identification and characterization of an interspersed repetitive dna fragment in plasmodium vivax...
TRANSCRIPT

www.elsevier.com/locate/yexpr
Experimental Parasitology 108 (2004) 81–88
Identification and characterization of an interspersed repetitiveDNA fragment in Plasmodium vivax with potential use
for specific parasite detection
Silvana Carnevalea,*, Jorge N. Velasquezb, Hernando del Portilloc,Jorge H. Labbea, Marta G. Cabreraa, Marcela Ferellad,
Bjorn Anderssond, Eduardo A. Guarneraa, Sergio O. Angela,e
a Departamento de Parasitologıa, Instituto Nacional de Enfermedades Infecciosas, ANLIS ‘‘Dr. Carlos G. Malbran,’’
Ciudad de Buenos Aires, Argentinab Hospital Municipal de Infecciosas ‘‘Dr. Francisco Javier Muniz,’’ Ciudad de Buenos Aires, Argentina
c Instituto de Ciencias Biomedicas II, Universidad de Sao Paulo, Sao Paulo, Brazild Center for Genomics and Bioinformatics, Karolinska Institute, Stockholm, Sweden
e Laboratorio de Parasitologıa Molecular, UB2, IIB-INTECH, Chascomus, Prov. de Buenos Aires, Argentina
Received 18 August 2003; received in revised form 26 July 2004; accepted 26 July 2004
Available online 2 September 2004
Abstract
We cloned and characterized a Plasmodium vivax repeat element of 7872 bp named PvRE7.8. Several internal tandem repeats
were found along the sequence. The repetitive nature of the PvRE7.8 element was confirmed by hybridization of a P. vivax
YAC library. Based on the data bank analysis and the presence of two contiguous putative genes that may encode proteins related
to DNA metabolism, PvRE7.8 could be considered an inactivated transposon-LINE element. By using Pv79 as probe or primers
derived from Pv79-flanking sequences, P. vivax DNA Could be detected from whole blood and mosquito samples. We consider that
the repeat element described here has potential for P. vivax malaria diagnosis and for epidemiological analysis of P. vivax transmis-
sion areas.
� 2004 Elsevier Inc. All rights reserved.
Index Descriptors and Abbreviations: DNA, deoxyribonucleic acid; PCR, polymerase chain reaction; PBS, phosphate-buffered saline; bp, base pairs;
dNTP, deoxynucleotide triphosphate; ORF, open reading frame; LINE, long interspersed nuclear element; SINE, small interspersed nuclear element
Keywords: Plasmodium vivax; Polymerase chain reaction; Repetitive element; LINE
1. Introduction
The malarial parasite Plasmodium vivax causes dis-ease in humans, including chronic infections and recur-
rent relapses, but the course of the infection is rarely
fatal (Garcia and Bruckner, 1993), unlike that caused
by Plasmodium falciparum. Whereas P. vivax is present
0014-4894/$ - see front matter � 2004 Elsevier Inc. All rights reserved.
doi:10.1016/j.exppara.2004.07.012
* Corresponding author. Fax: +54 11 4301 7437.
E-mail address: [email protected] (S. Carnevale).
in tropical and temperate areas, P. falciparum is only re-
stricted to tropical areas (Garcia and Bruckner, 1993).
In Argentina, malaria is actually associated with P. vi-
vax (Ministerio de Salud, 2002), but patients with P. fal-
ciparum infection are also detected as the result of
travel-related malaria.
In Argentina, endemic P. vivax malaria corresponds
to the northwestern area and it is transmitted by Anoph-
eles pseudopunctipennis. There is also an epidemic area,
in the northeast, related to the vector Anopheles darlingi

82 S. Carnevale et al. / Experimental Parasitology 108 (2004) 81–88
(Rey, 1991; Ministerio de Salud, 2002). Argentina
bounds in the north with Bolivia and Brazil, in which
P. vivax and P. falciparum parasites are established.
Thus, malaria in Argentina is a complex problem related
to poverty in areas with limited access and fluid migra-
tory movements at the bound areas (WHO, 2000;OPS, 2001). In spite of these important problems, up
to now, there were no adequate tools for epidemiologi-
cal studies and control activities.
The detection of malaria classically relies on the dem-
onstration of the pathogen in blood samples, being easy
to differentiate P. vivax from P. falciparum (WHO,
1991). However, this technique is not adequate if large
samples must be analyzed either for screening or epide-miological studies. For this reason, several molecular
analysis were developed, mainly to detect P. falciparum
(Tirasophon et al., 1991; Wataya et al., 1993).
Repetitive DNA elements have shown to be very use-
ful in the development of diagnostic molecular tools. This
repetitive feature allows the improvement of sensitivity
and, in general, these elements are very specific markers.
In nature, repetitive DNA elements have also been takenas important elements to generate variations in genome
structure and expression associated to species evolution
(Kidwell and Lisch, 1997). In this sense, the Plasmodium
spp. genomes showed to have tandem repeat DNA ele-
ments mainly related to subtelomeric regions which were
suggested to promote chromosome pairing, facilitating
meiotic recombination and gene conversion between telo-
mere and proximal genes (DeBruin et al., 1994).We cloned and characterized a P. vivax repeat DNA
element. The DNA fragment was sequenced and its
genomic organization was analyzed through genomic
programs. A highly specific P. vivax region was detected
along the DNA fragment, which was used as source of
DNA probes or PCR-target. Finally, its possible useful-
ness to detect parasite in human blood or mosquito sam-
ples was determined.
2. Materials and methods
2.1. Blood samples
Twenty blood samples from patients with P. vivax
infection and three from patients with P. falciparum
infection were obtained from a ‘‘Blood Bank,’’ orga-
nized by the laboratory of Malaria Diagnosis (Departa-
mento de Parasitologıa, ANLIS ‘‘Dr. Carlos G.
Malbran’’). Additionally, 60 blood samples from a
non-endemic/epidemic area were used as negative con-
trols. Malaria infection was determined by thin and
thick smears. The clinic history and Plasmodium spp./
ll counting were obtained for all positive blood samples.Blood samples were stored at 4 �C and processed in the
next 5 days after their collection.
2.2. Mosquito samples
Homogenates of 18 P. vivax- and six P. falciparum-
experimentally infected mosquitoes were kindly pro-
vided from Dr. J.B. Pereira Lima (Instituto Militar,
Rio de Janeiro, Brazil). These samples had been pre-pared by lysis in 0.01 PBS, pH 7.2, with addition of
3% non-fat milk and 0.5% Nonidet P-40. Sixteen Culex
spp. and eight Aedes spp. mosquitoes were used as neg-
ative controls.
2.3. Purification of parasites
Five milliliters of fresh blood samples infected with P.
vivax (n = 8) or with P. falciparum (n = 2) was centri-
fuged at 800g during 10 min. The pellet was resuspended
in 5 ml of 0.01 M PBS (pH 7.2), and filtered through Bio-
fil R filters (Biofil, Italy). The collected sample was centri-
fuged, the pellet resuspended in PBS and the infected
erythrocytes were separated by centrifugation of the sam-
ple in Nicodenz (57% v/v in PBS) gradient. Erythrocytes
were washed in PBS, resuspended in 0.1% saponin-PBSduring 5 min at room temperature, centrifuged at
10,000g at 4 �C, and washed. Thin smears of 1 ll sample
were dyed with Giemsa and parasites were counted.
2.4. Preparation of DNA samples
DNA from blood and parasites samples were ex-
tracted by Proteinase K/phenol–chloroform protocolas described (Angel et al., 1997). Before extraction,
blood samples were aliquoted in 300 ll. Finally, the
DNA was resuspended in 300 ll (for dot blot) or 10 ll(for PCR) of TE (10 mM Tris–HCl, pH 7.4, and
1 mM EDTA, pH 8.0).
The DNA from vectors was extracted as described
previously (Coulson et al., 1990). After extraction,
DNA was precipitated in absolute ethanol, centrifuged,washed with ethanol at 70 �C, and resuspended in 12 llTE. Homogenates of positive controls were treated by
the same procedure without rehydration steps.
2.5. Screening and obtaining of P. vivax probes
Purified P. vivax DNA was labeled by nick transla-
tion (Nick translation DNA Labeling Kit, Life Technol-ogies) with [32P]dCTP and used for screening a genomic
P.vivax gtWES library by colony hybridization (Mania-
tis et al., 1982). Hybridization was performed in 2· SSC
plus 0.25% non-fat dried milk at 65 �C, and washing at
high stringency in 0.1· SSC, 0.1% SDS at 68 �C. Positiveclones were re-screened for homogeneity.
To obtain Pv79 probe, clone Pv10 was digested with
several endonucleases and analyzed by agarose gel elec-trophoresis stained with ethidium bromide. Since diges-
tion with HaeIII produced smallest DNA fragments,

S. Carnevale et al. / Experimental Parasitology 108 (2004) 81–88 83
500 ng of Pv10 DNA was digested with HaeIII and
cloned into the SmaI site of pUC18 plasmid, and used
to transform DH5a strain of Escherichia coli. Pv10-
HaeIII mini-library was screened with [32P]dCTP radio-
labeled genomic P. vivax and P. falciparum.
To obtain PvE/C probe, clone Pv10 was digested withEcoRI and ClaI and cloned into EcoRI and ClaI sites of
pBluescript KS+ plasmid.
2.6. Sequence analysis
DNA sequencing was performed using a Perkin–El-
mer ABI 377 MegaBACE 1000 machine (Amersham
Biosciences) with a BigDye Terminator Cycle Sequenc-ing Kit (Perkin–Elmer) and a MegaBACE 1000 sequen-
cer and ET terminator Kit (Amersham Biosciences).
Several internal oligonucleotides were designed to allow
the complete sequencing. Sequence similarity and do-
main searches were performed using the Blast program
of the National Center for Biotechnology Information
(NCBI) web page (www.ncbi.nlm.nih.gov). Open read-
ing frame (ORF) searches were performed by ORF fin-der at the NCBI web page. G + C content and tandem
and inverted repeat analysis were performed by Geecee,
Einverted, Equicktandem, Etandem, Palindrome at
www.bioweb.pasteur.fr and as described by Benson
(1999). Curvature and bendability analysis were per-
formed at www3.icgeb.trieste.it/~dna/. The GenBank
Accession No. of PvRE7.8 is AF017049.
2.7. Genomic analysis
Filters containing 560 P. vivax genomic-YAC clones
(Camargo et al., 1997) were hybridized with random
priming (Random Priming Labeling Kit, Life Technolo-
gies) radiolabeled PvRE7.8, Pv79, Pv E/C and the single
copy MSP-1 gene (Premawansa et al., 1993) as well as
with internal oligonucleotide probes. When internal oli-gonucleotides were used, radiolabeling was done by
[c-32P]ATP by using 5 0Terminus Labeling Kit (Life
Technologies) and hybridization and washes were per-
formed at 42 �C.
2.8. Dot blot analysis
Genomic DNA isolated from either purified para-sites, blood or recombinant plasmid was spotted onto
nylon filters (Hybond, Amersham) and treated as de-
scribed (Maniatis et al., 1982). Hybridization and
washes of the filters were performed under the same con-
ditions described above. Probes were 32P-radio- or
digoxigenin-dUTP labeled. Digoxigenin-dUTP labeling
was performed using the DIG DNA Labeling Kit
(Boehringer–Mannheim). Hybridization and signaldevelopment conditions were established as described
previously (Angel et al., 1997). To quantify the levels
of the hybridization, filters were scanned by a Phospho
imager (Bio-Rad) and band-density was analyzed by
using the Molecular Analyst software (Bio-Rad).
2.9. PCR analysis
Amplifications were performed in a Gene Ataq ther-
mal cycler (Pharmacia) with 1 U of Taq DNA polymer-
ase (BRL) in a final volume of 50 ll with 50 mM KCl,
10 mM Tris–HCl, pH 9, 0.1% Triton X-100, 1.5 mM
MgCl2, 0.2 mM (each) dNTP, and 0.2 mM of each oli-
gonucleotide. When mosquito DNA was used as target,
0.2 mg/ml bovine serum albumin was added to the reac-
tion mixture. Samples were amplified for 25 (MSP-1F/Rprimers) or 35 (Pv79F/R and Pv411F/R primers) cycles
as follows: 1 min at 94 �C, 1 min at 42 �C (MSP-1 prim-
ers) (Premawansa et al., 1993), 43 �C (Pv411F/R prim-
ers, 5 0 TGTACATGGAAGCGCTAGCG 3 0 and 5 0
ACTTCCACTTGACGCAGAAG 3 0, respectively) or
55 �C (Pv79F/R primers, 5 0 CCCTGCCGCATCGGC
ACGAA 3 0 and 5 0 CCCACAGGGGGGACACCTTT
3 0, respectively), and 1 min at 72 �C. An initial step of1 min at 98 �C and a final step of 5 min at 72 �C were in-
cluded. PCR products were analyzed by ethidium bro-
mide dying in 1.5% agarose gel.
3. Results
3.1. Isolation of repetitive DNA sequences and cloning of
specific P. vivax DNA probe
To isolate P. vivax repetitive DNA sequences, a geno-
mic cgtWES library screening was screened with 32P-la-
beled P. vivax genomic DNA. One recombinant phage
was selected on the basis of its strongest signal, showing
a 7.8-kpb insert, which was called PvRE7.8 (data not
shown).To detect P. vivax-specific internal regions, PvRE7.8
was digested with HaeIII, their products cloned into
pUC18 plasmid, and the library was screened by repli-
cate plate with 32P-labeled P. vivax and P. falciparum
genomic DNAs. One recombinant pUC18 plasmid re-
leased a P. vivax-specific fragment of approximately
80 bp, which was named Pv79 (Fig. 3).
To confirm theP. vivax specificity of Pv79,P. vivax andP. falciparum genomic DNA obtained from blood sam-
ples of infected individuals were hybridized with both
PvRE7.8 and Pv79 probes. Clearly, Pv79 specifically rec-
ognized P. vivax DNA, whereas PvRE7.8 showed cros-
shybridizationwithP. falciparumDNA(data not shown).
3.2. Genomic characterization of PvRE7.8 and Pv79
A genomic P. vivax YAC library was hybridized with
PvRE7.8, Pv79 and the single copy MSP-1 gene. Several

Fig. 1. Determination of Pv79 copy number in the parasite genome.
Plasmid containing Pv79 was spotted from right to left in quantities of
1010–101 vector molecules, in successive 1:10 dilutions. Parasites were
spotted from right to left in quantities of 25,600 to 50 in successive 1:2
dilutions (107 signal intensity was equivalent to 3200 parasites).
Fig. 2. Schematic representation of the PvRE7.8 region sequence
(thick line). (A) Distribution of repeat DNA elements (boxes) along
the PvRE7.8 sequence. Inverted repeat (IR) sequences flanking the
Pv79 element are indicated as nucleotides. (B) The thin line represents
the nucleotide sequence of a P. knowlesi (PKN) DNA fragment and its
corresponding accession number is indicated above. Identity is
indicated as percentage. (C) Thin lines represent protein sequences
(aa, amino acidic residue). The P. falciparum (Pf) and P. yoeli yoeli
(Pyy) accession numbers corresponding to these proteins are indicated
above these lines. Identities are indicated as percentage.
84 S. Carnevale et al. / Experimental Parasitology 108 (2004) 81–88
clones were identified by using PvRE7.8 and Pv79
probes, but only 2 were detected with the MSP-1 probe,
indicating that the P. vivax probes are repetitive DNA
elements of the parasite (data not shown). As expectedPvRE7.8 and Pv79 probes shared almost all of the posi-
tive clones. However, Pv79 showed more positive signals
than the observed by using PvRE7.8 probe.
To determine the copy number of the P. vivax-specific
probe in the parasite genome, a quantitative assay was
developed (Fig. 1). It could be inferred by comparison
of signal intensities in dot blot hybridization that Pv79
has 1.5 · 103 copies/haploid genome.
3.3. Sequence analysis of PvRE7.8
The nucleotide sequence showed that PvRE7.8 is a
7872-bp element. It is possible to detect several internal
tandem repeats along the sequence that range between 2
and 6 monomers of 6 and 32 bp length (Fig. 3A). It was
also possible to detect several T/A and G/C stretches,and a %G + C of 55% uniformly distributed along the
sequence. The Pv79 repeat element showed to be flanked
by a 9–10 bp inverted repeat including the HaeIII site
(Fig. 2A).
The data Bank searching displayed low homology
with other sequences excepting those emerging of the
2021–2044 bp tandem repeat (element e, [TCTTCA]4),
which had >90% identity with similar elements fromDNAs of a wide range of animals and plants species
(data not shown). Since the ubiquitous feature of this re-
peat it was denominated UbiRE. UbiRE was also de-
tected in a wide number of mRNA and EST sequences
from different organisms (data not shown).
The search in Plasmodium spp. genome data bank
displayed its main identity with short fragments (about
500 bp) of P. vivax and a long fragment of P. knowlesi(Fig. 2B).
3.4. Detection of putative genes in PvRE7.8
When the analysis was performed by Blastx, long
fragments of PvRE7.8 showed a high identity with two
hypothetical proteins from other Plasmodium spp.
(Fig. 2C). In addition, several ORFs were detected alongthe PvRE7.8 sequences (data not shown). Interestingly
the deduced amino acid sequence of the five longest
ORFs (750, 996, 1302, 1431, and 1506 bp) correspondedto the sequences with identity to the P. falciparum and
P. yoelii yoelii hypothetical proteins (data not shown).
Blastp analysis with the amino acid sequences from
these five ORFs showed identities with the Plasmodium
sequences mentioned. To search in protein data bank,
two proteins were constructed based on these five ORFs
and the alignment with these hypothetical Plasmodium
protein sequences, one of them (PvRE7.8-1) with 912residues (from nucleotide 3225 to 494, reverse comple-
mentary sequence) and another (PvRE7.8-2) also with
912 residues (from the end to 4581, reverse complemen-
tary sequence). Some regions of PvRE7.8-1 presented
identity with a human replicative senescence downregu-
lated leo1-like protein, a family of proteins that are part
of the Paf1/RNA polymerase II complex; a 5-methylcy-
tosine G/T mismatch-specific DNA glycosylase, aPhotorhabdus luminescens transposase, and a P. falcipa-
rum b-adaptin (Fig 3A), whereas PvRE7.8-2 showed
identity with several proteins among them an Arabidop-
sis thaliana reverse transcriptase and a Bacillus halodu-

Fig. 3. Amino acid sequence analysis. (A) Blastp sequence alignment between PvRE7.8-1 deduced protein and Hum elong, a human leo1-like protein
elongase (Accession No. NP_620147); Gg DNA-Gl, a Gallus gallus 5-methylcytosine G/T mismatch-specific DNA glycosylase (Accession No.
AAF68981, Zhu et al., 2000); Pl transp, a Photorhabdus luminescens transposase (Accession No. AAN64216, Waterfield et al., 2002); and a P.
falciparum b-adaptin (AN: NP_703622). (B) Blastp sequence alignment between At RT, an Arabidopsis thaliana reverse transcriptase (Accession No.
NP_180710); Bh recomb, a Bacillus halodurans cassette chromosome recombinase B1 (Accession No. NP_241548, Takami et al., 2000); and a SbcC
Dom, a consensus domain corresponding to a SbcC protein (gnl/CDD/10293, COGO419). Points, identical amino acid; +, conservative amino acid.
S. Carnevale et al. / Experimental Parasitology 108 (2004) 81–88 85
rans recombinase B1 (Fig. 3B). In addition, a Blast do-
main analysis detected the presence of a SbcC protein,
an ATPase involved in DNA replication, recombina-
tion, and repair (Fig. 3B). The same analysis with Plas-
modium hypothetical proteins (Fig. 2C) only displayed
identity with themselves (data not shown).
3.5. Detection of parasite in blood and mosquito samples
The feasibility of detecting P. vivax in blood samples
obtained from an infected patient was studied by PCR
analysis using MSP-1F/R and Pv411F/R (F, positions
352–371 and R, positions 743–756 on PvRE7.8). DNA
from blood samples that amplified by using MSP-1primers were used to analyze DNA amplification with
Pv411F/R primers. The expected 411-bp amplification
band was observed in the three positive samples but
not with DNA of a patient with P. falciparum infection
or non-infected patient (Fig. 4A). The sensitivity analy-
sis of Pv411-PCR assay was determined by using blood
samples with a known quantity of P. vivax, resulting in
the detection up to a minimum of 100 parasites (Fig.4B), that is equivalent to 1.7 plasmodium/ll of blood.
The analysis of mosquito samples by dot blot using
non-isotopic-labeled Pv79 probe allowed the specific
detection of P. vivax infected mosquitoes, but with a
low level intensity signal (data not shown). To improve
the diagnosis, Pv411-PCR assay on purified mosquito
DNA was performed. Once again, PCR-amplificationwas observed only in positives samples but not in P.
falciparum infected or non-infected mosquito DNA
(Fig. 5).
4. Discussion
Contrasting that occurs with P. falciparum, up tonow little is known about P. vivax genome. According
to McCutchan et al. (1984), P. vivax genome presents
two G + C components, one of them of 18% and the an-
other one of 30%, but current estimates of the (G + C)
content of the P. vivax genome put it at approximately
45%, in comparison to the (G + C) content of P. falcipa-
rum which is 19% (http://www.tigr.org/tdb/e2k1/pva1/).
PvRE7.8 showed a 55% of G + C content, inferring thatit is an unusual sequence region of the parasite genome.
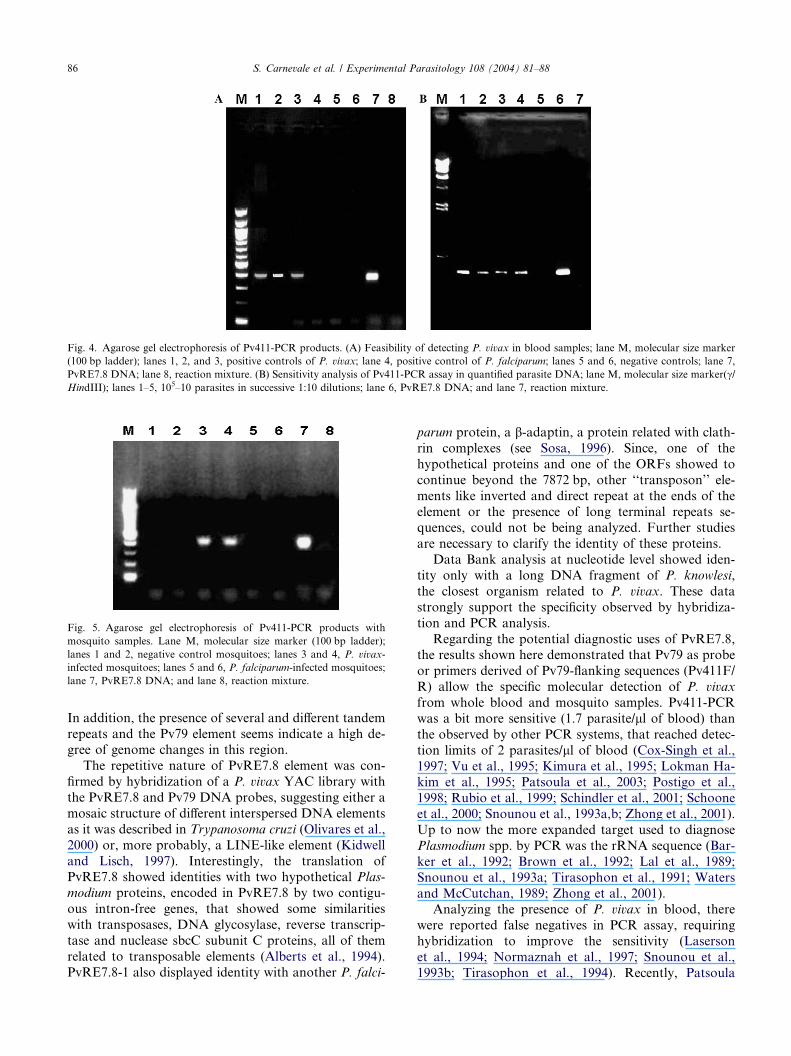
Fig. 4. Agarose gel electrophoresis of Pv411-PCR products. (A) Feasibility of detecting P. vivax in blood samples; lane M, molecular size marker
(100 bp ladder); lanes 1, 2, and 3, positive controls of P. vivax; lane 4, positive control of P. falciparum; lanes 5 and 6, negative controls; lane 7,
PvRE7.8 DNA; lane 8, reaction mixture. (B) Sensitivity analysis of Pv411-PCR assay in quantified parasite DNA; lane M, molecular size marker(c/HindIII); lanes 1–5, 105–10 parasites in successive 1:10 dilutions; lane 6, PvRE7.8 DNA; and lane 7, reaction mixture.
Fig. 5. Agarose gel electrophoresis of Pv411-PCR products with
mosquito samples. Lane M, molecular size marker (100 bp ladder);
lanes 1 and 2, negative control mosquitoes; lanes 3 and 4, P. vivax-
infected mosquitoes; lanes 5 and 6, P. falciparum-infected mosquitoes;
lane 7, PvRE7.8 DNA; and lane 8, reaction mixture.
86 S. Carnevale et al. / Experimental Parasitology 108 (2004) 81–88
In addition, the presence of several and different tandem
repeats and the Pv79 element seems indicate a high de-gree of genome changes in this region.
The repetitive nature of PvRE7.8 element was con-
firmed by hybridization of a P. vivax YAC library with
the PvRE7.8 and Pv79 DNA probes, suggesting either a
mosaic structure of different interspersed DNA elements
as it was described in Trypanosoma cruzi (Olivares et al.,
2000) or, more probably, a LINE-like element (Kidwell
and Lisch, 1997). Interestingly, the translation ofPvRE7.8 showed identities with two hypothetical Plas-
modium proteins, encoded in PvRE7.8 by two contigu-
ous intron-free genes, that showed some similarities
with transposases, DNA glycosylase, reverse transcrip-
tase and nuclease sbcC subunit C proteins, all of them
related to transposable elements (Alberts et al., 1994).
PvRE7.8-1 also displayed identity with another P. falci-
parum protein, a b-adaptin, a protein related with clath-
rin complexes (see Sosa, 1996). Since, one of the
hypothetical proteins and one of the ORFs showed to
continue beyond the 7872 bp, other ‘‘transposon’’ ele-
ments like inverted and direct repeat at the ends of the
element or the presence of long terminal repeats se-quences, could not be being analyzed. Further studies
are necessary to clarify the identity of these proteins.
Data Bank analysis at nucleotide level showed iden-
tity only with a long DNA fragment of P. knowlesi,
the closest organism related to P. vivax. These data
strongly support the specificity observed by hybridiza-
tion and PCR analysis.
Regarding the potential diagnostic uses of PvRE7.8,the results shown here demonstrated that Pv79 as probe
or primers derived of Pv79-flanking sequences (Pv411F/
R) allow the specific molecular detection of P. vivax
from whole blood and mosquito samples. Pv411-PCR
was a bit more sensitive (1.7 parasite/ll of blood) thanthe observed by other PCR systems, that reached detec-
tion limits of 2 parasites/ll of blood (Cox-Singh et al.,
1997; Vu et al., 1995; Kimura et al., 1995; Lokman Ha-kim et al., 1995; Patsoula et al., 2003; Postigo et al.,
1998; Rubio et al., 1999; Schindler et al., 2001; Schoone
et al., 2000; Snounou et al., 1993a,b; Zhong et al., 2001).
Up to now the more expanded target used to diagnose
Plasmodium spp. by PCR was the rRNA sequence (Bar-
ker et al., 1992; Brown et al., 1992; Lal et al., 1989;
Snounou et al., 1993a; Tirasophon et al., 1991; Waters
and McCutchan, 1989; Zhong et al., 2001).Analyzing the presence of P. vivax in blood, there
were reported false negatives in PCR assay, requiring
hybridization to improve the sensitivity (Laserson
et al., 1994; Normaznah et al., 1997; Snounou et al.,
1993b; Tirasophon et al., 1994). Recently, Patsoula

S. Carnevale et al. / Experimental Parasitology 108 (2004) 81–88 87
et al. (2003), developed a multiplex PCR assay, that al-
lowed to differentiate in a 4-h-assay P. falciparum and P.
vivax by differential amplification band length. The
Pv411-PCR developed here is also a single-step, accu-
rate, and reproducible assay that allows to detect specif-
ically P. vivax in a short time either from blood ormosquito samples. In addition, our assays showed that
parasites could be detected by a hybridization assay.
The development of simple PCR or an alternative
technique as dot blot assay could represent a very
important tool for malaria control programs. The detec-
tion of parasites in mosquitoes should be highly valu-
able since different areas could be studied to know the
transmission potential based on the detection of infectedvectors (Ramsey, 1988). In addition, a large number of
human blood samples of low volume could be also ana-
lyzed with epidemiological purposes. In conclusion, we
consider that the repeat element described here could
be an interesting tool for P. vivax malaria diagnosis
and for epidemiological analysis of P. vivax transmis-
sion areas. In addition, it showed to be the first LINE
element described in Plasmodium spp.
Acknowledgments
We acknowledge Dr. Jose Bento Pereira Lima from
Instituto Militar, Rio de Janeiro, Brazil, for kindly pro-
viding infected-mosquito samples. We thank Dr. Nico-
las Schweigmann, from Facultad de Ciencias Exactas yNaturales, Universidad de Buenos Aires, Argentina,
for providing negative mosquito controls. S.O. Angel
is a member of the Consejo Nacional de Investigaciones
Cientıficas y Tecnicas (CONICET) and of the Universi-
dad de Buenos Aires (Departamento de Fisiologıa y
Biologıa Molecular, FCEyN). This work was supported
by ANLIS Dr. Carlos G. Malbran.
References
Alberts, B., Bray, D., Lewis, J., Raff, M., Roberts, K., Watson, J.D.,
1994. Molecular Biology of the Cell, third ed. Garland Publishing
Inc., New York.
Angel, S.O., Matrajt, M., Margarit, J., Nigro, M., Illescas, E.,
Pszenny, V., Amendoeira, R., Guarnera, E., Garberi, J.C., 1997.
Screening for active toxoplasmosis in human patients by DNA
hybridizations with ABGTg7 probe in blood samples. Journal of
Clinical Microbiology 35, 591–595.
Barker, R.H., Banchongaksorn, T., Courval, J.M., Suwonkerd, W.,
Rimwungtragoon, K., Wirth, D.F., 1992. A simple method to
detect Plasmodium falciparum directly from blood samples using
PCR. The American Journal of Tropical Medicine and Hygiene 46,
416–426.
Benson, G., 1999. Tandem repeats finder: a program to analyze DNA
sequences. Nucleic Acids Research 27, 573–580.
Brown, A.E., Kain, K.C., Pipithkul, J., Webster, H.K., 1992.
Demonstration by the polymerase chain reaction of mixed Plas-
modium falciparum and P. vivax infections undetected by conven-
tional microscopy. Transactions of the Royal Society of Tropical
Medicine and Hygiene 86, 609–612.
Camargo, A.A., Fischer, K., Lanzer, M., del Portillo, H.A., 1997.
Construction and characterization of a Plasmodium vivax genomic
library in yeast artificial chromosomes. Genomics 42, 467–473.
Coulson, R.M.R., Curtis, C.F., Ready, P.D., Hill, N., Smith, D.F.,
1990. Amplification and analysis of human DNA present in
mosquito bloodmeals. Medical Veterinary Entomology 90, 357–
366.
Cox-Singh, J., Mahayet, S., Abdullah, M.S., Singh, B., 1997. Increased
sensitivity of malaria detection by nested polymerase chain reaction
using simple sampling and DNA extraction. International Journal
for Parasitology 27, 1575–1577.
DeBruin, D., Lanzer, M., Ravetch, J.V., 1994. The polymorphic
subtelomeric regions of Plasmodium falciparum chromosomes
contain arrays of repetitive sequence elements. Proceedings of the
National Academy of Sciences of the United States of America 91,
619–623.
Garcia, L.S., Bruckner, D.A., 1993. Malaria and Babesia spp. In:
Garcia, L.S., Bruckner, D.A. (Eds.), Diagnostic Medical Parasi-
tology. American Society for Microbiology, Washington, DC, pp.
113–138.
Kidwell, M.G., Lisch, D., 1997. Transposable elements as sources of
variation in animals and plants. Proceedings of the National
Academy of Sciences of the United States of America 94, 7704–
7711.
Kimura, M., Miyake, H., Kim, H.S., Tanabe, M., Arai, M., Kawai, S.,
Yamane, A., Wataya, Y., 1995. Species-specific PCR detection of
malaria parasites by microtiter plate hybridization: clinical study
with malaria patients. Journal of Clinical Microbiology 33, 2342–
2346.
Lal, A.A., Changkasiri, S., Hollingdale, M.R., McCutchan, T.F.,
1989. Ribosomal RNA-based diagnosis of Plasmodium falciparum
malaria. Molecular and Biochemical Parasitology 36, 67–71.
Laserson, K., Petralanda, Y., Hamlin, D., Almera, R., Fuentes, M.,
Carrasquel, A., Barker, R., 1994. Use of polymerase chain reaction
to directly detect malaria parasites in blood samples from the
Venezuelan Amazon. The American Journal of Tropical Medicine
and Hygiene 50, 169–180.
Lokman Hakim, S., Furuta, T., Noor Rain, A., Normaznah, Y.,
Mohd Zamri, M.R., Kojima, S., 1995. Diagnosis of malaria using a
plate hybridization method for the detection of polymerase chain
reaction products. Transactions of the Royal Society of Tropical
Medicine and Hygiene 89, 271–272.
Maniatis, T., Fritsch, E.F., Sambrook, J., 1982. Molecular Cloning: A
Laboratory Manual. Cold Spring Harbor Laboratory, Cold Spring
Harbor, New York.
McCutchan, T.F., Dame, J.B., Miller, L.H., Barnswell, J., 1984.
Evolutionary relatedness of Plasmodium species as determined by
the structure of DNA. Science 225, 808–811.
Ministerio de Salud, 2002. Boletın epidemiologico Nacional. Available
from: <http//www.direpi.vigia.org.ar>.
Normaznah, Y., Furuta, T., Saniah, K., Noor Rain, A., Kojima, S.,
Mak, J.W., 1997. Diagnosis of malaria by enzymatic detection of
polymerase chain reaction products. Southeast Asian Journal of
Tropical Medicine and Public Health 28, 432–433.
Olivares, M., Thomas, M.C., Lopez-Barajas, A., Requena, J.M.,
Garcıa-Perez, J.L., Angel, S.O., Alonso, C., Lopez, M.C., 2000.
Genomic clustering of the Trypanosoma cruzi non-long terminal
L1Tc retrotransposon with defined interspersed repeated DNA
elements. Electrophoresis 14, 2973–2982.
Organizacion Panamericana de la Salud, 2001. Boletın Epidemiolog-
ico, vol. 22, No. 1, OPS, Washington, DC.
Patsoula, E., Spanakos, G., Sofianatou, D., Parara, M., Vakalis, N.C.,
2003. A single-step, PCR-based method for the detection and
differentiation of Plasmodium vivax and P. falciparum. Annals of
Tropical Medical Parasitology 97, 15–21.

88 S. Carnevale et al. / Experimental Parasitology 108 (2004) 81–88
Postigo, M., Mendoza Leon, A., Perez, H.A., 1998. Malaria diagnosis
by the polymerase chain reaction: a field study in south-eastern
Venezuela. Transactions of the Royal Society of Tropical Medicine
and Hygiene 92, 509–511.
Premawansa, S., Snewin, V.A., Khouri, E., Mendis, K.N., David,
P.H., 1993. Plasmodium vivax: recombination between potential
allelic types of the merozoite surface protein MSP1 in parasites
isolated from patients. Experimental Parasitology 76, 192–199.
Ramsey, J.M., 1988. La deteccion de parasitos de la malaria en el
mosquito por observacion microscopica. In: Lopez Antunano, F.J.,
Schmunis, G., (Eds.), Diagnostico de Malaria. Publicacion
Cientıfica No. 512. OPS, Washington, DC, pp. 51–56.
Rey, L., 1991. Parasitologia. Segunda Edicao. Guanabara Koogan
S.A., Rio de Janeiro, Brazil.
Rubio, J.M., Benito, A., Roche, J., Berloza, P.J., Garcıa, M.L., Mico,
M., Edu, M., Alvar, J., 1999. Semi-nested, multiplex polymerase
chain reaction for detection of human malaria parasites and
evidence of Plasmodium vivax infection in Equatorial Guinea. The
American Journal of Tropical Medicine and Hygiene 60, 183–187.
Schindler, H.C., Montenegro, L., Montenegro, R., Carvalho, A.B.,
Abath, F.G., Jaureguiberry, G., 2001. Development and optimi-
zation of polymerase chain reaction-based malaria diagnostic
methods and their comparison with quantitative buffy coat assay.
The American Journal of Tropical Medicine and Hygiene 65, 355–
361.
Schoone, G.J., Oskam, L., Kroon, N.C.M., Schallig, H.D.F.H., Omar,
S.A., 2000. Detection and quantification of Plasmodium falciparum
in blood samples using quantitative nucleic acid sequence-based
amplification. Journal of Clinical Microbiology 38, 4072–4075.
Snounou, G., Viriyakosol, S., Jarra, W., Thaithong, S., Neil Brown,
K., 1993a. Identification of the four human malaria parasite species
in field samples by the polymerase chain reaction and detection of a
high prevalence of mixed infections. Molecular and Biochemical
Parasitology 58, 283–292.
Snounou, G., Viriyakosol, S., Zhu, X., Jarra, P., Pinheiro, W., do
Rosario, L., Thaithong, V.E., Brown, K.N., 1993b. High sensitivity
of detection of human malaria parasites by the use of nested
polymerase chain reaction. Molecular and Biochemical Parasitol-
ogy 61, 315–320.
Sosa, M.A., 1996. The adaptor complexes: a bridge between the
transmembrane proteins and clathrin lattices. Biocell 20, 301–305.
Takami, H., Nakasone, K., Takaki, Y., Maeno, G., Sasaki, R., Masui,
N., Fuji, F., Hirama, C., Nakamura, Y., Ogasawara, N., Kuhara, S.,
Horikoshi, K., 2000. Complete genome sequence of the alkaliphilic
bacterium Bacillus halodurans and genomic sequence comparison
with Bacillus subtilis. Nucleic Acids Research 28, 4317–4331.
Tirasophon, W., Ponglikitmongkol, M., Wilairat, P., Boonsaeng, V.,
Panyim, S., 1991. A novel detection of a single Plasmodium
falciparum in infected blood. Biochemical and Biophysics Research
Communications 175, 179–184.
Tirasophon, W., Rajkulchai, P., Ponglikitmongkol, M., Wilairat, P.,
Boonsaeng, V., Pnyim, S., 1994. A highly sensitive, rapid, and
simple polymerase chain reaction-based method to detect human
malaria (Plasmodium falciparum and Plasmodium vivax) in blood
samples. The American Journal of Tropical Medicine and Hygiene
51, 308–313.
Vu, T.T., Tran, V.B., Phan, N.T., Le, T.T., Luong, V.H., O�Brien, E.,Morris, G.E., 1995. Screening donor blood for malaria by
polymerase chain reaction. Transactions of the Royal Society of
Tropical Medicine and Hygiene 89, 44–47.
Wataya, Y., Arai, M., Kubochi, F., Mizukoshi, C., Kakutani, T., Ishii,
A., 1993. DNA diagnosis of falciparum malaria using a double
PCR technique; a field trial in the Solomon Islands. Molecular and
Biochemical Parasitology 58, 283–292.
Waterfield, N.R., Daborn, P.J., French-Constant, R.H., 2002. Geno-
mic islands in Photorhabdus. Trends in Microbiology 10, 541–545.
Waters, A.P., McCutchan, T.F., 1989. Rapid, sensitive diagnosis of
malaria based on ribosomal RNA. Lancet 17, 1343–1346.
World Health Organization, 1991. Basic Malaria Microscopy. WHO,
Geneva, Switzerland.
World Health Organization, 2000. WHO Expert Committee on
Malaria. Twentieth Report. WHO Technical Report Series 892.
WHO, Geneva, Switzerland.
Zhong, K.J.Y., Salas, C.J., Shafer, R., Gubanov, A., Gasser, R.A.,
Magill, A.J., Russ Forney, J., Kain, K.C., 2001. Comparison of
Iso-Code STIX and FTA Gene Guard collection matrices as
whole-blood storage and processing devices for diagnosis of
malaria by PCR. Journal of Clinical Microbiology 39, 1195–1196.
Zhu, B., Zheng, Y., Angliker, H., Schwarz, S., Thiry, S., Siegmann,
M., Jost, J.P., 2000. 5-Methylcytosine DNA glycosylase activity is
also present in the human MBD4 (G/T mismatch glycosylase) and
in a related avian sequence. Nucleic Acids Research 28, 4157–4165.