identity and typification of navicula hasta (bacillariophyceae
TRANSCRIPT

Identity and typification of Navicula hasta(Bacillariophyceae)Zlatko Levkov,1* Mark B. Edlund2 and Teofil Nakov1
1Institute of Biology, Faculty of Natural Sciences, 1000 Skopje, Macedonia, and 2St. Croix Watershed Research StationScience Museum of Minnesota, Marine on St. Croix, Minnesota 55047, USA
SUMMARYThe name Navicula hasta is regularly applied to large,heavily silicified Navicula specimens with lanceolateoutlines, protracted ends, and relatively coarse, radiate,and lineolate striae that are more irregularly spacedaround the central area. As such, Navicula hasta hasbeen reported from numerous localities such as brack-ish water fossil sediments, oligotrophic ancient lakesand postglacial lakes. When the data for distribution ofNavicula hasta are summarized, this species appears tobe geographically and temporally widespread with abroad range of ecologic tolerances. Published reportsfor size range show that diatomists during the lastcentury have differing concepts of the species startingwith the disagreement between the protologue and oneof the figures given by Pantocsek. Furthermore, aninformal ‘type’ of Navicula hasta reported in 1980 doesnot match the protologue and is not unambiguouslybased on material Pantocsek had in his hands whendescribing his new species. We studied publishedreports of Navicula hasta and investigated materialreported to contain Navicula hasta and its allies fromthe type locality (Köpecz), one of the fossil depositsreported to contain N. hasta (Bodos) and from ancientand other younger lakes worldwide. From our analyses,we document the variation and underappreciated diver-sity within the Navicula hasta group and set the stagefor formal recognition of new species in subsequentpapers. Herein, we formally propose a lectotype fromamong the original illustrations and designate anepitype for Navicula hasta based on Pantocsek’sKöpecz material. Additionally, two new species, Nav-icula krenneri spec. nov. and Navicula pseudohastaspec. nov. are described and a lectotype of Naviculasuperhasta Lange-Bertalot and Metzeltin is proposed.
Key words: Bodos, diatom, distribution, epitype,Köpecz, lectotype, Navicula hasta, Romania,typification.
INTRODUCTION
Navicula hasta Pantocsek was named and illustratedfrom Tertiary fossil deposits from Köpecz, Transylvania
(Pantocsek 1892, fig. 5: 74, 14: 213), while thedescription was given 13 years later (Pantocsek 1905,p. 69) which listed the distribution in ‘Bibarczfalva,Köpecz et Bodos in Transilvania’. However, measure-ments of the second illustration (Pantocsek 1892,fig. 14: 213) do not entirely fit Pantocsek’s descriptionof the species. The first illustration is 71 mm long and15 mm wide (Fig. 1), while the second figure illustratedis 85 mm long and 19 mm wide (Fig. 2); these measure-ments are outside the size range given in the descrip-tion. The problem has been further confounded by theinformal identification of the ‘type’ for N. hasta byKrenner (1980), who chose Kinker’s slide no. 122(discussed by Wojtal & Buczko 2004) from Bodos sug-gesting it contained ‘Pantocsek’s original type species’.This slide is not attributable to Pantocsek, Bodos is notthe type locality for N. hasta, nor does the specimen(Fig. 3) on this slide correspond to Pantocsek’sprotologue.
Since the original description, the name N. hastahas been applied to many specimens from throughoutthe world (Table 1). Part of these records originate fromdifferent fossil deposits in Romania (Hustedt 1934 inSchmidt et al. (1874–1959)), Portugal (Silva 1946),Sweden (Cleve-Euler 1953), and Ethiopia (Gasse1980), while the others are recent populations fromgeologically old lakes such as Aoki (Haraguchi 1997),Biwa (Skvortzow 1937; Ohtsuka & Tuji 2002), Baikal(Skvortzow 1936; Zabelina et al. 1951), Tanganyika(Cocquyt 1998), Ohrid (Jurilj 1954), Hövsgöl (Edlundet al. 2001) and other younger lakes such as LakeGeorge, Eastern Australia (Foged 1978), Lake Fodler,Tasmania (Hodgson et al. 1997), Laurentian GreatLakes (Stoermer et al. 1999) and Lake Begnas, Nepal(Rai 2000).
The aims of this project are to conduct a detailedrevision of populations of the N. hasta-complex withspecial emphasis on locations with high, cryptic, or
*To whom correspondence should be addressed.Email: [email protected] editor: S. Mayama.Received 7 December 2006; accepted 31 August 2007.doi: 10.1111/j.1440-1835.2008.00484.x
Phycological Research 2008; 56: 46–57

undiscovered diversity. In this first paper, we will sum-marize the existing data on distribution and taxonomyof N. hasta and its allies with special attention to theproblems surrounding the original and later circum-scriptions of this taxon. We will restrict our presenta-tion, however, to N. hasta populations from fossildeposits (Köpecz, Bodos and Rumänia) including thetype locality and related localities.
History and distribution of thenavicula hasta-complex
As noted in the introduction, confusion surroundingN. hasta (Table 1) began with the incongruence of Pan-tocsek’s illustrations (Pantocsek 1892) and protologue(Pantocsek 1905). Hustedt (1930, p. 306, fig. 541)broadened the concept of N. hasta compared with Pan-tocsek (1905) to generally accommodate longer valves.
Furthermore, in Schmidt et al. (1874–1959, fig. 395:10–15), Hustedt depicts two populations of N. hastafrom fossil deposits in Romania and Lake Aoki, thatdiffer by size and general outline of the valve.
Jurilj (1954), in his observation on N. hasta fromLake Ohrid, Macedonia, adopts Hustedt’s (1930)concept of the species, but doesn’t illustrate N. hasta.Jurilj (1954) compares Hustedt’s (1930) measure-ments to his newly described N. hastata Jurilj (Jurilj1954; p.139), one of the first taxa segregated from theN. hasta-complex.
Cleve-Euler (1953, p. 135) had a broader concept ofN. hasta, with a valve length of 57–130 mm and widthof 13–29 mm (figs 774a–e). Illustrations (figs 774a–e)presented by Cleve-Euler also have different patterns inthe central and axial areas than Pantocsek’s illustra-tions. Gasse (1980) gives a record of N. hasta forPlio-Pleistocene fossil deposits in Ethiopia. Shedescribes and illustrates narrower specimens comparedwith Pantocsek’s (1892) concept with differences inwidth of 11 mm versus 14–18 mm.
Lange-Bertalot (2001, p. 37) further expanded theconcept of N. hasta with regard to morphometric fea-tures and briefly discussed the taxonomic problemaround N. hasta. According to Lange-Bertalot (2001,p. 39) there are two groups with similar morphology,striae and lineola density, but they differ in size, espe-cially breadth (20–25 mm vs 15–20 mm).
Wojtal and Buczko (2004), using material fromPoland, Pantocsek’s material from Bodos, and slide122 (Budapest Natural History Museum), also givedifferent morphometric characteristics for N. hastathat accommodate larger, broader and coarser valves(Table 1).
MATERIALS AND METHODS
Pantocsek’s original material was obtained from theHungarian Natural History Museum (HNHM) (BP 87,88) and was analyzed for the presence of N. hastaspecimens. Sample no. 88 contained a higher numberof valves of N. hasta sensu lato and was further used inthe analysis of this complex in Köpecz. All valves pre-sented here (Figs 4–8 and 10–19) originate from thissample.
In addition to Pantocsek’s type material and pub-lished reports (Table 1), 29 collections were examinedin the present study. Several slides were prepared ofmaterial mounted in Naphrax (Brunel Microscopes Ltd.,Wiltshire, UK) or Zrax (MicrAP Enterprises, Philadel-phia, Pennsylvania, USA). Slides are deposited in theDiatom Collection at the Institute of Biology, Faculty ofNatural Sciences in Skopje, Macedonia. Additionally,slides and samples from the Alfred Wegener Institutefor Polar and Marine Research (Bremerhaven; BRM),Natural History Museum (London; BM), California
Fig. 1–3. Illustrations of Navicula hasta Pantocsek sensu lato.
Scale bar = 10 mm. 1,2. Pantocsek’s illustrations (images modi-
fied from Pantocsek 1892; fig. 5: 74, 14: 213). 1. Lecto-
type. 3. N. krenneri Levkov, Edlund & Nakov spec. nov.
(=N. hasta sensu Krenner) from slide BP 122, Hungarian Natural
History Museum (HNHM).
47Identity of Navicula hasta
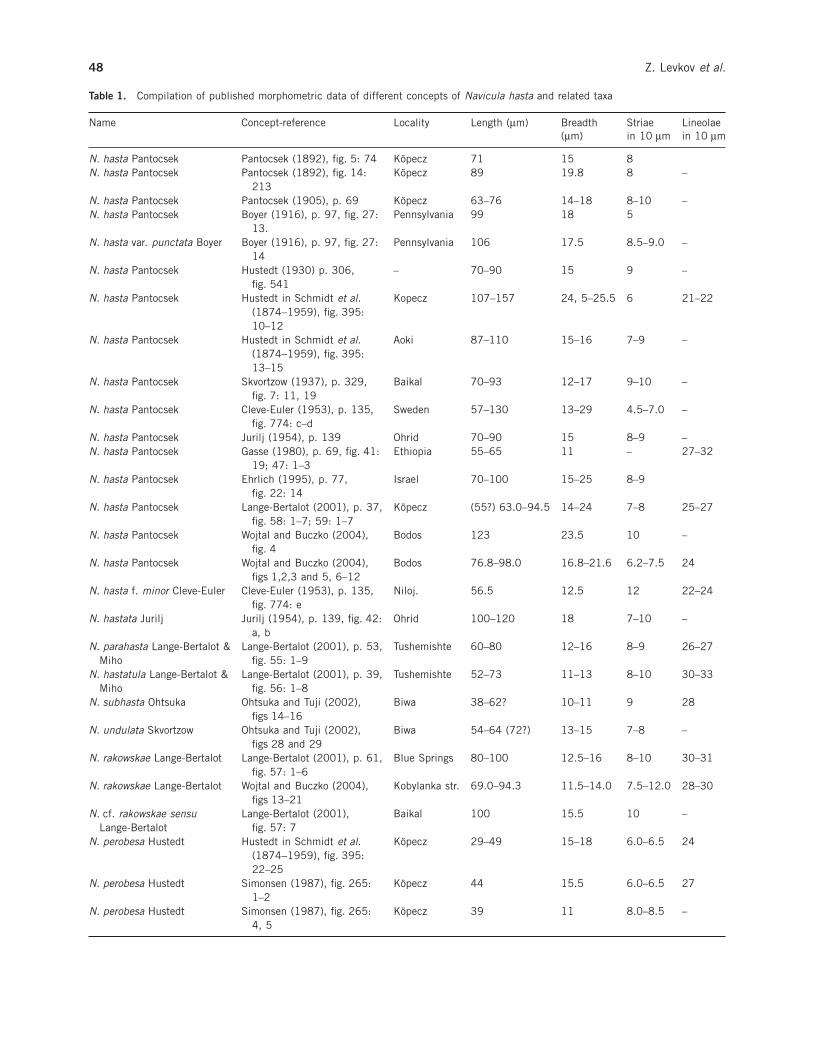
Table 1. Compilation of published morphometric data of different concepts of Navicula hasta and related taxa
Name Concept-reference Locality Length (mm) Breadth(mm)
Striaein 10 mm
Lineolaein 10 mm
N. hasta Pantocsek Pantocsek (1892), fig. 5: 74 Köpecz 71 15 8N. hasta Pantocsek Pantocsek (1892), fig. 14:
213Köpecz 89 19.8 8 –
N. hasta Pantocsek Pantocsek (1905), p. 69 Köpecz 63–76 14–18 8–10 –N. hasta Pantocsek Boyer (1916), p. 97, fig. 27:
13.Pennsylvania 99 18 5
N. hasta var. punctata Boyer Boyer (1916), p. 97, fig. 27:14
Pennsylvania 106 17.5 8.5–9.0 –
N. hasta Pantocsek Hustedt (1930) p. 306,fig. 541
– 70–90 15 9 –
N. hasta Pantocsek Hustedt in Schmidt et al.(1874-1959), fig. 395:10–12
Kopecz 107–157 24, 5–25.5 6 21–22
N. hasta Pantocsek Hustedt in Schmidt et al.(1874-1959), fig. 395:13–15
Aoki 87–110 15–16 7–9 –
N. hasta Pantocsek Skvortzow (1937), p. 329,fig. 7: 11, 19
Baikal 70–93 12–17 9–10 –
N. hasta Pantocsek Cleve-Euler (1953), p. 135,fig. 774: c–d
Sweden 57–130 13–29 4.5–7.0 –
N. hasta Pantocsek Jurilj (1954), p. 139 Ohrid 70–90 15 8–9 –N. hasta Pantocsek Gasse (1980), p. 69, fig. 41:
19; 47: 1–3Ethiopia 55–65 11 – 27–32
N. hasta Pantocsek Ehrlich (1995), p. 77,fig. 22: 14
Israel 70–100 15–25 8–9
N. hasta Pantocsek Lange-Bertalot (2001), p. 37,fig. 58: 1–7; 59: 1–7
Köpecz (55?) 63.0–94.5 14–24 7–8 25–27
N. hasta Pantocsek Wojtal and Buczko (2004),fig. 4
Bodos 123 23.5 10 –
N. hasta Pantocsek Wojtal and Buczko (2004),figs 1,2,3 and 5, 6–12
Bodos 76.8–98.0 16.8–21.6 6.2–7.5 24
N. hasta f. minor Cleve-Euler Cleve-Euler (1953), p. 135,fig. 774: e
Niloj. 56.5 12.5 12 22–24
N. hastata Jurilj Jurilj (1954), p. 139, fig. 42:a, b
Ohrid 100–120 18 7–10 –
N. parahasta Lange-Bertalot &Miho
Lange-Bertalot (2001), p. 53,fig. 55: 1–9
Tushemishte 60–80 12–16 8–9 26–27
N. hastatula Lange-Bertalot &Miho
Lange-Bertalot (2001), p. 39,fig. 56: 1–8
Tushemishte 52–73 11–13 8–10 30–33
N. subhasta Ohtsuka Ohtsuka and Tuji (2002),figs 14–16
Biwa 38–62? 10–11 9 28
N. undulata Skvortzow Ohtsuka and Tuji (2002),figs 28 and 29
Biwa 54–64 (72?) 13–15 7–8 –
N. rakowskae Lange-Bertalot Lange-Bertalot (2001), p. 61,fig. 57: 1–6
Blue Springs 80–100 12.5–16 8–10 30–31
N. rakowskae Lange-Bertalot Wojtal and Buczko (2004),figs 13–21
Kobylanka str. 69.0–94.3 11.5–14.0 7.5–12.0 28–30
N. cf. rakowskae sensuLange-Bertalot
Lange-Bertalot (2001),fig. 57: 7
Baikal 100 15.5 10 –
N. perobesa Hustedt Hustedt in Schmidt et al.(1874-1959), fig. 395:22–25
Köpecz 29–49 15–18 6.0–6.5 24
N. perobesa Hustedt Simonsen (1987), fig. 265:1–2
Köpecz 44 15.5 6.0–6.5 27
N. perobesa Hustedt Simonsen (1987), fig. 265:4, 5
Köpecz 39 11 8.0–8.5 –
48 Z. Levkov et al.

Academy of Sciences (San Francisco; CAS), AdolfCaljon Burundi Algae Collection (Meise, Belgium: BR),Academy of Natural Sciences of Philadelphia (ANSP),and the Lake Biwa Museum (Kusatsu-shi, Shiga), wereexamined on loan.
RESULTS
Typification and revised metrics ofNavicula hasta Pantocsek
As a starting point for resolving the systematic, taxo-nomic and biogeographic problems surrounding the‘Navicula hasta complex’, it is necessary to propose alectotype and epitype for N. hasta from Pantocsek’s(1892) original illustrations and material.
Krenner (1980), working on Pantocsek’s collection,identified a ‘type’ for N. hasta on slide 122 in theBudapest Museum of Natural History, suggesting itcontained ‘Pantocsek’s original type species’. This slidewas prepared by Kinker from Bodos material, one of thelocalities noted by Pantocsek (1905) to contain N.hasta. This specimen cannot represent the holotype assuggested by Wojtal and Buczko 2004), because Kink-er’s Slide 122 cannot be unambiguously attributed tohave been in Pantocsek’s hands, the specimen (Fig. 3)on Slide 122 assigned as the ‘type’ of N. hasta(Krenner 1980) does not correspond to Pantocsek’s
description of the species according to morphometricfeatures and central area pattern, and Bodos is not thetype locality for N. hasta.
Our analyses of Köpecz samples attributable to Pan-tocsek show that there are four species that are closelyrelated to N. hasta. We consider that the species pre-sented in Figures 4–9 is the most appropriate entity fortypification of N. hasta, because of its similarity toPantocsek’s (1892) illustrations. Because Pantocsektypified N. hasta using two illustrations, we must firstpropose a lectotype for N. hasta. We conclude that bothillustrations unambiguously belong to N. hasta sensustricto as a part of a diminution series; although Pan-tocsek’s second illustration cannot fit into the proto-logue (it is too large). Therefore we propose Pantocsek’sfirst illustration as the lectotype as it most closelyadheres to the protologue. However, valves with largersize and comparable morphology to both of Pantocsek’sillustrations were observed in Köpecz (Figs 4–8) andBodos samples (Fig. 9) and we conclude that thisspecies is present in at least two of the three localitieslisted by Pantocsek (1905). Finally, we suggest abroader range of length and breadth is appropriate forthe description of N. hasta s.s. length 60–105 mm andwidth 14.0–20.5 mm; striae 6.5–7.0 in 10 mm andlineolae 24–26 in 10 mm.
We further propose an epitype for N. hasta using aspecimen from Köpecz. The designation of an epitype
Fig. 4–9. Navicula hasta sensu
stricto. Scale bar = 10 mm. 4–8.
Slide Diatom Collection at Institute
of Biology (DCIBS) 001023,
Köpecz material BP 88 (HNHM),
Köpecz. 8. Epitype. 9. Bodos mat-
erial BP 27 (Hungarian Natural
History Museum (HNHM)).
49Identity of Navicula hasta
will allow us to begin a systematic revision of otherworldwide populations reported as N. hasta, and super-sede Krenner’s (1980) informally designated ‘type’(Fig. 3).
Navicula hasta Pantocsek, Beiträge zur Kenntnis derfossilen Bacillarien Ungarns. III. Süßwasser Bacilla-rien, T. V: fig. 74 and T. XIV: fig. 213. 1892.
Lectotypus (designated here): [icon] T. V: fig. 74 inPantocsek, Beiträge zur Kenntnis der fossilen Bacil-larien Ungarns. III. Süßwasser Bacillarien, 1892.(here reproduced as Fig. 1).
Locus typicus: Köpecz, Transsylvania, Rumania, fossildeposit.
Epitypus (designated here): Preparation 001023 fromKöpecz material (No. BP 88 from the HNHM) depos-ited in Diatom Collection at Institute of Biology,Faculty of Natural Sciences in Skopje (the valverepresenting the epitype is Fig. 8).
Isoepitypus: Preparation BP 1052 in HNHM.
Köpecz
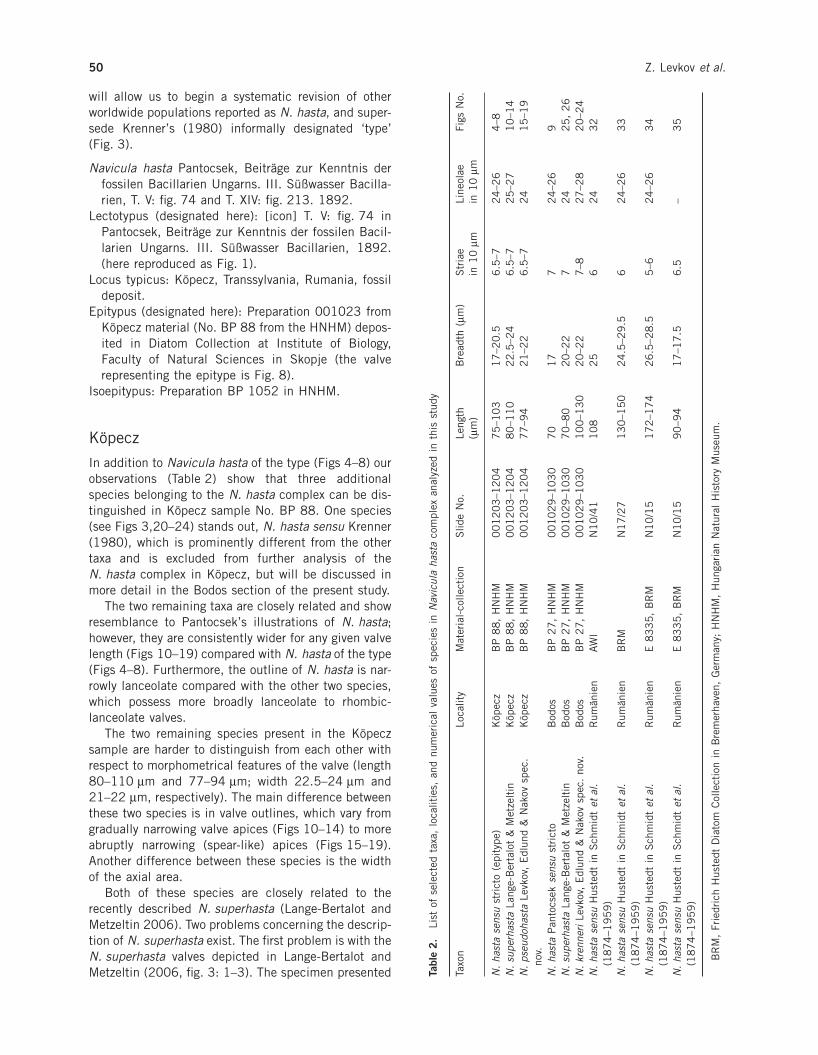
In addition to Navicula hasta of the type (Figs 4–8) ourobservations (Table 2) show that three additionalspecies belonging to the N. hasta complex can be dis-tinguished in Köpecz sample No. BP 88. One species(see Figs 3,20–24) stands out, N. hasta sensu Krenner(1980), which is prominently different from the othertaxa and is excluded from further analysis of theN. hasta complex in Köpecz, but will be discussed inmore detail in the Bodos section of the present study.
The two remaining taxa are closely related and showresemblance to Pantocsek’s illustrations of N. hasta;however, they are consistently wider for any given valvelength (Figs 10–19) compared with N. hasta of the type(Figs 4–8). Furthermore, the outline of N. hasta is nar-rowly lanceolate compared with the other two species,which possess more broadly lanceolate to rhombic-lanceolate valves.
The two remaining species present in the Köpeczsample are harder to distinguish from each other withrespect to morphometrical features of the valve (length80–110 mm and 77–94 mm; width 22.5–24 mm and21–22 mm, respectively). The main difference betweenthese two species is in valve outlines, which vary fromgradually narrowing valve apices (Figs 10–14) to moreabruptly narrowing (spear-like) apices (Figs 15–19).Another difference between these species is the widthof the axial area.
Both of these species are closely related to therecently described N. superhasta (Lange-Bertalot andMetzeltin 2006). Two problems concerning the descrip-tion of N. superhasta exist. The first problem is with theN. superhasta valves depicted in Lange-Bertalot andMetzeltin (2006, fig. 3: 1–3). The specimen presented Tabl
e2
.Li
stof
sele
cted
taxa
,lo
calit
ies,
and
num
eric
alva
lues
ofsp
ecie
sin
Nav
icul
aha
sta
com
plex
anal
yzed
inth
isst
udy
Taxo
nLo
calit
yM
ater
ial-
colle
ctio
nS
lide
No.
Leng
th(m
m)
Bre
adth
(mm
)S
tria
ein
10
mmLi
neol
aein
10
mmFi
gsN
o.
N.ha
sta
sens
ust
rict
o(e
pity
pe)
Köp
ecz
BP
88
,H
NH
M0
01
20
3–1
20
47
5–1
03
17
–20
.56
.5–7
24
–26
4–8
N.su
perh
asta
Lang
e-B
erta
lot
&M
etze
ltin
Köp
ecz
BP
88
,H
NH
M0
01
20
3–1
20
48
0–1
10
22
.5–2
46
.5–7
25
–27
10
–14
N.ps
eudo
hast
aLe
vkov
,E
dlun
d&
Nak
ovsp
ec.
nov.
Köp
ecz
BP
88
,H
NH
M0
01
20
3–1
20
47
7–9
42
1–2
26
.5–7
24
15
–19
N.ha
sta
Pan
tocs
ekse
nsu
stri
cto
Bod
osB
P2
7,
HN
HM
00
10
29
–10
30
70
17
72
4–2
69
N.su
perh
asta
Lang
e-B
erta
lot
&M
etze
ltin
Bod
osB
P2
7,
HN
HM
00
10
29
–10
30
70
–80
20
–22
72
42
5,
26
N.kr
enne
riLe
vkov
,E
dlun
d&
Nak
ovsp
ec.
nov.
Bod
osB
P2
7,
HN
HM
00
10
29
–10
30
10
0–1
30
20
–22
7–8
27
–28
20
–24
N.ha
sta
sens
uH
uste
dtin
Sch
mid
tet
al.
(18
74
-19
59
)R
umän
ien
AWI
N1
0/4
11
08
25
62
43
2
N.ha
sta
sens
uH
uste
dtin
Sch
mid
tet
al.
(18
74
-19
59
)R
umän
ien
BR
MN
17
/27
13
0–1
50
24
.5–2
9.5
62
4–2
63
3
N.ha
sta
sens
uH
uste
dtin
Sch
mid
tet
al.
(18
74
-19
59
)R
umän
ien
E8
33
5,
BR
MN
10
/15
17
2–1
74
26
.5–2
8.5
5–6
24
–26
34
N.ha
sta
sens
uH
uste
dtin
Sch
mid
tet
al.
(18
74
-19
59
)R
umän
ien
E8
33
5,
BR
MN
10
/15
90
–94
17
–17
.56
.5–
35
BR
M,
Frie
dric
hH
uste
dtD
iato
mC
olle
ctio
nin
Bre
mer
have
n,G
erm
any;
HN
HM
,H
unga
rian
Nat
ural
His
tory
Mus
eum
.
50 Z. Levkov et al.

in fig. 3: 1 is closely related to N. hasta sensu Krennerfrom Bodos (Figs 20–24). This is supported with thestriae pattern of the central area where some irregularlyshortened striae can be observed and the striae densitymidway between the center and ends of the valve (8–9in 10 mm) exceeds the stria density range provided(‘about 6 in 10 mm’) in the protologue of N. superhasta(Lange-Bertalot & Metzeltin 2006; p. 61). Lange-Bertalot and Metzeltin’s (2006) fig. 3: 2–3 resembleN. hasta sensu Hustedt 1934 in Schmidt et al. (1874–1959) and Köpecz specimens presented in this studyas Figures 10–14, which are characterized by widelyspaced central striae, 6 in 10 mm, in contrast to thefirst specimen illustrated by Lange-Bertalot and Met-
zeltin (2006, fig. 3: 1). Conspecificity of the specimensof N. hasta sensu Krenner (Figs 20–24) and N. hastasensu Hustedt (Fig. 33) based on morphological char-acteristics is very unlikely and the conspecificity of thethree specimens illustrated in Lange-Bertalot and Met-zeltin (2006; fig. 3: 1–3) is questionable. We concludethat the specimens typifying N. superhasta are not rep-resentative of a single morphospecies.
A second problem is found in the synonymy given forN. superhasta. According to Lange-Bertalot and Metzel-tin (2006, p. 59) the second illustration of N. hasta(Pantocsek 1892; fig. 14: 213) is ‘approximately108 mm long and 25 mm broad’. These measurementsare incorrect and cannot be associated with the speci-
Fig. 10–19. Navicula hasta sensu
lato. Scale bar = 10 mm. 10–14.
N. superhasta Lange-Bertalot & Met-
zeltin from slide Diatom Collection
at Institute of Biology (DCIBS)
001023, material BP 88 (Hungarian
Natural History Museum (HNHM)),
Köpecz. 15–19. N. pseudohasta
Levkov, Edlund & Nakov spec. nov.
from slide DCIBS 001023, mat-
erial BP 88 (HNHM), Köpecz. 17.
Holotype.
51Identity of Navicula hasta

men illustrated by Pantocsek (1892, fig. 14: 213), butthey can be associated with Hustedt 1934 in Schmidtet al. (1874–1959) drawing (fig. 395: 11). The correctmeasurements of Pantocsek’s (1892) fig. 14: 213 arec. 89 mm long and 19.8 mm broad. Pantocsek’s (1892)fig. 14: 213 can not be treated as part of N. superhastaaccording to the protologue given by Lange-Bertalot andMetzeltin (2006), and it should not be considered as asynonym of N. superhasta.
As a result of our observations, a formal second steplectotypification of N. superhasta is carried out andN. pseudohasta spec. nov. is newly described.
Navicula superhasta Lange-Bertalot and Metzeltin(2006)
Lectotype (designated here): the single individualspecimen on preparation no. 400/11b in Coll. Hustedt,Bremerhaven, depicted in fig. 3: 2 in Lange-Bertalotand Metzeltin (2006).
Navicula pseudohasta Levkov, Edlund & Nakov spec.nov. Figures 15–19.
Descriptio: Valvae late lanceolatae apicibus abrupteattenuatis, 77–94 mm longae, 21–22 mm latae. Raphelateralis, poris centralibus distinctis. Area axialismodice angusta, linearis, area centralis parva vel inter-dum indistinctis. Striae transapicales radiantes 6.5–7.5 in 10 mm, mediis distantius positis, crassepunctatae. Lineolae striarum, distincte discernandae inmicroscopio photonico, 24–26 in 10 mm.
Holotype: Slide no. 001203 Diatom Collection at Insti-tute of Biology in Skopje (DCIBS), specimen repre-senting the holotype is Figure. 17.
Isotypes: Slide no. 001204 DCIBS, ZU6/31 in BRM;BP 1052 in HNHM.
Type locality: Köpecz, Neogene deposits.Original material: BP 88, HNHM.Etymology: the species is named as a false N. hasta.
Description: Valves broadly lanceolate, ends abruptlynarrowing (spear-like), valve length 77–94 mm, valvebreadth 21–22 mm. Raphe lateral with distinct proxi-mal raphe ends. Axial area linear moderately narrow,
Fig. 20–24. Navicula krenneri
Levkov, Edlund & Nakov spec. nov.
from material BP 27, Bodos. Scale
bar = 10 mm. 21. Holotype.
52 Z. Levkov et al.

central area small round or sometimes indistinct. Striaeradial throughout, more widely spaced in the middle6.5–7.5 in 10 mm, coarsely punctuated. Lineolaeclearly visible in LM, 24–26 in 10 mm.
Differential diagnosis: N. pseudohasta can be differ-entiated from N. superhasta by width of axial area.N. superhasta always possesses a wider, more irregular-shaped axial area, whereas N. pseudohasta possesses anarrower, more linear axial area. Another difference isthe shape of the valve apices (gradually narrowing inN. superhasta versus abruptly narrowing in N.pseudohasta). N. hastata Jurilj although possessingspear-like apices, is always narrower (16–18 mm) thanN. superhasta and N. pseudohasta.
Bodos
Navicula hasta was also recorded from material thatoriginated from Bodos (Pantocsek 1905). Wojtal andBuczko (2004) located Pantocsek’s Bodos material in
the Budapest Museum of Natural History (AccessionNo. BP 27 in HNHM). Analysis of slides made fromBodos material shows the presence of several taxa thatbelong to the ‘hasta’ complex. The first taxon observedin Bodos (Figs 20–24) is very close to the ‘type’ chosenby Krenner (1980), here depicted in Figure 3, whereasother Bodos specimens (Figs 25,26) resemble N.superhasta (Figs 10–14). It is unlikely that these twoBodos taxa are conspecific, due to large differencesin valve and central area morphology and numericalfeatures. We argue that N. hasta sensu Krenner(1980; here as Fig. 3) and valves depicted here asFigures 20–24 are conspecific and do not representN. hasta in the strict sense, but are closely related to (ifnot the same as) Navicula (?nov) spec. sensu Lange-Bertalot (2001; fig. 60: 1) or Navicula superhasta proparte (fig. 3: 1 in Lange-Bertalot & Metzeltin 2006).Bearing in mind the above-mentioned difficulties in thedescription of N. superhasta and the need to avoidany further confusion regarding taxa sympatric with
Fig. 25–32. Different Navicula
hasta related species from material
BP27, Bodos. Scale bar = 10 mm.
25,26. N. superhasta Lange-
Bertalot & Metzeltin. 27,28. Navi-
cula sp. 29. Navicula perobesa
Hustedt sensu lato. 30–32. Navi-
cula perobesa Hustedt.
53Identity of Navicula hasta

N. hasta, we feel it is most appropriate to describe thismorphodeme as a new species.
Navicula krenneri Levkov, Edlund & Nakov spec. nov.Figures 3, 20–24
Synonym: Navicula hasta Pantocsek sensu Wojtal andBuczko (2004), partim fig. 4
Synonym: Navicula superhasta Lange-Bertalot and Met-zeltin (2006), partim fig. 3: 1
Descriptio: Valvae lanceolatae ad anguste lanceolatae,apicibus gradatim attenuatis, longitudo 100–130 mm,latitudo 20–22 mm. Raphe distincte lateralis, sternoraphis lato distincte conturato. Poris centralibus dis-tinctis et dilatatis. Area axialis linearis, gradatimattenuatis ad apicibus. Area centralis parva et irregu-lariter transverse dilatata. Striae transapicales radi-antes et irregulariter abbreviates ad aream centralem,7–9 in 10 mm. Lineolae striarum discernandae inmicroscopio photonico, 27–28 in 10 mm.
Holotype: Slide no. 001029 DCIBS, specimen repre-senting the holotype is Figure 21.
Isotypes: Slide no. 001030 DCIBS, ZU6/32 in BRM,and BP 1053 in HNHM.
Type locality: Bodos, Neogene deposits.Original material: BP 27, HNHM.Etymology: the species is dedicated to J. A. Krenner.
Description: Valves lanceolate to narrowly lanceolate,ends gradually narrowing. Valve length 100–130 mm,breadth 20–22 mm. The raphe is distinctly lateralaccompanied by a strongly contoured sternum. Proxi-mal raphe ends large, clearly expanded. The axial areais linear tapering to the ends. The central area is small,transapically irregularly widened. The stria are radiatethroughout and shortened in irregular fashion in thecentral area, 7–9 in 10 mm. Lineolae discernible withlight microscopy, 27–28 in 10 mm.
Additionally, two other taxa that belong to the ‘hasta’complex are present in the Bodos sample (Figs 27–32).The valves depicted in Figures 27 and 28 closelyresemble Navicula spec (?nov) sensu Lange-Bertalot(2001; fig. 62: 4, 5), and differ from N. hasta sensustricto by having smaller valves and wider spaced lin-eolae. The valves depicted in Figures 29–32 resembleN. perobesa Hustedt sensu Lange-Bertalot.
Rumänien/Romania
Apart from the original material, and to facilitate com-parison and further characterization of the ‘hastacomplex’, we also observed slides from other fossildeposits from Romania that have been reported tocontain N. hasta (Hustedt 1934 in Schmidt et al.1874–1959). Analyses of these slides show the pres-ence of a few other Navicula taxa that more or lessresemble N. hasta. On the slide labeled as ‘N. 10/41.
Rumänien’ with mark ‘Navicula hasta Pant’ (BRM –Acc. no. 10/41) there is only one valve and it is about108 mm long and 25 mm wide (Fig. 33). This valve is inconcordance with Hustedt’s concept of N. hasta proparte (Hustedt 1934 in Schmidt et al. 1874–1959)from populations in Romanian fossil deposits. We con-clude that this valve (Fig. 34) belongs to the diminutionseries of N. superhasta observed in Pantocsek’s Köpeczsample.
On the slide labeled Rumänien (BRM – Acc. No.N17/27), a taxon with lanceolate outline, length about150 mm and breadth 24.5–29.5 mm is found (Fig. 34).According to valve shape and structure the taxon isclose to N. hasta pro parte sensu Hustedt (1934 inSchmidt et al. 1874–1959, fig. 395: 10). On the slidelabeled N. 10/15, Rumänien 5 Oberdaz 1957 withmark ‘Navicula hasta Pant, N. platystoma E.’ two dif-ferent taxa that resemble N. hasta were observed. Thefirst taxon, with length of 172–174 mm and width of
Fig. 33–36. Navicula hasta sensu lato from Rumänia. 33.
N. superhasta Lange-Bertalot & Metzeltin, from slide N. 10/41
Bremerhaven (BRM). 34. N. hasta sensu lato, fossil, from slide
N17/27 BRM. 35. N. hasta sensu lato fossil, from slide N.
10/15 BRM. 36. N. hasta sensu lato from slide N. 1015 BRM.
54 Z. Levkov et al.

26.5–28.5 mm (Fig. 35), is different than Hustedt’s(1930), Lange-Bertalot’s (2001), and the Levkov et al.(2007) concept of N. hasta, but it is very similar tovalves of N. hasta pro parte sensu Hustedt (1934 inSchmidt et al. 1874–1959) and the valve depicted inFigure 34. Both valves (Figs 34 and 35) share similarmorphometrical features and it is possible that theybelong to the same species.
The second species on BRM N. 10/15 has lan-ceolate outline with protracted ends and length90–94 mm and breadth 16–17 mm (Fig. 36). Accordingto morphometric features, this species is similar toHustedt’s (1930, fig. 541) concept, but it has a slightlydifferent outline that is closer to N. rakowskae Lange-Bertalot emend. Levkov & Nakov (in Levkov et al.2007) and Navicula (?nov) spec. sensu Lange-Bertalot(2001, fig. 62: 4, 5). This taxon should be treated aspart of the ‘N. hasta-complex’, but likely belongs toanother species separate from N. hasta sensu stricto.
DISCUSSION
In general, our concept of N. hasta includes large,heavily silicified Navicula sensu stricto that have pro-tracted ends, lateral raphe branches, and relativelycoarse striae that are radiate near the central area andbecome parallel only near the valve ends. The striae areless regularly and more distantly spaced around thecentral area and are usually clearly lineolate in the lightmicroscope. It is this group of taxa that we consider tobe members of the ‘N. hasta-complex’. Although theabove description of the complex circumscribes mostspecimens assigned by authors to N. hasta, there isclearly confusion surrounding Pantocsek’s originalconcept of this taxon and the treatment of N. hasta bysubsequent authors. Furthermore, when populationsof ‘N. hasta’ from around the world are more closelystudied, it is clear that this complex contains manyforms that are morphologically, ecologically, and/or geo-graphically discrete. Earlier authors recognized a fewmembers of the N. hasta-complex as entities deservingspecies status (Hustedt 1934; Skvortzow 1936, 1937;Jurilj 1954), but only recently has the diversity ofthe N. hasta-complex been further scrutinized (Lange-Bertalot 2001; Ohtsuka & Tuji 2002). These recentworks have answered some questions but raised addi-tional problems. There has been a significant influx ofnames in this group (Lange-Bertalot 2001; Lange-Bertalot & Metzeltin 2006; Levkov et al. 2007) andsome of them require more attention and work. This isin part due to the unresolved typification of N. hasta,which is undoubtedly at the base of this continuingpuzzle.
More recently Levkov et al. (2007), shed new lighton the diversity of this complex in ancient lakes Ohridand Prespa. Although, several new N. hasta-complex
forms have been described (N. hastata Jurilj, N. para-hasta Lange-Bertalot & Miho, N. hastatula Lange-Bertalot & Miho, N. subhastatula Levkov & Metzeltinand N. krsticii Levkov), the presence of N. hasta sensustricto (reported by Jurilj 1954) has not been con-firmed. Interestingly and contrary to the presumablyextinct N. hasta s.s., other taxa primarily describedfrom Köpecz (e.g. Navicula turris Pantocsek) haveremained living in Lake Ohrid. The situation has beenconfirmed in the opposite direction also. Navicula rein-hardtii var. rhomboelliptica Levkov, Krstic & Nakov wasdescribed recently as living from lakes Ohrid andPrespa and has also been observed in Köpecz (Levkovand Nakov 2007 unpubl. data).
Members of the Navicula hasta species complexhave been reported from all continents except SouthAmerica and Antarctica. They have broad temporal rep-resentation in the fossil record in Tertiary to Recentdeposits. They also have apparent broad ecologic toler-ances, especially along salinity gradients, and havebeen reported from oligotrophic to brackish habitats.
Foged’s (1978, fig. 28: 4) report of N. hasta forEastern Australia has to be excluded as a misidentifi-cation. The image presented clearly represents a ‘heri-baudii’ form of a Craticula species. Hodgson et al.’s(1997) identification of N. hasta in Lake Fodler, Tas-mania is also questionable. After examination of loanedmaterial, no ‘hasta-like’ specimens were observed andthe large celled Navicula species present in the mate-rial is probably more closely related to N. peregrina(Ehrenberg) Kützing (for further details see Levkovet al. 2007; fig. 220: 1–4). Cocquyt (1998) alsoreports N. hasta for Lake Tanganyika. After examinationof slides from the ACBUA collection we have failed todocument any N. hasta sensu stricto specimens or anyof the more closely related European morphodemes.The valves present in Tanganyika, although superficiallyresembling N. hasta differ with respect to outline, striaearrangement and sternum structure and are in need ofdescription. Boyer (1916), Patrick and Reimer (1966),Stoermer and Kreis (1978), Lowe and Kociolek (1984),Stoermer et al. (1999) and Edlund (2007 unpubl.data) report N. hasta for different fossil and recentlocalities in North America. After examination of someof the existing material we have concluded thatalthough ‘hasta-like’ specimens are present, none ofthem can be clearly associated with the N. hasta of thetype. These specimens are probably more closelyrelated to the N. hasta s.l. populations from the ancientlakes in Japan and populations from the Baikal RiftZone lakes.
In our future work on the Navicula hasta speciescomplex, a critical approach will be understanding thedistribution and biogeography of these taxa. From ourinitial assessment, we think the presence of the typicalN. hasta outside of the Neogene deposits is doubtful. In
55Identity of Navicula hasta

addition to determining the identity of the type andstudying the full range of morphological variability, theother main problem that produces reports of widelydistributed (temporal, geographic and ecologic) ‘cos-mopolitan’ taxa is the use of widely available and well-illustrated European floras. For example, N. hastasensu Krammer and Lange-Bertalot (1986, fig. 26: 1)is illustrated from a Japanese lake and clearly does notcorrespond to Pantocsek’s concept of N. hasta.
The full diversity within the N. hasta complex is stillunknown. This paper takes a first step by clearly iden-tifying a lectotype and epitype for N. hasta from amongPantocsek’s original illustrations, description and mate-rial. But from Köpecz alone, we have observed at leastthree more morphologically different species that aresympatric with the type: N. krenneri, N. superhasta andN. pseudohasta. Similarly, Lange-Bertalot and Metzel-tin (2006) report two size classes of N. hasta sensu latofrom Sofia Neogene deposits in Bulgaria. Whether thislevel of diversity and these patterns of variation and sizeclasses are repeated elsewhere in the N. hasta-complexis yet to be determined.
ACKNOWLEDGMENTS
The authors express their gratitude to Dr David M.Williams, Dr Taisuke Ohtsuka, Dr Kazuo Haraguchi,Tetsuya Wakabayasi, Satoshi Ichise, Dr KrisztinaBucko, Dr Christine Cocquyt, Dr Richard Crawford, DrDominic Hodgson, Dr Agata Wojtal, Dr N. Soninkhishig,Dr Eugene Stoermer and Dr Norman Andresen for sup-plying the material and slides necessary for analyses ofthe Navicula hasta complex. This material is basedpartly on work supported by the National Park Service(NPS) Great Lakes Monitoring Network CooperativeAgreement H6000C02000 and the National ScienceFoundation (NSF) under grant DEB-0316503 to MBE.Any opinions, findings, and conclusions or recommen-dations expressed in this material are those of theauthor(s) and do not necessarily reflect the views of theNSF or NPS.
REFERENCES
Boyer, C. S. 1916. The Diatomaceae of Philadelphia andVicinity. J.B. Lippincott Co, Philadelphia, PA.
Cleve-Euler, A. 1953. Die Diatomeen von Schweden undFinnland. Teil III. Monoraphideae, Biraphideae 1. Kungl.Svenska Vetskapaka-demiens Handlingar Fjärde Serienn 4:1–255, 41 Tafeln. Stockholm.
Cocquyt, C. 1998. Diatoms from the Northern Basin of LakeTanganyika. Bibliotheca Diatomologica 39: 1–275.
Edlund, M. B., Soninkhishig, N., Williams, R. M. and Sto-ermer, E. F. 2001. Biodiversity in Mongolia: Checklist ofdiatoms, including new distributional reports of 31 taxa.Nova Hedwigia 72: 59–90.
Ehrlich, A. 1995. Atlas of the Inland Water Diatom Flora ofIsrael. Israel Academy of Sciences and Humanities,Jerusalem, 217 pp.
Foged, N. 1978. Diatoms in Eastern Australia. BibliothecaPhycologica 41: 1–243.
Gasse, F. 1980. Les diatomées lacustres plio-pleistocenes duGadeb (Ethiopie). Systematique, paléoécologie, biostratyg-raphie. Rev. Algol. Mém. Sér. 3: 1–249.
Haraguchi, K. 1997. Diatoms from Lake Aoki, Nagano Pre-fecture, Central Japan. Diatom 13: 215–31.
Hodgson, D., Vyverman, W. and Tyler, P. 1997. Diatoms ofmeromictic lakes adjacent to the Gordon River, and of theGordon River estuary in south-west Tasmania. BibliothecaDiatomologica 35: 1–173.
Hustedt, F. 1930. Bacillariophyta (Diatomeae). In Pascher, A.(Ed.) Süßwasserflora von Mitteleuropas, Heft 10. GustavFischer Verlag, Jena, p. 466.
Jurilj, A. 1954. Flora and Vegetation of diatoms in Ochridlake. Prirodslovna istraživanja 26: 99–190.
Krammer, K. and Lange-Bertalot, H. 1986. Bacillariophyceae.1: Naviculaceae. In Ettl, H., Gerloff, J., Heynig, H. andMollenhauer, D. (Eds) Süßwasserflora Von Mitteleuropa,2/1. Gustav Fischer Verlag, Stuttgart.
Krenner, J. A. 1980. The post-war remains of the diatomcollection of Dr. József Pantocsek. Studia Botanica Hun-garica 14: 9–28.
Lange-Bertalot, H. 2001. Navicula sensu stricto, 10 generaseparated from Navicula sensu lato, Frustulia. In Lange-Bertalot, H. (Ed.) Diatoms of Europe. Diatoms of theEuropean Inland Waters and Comparable Habitats, Vol. 2.A. R. G. Gantner Verlag K. G., Königstein, pp. 1–526.
Lange-Bertalot, H. and Metzeltin, D. 2006. Comparison ofseven species of Navicula sensu stricto. In Ognjanova-Rumenova, N. and Manoylov, K. (Eds) Advances in Phy-cological Studies. Festschrift in Honour of Prof. DobrinaTemniskova-Topalova. Pensoft Sofia, pp. 55–72.
Levkov, Z., Krstic, S., Metzeltin, D. and Nakov, T. 2007.Diatoms from Lakes Prespa and Ohrid. Iconographia Diato-mologica 16: 1–603.
Lowe, R. L. and Kociolek, J. P. 1984. New and rare diatomsfrom Great Smoky Mountains National Park. Nova Hedwi-gia 39: 465–76.
Ohtsuka, T. and Tuji, A. 2002. Lectotypification of somepennate diatoms described by Skvortzow in 1936 fromLake Biwa. Phycol. Res. 50: 243–9.
Pantocsek, J. 1892.Beiträge Zur Kenntniss der FossilenBacillarien Ungarns. III. Süßwasser Bacillarien. Anhang:Analysen 15 neuer Depots von Bulgarien, Japan, Mähren,Rublands und Ungarn, 42 Taf. Nagytapolcsány.
Pantocsek, J. 1905. Beiträge zur Kenntniss der fossilen Bacil-larien Ungarns. III. Beschreibung der auf Tafel 1–4 abge-bilden Arten, Pozsoni.
Patrick, R. M. and Reimer, C. W. 1966. The Diatoms of theUnited States Exclusive of Alaska and Hawaii, Vol. I. Fragi-lariaceae, Eunotiaceae, Achnanthaceae and Naviculaceae.
56 Z. Levkov et al.

Monograph No. 13. Academy of Natural Sciences of Phila-delphia, Philadelphia, PA.
Rai, A. K. 2000. Evaluation of natural food for planktivorousfish in Lakes Phewa, Begans, and Rupa in Pokhara Valley,Nepal. Limnology 1: 81–9.
Schmidt, A. Schmidt, M., Fricke, M. et al. (1874–1959).Atlas der Diatomaceen – Kunde, Tafeln 1–460. Aschersle-ben and Leipzig.
Silva, A. 1946. Diatomáceas fósseis de Portugal. Bol. Soc.Geol. Portugal 4: 1–166.
Simonsen, R. 1987. Atlas and Catalogue of the Diatom Typesof Friedrich Hustedt. Volume 1 Catalogue, Volume 2 Atlas,Plates 1–395, Volume 3 Atlas, Plates 396–772. Verlag J,Cramer in der Gebrueder Borntraeger Verlagsbuchhandlung.
Skvortzow, B. W. 1936. Diatoms from Biwa Lake, HonshuIsland, Nippon. Philippine J. Sci. 61: 253–96.
Skvortzow, B. W. 1937. Bottom Diatoms from Olhon Gate ofBaikal Lake, Siberia. Philippine J. Sci. 62: 293–377.
Stoermer, E. F. and Kreis, R. G. Jr. 1978. Preliminary checklist of diatoms (Bacillariophyta) from the Laurentian GreatLakes. J. Great Lakes Res. 4: 149–69.
Stoermer, E. F., Kreis, R. G. Jr and Andersen, A. 1999.Checklist of diatoms from the Laurentian Great Lakes. II.J. Great Lakes Res. 25: 515–66.
Wojtal, A. and Buczko, K. 2004. Comparative study of Nav-icula hasta and N. rakowskae: their morphology and distri-bution. Studia Botanica Hungarica 35: 33–44.
Zabelina, M. M., Kieselev, A. I., Proskina-Lavrenko, A. I. andSheshukova, V. S. 1951. Inventory of Freshwater Algae ofthe USSR, Vol. 4. Diatoms. Sov. Nauka, Moscow.
57Identity of Navicula hasta