improvement of a serodiagnostic strategy for foot-and-mouth disease virus surveillance in cattle...
TRANSCRIPT

Arch Virol (2000) 145: 473–489
Improvement of a serodiagnostic strategy for foot-and-mouthdisease virus surveillance in cattle under systematic vaccination:
a combined system of an indirect ELISA-3ABC with anenzyme-linked immunoelectrotransfer blot assay
I. E. Bergmann1, V. Malirat 1,∗, E. Neitzert1, E. Beck2, N. Panizzutti3,C. Sánchez4, andA. Falczuk5
1Pan-American Foot-and-Mouth Disease Center (PAHO/WHO), Rio de Janeiro, Brazil2Biochemisches Institut am Klinikum der Justus-Liebig Universität, Giessen, Germany
3Laboratório Regional de Apoio Animal-LARA/RS, Porto Alegre, Brazil4Instituto Colombiano de Agropecuaria, ICA, Santafé de Bogotá, Colombia
5Gerencia de Laboratorios/Servicio Nacional de Sanidad Animal Gelab,SENASA, Buenos Aires, Argentina
Accepted October 24, 1999
Summary.Foot-and-mouth disease (FMD) recombinant non-capsideal viral anti-gens 3A, 3B, 2C, 3D and 3ABC were assessed individually in an indirect enzyme-linked immunosorbent assay (I-ELISA) for their ability to screen for persistentinfection-specific antibodies in cattle, regardless of vaccination condition. Resultsof sequential serum samples from non-vaccinated animals with experimentallyinduced persistent infection, and their correlation with virus isolation, indicatedthat the polypeptides 3A, 3B and 3ABC showed the most adequate characteristicsfor further field studies.
Reliable performance of the I-ELISA with the selected antigen 3ABC wasindicated by the distinct patterns observed for the frequency distribution valuesof naive and true positive samples. For regularly vaccinated livestock, a clearnegative profile was proved in samples from regions without recent history ofFMD. In contrast, at 90 and 900 days post-outbreak, coexistence of a positive anda negative population was established. These findings indicated that, irrespectiveof vaccination, the test allowed a classification of the herd-disease status.
A high degree of agreement was observed between I-ELISA-3ABC andEITB results for clearly reactive and non-reactive sera. For samples with reactivity
∗Current address: Departamento de Virologia, Instituto de Microbiologia, UniversidadeFederal do Rio de Janeiro, CCS, Ilha do Fundão, Brazil.

474 I. E. Bergmann et al.
values close to that of the cut-off, the EITB profiles upheld the definition of theinfection condition. On this basis, screening by I-ELISA-3ABC, together withconfirmation of suspect or positive samples by EITB is proposed as an adequateand accurate approach for large-scale epidemiological surveillance.
Introduction
Since 1988, when foot-and-mouth disease (FMD) eradication programs wereimplemented in South American countries, continuous progress of the epidemi-ological situation in the region emphasised the need for the development andapplication of rapid and accurate serodiagnostic approaches to assess residualinfection-specific antibodies in a livestock population. This is of great relevancefor surveillance purposes and for the safety of animal movement.
Since control programs in regions under eradication campaigns are dependenton systematic immunisation, assays should be based on the detection of antibod-ies against non-capsideal proteins which, in principle, are induced only duringinfection and not upon vaccination [2, 9, 13]. In this regard, we reported the useof recombinant DNA-derived non-capsideal FMD virus (FMDV) antigens 3A,3B, 2C, 3D and 3ABC in an enzyme-linked immunoelectrotransfer blot assay(EITB) for detection of infection-specific antibodies in sera from cattle with per-sistent infection [3, 4, 13]. Field application of this approach for identification ofFMD-free regions was described [5, 6].
The use of bioengineered non-capsideal viral proteins has been extended toELISA-based assays using either individual antigens [7, 11, 12, 14, 20–22] orpanels of them [10, 19]. Although evaluation has been conducted using experi-mental model systems or limited models under natural conditions, no significantfield studies have been carried out to allow for accurate evaluation of the test per-formance when applied to the various epidemiological situations that arise withinFMD virus surveillance and monitoring programs.
The objective of the study reported here was to compare the effectiveness ofthe individual non-capsideal antigens 3A, 3B, 2C, 3D and 3ABC for use in anindirect ELISA (I-ELISA)-based assay, in order to select the most suitable one fordetection of FMDV infection-specific antibodies in asymptomatic cattle. Evalu-ation of the performance of the I-ELISA with the selected antigen under variousfield conditions, including areas under intense vaccination programs, and corre-lation of test results with those achieved through the EITB test, which allowsfor the simultaneous analysis of all the antigens in a single assay, was under-taken to design the most adequate and accurate diagnostic approach for FMDVsurveillance.
Materials and methods
Test samples
For preliminary sensitivity and specificity studies, population of known positive or negativesamples were examined. Negative sera (250), derived from various FMD-free regions with-out vaccination, did not contain antibodies to the capsideal proteins, as measured by liquid

Serodiagnostic strategy for FMDV surveillance 475
phase blocking sandwich ELISA [8] and immunodot analysis [16]. Positive sera included105 animals involved in field outbreaks and of proven disease status, collected 20 days af-ter detection of the episode. The FMDV-positive population also covered 80 samples fromnon-vaccinated cattle experimentally infected with either one of the three South Americanserotypes of FMDV, collected 14 days after infection.
For the analysis of the persistent state, a set of sera was selected that included serialblood samples collected from cattle with experimentally induced persistent infection [3].Oesophageal pharyngeal (OP) fluid and serum samples obtained from each animal at 2-or 4-week intervals were analysed for virus recovery in cell culture and for presence ofnon-capsideal proteins by use of the EITB test, respectively, during 106 weeks. The cattlewere non-vaccinated Zebu steers from a farm that had been FMD-free for several years.Serum samples screened prior to infection were EITB-negative and gave I-ELISA test/cut-off absorbance values close to 0.1 for all the antigens. Results obtained for virus isolationand for EITB have been published elsewhere [3].
Frequency distribution of positive and negative samples was established by examiningpanels of sera from model systems representing different field situations. These included: 833sera from various free areas without vaccination; 28 samples from animals experimentallyinfected with a representative strain of one of the three South American serotypes, collected20 days after inoculation; 119 sera corresponding to cattle involved in FMD field outbreaks,in which reports indicated that most of the animals showed clinical signs of the disease,bled between 20 and 40 days after detection of the episode; 84 sera from cattle in FMD-free regions, collected 30 days after primovaccination with trivalent binary ethyleneimine(BEI)-inactivated commercial vaccines, from various producers; 4396 sera from differentgeographic regions without clinical FMD for over 4 years, which were still under intensevaccination programs, collected as in regular surveys 5–6 months after the last round ofvaccination, from 3754 and 642 animals under and over two years of age, respectively. Mostof these sera had been collected in support of the OIE requirements for declaration of FMD-free regions without vaccination. Samples were also assessed from cattle in a farm where anepisode occurred after having been FMD-free for at least the past 6 years. The livestock hadbeen and continued to be systematically vaccinated every 6 months with a vaccine known tolack interference even when applied under conditions that should lead to increased overallantibody response [3]. Forty samples, 20 of which were derived from cattle with clinicaldisease, were collected at 90 days post-outbreak (DPO). Out of the 40 animals studied, 12became persistently infected as indicated by virus recovery from OP fluid, 6 of which didnot show clinical symptoms. Another set of 40 serum samples was taken at 900 DPO. LastOP positive viral isolation in the herd was at 270 DPO.
Immunoassays
I-ELISAAll bioengineered FMDV antigens were expressed inE. coli and purified to electrophoretichomogeneity for use as serologic probes as described [13]. The assay is used as a singledilution screening test. ELISA plates (Immulon II, Dynatech Laboratories Inc.) were coatedby overnight incubation at 4◦C of optimal dilutions of non-capsideal proteins (100 ng/well)in carbonate/bicarbonate buffer pH 9.6. Plates were washed six times between stages withphosphate buffered saline (PBS)/0.05% Tween 20. A 1:20 dilution of test or reference sera inblocking buffer (PBS/0.05% Tween 20/5% non-fat dry milk/10% of equine sera/0.1%E. colilysate) was added to the plate. Sera were incubated for 30 min at 37◦C. Following washing,bound antibodies were detected by incubation for 30 min at 37◦C of an optimal dilutionin blocking buffer of rabbit anti-bovine IgG peroxidase conjugate (Sigma Chemical Co.).After the final wash, detection was made using 42 nM 3,3′,5,5′-tetramethylbenzidine plus
476 I. E. Bergmann et al.
0.004% (w/v) H2O2 in phosphate-citrate buffer, pH 5.5. Colour development was stoppedafter 15 min by the addition of 0.5M H2SO4. The optical density of each well was read at450 nm and at 620 nm for background correction. In order to control inter-assay variation,reference sera were included on every plate. For 3ABC, the intra-assay coefficient of variation(CV) was≤10%, and the inter-assay CV≤ 20% [11]. Each plate contains two or more blankwells, consisting of diluent without added serum, and a set of reference standards as definedfor the EITB assay [4, 6]. The cut-off control serum is derived from a pool of sera and/ordilution of positive sera, representing background EITB reactivities observed individuallyfor the different antigens in non-vaccinated animals from FMD-free regions. These controlsare secondary standards, validated against a primary standard corresponding to a serum froman animal at 714 days after experimental infection from which no virus had been recoveredfrom OP fluid for the past 364 days [3].
For the test system to be valid, the following performance criteria are applied. Theabsorbance of negative controls should be< 0.10 after correction for absorbance blank wells.The cut-off serum should give absorbance values of 0.15–0.40. Results are expressed as anindex derived by dividing the absorbance value of the serum tested (T) by that of the cut-offcontrol (C). The ratio of the weak positive/cut-off controls should be 2.5 with a coefficientof variation< 20%. Using this ratio the test sera are classified as non-reactive (T/C≤ 0.8),suspect (0.8< T/C≤ 1.0) or reactive (T/C> 1.0).
EITB
The EITB test was performed according to the procedure described previously [6, 13].
Results
Preliminary performance of the I-ELISAs
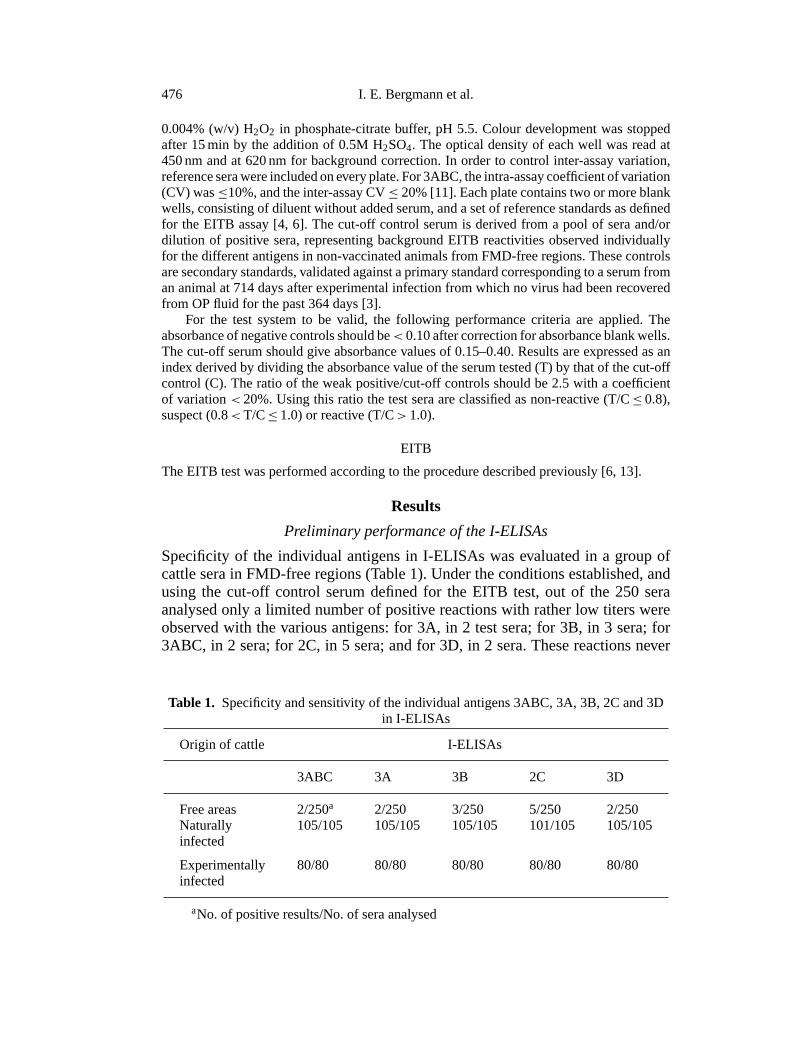
Specificity of the individual antigens in I-ELISAs was evaluated in a group ofcattle sera in FMD-free regions (Table 1). Under the conditions established, andusing the cut-off control serum defined for the EITB test, out of the 250 seraanalysed only a limited number of positive reactions with rather low titers wereobserved with the various antigens: for 3A, in 2 test sera; for 3B, in 3 sera; for3ABC, in 2 sera; for 2C, in 5 sera; and for 3D, in 2 sera. These reactions never
Table 1. Specificity and sensitivity of the individual antigens 3ABC, 3A, 3B, 2C and 3Din I-ELISAs
Origin of cattle I-ELISAs
3ABC 3A 3B 2C 3D
Free areas 2/250a 2/250 3/250 5/250 2/250Naturally 105/105 105/105 105/105 101/105 105/105infected
Experimentally 80/80 80/80 80/80 80/80 80/80infected
aNo. of positive results/No. of sera analysed

Serodiagnostic strategy for FMDV surveillance 477
Fig. 1 (continued)
478 I. E. Bergmann et al.

Serodiagnostic strategy for FMDV surveillance 479
occurred simultaneously in a single sample with more than 2 antigens, and usuallywere reflected as individual bands in the EITB test.
Sensitivity of the assay using the aforementioned antigens was evaluated byexamining 105 sera from animals involved in field outbreaks, and of provendisease status. As can be seen in Table 1, all the I-ELISAs, except for 2C, correctlyidentified sera from infected cattle as positive. Another group of 80 non-vaccinatedanimals inoculated with either one of the three South American serotypes ofFMDV was assessed. Sera before inoculation were I-ELISA negative, but at 14days post-infection (PI) all of them yielded positive results with each one of theantigens. The degree of the antibody response was not related to the virus serotypewith which the animal had been infected.
Evaluation of antibody responses during experimentallyinduced persistent infection
The results mentioned above indicated that, except for antigen 2C, sensitivity ofthe I-ELISAs for all antigens is close to 100%, at least for detection of FMDVinfection during the acute phase of the disease. The question still remains as tothe sensitivity of the methods to recognise FMDV infection-specific antibodies inanimals at late stages of the persistent infection, during which sera titers may bequite low, and the infection status of the animal is difficult to assess with certaintyby the currently available procedures.
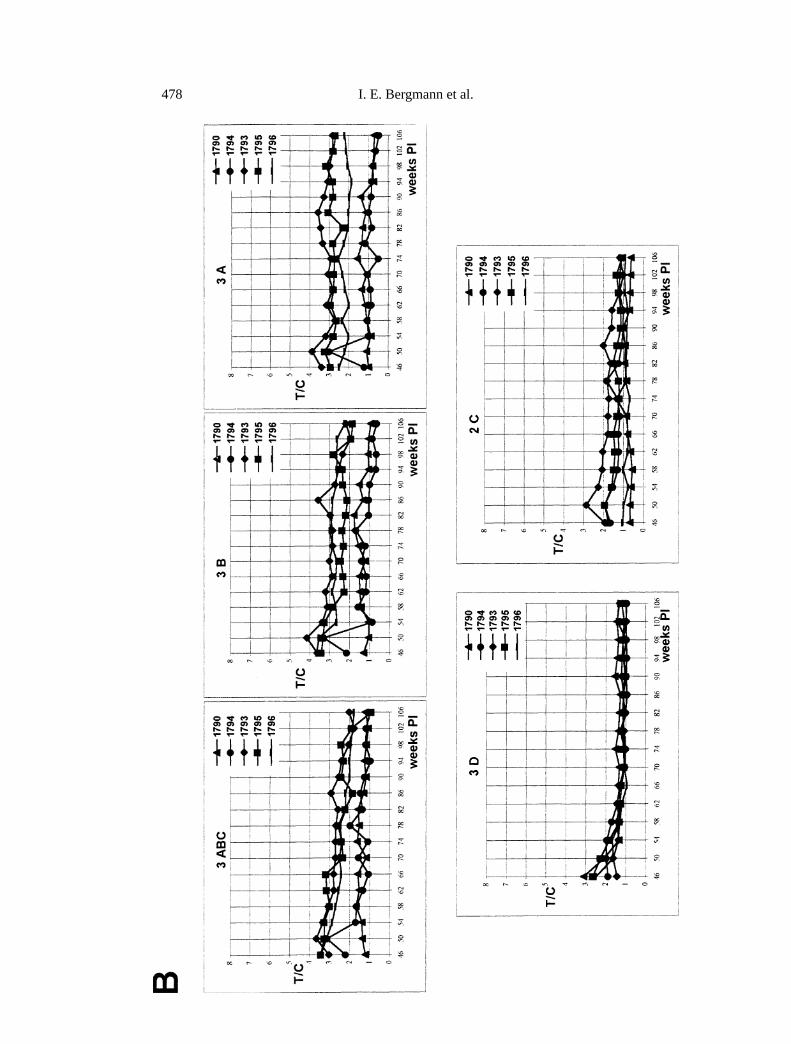
To this aim, and to compare the reliability of the I-ELISAs using individualpolypeptides 3A, 3B, 2C, 3D and 3ABC to detect infection-specific antibodies insera from cattle with persistent FMDV infection, evaluation was made of the timecourse of the antibody response to these antigens in sera obtained after experi-mental infection of various non-vaccinated animals. The results were comparedwith those obtained by virus isolation procedures and by EITB. Serum samplesfrom four cattle were screened immediately prior to infection, at one week PI,and thereafter serially at two-week intervals for up to 40 weeks (animals 973,982, 987) or 30 weeks (animal 977) (Fig. 1A). In general, and in agreement withresults by EITB, in the case of I-ELISAs using 3ABC, 3B and 3A, high antibodytiters were observed by the second week PI, which lasted throughout the study.
bFig. 1. Time course of the antibody response of experimentally infected cattle to individualantigens 3ABC, 3A, 3B, 3D, and 2C by I-ELISAs.A Sera from four infected cattle (# 973,977, 982 and 987) were tested against the indicated recombinant antigen at various weeksPI. Virus recovery from OP fluid was positive at 1, 2, 4, 6, 8, 10, 12, 14, 32 and 40 weeks PIfor animal 973; at 1, 2, 4, 6, 8, 10, 12, 14, 18, 20, 22, 30, 32, 36 and 40 weeks PI for animal977; at 1, 2, 4, 14, 34, 38 and 40 weeks PI for animal 982; and at 1, 6, 8, 12, 14, 16, 20, 34,36, 38 and 40 weeks PI for animal 987.B Sera from five infected cattle (# 1790, 1794, 1793,1795 and 1796) were tested against the indicated recombinant antigen at various weeks PI.Virus recovery from OP fluid was positive at 47 weeks PI for animal 1790; at 50 weeks PIfor animal 1794; at 46, 54 and 78 weeks PI for animal 1793; at 46, 66, 78, 86 and 90 weeksPI for animal 1795; and at 46, 50, 78 and 86 weeks PI for animal 1796

480 I. E. Bergmann et al.
A considerable increase in the absorbance values, with respect to those priorto inoculation could be detected at about 7 days after experimental infection,reaching values close to that of the cut-off (T/C∼1.0), and increasing with timeafter inoculation to a peak somewhere between 6 and 8 weeks PI. Subsequently, aslight and gradual decrease in the antibody response was registered as the infectionprogressed. For the I-ELISA-3D similar reactivity patterns were observed duringthe first 10–12 weeks PI. From then on, reactivity decreased appreciably, yieldingrather low ratios as early as 20–24 weeks PI, which were maintained thereafter.The ELISA-2C showed relatively low T/C values throughout the study.
Sensitivity of the I-ELISAs using the aforementioned antigens was furtherexamined in sera from five infected cattle at late stages of persistent infectionduring which virus was not recovered from the OP fluid or was only occasionallyrecovered (Fig. 1B). Serum samples were analysed serially at 4-week intervalscovering a period between 46 to 106 weeks PI. Overall, and in agreement withresults obtained by EITB, I-ELISA reactivities were still registered at 106 weeksPI for all the antigens. For I-ELISAs using 3ABC, 3B and 3A, higher T/C valueswere observed in sera from animals 1793, 1795 and 1796, in which virus wasoccasionally isolated from OP fluid. Lower ratios were established in sera fromcattle 1790 and 1794, in which virus was last recovered from OP fluid at 47 and50 weeks PI, respectively. In contrast, antigens 3D and 2C gave values very nearto those of the cut-off control sera for all the animals.
Frequency distribution of test results from sera of animalswith various status in the I-ELISA-3ABC
From the above-mentioned data, 3A, 3B and 3ABC appeared as the most ade-quate antigens for detection of infection-specific antibodies. On this basis, and forpractical reasons, the I-ELISA using polyprotein 3ABC was selected to extendthe performance characteristics and to determine whether the cut-off sera appliedremained appropriate for use under extensive field conditions. To this aim, fre-quency distribution of T/C values was recorded for sera from various regions withdefined epidemiological situation.
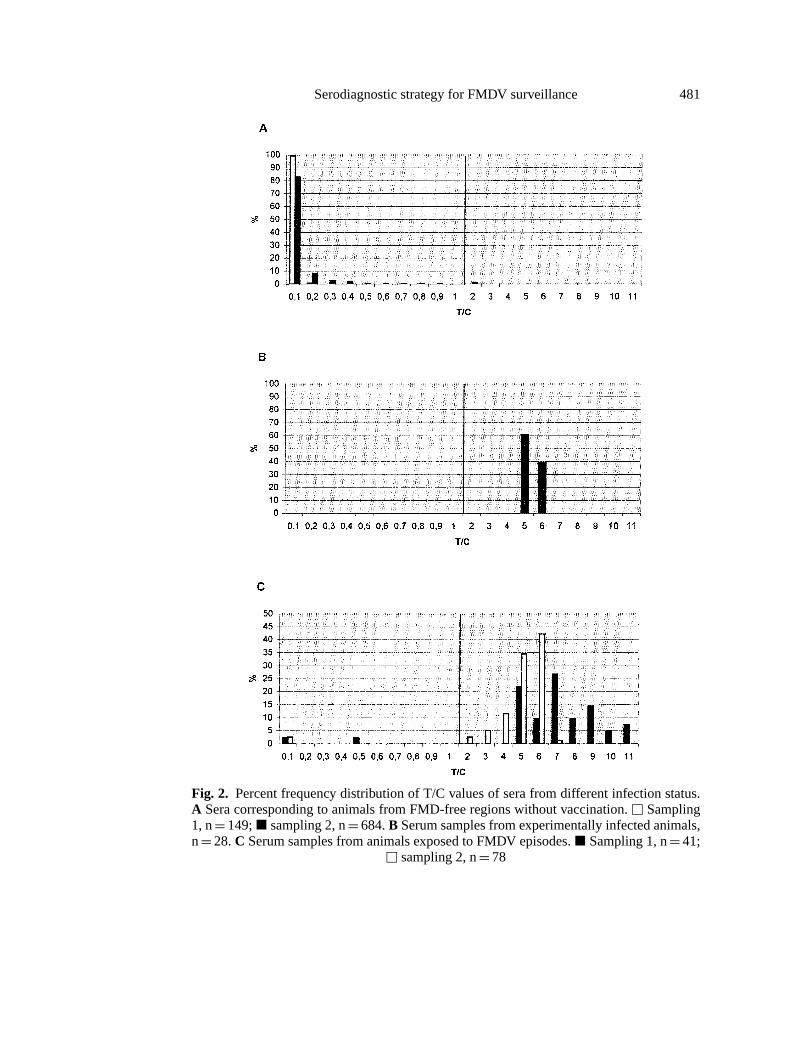
The results plotted on the frequency distribution chart in Fig. 2 clearly showthat there was a distinct separation between the true positive and the true negativepopulation. Most of the 833 sera representing the negative population, free regionswithout vaccination, is resolved in T/C≤ 0.1 (99.3% and 83.0% for sampling 1and 2, respectively), with none (sampling 1) or only 1.4% (sampling 2) of thesera with T/C values> 0.8 but≤ 2.0 (Fig. 2A). In contrast, all of the 28 sera fromthe true positive population, collected 20 days after experimental infection, aredistributed in T/C values> 4.0 (Fig. 2B). As expected, a more heterogeneousdistribution of T/C reactive values is observed in sera from cattle populationsinvolved in field outbreaks collected 20–40 days after detection of the episode(Fig. 2C). High T/C ratios (> 4.0) are attained in all of the reactive samples fromepisode 1, and in 80.2% of those from outbreak 2. Under field conditions, a portionof the cattle will probably not become infected, which could explain the negative
Serodiagnostic strategy for FMDV surveillance 481
Fig. 2. Percent frequency distribution of T/C values of sera from different infection status.A Sera corresponding to animals from FMD-free regions without vaccination.u Sampling1, n= 149;j sampling 2, n= 684.B Serum samples from experimentally infected animals,n= 28.C Serum samples from animals exposed to FMDV episodes.j Sampling 1, n= 41;
u sampling 2, n= 78

482 I. E. Bergmann et al.
results observed in these populations (4.8% in sampling 1 and 2.6% in sampling2).
Comparison with results obtained by EITB demonstrated a high degree ofagreement between the I-ELISA and EITB for sera that were either clearly positiveor clearly negative. Rarely, borderline results could be observed with the individ-ual antigens in the EITB for samples with I-ELISA T/C ratios≤ 0.8, whereas allsera with T/C values> 3.0 elicited the typical profile of EITB infection-specificantibodies, represented by the four antigens 3A, 3B, 3D and 3ABC,± 2C. InFMD-free areas, all of the 10 I-ELISA suspect sera (1.2% of the overall populationwith T/C> 0.8 but still≤ 2.0) were negative by EITB. Reactions against singlebands observed in the latter test may account for the I-ELISA responses. Screen-ing of antibodies to capsideal proteins of serotypes O1Campos, A24 Cruzeiro, orC3 Indaial in such sera gave negative results by ELISA and immunodot anal-ysis, indicating that observed I-ELISA/EITB reactivities represent non-specificresults. On the other hand, out of the six sera with I-ELISA responses> 1.0 but≤ 3.0 from cattle in episode 2, five reacted with all bands in the EITB (typi-cal infection profile), and only one sample reacted weakly with 3 out of the 5antigens (indeterminate category in the EITB), possibly in the seroconversionphase.
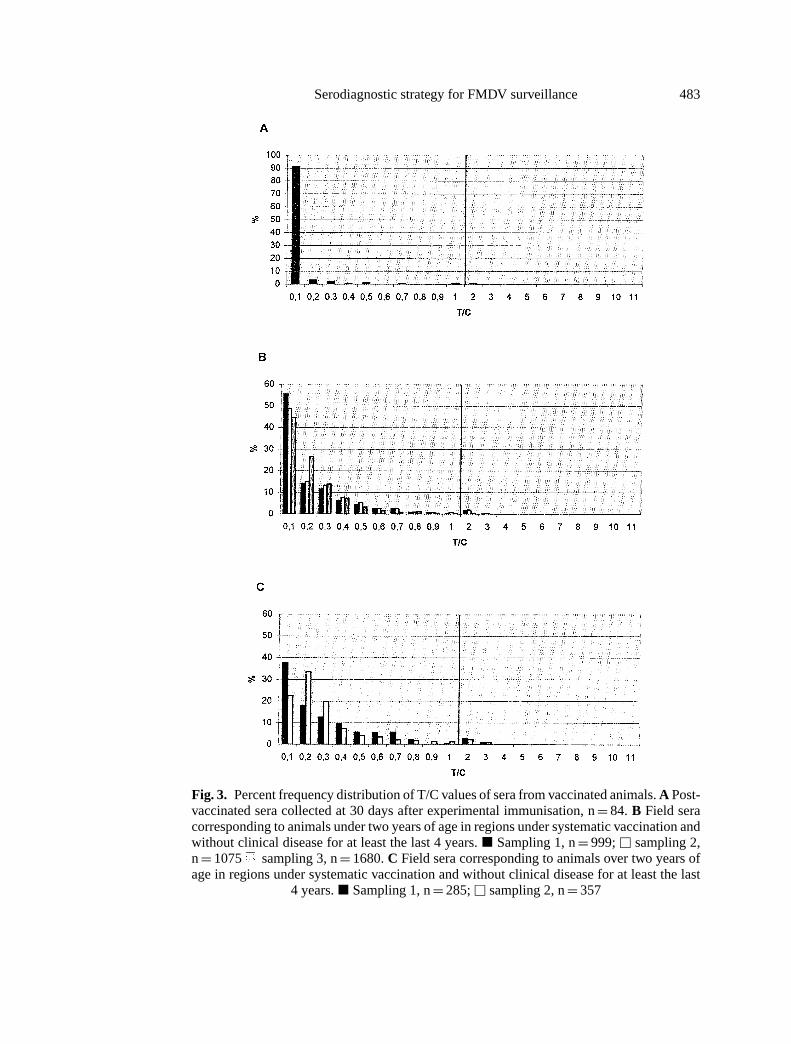
The presence of residual non-capsideal FMDV antigens in vaccine prepara-tions that could induce antibody responses against antigen 3ABC was furtherexamined. Figure 3A shows the distribution of results from 84 sera obtained fromvaccinated cattle immunised with various commercial vaccines collected 30 daysafter vaccination. As can be seen, negative results resolved under a rather similardistribution as to that observed in FMD-free areas (Fig. 2A). All sera, includingthe one with T/C= 1.1, were confirmed negative in the EITB test.
Methods for antigen purification and concentration used to manufacture vac-cines in laboratories in South America may vary widely. The goal of this devel-opment was mainly to monitor regions under advanced eradication campaignswhere high immunological coverage is expected. On this basis, and taking intoaccount that under field conditions, traces of non-capsideal viral antigens mayinduce responses after multiple vaccination cycles, or even act as boosters in cat-tle with low titer of antibodies to replicating aphthovirus, the potential effect ofresidual 3ABC in the various vaccine preparations when used under the complexfield situation found in South America, was further examined. To this effect, 4396sera from different regions with no history of FMD during at least the past 4 yearsprior to collection of the samples, corresponding to 3754 and 642 animals underand over two years of age, respectively subjected to regular vaccination programswere analysed. As shown in Figs. 3B and 3C, profiles were comparable to thosefrom populations of free regions without vaccination (Fig. 2A), as well as to thosefrom experimentally vaccinated cattle (Fig. 3A). Although there was a clear res-olution of the negative population, with a considerable number of samples withT/C≤ 0.1, long tails were detected, reaching values> 0.8 in 2.1% and 4.8% ofthe population under and over two years of age, respectively. No T/C values> 3were registered. Whereas in the true naive population, and in the experimentally
Serodiagnostic strategy for FMDV surveillance 483
Fig. 3. Percent frequency distribution of T/C values of sera from vaccinated animals.A Post-vaccinated sera collected at 30 days after experimental immunisation, n= 84. B Field seracorresponding to animals under two years of age in regions under systematic vaccination andwithout clinical disease for at least the last 4 years.j Sampling 1, n= 999;u sampling 2,n= 1075 sampling 3, n= 1680.C Field sera corresponding to animals over two years ofage in regions under systematic vaccination and without clinical disease for at least the last
4 years.j Sampling 1, n= 285;u sampling 2, n= 357

484 I. E. Bergmann et al.
vaccinated cattle, in average, 86.8% of the samples were defined in T/C≤ 0.1,in the populations from regions under systematic vaccination campaigns, 48.7%and 29.1% of the sera from animals under and over two years of age, respectivelyare resolved in this category. Distribution of the rest of the sera showed frequencyvalues decreasing as a function of increased T/C.
Comparison with results obtained by EITB indicated that all I-ELISA-negativeresults were confirmed negative in the EITB assay. For the population under twoyears of age, 70 (87.5%) of the 80 samples with T/C> 0.8 gave negative resultsin the EITB assay. Indeterminate EITB reactivities could be established in 9 seraof this population, whereas only one of the sera in this category showed reactivitynear that of the cut-off control serum, representing 0.026% of the overall popu-lation under two years of age. Among the population over two years of age, ofthe 31 samples showing T/C> 0.8, 22 were EITB negative, 7 gave indeterminatereactivities and only 2 samples gave EITB-positive reactivities (0.3% of the totalover two year-old category).
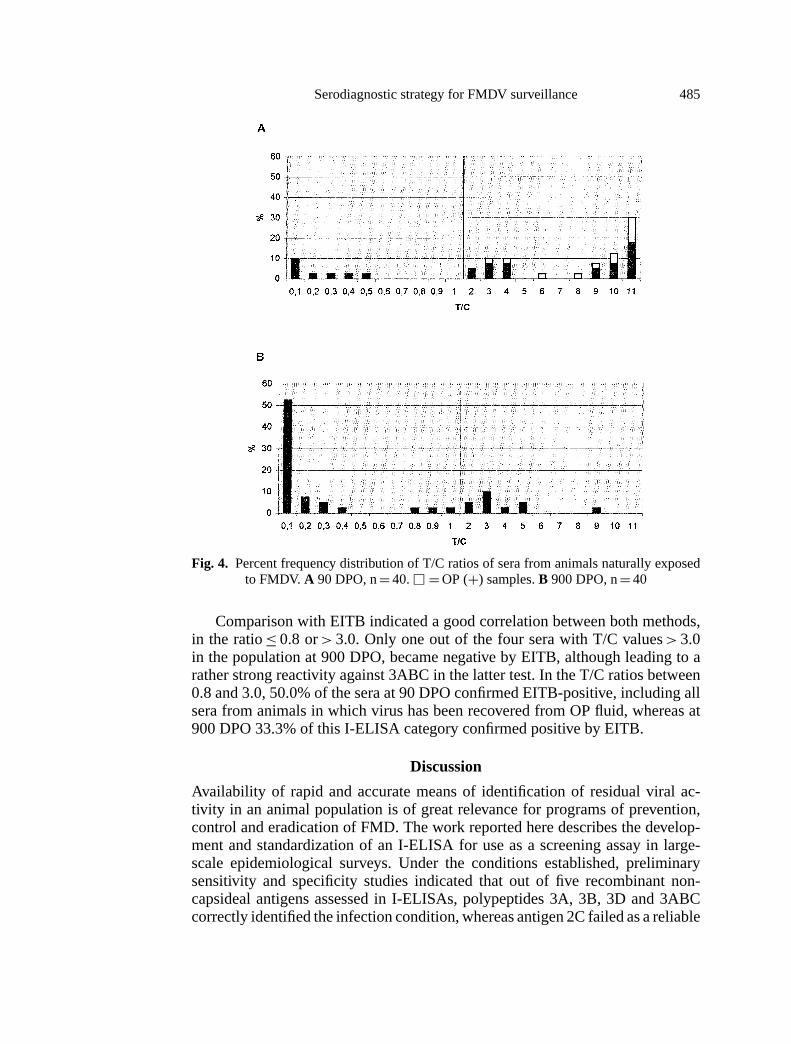
Although background or weak-positive reactions demonstrated in regions un-der regular vaccination with no recent history of FMD could be attributed to vac-cinations, antibody scars from previous infection or even subclinical infection,particularly in the over two year-old population, cannot be ruled out. To furtherevaluate the origin of these reactions, persistence of antibody responses against3ABC was examined in sera from regularly vaccinated animals obtained at differ-ent times after natural exposure to FMDV. To this effect, sera from systematicallyvaccinated cattle were analysed at 90 and 900 days post-outbreak (DPO) (Fig. 4).The frequency distribution of T/C values for these populations was markedlydifferent (bimodal), from that described above. In this case, a distinct negativepopulation coexisted with samples with T/C> 0.8, including sera with high values(T/C> 3.0). At 90 DPO (Fig. 4A), 80.0% of the sera yielded I-ELISA-positiveresults, reflecting the recent exposure of the herd. Within this positive population,high titer sera were the predominant feature. In addition, the assay can differen-tiate an antibody negative population (20.0% of the herd), that probably did notbecome infected. Correlation with OP-positive recoveries also indicated a cleardistribution between the persistently infected and non-infected populations. All ofthe OP-positive samples, including those from the 6 asymptomatic animals, gaveI-ELISA-positive results. Only one of the samples derived from the asymptomaticOP-positive cattle, resolved with relatively low titer (T/C= 2.5). Frequency dis-tribution established for sera at 900 DPO (Fig. 4B) still resolved as two distinctpopulations. In this case and in contrast to the distribution at 90 DPO, 70.0% ofthe sera gave T/C values≤0.8, indicating a clear shift of the positive populationtowards the negative range as a function of time after exposure. Variation wasobserved both in the total number of reactive samples, as well as in the magnitudeof the overall response against 3ABC. Positive values could still be detected in25% of the samples, although the last virus recovery in the population occurred at270 DPO. Thus, it seems that, as established in the experimental non-vaccinatedpersistently infected animals (Fig. 1B), antibodies from previous infection can bemaintained longer than the virus in the OP fluid.
Serodiagnostic strategy for FMDV surveillance 485
Fig. 4. Percent frequency distribution of T/C ratios of sera from animals naturally exposedto FMDV. A 90 DPO, n= 40.u = OP (+) samples.B 900 DPO, n= 40
Comparison with EITB indicated a good correlation between both methods,in the ratio≤ 0.8 or> 3.0. Only one out of the four sera with T/C values> 3.0in the population at 900 DPO, became negative by EITB, although leading to arather strong reactivity against 3ABC in the latter test. In the T/C ratios between0.8 and 3.0, 50.0% of the sera at 90 DPO confirmed EITB-positive, including allsera from animals in which virus has been recovered from OP fluid, whereas at900 DPO 33.3% of this I-ELISA category confirmed positive by EITB.
Discussion
Availability of rapid and accurate means of identification of residual viral ac-tivity in an animal population is of great relevance for programs of prevention,control and eradication of FMD. The work reported here describes the develop-ment and standardization of an I-ELISA for use as a screening assay in large-scale epidemiological surveys. Under the conditions established, preliminarysensitivity and specificity studies indicated that out of five recombinant non-capsideal antigens assessed in I-ELISAs, polypeptides 3A, 3B, 3D and 3ABCcorrectly identified the infection condition, whereas antigen 2C failed as a reliable

486 I. E. Bergmann et al.
indicator of the infection status. In general these findings are in agreement withresults published previously [10, 19], but differ from those reported for a bac-ulovirus expressed 2C [12]. Differences in protein configuration could accountfor the discrepancies observed.
High sensitivity of all the I-ELISAs for detecting persistent antibodies wasindicated by reactivities observed in sera from the experimentally non-vaccinatedpersistently infected animals, even at 106 weeks PI. The assays using either 3A,3B or 3ABC were found to provide the best results to detect specific antibodiesin animals with aphthovirus persistence at late stages of infection. Correlationbetween virus recovery from OP fluid and serum antibody titer against theseantigens demonstrated that the strongest reactions were observed during the earlyphases of persistence, when viral replication and consequently virus recovery ismaximal. In addition, relatively higher titers were observed in sera of cattle fromwhich virus was occasionally isolated, compared with those from which viruswas no longer recovered. The latter samples had titers close to cut-off levels, evenat 300 days after the last virus isolation from OP fluid.
Based on these findings and on previous experiences [3, 10, 19] polyprotein3ABC was selected to extend the validation criteria of the I-ELISA for widespreaduse in the field. Under the conditions established, distinct distribution of serafrom naive and infected cattle, including those with proven persistent infectionirrespective of whether or not clinical signs were observed, was attained. Hence,the cut-off selected allowed classification of test results into positive or negativecategories. However, 0.8% of the naive cattle yielded T/C values> 1.0 yet≤ 2.0.
I-ELISA-weak reactivities were more frequent in regions with no recent his-tory of FMD, subjected to regular vaccination campaigns. Previous studies withother tests had also documented antibody reactivities against non-capsideal pro-teins in sera from animals after vaccination or revaccination [1, 3, 17]. Antibodieselicited upon immunisation in the field might be affected by factors such as animalage and boosters from previous infection. In fact, a higher percentage of weak-positive results was observed in sera from animals over two years of age, whencompared to that in the younger population, born after the last FMD episode.Thus, sampling analysis considering an age distribution constitutes an additionalindicator for correct interpretation of the surveys, and particularly for accurateidentification of FMD-free regions. Since these samplings encompassed popula-tions from various geographic regions, evaluation of potential vaccine interferencepossibly included various manufacturers. Modern vaccines may comprise moreelaborated procedures for antigen concentration and purification, and therefore,continuous evaluation of potential infield interference of vaccines is advisable.
Alternatively to vaccine interference, weak reactivities observed in the I-ELISA-3ABC, particularly for the population over two years old, may representantibody scars from previous infection, or subclinical infection, as indicated byresults from the experimental model as well as after 90 and 900 DPO. In this re-gard, the application of the EITB was critical since it helped elucidate whether serawith T/C values between 0.8–3.0, occurring under the various field situations de-scribed, are or are not derived from animals with previous FMDV infection. This

Serodiagnostic strategy for FMDV surveillance 487
may be accounted for by the ability of the EITB to identify infection-specificantibodies against several viral proteins in a single assay. Our experience overmany years from EITB surveys has provided clear evidence that no animals werefound positive by virus recovery without antibody reactivity against the four anti-gens± 2C, and that single bands in the EITB are not indicative of past FMDVinfection. Use of the EITB reduced significantly the total positivity obtained byI-ELISA. In FMD-free regions without vaccination the percentage of reactivesamples declined from 1.2 to 0, and in FMD-free regions with vaccination from2.1% and 4.8% to 0.026% and 0.3% in cattle under and over two years of age,respectively. In contrast, in sera from animals during outbreaks and after exposureto an episode at 90 and 900 DPO, the EITB confirmed positive in 83.3%, 50.0%and 33.3%, respectively of the I-ELISA-weakly reactive sera.
Application of the EITB as a confirmatory method for suspect or positive I-ELISA responses allowed for the design of a diagnostic approach which is capableof using the I-ELISA with its maximum sensitivity, under the assumption that theconsequent loss of specificity will be resolved by EITB. A similar approach wasreported for the detection of serum antibodies against HTLV I, HTLV II and HIV[15, 18].
In summary, screening by I-ELISA-3ABC together with confirmation of sus-pect and positive samples by EITB is proposed as an adequate and accurateapproach for large-scale seroepidemiological surveys. The performance charac-teristics of the I-ELISA-3ABC validated under the conditions described, as wellas its economic, simple, rapid and easy throughput, which enables the process-ing of many samples, make it adequate as a screening assay to assess residualinfection-specific antibodies. The application of the EITB assay has grown be-yond the initial aim of defining the antibody profile of infection vs. vaccinationstatus. It now serves as a confirmatory test for FMD serosurveillance in cattleunder systematic vaccination.
Since the analytical method applied is indirect, based on the detection ofantibodies as indicators of FMDV exposure, it is not recommended as a stand-alone tool for assessing persistent infection in individual animals, for exampleduring animal trade. In this case, however, it serves as input in the evaluationof risk analysis for selection of import/export animals. A careful assessment ofthe epidemiological situation of the regions from which cattle are to be moved,including confirmation of absence of viral activity by EITB, is essential.
Acknowledgements
The authors thank Dr. José Germán Rodrı́guez Torres for continuous support and fruitfuldiscussions and Mr. PJ Vieira Pereira, LC Amparo dos Santos, A.T Botelho Freire and A.C.Ferraz de Chagas for excellent technical assistance.
References
1. Alonso AF, Gomes I, Bahnemann HG (1988) The induction of antibodies against VIAAin cattle vaccinated and revaccinated with inactivated foot-and-mouth disease vaccine.Bol Cent Panam Fiebre Aftosa 54: 43–50

488 I. E. Bergmann et al.
2. Berger HG, Straub OC, Ahl R, Tesar M, Marquardt O (1990) Identification of foot-and-mouth disease virus replication in vaccinated cattle by antibodies to nonstructural virusproteins. Vaccine 8: 213–216
3. Bergmann IE, Augé de Mello P, Neitzert E, Beck E, Gomes I (1993) Diagnosis of per-sistent aphthovirus infection and its differentiation from vaccination response in cattleby use of enzyme-linked immunoelectrotransfer blot analysis with bioengineered non-structural viral antigens. Am J Vet Res 54: 825–831
4. Bergmann IE, Malirat V (1995) Performance of a rapid enzyme-linked immunoelectro-transfer blot assay for detection of cattle exposed to foot-and-mouth disease virus. BolCentr Panam Fiebre Aftosa 61: 40–44
5. Bergmann IE, Malirat V, Dias LE, Dilandro R (1996) Identification of foot-and-mouthdisease virus-free regions by use of a standardized enzyme-linked immunoelectrotransferblot assay. Am J Vet Res 57: 972–974
6. Bergmann IE, Astudillo V, Malirat V, Neitzert E (1998) Serodiagnostic strategy for esti-mation of foot-and-mouth disease viral activity through highly sensitive immunoassaysusing bioengineered nonstructural proteins. Vet Q 20: S6–S9
7. De Diego M, Brocchi E, Mackay D, De Simone F (1997) The non-capsideal polyprotein3ABC of foot-and-mouth disease virus as a diagnostic antigen in ELISA to differentiateinfected from vaccinated cattle. Arch Virol 142: 2 021–2 033
8. Hamblin C, Barnett I, Hedger R (1986) A new enzyme-linked immunosorbent assay(ELISA) for the detection of antibodies against FMDV. I. Development and method ofELISA. J Immunol Methods 93: 115–121
9. Lubroth J, Brown F (1995) Identification of native foot-and-mouth disease virus non-capsideal protein 2C as a serological indicator to differentiate infected from vaccinatedlivestock. Res Vet Sci 59: 70–78
10. Mackay DKJ, Forsyth MA, Davies PR, Berlinzani A, Belsham GJ, Flint M, Ryan MD(1998) Differentiating infection from vaccination in foot-and-mouth disease using apanel of recombinant, non-capsideal proteins in ELISA. Vaccine 16: 446–459
11. Malirat V, Neitzert E, Bergmann IE, Maradei E, Beck E (1998) Detection of cattleexposed to foot-and-mouth disease virus by means of an indirect ELISA test usingbioengineered nonstructural polyprotein 3ABC. Vet Q 20: S24–S26
12. Meyer RF, Babcock GD, Newman JFE, Burrage TG, Toohey K, Lubroth J, Brown F(1997) Baculovirus expressed 2C of foot-and-mouth disease virus has the potential fordifferentiating convalescent from vaccinated animals. J Immunol Methods 65: 33–43
13. Neitzert E, Beck E, Augé de Mello P, Gomes I, Bergmann IE (1991) Expression of theAphthovirus RNA polymerase gene in Escherichia coli and its use together with otherbioengineered nonstructural antigens in detection of late persistent infection. Virology184: 799–804
14. O’Donnel VK, Boyle DB, Sproat K, Fondevila NA, Forman A, Schudel AA, SmitsaartEN (1996) Detection of antibodies against foot-and-mouth disease virus using a liquid-phase blocking sandwich ELISA (LPBE) with a bioengineered 3D protein. J Vet DiagnInvest 8: 143–150
15. Papadopulos-Eleopulos E, Turner VF, Papadimitriou JM (1993) Is a positive Westernblot proof of HIV infection? Bio/Technology 11: 696–707
16. Pappas MG, Hajkowski R, Hockmeyer WT (1983) Dot-enzyme-linked immunosorbentassay (Dot-ELISA): a microtechnique for the rapid diagnosis of visceral Leishmaniasis.J Immunol Methods 64: 205–214
17. Pinto AA, Garland AJM (1979) Immune response to virus-infection-associated (VIAA)antigen in cattle repeatedly vaccinated with foot-and-mouth disease virus inactivated byformalin or acetylethylenimine. J Hyg 82: 41–50

Serodiagnostic strategy for FMDV surveillance 489
18. Roberts BD, Foung SKH, Lipka JJ, Kaplan JE, Hadlock KG, Reyer GR, Chan L, HeneineW, Khabbaz RF (1993) Evaluation of an immunoblot assay for serological confirmationand differentiation of human T-cell lymphotropic virus types I and II. J Clin Microbiol31: 260–264
19. Rodríguez A, Dopazo J, Sáiz JC, Sobrino F (1994) Immunogenicity of non-capsidealproteins of foot-and-mouth virus: differences between infected and vaccinated swine.Arch Virol 136: 123–131
20. Silberstein E, Kaplan G, Taboga O, Duffy S, Palma E (1997) Foot-and-mouth diseasevirus-infected but not vaccinated cattle develop antibodies against recombinant 3AB1nonstructural protein. Arch Virol 142: 795–805
21. Sørensen KJ, Madsen KG, Madsen ES, Salt JS, Nqindi J, Mackay DKJ (1998) Differ-entiation of infection from vaccination in foot-and-mouth disease by the detection ofantibodies to the non-capsideal proteins 3D, 3AB and 3ABC in ELISA using antigensexpressed in baculovirus. Arch Virol 143: 1 461–1 476
22. Villinger F, Mueller HK, Bruckner L, Ackermann M, Kihm U (1989) Antibodies tofoot-and-mouth disease virus infection associated (VIA) antigen: use of a bioengineeredVIA protein as antigen in an ELISA. Vet Microbiol 20: 235–246
Authors’ address: Dr. I. E. Bergmann, Pan-American Foot-and-Mouth Disease Center((PAHO/WHO), Caixo Postal 589, 20001, Rio de Janeiro, Brazil.
Received May 3, 1999