human adaptation to the hypoxia of high altitude - cyberleninka
TRANSCRIPT

1
Human adaptation to the hypoxia of high altitude – the Tibetan paradigm 1
from the pre-genomic to the post-genomic era 2
Nayia Petousi and Peter A. Robbins 3
Department of Physiology, Anatomy and Genetics, University of Oxford, Oxford OX1 3PT, 4
United Kingdom 5
6
*Correspondence regarding publication to: Professor Peter A Robbins, 7
Department of Physiology, Anatomy & Genetics, Sherrington Building, Parks Road, Oxford 8
OX1 3PT, United Kingdom; Email: [email protected] 9
10
Running Head: High altitude adaptation in Tibetans 11
12
Articles in PresS. J Appl Physiol (November 7, 2013). doi:10.1152/japplphysiol.00605.2013
Copyright © 2013 by the American Physiological Society.

2
Abstract 13
The Tibetan Plateau is one of the highest regions on Earth. Tibetan highlanders are adapted to 14
life and reproduction in a hypoxic environment and possess a suite of distinctive 15
physiological traits. Recent studies have identified genomic loci that have undergone natural 16
selection in Tibetans. Two of these loci – EGLN1 and EPAS1 – encode major components of 17
the hypoxia-inducible factor (HIF) transcriptional system, which has a central role in oxygen 18
sensing and coordinating an organism’s response to hypoxia, as evidenced by studies in 19
humans and mice. An association between genetic variants within these genes and 20
hemoglobin concentration in Tibetans at high altitude was demonstrated in some of the 21
studies (8, 80, 96). Nevertheless, the functional variants within these genes and the 22
underlying mechanisms of action are still not known. Furthermore, there are a number of 23
other possible phenotypic traits, besides hemoglobin concentration, upon which natural 24
selection may have acted. Integration of studies at the genomic level with functional 25
molecular studies and studies in systems physiology has the potential to provide further 26
understanding of human evolution in response to high-altitude hypoxia. The Tibetan 27
paradigm provides further insight on the role of the HIF system in humans in relation to 28
oxygen homeostasis. 29
30
Keywords: hypoxia-inducible factor, EPAS1, EGLN1, evolution, Tibetan, adaptation 31
32
33

3
Introduction 34
The severely reduced oxygen availability at high altitude, termed hypobaric hypoxia, presents 35
a significant challenge to the ability of humans residing there to live and reproduce. As such, 36
it is likely to have acted as an agent of natural selection. Three human populations have lived 37
at high altitude for millenia – Andeans on the Andean Altiplano, Tibetans on the Tibetan 38
Plateau and Ethiopians on the Semian Plateau. Of these, Tibetans appear to have lived at high 39
altitude the longest. According to archeological and other data, e.g. genetic studies, humans 40
have lived on the Tibetan Plateau – one of the highest regions on Earth with an average 41
elevation of ~ 4000 m, a barometric pressure of less than 500 mmHg and an inspired partial 42
pressure of oxygen of ~ 80 mmHg – for at least 25 000 years (1) (2) (101) (71). In contrast, it 43
is estimated that the Andean Altiplano was first populated approximately only 11000 years 44
ago (2), while the Amhara population in Ethiopia is assumed to have settled at high altitude 45
for ~ 5000 years (3). This suggests that Tibetans will have had more time and opportunity for 46
natural selection in response to a hypoxic environment than any other high-altitude human 47
population. 48
49
Over the last few decades, much research has been done to investigate how Tibetans are 50
adapted to life in a hypoxic environment. In comparison with other high-altitude populations 51
and low-land natives who emigrated to high altitude, Tibetans were found to exhibit 52
distinctive physiological traits and to be resistant to certain pathophysiological processes. 53
While in a number of studies these traits exhibited a great degree of heritability (11) (12) (7) 54
(20), it was largely unknown whether the distinctive high-altitude Tibetan phenotype was the 55
result of natural selection. It is only recently, within the last few years, i.e. in the post-56
genomic era, that advances in genomic technology and science have made it possible to 57
gather evidence of natural selection in Tibetans and to implicate certain genes in their genetic 58

4
adaptation. Nevertheless, the functional variants within these genes and the underlying 59
mechanisms involved are still unknown. 60
61
In this review we highlight the dominant physiological features of Tibetans living at high 62
altitude. We then review the findings of recent studies investigating genetic selection in 63
Tibetans. We discuss how the genes identified as strong candidates for a role in Tibetans’ 64
evolutionary adaptation to high altitude are involved in human physiological processes 65
related to oxygen homeostasis. We discuss and speculate on the potential physiological 66
trait(s) upon which natural selection has acted. Finally, we consider how the evolutionary 67
paradigm of the Tibetan high altitude adaptation provides insights and raises questions on the 68
role of the HIF system in humans that live at sea level. 69
70
Tibetan physiology at high altitude 71
The human response to hypoxia is characterised by systemic changes in cardiovascular, 72
respiratory and hematopoietic physiology which affect convective oxygen transport. Central 73
to the Tibetan high-altitude phenotype is the fact that certain components of convective 74
oxygen transport are markedly different in Tibetans compared with other humans living at 75
high altitude. In this review, most comparisons are made between Tibetans and Andeans, as 76
the physiology of these two populations has been extensively studied. In contrast, 77
significantly less is known about the physiology of the Ethiopian highlanders. 78
79
An immediate rise in ventilation is the most obvious response of human lowlanders exposed 80
to acute high-altitude hypoxia, followed by more complex time-dependent changes with 81
prolonged exposure (69). Tibetan highlanders resemble acclimatised newcomers in that they 82
maintain a high resting ventilation and brisk hypoxic ventilatory sensitivities (32) (36), 83

5
whereas Andeans exhibit a considerable degree of ventilatory blunting (78) (46). Indeed, 84
Tibetan highlanders had elevated resting ventilation and augmented acute hypoxic ventilatory 85
responses when compared directly with Han Chinese long-term residents of the same altitude 86
(102) or Bolivian Aymaras resident in the Andes (11). 87
88
Hypoxia constricts rather than dilates the human pulmonary vasculature (59). Regional 89
hypoxic pulmonary vasoconstriction (HPV) distributes blood away from hypoxic regions of 90
the lung and can be beneficial in maintaining perfusion-ventilation matching. However, a 91
consequence of HPV is that global hypoxia, such as that experienced at high altitude, leads to 92
pulmonary arterial hypertension and right heart strain. Tibetan highlanders, in contrast to 93
their Andean counterparts or other high-altitude residents, exhibit a relative resistance to 94
developing pulmonary hypertension. This was demonstrated directly in a study of 5 Tibetans 95
resident at 3658 m who had pulmonary arterial pressures within sea-level norms which were 96
little changed by additional hypoxia (27). Consistent with this is the demonstration of 97
elevated nitric oxide – a vasodilator – in the lungs of Tibetans as compared with Andean 98
highlanders and lowlanders at sea level (10) (34). Furthermore, a study demonstrated lack of 99
smooth muscle in the small pulmonary arteries in Tibetan men at 3600 m consistent with the 100
absence of pronounced pulmonary vascular remodelling and hypertrophy that accompanies 101
pulmonary hypertension (30). 102
103
A hallmark of high-altitude hypoxic exposure in humans is the rise in red cell mass 104
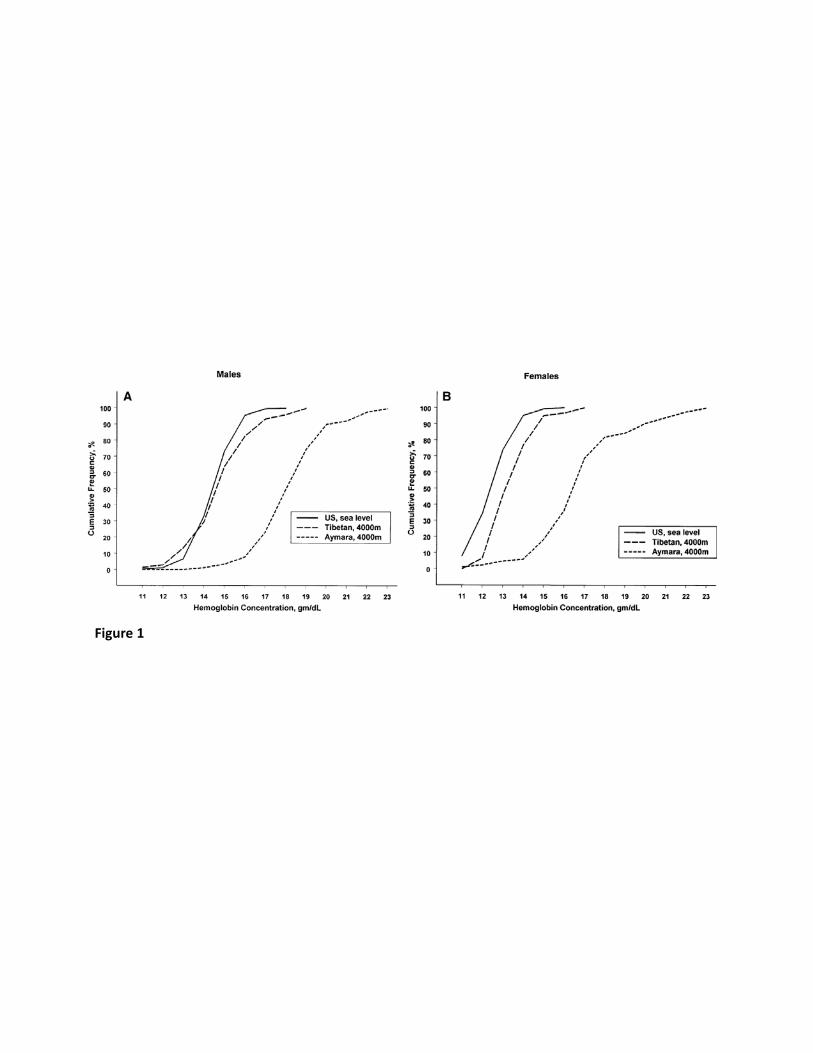
accompanied by elevated hemoglobin and hematocrit, which is associated with a rise in blood 105
erythropoietin (Epo) levels (73). Perhaps the most striking phenotypic difference between 106
Tibetan highlanders and other high-altitude residents is their blunted erythropoietic response 107
to hypoxia. This is evident by the fact that, at similar high altitudes, Tibetans have 108

6
significantly lower hemoglobin concentrations – typically 1 to 3.5 g/dl lower – than their 109
Andean counterparts or Han Chinese migrants to high altitude (7, 26, 55, 90). Tibetans 110
exhibit little or no increase in hemoglobin concentration with increasing altitude (6) (91) and 111
at 4000 m, both male and female Tibetans have hemoglobin concentrations comparable to 112
those of US sea-level residents (7), see Fig. 1. 113
114
Chronic mountain sickness (CMS) is a high-altitude disorder linked with the above 115
components of oxygen transport. It is a syndrome of adaptive failure to chronic hypoxia 116
characterised by excessive erythrocytosis with abnormally high hemoglobin and hematocrit 117
levels, hypoventilation, pulmonary hypertension and eventually right heart failure (53) (47). 118
It occurs in adults after prolonged residence at high altitude and is associated with significant 119
morbidity and mortality (60) (40). Of interest is the remarkably low prevalence of chronic 120
mountain sickness (CMS) in Tibetan highlanders in comparison with other high-altitude 121
populations such as Andeans or Han Chinese migrants to high altitude (54). The overall 122
prevalence of CMS in Tibetans living on the Qinghai-Tibetan plateau was 1.2 % compared 123
with 5.6 % in Han Chinese (92). Using the same criteria as Monge et al (53) to define CMS – 124
Hb >21.3 g/dl and SaO2 < 83 % – a lower prevalence was reported in Tibetans (0.9 %) at 125
4300 m (Madu) (93) in comparison with Peruvian Quechuas (15.6 %) at the same altitude 126
(Cerro De Pasco) (53). 127
128
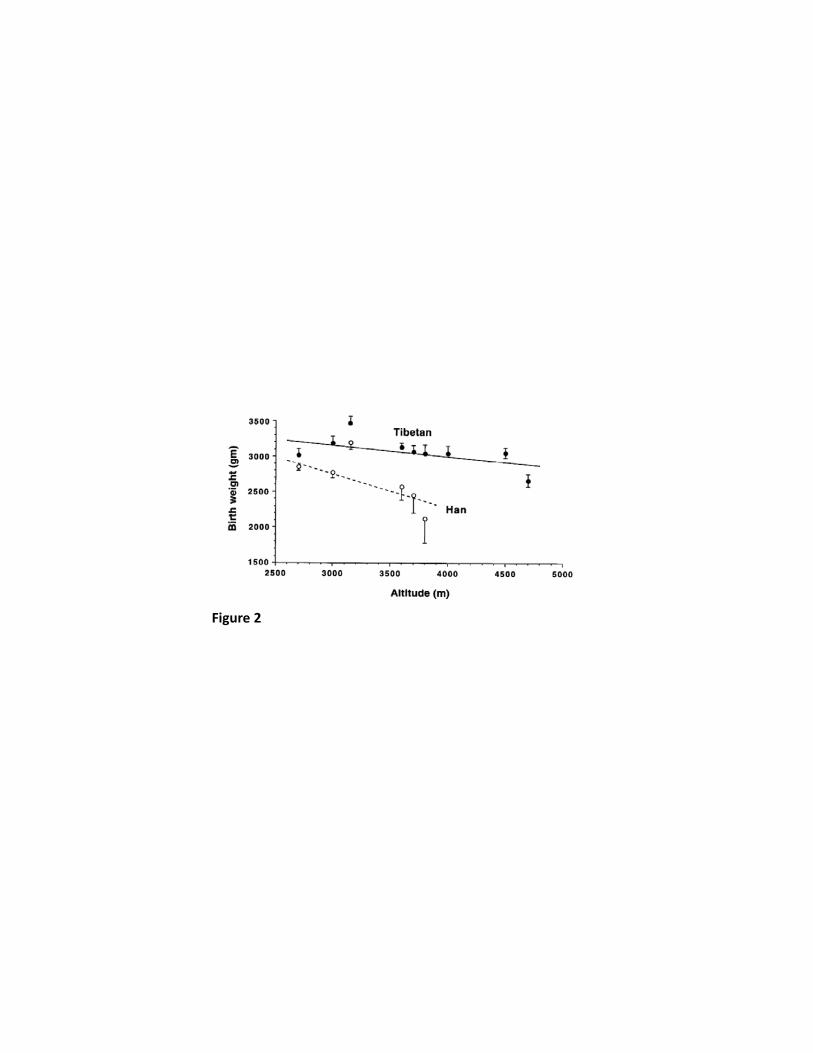
Another characteristic of the Tibetan population is a relative protection against the occurrence 129
of intra-uterine growth restriction (IUGR), which is associated with low birth weight at high 130
altitude. It is well recognised that reproductive success is more difficult at high than low 131
altitude, especially amongst non-natives (43). Birth weight progressively reduces with 132
increasing altitude across populations (41, 56) (97). However, the magnitude of this fall in 133

7
birth weight varies amongst populations; it is smaller in populations that have lived for 134
multiple generations at high altitude (31) (57) (99), such as Tibetans and Andeans, suggesting 135
that genetic factors play a role. Not only were heavier birth weights observed in Tibetans 136
compared with Han Chinese at the same altitude but the pre- and post-natal mortality was 137
threefold higher in Han Chinese than Tibetans (57). Figure 2 shows the birth weight 138
reduction with altitude for Tibetan and Han Chinese. To explain these differences, various 139
studies have focused on oxygen delivery to the placenta, and found that they are associated 140
with differences in uterine (UA) blood flow, as opposed to changes in ventilation, 141
hemoglobin concentration or hemoglobin saturation in pregnant women. Higher UA flow 142
velocities (58) and larger UA diameters (17) were observed in pregnant Tibetan compared 143
with Han Chinese women. Analogous results have been found in studies of Andean 144
populations; a doubling of the UA diameter during pregnancy was demonstrated in Andean 145
women but not in European women at high altitude, an effect seen only under circumstances 146
of chronic hypoxia but not at low altitude (89) (42) (100). The underlying mechanisms for 147
these observations – and whether they are the same in Tibetans and Andeans – are not 148
currently known. 149
150
The phenotypic characteristics of Ethiopian highlanders have not been examined as 151
extensively. Amhara Ethiopians at 3500 m were reported to have low haemoglobin 152
concentrations comparable with those of lowlanders (9) (3), resembling Tibetans; however, 153
Scheinfeldt et al (75) observed higher hemoglobin concentration in high-altitude Amharas 154
compared to lowlanders. Pulmonary arterial pressure, on the other hand, was found to be 155
elevated in Ahmara highlanders (35). 156
157

8
The distinctive oxygen-transport traits found in Tibetans in comparison with other high-158
altitude populations, together with reports of intra-population heritability, have been 159
interpreted as demonstrating the presence of genetic influences on high-altitude adaptation. 160
Moreover, the association of these traits to disorders such as CMS and IUGR, which have the 161
potential to influence reproduction and survival, suggests that some of these phenotypic 162
characteristics, alone or in combination, may be offering an evolutionary survival benefit to 163
Tibetans. 164
165
Studies of the Tibetan genome – evidence of natural selection 166
Recent genome-wide analyses in Tibetans provided the first lines of evidence that this 167
population has undergone genetic adaptation to high altitude. These studies used different 168
methodologies to compare the Tibetan genome with genomes of closely-related ethnic 169
backgrounds, such as the Han Chinese (8, 13, 62, 80, 88, 95, 96). The rationale was that any 170
differences captured in the genetic structure between the two populations are likely to have 171
arisen through natural selection in response to the hypoxic environment inhabited by the 172
Tibetans. These studies have been extensively reviewed (79) (5, 51, 76). The studies are 173
summarised in Table 1 and the key findings are discussed below. 174
175
Simonson et al (80) performed a genome-wide comparison of Tibetan and Han Chinese or 176
Japanese populations. They identified chromosomal regions of positive selective sweeps by 177
calculating two separate genomic statistics: the cross population extended haplotype 178
homozygosity (XP-EHH) statistic and the integrated haplotype score (iHS) for 200 kb non-179
overlapping genomic regions. The basis of these and other techniques are described in recent 180
reviews (15, 82). They then scanned the positive regions for 247 a priori selected candidate 181
genes (based on their involvement in oxygen homeostasis) and found 10 genes that were 182

9
located in or near these regions. Among these genes were EPAS1, EGLN1 and PPARA, with 183
EGLN1 identified by both genomic statistics. In addition, in a cohort of 29 Tibetans, genetic 184
variants in the latter two genes were associated with hemoglobin concentration such that the 185
selected haplotype in each gene was correlated with low hemoglobin concentration. 186
However, people with excessive erythrocytosis or anemia were not excluded from this 187
genotype-phenotype analysis. 188
189
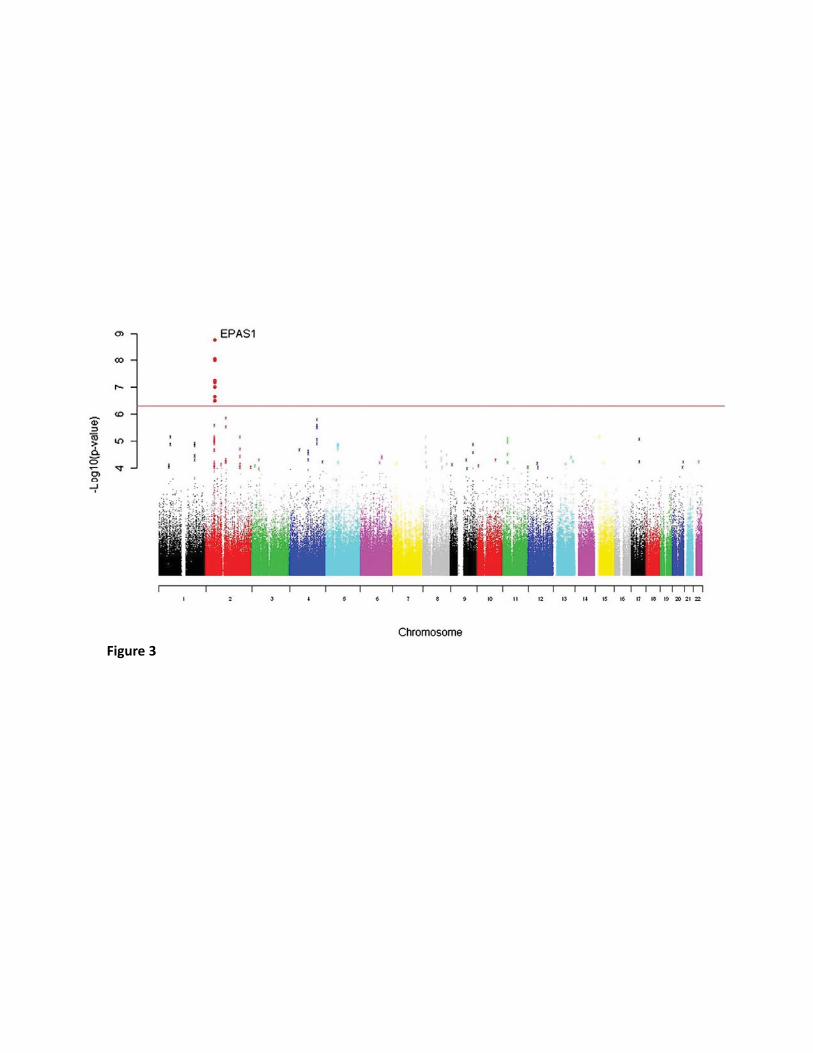
Beall et al (8) used a genome-wide allelic differentiation scan (GWADS) to compare ~ 190
500,000 SNPs between Tibetan high-altitude natives (from the Yunnan province) and low-191
land Han Chinese (see Fig. 3). This provided a completely unbiased genome-wide analysis 192
investigating genetic selection, with no a priori assumptions or hypotheses. They found 8 193
SNPs that achieved genome-wide significance (p values ranging from 2.81 × 10−7 to 1.49 × 194
10−9) in terms of their divergence between the two ethnic groups, which clustered within 195
~235 kb on chromosome 2, just downstream of EPAS1. These SNPs were found to be in high 196
linkage disequilibrium (LD) with one another – i.e. they were associated with each other in a 197
non-random way such that they occurred together more often than expected by chance. This 198
extended haplotype was present at high frequencies in Tibetans (46 %) and low frequencies 199
(2 %) in Han Chinese. In addition, through candidate gene approaches in two further separate 200
cohorts of Tibetans they identified ~30 single nucleotide polymorphisms (SNP) in EPAS1 in 201
high LD (consistent with a dominant variant hypothesis) that correlated significantly with 202
hemoglobin concentration. The correlation was such that major allele (alias “Tibetan” allele) 203
homozygotes averaged a Hb concentration of ~ 1 g/dl lower than the heterozygotes in each 204
separate cohort. These EPAS1 SNPs were found at higher frequencies in Tibetans than in Han 205
Chinese and were in high LD with the signal from the GWADS, thus providing evidence for 206
selection on EPAS1 associated with low hemoglobin concentration in Tibetan highlanders. 207

10
208
Yi et al (96) sequenced 50 Tibetan exomes and compared the sequencing data with Han 209
Chinese and Danish populations to identify changes in allelic frequencies consistent with 210
genetic adaptation in Tibetans. Using population branch statistics (PBS) they identified 211
EPAS1 as the strongest candidate gene for natural selection. In addition, the most 212
differentiated (in terms of allelic frequency) EPAS1 variant was correlated with erythrocyte 213
count within a larger Tibetan cohort. This was an intronic EPAS1 SNP which was captured 214
by this exome-targeted approach and was found at 87 % frequency in Tibetans compared 215
with 9 % in Han Chinese. Interestingly, no coding genetic variants were identified to be 216
highly differentiated between the populations. This leads to suggest that adaptation to high 217
altitude has not proceeded by way of selection on coding variants that might be expected to 218
alter protein structure and function. Based on their analyses, they estimated a Tibetan-Han 219
divergence time of 2750 years, which is far more recent than previously estimated based on 220
archeological data or other analyses (1) (101) (62). This short divergence time has been 221
disputed; it has been argued that it may be related to a complex mosaic of the Tibetan 222
population created by multiple migrations at different times into the Plateau and to the small 223
sample sizes in the study (1). 224
225
Xu et al (95) compared genome-wide allelic frequencies between Tibetan and Han Chinese 226
and identified 6 EGLN1 SNPs and 25 EPAS1 SNPs among the top differentiated SNPs (top 227
0.0001% SNPs with FST statistic > 0.3). They went on to identify positive selective sweeps 228
and dominant haplotypes in EPAS1 and EGLN1 genes in Tibetans and also showed a 229
correlation of the prevalence of the dominant haplotypes for these genes with altitude of 230
residence. 231
232

11
Peng et al (62) SNP-typed 50 Tibetan individuals and compared the results with data from 233
HapMap Han Chinese, identifying EPAS1 as one of the genes that underwent a selective 234
sweep in Tibetans. They also sequenced the full length of the EPAS1 gene in these 235
individuals and showed marked allelic differentiation at this locus between Tibetans and Han 236
Chinese. They constructed haplotypes and further analysis allowed an estimation of a 237
divergence time for Tibetans which was six-fold greater than that of Yi et al in (96) and 238
consistent with previous archeological and genetic reports (1) (2, 101). They also genotyped 239
3 SNPs from each of EPAS1, EGLN1 and PPARA in a large cohort of 1334 Tibetans. They 240
found significant differences in allele frequencies for the EPAS1 SNPs between Tibetan and 241
Han Chinese/Japanese and also for one of the SNPs in EGLN1. 242
243
Wang et al (88) performed a number of tests of selection using genome-wide SNP data from 244
30 Tibetans living near Lhasa, and identified the EPAS1 locus as the strongest signal of 245
positive selection in the Tibetan genome. The second most significant genomic region with 246
evidence of positive selection was a region containing the EGLN1 gene. 247
248
Bigham et al (13) performed high density genome scans and applied four population genetic 249
statistics to identify selection-nominated candidate genes and candidate regions for two high-250
altitude populations, Andeans and Tibetans. These two populations were studied separately. 251
They found different patterns of positive selection for the two populations. In Tibetans, a 252
genomic region containing EPAS1 exhibited significant variation between Tibetans and the 253
lowland comparators, HapMap Asians. Interestingly, EGLN1 showed evidence of positive 254
selection in both Tibetans and Andeans although the SNP frequencies and haplotypes are 255
different between the two populations. 256
257

12
While EGLN1 was identified as undergoing selection in both Tibetans and Andeans, neither 258
EPAS1 or EGLN1 featured as candidates in recent genomic studies in Ethiopian highlanders 259
(3) (75) (37). Other – different – candidate genes have been found in these studies. PPARA, 260
however, which was previously reported as a selection candidate in Tibetans (80), was also 261
identified as a selection candidate gene in highland Ethiopians with a marginal association 262
with Hb concentration (75). Thus, different high-altitude populations evolved independently 263
to adapt to the challenges of a hypoxic environment. However, of interest is that in the study 264
by Huerta-Sanchez et al, the top candidate gene, DEC1, is functionally related to the oxygen-265
sensing pathway; it is transcriptionally regulated by HIF1α, directly binds HIF1α, 266
downregulates HIF1α and HIF2α protein expression and represses many HIF-target genes 267
(37). It is likely these inter-population differences in their genetic adaptation relate to the 268
length of high-altitude inhabitation, the severity of the hypoxic stimulus, the different genetic 269
backgrounds, different opportunities for admixture with lowlanders and evolutionary chance. 270
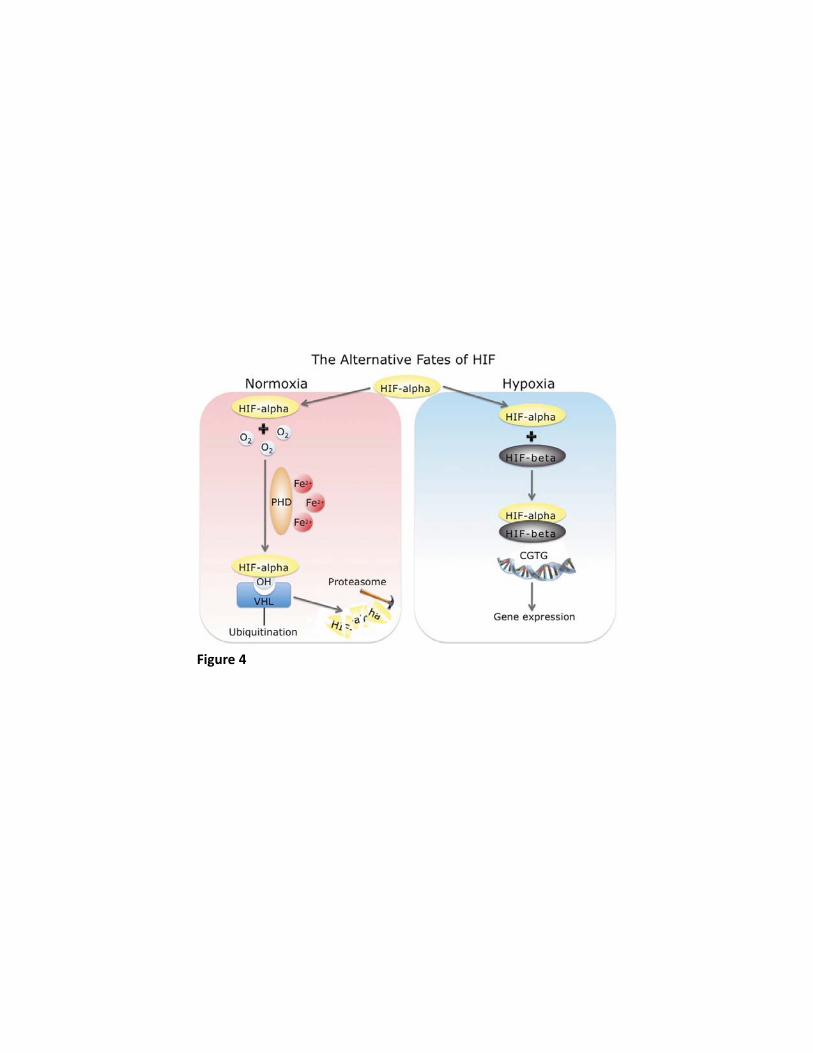
271
Additional genes which may have undergone selection in Tibetans with rather weaker signals 272
of selection do exist and have been reported in the various studies (29) (80) (96) . However, 273
what is striking when considering the convergent findings of all the studies to date, is the 274
independent identification of positive selection in Tibetans (from different geographic 275
locations) at genomic loci that involve EPAS1 (which codes for HIF2α) and EGLN1 (which 276
codes for PHD2), both key components of the hypoxia-inducible factor (HIF) pathway, thus 277
implicating these genes in the genetic adaptation of Tibetans to high altitude. 278
279
EPAS1 and EGLN1 – The HIF transcriptional pathway 280
Hypoxia-inducible factors (HIFs) are a family of transcription factors which coordinate 281
oxygen sensing and intracellular responses to hypoxia through regulation of expression of 282

13
hundreds of genes belonging to biological pathways such as energy metabolism, 283
angiogenesis, erythropoiesis, iron homeostasis, apoptosis and others (77). 284
285
EPAS1 encodes HIF2α (87), which is one of three HIF-alpha (α) subunit isoforms. In the 286
presence of oxygen, HIF-α is hydroxylated at 2 proline residues by prolyl hydroxylase 287
domain (PHD) enzymes, of which there are 3 isoforms – PHD1, PHD2 (coded by the EGLN1 288
gene) and PHD3 – in an oxygen-dependent manner (22, 38, 39). This hydroxylation enhances 289
the binding of the von Hippel-Lindau (VHL) protein to HIF-α which in turn enhances HIF-α 290
ubiquitination, resulting in its subsequent rapid proteasomal degradation (18, 39). In hypoxia, 291
the reduced hydroxylase activity of the PHD enzymes results in the stabilisation and 292
accumulation of the HIF-α subunits, which then heterodimerise with HIF-β and 293
transcriptionally activate target genes (Fig. 4). .Evidence for a central role of the HIF 294
pathway – and more specifically for EPAS1 (HIF2α) and EGLN1 (PHD2) – in systemic 295
erythropoiesis but also in other aspects of integrative physiology related to oxygen 296
homeostasis comes from studying patients with genetic abnormalities in the HIF system and 297
from genetic mouse models. The most extensively studied patients are those with Chuvash 298
polycythemia, who are homozygous for a hypomorphic allele of the von Hippel-Lindau 299
(VHL) tumor suppressor which impairs HIF1α and HIF2α degradation (4). These patients 300
have excessive erythrocytosis with high hemoglobin and hematocrit levels due to an up-301
regulation of erythropoietin production. . Apart from VHL, gain-of-function mutations in 302
EPAS1 leading to stabilisation of HIF2α (64, 65, 67) and mutations in EGLN1 associated 303
with diminished activity of PHD2 (45) (66) have also been identified in patients with 304
excessive erythrocytosis. 305
306

14
Further phenotyping has shown that patients with Chuvash Polycythemia have abnormal 307
cardiopulmonary physiology. They have pulmonary hypertension, increased resting 308
ventilation and upon exposure to acute systemic hypoxia they exhibit an exaggerated 309
pulmonary vascular response and an exaggerated ventilatory sensitivity (81). In addition, they 310
have abnormal metabolism during exercise with a reduced maximal exercise capacity, an 311
increased exercise-induced lactate accumulation and an early and marked phosphocreatine 312
depletion and acidosis in skeletal muscle (24). In contrast, patients with gain-of-function 313
HIF2α mutations have a more restricted cardiopulmonary phenotype of pulmonary 314
hypertension and exaggerated pulmonary vascular responses to acute hypoxia (23) (25). 315
316
Studies in genetically-modified mice further elucidated the role of the HIF pathway in 317
systemic physiology. Both HIF1α- and HIF2α-null mice die during embryogenesis (61, 74). 318
Postnatal deletion of HIF2α in a conditional mouse model resulted in anemia and 319
demonstrated that HIF2α – rather than HIF1α – is the critical isoform regulating 320
erythropoiesis in adults (28). Mice with heterozygous deficiency for functional HIF genes – 321
either HIF1α (98) or HIF2α (16) – developed less polycythemia and pulmonary hypertension 322
in response to hypoxia. Mouse models of Chuvash Polycythemia confirmed the phenotype of 323
increased ventilation and pulmonary hypertension found in humans, and further demonstrated 324
that the effects seen in the disease are primarily HIF2α-driven as opposed to HIF1α-driven 325
(33). Similarly, a mouse model of the human HIF2α gain-of-function mutation recapitulated 326
the phenotype of excessive erythrocytosis and pulmonary hypertension in a dose-dependent 327
manner (86). Knockout mice with heterozygous loss of HIF1α had depressed carotid body 328
oxygen sensitivity and abnormal ventilatory acclimatization to chronic hypoxia (44). In 329
contrast, HIF2α+/- mice manifested heightened carotid responses to hypoxia, highlighting 330
potentially different roles of HIF1 and HIF2 in systemic physiology (63). Germline PHD2 -/- 331

15
mice are not viable (85). PHD2 conditional knockout mice developed excessive 332
erythrocytosis and increased vascular growth (83, 84) and mice heterozygous for PHD2 had 333
increased ventilatory sensitivity to hypoxia and carotid body hyperplasia (14). 334
335
The above findings highlight the key role of the HIF pathway as a master controller of 336
oxygen sensing and regulator of systemic physiology in humans and mice. It is evident that 337
perturbations of the HIF pathway, whether leading to an upregulation or a downregulation of 338
the hypoxic HIF response, act on components of convective oxygen transport such as 339
ventilation, hypoxic pulmonary vascular response and erythropoiesis and produce phenotypic 340
changes. Interestingly, it is these components of oxygen transport that are distinctive in 341
Tibetan highlanders and make up their high-altitude phenotype, as previously mentioned, 342
alluding to a role of the HIF pathway in their adaptation to hypoxia. This is discussed below, 343
where we consider the potential physiological trait(s) upon which natural selection may have 344
acted. 345
346
Conclusions and future considerations 347
We have briefly reviewed the evidence that genetic variation in the HIF pathway affects 348
heamatopoietic and cardiopulmonary physiology in humans. The similarities between the 349
phenotype of lowland patients with genetic mutations in the HIF pathway that “upregulate” 350
the HIF response with the phenotype of patients with CMS at high altitude are remarkable. 351
Coupling these observations with the recent studies identifying EPAS1 and EGLN1 as loci 352
which have undergone recent positive selection in Tibetans (8, 13, 62, 80, 95, 96) suggests 353
that natural selection to hypoxia in Tibetans may have occurred through genetic variation in 354
the HIF pathway which operated on related integrative-physiology phenotypes. Several 355
questions remain unanswered. 356

16
357
One question relates to the identification of the “substrate” of natural selection, in other 358
words, the phenotype on which selection occurred. In some reports, the putatively selected 359
“Tibetan” genetic variants were associated with lower hemoglobin concentrations at high 360
altitude (8, 80, 96). Several possibilities exist. One is that blunted erythropoiesis may be 361
beneficial for highlanders and that low hemoglobin concentration may be the “selected” 362
phenotype. Arguments in favor of this are the association of excessive erythrocytosis with 363
CMS – which poses significant morbidity and mortality risks – and the remarkably low 364
prevalence of CMS in the Tibetan population. On the other hand, the higher hemoglobin and 365
hematocrit levels found in the Andean populations are not always associated with disease and 366
do not preclude the Andeans from being a growing population at high altitude, suggesting 367
that blunted erythropoiesis alone may not be a sufficient enough advantage to drive genetic 368
selection in Tibetans. Another possibility is that the phenotype of low hemoglobin 369
concentration is selected only as part of a more complex integrative phenotype that offers a 370
survival advantage at high altitude, owing to the pleiotropic effects of the HIF pathway, and 371
of EPAS1 in particular. It is even possible that the hematopoietic phenotype may actually be a 372
secondary outcome – or even a “side effect” – of the selection process which may have acted 373
upon some other aspect of the phenotype. Apart from affecting cardiopulmonary physiology, 374
EPAS1 plays roles in placenta and embryonic development (70) (52) (19) and may be 375
involved in IUGR (21). Of particular interest is that the placenta is one of the tissues where 376
high expression of HIF2α is found (72). The increased reproductive ability of Tibetans as 377
evident by the low pre- and post-natal mortality in comparison to Han Chinese high-altitude 378
residents, coupled with their resistance to IUGR, suggests that natural selection on EPAS1 379
may have also operated via effects during pregnancy on fetal growth. The study of 380

17
associations between birth weight at high altitude and genotype is a potentially fruitful area of 381
investigation. 382
383
A further question to consider is whether the effects of the genetic variation in EPAS1 and 384
EGLN1 selected for in Tibetans are functional and operate in generating a phenotype only in 385
conditions of severe and ongoing – i.e. for years – environmental hypoxia. It is of interest 386
that, while genetic variants in EPAS1 and EGLN1 were significantly correlated with 387
hemoglobin concentration in Tibetan highlanders, neither EPAS1 nor EGLN1 featured in any 388
of a number of genome-wide studies exploring genetic determinants of hemoglobin 389
concentration at sea level [22-24]. This suggests that either the genetic variation in EPAS1 or 390
EGLN1 relates to a hematopoietic phenotype only in conditions of severe and prolonged 391
environmental hypoxia or that the “functional” genetic variants in these genes are unique to 392
Tibetans, naturally selected by living for generations in hypoxia. This raises an interesting 393
question as to whether lessons can be learnt from the Tibetan paradigm on the role of the HIF 394
system near sea level. An ensuing fundamental question is to what degree the physiological 395
responses that lowland humans exhibit when exposed to high altitude – e.g. erythropoietic 396
and cardiopulmonary – can be considered adaptive to a reduction in ambient oxygen as 397
opposed to “mishaps” arising from oxygen sensing mechanisms that have been optimised to 398
operate at “normal” oxygen levels found at sea level. Accordingly, the HIF system in humans 399
may have been optimised to sculpt structure and physiological function using oxygen as a 400
guiding signal rather than to respond to environmental hypoxic challenges. For example, 401
reduced oxygen delivery to the kidneys is used as a guiding signal to stimulate erythropoietin 402
production through an upregulated HIF system in order to restore reductions in hemoglobin 403
or hematocrit levels, e.g. in anemia or hemorrhage, rather than to respond to environmental 404
oxygen changes. Similarly, hypoxic pulmonary vasoconstriction, in which the HIF system 405

18
plays a role, is a process evolved at sea level and is fundamental for the fetal circulation in 406
utero but it can be maladaptive in response to reductions in environmental oxygen. 407
408
Recently, Petousi et al (68) studied ethnic Tibetans living at sea level in the UK and provided 409
a first report of their phenotype in the absence of on-going, severe environmental hypoxia. In 410
this study Tibetans resident at sea level had a lower hemoglobin concentration, a higher 411
resting ventilation and blunted pulmonary vascular responses to both acute (minutes) and 412
sustained (8 hours) hypoxic challenges in comparison with Han Chinese lowlanders. The 413
physiological studies were complemented with cellular studies which showed that the relative 414
expression and the hypoxic induction of HIF-regulated genes were significantly lower in 415
peripheral blood lymphocytes from Tibetans compared with Han Chinese. Thus, this study 416
provided evidence that Tibetans possess a hypo-responsive HIF system and demonstrated that 417
a phenotype is present at sea level in the absence of on-going environmental hypoxia to 418
activate the HIF system. Indeed, Tibetans are the first humans in whom a hypo-responsive 419
HIF system has been demonstrated and this may represent an evolutionary re-setting of the 420
HIF system to operate within a hypoxic environment. 421
422
Nonetheless, for the genes that have undergone natural selection in Tibetans, neither the 423
functional variants nor the mechanisms by which they may be operating have been identified 424
to date. In the case of EPAS1, all the genetic variants reported in Tibetans to date are found in 425
non-coding regions, either within introns of EPAS1 or downstream of EPAS1; whole exome 426
sequencing (96) or targeted sequencing of the full-length EPAS1 gene (62) in Tibetans failed 427
to reveal any coding genetic variants as functional candidates. Thus, it is likely that 428
“functional” genetic variation in Tibetans might lie in regulatory regions and actually affect 429
transcription of the EPAS1 gene itself, as opposed to downstream HIF2α structure and 430

19
function. In keeping with such a possibility, Petousi et al (68) reported lower HIF2α mRNA 431
expression in peripheral blood lymphocytes from Tibetan than from Han Chinese volunteers, 432
which may be related to lower levels of transcription of EPAS1. In the case of EGLN1, two 433
coding variants have been described of particularly high frequencies in Tibetans (49) (48) 434
(94) (50) although their effect on PHD2 protein structure and function are not yet known. 435
Functional molecular studies are required to unravel the “causal” variants in the candidate 436
genes and their mechanism of action. Further exploration of Tibetan physiology by 437
integrating studies at the genomic level with functional molecular studies and whole-system 438
phenotyping has the potential to yield important additional insights into human evolution in 439
response to the environment of high-altitude hypoxia. 440
441
Acknowledgements 442
Nayia Petousi was funded by a Wellcome Trust Clinical Training Research Fellowship (grant 443
089457/Z/09/Z). 444
445
References 446
1. Aldenderfer M. Peopling the Tibetan plateau: insights from archaeology. High Alt Med Biol 447 12: 141-147, 2011. 448 2. Aldenderfer MS. Moving up in the world. American Scientist 91: 542-549, 2003. 449 3. Alkorta-Aranburu G, Beall CM, Witonsky DB, Gebremedhin A, Pritchard JK, and Di Rienzo 450 A. The genetic architecture of adaptations to high altitude in Ethiopia. PLoS Genet 8: e1003110, 451 2012. 452 4. Ang SO, Chen H, Hirota K, Gordeuk VR, Jelinek J, Guan Y, Liu E, Sergueeva AI, Miasnikova 453 GY, Mole D, Maxwell PH, Stockton DW, Semenza GL, and Prchal JT. Disruption of oxygen 454 homeostasis underlies congenital Chuvash polycythemia. Nat Genet 32: 614-621, 2002. 455 5. Beall CM. Genetic changes in Tibet. High Alt Med Biol 12: 101-102, 2011. 456 6. Beall CM. Two routes to functional adaptation: Tibetan and Andean high-altitude natives. 457 Proc Natl Acad Sci U S A 104 Suppl 1: 8655-8660, 2007. 458 7. Beall CM, Brittenham GM, Strohl KP, Blangero J, Williams-Blangero S, Goldstein MC, 459 Decker MJ, Vargas E, Villena M, Soria R, Alarcon AM, and Gonzales C. Hemoglobin concentration of 460 high-altitude Tibetans and Bolivian Aymara. Am J Phys Anthropol 106: 385-400, 1998. 461 8. Beall CM, Cavalleri GL, Deng L, Elston RC, Gao Y, Knight J, Li C, Li JC, Liang Y, McCormack M, 462 Montgomery HE, Pan H, Robbins PA, Shianna KV, Tam SC, Tsering N, Veeramah KR, Wang W, 463 Wangdui P, Weale ME, Xu Y, Xu Z, Yang L, Zaman MJ, Zeng C, Zhang L, Zhang X, Zhaxi P, and Zheng 464

20
YT. Natural selection on EPAS1 (HIF2alpha) associated with low hemoglobin concentration in Tibetan 465 highlanders. Proc Natl Acad Sci U S A 107: 11459-11464, 2010. 466 9. Beall CM, Decker MJ, Brittenham GM, Kushner I, Gebremedhin A, and Strohl KP. An 467 Ethiopian pattern of human adaptation to high-altitude hypoxia. Proc Natl Acad Sci U S A 99: 17215-468 17218, 2002. 469 10. Beall CM, Laskowski D, Strohl KP, Soria R, Villena M, Vargas E, Alarcon AM, Gonzales C, 470 and Erzurum SC. Pulmonary nitric oxide in mountain dwellers. Nature 414: 411-412, 2001. 471 11. Beall CM, Strohl KP, Blangero J, Williams-Blangero S, Almasy LA, Decker MJ, Worthman 472 CM, Goldstein MC, Vargas E, Villena M, Soria R, Alarcon AM, and Gonzales C. Ventilation and 473 hypoxic ventilatory response of Tibetan and Aymara high altitude natives. Am J Phys Anthropol 104: 474 427-447, 1997. 475 12. Beall CM, Strohl KP, Blangero J, Williams-Blangero S, Decker MJ, Brittenham GM, and 476 Goldstein MC. Quantitative genetic analysis of arterial oxygen saturation in Tibetan highlanders. 477 Hum Biol 69: 597-604, 1997. 478 13. Bigham A, Bauchet M, Pinto D, Mao X, Akey JM, Mei R, Scherer SW, Julian CG, Wilson MJ, 479 Lopez Herraez D, Brutsaert T, Parra EJ, Moore LG, and Shriver MD. Identifying signatures of natural 480 selection in Tibetan and Andean populations using dense genome scan data. PLoS Genet 6: 2010. 481 14. Bishop T, Talbot NP, Turner PJ, Nicholls LG, Pascual A, Hodson EJ, Douglas G, Fielding JW, 482 Smith TG, Demetriades M, Schofield CJ, Robbins PA, Pugh CW, Buckler KJ, and Ratcliffe PJ. Carotid 483 body hyperplasia and enhanced ventilatory responses to hypoxia in mice with heterozygous 484 deficiency of PHD2. J Physiol 591: 3565-3577, 2013. 485 15. Biswas S, and Akey JM. Genomic insights into positive selection. Trends Genet 22: 437-446, 486 2006. 487 16. Brusselmans K, Compernolle V, Tjwa M, Wiesener MS, Maxwell PH, Collen D, and 488 Carmeliet P. Heterozygous deficiency of hypoxia-inducible factor-2alpha protects mice against 489 pulmonary hypertension and right ventricular dysfunction during prolonged hypoxia. J Clin Invest 490 111: 1519-1527, 2003. 491 17. Chen D, Zhou X, Zhu Y, Zhu T, and Wang J. [Comparison study on uterine and umbilical 492 artery blood flow during pregnancy at high altitude and at low altitude]. Zhonghua Fu Chan Ke Za Zhi 493 37: 69-71, 2002. 494 18. Cockman ME, Masson N, Mole DR, Jaakkola P, Chang GW, Clifford SC, Maher ER, Pugh CW, 495 Ratcliffe PJ, and Maxwell PH. Hypoxia inducible factor-alpha binding and ubiquitylation by the von 496 Hippel-Lindau tumor suppressor protein. J Biol Chem 275: 25733-25741, 2000. 497 19. Cowden Dahl KD, Fryer BH, Mack FA, Compernolle V, Maltepe E, Adelman DM, Carmeliet 498 P, and Simon MC. Hypoxia-inducible factors 1alpha and 2alpha regulate trophoblast differentiation. 499 Mol Cell Biol 25: 10479-10491, 2005. 500 20. Curran LS, Zhuang J, Sun SF, and Moore LG. Ventilation and hypoxic ventilatory 501 responsiveness in Chinese-Tibetan residents at 3,658 m. J Appl Physiol 83: 2098-2104, 1997. 502 21. Dai SY, Kanenishi K, Ueno M, Sakamoto H, and Hata T. Hypoxia-inducible factor-2alpha is 503 involved in enhanced apoptosis in the placenta from pregnancies with fetal growth restriction. 504 Pathol Int 54: 843-849, 2004. 505 22. Epstein AC, Gleadle JM, McNeill LA, Hewitson KS, O'Rourke J, Mole DR, Mukherji M, 506 Metzen E, Wilson MI, Dhanda A, Tian YM, Masson N, Hamilton DL, Jaakkola P, Barstead R, Hodgkin 507 J, Maxwell PH, Pugh CW, Schofield CJ, and Ratcliffe PJ. C. elegans EGL-9 and mammalian homologs 508 define a family of dioxygenases that regulate HIF by prolyl hydroxylation. Cell 107: 43-54, 2001. 509 23. Formenti F, Beer PA, Croft QP, Dorrington KL, Gale DP, Lappin TR, Lucas GS, Maher ER, 510 Maxwell PH, McMullin MF, O'Connor DF, Percy MJ, Pugh CW, Ratcliffe PJ, Smith TG, Talbot NP, 511 and Robbins PA. Cardiopulmonary function in two human disorders of the hypoxia-inducible factor 512 (HIF) pathway: von Hippel-Lindau disease and HIF-2{alpha} gain-of-function mutation. FASEB J 25: 513 2001-2011, 2011. 514

21
24. Formenti F, Constantin-Teodosiu D, Emmanuel Y, Cheeseman J, Dorrington KL, Edwards 515 LM, Humphreys SM, Lappin TR, McMullin MF, McNamara CJ, Mills W, Murphy JA, O'Connor DF, 516 Percy MJ, Ratcliffe PJ, Smith TG, Treacy M, Frayn KN, Greenhaff PL, Karpe F, Clarke K, and Robbins 517 PA. Regulation of human metabolism by hypoxia-inducible factor. Proc Natl Acad Sci U S A 107: 518 12722-12727, 2010. 519 25. Gale DP, Harten SK, Reid CD, Tuddenham EG, and Maxwell PH. Autosomal dominant 520 erythrocytosis and pulmonary arterial hypertension associated with an activating HIF2 alpha 521 mutation. Blood 112: 919-921, 2008. 522 26. Garruto RM, Chin CT, Weitz CA, Liu JC, Liu RL, and He X. Hematological differences during 523 growth among Tibetans and Han Chinese born and raised at high altitude in Qinghai, China. Am J 524 Phys Anthropol 122: 171-183, 2003. 525 27. Groves BM, Droma T, Sutton JR, McCullough RG, McCullough RE, Zhuang J, Rapmund G, 526 Sun S, Janes C, and Moore LG. Minimal hypoxic pulmonary hypertension in normal Tibetans at 3,658 527 m. J Appl Physiol 74: 312-318, 1993. 528 28. Gruber M, Hu CJ, Johnson RS, Brown EJ, Keith B, and Simon MC. Acute postnatal ablation of 529 Hif-2alpha results in anemia. Proc Natl Acad Sci U S A 104: 2301-2306, 2007. 530 29. Gu M, Dong X, Shi L, Lin K, Huang X, and Chu J. Differences in mtDNA whole sequence 531 between Tibetan and Han populations suggesting adaptive selection to high altitude. Gene 496: 37-532 44, 2012. 533 30. Gupta ML, Rao KS, Anand IS, Banerjee AK, and Boparai MS. Lack of smooth muscle in the 534 small pulmonary arteries of the native Ladakhi. Is the Himalayan highlander adapted? Am Rev Respir 535 Dis 145: 1201-1204, 1992. 536 31. Haas JD. Human adaptability approach to nutritional assessment: a Bolivian example. Fed 537 Proc 40: 2577-2582, 1981. 538 32. Hackett PH, Reeves JT, Reeves CD, Grover RF, and Rennie D. Control of breathing in Sherpas 539 at low and high altitude. J Appl Physiol 49: 374-379, 1980. 540 33. Hickey MM, Richardson T, Wang T, Mosqueira M, Arguiri E, Yu H, Yu QC, Solomides CC, 541 Morrisey EE, Khurana TS, Christofidou-Solomidou M, and Simon MC. The von Hippel-Lindau 542 Chuvash mutation promotes pulmonary hypertension and fibrosis in mice. J Clin Invest 120: 827-839, 543 2010. 544 34. Hoit BD, Dalton ND, Erzurum SC, Laskowski D, Strohl KP, and Beall CM. Nitric oxide and 545 cardiopulmonary hemodynamics in Tibetan highlanders. J Appl Physiol 99: 1796-1801, 2005. 546 35. Hoit BD, Dalton ND, Gebremedhin A, Janocha A, Zimmerman PA, Zimmerman AM, Strohl 547 KP, Erzurum SC, and Beall CM. Elevated pulmonary artery pressure among Amhara highlanders in 548 Ethiopia. Am J Hum Biol 23: 168-176, 2011 549 550 36. Huang ZR, Zhu SC, Ba ZF, and Hu ST. Ventilatory Control in Tibetan highlanders. In: 551 Geological and Ecological Studies og Qinghai-Xizang Plateau, edited by Liu DS. Beijing, PRC: Science 552 Press, 1981, p. 1363-1369. 553 37. Huerta-Sanchez E, Degiorgio M, Pagani L, Tarekegn A, Ekong R, Antao T, Cardona A, 554 Montgomery HE, Cavalleri GL, Robbins PA, Weale ME, Bradman N, Bekele E, Kivisild T, Tyler-Smith 555 C, and Nielsen R. Genetic signatures reveal high-altitude adaptation in a set of ethiopian 556 populations. Mol Biol Evol 30: 1877-1888, 2013. 557 38. Ivan M, Haberberger T, Gervasi DC, Michelson KS, Gunzler V, Kondo K, Yang H, Sorokina I, 558 Conaway RC, Conaway JW, and Kaelin WG, Jr. Biochemical purification and pharmacological 559 inhibition of a mammalian prolyl hydroxylase acting on hypoxia-inducible factor. Proc Natl Acad Sci U 560 S A 99: 13459-13464, 2002. 561 39. Jaakkola P, Mole DR, Tian YM, Wilson MI, Gielbert J, Gaskell SJ, Kriegsheim A, Hebestreit 562 HF, Mukherji M, Schofield CJ, Maxwell PH, Pugh CW, and Ratcliffe PJ. Targeting of HIF-alpha to the 563 von Hippel-Lindau ubiquitylation complex by O2-regulated prolyl hydroxylation. Science 292: 468-564 472, 2001. 565

22
40. Jaillard AS, Hommel M, and Mazetti P. Prevalence of stroke at high altitude (3380 m) in 566 Cuzco, a town of Peru. A population-based study. Stroke 26: 562-568, 1995. 567 41. Jensen GM, and Moore LG. The effect of high altitude and other risk factors on birthweight: 568 independent or interactive effects? Am J Public Health 87: 1003-1007, 1997. 569 42. Julian CG, Wilson MJ, Lopez M, Yamashiro H, Tellez W, Rodriguez A, Bigham AW, Shriver 570 MD, Rodriguez C, Vargas E, and Moore LG. Augmented uterine artery blood flow and oxygen 571 delivery protect Andeans from altitude-associated reductions in fetal growth. Am J Physiol Regul 572 Integr Comp Physiol 296: R1564-1575, 2009. 573 43. Julian CG, Wilson MJ, and Moore LG. Evolutionary adaptation to high altitude: a view from 574 in utero. Am J Hum Biol 21: 614-622, 2009. 575 44. Kline DD, Peng YJ, Manalo DJ, Semenza GL, and Prabhakar NR. Defective carotid body 576 function and impaired ventilatory responses to chronic hypoxia in mice partially deficient for 577 hypoxia-inducible factor 1 alpha. Proc Natl Acad Sci U S A 99: 821-826, 2002. 578 45. Ladroue C, Hoogewijs D, Gad S, Carcenac R, Storti F, Barrois M, Gimenez-Roqueplo AP, 579 Leporrier M, Casadevall N, Hermine O, Kiladjian JJ, Baruchel A, Fakhoury F, Bressac-de Paillerets B, 580 Feunteun J, Mazure N, Pouyssegur J, Wenger RH, Richard S, and Gardie B. Distinct deregulation of 581 the hypoxia inducible factor by PHD2 mutants identified in germline DNA of patients with 582 polycythemia. Haematologica 97: 9-14, 2012. 583 46. Lahiri S, Kao FF, Velasque.T, Martinez C, and Pezzia W. Irreversible Blunted Respiratory 584 Sensitivity to Hypoxia in High Altitude Natives. Respiration Physiology 6: 360-&, 1969. 585 47. Leon-Velarde F, Maggiorini M, Reeves JT, Aldashev A, Asmus I, Bernardi L, Ge RL, Hackett 586 P, Kobayashi T, Moore LG, Penaloza D, Richalet JP, Roach R, Wu T, Vargas E, Zubieta-Castillo G, and 587 Zubieta-Calleja G. Consensus statement on chronic and subacute high altitude diseases. High Alt 588 Med Biol 6: 147-157, 2005. 589 48. Lorenzo FPV, Swierczek S, Huff CD, and Prchal JT. The Tibetan PHD2 Polymorphism Asp4Glu 590 Is Associated with Hypersensitivity of Erythroid Progenitors to EPO and Upregulation of HIF-1 591 Regulated Genes Hexokinase (HK1) and Glucose Transporter 1 (SLC2A/GLUT1). In: 53rd ASH Annual 592 Meeting and Exposition, 2011. 593 49. Lorenzo FRV, Simonson TS, Yang Y, Ge R, and Prchal JT. A Novel PHD2 Mutation Associated 594 with Tibetan Genetic Adaptation to High Altitude Hypoxia. In: 52nd ASH Annual Meeting and 595 Exposition, 2010. 596 50. Lorenzo FRV ST, Yang Y, Ge R, Prchal JT. A Novel PHD2 Mutation Associated with Tibetan 597 Genetic Adaptation to High Altitude Hypoxia. In: 54th ASH Annual Meeting and Exposition2012. 598 51. MacInnis MJ, and Rupert JL. 'ome on the Range: altitude adaptation, positive selection, and 599 Himalayan genomics. High Alt Med Biol 12: 133-139, 2011. 600 52. Meade ES, Ma YY, and Guller S. Role of hypoxia-inducible transcription factors 1alpha and 601 2alpha in the regulation of plasminogen activator inhibitor-1 expression in a human trophoblast cell 602 line. Placenta 28: 1012-1019, 2007. 603 53. Monge C, Arregui CA, and Leonvelarde F. Pathophysiology and Epidemiology of Chronic 604 Mountain-Sickness. International Journal of Sports Medicine 13: S79-S81, 1992. 605 54. Moore LG, Armaza F, Villena M, and Vargas E. Comparative aspects of high-altitude 606 adaptation in human populations. Adv Exp Med Biol 475: 45-62, 2000. 607 55. Moore LG, Curran-Everett L, Droma TS, Groves BM, McCullough RE, McCullough RG, Sun 608 SF, Sutton JR, Zamudio S, and Zhuang JG. Are Tibetans better adapted? Int J Sports Med 13 Suppl 1: 609 S86-88, 1992. 610 56. Moore LG, Shriver M, Bemis L, Hickler B, Wilson M, Brutsaert T, Parra E, and Vargas E. 611 Maternal adaptation to high-altitude pregnancy: an experiment of nature--a review. Placenta 25 612 Suppl A: S60-71, 2004. 613 57. Moore LG, Young D, McCullough RE, Droma T, and Zamudio S. Tibetan protection from 614 intrauterine growth restriction (IUGR) and reproductive loss at high altitude. Am J Hum Biol 13: 635-615 644, 2001. 616

23
58. Moore LG, Zamudio S, Zhuang J, Sun S, and Droma T. Oxygen transport in tibetan women 617 during pregnancy at 3,658 m. Am J Phys Anthropol 114: 42-53, 2001. 618 59. Motley HL, Cournand A, and et al. The influence of short periods of induced acute anoxia 619 upon pulmonary artery pressures in man. Am J Physiol 150: 315-320, 1947. 620 60. Penaloza D, and Arias-Stella J. The heart and pulmonary circulation at high altitudes: 621 healthy highlanders and chronic mountain sickness. Circulation 115: 1132-1146, 2007. 622 61. Peng J, Zhang L, Drysdale L, and Fong GH. The transcription factor EPAS-1/hypoxia-inducible 623 factor 2alpha plays an important role in vascular remodeling. Proc Natl Acad Sci U S A 97: 8386-8391, 624 2000. 625 62. Peng Y, Yang Z, Zhang H, Cui C, Qi X, Luo X, Tao X, Wu T, Ouzhuluobu, Basang, 626 Ciwangsangbu, Danzengduojie, Chen H, Shi H, and Su B. Genetic variations in Tibetan populations 627 and high-altitude adaptation at the Himalayas. Mol Biol Evol 28: 1075-1081, 2011. 628 63. Peng YJ, Nanduri J, Khan SA, Yuan G, Wang N, Kinsman B, Vaddi DR, Kumar GK, Garcia JA, 629 Semenza GL, and Prabhakar NR. Hypoxia-inducible factor 2alpha (HIF-2alpha) heterozygous-null 630 mice exhibit exaggerated carotid body sensitivity to hypoxia, breathing instability, and hypertension. 631 Proc Natl Acad Sci U S A 108: 3065-3070, 2011. 632 64. Percy MJ, Beer PA, Campbell G, Dekker AW, Green AR, Oscier D, Rainey MG, van Wijk R, 633 Wood M, Lappin TR, McMullin MF, and Lee FS. Novel exon 12 mutations in the HIF2A gene 634 associated with erythrocytosis. Blood 111: 5400-5402, 2008. 635 65. Percy MJ, Chung YJ, Harrison C, Mercieca J, Hoffbrand AV, Dinardo CL, Santos PC, Fonseca 636 GH, Gualandro SF, Pereira AC, Lappin TR, McMullin MF, and Lee FS. Two new mutations in the 637 HIF2A gene associated with erythrocytosis. Am J Hematol 87: 439-442, 2012. 638 66. Percy MJ, Furlow PW, Beer PA, Lappin TR, McMullin MF, and Lee FS. A novel 639 erythrocytosis-associated PHD2 mutation suggests the location of a HIF binding groove. Blood 110: 640 2193-2196, 2007. 641 67. Percy MJ, Furlow PW, Lucas GS, Li X, Lappin TR, McMullin MF, and Lee FS. A gain-of-642 function mutation in the HIF2A gene in familial erythrocytosis. N Engl J Med 358: 162-168, 2008. 643 68. Petousi N, Croft QPP, Cavalleri GL, Cheng HY, Formenti F, Ishida K, Lunn D, McCormack M, 644 Shianna KV, Talbot NP, Ratcliffe PJ, and Robbins PA. Tibetans living at sea level have a 645 hyporesponsive hypoxia-inducible factor (HIF) system and blunted physiological responses to 646 hypoxia. 2013. 647 69. Powell FL, Milsom WK, and Mitchell GS. Time domains of the hypoxic ventilatory response. 648 Respir Physiol 112: 123-134, 1998. 649 70. Pringle KG, Kind KL, Sferruzzi-Perri AN, Thompson JG, and Roberts CT. Beyond oxygen: 650 complex regulation and activity of hypoxia inducible factors in pregnancy. Hum Reprod Update 16: 651 415-431, 2010. 652 71. Qi X, Cui C, Peng Y, Zhang X, Yang Z, Zhong H, Zhang H, Xiang K, Cao X, Wang Y, 653 Ouzhuluobu, Basang, Ciwangsangbu, Bianba, Gonggalanzi, Wu T, Chen H, Shi H, and Su B. Genetic 654 evidence of paleolithic colonization and neolithic expansion of modern humans on the tibetan 655 plateau. Mol Biol Evol 30: 1761-1778, 2013. 656 72. Rajakumar A, and Conrad KP. Expression, ontogeny, and regulation of hypoxia-inducible 657 transcription factors in the human placenta. Biol Reprod 63: 559-569, 2000. 658 73. Richalet JP, Souberbielle JC, Antezana AM, Dechaux M, Le Trong JL, Bienvenu A, Daniel F, 659 Blanchot C, and Zittoun J. Control of erythropoiesis in humans during prolonged exposure to the 660 altitude of 6,542 m. Am J Physiol 266: R756-764, 1994. 661 74. Ryan HE, Lo J, and Johnson RS. HIF-1 alpha is required for solid tumor formation and 662 embryonic vascularization. EMBO J 17: 3005-3015, 1998. 663 75. Scheinfeldt LB, Soi S, Thompson S, Ranciaro A, Woldemeskel D, Beggs W, Lambert C, Jarvis 664 JP, Abate D, Belay G, and Tishkoff SA. Genetic adaptation to high altitude in the Ethiopian highlands. 665 Genome Biol 13: R1, 2012. 666

24
76. Scheinfeldt LB, and Tishkoff SA. Living the high life: high-altitude adaptation. Genome Biol 667 11: 133, 2010. 668 77. Schofield CJ, and Ratcliffe PJ. Oxygen sensing by HIF hydroxylases. Nat Rev Mol Cell Biol 5: 669 343-354, 2004. 670 78. Severinghaus JW, Bainton CR, and Carcelen A. Respiratory insensitivity to hypoxia in 671 chronically hypoxic man. Respir Physiol 1: 308-334, 1966. 672 79. Simonson TS, McClain DA, Jorde LB, and Prchal JT. Genetic determinants of Tibetan high-673 altitude adaptation. Hum Genet 131: 527-533. 674 80. Simonson TS, Yang Y, Huff CD, Yun H, Qin G, Witherspoon DJ, Bai Z, Lorenzo FR, Xing J, 675 Jorde LB, Prchal JT, and Ge R. Genetic evidence for high-altitude adaptation in Tibet. Science 329: 676 72-75, 2010. 677 81. Smith TG, Brooks JT, Balanos GM, Lappin TR, Layton DM, Leedham DL, Liu C, Maxwell PH, 678 McMullin MF, McNamara CJ, Percy MJ, Pugh CW, Ratcliffe PJ, Talbot NP, Treacy M, and Robbins 679 PA. Mutation of von Hippel-Lindau tumour suppressor and human cardiopulmonary physiology. PLoS 680 Med 3: e290, 2006. 681 82. Suzuki Y. Statistical methods for detecting natural selection from genomic data. Genes Genet 682 Syst 85: 359-376, 2010. 683 83. Takeda K, Aguila HL, Parikh NS, Li X, Lamothe K, Duan LJ, Takeda H, Lee FS, and Fong GH. 684 Regulation of adult erythropoiesis by prolyl hydroxylase domain proteins. Blood 111: 3229-3235, 685 2008. 686 84. Takeda K, Cowan A, and Fong GH. Essential role for prolyl hydroxylase domain protein 2 in 687 oxygen homeostasis of the adult vascular system. Circulation 116: 774-781, 2007. 688 85. Takeda K, Ho VC, Takeda H, Duan LJ, Nagy A, and Fong GH. Placental but not heart defects 689 are associated with elevated hypoxia-inducible factor alpha levels in mice lacking prolyl hydroxylase 690 domain protein 2. Mol Cell Biol 26: 8336-8346, 2006. 691 86. Tan Q, Kerestes H, Percy MJ, Pietrofesa R, Chen L, Khurana TS, Christofidou-Solomidou M, 692 Lappin TR, and Lee FS. Erythrocytosis and Pulmonary Hypertension in a Mouse Model of Human 693 HIF2A Gain-of-Function Mutation. J Biol Chem 2013. 694 87. Tian H, McKnight SL, and Russell DW. Endothelial PAS domain protein 1 (EPAS1), a 695 transcription factor selectively expressed in endothelial cells. Genes Dev 11: 72-82, 1997. 696 88. Wang B, Zhang YB, Zhang F, Lin H, Wang X, Wan N, Ye Z, Weng H, Zhang L, Li X, Yan J, 697 Wang P, Wu T, Cheng L, Wang J, Wang DM, Ma X, and Yu J. On the origin of Tibetans and their 698 genetic basis in adapting high-altitude environments. PLoS One 6: e17002, 2011. 699 89. Wilson MJ, Lopez M, Vargas M, Julian C, Tellez W, Rodriguez A, Bigham A, Armaza JF, 700 Niermeyer S, Shriver M, Vargas E, and Moore LG. Greater uterine artery blood flow during 701 pregnancy in multigenerational (Andean) than shorter-term (European) high-altitude residents. Am J 702 Physiol Regul Integr Comp Physiol 293: R1313-1324, 2007. 703 90. Winslow RM, Chapman KW, Gibson CC, Samaja M, Monge CC, Goldwasser E, Sherpa M, 704 Blume FD, and Santolaya R. Different hematologic responses to hypoxia in Sherpas and Quechua 705 Indians. J Appl Physiol 66: 1561-1569, 1989. 706 91. Wu T, Wang X, Wei C, Cheng H, Li Y, Ge D, Zhao H, Young P, Li G, and Wang Z. Hemoglobin 707 levels in Qinghai-Tibet: different effects of gender for Tibetans vs. Han. J Appl Physiol 98: 598-604, 708 2005. 709 92. Wu TY, Li W, Li Y, Ge RL, Cheng Q, Wang S, Zhao G, Wei L, Jin Y, and Gen-Don. Epidemiology 710 of Chronic Mountain Sickness: Ten years' study in Qinghai, Tibet. Progress in Mountain Medicine and 711 High Altitude Physiology. In: 3rd World Congress in Mountain Medicine and High altitude Physiology, 712 edited by Committee P. Matsumoto, Japan: 1998a, p. 120-125. 713 93. Wu TY, Zhang Q, Jin B, Xu F, Cheng Q, and Wan X. Chronic Mountain Sickness (Monge's 714 disease): An observation in Qinghai-Tibet plateau. In: High Altitude Medicine, edited by Ueda G. 715 Matsumoto: Shinshu University Press, 1992, p. 314-324. 716

25
94. Xiang K, Ouzhuluobu, Peng Y, Yang Z, Zhang X, Cui C, Zhang H, Li M, Zhang Y, Bianba, 717 Gonggalanzi, Basang, Ciwangsangbu, Wu T, Chen H, Shi H, Qi X, and Su B. Identification of a 718 Tibetan-Specific Mutation in the Hypoxic Gene EGLN1 and Its Contribution to High-Altitude 719 Adaptation. Mol Biol Evol 30: 1889-1898, 2013. 720 95. Xu S, Li S, Yang Y, Tan J, Lou H, Jin W, Yang L, Pan X, Wang J, Shen Y, Wu B, Wang H, and Jin 721 L. A genome-wide search for signals of high-altitude adaptation in Tibetans. Mol Biol Evol 28: 1003-722 1011, 2011. 723 96. Yi X, Liang Y, Huerta-Sanchez E, Jin X, Cuo ZX, Pool JE, Xu X, Jiang H, Vinckenbosch N, 724 Korneliussen TS, Zheng H, Liu T, He W, Li K, Luo R, Nie X, Wu H, Zhao M, Cao H, Zou J, Shan Y, Li S, 725 Yang Q, Asan, Ni P, Tian G, Xu J, Liu X, Jiang T, Wu R, Zhou G, Tang M, Qin J, Wang T, Feng S, Li G, 726 Huasang, Luosang J, Wang W, Chen F, Wang Y, Zheng X, Li Z, Bianba Z, Yang G, Wang X, Tang S, Gao 727 G, Chen Y, Luo Z, Gusang L, Cao Z, Zhang Q, Ouyang W, Ren X, Liang H, Huang Y, Li J, Bolund L, 728 Kristiansen K, Li Y, Zhang Y, Zhang X, Li R, Yang H, Nielsen R, and Wang J. Sequencing of 50 human 729 exomes reveals adaptation to high altitude. Science 329: 75-78, 2010. 730 97. Yip R. Altitude and birth weight. J Pediatr 111: 869-876, 1987. 731 98. Yu AY, Shimoda LA, Iyer NV, Huso DL, Sun X, McWilliams R, Beaty T, Sham JS, Wiener CM, 732 Sylvester JT, and Semenza GL. Impaired physiological responses to chronic hypoxia in mice partially 733 deficient for hypoxia-inducible factor 1alpha. J Clin Invest 103: 691-696, 1999. 734 99. Zamudio S, Droma T, Norkyel KY, Acharya G, Zamudio JA, Niermeyer SN, and Moore LG. 735 Protection from intrauterine growth retardation in Tibetans at high altitude. Am J Phys Anthropol 91: 736 215-224, 1993. 737 100. Zamudio S, Postigo L, Illsley NP, Rodriguez C, Heredia G, Brimacombe M, Echalar L, Torricos 738 T, Tellez W, Maldonado I, Balanza E, Alvarez T, Ameller J, and Vargas E. Maternal oxygen delivery is 739 not related to altitude- and ancestry-associated differences in human fetal growth. J Physiol 582: 740 883-895, 2007. 741 101. Zhao M, Kong QP, Wang HW, Peng MS, Xie XD, Wang WZ, Jiayang, Duan JG, Cai MC, Zhao 742 SN, Cidanpingcuo, Tu YQ, Wu SF, Yao YG, Bandelt HJ, and Zhang YP. Mitochondrial genome 743 evidence reveals successful Late Paleolithic settlement on the Tibetan Plateau. Proceedings of the 744 National Academy of Sciences of the United States of America 106: 21230-21235, 2009. 745 102. Zhuang J, Droma T, Sun S, Janes C, McCullough RE, McCullough RG, Cymerman A, Huang 746 SY, Reeves JT, and Moore LG. Hypoxic ventilatory responsiveness in Tibetan compared with Han 747 residents of 3,658 m. J Appl Physiol 74: 303-311, 1993. 748 749 750 751 752 Figure legends 753 754 Figure 1: Cumulative frequency distribution of hemoglobin concentrations in US residents at 755
sea level and Tibetan and Bolivian Aymara natives at 4000 m. (A) displays results for males 756
and (B) for females. (Reproduced from Beall et al (7) with permission from the American 757
Journal of Physical Anthropology.) 758
759
Figure 2: Birth weight plotted against altitude of residence for Tibetans and Han Chinese, 760
showing a significantly smaller birth weight fall with increasing altitude in Tibetans than in 761

26
Han Chinese. Values are means and error bars represent S.E.M. (Reproduced from Moore et 762
al (57) with permission from the American Journal of Human Biology.) 763
764
Figure 3: Genome-wide allelic differentiation scans comparing Tibetan highlanders and 765
HapMap Han Chinese. Eight SNPs near EPAS1 achieved genome-wide significance. 766
(Reproduced from Beall et al (8) with permission from PNAS.) 767
768
Figure 4: Schematic representation of the regulation of HIF-alpha by hypoxia. In the 769
presence of oxygen (O2) and iron (Fe2+), prolyl hydroxylase domain (PHD) enzymes 770
hydroxylate specific proline residues in HIF-alpha, increasing binding of the Von Hippel-771
Lindau tumor suppressor protein (VHL). This targets HIF-alpha for ubiquitination and 772
mediates its proteosmal degradation. In hypoxia, HIF-alpha accumulates, dimerises with HIF-773
beta, binds to DNA and transcriptionally regulates hypoxia-responsive genes. (Courtesy of Dr 774
Federico Formenti, University of Oxford). 775
Figure�1
Figure�2
Figure�3
Figure�4
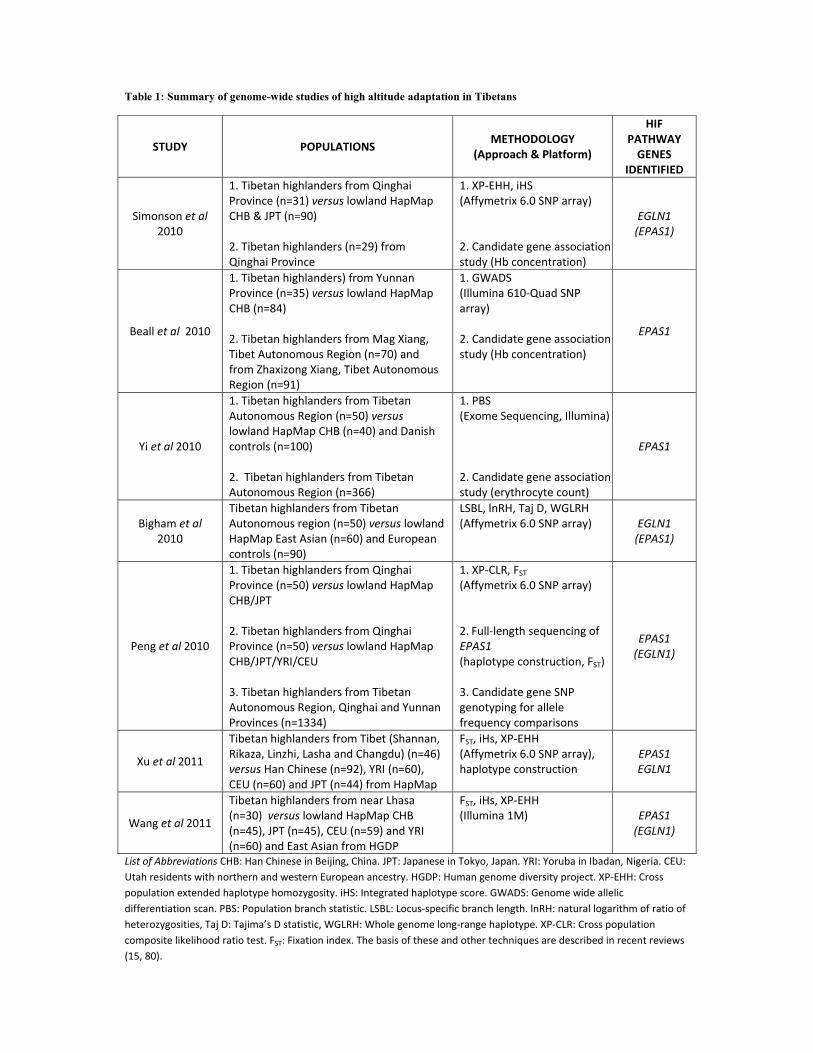
Table 1: Summary of genome-wide studies of high altitude adaptation in Tibetans
STUDY POPULATIONS METHODOLOGY
(Approach & Platform)
HIF PATHWAY
GENES IDENTIFIED
Simonson et al 2010
1. Tibetan highlanders from Qinghai Province (n=31) versus lowland HapMap CHB & JPT (n=90) 2. Tibetan highlanders (n=29) from Qinghai Province
1. XP-EHH, iHS (Affymetrix 6.0 SNP array) 2. Candidate gene association study (Hb concentration)
EGLN1 (EPAS1)
Beall et al 2010
1. Tibetan highlanders) from Yunnan Province (n=35) versus lowland HapMap CHB (n=84) 2. Tibetan highlanders from Mag Xiang, Tibet Autonomous Region (n=70) and from Zhaxizong Xiang, Tibet Autonomous Region (n=91)
1. GWADS (Illumina 610-Quad SNP array) 2. Candidate gene association study (Hb concentration)
EPAS1
Yi et al 2010
1. Tibetan highlanders from Tibetan Autonomous Region (n=50) versus lowland HapMap CHB (n=40) and Danish controls (n=100) 2. Tibetan highlanders from Tibetan Autonomous Region (n=366)
1. PBS(Exome Sequencing, Illumina) 2. Candidate gene association study (erythrocyte count)
EPAS1
Bigham et al 2010
Tibetan highlanders from Tibetan Autonomous region (n=50) versus lowland HapMap East Asian (n=60) and European controls (n=90)
LSBL, lnRH, Taj D, WGLRH (Affymetrix 6.0 SNP array)
EGLN1 (EPAS1)
Peng et al 2010
1. Tibetan highlanders from Qinghai Province (n=50) versus lowland HapMap CHB/JPT 2. Tibetan highlanders from Qinghai Province (n=50) versus lowland HapMap CHB/JPT/YRI/CEU 3. Tibetan highlanders from Tibetan Autonomous Region, Qinghai and Yunnan Provinces (n=1334)
1. XP-CLR, FST
(Affymetrix 6.0 SNP array) 2. Full-length sequencing of EPAS1 (haplotype construction, FST) 3. Candidate gene SNP genotyping for allele frequency comparisons
EPAS1 (EGLN1)
Xu et al 2011
Tibetan highlanders from Tibet (Shannan, Rikaza, Linzhi, Lasha and Changdu) (n=46) versus Han Chinese (n=92), YRI (n=60), CEU (n=60) and JPT (n=44) from HapMap
FST, iHs, XP-EHH(Affymetrix 6.0 SNP array), haplotype construction
EPAS1 EGLN1
Wang et al 2011
Tibetan highlanders from near Lhasa (n=30) versus lowland HapMap CHB (n=45), JPT (n=45), CEU (n=59) and YRI (n=60) and East Asian from HGDP
FST, iHs, XP-EHH(Illumina 1M) EPAS1
(EGLN1)
List of Abbreviations CHB: Han Chinese in Beijing, China. JPT: Japanese in Tokyo, Japan. YRI: Yoruba in Ibadan, Nigeria. CEU: Utah residents with northern and western European ancestry. HGDP: Human genome diversity project. XP-EHH: Cross population extended haplotype homozygosity. iHS: Integrated haplotype score. GWADS: Genome wide allelic differentiation scan. PBS: Population branch statistic. LSBL: Locus-specific branch length. lnRH: natural logarithm of ratio of heterozygosities, Taj D: Tajima’s D statistic, WGLRH: Whole genome long-range haplotype. XP-CLR: Cross population composite likelihood ratio test. FST: Fixation index. The basis of these and other techniques are described in recent reviews (15, 80).