscle. J. Anim. Sci. 66: 255-264.
Solomon, M.B., R.G. Campbell, and N.C. Steele (1990): Effect of sex and exogenous porcine somatotropin on longissimus muscle fiber characteristics of growing pigs. J. Anim. Sci. 68: 1176-1181.
Soffner, P., A. Scholz, S. Nüske, M. Förster (2000): Erste Ergebnisse von Wachstumstudien am Schwein mit Hilfe der Dualenergie-Röntgenabsorptiometrie am Lehr- und Versuchsgut Oberschleißheim. DGfZ/GfT-Vortragstagung in Kiel, 20./21. September, A02.
Soffner, P., A. Scholz, E. Littmann, W. Peschke und M. Förster (2001): Untersuchungen zur Genauigkeit der Dualenergie-Röntgenabsorptiometrie für die Schlachtkörperanalyse beim Schwein anhand der EU-Referenzzerlegung. DGfZ/GfT-Gemeinschaftstagung, 12./13. September, Weihenstephan, D25.
Sonesson, A.K., K.H. de Greef, T.H.E. Meuwissen (1998): Genetic parameters and trends of meat quality, carcass composition and performance traits in two selected lines of Large White pigs. Livest. Prod. Sci. 57: 23-32.
Sonstegard, T.S., Rohrer, G.A., and Smith, T.P.L. (1998): Myostatin maps to porcine chromosome 15 by linkage and physical analyses. Animal Genetics 29:19-22.
Sorrentino, V., Virginia Barone, and Daniela Rossi (2000): Intracellular Ca2+ release channels in evolution. Current Opinion in Genetics & Development 10: 662-667.
Sosnicki, A. (1987): Histopathological observation of stress myopathy in M. longissimus in the pig and
325

Literaturverzeichnis
relationships with meat quality, fattening and slaughter traits. J. Anim. Sci. 65: 584-596.
Sönnichsen, Monika, A. Dobrowolski und R. Höreth (2001): Videobildauswertung an Schweineschlachtkörpern und Schlachtkörperanschnitten. Kurzfassungen der Fachvorträge, 36. Kulmbacher Woche, 8./9. Mai 2001. http://www.bfa-fleisch.de/36wochekurz.htm#Videobildauswertung
Spady, D.W., L.J. Filipow, T.R. Overton, W.A. Szymanski (1986): Measurement of total body potassium in premature infants by means of a whole-body counter. J. Pediatr. Gastroenterol. Nutr. 5: 750-755.
Speakman, J.R., D. Booles and R. Butterwick (2001): Validation of dual energy X-ray absorptiometry (DXA) by comparison with chemical analysis of dogs and cats. Intl. J. Obesity 25: 439-447.
Speer, V.C. (1988): Altering Carcass Measurements and Composition of the Pig. In: Designing Foods: Animal Product Options in the Marketplace. National Academy Press, Washington, DC: 273-277. http://books.nap.edu/books/0309037956/html/R1.html Spurlock, M.E., G.R. Frank, St.G: Cornelius, S. Ji, G.M. Willis, and Ch.A. Bidwell (1998): Obese Gene Expression in Porcine Adipose Tissue is reduced by Food Deprivation but not by Maintenace or Submaintenance Intake. J. Nutr. 128: 677-682.
Stalder, K.J., J. Maya, L.L. Christian, S.J. Moeller, and K.J. Prusa (1998): Effects of Preslaughter Management on the Quality of Carcasses from Porcine Stress Syndrome Heterozygous Market Hogs. J. Anim. Sci. 76: 2435-2433.
Standal, N. (1992): Live sheep evaluation using computerized tomography and other sophisticated methods. 43rd Annual Meeting of the EAAP, Madrid, 13-17 September, (paper copy) 9 p.
Stanford, K., S.D.M. Jones, M.A. Price (1998a): Methods of predicting lamb carcass composition: A review. Small Ruminant Research 29: 241-254.
Stanford, K., R.J. Richmond, S.D.M. Jones, W.M: Robertson, M.A. Price, and A.J. Gordon (1998b): Video image analysis for on-line classification of lamb carcasses. Animal Science 67: 311-316.
Steele, N.C. and L.T. Frobish (1976): Selected lipogenic enzyme activities of swine adipose tissue as influenced by genetic phenotype, age, feeding frequency and dietary energy source. Growth 40: 369-378.
Stern, S., L. Rhydmer, N. Lundeheim, P-E. Nyström, K. Johansson, and K. Andersson (1994): Performance, Puberty and Carcass Traits of Pigs Selected for High Lean Tissue Growth Rate on High or Low Protein Diets. Proc. 5th World Congr. Genet. Appl. Live. Prod., Guelph, Vol. 19: 49–52.
326

Literaturverzeichnis
Stiewe, H. (1986): Futteraufnahme und Futterverwertung – Die richtigen Tiere zur Mast einstellen. Schweinezucht und Schweinemast 10: 308-310 und 11: 348-351.
Stouffer, J.R., M.V. Wallentine, G.H. Wellington, and A. Diekmann (1961): Development and application of ultrasonic methods for measuring fat thickness and rib-eye area in cattle and hogs. J. Anim. Sci. 20: 759-767.
Stouffer, J.R. (1991): Using Ultrasound to Objectively Evaluate Composition and Quality of Livestock. 21st Century Concepts Important to Meat-Animal Evaluation, University of Wisconsin, Madison, WI: 49-54.
Streitz, E. (1995): Schätzung der Körperzusammensetzung wachsender Lämmer mit Hilfe der Magnet-Resonanz-Tomographie (MRT). Diss. Georg-August-Universität Göttingen, Landbauforschung Völkenrode, Sonderheft 156: 109 S.
Streitz, E., U. Baulain und E. Kallweit (1995): Untersuchungen zur Körperzusammensetzung wachsender Lämmer mit Hilfe der Magnet-Resonanz-Tomographie (MRT). Züchtungskunde 67: 392-403.
Strosberg, A.D. and T. Issad (1999): The involvement of leptin in humans revealed by mutations in leptin and leptin receptors. TiPS 20: 227-230.
Stroud, M.A., P. Ritz, W.A. Coward, M.B. Sawyer, D. Constantin-Teodosiu, P.L. Greenhaff, I.A. Macdonald (1997): Energy expenditure using isotope-labelled water (2H2 18O), exercise performance, skeletal muscle enzyme activities and plasma biochemical parameters in humans during 95 days of endurance exercise with inadequate energy intake. Eur. J. Appl. Physiol. 76: 243-252.
Sudi, K.M., R. Möller, E. Tafeit, K. Vrecko, R. Horejsi, G. Reibneger, P. Hofmann (1998): Die Beschreibung der subkutanen Fett-Topographie (SFTop) mittels Lipometer als Bindeglied zu traditionellen Bestimmungsmethoden der Körperzusammensetzung. Eine kritsiche Übersicht zu einem neuen Verfahren. Deutsche Zeitschrift für Sportmedizin 49: 46-52.
Sunoo, S., K. Asano, F. Mitsumori (1996): 31P nuclear magnetic resonance study on changes in phosphocreatine and intracellular pH in rat skeletal muscle during exercise at various inspired oxygen contents. Eur. J. Appl. Physiol. 74: 305-310.
Susenbeth, A. and K. Keitel (1988): Partition of Whole Body Protein in Different Body Fractions and Some Constants in Body Composition in Pigs. Livest. Prod. Sci. 20: 37-52.
Suster, D., B.J. Leury, J.D. Wark, D.J. Kerton, E. Ostrowska, and F. Dunshea (2000): Dual energy X-ray absorptiometry accurately predicts whole body and carcass composition in pigs. J. Anim. Sci. 78 (Suppl. 1) /J. Dairy Sci. 83 (Suppl. 1): 145.
Sutcliffe, J.F. (1996): A review of in vivo experimental methods to determine the composition of the human body. Phys. Med. Biol. 41:791-833.
327

Literaturverzeichnis
Sutcliffe, J.F., A.H. Smith, M.C.J. Barker, and M.A. Smith (1993): A theoretical analysis using ratios of the major elements measured by neutron activation analysis to derive total body water, protein, and fat. Med. Phys. 20: 1129-1134.
Sutcliffe, J.F., S.W. Smye, and M.A. Smith (1995): A further assessment of an electromagnetic method to measure body composition. Phys. Med. Biol. 40: 659-670.
Suzuki, H., J.D. Rounds, and D.W. Wilmore (1996): Does multifrequency bioelectrical impedance relate to body composition? J. Surg. Res. 65: 63-69.
Svendson, O.L., J. Haarbo, Ch. Hassager, C. Christiansen (1993): Accuracy of measurements of body composition by dual-energy x-ray absorptiometry in vivo. Am. J. Clin. Nutr. 57: 605-608.
Swantek, P.M., J.D. Crenshaw, M.J. Marchello, H.C. Lukaski (1992): Bioelectrical impedance: A nondestructive method to determine fat-free mass of live market swine and pork carcasses. J. Anim. Sci. 70: 169-177.
Swantek, P.M., M.J. Marchello, J.E. Tilton, J.D. Crenshaw (1999): Prediction of fat-free mass of pigs from 50 to 130 kilograms live weight. J. Anim. Sci. 77: 893-897.
Swatland, H.J. (1993): Growth Physiology and Post-mortem Metabolism in Porcine Muscle. In: Pork Quality: Genetic and Metabolic Factors (Eds.: E. Poulanne, D.J. Demeyer, M. Ruusunen and S. Ellis), CAB International, Wallingford, UK: 115-139.
Swatland, H.J. (1994): Structure and Development of Meat Animals and Poultry (2nd edition). Technomic Publishing, Lancaster, PA, USA.
Swensen K., M. Ellis, M.S. Brewer, J. Novakofski, F.K. McKeith (1998): Pork carcass composition: I. Interrelationships of compositional end points. J. Anim. Sci. 76: 2399-2404.
Szabo, G., G. Dallmann, G. Müller, L. Patthy, M. Soller, L. Varga (1998): A deletion in the myostatin gene causes the compact (Cmpt) hypermuscular mutation in mice. Mammalian Genome 9: 671-672.
Szabo, Cs., M.W. Verstegen, L. Babinszky, A.J.M. Jansman, O. Vangen, E. Kanis, P. Horn, R. Romvari, I: Takacs (1998): The possibility to estimate protein and fat content of pigs by computer tomography. J. Anim. Sci. 76 (Suppl. 1): 167.
Szabo, Cs., L. Babinszky, M.W.A. Verstegen, O. Vangen, A.J.M. Jansman, and E. Kanis (1999): The application of digital imaging techniques in the in vivo estimation of the body composition of pigs: a review. Livest. Prod. Sci. 60: 1-11.
Szendrõ, Zs., Horn, P., Kövér, Gy., Berényi, E., Radnai, I., Bíróné Németh, E. (1992): In vivo measurment of the carcass traits of meat type rabbits by X-ray computerised tomography. J. Appl. Rabbit. Res. 15: 799-809.
328

Literaturverzeichnis
Talbot, C., T. Preston, B.W. East (1986): Body composition of Atlantic salmon (Salmo salar L.) studied by neutron activation analysis. Comp. Biochem. Physiol. A 85: 445-450.
Tang, H., J. Vasselli, E. Wu, and D. Gallagher (2000): In vivo determination of body composition of rats using magnetic resonance imaging. Ann. N. Y. Acad. Sci. 904: 32-41.
Taylor, D.J., P. Styles, P.M. Mathews, D.L. Arnold, D.G. Gadian, P. Bore, G.K. Radda (1986): Energetics of human muscle: exercise-induced ATP depletion. Magn. Reson. Med. 3: 44-54.
Te Pas, M.F.W., A. Soumillion, F. L. Harders, F. J. Verburg, T. J. van den Bosch, P. Galesloot, and T.H.E. Meuwissen (1999): Influences of Myogenin Genotypes on Birth Weight, Growth Rate, Carcass Weight, Backfat Thickness, and Lean Weight of Pigs. J. Anim. Sci. 77: 2352–2356.
Terada, K., T. Miura, N. Kikuchi (1997): Digital image-based modeling applied to the homogenization analysis of composite materials. Computational Mechanics 20: 331-346.
Tess, M.W., G.E. Dickerson, J.A. Nienaber and C.L. Ferrell (1986): Growth, development and body composition in three genetic stocks of swine. J. Anim. Sci. 62: 968-979.
Thaller, G., L. Dempfle, Anna Schlecht, Sabine Wiedemann, H. Eichinger and R. Fries (2000): Effects of the MHS locus on growth, carcass and meat quality traits in F2 crosses between Mangalitza and Piétrain breeds. Arch. Tierz. (Dummerstorf) 43: 263-275.
Thielebein, J. (1994): Kernspintomographie an einem juvenilen Kragenbären nach Schädelimpressionsfraktur. Verh.ber. Erkrg. Zootiere 36: 175-180.
Thiet, W. und U. Baulain (1992): Technik und Anwendungsmöglichkeiten der Kernspintomographie beim Hund. Kleintierpraxis 37: 695-700.
Tholen, E., W. Peschke, U. Baulain und K. Schellander (1998a): Objektive Erfassung des Fleischanteils im Schweinebauch. 1. Mitteilung: Entwicklung von Schätzgleichungen aus Schlachtkörpermaßen. Züchtungskunde 70: 196-204.
Tholen, E., W. Peschke, U. Baulain und K. Schellander (1998b): Fleischanteil im Bauch objektiv erfassen. Schweinezucht und Schweinemast 46/4: 12-14.
Thomas, E. Louise, N. Saeed, J.V. Hajnal, A. Brynes, A.P. Goldstone, G. Frost, and J.D. Bell (1998): Magnetic resonance imaging of total body fat. J. Appl. Phys. 85: 1778-1785.
Thompson, J.M. and B.P. Kinghorn (1992): CATMAN - A program to measure CAT-Scans for prediction of body composition in live animals. Proc. of the 10th Conf. of Austr. Ass. of Animal Breeding and Genetics, Rockhampton, Australia, 5 pp.
329

Literaturverzeichnis
Thompson, J.M., B.P. Kinghorn and R.G. Banks (1996): Application of non-invasive techniques for carcass evaluation in breeding. 47th Annual Meeting of the EAAP, Lillehammer, Norway, August 25-29, 8 pp.
Thompson, A., A. Damyanovich, Annie Madapallimattam, D. Mikalus, Johane Allard, and Khursheed N. Jeejeebhoy (1998): 31P-nuclear magnetic resonance studies of bioenergetic changes in skeletal muscle in malnourished human adults. Am. J. Clin. Nutr. 67: 39–43.
Tibau, J., J. Soler, X. Puigvert, M. Gispert, X. Cufi, A. Sanchez (1996): INVIVO: A software application for carcass evaluation in live pigs. 47th Annual Meeting of the EAAP, August, Lillehammer, Norway, (paper copy, 6 pages).
Tölli, J., B.-Å. Bengtson, I. Bosaeus, G. Johansson, and M. Alpstein (1998): A Comparison of Different Methods to Measure Body Composition in Patients. Appl. Radiat. Isot. 49: 469-472.
Topel, D.G. and R. Kauffman (1988): Live Animal and Carcass Measurement. In: Designing Foods: Animal Product Options in the Marketplace. National Academy Press, Washington, DC, pp. 258-272. http://books.nap.edu/books/0309037956/html/R1.html Tothill, P., A. Avenell, J. Love, and D.M. Reid (1994): Comparisons between Hologic, Lunar and Norland dual-energy X-ray absorptiometers and other techniques used for whole-body soft tissue measurements. European Journal of Clinical Nutrition 48: 781-794.
Tothill, P., W.J. Hannan, S. Cowen, C.P. Freeman (1997): Anomalies in the Measurement of Changes in Total-Body Bone Mineral by Dual-Energy X-Ray Absorptiometry During Weight Change. Journal of Bone and Mineral Research 12: 1908-1921.
Tornsten, A., A. Wraith, D. Larhammer, and B.P. Chowdhary (1998): FISH mapping of the porcine NPY5 gene to chromosome 8p11. Mammalian Genome 9: 262-263.
Toussant, J.-F., K.K. Kwong, F. M’Kparu, R.M. Weisskopf, P.J. LaRaia, and H.L. Kantor (1996): Interrelationship of oxidative metabolism and local perfusion demonstrated by NMR in human skeletal muscle. J. Appl. Physiol. 81: 2221-2228.
Towner, R.A., A. Lirette, R.J. Etches, S. Chu, A. Rath, J. Millam, B. Cutler (1991): Phosphorous-31 Localized Spectroscopy of growing chicken embryos (Days 2-19). Proc. 12th Symposium Energy Metabolism of Farm Animals, Kartause Ittingen, Switzerland, 1-7 September: 234-237.
Trattnig, S., G. Kontaxis, M. Breitenseher, Ch. Czerny, Th. Rand, K. Turetschek, M. Barth, und H. Imhof (1997): MRT auf Niederfeldtomographen (0,2 Tesla): Ein quantitativer Vergleich mit einem Gerät mittlerer Feldstärke (1,0 Tesla). Der Radiologe 37: 773-777.
Treitl, U. (1996): Applicability of the dilution method using D2O to determine body composition of growing kids. Proc. 47th Annual Meeting of the EAAP, Lillehammer, Norway, August 25-29, 277.
330

Literaturverzeichnis
Treitl, U. (1998): Untersuchungen zur Bestimmung der Körperzusammensetzung wachsender Ziegenlämmer in vivo mit Hilfe der D20-Verdünnungsmethode. Diss. Humboldt-Universität zu Berlin.
Trippo, U. (2000): Körperbau, Körperzusammensetzung und Ernährungsgewohnheiten bei Erwachsenen in Abhängigkeit von Alter und Geschlecht. Diss., Universität Potsdam.
Tschümperlin, K. (1996): Schätzung der Körperzusammensetzung am lebenden Rind. Thesis, Nr. 11447, ETH Zürich, Schweiz.
Tuchscherer, Margret und G. Manteuffel (2000): Die Wirkung von psychischem Streß auf das Immunsystem. Ein weiterer Grund für tiergerechte Haltung (Übersichtsreferat). Arch. Tierzucht (Dummerstorf) 43: 547-560.
Uhrin, P. and Liptaj, T. (1990): 31P-NMR study of metabolism of the pig muscles post mortem. Proc. 41st Annual Meeting of European Association of Animal Production, Toulouse, July, 9-12: 312.
Ulin, K. and R.G. Zamenhof (1986): Measurement of total-body oxygen, nitrogen, and carbon in vivo by photon activation analysis. Med. Phys. 13: 887-897.
Valle-Zarate, Anne (1981): Schätzung von Populationsparametern für die Futteraufnahme bei Schweinen der Deutschen Landrasse. Diss. Univ. Hohenheim.
Van Betteray, K. (1994): Einsatz des Ultraschall-B-Scan-Verfahrens in der Schlachtindustrie. Landbauforschung Völkenrode, Sonderheft 145: 74-78.
Van den Bergh, A., S. Houtman, A. Heerschap, Nancy J. Reherer, H.J. van den Boogert, B. Oeseburg, and Maria T.E. Hopman (1996): Muscle glycogen recovery after exercise during glucose and fructose intake monitored by 13C-NMR. J. Appl. Physiol. 81: 1495-1500.
Van den Thillart, G., F. Körner, A. van Waarde, C. Erkelens, and J. Lugtenburg (1989): A Flow-Through Probe for in Vivo 31P NMR Spectroscopy of Unanesthetized Aquatic Vertebrates at 9,4 Tesla. J. Magn. Resonance 84: 573-579.
Van Erp, A.J.M, F. Gerbens, J.H. Veerkamp, M.F.W. te Pas, J.W.M. Merks and T.H.E. Meuwissen (1997): Effects of fatty acid-binding protein (H-FABP) on intramuscular fat and other production traits in pigs. 48th Annual Meeting of the European Association for Animal Production, Book of Abstracts No. 3: 4.
Van Logtestijn, J.G. und W. Sybesma (1967): Muskel- und Fleischdegenerationen bei Schweinen. Dtsch. Tierärztl. Wschr. 74: 377-385.
Van Steenbergen, E. J. (1990): Relevance of exterior appraisal in pig breeding. Doctoral thesis, Wageningen, Netherlands.
Van Waarde, A., G. van den Thillart, C. Erkelens, A. Addink, and J. Lugtenburg (1990):
331

Literaturverzeichnis
Functional Coupling of Glycolysis and Phosphocreatine Utilization in Anoxic Fish Muscle – AN IN VIVO 31P NMR STUDY. J. Biol. Chem. 265: 914-923.
Vangen, O. (1988a): X-Ray CT for Body Composition. In: E. Kallweit - M. Henning - E. Groeneveld ”Application of NMR Techniques on the Body Composition of Live Animals”. - EC-Seminar, Mariensee, 14-15 June, Elsevier Applied Science, London, New York: 91-99.
Vangen, O. (1988b): Experience from several years of using computerized tomography (CT) in animal breeding research. Proc. VI World Conference on Animal Production, Helsinki, Finland, 27th June - 1st July, P4.37.
Vangen, O. (1992): Assessing body composition of pigs by computer assisted tomography. Pig News and Information 13: 159N-162N.
Vangen, O. and H. Enting (1990): Intramuscular fat in different Duroc-crosses estimated by computerized tomography (CT). Proc. Seminar on Meat Quality in Slaughter Animals, NJF-Seminar 183: 243-252.
Vangen, O. and J.M. Thompson (1992): The use of CAT-scanning to measure maintenance efficiency in genetic lines of sheep. 43rd Annual Meeting of the EAAP, Madrid, 14-17 September, 10 pp.
Vangen, O. and N.B. Jopson (1996): Research application of non-invasive techniques for body composition. 47th Annual Meeting of the EAAP, Lillehammer, Norway, August 25-29, 8 pp.
Varga, L., G. Szabó, A. Darvasi, G. Müller, M. Sass, and M. Soller (1997): Inheritance and Mapping of Compact (Cmpt), a New Mutation Causing Hypermuscularity in Mice. Genetics 147: 755-764.
Vartsky, D., K.J. Ellis, S.H. Cohn (1979): In vivo measurement of body nitrogen by analysis of prompt gammas from neutron capture. J. Nucl. Med. 20: 1158-1165.
Vartsky, D., M.B. Goldberg, D. Bar, A. Goldschmidt, G. Feldman, E. Sayag, D. Katz, I.M. Stronach, J.W. Stark, W.V. Prestwich, F.E. McNeill, D.R. Chettle (2000): Gamma ray nuclear resonance absorption: an alternative method for in vivo body composition studies. Ann. N. Y. Acad. Sci. 904: 236-246.
Verburg, M.P., F.T.J.J. Oerlemans, C.A. van Bennekom, M.J.M. Gielen, C.H.M.M. de Bruyn and J.F. Crul (1984): In vivo induced malignant hyperthermia in pigs. I. Physiological and biochemical changes and the influenec of Dantrolene Sodium. Acta Anaesthesiol. Scand. 28: 1-8.
Villé, H., C. Decanniere, R. Geers, P. van Hecke, V. Goedseels, G. Maes, L. Bosschaerts, J. Deley, S. Janssens (1991): In vivo 1H-NMR-spectroscopy on muscle tissue of piglets: determination of intramuscular fat content. Proc. 12th Symposium Energy Metabolism of Farm Animals, Kartause Ittingen, Switzerland, 1-7 September: 218-221.
Villé, H., G. Rombouts, P. van Hecke, S. Perremans, G. Maes, G. Spincemaille, R. Geers (1997): An Evaluation of Ultrasound and Nuclear Magnetic Resonance Spectroscopy to
332

Literaturverzeichnis
Measure In Vivo Intramuscular Fat Content of Longissimus Muscle of Pigs. J. Anim. Sci. 75: 2942-2949.
Vincent, A.L., L. Wang and M.F. Rothschild (1997): Linkage mapping of the leptin receptor gene to pig chromosome 6. J. Anim. Sci. 75 (Suppl. 1): 145.
Virgili, F., G. Maiani, Z.H. Zahoor, D. Ciarapica, A. Raguzzini, A. Ferro-Luzzi (1994): Relationship between fat-free mass and urinary excretion of creatinine and 3-methylhistidine in adult humans. J. Appl. Physiol. 76: 1946-1950.
VNMR (1992-1994): Software Versionen 4.1 - 4.3, Varian NMR Systems, 3120 Hansen Way, Palo Alto, CA 94304.
Vogel Hertzel, Ann and D.A. Bernlohr (2000): The Mammalian Fatty Acid-binding Protein Multigen Family: Molecular and Genetic Insights into Function. Trends in Endocrinology and Metabolism 11: 175-180.
Von Borell, E. (2000): Stress and coping in farm animals. Arch. Tierz. (Dummerstorf) 43 (Sonderheft): 144-152.
Von Felde, A., R. Röhe, E. Kalm (1996): Genetische Analyse von Merkmalen der Mastleistung und des Schlachtkörperwertes stationsgeprüfter Jungeber in Einzel- und Gruppenhaltung mittels Gibbs-Sampling. Züchtungskunde 68: 305-318.
Von Lengerken, G., S. Maak, M. Wicke, I. Fiedler, K. Ender (1994): Suitability of structural and functional traits of skeletal muscle for the genetic improvement of meat quality in pigs. Arch. Tierzucht (Dummerstorf) 37: 133-143.
Von Lengerken, G., M. Wicke und S. Maak (1997): Streßempfindlichkeit und Fleischqualität - Stand und Perspektiven in Praxis und Forschung. Arch. Tierzucht (Dummerstorf) 40 (Sonderheft): 163-171.
Wäfler, P., P. Morel und C. Herwig (1984): Die Bedeutung des Futterverzehrs in der Schweinezucht. - 1. Zusammenhänge zwischen Appetit und den Mast- und Schlachtleistungen bei Schweinen unter ad libitum Fütterung. Schweizer Landwirtschaftliche Monatshefte 62: 337-343.
Wagenknecht, T. and M. Radermacher (1997): Ryanodine receptors: structure and macromolecular interactions. Current Opinion in Structural Biology 7: 258-265.
Wagner, J.R., A.P. Schinckel, W. Chen, J. C. Forrest, and B.L. Coe (1999): Analysis of Body Composition Changes of Swine During Growth and Development. J. Anim. Sci. 77: 1442–1466.
Walling, G.A., A.L. Archibald, P.M. Visscher, C.S. Haley (1998): Mapping of quantitative trait loci on chromosome 4 in a Large White x Meishan pig F2 population. Proc. 6th World Congress on Genetics Applied to Livestock Production, Armidale, Australia, Vol. 23: 519-522.
Walstra, P. (1980): Growth and carcass composition from birth to maturity in relation to feeding level and sex in Dutch Landrace pigs. Mededelingen Landbouwhogeschool, Wageningen, NL. 80 (4): 1-206.
333

Literaturverzeichnis
Walstra, P. and G.S.M. Merkus (1995): Procedure for the assessment of the lean meat percentage as a consequence of the new EU reference dissection method in pig carcass classification. Paper based on discussions in the EU Management Committee on Pig Meat and on discussions with dissection experts during a meeting on May 18-19, 1994 at Zeist, NL, DLO-Research Institute for Animal Science and Health (ID-DLO), Zeist, NL, p. 24.
Wang, J. and P. Deurenberg (1996): The validity of predicted body composition in Chinese adults from anthropometry and bioelectrical impedance in comparison with densitometry. Brit. J. Nutr. 76: 175-182.
Wang, L., T.P. Yu, H.W. Chung, C.K. Tuggle and M.F. Rothschild (1997): Pinpointing QTLs on Chromosome 7 for Performance Traits in the Pig. J. Anim. Sci. 75 (Suppl. 1): 145.
Wang, Z., R.N. Pierson Jr., S.B. Heymsfield (1992): The five level model: a new approach to organizing body composition research. Am. J. Clin. Nutr. 56:19-28.
Wang, Z., P. Deurenberg, SS. Guo, A. Pietrobelli, J. Wang, R.N. Pierson Jr., S.B. Heymsfield (1998a): Six-compartment body composition model: Inter-method comparisons of total body fat measurement. Intl. J. Obesity 22: 329-337.
Wang, Z., P. Deurenberg, W. Wang, R.N. Pierson Jr., and S.B. Heymsfield (1998b): Fraction of Carbon-Free Body Mass as Oxygen is a Constant Body Composition Ratio in Men. J. Nutr. 128: 1008-1010.
Wang, Z., P. Deurenberg, D.E. Matthews, and S.B. Heymsfield (1998c): Urinary 3-methylhistidine excretion: association with total body skeletal muscle mass by computerized axial tomography. J. Parenter. Enteral. Nutr. 22: 82-86.
Wang, Z., P.Deurenberg, W. Wang, A. Pietrobelli, R.N. Baumgartner, and St. B. Heymsfield (1999): Hydration of fat-free body mass: new physiological approach. Am. J. Physiol. 276: E995-E1003.
Wang, Z., P. Deurenberg, and S.B. Heymsfield (2000): Cellular-Level Body Composition Model. A New Approach to Studying Fat-free Mass Hydration. Annals of the New York Academy of Sciences (In Vivo Body Composition Studies, Editors: S. Yasumuru, J. Wang, and R.N. Pierson Jr.) 904: 306-311.
Wappler, O. (1997): Vergleichende Untersuchungen zur Aussage des Harnstoffverteilungsraumes, der Lebendmasse und der Rückenfettdicke zum Körperfettgehalt der Milchkuh. Dissertation, Freie Universität Berlin.
Warden, C.H., J.S. Fisler, S.M. Shoemaker, P.Z. Wen, K.L. Svenson, M.J. Pace and A.J. Lusis (1995): Identification of four chromosomal loci determining obesity in a multifactorial mouse model. J. Clin. Invest. 95: 1545-1552.
Warriss, P.D., S.N. Brown, J.G. Franklin, and S.C. Ketsin (1990): The Thickness and Quality of Backfat in Various Pig Breeds and Their Relationship to Intramuscular Fat and the Setting of Joints from the Carcasses. Meat Sci. 38: 21-29.
334

Literaturverzeichnis
Weaver, S.A., W.T. Dixon, and A.L. Schaefer (2000): The effects of mutated skeletal ryanodine receptors on hypothalamic-pituitary-adrenal axis function in boars. J. Anim. Sci. 78: 1319-1330.
Webb, A.J. (1989): Genetics of food intake in the pig. - in: The Voluntary Food Intake of Pigs. Occasional Public. No. 13 - British Society of Animal Production (Eds.: Forbes, J.M.; Varley, M.A.; Lawrence, T.L.J.).
Webb, A.J. and M.K. Curran (1986): Selection regime by production system interaction in pig improvement: a review of possible causes and solutions. Livest. Prod. Sci. 14: 41 – 54.
Webb, A.J., B. Grundy, P. Kitchin (1994): Within-litter effect of the HAL-1843 heterozygote on lean growth in pigs. Proc. 5th World Congress on Genetics Applied to Livestock Production, Guelph, Canada, Vol. 17: 421-424.
Weigle, D.S. and J.L. Kuijper (1996): Obesity genes and the regulation of body fat content. BioEssays 18: 867-874.
Weiß, D. (2001a): BSE-Krise erfasst ganz Europa. Marktkommentar - ZMP-Zentralbericht (01.03.2001). http://www.zmp.de/presse/zb/zbko9.pdf (Stand: 7.7.2001)
Weiß, D. (2001b): MKS bedroht den europäischen Schweinemarkt. Marktkommentar - ZMP-Zentralbericht (30.03.2001). http://www.zmp.de/presse/zb/zbko13.pdf (Stand: 7.7.2001)
Wells, P.N.T. (1991): The description of animal form and function. Livest. Prod. Sci. 27: 19-33.
Wells, J.C.K., I: Douros, N.J. Fuller, M. Elia, and L. Dekker (2000): Assessment of Body Volume Using Three-Dimensional Photonic Scanning. Annals of the New York Academy of Sciences (In Vivo Body Composition Studies, Editors: S. Yasumuru, J. Wang, and R.N. Pierson Jr.) 904: 247-254.
Wenk, C. (2000): Meat consumption and meat quality – How important is it? In: Quality of meat and fat in pigs as affected by genetics and nutrition (Eds.: C. Wenk, J.A. Fernández, and M. Dupuis). EAAP publication No. 100: 7-12.
Wentzel, G. (1994): Erläuterungen zur MR-Spektroskopie. Landbauforschung Völkenrode. Sonderheft 145: 79-83.
West, D.B., J. Goudey-Lefevre, B. York and G.E. Truett (1994): Dietary obesity linked to genetic loci on chromosome 9 and 15 in a polygenic mouse model. J. Clin. Invest. 94: 1410-1416.
West, D.B., J. Waguespack, B. York, J. Goudey-Lefevre and R.A. Price (1994): Genetics of dietary obesity in AKR/J x SWR/J mice: segregation of the trait and identification of a linked locus on chromosome 4. Mamm. Genome 5: 546-552.
White, B.R., Y.H. Lan, F.K. McKeith, J. Novakofski, M.B. Wheeler, and D.G. McLaren (1995):
335

Literaturverzeichnis
Growth and Body Composition of Meishan and Yorkshire Barrows and Gilts. J. Anim. Sci. 73: 738-749.
Whittemore, C.T. (1985): The application of the principles of nutrition to the feeding of breeding sows and the production of meat from growing pigs. S. Afr. J. Anim. Sci. 15: 97-101.
Whittemore, C.T. (1987): Elements of PIG SCIENCE. Longman handbooks in agriculture.
Whittemore, C.T., J.B. Tullis, and G.C. Emmans (1988): Protein growth in pigs. Anim. Prod. 46: 437-445.
Whittemore, C.T., J.C. Kerr & N.D. Cameron (1995): An Approach to Prediction of Feed Intake in Growing Pigs Using Simple Body Measurements. Agricultural Systems 47: 235-244.
Wicke, M., G. von Lengerken, I. Fiedler, M. Altmann, und K. Ender (1991): Einfluß der Selektion nach Merkmalen der Muskelstruktur des M. longissimus dorsi auf Belastungsempfindlichkeit und Fleischbeschaffenheit beim Schwein. Fleischwirtschaft 71: 437-442.
Wicke, M., G. von Lengerken, S. Maak und I. Fiedler (1993): Vorhersage von PSE-Fleisch mittels biochemsicher und morphologischer Merkamle der Skelettmuskulatur am lebenden Schwein. Arch. Tierzucht (Dummerstorf) 36: 631-638.
Wiederhold, S. (1996): In vivo Messungen ausgewählter Merkmale der Körperzusammensetzung bei Wassergeflügel mit Hilfe der Magnet-Resonanz-Tomographie (MRT). Diss. Halle (Saale), Mariensee, 121 S.
Wilmore, K.M., P.J. McBride, and J.H. Wilmore (1994): Comparison of bioelectric impedance and near-infrared interactance for body composition assessment in a population of self-perceived overweight adults. Intl. J. Obesity 18: 375-381.
Wilson, B.D., M.M. Ollmann, and G.S. Barsh (1999): The role of agouti-related protein in regulating body weight. Molecular Medicine Today 5: 250-256.
Windhorst, H.-W. (1998): Host country number one in EU. But, supply shortfalls demand immediate attention. Pig Progress 14: 9-14.
Windisch, W., G.G. Gotterbarm, F.X. Roth, D. Schams und M. Kirchgessner (2000): Einfluss der genetischen Herkunft von Mastschweinen (Deutsche Landrasse, Pietrain) auf Kenndaten des Proteinstoffwechsels. Züchtungskunde 72: 379-388.
Wishmeyer D.L., G.D. Snowder, D.H. Clark, and N.E. Cockett (1994): Prediction of live lamb chemical composition utilizing electromagnetic scanning (ToBEC). J. Anim. Sci. 74: 1864-1872.
Wittenberg, Beatrice A. and J.B. Wittenberg (1989): Transport of Oxygen in muscle. Annu. Rev. Physiol. 51: 857-878.
Wittmann, W., K.-U. Götz, W. Peschke, J.-P. Lindner und M. Hauser (1998): Einfluß des MHS-Genotyps auf die Mast- und Schlachtleistung von Pietrainschweinen
336

Literaturverzeichnis
und PI x DL-Mastendprodukten in der Stationsprüfung. Arch. Tierzucht (Dummerstorf) 42: 139-147.
Wolf, E., H. Lahm, M. Wu, R. Wanke, A. Hoeflich (2000): Effects of IGFBP-2 overexpression in vitro and in vivo. Pediatr. Nephrol. 14: 572-578.
Wolf-Schwerin, Chr. und E. Kallweit (1991): Belastungsreaktionen und Leistungsmerkmale von definierten Halothangenotypen der Deutschen Landrasse. Züchtungskunde 63: 51-64.
Wolz, W. (1999): Molekulargenetische Analyse der Central Core Disease. Dissertation, Bayerische Julius-Maximilians-Universität Würzburg. http://erl.bibliothek.uni-wuerzburg.de/DISS/Biologie/1999/X116679/html/index.htm
Wood, J.D. and N.D. Cameron (1994): Genetics of Meat Quality in Pigs. Proc. 5th World Congress on Genetics Applied to Livestock Production, Guelph, Canada, Vol. 19: 458-464.
Wolter, Maren (1997): Schnittbildanatomie des Rumpfbereiches des Schweines anhand der Magnetresonanztomographie. Diss., Justus-Liebig-Univ. Gießen.
Wray-Cahen, C.D., D.E. Kerr, Ch.M. Evock-Clover, and N.C. Steele (1998): Redefining Body Composition: Nutrients, Hormones, and Genes in Meat Production. Annu. Rev. Nutr. 18: 63-92.
Xie, C., E. Albrecht, J. Wegner, G.A. Brockmann, C. Kazala, R.J. Weselake, K. Ender (1999): Leptin, ein Molekül für die Schmackhaftigkeit? - Ein Überblick. Arch. Tierzucht (Dummerstorf) 42: 191-199.
Xiong, Youling L. (1994): Myofibrillar Protein from Different Muscle Fiber Types: Implications of Biochemical and Functional Properties in Meat Processing. Critical Reviews in Food Science and Nutrition 34: 293-320.
Yasumura, S., K. Jones, P. Spanne, G. Schidlovsky, L. Wielopolski, X. Ren, D. Glaros, and Y. Xatzikonstantinou (1993): In vivo animal models of body composition in aging. J. Nutr. 123 (2 Suppl.): 459-464.
Yasumura S., I.E. Stamatelatos, C.N. Boozer, R. Moore, and R. Ma (1998): In vivo body composition studies in rats: assessment of total body protein. Appl. Radiat. Isot. 49: 731-732.
York, B., K. Lei and D.B. West (1996): Sensitivity to dietary obesity linked to a locus on chromosome 15 in a CAST/Ei x C57BL/6J F2 intercross. Mamm. Genome 7: 677-681.
Yoshiyama, M., M. Ishikawa, I. Miura, K. Takeuchi and T. Takeda (1994): Time course of the recovery of adenosine triphosphate content with adenosine in post-ischemic hearts - A 31P Magnetic Resonance Spectroscopy Study -. Japanese Circulation Journal 58: 662-670.
Yu, T.P., C.K. Tuggle, C.B. Schmitz, M.F. Rothschild (1995): Association of PIT1 Polymorphisms with Growth and Carcass Traits in Pigs. J. Anim. Sci. 73: 1282-1288.
337

Literaturverzeichnis
Youatt, W. and W.C.L. Martin (1865): The Hog; A treatise on the breeds, management, feeding and medical treatment of swine; with directions for salting pork and curing and baking of hams. (edited by A. Stevens) Orange Judd, New York. http://cdl.library.cornell.edu/dienst/nehbrowse.fly/MOA-JOURNALS2:youa-0001/15/1:TIFF2GIF:100?,,
Zemel, B.S., E.M. Riley, V.A. Stallings (1997): EVALUATION OF METHODOLOGY FOR NUTRITIONAL ASSESSMENT IN CHILDREN: Anthropometry, Body Composition, and Energy Expenditure. Annu. Rev. Nutr. 17: 211-235.
Žgur, S. (1991): Histologische und biochemische Veränderungen im Muskelgewebe wachsender Schweine. Diss. Göttingen, 83 S.
Zhang, Y., R. Proenca, M. Maffei, M. Barone, L. Leopold, J.M. Friedman (1994): Positional cloning of the mouse obese gene and its human homologue. Nature 372: 425-432.
Zupke, C. and B. Foy (1995): Nuclear magnetic resonance analysis of cell metabolism. Current Opinion in Biotechnology 6: 192-197.
338

Anhang I
339
Anhang I
Zusätzliche 1H-MRT-Ergebnisse
Nach Rassezusammensetzung (30, 60, 90 kg Lebendmasse)
30 kg Gruppe
Tab. A1: Volumen des Schinkenmagergewebes einschließlich Knochen (SMV) und des
subkutanen Fettes einschließlich Haut (SFETTV) sowie des gesamten Schinkens (SGV)
und das Fett-zu-Muskel-Verhältnis (SFMV) in einer 15 cm MRT-Region für Schweine
mit unterschiedlicher Rassezusammensetzung bei ca. 30 kg Lebendmasse (LSM ±
SEE#)
n SMV (cm3) SFETTV (cm3) SFMV SGV (cm3)
Du 21 3920,98 ± 47,01 a 756,79 ± 19,17 a 0,195 ± 0,006 ac 4677,77 ± 47,37 a
Ha 8 3853,29 ± 80,52 a 674,62 ± 32,83 b 0,176 ± 0,010 a 4527,91 ± 81,14 ac
LR 8 3702,70 ± 96,19 a 668,86 ± 39,22 a 0,182 ± 0,012 a 4371,55 ± 96,93 bc
PCh x LR 7 3453,28 ±102,17 b 756,83 ± 41,66 ab 0,217 ± 0,013 bc 4210,11 ±102,96 b
S 3 3656,90 ±128,86 ab 940,17 ± 52,54 c 0,259 ± 0,016 b 4597,06 ±129,86 ac
#) Kleinste Quadrate Mittelwerte mit unterschiedlichen Buchstaben sind signifikant verschieden
(p ≤ 0,05).
Tab. A2: Volumen der Mm. longissimi dorsi (MLDV) und des subkutanen Fettes
einschließlich Haut (FETTV) sowie das Fett-zu-Muskel-Verhältnis (FMV) in einer
10 cm MRT-Region für Schweine mit unterschiedlicher Rassezusammensetzung bei ca.
30 kg Lebendmasse (LSM ± SEE#)
n MLDV (cm3) FETTV (cm3) FMV
Du 21 286,76 ± 9,65 a 92,78 ± 4,67 a 0,338 ± 0,018 a
Ha 8 351,62 ± 16,53 b 90,64 ± 8,00 a 0,265 ± 0,031 ab
LR 8 326,68 ± 19,77 ab 79,96 ± 9,57 a 0,233 ± 0,037 b
PCh x LR 7 282,17 ± 21,09 a 89,43 ± 10,21 a 0,310 ± 0,039 ab
S 3 289,88 ± 31,43 ab 142,09 ± 15,21 b 0,512 ± 0,059 c
#) Kleinste Quadrate Mittelwerte mit unterschiedlichen Buchstaben sind signifikant verschieden
(p ≤ 0,05).
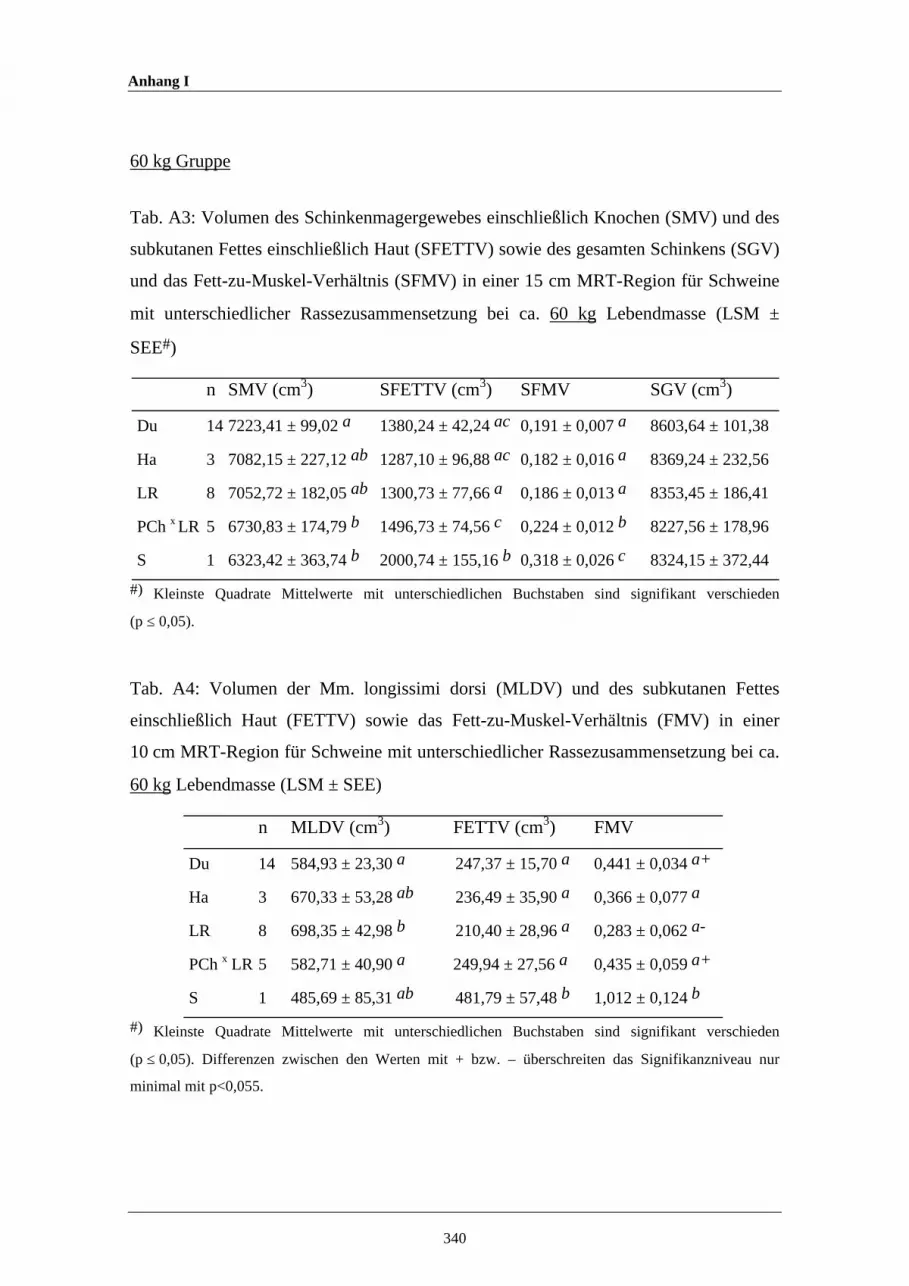
Anhang I
340
60 kg Gruppe
Tab. A3: Volumen des Schinkenmagergewebes einschließlich Knochen (SMV) und des
subkutanen Fettes einschließlich Haut (SFETTV) sowie des gesamten Schinkens (SGV)
und das Fett-zu-Muskel-Verhältnis (SFMV) in einer 15 cm MRT-Region für Schweine
mit unterschiedlicher Rassezusammensetzung bei ca. 60 kg Lebendmasse (LSM ±
SEE#)
n SMV (cm3) SFETTV (cm3) SFMV SGV (cm3)
Du 14 7223,41 ± 99,02 a 1380,24 ± 42,24 ac 0,191 ± 0,007 a 8603,64 ± 101,38
Ha 3 7082,15 ± 227,12 ab 1287,10 ± 96,88 ac 0,182 ± 0,016 a 8369,24 ± 232,56
LR 8 7052,72 ± 182,05 ab 1300,73 ± 77,66 a 0,186 ± 0,013 a 8353,45 ± 186,41
PCh x LR 5 6730,83 ± 174,79 b 1496,73 ± 74,56 c 0,224 ± 0,012 b 8227,56 ± 178,96
S 1 6323,42 ± 363,74 b 2000,74 ± 155,16 b 0,318 ± 0,026 c 8324,15 ± 372,44
#) Kleinste Quadrate Mittelwerte mit unterschiedlichen Buchstaben sind signifikant verschieden
(p ≤ 0,05).
Tab. A4: Volumen der Mm. longissimi dorsi (MLDV) und des subkutanen Fettes
einschließlich Haut (FETTV) sowie das Fett-zu-Muskel-Verhältnis (FMV) in einer
10 cm MRT-Region für Schweine mit unterschiedlicher Rassezusammensetzung bei ca.
60 kg Lebendmasse (LSM ± SEE)
n MLDV (cm3) FETTV (cm3) FMV
Du 14 584,93 ± 23,30 a 247,37 ± 15,70 a 0,441 ± 0,034 a+
Ha 3 670,33 ± 53,28 ab 236,49 ± 35,90 a 0,366 ± 0,077 a
LR 8 698,35 ± 42,98 b 210,40 ± 28,96 a 0,283 ± 0,062 a-
PCh x LR 5 582,71 ± 40,90 a 249,94 ± 27,56 a 0,435 ± 0,059 a+
S 1 485,69 ± 85,31 ab 481,79 ± 57,48 b 1,012 ± 0,124 b
#) Kleinste Quadrate Mittelwerte mit unterschiedlichen Buchstaben sind signifikant verschieden
(p ≤ 0,05). Differenzen zwischen den Werten mit + bzw. – überschreiten das Signifikanzniveau nur
minimal mit p<0,055.

Anhang I
341
90 kg Gruppe
Tab. A5: Volumen des Schinkenmagergewebes einschließlich Knochen (SMV) und des
subkutanen Fettes einschließlich Haut (SFETTV) sowie des gesamten Schinkens (SGV)
und das Fett-zu-Muskel-Verhältnis (SFMV) in einer 15 cm MRT-Region für Schweine
mit unterschiedlicher Rassezusammensetzung bei ca. 90 kg Lebendmasse (LSM ±
SEE#)
n SMV (cm3) SFETTV (cm3) SFMV SGV (cm3)
Du 9 9042,68 ± 120,33 a 1674,52 ± 107,48 0,187 ± 0,013 10717,19 ± 151,55
Ha 2 8906,33 ± 265,98 ab 1776,77 ± 237,57 0,200 ± 0,027 10683,11 ± 334,99
.LR 2 9230,13 ± 297,48 a 1896,04 ± 265,70 0,205 ± 0,031 11126,16 ± 374,65
PCh x LR 4 8424,02 ± 179,89 b 2031,16 ± 160,67 0,240 ± 0,019 10455,20 ± 226,56
#) Kleinste Quadrate Mittelwerte mit unterschiedlichen Buchstaben sind signifikant verschieden
(p ≤ 0,05).
Tab. A6: Volumen der Mm. longissimi dorsi (MLDV) und des subkutanen Fettes
einschließlich Haut (FETTV) sowie das Fett-zu-Muskel-Verhältnis (FMV) in einer
10 cm MRT-Region für Schweine mit unterschiedlicher Rassezusammensetzung bei ca.
90 kg Lebendmasse (LSM ± SEE#)
n MLDV (cm3) FETTV (cm3) FMV
Du 9 764,76 ± 21,83 374,54 ± 35,76 0,519 ± 0,048
Ha 2 842,76 ± 48,26 349,06 ± 79,05 0,397 ± 0,107
.LR 2 912,22 ± 53,97 226,57 ± 88,41 0,210 ± 0,119
PCh x LR 4 799,40 ± 32,64 380,89 ± 53,46 0,466 ± 0,072
#) Kleinste Quadrate Mittelwerte mit unterschiedlichen Buchstaben sind signifikant verschieden
(p ≤ 0,05).

Anhang I
342
Nach Geschlecht
30 kg Gruppe
Tab. A7: Volumen des Schinkenmagergewebes einschließlich Knochen (SMV) und des
subkutanen Fettes einschließlich Haut (SFETTV) sowie des gesamten Schinkens (SGV)
und das Fett-zu-Muskel-Verhältnis (SFMV) in einer 15 cm MRT-Region für männliche
Kastraten (M) und weibliche Schweine (W) bei ca. 30 kg Lebendmasse (LSM ± SEE#)
n SMV (cm3) SFETTV (cm3) SFMV SGV (cm3)
M 17 3629,75 ± 68,25 a 753,97 ± 27,25 0,208 ± 0,009 4383,72 ± 68,78 a
W 30 3805,11 ± 49,07 b 764,94 ± 20,01 0,204 ± 0,006 4570,05 ± 49,45 b
#) Kleinste Quadrate Mittelwerte mit unterschiedlichen Buchstaben sind signifikant verschieden
(p ≤ 0,05).
Tab. A8: Volumen der Mm. longissimi dorsi (MLDV) und des subkutanen Fettes
einschließlich Haut (FETTV) sowie das Fett-zu-Muskel-Verhältnis (FMV) in einer
10 cm MRT-Region für männliche Kastraten (M) und weibliche Schweine (W) bei ca.
30 kg Lebendmasse (LSM ± SEE#)
n MLDV (cm3) FETTV (cm3) FMV
M 17 307,25 ± 14,32 98,94 ± 6,93 0,328 ± 0,027
W 30 307,60 ± 10,64 99,93 ± 5,15 0,235 ± 0,020
#) Kleinste Quadrate Mittelwerte mit unterschiedlichen Buchstaben sind signifikant verschieden
(p ≤ 0,05).

Anhang I
343
60 kg Gruppe
Tab. A9: Volumen des Schinkenmagergewebes einschließlich Knochen (SMV) und des
subkutanen Fettes einschließlich Haut (SFETTV) sowie des Schinkengesamtvolumens
(SGV) und das Fett-zu-Muskel-Verhältnis (SFMV) in einer 15 cm MRT-Region für
männliche Kastraten (M) und weibliche Schweine (W) bei ca. 60 kg Lebendmasse
(LSM ± SEE#)
n SMV (cm3) SFETTV (cm3) SFMV SGV (cm3)
M 9 6795,33 ±140,46 1522,15 ± 59,92 0,227 ± 0,010 8317,48 ±143,82
W 22 6969,68 ±126,21 1464,06 ± 53.83 0,213 ± 0,009 8433,74 ±129,22
#) Kleinste Quadrate Mittelwerte mit unterschiedlichen Buchstaben sind signifikant verschieden
(p ≤ 0,05).
Tab. A10: Volumen der Mm. longissimi dorsi (MLDV) und des subkutanen Fettes
einschließlich Haut (FETTV) sowie das Fett-zu-Muskel-Verhältnis (FMV) in einer 10
cm MRT-Region für männliche Kastraten (M) und weibliche Schweine (W) bei ca. 60
kg Lebendmasse (LSM ± SEE#)
n MLDV (cm3) FETTV (cm3) FMV
M 9 574,60 ± 32,94 294,99 ± 22,19 0,551 ± 0,047
W 22 634,21 ± 29,59 275,41 ± 19,93 0,464 ± 0,043
#) Kleinste Quadrate Mittelwerte mit unterschiedlichen Buchstaben sind signifikant verschieden
(p ≤ 0,05).

Anhang I
344
90 kg Gruppe
Tab. A11: Volumen des Schinkenmagergewebes einschließlich Knochen (SMV) und
des subkutanen Fettes einschließlich Haut (SFETTV) sowie des
Schinkengesamtvolumens (SGV) und das Fett-zu-Muskel-Verhältnis (SFMV) in einer
15 cm MRT-Region für männliche Kastraten (M) und weibliche Schweine (W) bei ca.
90 kg Lebendmasse (LSM ± SEE#)
n SMV (cm3) SFETTV (cm3) SFMV SGV (cm3)
M 7 8663,04 ± 168,48 a 1910,58 ± 150,48 0,220 ± 0,018 10573,62 ± 212,19
W 10 9138,55 ± 128,81 b 1778,67 ± 115.05 0,196 ± 0,013 10917,21 ± 162,22
#) Kleinste Quadrate Mittelwerte mit unterschiedlichen Buchstaben sind signifikant verschieden
(p ≤ 0,05).
Tab. A12: Volumen der Mm. longissimi dorsi (MLDV) und des subkutanen Fettes
einschließlich Haut (FETTV) sowie das Fett-zu-Muskel-Verhältnis (FMV) in einer
10 cm MRT-Region für männliche Kastraten (M) und weibliche Schweine (W) bei ca.
90 kg Lebendmasse (LSM ± SEE#)
n MLDV (cm3) FETTV (cm3) FMV
M 7 756,48 ± 30,57 a 347,90 ± 50,07 0,454 ± 0,068
W 10 903,09 ± 23,37 b 317,64 ± 38,28 0,342 ± 0,052
#) Kleinste Quadrate Mittelwerte mit unterschiedlichen Buchstaben sind signifikant verschieden
(p ≤ 0,05).
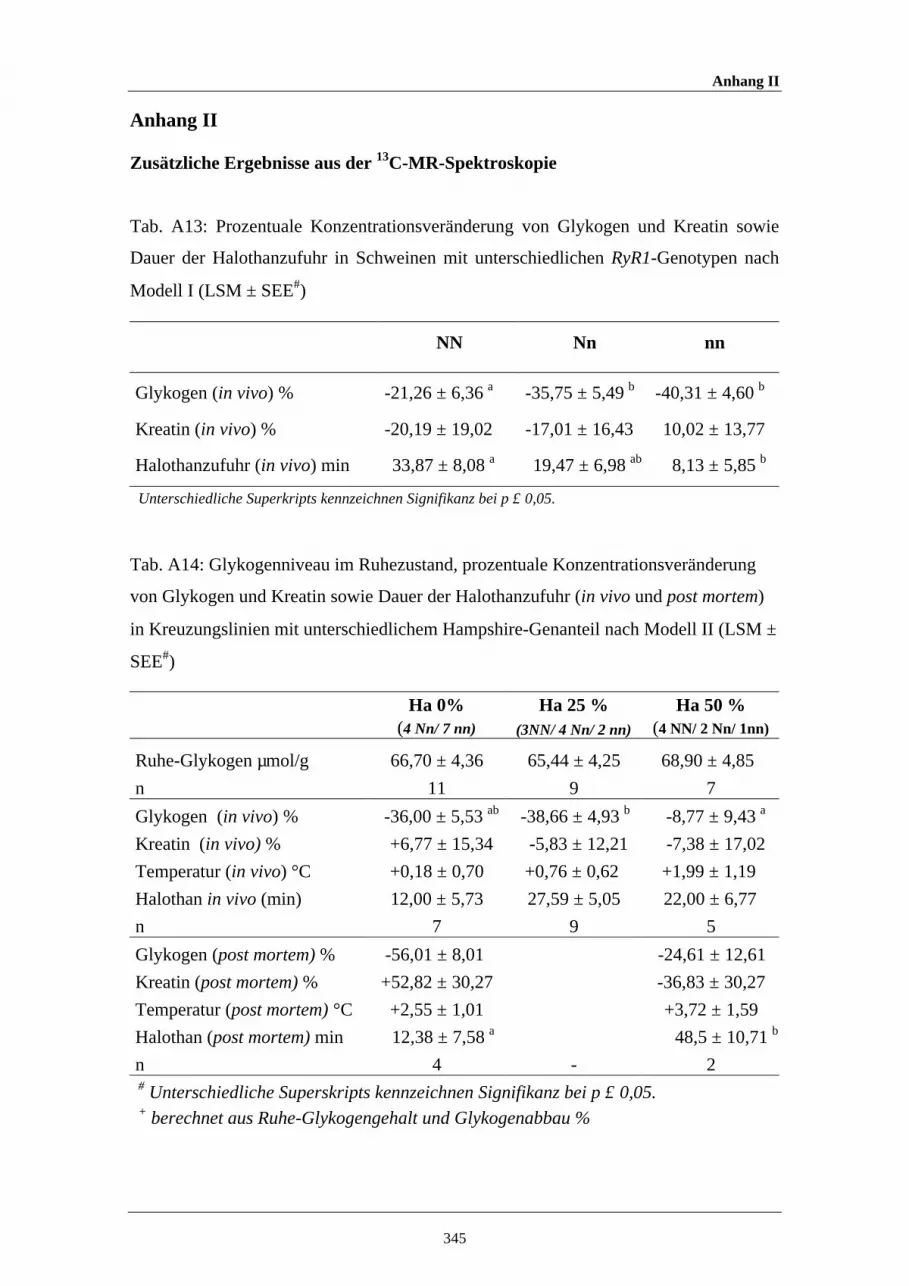
Anhang II
345
Anhang II
Zusätzliche Ergebnisse aus der 13C-MR-Spektroskopie
Tab. A13: Prozentuale Konzentrationsveränderung von Glykogen und Kreatin sowie
Dauer der Halothanzufuhr in Schweinen mit unterschiedlichen RyR1-Genotypen nach
Modell I (LSM ± SEE#)
NN Nn nn
Glykogen (in vivo) % -21,26 ± 6,36 a -35,75 ± 5,49 b -40,31 ± 4,60 b
Kreatin (in vivo) % -20,19 ± 19,02 -17,01 ± 16,43 10,02 ± 13,77
Halothanzufuhr (in vivo) min 33,87 ± 8,08 a 19,47 ± 6,98 ab 8,13 ± 5,85 b
Unterschiedliche Superkripts kennzeichnen Signifikanz bei p ≤ 0,05.
Tab. A14: Glykogenniveau im Ruhezustand, prozentuale Konzentrationsveränderung
von Glykogen und Kreatin sowie Dauer der Halothanzufuhr (in vivo und post mortem)
in Kreuzungslinien mit unterschiedlichem Hampshire-Genanteil nach Modell II (LSM ±
SEE#)
Ha 0% Ha 25 % Ha 50 %(4 Nn/ 7 nn) (3NN/ 4 Nn/ 2 nn) (4 NN/ 2 Nn/ 1nn)
Ruhe-Glykogen µmol/g 66,70 ± 4,36 65,44 ± 4,25 68,90 ± 4,85
n 11 9 7
Glykogen (in vivo) % -36,00 ± 5,53 ab -38,66 ± 4,93 b -8,77 ± 9,43 a
Kreatin (in vivo) % +6,77 ± 15,34 -5,83 ± 12,21 -7,38 ± 17,02
Temperatur (in vivo) °C +0,18 ± 0,70 +0,76 ± 0,62 +1,99 ± 1,19
Halothan in vivo (min) 12,00 ± 5,73 27,59 ± 5,05 22,00 ± 6,77
n 7 9 5
Glykogen (post mortem) % -56,01 ± 8,01 -24,61 ± 12,61
Kreatin (post mortem) % +52,82 ± 30,27 -36,83 ± 30,27
Temperatur (post mortem) °C +2,55 ± 1,01 +3,72 ± 1,59
Halothan (post mortem) min 12,38 ± 7,58 a 48,5 ± 10,71 b
n 4 - 2# Unterschiedliche Superskripts kennzeichnen Signifikanz bei p ≤ 0,05.+ berechnet aus Ruhe-Glykogengehalt und Glykogenabbau %

Anhang III
346
P C R Protocol for the MHS gene test (by Wiebke Scholz, 1994)
(PCR = polymerase chain reaction)
E x t r a c t i o n of DNA for PCR from whole blood
This procedure dissolves the cytoplasmic membrane and pellets the nuclei. Therefore, cytoplasmic DNA and RNA are lost.It should yield about 10 µg of DNA.
-Preserve the whole blood with EDTA.-Mix 0.25 ml blood with 0.25 ml lysis buffer in a 1.5 ml micro centrifuge tube.-Centrifuge 13,000ΧG for 20 sec.-Remove supernatant with pipette and resuspend pellet using vortex mixer in 1.0 ml lysis buffer (see above). Centrifuge 13,000XG for 20 sec.-Repeat previous step at least twice or until colorless.-Centrifuge 13,000ΧG for 20 sec., remove supernatant, and resuspend in 0.25 ml of "PCR buffer with nonionic detergents and Proteinase K"(see above).-Incubate at 50-60ºC in water bath for 1 hr (digest protein, DNA into solution).-Incubate at 95ºC for 10 min to inactivate the protease (kills Proteinase K).-Store frozen.
PCR - r e a c t i o n
-Example for 40 µl PCR-solution for 1 test :-Prepare a master mix of in order :
22 µl dH2O+ 4 µl 10x buffer (PCR 103) -- GeneAmp® PERKIN ELMER+ 4 µl 2 mM dNTP+ 1 µl 15 mM MgCl2+ 1 µl primer I (1:7) (BioServe -- Oligo Name: 17824)+ 1 µl primer II (1:7) (BioServe -- Oligo Name: 17825)
+ 1 µl Taq (AmpliTaq™ -- PERKIN ELMER) DNA polymerase (under ice)
-Work fast, mix all reagents with pipette.
-Add 6 µl DNA-suspension to master mix.-Mix carefully all reagents of PCR-solution.-Cover with 2 drops of mineral oil.-Place in PCR - machine (currency ca. 4½ hr).
-Program cycles (35x)

Anhang III
347
→Denature of the DNA strands to be matrix for primer and DNA polymerase at 94ºC (1 min)→Annealing of the primers on the complementary strands at 55ºC (1 min 15 sec)→Extension of primers by Taq DNA polymerase (DNA synthese) at 72ºC (1 min 15 sec)→1x 72°C (4 min)→1x 4°C indefinitely
T e s t i n g of PCR - reaction with electrophoresis
-Fill ethidium bromide containing 2% agarose gel in carrier of electrophoresis after placing of comb. Cooling is necessary until solidified!-Fill gel sub with 1x TAE (TBE) as buffer.-Pipet carefully through the oil to get 12 µl PCR product! Don't mix oil with PCR- product!-Mix 2 µl 10x loading buffer + 12 µl PCR-product. Fill in wells of gel.-Fill 10 µl molecular weight marker in an additional well for orientation.-Currency of electrophoresis is ca.60 min. by 90V.-Result: Expect a single 134 bp band visible under UV-light.
T y p i n g of DNA
Restriction enzyme Cfo1 (GIBCO BRL) cuts DNA-fragments in PCR suspension that canbe visualized on a 3% agarose ethidium bromide gel with UV-light.
-Example for "typing-solution" for 1 test:-20 µl PCR-product-Master mix under ice:
-2 µl Cfo1-buffer (for reaction of Cfo1 necessary)-2 µl Cfo1
-Incubate after mixing of all reagents in 37°C water bath immediately!-Digest over night( at least 2 hr).-Fill 3% agarose ethidium bromide gel in carrier of electrophoresis. Cooling is necessary until solidified.-Fill gel sub with 1x TAE (TBE) as buffer.-Mix 2 µl 10x loading buffer + 20 µl PCR-suspension. Fill in wells of gel.-Fill 10 µl molecular weight marker (Boehringer Mannheim Biochemica) in an
additional well for orientation.-Currency of electrophoresis is ca. 60 min. by 90V.-Result :
Because of the cutting of the DNA-fragments by Cfo1 you can find severalbands depending on the MHS-genotype (PSS) :

Anhang III
348
-homozygote positive (nn):1 band: 134 bp(base pairs)
-homozygote negative (NN):2 bands: 84 bp, 50 bp
-heterozygote (Nn):3 bands: 134 bp, 84 bp, 50 bp
R e a g e n t s
For prevention of coagulation of fresh whole blood
EDTA = ethylenediaminetetraacetic acid
For extraction of DNA for PCR from whole blood
lysis buffer 5.5 g 1M sucrose+ 5 ml 100mM Tris-HCl
(trizma hydrochloride)+ 250 µl 1M MgCl2+ 0.5 ml Triton X-100___________________________add sterile H20 up to 50 ml
"PCR 101-buffer with nonionic detergent and Proteinase K"-PCR 101-buffer with nonionic detergent :
0.125 ml 1M MgCl2(20.3 g into 100 ml dH2O = 1M)
+ 2.5 ml 1M KCl(7.45 g into 100 ml dH2O = 1M)
+ 0.5 ml 1M Tris-HCl(15.76 g into 100 ml dH2O = 1M)
+ 0.25 ml of 20 mg/ml gelatine(0.1 g in 5 ml dH2O)
+ 0.225 ml Nonidet P40 (0.45%)+ 0.225 ml Tween 20 (0.45%)_______________________add sterile H2O up to 50 ml
-Autoclave and store frozen.-Add 0.6 µl of 10mg/ml Proteinase K per 100 µl solution after thawing.

Anhang III
349
For PCR-reaction
10x buffer (PCR 103) - without MgCl2-Provides pH and ionic strength for PCR amplification reaction.-100 mM Tris-HCl (pH 8.3); 500 mM KCl
dNTP (deoxynucleoside triphosphates) :-These enzymes split the DNA in oligonucleotide.-dNTP consists of :
-dATP (2 deoxyadenosine 5 triphosphate)-dCTP (2 deoxycytidine 5 triphosphate)-dGTP (2 deoxyguanosine 5 triphosphate)-dTTP (2 deoxythymidine 5 triphosphate)
-Solution for PCR : 250 µl 2 mM dNTP's : 50 µl 10mM dATP+50 µl 10mM dCTP+50 µl 10mM dGTP+50 µl 10mM dTTP+50 µl dH2O
MgCl2 (magnesium chloride)-Required to achieve optimal PCR amplification is dependent on the specific set of primers and template used / Supposition for function of Taq DNA polymearse (coenzyme)
Solution for PCR : 15 mM MgCl2 (1:3) →1 µl 45 nM MgCl2 + 2 µl dH2O (Storing in freezer!)
Primer-For start of polymerase reaction-Primers anneal on both sides of single DNA strand.-Concentration of the primers for PCR :20 pmol after dilution.
1:7 (1 µl primer + 6 µl dH2O)primer I (sequence number:17824) :
5'> GTG CTG GAT GTC CTG TGT TCC CT <3'primer II (sequence number:17825) :
5'> CTG GTG ACA TAG TTG ATG AGG TTT G <3'
Taq DNA polymerase-Concentration for PCR : 5 U/µl (1 unit characterizes synthesized DNA quantity under normal conditions.)
Mineral oil-On top of the PCR reaction to prevent evaporation.

Anhang III
350
For testing of PCR-reaction with electrophoresis
2% agarose ETBr 1.6 g agarose
(MetaphorTM agarose orMolecular Biology certified agarose Ultra Pur DNA Grade Agarose)
+1.6 ml 50x TAE 121 g tris base(TRIZMA Base+28.55 ml glacial acidic acid+50 ml 0.5 M EDTA(pH 8.0)
(or 13.75 g EDTA)__________________________add sterile H2O up to 500 ml
+80 ml dH2O+4 µl ETBr (ethidium bromide)
→ U s e g l o v e s !!!
-Heat prepared agarose gel carefully until viscous and in solution.-Pour gel in prepared gel carrier.-Remove comb (wells for probes) after gel is solidified.
1x TAE(TBE) as buffer : 10ml of 50xTAE+490ml dH2O
10x gel loading buffer 0.2g Ficoll 400 (20%)+0.2ml 0.5M Na2EDTA (pH8)+0.1ml SDS (Sodium dodecyl sulfate) (10%)+0.25ml Xylene Brilliant (1%)
(or 0.01g in 1ml dH2O)+0.45ml dH2O
DNA molecular weight marker VIII-Solution in 10mmol/l Tris-HCl, 1mmol/l EDTA(pH8)-Fragment mixture prepared by cleavage of pUCBM21 DNA with restriction endonuclease Hpa II and Dra I plus Hind III.-The mixture contains DNA-fragments with the following base pair (bp) lengths (1bp = 660 daltons) : 1114,900,692,501,489,404,320,242,190,147,124,110,67,37,34(2x),26,19.-The DNA fragment mixture shows the typical pattern with 13 bands in agarose gel electrophoresis.-Use 10µl of following mixture per well :
40µl marker+20µl 10x loading buffer+140µl dH2O

Anhang III
351
For typing of DNA
Cfo1-Cfo1 is isolated from Clostridium formicoaceticum.-1 unit is the amount of enzyme required to cleave 1µg of λ DNA in 1 hr at 37°C in appropriate buffer.
Storage buffer (Cfo1-buffer)50mM Tris-HCl (pH 7.4)100mM NaCl6mM 2-mercaptoethanol0.1mM PMSF500µg/ml BSA50%(v/v) glycerol
3% agarose ETBr gel 2.4g agarose+1.6ml 50x TAE+80ml dH2O+4µl ETBr
-Look under 2% agarose ETBr gel !
1x TAE as buffer → See above !10x gel loading buffer → See above !marker → See above !

Anhang IV
352
Anhang IV
Konferenzbeiträge 1. Konferenz der SAS-Benutzer in Forschung und Entwicklung(KSFE), Humboldt-Universität zu Berlin, 20./21. Februar 1997, Herausgeber:
Rechenzentrum der Humboldt-Universität zu Berlin (S. 295 – 308)
Videobildanalyse zur Bestimmung des Marmorierungsgrades im Kotelettmit Hilfe verschiedener SAS-Prozeduren
Armin Scholz
http://www.hu-berlin.de/rz/ksfe/inhalt.html [ftp://ftp.rz.hu-berlin.de/pub/docs/ksfe/FR_A1130.EXE]
Einleitung
Ein wichtiges Kriterium für den Genusswert des Schweinefleisches, den Geschmack und
die Beschaffenheit stellt das intramuskuläre Fett bzw. die Marmorierung im Kotelettmuskel
(M. longissimus dorsi) dar. In der praktischen Schweinezucht wurde aufgrund der
Preisgestaltung für Schlachtschweine der Hauptselektionserfolg im Magerfleischanteil
erzielt. Infolgedessen weist der intramuskuläre Fettgehalt (IMF) im Kotelettmuskel der in
Deutschland hauptsächlich zur Fleischerzeugung eingesetzten Schweinerassen bzw.
Kreuzungen bei einer Variation von durchschnittlich 0,6 - 1,8 % ein sehr niedriges Niveau
auf. Die einzige Ausnahme bildet die Rasse Duroc mit einem IMF von ca. 2,5%. Das
Optimum für den intramuskulären Fettgehalt wird jedoch bei 1,5 - 2,5 % angegeben.
Ein weiteres Problem liegt in der ungenauen Bestimmung des intramuskulären
Fettgehaltes bzw. des Marmorierungsgrades. Gegenwärtig wird in den deutschen Schweine-
Leistungsprüfanstalten nur eine subjektive Bonitierung der Marmorierung (Noten 1 - 6) am
Kotelettanschnitt zwischen 13. und 14. Brustwirbel vorgenommen.
Eine Umkehr bei der Entwicklung des intramuskulären Fettgehaltes ist erst durch
Einführung einer objektiven Leistungsprüfung und die Einbeziehung dieses Merkmals in
die Selektion zu erwarten. Zur Bestimmung des intramuskulären Fettgehaltes am
Schlachtkörper bzw. am lebenden Tier werden verschiedene chemische und physikalische
Methoden genutzt, wie z. B. die Soxhlet-Extraktion, Ultraschall-Technik,
Reflexionsmessungen, Computer-Tomographie, 1H-NMR- bzw. Infrarot-Spektroskopie.

Anhang IV
353
Da in einigen Prüfstationen die Videobildanalyse bereits zur Ermittlung der Fett- und
Fleischfläche am Kotelettanschnitt eingesetzt wird, war es naheliegend, diese Technik
ebenfalls für die Ermittlung des Marmorierungsgrades zu nutzen.
Material und Methode
Für die Untersuchung wurden 130 Koteletts (13./14. Brustwirbel) von Schweinen
verschiedener Genotypen aus der Leistungsprüfanstalt Ruhlsdorf genutzt. Die Koteletts
wurden mit einer 3 Chip R G B - Color - Kamera (Hitachi - HV-C10) beidseitig cranial
bzw. caudal aufgenommen und über drei Analog - Digital - Wandler (ITI - Color Frame
Grabber) auf einer wiederbeschreibbaren Magneto-Optischen-Diskette (600 MB)
gespeichert. Ein Bild umfasst 768*512 Bildpunkte bei einer Datentiefe von 24 bit, die auf
die drei Farbkanäle ROT, GRÜN und BLAU (R G B) aufgeteilt sind. Für jeden Bildpunkt,
der eine Fläche von ca. 0,05 mm2 einnimmt, liegen folglich drei Grauwertinformationen
jeweils im Bereich von 0 - 255 vor.
Videobild eines Koteletts mit 3-Chip-RGB-Kamera
(768*512 Pixel)

Anhang IV
354
Mit Hilfe des Bildverarbeitungsprogrammes BioScan-OPTIMAS wurden die
interessierenden Kotelettregionen, sogenannte Regions Of Interest (ROI's), halbautomatisch
eingegrenzt, maskiert und als neue TIF-Datei gespeichert.
Maskieren (Ausschneiden) der Region of Interest (ROI)
Die Kantendetektion beruht dabei auf den Grauwertunterschieden zwischen Fleisch- und
Fettgewebe. Niedrige (dunkle) Grauwerte charakterisieren Fleischgewebe und hohe
Grauwerte kennzeichnen Fettgewebe.

Anhang IV
355
Ergebnis der Maskierung
Die Weiterverarbeitung der TIF-Bilddatei erfolgte mit SAS 6.11. Anhand einer Beispiel-
TIF-Datei (f26lmsk.tif) wird der Ablauf nachfolgend näher beschrieben.
/* 1. Schritt: Bildinformationen filtern - Bildaufbau in Tags, Stripes und Scoreskodiert */
%let file=d:\mp120193\f26lmsk.tif;
libname f26lmsk "d:\asda";filename tif "&file";title "&file";data tags (keep=tag text type typ length score comment) stripes (keep=stripa stripe stripl) scores (keep=subfile xpixels ypixels bits compress photo nrows minval maxval xres yres planar resunit nstrips); length text comment modus $32; length default=4; retain ifd ntags (2 0); infile tif recfm=n; /* ermöglicht das Importieren von beliebigen internen Binär - Daten */ /* header */ input fa $char2. vers pib2.; if fa='II' then firma='Intel '; if fa='MM' then firma='Motorola';
a: iifd+1; iifd+1; input @(ifd+12*ntags+3) ifd pib4.; if ifd=0 then stop; /* ifd */ input @(ifd+1) ntags pib2.; do itag=1 to ntags; /* tag */ comment=''; input @(ifd+12*itag-9) tag pib2. type pib2. length pib4. score pib4.; select (tag); when (254) text='NewSubfile';

Anhang IV
356
when (255) do; text='SubfileType'; select (score); when (1) modus='FullResolution'; when (2) modus='ReducedResolution'; when (3) modus='SinglePageImage'; otherwise modus='unknown'; end; comment=modus; subfile=score; end; when (256) do; /* Breite */ text='ImageWidth'; comment='Xpixels'; xpixels=score; end; when (257) do; /* Höhe */ text='ImageLength'; comment='Ypixels'; ypixels=score; end; when (258) do; text='BitsPerSample'; bits=score; end; when (259) do; text='Compression'; select (score); when (1) modus='Uncompressed'; when (2) modus='Huffman RLE'; when (3) modus='FAX CCITT Group3'; when (4) modus='FAX CCITT Group4'; when (5) modus='LZW'; when (6) modus='JPEG'; otherwise modus='unknown'; end; comment=modus; compress=score; end; when (262) do; text='PhotometricInterpretation'; select (score); when (0) modus='MinSampleValue=White'; when (1) modus='MaxSampleValue=White'; when (2) modus='RGB'; when (3) modus='PaletteColor'; when (4) modus='TransparencyMask'; when (6) modus='YCbCrColorSpace'; when (8) modus='1976CIEL*a*b ColorSystem'; otherwise modus='unknown'; end; comment=modus; photo=score; end; when (263) text='Tresholding'; when (264) text='CellWidth';

Anhang IV
357
when (265) text='CellLength'; when (266) text='FillOrderTag'; when (269) text='DocumentName'; when (270) text='ImageDescription'; when (271) text='ScannerMake'; when (272) text='ScannerModel'; when (273) do; text='StripOffsets'; nstrips=length; kstrips=score; end; when (274) text='Orientation'; when (277) text='SamplesPerPixel'; when (278) do; text='RowsPerStrip'; nrows=score; end; when (279) do; text='StripByteCounts'; lstrips=score; end; when (280) do; text='MinSampleValue'; minval=score; end; when (281) do; text='MaxSampleValue'; maxval=score; end; when (282) do; text='XResolution'; input @(score+1) z pib4. n pib4.; xres=z/n; comment=left(put(xres,best12.3)); end; when (283) do; text='YResolution'; input @(score+1) z pib4. n pib4.; yres=z/n; comment=left(put(yres,best12.3)); end; when (284) do; text='PlanarConfiguration'; planar=score; select (score); when (1) modus='NoPlaneSplitting'; when (2) modus='PlaneSplitting'; otherwise modus='unknown'; end; comment=modus; end; when (285) text='PageName'; when (286) text='XPosition'; when (287) text='YPosition'; when (288) text='FreeOffsets'; when (289) text='FreeByteCounts';

Anhang IV
358
when (290) text='GrayResponseUnit'; when (291) text='GrayResponseCurve'; when (292) text='T4Options'; when (293) text='T6Options'; when (296) do; text='ResolutionUnit'; resunit=score; select (score); when (1) unit='Aspect '; when (2) unit='Inch '; when (3) unit='Cm '; otherwise unit='unknown'; end; comment=unit; end; when (297) text='PageNumber'; when (300) text='ColorResponseUnit'; when (301) text='TransferFunction'; when (305) text='Software'; /* Ersteller Software */ when (306) text='DateTime'; /* Erstellung am um */ when (315) text='Artist'; /* Macher Name */ when (316) text='HostComputer'; /* Macher Computer */ when (317) text='Predictor'; /* nur bei LWZ-Compression=5 */ when (318) text='WhitePoint'; when (319) text='PrimaryCromaticities'; when (320) text='ColorMap'; when (321) text='HalftoneHints'; when (322) text='TileWidth'; when (323) text='TileLength'; when (324) text='TileOffsets'; when (325) text='TileByteCount'; when (332) text='InkSet'; when (333) text='InkNames'; when (334) text='NumberOfInks'; when (336) text='DotRange'; when (337) text='TargetPrinter'; when (338) text='ExtrasSamples'; when (339) text='SampleFormat'; when (340) text='SMinSampleValue'; when (340) text='SMinSampleValue'; when (341) text='SMaxSampleValue'; when (342) text='TransferRange'; when (512) text='JPEGProc'; when (513) text='JPEGInterchangeFormat'; when (514) text='JPEGInterchangeFormatLength'; when (515) text='JPEGRestartInterval'; when (517) text='JPEGLossLessPredictors'; when (518) text='JPEGPointTransforms'; when (519) text='JPEGQTables'; when (520) text='JPEGDCTables'; when (521) text='JPEGACTables'; when (529) text='YCbCrCoefficient'; when (530) text='YCbCrSubSampling'; when (531) text='YCbCrPositioning'; when (532) text='ReferenceBlackWhite'; otherwise text='UnknownTag';

Anhang IV
359
end; select (type); when ( 1) typ='Byte '; /* 8 Bit-Byte */ when ( 2) typ='ASCII '; /* 8 Bit-ASCII-Code */ when ( 3) typ='SHORT '; /* 16 Bit unsigned Integer */ when ( 4) typ='LONG '; /* 32 Bit unsigned Integer */ when ( 5) typ='RATIONAL '; /* 2 LONGs: Fraction/Denominator */ when ( 6) typ='SByte '; /* 8 Bit signed Integer */ when ( 7) typ='UNDEFINED'; /* 8 Bit Anything */ when ( 8) typ='SSHORT '; /* 16 Bit signed Integer */ when ( 9) typ='SLONG '; /* 32 Bit signed Integer */ when (10) typ='RATIONAL '; /* 2 SLONGs: Fraction/Denominator */ when (11) typ='FLOAT '; /* 4 Byte IEEE floating point */ when (12) typ='DOUBLED '; /* 8 Byte IEEE floating point */ otherwise typ='Unknown '; end; output tags; end; do istrip=1 to nstrips; input @(4*istrip+kstrips-3) stripa pib4. @(4*istrip+lstrips-3) stripl pib4.; stripa=stripa+1; stripe=stripa+stripl-1; output stripes; end; output scores; goto a;run;
WORK.SCORES -- Auszug C X Y O N R P P M S E I I P P T N S X X B R H R R X Y U E E I E O I O R R N L L T S T P W E E I S S S S O S S S S T
465 320 446606 1 2 30 11 299 288 2

Anhang IV
360
WORK.TAGS
OBS TEXT COMMENT TAG TYPE LENGTH SCORESCORE TYP
1 NewSubfile 254 4 1 0 LONG 2 ImageWidth Xpixels 256256 4 1 465465 LONG 3 ImageLength Ypixels 257257 4 1 320320 LONG 4 BitsPerSample 258258 3 3 446606 SHORT 5 Compression Uncompressed 259259 3 1 11 SHORT 6 PhotometricInterpretation R G B R G B 262262 3 1 22 SHORT 7 ScannerModel 272 2 10 446612 ASCII 8 StripOffsets 273 4 30 446622 LONG 9 SamplesPerPixel 277277 3 1 33 SHORT10 RowsPerStrip 278278 4 1 1111 LONG11 StripByteCounts 279 4 30 446742 LONG12 XResolution 299299 282282 5 1 446862 RATIONAL13 YResolution 288288 283283 5 1 446870 RATIONAL
14 ResolutionUnit Inch 296 3 1 2 SHORT15 Software 305 2 25 446878 ASCII16 DateTime 306 2 26 446903 ASCII
WORK.TAGS
OBS STRIPA STRIPL STRIPE
1 9 15345 15353 2 15354 15345 30698 3 30699 15345 46043 4 46044 15345 61388 5 61389 15345 76733 6 76734 15345 92078 7 92079 15345 107423 8 107424 15345 122768 9 122769 15345 138113 10 138114 15345 153458 11 153459 15345 168803 12 168804 15345 184148 13 184149 15345 199493 14 199494 15345 214838 15 214839 15345 230183
OBS STRIPA STRIPL STRIPE
16 230184 15345 245528 17 245529 15345 260873 18 260874 15345 276218 19 276219 15345 291563 20 291564 15345 306908 21 306909 15345 322253 22 322254 15345 337598 23 337599 15345 352943 24 352944 15345 368288 25 368289 15345 383633 26 383634 15345 398978 27 398979 15345 414323 28 414324 15345 429668 29 429669 15345 445013 30 445014 1395 446408
WORK.STRIPES
/* DATA STEP zur Erzeugung der SAS-Datei (.RGB) */
data f26lmsk.rgb (keep=x y r g b); /* PhotometricInterpretation=RGB */ length default=3; infile tif recfm=n; set scores; y=ypixels+1; do i=1 to nstrips;

Anhang IV
361
set stripes; k=stripa-3; do j=1 to nrows; do y=y-1 while(y>0); do x=1 to xpixels; k+3; input @k R pib1. G pib1. B pib1.; if R+G+B then output; end; end; end; end;run;
OBS Y X R G B
87783 76 379 128 107 117
87784 76 380 121 103 108
87785 76 381 125 112 114
87786 76 382 140 133 137
87787 75 64 148 139 154
87788 75 65 145 139 149
F26LMSK.RGB -- Auszug
/* Erzeugung einer SEED-Datei für die anschließende Clusteranalyse */
libname scholz "d:\mp120193";
DATA SCHOLZ.SEED; INPUT R G B CLUSTER; CARDS; 165 150 155 1 155 135 140 2 145 125 135 3 135 115 120 4 125 110 115 5 115 100 105 6 105 95 100 7 ;RUN;
/* Procedur Clusteranalyse */
PROC FASTCLUS DATA = f26lmsk.RGB (WHERE=(R+G+B)) SEED =SCHOLZ.SEED MAXC =7 MAXITER =99 DRIFT REPLACE =PART CONVERGE=0.001 OUT =f26lmsk.CLUST; VAR R G B;RUN;

Anhang IV
362
OBS Y X R G B CLUSTER DISTANCE
102523 3 303 130 107 115 6 4.8076 102524 3 304 132 110 117 5 6.4517 102525 3 305 130 110 117 5 7.4931 102526 3 306 132 114 118 4 3.8475 102527 3 307 139 122 126 4 3.2864 102528 3 308 142 124 133 4 4.7908 102529 3 310 137 120 128 4 4.6087 102530 3 311 134 116 123 5 2.7512 102531 3 312 133 113 115 5 6.1770 102532 3 313 129 110 111 6 4.8802 102533 3 314 143 126 130 4 4.4330 102534 3 315 148 132 135 3 2.5921
F26LMSK.CLUST -- Auszug
/* Procedur Diskriminanzanalyse */
PROC DISCRIM DATA =f26lmsk.CLUST METHOD=NORMAL POOL =YES OUT =f26lmsk.DISKRI; CLASS CLUSTER; VAR R G B;RUN ;
OBS OBS Y X R G B CLUSTER DISTANCE _1 ... _7 _INTO_ Y X R G B CLUSTER DISTANCE _1 ... _7 _INTO_
102307 11 308 155 151 153 22 14.1864 0.54274 0.0000000 11
102308 11 309 147 136 146 2 7.3491 0.00000 0.0000000 2
102309 11 310 137 118 130 4 5.9974 0.00000 0.0000000 4
102310 11 311 131 111 119 5 5.3652 0.00000 0.0000000 5
102311 11 312 129 107 114 6 3.5556 0.00000 0.0000076 6
F26LMSK.DISKRI -- AUSZUG

Anhang IV
363
/* ERZEUGUNG des Clusterbildes */
libname scholz "d:\mp120193";title "create the ANNOTATE - scholz02s.sas";
data anno; attrib x y length=4; length default=4; array farbe (0:15) $8 _temporary_ ('gray' 'white' 'blue' 'green' 'cyan' 'yellow' 'magenta' 'orange' 'red' 'brown' 'lime' 'gold' 'tan' 'purple' 'pink' 'steel'); retain function 'symbol'; retain text 'dot'; retain xsys ysys '2'; retain size .023; set f26lmsk.DISKRI (where=(_into_)); color=farbe(mod(_into_,16)); keep function color text xsys ysys size x y;run;
OBS X Y FUNCTION TEXT XSYS YSYS SIZE COLOR 87501 97 76 symbol dot 2 2 0.023000 white 87502 98 76 symbol dot 2 2 0.023000 blue 87503 99 76 symbol dot 2 2 0.023000 red 87504 100 76 symbol dot 2 2 0.023000 green 87505 101 76 symbol dot 2 2 0.023000 green 87506 102 76 symbol dot 2 2 0.023000 blue
WORK.ANNO -- Auszug
goptions reset=all dev=win;goptions colors=('gray' 'white' 'blue' 'green' 'cyan' 'yellow' 'magenta' 'orange' 'red' 'brown' 'lime' 'gold' 'tan' 'purple' 'pink' 'steel') cback=black;proc ganno anno=_last_ datasys;run;

Anhang IV
364
7-Cluster-Bildweiß charakterisiert intramuskuläres Fett
Ergebnisse aus der Cluster- bzw. Diskriminanzanalyse
Cluster Means
Cluster R G B
1 163.980 155.374 163.413 2 153.553 138.978 145.278 3 146.959 129.780 136.092 4 140.810 122.177 128.498 5 134.524 114.412 120.608 6 127.470 105.206 111.080 7 118.608 92.853 97.597
Cluster Standard Deviations
Cluster R G B
1 6.00447 8.18150 8.94079 2 3.53037 3.86297 4.31256 3 3.05880 3.00326 3.33203 4 2.95175 2.84208 3.23728 5 3.05158 3.03086 3.45572 6 3.33078 3.56051 4.08508 7 4.45324 5.25349 5.79337
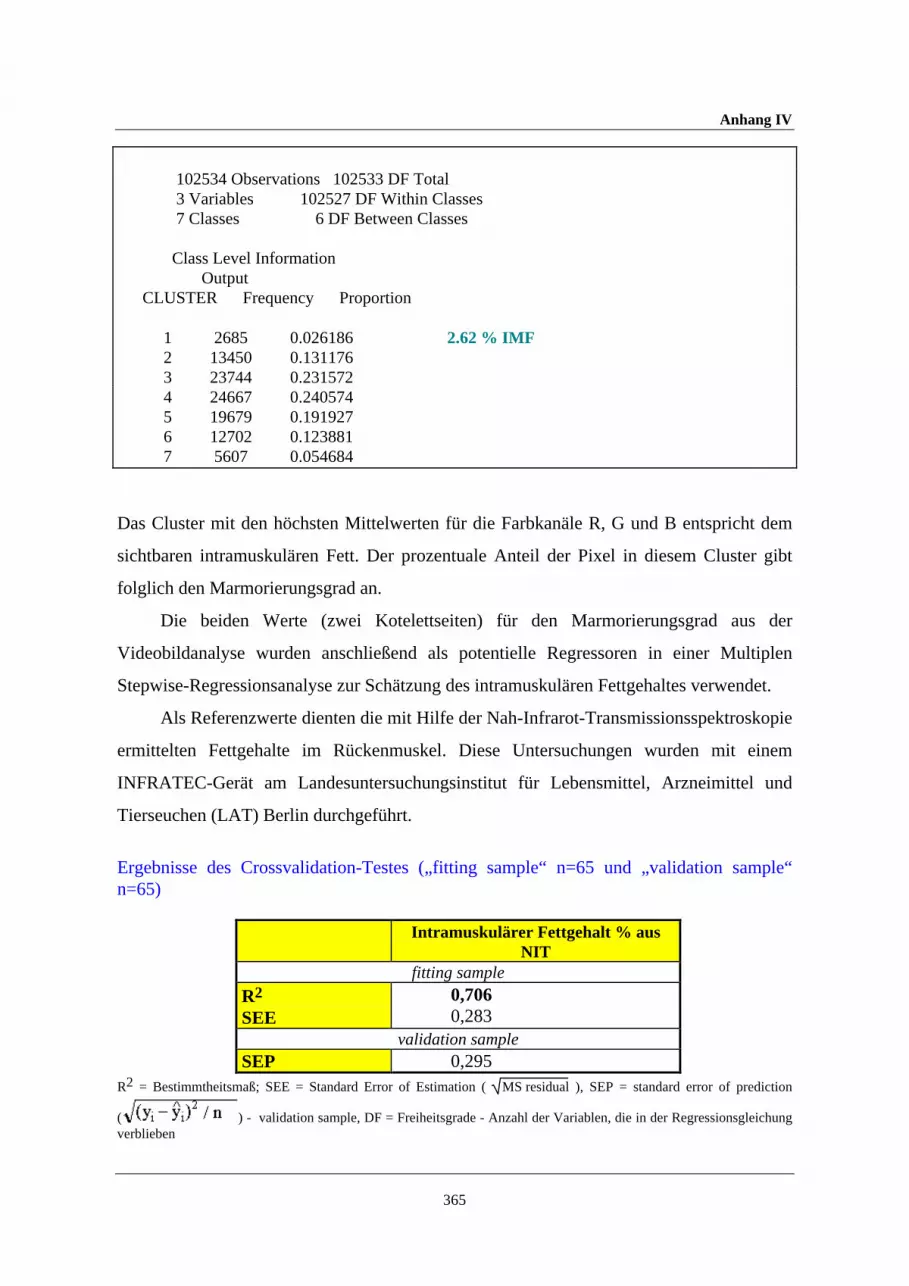
Anhang IV
365
102534 Observations 102533 DF Total 3 Variables 102527 DF Within Classes 7 Classes 6 DF Between Classes
Class Level Information Output CLUSTER Frequency Proportion
1 2685 0.026186 2.62 % IMF 2 13450 0.131176 3 23744 0.231572 4 24667 0.240574 5 19679 0.191927 6 12702 0.123881 7 5607 0.054684
Das Cluster mit den höchsten Mittelwerten für die Farbkanäle R, G und B entspricht dem
sichtbaren intramuskulären Fett. Der prozentuale Anteil der Pixel in diesem Cluster gibt
folglich den Marmorierungsgrad an.
Die beiden Werte (zwei Kotelettseiten) für den Marmorierungsgrad aus der
Videobildanalyse wurden anschließend als potentielle Regressoren in einer Multiplen
Stepwise-Regressionsanalyse zur Schätzung des intramuskulären Fettgehaltes verwendet.
Als Referenzwerte dienten die mit Hilfe der Nah-Infrarot-Transmissionsspektroskopie
ermittelten Fettgehalte im Rückenmuskel. Diese Untersuchungen wurden mit einem
INFRATEC-Gerät am Landesuntersuchungsinstitut für Lebensmittel, Arzneimittel und
Tierseuchen (LAT) Berlin durchgeführt.
Ergebnisse des Crossvalidation-Testes („fitting sample“ n=65 und „validation sample“n=65)
Intramuskulärer Fettgehalt % ausNIT
fitting sample
R2
SEE0,7060,283
validation sampleSEP 0,295
R2 = Bestimmtheitsmaß; SEE = Standard Error of Estimation ( MS residual ), SEP = standard error of prediction
( ) - validation sample, DF = Freiheitsgrade - Anzahl der Variablen, die in der Regressionsgleichungverblieben

Anhang IV
366
SchlußfolgerungenDie Videobildauswertung mit Hilfe der Cluster- bzw. Diskriminanzanalyse eröffnet dieMöglichkeit, den Marmorierungsgrad im Kotelettmuskel beim Schwein oder anderenTierarten relativ einfach, schnell und objektiv zu bestimmen. Voraussetzungen für eineerfolgreiche Verwendung dieser Technik sind die Einhaltung standardisierter Bedingungenbei der Bildaufnahme unter weitgehender Vermeidung von Oberflächenreflexionen. Essollte eine möglichst hochauflösende Digitalkamera verwendet werden.
Literatur:
Referenzhandbuch DateiformateGünter BornAdison Wesley, 4. Aufl. 1996, 557 - 592
Johnson, B. and W. Johnson (1992): Importing X-WindowSystem Raster Images into SAS-Graph / Software Output.SUGI 17, Hawai, 543 – 548
Herzlicher Dank gebührt Herrn W. Lesener (Rechenzentrum, HU Berlin).

Anhang V
367
Anhang V
Quantitative Markergenorte für den Körperfettgehalt
Tab. A15: Auswahl potentieller „Quantitativer Markergenorte“ (QTL) für denKörperfettgehalt bei Schwein, Maus und Mensch (eigene modifizierte Tabelle ausMitchell et al., 2001)
Quantitative TraitLoci (Major- oderKandidatengene,Microsatelliten)
Chromosomenlokalisation(Referenz)
beeinflusste Merkmale,Rezeptoren,Metaboliten
Schwein Maus Menschmehrere Marker 1
(34, 47, 48,50)
Körperfettgehalt,Rückenspeckdicke,Wachstum
Melanocortin-4-Rezeptor (MC4R)
1q22-q27(34, 75, 76)
- 18q22(70, 76)
Körperfettgehalt,Rückenspeckdicke
„Insulin-like growthfactor 1“-Rezeptor(IGF1R)
1q17-q22(34, 77)
7(70)
15q25-q26(70)
Körperfettgehalt,Rückenspeckdicke,Wachstum
Proopiomelanocortin(POMC)
3q25(34, 81)
12(70)
2p23.3(70, 81)
Serumleptinniveau –Fettmasse,Energieregulation,Pigmentierung
Glucokinaseregulatory protein(GKRP)
2p23.3-p23.2(8, 70)
Glukose-Homöostase,Insulin-Sensitivität
S0175, verschiedeneMarker, z.B.abdominal fatdeposition (AFD);total growth (TGR);
4(6, 7, 34, 47,48)
subkutane Fettdickeund Abdominalfett-anteil,Körperfettverteilung
Adipocyte-specificfatty acid-bindingprotein (A-FABP,FABP4, H-ALBP)
4(34, 79)
8q12(70)
subkutane Fettdickeund Abdominalfett-anteil,Körperfettverteilung
Ryanodin-Rezeptor-1(RyR1 -- Majorgen)
6q11-q12(34, 35, 47)
7a2-b3(34, 37)
19q13.1(34,36)
MalignesHyperthermieSyndrom,Magerfleischigkeit
Glukosephosphat-isomerase (GPI)
6q12(34, 69)
7(70)
19q13.1(70)
GPI katalysiert diewechselseitige Um-wandlung vonGlukose-6-Phosphatin Fruktose-6-Phosphat
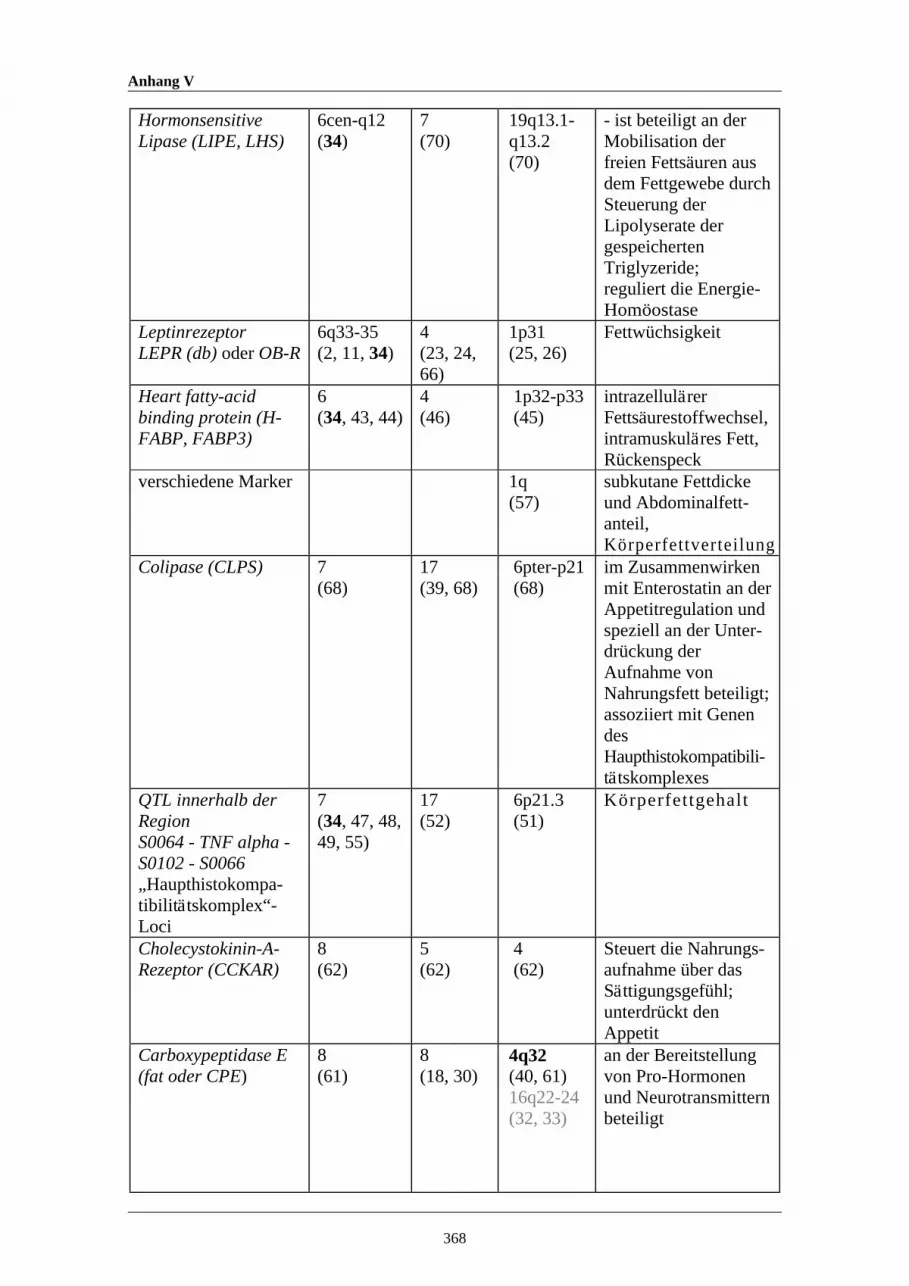
Anhang V
368
HormonsensitiveLipase (LIPE, LHS)
6cen-q12(34)
7(70)
19q13.1-q13.2(70)
- ist beteiligt an derMobilisation derfreien Fettsäuren ausdem Fettgewebe durchSteuerung derLipolyserate dergespeichertenTriglyzeride;reguliert die Energie-Homöostase
LeptinrezeptorLEPR (db) oder OB-R
6q33-35(2, 11, 34)
4(23, 24,66)
1p31(25, 26)
Fettwüchsigkeit
Heart fatty-acidbinding protein (H-FABP, FABP3)
6(34, 43, 44)
4(46)
1p32-p33 (45)
intrazellulärerFettsäurestoffwechsel,intramuskuläres Fett,Rückenspeck
verschiedene Marker 1q(57)
subkutane Fettdickeund Abdominalfett-anteil,Körperfettverteilung
Colipase (CLPS) 7(68)
17(39, 68)
6pter-p21 (68)
im Zusammenwirkenmit Enterostatin an derAppetitregulation undspeziell an der Unter-drückung derAufnahme vonNahrungsfett beteiligt;assoziiert mit GenendesHaupthistokompatibili-tätskomplexes
QTL innerhalb derRegionS0064 - TNF alpha -S0102 - S0066„Haupthistokompa-tibilitätskomplex“-Loci
7(34, 47, 48,49, 55)
17(52)
6p21.3 (51)
Körperfettgehalt
Cholecystokinin-A-Rezeptor (CCKAR)
8(62)
5(62)
4 (62)
Steuert die Nahrungs-aufnahme über dasSättigungsgefühl;unterdrückt denAppetit
Carboxypeptidase E(fat oder CPE)
8(61)
8(18, 30)
4q32(40, 61)16q22-24(32, 33)
an der Bereitstellungvon Pro-Hormonenund Neurotransmitternbeteiligt

Anhang V
369
NEUROPEPTID YREZEPTOR 5 (NPY5R)
8p11(34, 67)
8(70)
4q31-q32 (70)
Das Neuropeptid Ystimuliert den Appetit;schränkt den Energie-verbrauch ein undsteigert dieLipogenese. DieRezeptoren NPY5Rund NPY1R befindensich in derselbenChromosomenregion.
Mitochondrialuncoupling protein(UCP2)
9p21-p24(34, 78)
7 (31) 11 (31) Körpergewicht,Energiebalance
Tubby (TUB) 9 ? 7(19, 28,30, 41,66)
11p15.4(29)17p15(28)16q#) (21)2p#) (22)
Körpergröße,Fettansatz,#) beim MenschenBezug zum Bardet-Biedl- und AlstroemSyndrom)
Myogenin (MYOG,MYF4)
9q21-q26(34, 73)
1(70)
1q31-q41(70)
MuskelspezifischerTranskriptionsfaktorreguliertMuskelfaserzahl,Muskelmasse.
Growth Hormone 1(GH1)
12p14(34)
11(70)
17q22-q24(70)
Wachstum
Pituitary-specifictranscription factors(PIT-1/GHF-1)
13(34, 48, 54,56)
16(54)
3p (53)
Körpergröße,Aktivierung desWachstumshormons
Myostatin (GDF 8) 15q23-q24(34, 65)
1(63, 64)
2q32 (63)
Muskelhyperplasie
Growth HormoneReceptor (GHR)
16q12-q13(34)
15(70)
5p13-p12(70)
Regulation desWachstumshormonsund der Synthese vonIGF1
Mahogany (Attractin,ATRN)
17(71)
2(70, 71)
20p13(70, 71)
Serum-Glykoprotein:unterdrückt einenahrungsinduzierteVerfettung
Agouti 17q21(74)
2(9, 20,42)
20p11.23-q11.2(20,27,70)
Körpergewicht,Fettwüchsigkeit,‘agouti’-Locus-Protein beeinflusstMelanozyten-stimulierendesHormon

Anhang V
370
Leptin, Obese(lep oder ob oder obs)(S0062)
18q13-q21(3, 4, 34,59)
6(1, 15,66)
7q31-q32(3, 5, 15,16, 17)
Leptinniveau,Körperfettdepots,Körpergewicht,Futteraufnahme,Sättigungsgefühl,beim Menschen in derNähe des Genortes fürLipoamid-dehydrogenase
Growth hormonereleasing hormonereceptor (GHRHR)
18q24(34)
6(70)
7p15-p14(70)
Wachstum
NEUROPEPTID Y 18q24(34)
6(70)
7p15.1(70)
Das Neuropeptid Ystimuliert den Appetit;schränkt den Energie-verbrauch ein undsteigert dieLipogenese.
Glucokinase(GK)
X(80)
Glukose-Homöostase
Mob1 7(10, 38,41)
multigeneFettwüchsigkeit,Cholesterolgehalt imPlasma,Schlachtkörperfett-gehalt
Mob2 6(38)
multigeneFettwüchsigkeit,Cholesterolgehalt imPlasma
Dob1 4(13)
1p31-(58)see LEPR
gekoppelt mitabdominalemFettdepotgewicht,ernährungsbedingteFettwüchsigkeit, distalzu db
Dob2 9(12)
Mesenterium-Fettdepotgewicht -ernährungsbedingteFettwüchsigkeit
Dob3, Dob4, Mob4 15(12, 14)
Mesenterium-Fettdepotgewicht -ernährungsbedingteFettwüchsigkeit
Mob3, Mob4 6,7,12,15(12, 14,39)
Körperfett, spezifischeFettdepots,multigeneFettwüchsigkeit

Anhang V
371
BW4 11(60)
17 (60)
assoziiert mitGenorten für Myosinheavy chain (Myhs)undGlukosetransporter 4(GLUT4) sowie dem„Early B Cell“ Faktor(Ebf) bzw. derinsulinabhängigenDiabetesanfälligkeit(Idd4) mit einemEffekt auf dieKörperfett-anreicherung
Literatur zu Tab. A15:
1) Friedman, J.M. (1991): Molecular mapping of the mouse ob mutation. Genomics 11:1054-1062.
2) Ernst, C.W., P. Kapke, M.Yerle and M.F. Rothschild (1997): The leptin receptorgene (LEPR) maps to porcine Chromosome 6. Mammalian Genome 8: 226.
3) Neuenschwander, S., G. Rettenberger, E. Meijerink, H. Jörg and G. Stranzinger(1996): Partial characterization of porcine obesity gene (OBS) and its localizationto chromosome 18 by somatic cell hybrids. Animal Genetics 27: 275-278.
4) Sasaki, S., A.C. Clutter and D. Pomp (1996): Assignment of the porcine obese(leptin) gene to Chromosome 18 by linkage analysis of a PCR-based product.Mammalian Genome 7: 471-472.
5) Weigle, D.S. and J.L. Kuijper (1996): Obesity genes and the regulation of body fatcontent. BioEssays 18: 867-874.
6) Andersson, L., C.S. Haley, H. Ellegren, S.A. Knott, M. Johansson, K. Andersson, L.Andersson-Eklund, I. Edfors-Lilja, M. Fredholm, I. Hansson, J. Håkansson and K.Lundstr öm. (1994): Genetic mapping of quantitative trait loci for growth andfatness in pigs. Science 263: 1771-1774.
7) Andersson, L., L: Marklund, P.-E. Nystr öm and S. Stern (1996): Identification of amajor QTL for fatness on pig chromosome 4. 47th Annual Meeting of EAAP,Lillehammer, Norway, p. 37.
8) Comuzzie, A.G., J.E. Hixson, L. Almasy, B.D. Mitchell, M.C. Mahaney, T.D. Dyer,M.P. Stern, J.W. MacCluer and J. Blangero (1997): A major quantitative traitlocus determining serum leptin levels and fat mass is located on human chromosome 2. Nat. Genet. 15: 273-276.
9) Drudik, D.K., D. Pomp, Z-B. Zeng and E.J. Eisen (1995): Identification of majorgenes controlling body weight and fat percentage on mouse chromosome 2. J.Anim. Sci. 73 (Suppl. 1): 110.
10) Taylor, B.A. and S.J. Phillips (1996): Detection of obesity QTLs on mousechromosome 1 and 7 by selective DNA pooling. Genomics 34: 389-398.

Anhang V
372
11) Vincent, A.L., L. Wang and M.F. Rothschild (1997): Linkage mapping of the leptinreceptor gene to pig chromosome 6. J. Anim. Sci. 75 (Suppl. 1) 145.
12) West, D.B., J. Goudey-Lefevre, B. York and G.E. Truett (1994): Dietary obesitylinked to genetic loci on chromosome 9 and 15 in a polygenic mouse model. J.Clin. Invest. 94: 1410-1416.
13) West, D.B., J. Waguespack, B. York, J. Goudey-Lefevre and R.A. Price (1994): Genetics of dietary obesity in AKR/J x SWR/J mice: segregation of the trait and identification of a linked locus on chromosome 4. Mamm. Genome 5: 546-552.
14) York, B., K. Lei and D.B. West (1996): Sensitivity to dietary obesity linked to alocus on chromosome 15 in a CAST/Ei x C57BL/6J F2 intercross. Mamm.Genome 7: 677-681.
15) Zhang, Y., Proenca, R., Maffei, M., Barone, M., Leopold, L. and Friedman, J.M. (1994): Positional cloning of the mouse obese gene and its human homologue. Nature 372: 425-432.
16) Geffroy, S., P. De Vos, B. Staels, B. Duban, J. Auwerx and B. Martinville (1995):Localization of the Human OB gene (OBS) to chromosome 7q32 by fluorescencein situ hybridization. Genomics 28: 603-604.
17) Scherer, S.W., G. Otulakowski, B.H. Robinson and L.-C. Tsui (1991): Localization of the human dihydrolipoamide dehydrogenase gene (DLD) to 7q31->q32.Cytogenetics and Cell Genetics 56: 176-177.
18) Naggert, J.K., L.D. Fricker, O. Varlamov, P.M. Nishina, Y. Rouille, D.F. Steiner,R.J. Carroll, B.J. Paigen, E.H. Leiter (1995): Hyperproinsulinaemia in obesefat/fat mice associated with a carboxypeptidase E mutation which reduces enzymeactivity. Nature Genetics 10: 135-142.
19) Noben-Trauth, K., J.K. Naggert, M.A. North, P.M. Nishina (1996): A candidate gene for the mouse mutation tubby. Nature 380: 534-538.
20) Kwon, H.Y., S.J. Bultman, C. Loffler, W.J. Chen, P.J. Furdon, J.G. Powell, A.L.Usala, W. Wilkison, I. Hansmann, R.P. Woychik (1994): Molecular structure andchromosomal mapping of the human homolog of the agouti gene. Proc. Natl.Acad. Sci. USA 91: 9760-9764.
21) Kwitek-Black, A.E., R. Carmi, G.M. Duyk, K.H. Buetow, K. Elbedour, R. Parvari,C.N. Yandava, E.M. Stone, V.C. Sheffield (1993): Linkage of Bardet-Biedlsyndrome to chromosome 16q and evidence for non-allelic genetic heterogeneity.Nature genetics 5: 392-396.
22) Collin, G.B., J.D. Marshall, L.R. Cardon, P.M. Nishina (1997): Homozygositymapping of Alstroem syndrome to chromsome 2p. Hum. Mol. Genet. 6: 213-220.
23) Tartaglia, L.A., M. Dembski, X. Weng, N. Deng, J. Culpepper, R. Devos, G.J. Richards,L.A. Campfield, F.T. Clark, J. Deeds (1995): Identification and expression cloning of a leptin receptor, OB-R. Cell 83: 1263-1271.
24) Chen, H., O. Charlat, L.A. Tartaglia, E.A: Woolf, X. Weng, S.J. Ellis, N.D. Lakey,J. Culpepper, K.J. Moore, R.E. Breitbart,.G.M. Duyk, R.I. Tepper, J.P.Morgenstern (1996): Evidence that the diabetes gene encodes the leptin receptor:identification of a mutation in the leptin receptor gene in db/db mice. Cell 84:491-495.

Anhang V
373
25) Winick, J.D., M. Stoffel, J.M. Friedman (1996): Identification of microsatellitemarkers linked to the human leptin receptor gene on chromosome 1. Genomics36: 221-222.
26) Chung, W.K., L. Power-Kehoe, M. Chua, R.L. Leibel (1996): Mapping of the OBreceptor to 1p in a region of nonconserved gene order from mouse and rat tohuman. Genome Res. 6: 431-438.
27) Wilson, B.D., M.M. Ollmann, L. Kang, M. Stoffel, G.I. Bell, G.S. Barsh (1995):Structure and function of ASP, the human homolog of the mouse agouti gene.Hum. Mol. Genet. 4: 223-230.
28) Chung, W.K., J. Goldber-Berman, L. Power-Kehoe, R.L. Leibel (1996): Molecularmapping of the tubby (tub) mutation on mouse chromosome 7. Genomics 32: 210-217.
29) North, M.A., J.K. Naggert, Y. Yan, K. Noben-Trauth, P.M. Nishina (1997):Molecular characterization of TUB, TULP1, and TULP2, members of the novel tubby gene family and their possible relation to ocular diseases. Proc. Natl.Acad. Sci. USA 94: 3128-3133.
30) Coleman, D.L. and E.M. Eicher (1990): Fat (fat) and tubby (tub): two autosomalrecessive mutations causing obesity syndromes in the mouse. J. Hered. 81: 424-427.
31) Fleury, Ch., M. Neverova, S. Collins, S. Raimbault, O. Champigny, C. Levi-Meyrueis, F. Bouillaud, M.F. Seldin, R.S. Surwit, D. Ricquier, C.H. Warden(1997): Uncoupling protein-2: a novel gene linked to obesity andhyperinsulinemia. Nature Genetics 15: 269-272.
32) Friedman, J.M., R.L. Leibel, N. Bahary (1991): Molecular mapping of obesitygenes. Mammalian Genome 1: 130-144.
33) Bouchard, C and L. Pérusse (1993): Genetics of obesity. Annu. Rev. Nutr. 13: 337-354.
34) Pig (Animal) Genome Database - synteny conservation.http://ws4.niai.affrc.go.jp/und http://www.genome.iastate.edu/pig (Stand 31.07.2001)http://www.genome.iastate.edu/maps/reprogenes.html (Stand 31.07.2001)http://www.toulouse.inra.fr/lgc/pig/compare/table.htm (Stand 31.07.2001)http://www.toulouse.inra.fr/lgc/pig/cyto/gene/chromo/SSCG6.htm (z.B. fürChromosom 6, Stand 31.07.2001)
35) Bolt, R., P. Vogeli, R. Fries (1993): A Polymorphic Microsatellite at the RYR1Locus in Swine. Anim. Genet. 24: 72.
36) MacLennan, D.H., C. Duff, F. Zorzato, J. Fujii, M. Phillips, R.G. Korneluk, W.Frodis (1990): Ryanodine receptor gene is a candidate for predisposition tomalignant hyperthermia. Nature 343: 559-561.
37) Mattei, M.G., G. Giannini, F. Moscatelli, V. Sorrentino (1994): Chromosomallocalization of murine ryanodine receptor genes RYR1, RYR2, and RYR3 by insitu hybridization. Genomics 22: 202-204.
38) Warden, C.H., J.S. Fisler, M.J. Pace, K.L. Svenson, A.J. Luis (1993): Coincidenceof genetic loci for plasma cholesterol levels and obesity in a multifactorial mousemodel. J. Clin. Invest. 92: 773-779.

Anhang V
374
39) Warden, C.H., J.S. Fisler, S.M. Shoemaker, P.Z. Wen, K.L. Svenson, M.J. Pace,A.J. Lusis (1995): Identification of four chromosomal loci determining obesity ina multifactorial mouse model. J. Clin. Invest. 95: 1545-1552.
40) Hall, C., E. Manser, N.K. Spurr, L. Lim (1993): Assignment of the humancarboxypeptidase E (CPE) gene to chromosome 4. Genomics 15: 461-463.
41) Kleyn P.W., W. Fan, S.G. Kovats, J.J Lee, J.C. Pulido, Y. Wu, L.R. Berkemeier,D.J. Misumi, L. Holmgren, O. Charlat, E.A. Woolf, O. Taybe, T. Brody, P. Shu,F. Hawkins, B. Kennedy, L. Baldini, C. Ebeling, G.D. Alperin, J. Deeds, N.D.Lakey, J. Culpepper, H. Chen, M.A. Glucksmann-Kuis (1996): Identification andcharacterization of the mouse obesity gene tubby: a member of a novel genefamily. Cell 85: 281-290.
42) Vage D.I., D.S. Lu, H. Klungland, S. Lien, S. Adalsteinsson, R.D. Cone (1997): Anon-epistatic interaction of agouti and extension in the fox, Vulpes vulpes. NatureGenetics 15: 311-315.
43) Gerbens, F., G. Rettenberger, J.A. Lenstra, J.H. Veerkamp, M.F. te Pas (1997):Characterization, chromosomal localization, and genetic variation of the porcineheart fatty acid-binding protein gene. Mamm Genome 8: 328-32.
44) Gerbens, F., A:J.M. van Erp, T.H.E. Meuwissen, J.H. Veerkamp, M.F.W. te Pas(1998): Heart fatty-acid binding protein gene variants are associated withintramuscular fat content and back fat thickness in pigs. Proceeding of the 6thWorld Congress on Genetics Applied to Livestock Production, Armidale,Australia, January 11-16, Volume 26: 187-190.
45) Prinsen, C.F., D.O. Weghuis, A.G. Kessel, J.H. Veerkamp (1997): Identification ofa human heart FABP pseudogene located on chromosome 13. Gene 193: 245-251.
46) Treuner, M., C.A. Kozak, D. Gallahan, R. Grosse, T. Müller (1994): Cloning and characterization of the mouse gene encoding mammary-derived growth inhibitor/heart-fatty acid-binding protein. Gene 147: 237-242.
47) Moser, G., E. Mueller, P. Beekmann, G. Yue, H. Geldermann (1998): Mapping ofQTLS in F2 Generations of Wild Boar, Pietrain and Meishan pigs. Proceeding ofthe 6th World Congress on Genetics Applied to Livestock Production, Armidale,Australia, January 11-16, Volume 26: 478-481.
48) Rothschild, M.F. (1998): Identification of Quantitative Trait Loci and InterestingCandidate Genes in the Pig: Progress and Prospects. Proceeding of the 6th WorldCongress on Genetics Applied to Livestock Production, Armidale, Australia,January 11-16, Volume 26: 403-409.
49) Milan, D. J.P. Bidanel, P.Le Roy, C. Chevalet, N. Woloszyn, J.C. Caritez, J.Gruand, H.Lagant, M. Bonneau, L. Lefaucheur, C. Renard, M. Vaiman, P.Mormède, C. Désautés, Y. Amigues, F. Bourgeois, J. Gellin, L.Ollivier (1998):Current status of QTL detection in Large White x Meishan Crosses in France.Proceeding of the 6th World Congress on Genetics Applied to LivestockProduction, Armidale, Australia, January 11-16, Volume 26: 414-417.
50) Koning, de D.J., L.L.G. Janss, J.A.M. Arendonk, P.A.M. van Oers, M.A.M.Groenen (1998): Mapping major genes affecting meat quality in Meishancrossbreds using standard linkage software. Proceeding of the 6th World Congresson Genetics Applied to Livestock Production, Armidale, Australia, January 11-16,Volume 26: 410-413.

Anhang V
375
51) Norman, R.A., C. Bogardus, E. Ravussin (1995): Linkage between obesity and amarker near the tumor necrosis factor-alpha locus in Pima Indians. J. Clin. Invest.96: 158-162.
52) Lawton, P., J. Nelson, R. Tizard, J.L. Browning (1995): Characterization of themouse lymphotoxin-beta gene. J. Immunol. 154: 239-246.
53) Raskin, S., J.D. Cogan, M.L. Summar, A. Moreno, M.R. Krishnamani, J.A. Phillips3rd (1996): Genetic mapping of the human pituitary-specific transcriptional factorgene and its analysis in familial panhypopituitary dwarfism. Hum. Genet. 98: 703-705.
54) Reeves, R.H., E.E. Rue, M.P. Citron, D.E. Cabin (1997): High-resolutionrecombinational map of mouse chromosome 16. Genomics 43: 202-208.
55) Wang, L., T.P. Yu, H.W. Chung, C.K. Tuggle and M.F. Rothschild (1997):Pinpointing QTLs on Chromosome 7 for Performance Traits in the Pig. J. Anim.Sci. 75 (Suppl. 1) 145.
56) Yu, T.P., C.K. Tuggle, C.B. Schmitz, M.F. Rothschild (1995): Association of PIT1Polymorphisms with Growth and Carcass Traits in Pigs. J. Anim. Sci. 73: 1282-1288.
57) Hegele, R.A., J.H. Brunt, P.W. Connelly (1995): Genetic variation on chromosome1 associated with variation in body fat distribution in men. Circulation 92: 1089-1093.
58) Chagnon, Y.C., L. Pérusse, M. Lamothe, M. Chagnon, A. Nadeau, F.T. Dionne,J. Gagnon, W.K. Chung, R.L. Leibel, C. Bouchard (1997): Suggestive linkagesbetween markers on human 1p32-p22 and body fat and insulin levels in theQuebec Family Study. Obes. Res. 5: 115-121.
59) Dai, R., N. Li, Ch. Wu (1998): Molecular cloning of ”ob” cDNA and polymorphicanalysing on ”ob” locus in swine. Proc. 6th World Congress on Genetics Appliedto Livestock Production, Armidale, Australia, Vol. 23: 612-615.
60) Brockmann, G.A., Ch.S. Haley, U. Renne, S.A. Knott, and M. Schwerin (1998):Quantitative Trait Loci Affecting Body Weight and Fatness From a Mouse LineSelected for Extreme High Growth. Genetics 150: 369-381.
61) Cargill, E.J., L.C. Baskin, and D.Pomp (1998): Rapid Communication: Localizationof the Porcine Carboxypeptidase-E Gene by Linkage Analysis Further Extendsthe region of Synteny Between Human Chromosome 4 and Porcine Chromosome8. J. Anim. Sci. 76: 2211-2212.
62) Clutter, A.C., S. Sasaki, and D. Pomp (1998): Rapid Communication: TheCholecystokinin Type-A Receptor (CCKAR) Gene Maps to Porcine Chromosome8. J. Anim. Sci. 76: 1983-1984.
63) McPherron, A.C., and S.-J. Lee (1997): Double muscling in cattle due to mutationsin the myostatin gene. Proc. Natl. Acad. Sci. USA 94: 12457-12461.
64) Szabo, G., G. Dallmann, G. Müller, L. Patthy, M. Soller, L. Varga (1998): Adeletion in the myostatin gene causes the compact (Cmpt) hypermuscularmutation in mice. Mammalian Genome 9: 671-672.
65) Sonstegard, T.S., G.A. Rohrer and T.P.L Smith (1998): Myostatin maps to porcinechromosome 15 by linkage and physical analyses. Animal Genetics 29: 19-22.

Anhang V
376
66) Keightley, P.D., T. Hardge, L. May, G. Bulfield (1996): A Genetic Map ofQuantitative Trait Loci for Body Weight in the Mouse. Genetics 142: 227-235.
67) Tornsten, A., A. Wraith, D. Larhammer, and B.P. Chowdhary (1998): FISHmapping of the porcine NPY5 gene to chromosome 8p11. Mammalian Genome 9:262-263.
68) Baskin, L.C. and D. Pomp (1998): Rapid Communication: Mapping of the PorcineColipase Gene to Chromosome 7 Using Linkage Analysis. J. Anim. Sci. 76:1241–1242.
69) Jiang, Z.H. and J.P. Gibson (1998): Rapid Communication: A PCR AmplifiedProduct Length Polymorphism (APLP) Marker at the Porcine GlucosephosphateIsomerase Locus Shows Between-Population Frequency Variation. J. Anim. Sci.76: 1718–1719.
70) Human Genome Database:http://www.ncbi.nlm.nih.gov/htbin-post/Omim/getmap (Stand 31.07.2001)
71) Edeal, B., J. M. Rumph, R. Mass, K. Killinger, N. Jerez, S. Elnagar, T. McDaneld,R. Aljumaah, U. Pithpongsiriporn, K. Nephawe, G. Martinez, C. D. Gladney, M.F. Allan, and D. Pomp (2000):Rapid communication: Linkage mapping of theMahogany (attractin) locus in cattle and pigs. J. Anim. Sci. 78: 2479–2480.
72) Te Pas, M.F.W., A. Soumillion, F. L. Harders, F. J. Verburg, T. J. van den Bosch, P.Galesloot, and T.H.E. Meuwissen (1999): Influences of Myogenin Genotypes onBirth Weight, Growth Rate, Carcass Weight, Backfat Thickness, and Lean Weightof Pigs. J. Anim. Sci. 77: 2352–2356.
73) Ernst, C.W., E.A. Mendez, A. Robic, M. Rothschild (1998). Rapid communication:myogenin (MYOG) physically maps to porcine chromosome 9q2.1-q2.6. J. Anim.Sci. 76: 328.
74) Kim, K.S., E.A. Mendez, S. Marklund A.C. Clutter, D. Pomp, and M.F. Rothschild(2000): Rapid communication: Linkage mapping of the porcine Agouti gene. J.Anim. Sci. 78: 1395–1396.
75) Kim, K.S., N. Larsen, T. Short, G. Plastow, M.F. Rothschild (2000): A missensevariant of the porcine melanocortin-4 receptor (MC4R) gene is associated withfatness, growth, and feed intake traits. Mammalian Genome 11: 131-135.
76) Kim, K.S., N. Larsen, and M.F. Rothschild (2000): Rapid communication: Linkageand physical mapping of the porcine melanocortin-4 receptor (MC4R) gene. J.Anim. Sci. 78: 791–792.
77) Lahbib-Mansais, Y., M. Yerle, J. Gellin (1995): Localization of IGF1R and EDN1genes to pig chromosomes 1 and 7 by in situ hybridisation. Cytogenet. Cell Genet.71: 225-227.
78) Werner, P., S. Neuenschwander, G. Stranzinger (1999): Characterization of theporcine uncoupling proteins 2 and 3 (UCP2 - UCP3) and their localization tochromosome 9p by somatic cell hybrids. Anim. Genet. 30: 221-224.
79) Gerbens, F., A. Jansen, A.J.M. van Erp, F.L. Harders, T.H.E. Meuwissen, G.Rettenberger, J.H. Veerkamp, M.F.W. te Pas (1998): The adipocyte fatty acid-binding protein locus: characterization and association with intramuscular fatcontent in pigs. Mammalian Genome 9: 1022-1026.

Anhang V
377
80) Niswender, K.D, Catherine Postic, T.L. Jetton, B.D. Bennett, D.W. Piston, S. Efrati,and M.A. Magnuson (1997): Cell-specific Expression and Regulation of aGlucokinase Gene Locus Transgene. J. Biol. Chem. 272: 22564–22569.
81) Messer, L.A., E.J. Cargill, and D. Pomp (2001): Rapid communication: Mapping ofthe pro-opiomelanocortin (POMC) gene to porcine chromosome by linkageanalysis using a PCR-RFLP. J. Anim. Sci. 79: 2241–2242.

378

Danksagung
379
Danksagung
Mein ganz besonderer Dank gilt Dr. Alva D. Mitchell vom Growth Biology Laboratory(GBL) des Beltsville Agricultural Research Center (BARC) des United StatesDepartment of Agriculture (USDA) in Beltsville, Maryland, USA. Die hervorragendewissenschaftliche und freundschaftliche Zusammenarbeit mit Dr. Mitchell legte denwesentlichen Grundstein für diese Arbeit. Daneben gilt mein Dank Dr. Norman Steele,der in seiner damaligen Funktion als „Research Leader“ des GBL alle nötigenForschungsmittel schnell und komplikationslos zur Verfügung stellte und damit meineArbeit entscheidend voran trieb.
Ohne die finanzielle Unterstützung der Alexander von Humboldt-Stiftung (AvH),Bonn, die mein Projekt mit einem Feodor-Lynen-Stipendium (05/1994-10/1995)förderte, wäre die gesamte Arbeit jedoch nicht zustande gekommen. In diesemZusammenhang danke ich Prof. Dr. Raymond Bernor (College of Medicine,Department of Anatomy, Howard University, Washington, DC), der meinen Aufenthaltin den USA als Gastgeber im Namen der AvH überhaupt erst ermöglichte.
Ein weiteres Stipendium zur Fortführung meiner Arbeiten am USDA in Beltsvilleund an der Howard University wurde mir durch die Organisation für ÖkonomischeZusammenarbeit und Entwicklung (OECD), Paris, Frankreich, im Rahmen desProgramms „Biological Resource Management for Sustainable Agricultural Systems:Quality of Animal Products and Safety of Food“ gewährt (06-07/1996).
Am „College of Medicine, Department of Radiology“ der Howard University,Washington, DC, erfuhr ich im Verlauf der Magnetresonanz-Studien eine sehrzuvorkommende Unterstützung durch Prof. Paul C. Wang, „Research Leader“ des„Biomedical NMR Research Facility“ sowie durch seine Mitarbeiter Huafu Song undZhenjin Yan.
Mein herzlicher Dank gilt ebenso Joan K. Lunney (Ph.D), „Research Leader“ amImmunology and Disease Resistance Laboratory (BARC, USDA) sowie ihrenMitarbeitern Dr. David Grimm und Rhoanda Shanks, die die Arbeiten zur DNA-Analyse durch meine Frau, Wiebke Scholz, in ihrem Labor sehr hilfsbereitunterstützten.
Weiterhin danke ich Dr. M.B. Solomon, „Research Leader“ des „Meat ScienceLaboratory“ des USDA in Beltsville und seiner Mitarbeiterin Janet Eastridge für diesehr entgegenkommende Hilfe bei der Konservierung und Präparation derMuskelbiopsie-Proben.
Nicht zuletzt danke ich Nancy Falconer, Mitarbeiterin von Dr. Mitchell, für ihrentatkräftigen „Einsatz“ bei der chemischen Analyse der Gewebeproben.
Einen sehr großen Beitrag bei der unermüdlichen Literaturbeschaffung leisteteFrau Dr. Ulrike Funke vom Institut für Nutztierwissenschaften der Humboldt-Universität zu Berlin. Herzlichen Dank.
Prof. Dr. E. Kallweit, Dr. U. Baulain und Frau Dr. Martina Henning vom Institutfür Tierzucht und Tierverhalten der Bundesforschungsanstalt für Landwirtschaft inMariensee danke ich speziell dafür, mir den Weg in das Forschungsgebiet „In-vivo-Analyse komplexer biologischer Systeme“ im Rahmen eines ebenfalls durch die AvHgeförderten Forschungsaufenthaltes von 05-10/1991 eröffnet zu haben.
Außerdem danke ich den Herren Prof. Dr. G. Schönmuth und Prof. Dr. G.Triebler, ehemaliges Institut für Tierzucht und Haustiergenetik der Humboldt-Universität zu Berlin, Prof. Dr. W. Schlote, Institut für Nutztierwissenschaften derHumboldt-Universität zu Berlin und Prof. Dr. M. Förster, Institut für Tierzucht undAllgemeine Landwirtschaftslehre der Ludwig-Maximilians-Universität München für diestetige Förderung meiner wissenschaftlichen Arbeiten.