heterogeneity in the progeny of single human muscle satellite cells
TRANSCRIPT

&p.1:Abstract We demonstrate that self-renewing myoblastscan be identified in the progeny of single human musclesatellite cells (HMSC) in culture. We show, using cyto-skeletal proteins and cell size as markers, that self-re-newing myoblasts are phenotypically different from oth-er myoblasts, but similar to native HMSC. Native des-min-positive HMSC, cultured as single cells, yielded twomajor populations of myoblasts, α-sarcomeric (α-SR)-actin-positive myoblasts and desmin-positive myoblasts.In appropriate culture conditions, α-SR-actin-positivemyoblasts fused into myotubes, whereas a population ofdesmin-positive non-fusing myoblasts (NFMB) persistedfor weeks among the myotubes. Upon isolation frommyotubes, some of the NFMB resumed proliferation andtheir progeny included fusing and non-fusing myoblasts,with the same cytoskeletal phenotypes as the progeny ofnative HMSC. This self-renewal cycle could be repeated,yielding four cohorts of myoblasts. The yield of self-re-newing cells appeared to decrease with the number ofcycles. These results suggest that stem cells are presentamong NFMB. Moreover, we find that these presumptivestem cells are already segregated during myoblast prolif-eration. They are small, phenotypically similar to nativeHMSC, and do not divide unless they are isolated fromtheir sister progeny and cultured alone. Enriched prepa-rations of cells with stem cell-like properties can be ob-tained from proliferating myoblasts by flow cytometryon the basis of size and nucleocytoplasmic ratio.&bdy:
Introduction
When muscle injury occurs, satellite cells (SC) are stim-ulated and undergo extensive proliferation for a few days
before beginning to fuse, thereby regenerating new mus-cle fibers or adding nuclei to pre-existing fibers [3, 7].During this process, a new population of SC is formedalong with regenerated fibers [35]. This can be repeatedover several cycles of degeneration/regeneration [13, 26,36]. SC activation is also needed for muscle hypertrophy[32].
A series of papers studying myogenesis in chick em-bryo cultures [29, 30] and also in cultures of post-natalchicken SC [31] led to a better understanding of the cel-lular basis of muscle regeneration. The results of thesestudies were explained by the presence of both self-re-newing cells and cells comitted to terminal differentia-tion [31]. Also, this process of self-renewal could be ob-served during successive subcloning of a same popula-tion of embryonic myoblasts [31]. These studies on self-renewal, in which subcloning is performed in differenti-ated cultures, should be distinguished from those demon-strating that proliferating myoblasts can be repeatedlysubcultured [5, 19]. Thus, embryonic myoblasts can beconsidered as cells with stem cell-like properties: theyare able to self-renew and they yield in their progenyboth cells comitted to terminal differentiation and cellssimilar to the mother cells (see Hall and Watt [15] for adefinition of stem cells).
Cells with stem cell-like properties are obviously cru-cial for the regeneration potential of human muscle. Themultiplication potential of these cells is limited and theirdepletion leads to the incapacity of muscle repair ob-served in severe dystrophies [19, 41]. Also, in view ofthe potential use of human muscle satellite cells (HMSC)for transplantation and ex vivo somatic gene therapy, itmay be important to obtain enriched population of thesecells.
The goal of the present study was to examine the pos-sibility of identifying cells with stem-cell-like propertiesand the possibility of getting enriched preparations ofthese cells in cultures derived from purified HMSC. Ourhypothesis was that cells with stem-cell-like propertiesshould be similar to native HMSC, i.e. SC freshly col-lected from human muscle. In previous work, we found
A. Baroffio · M. Hamann · L. Bernheim · C. Bader (✉)Dept. of Physiology, University Medical Center,Division de Recherche Clinique Neuromusculaire,1, rue Michel Servet, CH-1211 Geneva 4, Switzerland
M.L. Bochaton-Piallat · G. GabbianiDepartment of Pathology, University Medical Center,University Hospital, Geneva, Switzerland
Differentiation (1996) 60:47–57 © Springer-Verlag 1996
O R I G I N A L A RT I C L E
&roles:Anne Baroffio · Martine Hamann · Laurent BernheimMarie-Luce Bochaton-Piallat · Giulio GabbianiCharles R. Bader
Identification of self-renewing myoblasts in the progenyof single human muscle satellite cells
&misc:Accepted in revised form: 30 November 1995

that HMSC are characterized by the presence of desmin[1], the absence of α-sarcomeric actin (α-SR actin) [2],and by the absence of both a voltage-gated potassiumcurrent [17, 42] and an ACh-induced current [22].HMSC can be cultured in two different types of media[16, 39], a growth medium (GM) in which the myoblastsderived from these cells can be maintained in a prolifer-ating state, and a differentiation medium (DM) in whichcells are induced to differentiate. In the latter medium,most but not all cells fuse and form myotubes. We ob-served that even after a long period of exposure to a DM,there are cells that remain mononucleated. These cells,contrary to fused cells, were immunocytochemically [2]and electrophysiologically [22, 42] similar to nativeHMSC. Therefore, we examined the properties of thesenon-fusing cells.
We found that at least a fraction of these cells has in-deed stem-cell-like properties. We also demonstrate thatcells with stem-cell-like properties withdraw rapidlyfrom the cell cycle. Their morphological propertiesmakes it possible to obtain enriched preparation of thesecells by flow cytometry sorting of proliferating humanmyoblasts.
Methods
Muscle biopsies. &p.2:Thirty skeletal muscle samples were obtainedfrom young patients (1-19 years old) during corrective orthopedicsurgery. Biopsies were made in accordance with the guidelines ofthe ethical committee of the University Hospital in Geneva. Thesamples were obtained from various muscles (back, abdomen, legand foot).
Preparation of cell suspensions. &p.2:Each biopsy was dissociated ac-cording to a modified procedure of a method first described byBlau and Webster [5] (see Baroffio et al. [1] for further details).SC from the cell suspension could either be directly purified byflow cytometry or be cultured as clones.
Flow cytometry analysis and sorting. &p.2:SC were purified from fresh-ly dissociated muscle cell suspensions by flow cytometry sortingas described previously [1]. Flow cytometry was also used to sepa-rate populations of proliferating myoblasts on the basis of theirsize in order to identify putative stem cells among proliferatingmyoblasts. This was done with a FACStar plus (Becton Dickinson)cell sorter equipped with an argon laser set at a wavelength of 488nm. The sorting was performed with the sorting enhancementmodule and the windows were set for forward light scatter (size)and right-angle light scatter (nucleocytoplasmic ratio). Sortingspeed was maximally 1,500 events/s. Data acquisition and analys-es were performed with the Lysis II Software (Version 1.1 BectonDickinson).
Manual cloning. &p.1:Single cells in suspensions obtained from musclebiopsies were manually collected with a micropipette under a mi-croscope, and cultured individually in culture wells (96-wellplates, Costar) containing GM. This technique ensures that a sin-gle cell is put in culture.
Culture media. &p.2:SC were cultured in two different media: (1) a GMin which cells remain mononucleated and actively proliferate, and(2) a DM, in which SC can fuse to form myotubes (MT). Compo-sition of these media is described by Baroffio et al. [1]. MT some-times present cross striations and often constract spontaneously.Half of the medium was replaced 3 times a week. Each clone was
tested for its myogenicity by transferring confluent cultures to DMand checking formation of MT.
Immunocytochemical procedures. &p.2:Cells were fixed for 5 min inmethanol at −20° C. Double immunofluorescence staining wascarried out using an affinity-purified rabbit polyclonal IgG specificfor desmin, diluted 1/3 [21], and a mouse monoclonal IgM specif-ic for α-SR actin, diluted 1/20, which recognizes both α-skeletaland α-cardiac actin isoforms (anti- αSR-1 [38]). Tetramethylrho-damin isothiocyanate (TRITC)-conjugated goat anti-rabbit IgG(diluted 1/20) and fluorescein isothiocyanate (FITC)-conjugatedgoat anti-mouse IgG (diluted 1/30) (Nordic Immunology Labora-tory) were used as second andibodies. Cells were incubated for 30min. Preparations were observed with a Zeiss Axiophot photomi-croscope equipped with epi-illumination and specific filters forrhodamine and fluorescein (Carl Zeiss). Photographs were takenon Tmax film (400 ASA, Eastman Kodak) using plan apochromatex40/1.0 and x63/1.40 objectives.
Determination of the fusion index. &p.2:Fifteen clones from eight biop-sies were cultivated in DM for different times ranging from 1–10days. Then they were fixed in ethanol for 5 min at −20° C andstained with hematoxylin for 10 min at room temperature. The fu-sion index was established by dividing the number of nuclei with-in MT by the total number of nuclei in 5–10 microscopic fieldsthat were randomly chosen in each culture. One microscopic fieldincluded 50–150 nuclei.
Subcultures of HMSC clones (see scheme a of Fig. 1). &p.2:It is impor-tant to mention, as a preliminary comment, that in these studiescultures were grown for relatively long periods. Our previous stud-ies indicated that SC from children and young adults can be grownfor more than 11 weeks before their proliferation potential pro-gressively diminishes and until divisions stop at approximately 17weeks [42]. At that time cells are senescent and have a characteris-tic morphology (very flat and fibroblast-like) by contrast with thatof proliferating myoblasts (fusiform).
In the subculture experiments, clones of HMSC, previouslycultured for 7 weeks in GM (one clone was previously cultured for9 weeks), were replated in 35 mm culture dishes (Nunc), grown toconfluency (this took less than 1 week), and then transferred to
48
Fig. 1 Schematic summary of the experimental procedure for thesubcultures (a) and subcloning (b). MB0, native, freshly isolatedHMSC; MB, myoblasts proliferating in GM; fusion is induced byexposure to DM; MT, myotubes; NFMB, non-fusing myoblasts;numbers 1–4 refer to successive cohorts. See text for details&/fig.c:

DM and cultured for 7 days. At that time, large MT were presenttogether with non-fusing myoblasts (NFMB). The cultures weretrypsinized (Boehringer Mannheim) and MT were eliminated bydecantation for 30 min. The supernatant, which consisted mainlyof NFMB (first cohort NFMB1), was replated in several culturedishes. Cells in two dishes were fixed as soon as they had spreadout in order to study cytoskeletal characteristics of NFMB1 by im-munocytochemical procedures. Two dishes were allowed to prolif-erate in GM for 7 days and were fixed in order to study cytoskele-tal expression in the new cohort of MB (MB2). Three dishes wereallowed to proliferate to reach confluency and were transferred toDM for 7 days. Two of these latter dishes were fixed for studyingcytoskeletal features of MT2 by immunocytochemistry, whereasthe third dish was again trypsinized in order to separate MT2 andNFMB2 (second cohort). The whole procedure was repeated untilcells stopped proliferating.
Subclones of HMSC clones (see scheme b of Fig. 1). &p.1:NFMB1 wereisolated by the procedure described above. NFMB1 were manuallycloned in individual wells (96-well plates, Costar). Clones of thesecond cohort (MB2 subclones) were grown to confluency, andsome of them were trypsinized and separated in two wells. In or-der to assess the myogenicity of the subclone, cells in one of thewells were transferred to DM when the cultures became confluent.After a few days, the content of the second well was trypsinizedand replated in four 35-mm culture dishes (Nunc) to study cyto-skeletal characteristics by immunocytochemistry; two dishes weregrown in GM and fixed at low confluency, whereas the other twowere grown to confluency, further cultured in DM for 7 days, andfixed.
Autoradiography. &p.1:In order to estimate the proportion of dividingcells, cultures were exposed to 1µCi/ml [3H]-thymidine (specificactivity 25 Ci/mmol, Amersham) dissolved in the culture mediumfor the times indicated in the relevant section. After flow cytome-try sorting, small and large cells were immediately cytocentri-fuged at 125g for 5 min (Shandon). Cells were then fixed 5 minwith methanol at −20° C, rinsed with distilled water, and coatedwith NTB3 autoradiographic emulsion (Kodak). After 14 days ofexposure, plates were developed at 20° C with Kodak D19 for 5min, rinsed in water, and fixed. The percentage of dividing cellswas established by counting a minimum of 100 cells.
Statistics. &p.2:Results are expressed as means±SEM.
Results
Our previous work on NFMB indicated that these cellsresemble HMSC both immunocytochemically and elec-trophysiologically [2, 22, 42]. In what follows, we shallexamine other properties of these NFMB, in particulartheir self-renewal properties. However, to ensure thatNFMB are in a steady state of failure to fuse despite thepresence of the fusion-promoting DM, our first goal willbe to assess that the fusion process is indeed completed.
The fusion index was evaluated in a total of 44 clonalcultures (15 different clones from eight biopsies) thathad been fixed after 1–10 days in DM. The average fu-sion index increased from 0 to about 70% in 5 days andthen remained stable (Fig. 2a). Some clones had fasterrates of fusion than others. The maximal fusion indexwas reached by individual clones between day 3 and 7but it always reached a value comprised between 60%and 80%. Thus, NFMB account for approximately 30%of the whole progeny of each SC (Fig. 2b). NFMB re-mained mononucleated and did not proliferate as long as
they stayed in DM (up to 3 months at least). These near-ly confluent cultures remained remarkably stable for upto 3 months, and we never observed the mitosis that areso frequently seen in GM.
NFMB have self-renewal properties
The next step was to demonstrate the self-renewal abilityof NFMB. Experiments were done either with mass cul-tures of NFMB (“subcultures”; see Fig. 1a) or by puttinga single NFMB in a dish (“subclones”; see Fig. 1b). Thefusion index and the cytoskeletal characteristics of theprogenies obtained in the subcultures or subclones wereanalyzed and compared from one cohort to the next.
Studies on subcultures of NFMB
In 7 of the 8 clones of HMSC studied, pooled NFMB1(Fig. 1a) were able to resume proliferation and gave riseto a second cohort of NFMB (NFMB2). This cycle couldbe repeated up to four successive cohorts (see Fig. 3f, in-set). Thus, experiments on subcultures indicate that atleast a fraction of the NFMB reproduce the behavior typ-ical of native HMSC, when these are first put in culture.
Studies on subclones and estimation of the proportionof cells with stem cell-like properties
In order to get a more precise idea of the number of cellswith stem cell-like properties in the NFMB population,
49
Fig. 2 aEvolution of the fusion index with time in DM. A total of44 clonal cultures (15 different clones from eight biopsies) weretransferred to DM on day 0 and were fixed after different timesranging from 1–10 days. Fusion index was estimated as describedunder experimental procedures. Results are expressed asmeans±SEM. Numbers in parenthesesrepresent the number ofcultures. b Phase contrast micrograph of a clonal culture fixed andstained with hematoxylin after 7 days in DM. Star, MT; arrows,NFMB. ×500&/fig.c:

we conducted experiments in which the progeny of a sin-gle NFMB was examined. NFMB1 were obtained fromfive different clones that had reached a steady-state ofthe fusion process. Depending upon their rates of fusion,the cells of the various clones tested were cultured for3–9 days in DM. The final average fusion index was76±2% (Table 1). A total of 280 NFMB were manuallycloned and cultured as single cells in individual wells(Fig. 1b). Of these 280 NFMB1, 21 qualified as trulyself-renewing cells, as they resumed proliferation andgave rise to an actively proliferating progeny (i.e., under-
going more than ten divisions) that was able to fusewhen transferred to DM. In the five individual subclon-ing experiments, the percentage of self-renewing cellsamong NFMB1 varied between 3% and 20%, with an av-erage of 9% ±3.
Taking into account the fraction of NFMB1 in eachexperiment (100% minus the fusion index before sub-cloning; see Table 1), we estimated the proportion of pu-tative stem cells within the progeny of a single HMSCunder our experimental conditions. For example, forclone 1, the fraction of putative stem cells would be
50
Fig. 3a–f Quantitative andqualitative evaluation of sub-clones and subcultures in theproliferation state. Results areexpressed as means±SEM.Numbers in parenthesesare thenumber of clones studied. a, bDouble immunofluorescencestainings of a proliferating sub-clone (MB2) for α-SR actin (a)and desmin (b), revealing α-SR-positive, desmin-positive(arrowhead) and α-SR-nega-tive, desmin-positive (arrow)cells. ×650. c, d Double immu-nofluorescence stainings of thefourth cohort of MB in subcul-tures (MB4) for α-SR actin (c)and desmin (d). Note that thethree cells were doubly labeled×650. e Relative percentages ofα-SR-positive, desmin-positive(white) and of α-SR-negative,desmin-positive (gray) cellsamong MB at each cohort. Thetotal percentage of αSR+ andαSR– cells is less than 100%(see reference 2) f Relation be-tween the proportion of α-SR-positive cells in the first cohortof MB and the number of co-horts that a given clone couldyield. Histogram of the numberof clones that yielded one tofour cohorts (inset) &/fig.c:
Table 1 Proportion of self-renewing cells among NFMB&/tbl.c:&tbl.b:
Clone no. Days in Fusion index Number Number of Fraction of Fraction ofDM before of cells myogenic self-renewing cells self-renewing cells
subcloning subcloned subclones in NFMB in the whole(%) (%) progeny
(%)
1 9 78 40 8 20.0 4.42 4 80 40 1 3.0 0.63 3 70 40 2 5.0 1.54 7 80 40 5 12.5 2.55 7 71 120 5 4.2 1.2
76±2a 280b 21b 9±3a 2±1a
a Means±SEM; b Total number&/tbl.b:

20%×(100−78%)=4.4%. Similar calculations in the otherclones lead to a mean estimate of the fraction of stemcells in the overall progeny of a native HMSC of 2±1%(range 0.6–4.4%) after a total of 8 weeks in proliferationmedium. This fraction may vary for different times ofproliferation, but this was not examined here.
Cytoskeletal expression and fusion index in each cohort
We analyzed the expression of desmin and α-SR actin insuccessive cohorts of myoblasts (MB), MT and NFMB.The fusion index was also determined in successive co-horts of MT.
We described recently the various cell populationpresent in a clone of proliferating MB [2], the predomi-nant ones being: desmin-positive/ α-SR actin-positivecells (we shall call these cells predifferentiated MB) anddesmin-positive/α-SR actin-negative cells (undifferenti-ated MB). It is important to note that all of the MB phe-notypes we found in SC or in their MB progeny grown invitro were observed in vivo (desmin [46]; α-cardiac andα-skeletal actin, see for example [14, 33]; α-smooth ac-tin [6, 34]).
In each cohort, predifferentiated MB and undifferenti-ated MB were present (Fig. 3a–d), but their relative pro-portions varied with the cohorts (Fig. 3e). The proportionof undifferentiated MB was 65% in the first cohort andthen decreased to 10% in the fourth cohort. Concomi-tantly, the proportion of predifferentiated MB that was29% in the first cohort gradually increased to 80% in thefourth cohort. These observations combined with the factthat the potential of self-renewal decreased at each co-hort (see Fig. 3, inset) suggested that the potential ofself-renewal of a given population of MB may be in-versely correlated with the level of predifferentiation tak-ing place in that population.
To test this hypothesis we investigated whether thenumber of cohorts that a given clone derived from a sin-gle HMSC would yield was correlated with the propor-tion of predifferentiated MB present in the MB1 of thatclone (Fig. 3). We observed that for the clone thatreached the fourth cohort, only 5% of the MB1 cellswere positive for α-SR actin. In contrast, the clonewhich failed to give a second cohort of MB possessed50% of predifferentiated MB at the stage MB1. The sixclones that reached the second or third cohort had an in-termediate proportion of 20% of predifferentiated MB intheir MB1 population. Thus, it appears that there is aninverse relationship (r=−0.92) between the proportion ofαSR actin-positive predifferentiated MB and the self-re-newal potential of a given clone.
In the presence of DM (Fig. 4), MT were formed ineach cohort and displayed the expected desmin-positive,α-SR actin-positive phenotype (Fig. 4a–d). However, thefusion index tended to decrease with the cohorts (Fig.4e). Like proliferating MB, NFMB were made up of twomajor populations: desmin-positive/ α-SR actin-negativeand desmin-positive/ α-SR actin-positive cells (Fig. 4f).
In the first cohort, the undifferentiated phenotype wasclearly predominant, as already mentioned, but it de-creased with cohorts. Thus, NFMB reproduce in theirprogeny the cytoskeletal patterns that are observed in theprogeny of parental native HMSC directly isolated fromthe muscle. However, for each clone analyzed, the levelof predifferentiation increased over cohorts, and this wascorrelated with a marked decrease of the self-renewalpotential and a smaller reduction of the fusion potential.
Stem cells among proliferating MB
We wondered whether the self-renewing cells that wefind among NFMB were segregated during exposure toDM at the time MT form, or whether the segregation pro-cess took place already while MB proliferate in GM. Wehave already mentioned that proliferating MB containpopulations of predifferentiated and undifferentiated cellswith respect to actin expression [2]. These two subpopu-lations appeared very early at the onset of the prolifera-tion of a HMSC. Similarly, our electrophysiological ex-amination of the response to applied ACh indicated thattwo types of responses were observed when ACh was ap-plied on proliferating MB [22]. This suggests the pres-ence of two populations of MB with respect to ACh sen-sitivity. In addition, as we observed that ACh sensitivityincreased with differentiation [22], it suggested that ACh-sensitive cells belong to the predifferentiated MB.
Our strategy for attempting to identify putative stemcells in the whole population of proliferating MB wasthe following. Native HMSC, which are in the G0 phaseof the cell cycle, are known to be small cells [1, 17].Moreover, physical features such as different size andlight scatter properties distinguish hematopoietic or kera-tinocytes stem cells from differentiated cells [8, 9, 12,28]. Finally, a characteristic of stem cells, as seen inmany different types of renewing cell populations, is thatunder steady-state conditions, they are non-dividing [25].Thus, we thought that the putative stem cells in our cul-tures might be smaller than most proliferating MB. Inaddition, they might be distinguished from the fractionof small proliferating MB (G1 phase), by a long-term ex-posure to 3H-thymidine (stem cells should not be la-beled).
Therefore, we collected cells proliferating in GM(MB1, Fig. 1) and analyzed them on the basis of theirsize and nucleocytoplasmic ratio with a cell sorter. Eightclonal cultures were examined after 8–9 weeks of prolif-eration. The analysis clearly revealed the presence of twosubpopulations of cells with distinct sizes (forward scat-ter) and nucleocytoplasmic ratio (log right angle scatter)(Fig. 5a). The diameters of these populations were10±0.4µm (n=53) and 20±0.3µm (n=75), respectively.The proportion of the small cell subpopulation was34±8% at the particular time in culture studied here (8–9weeks of proliferation). We did not examine the percent-age of the small cell population at different time of pro-liferation in culture.
51
In order to determine whether the subpopulation ofsmall and large cells were different, we compared (1)their proliferation abilities, (2) their level of cytoskeletaldifferentiation, and (3) their self-renewal capacity. Theproliferation state was investigated by incubating prolif-erating MB for 2 days with a S-phase-specific agent([3H]-thymidine) before sorting them according to theirsize and nucleocytoplasmic ratio (average cell cycle du-ration of 30 h [42]). Subsequently the presence of [3H]-thymidine was examined in both populations by autora-diography (Fig. 5b). The small cells did not incorporate[3H]-thymidine, whereas 79±1% of the large cells werelabeled (three clones tested). Longer exposures only
52
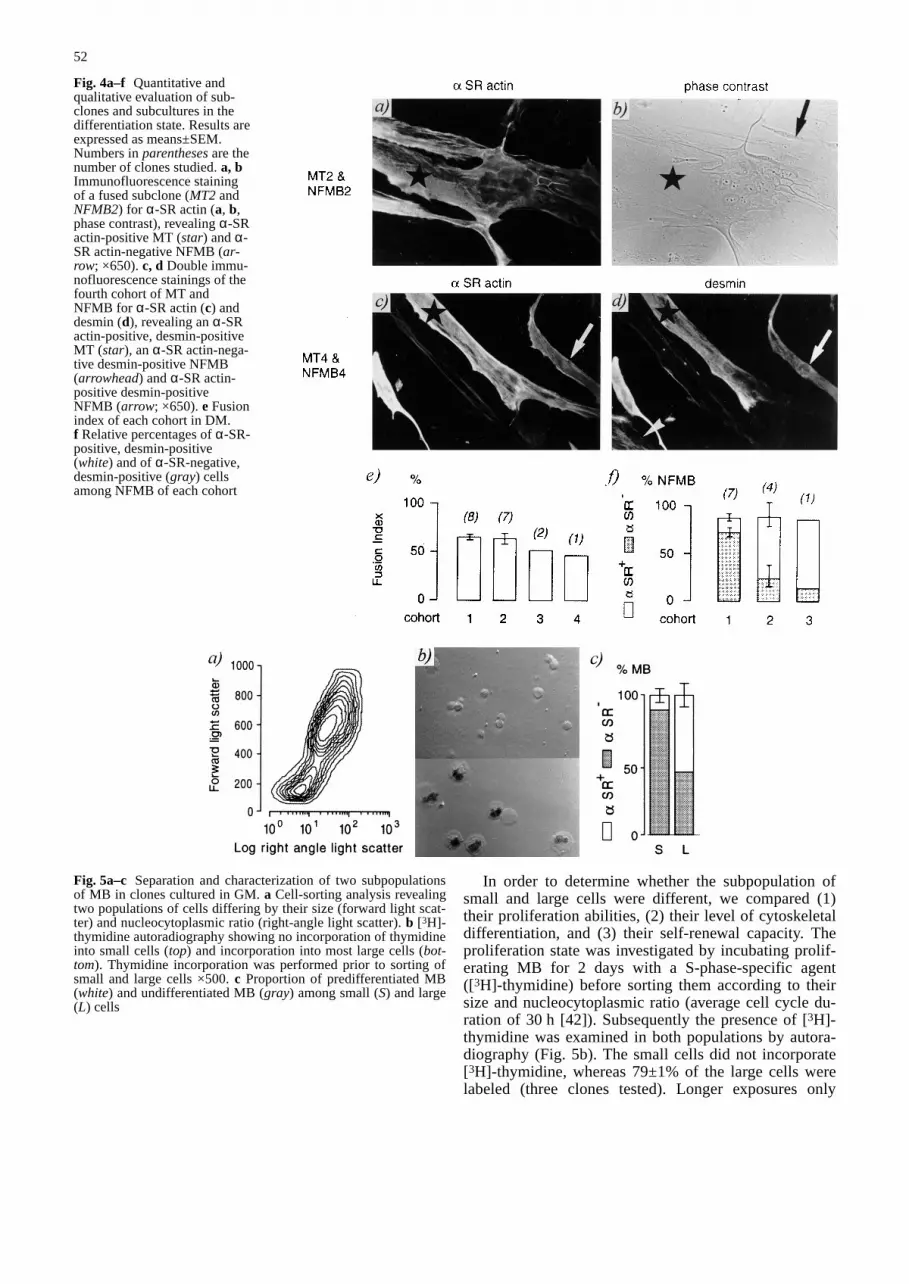
Fig. 4a–f Quantitative andqualitative evaluation of sub-clones and subcultures in thedifferentiation state. Results areexpressed as means±SEM.Numbers in parenthesesare thenumber of clones studied. a, bImmunofluorescence stainingof a fused subclone (MT2 andNFMB2) for α-SR actin (a, b,phase contrast), revealing α-SRactin-positive MT (star) and α-SR actin-negative NFMB (ar-row; ×650). c, d Double immu-nofluorescence stainings of thefourth cohort of MT andNFMB for α-SR actin (c) anddesmin (d), revealing an α-SRactin-positive, desmin-positiveMT (star), an α-SR actin-nega-tive desmin-positive NFMB(arrowhead) and α-SR actin-positive desmin-positiveNFMB (arrow; ×650). e Fusionindex of each cohort in DM.f Relative percentages of α-SR-positive, desmin-positive(white) and of α-SR-negative,desmin-positive (gray) cellsamong NFMB of each cohort&/fig.c:
Fig. 5a–c Separation and characterization of two subpopulationsof MB in clones cultured in GM. a Cell-sorting analysis revealingtwo populations of cells differing by their size (forward light scat-ter) and nucleocytoplasmic ratio (right-angle light scatter). b [3H]-thymidine autoradiography showing no incorporation of thymidineinto small cells (top) and incorporation into most large cells (bot-tom). Thymidine incorporation was performed prior to sorting ofsmall and large cells ×500. c Proportion of predifferentiated MB(white) and undifferentiated MB (gray) among small (S) and large(L) cells

marginally increased the fraction of cells labeled with[3H]-thymidine; after 7 days of exposure, 2% of thesmall cells incorporated [3H]-thymidine, whereas 77% ofthe large cells were labeled (one clone tested). This dem-onstrates that cycling cells belonged to the population oflarge cells, whereas small cells were either quiescent orcycling very slowly.
Our hypothesis was that small cells would have thecytochemical characteristics of native HMSC and thatlarge cells would include the predifferentiated MB. Wefound (Fig. 5c) that about 90% of the smaller cells wereundifferentiated MB, whereas about 55% of the largercells were predifferentiated MB (three clones, seven de-terminations).
By definition, cells with stem cell-like propertiesshould have the capacity of self-renewal [15]. This abili-ty can be best evaluated by clonal analysis, which allowsthe properties of individual small and large cells to bestudied. We studied the survival and the self-renewal po-tential of both populations by subcloning 150 small cellsand 140 large cells and plating them as single cells in in-dividual wells (n=4 clones; Table 2). The survival (de-fined as a subclone comprising at least one cell) was thesame (62%) for both types of cells. Among the survivingcells, about 70% of the small cells reentered the cell cy-cle (subclone constituted of at least two cells). In con-trast, more than 90% of the large cells remained as singlecells in the wells; only 9% of them reentered the cell cy-cle, but they did not divide more than seven times anddid not give rise to MT. It is important to emphasize thatthese large cells fail to proliferate and fuse only whenthey are cultured alone. Mass cultures of sorted largeMB rapidly fuse when put in culture in DM.
In contrast, 12% of the surviving small cells (7% ofsubcloned cells) were able to proliferate actively (i.e.,more than ten divisions) and gave rise to large myogenic
clones. Therefore, true self-renewing cells were exclu-sively found among small cells. Thus, as a HMSC startsproliferating in GM, a fraction of its clonal progenykeeps the phenotype of the native cell and appears towithdraw from the cell cycle. When isolated from theirsister progeny, some of these cells resume proliferationand are able to give rise to large myogenic subclones. Weconsider that these cells show a capacity for self-renewaland become equivalent to native muscle SC. On the otherhand, another part of the clonal progeny acquires a moredifferentiated phenotype, proliferates in the presence ofthe sister progeny, and never gives rise to a myogenicsubclone when cultured as a single cell. However, thispredifferentiated progeny is able to fuse rapidly even af-ter cell sorting, provided it is cultured at an adequate celldensity.
Taking into account the fraction of small cells in pro-liferating MB (34%), the average percentage of stemcells is estimated to be 2% in a clonal culture of myo-blasts (i.e., 7% stem cells in the population of small cells×34% small cells in the overall clonal population) after8–9 weeks of proliferation in culture. As we mentionedalready in the experiments with subclones, this fractioncould be different for other durations of the proliferationperiod.
Discussion
On the basis of (1) an immunocytochemical and electro-physiological characterization of HMSC and of NFMBand of (2) the ability to study either proliferating MB ordifferentiated cultures, we could confirm, in human cells,previous observations on the presence of cells with stemcell-like properties in embryonic and postnatal myogeniccells. Quinn et al. [30, 31] gave evidence of the existence
53
Table 2 Proportion of self-re-newing cells among the prolif-erating myoblasts (MB1)&/tbl.c:&tbl.b:
Clone no. Cell size Number Number Number of Number ofof cells of proliferating myogenicsubcloneda surviving cells subclones
cells
1 Small 30 24 10 2Large 30 22 6 0
2 Small 60 36 33 4Large 50 32 1 0
3 Small 30 10 5 2Large 30 15 1 0
4 Small 30 22 20 2Large 30 18 0 0
Total Small 150 92 68 10Large 140 87 8 0
Mean Small 62±10 43±11 7±0(in % of Large 62±5 6±5 0subclonedcells)
Mean Small 69±13 12±3(in % of Large 9±6 0survivingcells)
a Surviving cell=subclone ≥1cell; proliferating cell=sub-clone ≥2 cells; self-renewingcell=large myogenic subclone&/tbl.b:

of stem cells among chicken embryonic MB in culture.This observation has been extended to chicken SC [31,43]. Recently, experiments performed in vivo [45]showed that implanted MB were giving rise to both afusing progeny and to a new pool of precursor cells.
In addition, the present study demonstrates that atleast part of the SC that are born in vitro are already seg-regated during the proliferation phase of MB and thatthey withdraw from the cell cycle. Another interestingobservation is that cells with stem cell-like properties be-long to a cell population characterized by its small size,allowing for enriched preparations of these cells. Otherfindings, which may also be of therapeutical relevance,are an estimate of the stem cell contents in the progenyof HMSC and the ability to predict this stem cell contenton the basis of the immunocytochemical characteristicsof proliferating MB.
HMSC are a kind of stem cells
In the present study we propose that the progeny of eachsingle HMSC includes both predifferentiated cells (giv-ing rise to MT in DM) and cells that maintain the samephenotype as the mother cell (small undifferentiated MBin GM, and NFMB in DM). The undifferentiated MB ap-pear to “protect” their mitotic potential, as they stop pro-liferating relatively rapidly. They have a self-renewalability, as they reproduce the same cycle as the mothercell, yielding again both types of progenies under appro-priate culture conditions. Thus, we believe that stemcells are found among small undifferentiated MB. It isworth noting that the NFMB do not respond to ACh ap-plication [22], which is another indication that SC bornin vitro are identical to native SC.
Can we exclude that predifferentiated cells behave asstem cells when cultured as single cells? Direct proof isdifficult to obtain, as labeling a cell to detect the pres-ence of α-SR-actin requires fixing it. However, severalfacts argue against this possibility. First, we would ex-pect stem cell in culture to have the same phenotype asnative SC, and SC are α-SR actin-negative. Thus, to sat-isfy our criterion, predifferentiated cells would have tode-differentiate. This behavior is usually not seen in nor-mal, untransformed cells. Second, in our experimentswith proliferating MB, candidate stem cells clearly be-long to a population of small cells, of which 90% are α-SR actin-negative. Subcloned large cells of the growingMB population never resulted in MT, and 50% of theselarge cells are predifferentiated cells. Third, if α-SR ac-tin-positive cells were the stem cells born in culture, itwould be difficult to explain why the stem cell potentialruns down, as the fraction of α-SR actin-positive cellsincreases during successive subcloning experiments (seeFigs. 3e, f and 4f). Therefore, on the basis of these argu-ments we propose that it is very likely that only the smallα-SR actin-negative/desmin-positive cell behave as stemcells.
Nature and fate of the large MB in clonal cultures
It is interesting to mention that most of the larger MBwere only able to proliferate in GM as long as they werecultured with the entire progeny of a HMSC. As soon asthese MB were segregated from the whole populationand cultured alone, they either stopped dividing or prolif-erated only a few times. This observation suggests thatinteractions may exist between the pre- and un-differen-tiated human myoblast populations. In particular, thepopulation of larger predifferentiated MB might be pro-liferating in response to soluble factors produced by theirsister cells [10] or as a consequence of direct cell-cell in-teractions. Under appropriate conditions these cells willdifferentiate into muscle fibers. These conditions are ob-viously not present in our clonal cultures, in particularbecause of the small number of cells present. Indeed,mass cultures of sorted large MB differentiated andfused rapidly.
What are the larger MB in the proliferating MB popu-lation that were α-SR actin-negative? Why did thesecells not behave as stem cells? We propose that theselarge α-SR actin-negative cells may be already partiallypredifferentiated, as a population of desmin-positive/ α-SR actin-negative MB start expressing α-SR actin andfuse immediately when put in DM [2]. This suggests thatthe whole population of larger MB is destined to fuse.
Percentage of small cells that behave as stem cells
As already mentioned, cells with stem cell properties inthe proliferating MB preparation were present exclusive-ly in the population of small cells. Of 100 small cells,however, only 7 qualified as stem cells, having an exten-sive growth potential and yielding MT. What can be saidabout the 93% of small cells that were cloned and thatdid not form large colonies? In our analysis, we distin-guished four categories: (1) stem cells (7%) (2) cells thatdid not survive because of the sorting and culture condi-tions (38%), (3) cells that survived but did not proliferate(19%; predifferentiated and/or senescent cells), and (4)cells with only a small progeny on which fusion couldnot be assessed (36%, abortive or predifferentiated cells).What follows is an explanation on these categories.
The fate of the cells that did not survive the cloningprocedure (38%) could not be assessed. It must be re-called that all cells were submitted to resuspension fol-lowing proliferation, to flow cytometry cell sorting, andto culture in isolation. It is not surprising that all cellswould not have survived the procedure. The remaining55% of small cells (Table 2, 62% surviving cells minus7% myogenic subclones) that did not behave as stemcells can be classified as follows. Nineteen percent sur-vived as single cells (Table 2, 62% surviving cells minus43% proliferating cells), but did not reenter the cell cy-cle. Half of these (i.e., 10%) could be predifferentiatedcells, corresponding to the 10% of α-SR-positive cells inthe small cell population (see Fig. 5c) that cannot resume
54

proliferation as single cells; the remaining 9% could re-present senescent cells that have definitively withdrawnfrom the cell cycle. Senescent cells are often described tobe larger than dividing cells [11]. In our cultures, howev-er, senescent cells may be smaller, as flow cytometryanalysis of old clonal cultures that had stopped dividingafter 50–60 divisions [42] indicated that these cultureswere made up of about 80% of small cells (data notshown). The remaining 36% of small cells (Table 2, 43%proliferating minus 7% myogenic subclones), reenteredthe cell cycle, but had a very small progeny. These couldbe “abortive” stem cells that were not killed but affectedby the entire procedure and did not provide their progenywith enough resources to proliferate in our culture condi-tions. Alternatively, they could represent cells that are al-ready at an early stage of predifferentiation but that donot yet express α-SR actin.
Based on our results with both NFMB and proliferat-ing MB, we find that after 8–9 weeks of proliferation ap-proximately 2% of the progeny of a single HMSC hasthe capacity of self-renewal. This may be a lower esti-mate, as there are reasons to believe that our culture con-ditions are not optimal for the survival of self-renewingcells (see preceding paragraph). On the other hand, it isalso possible, even if very unlikely, that our culture con-ditions promote the development of stem cells. In theirstudy, Yao and Kurachi [45] estimated that 0.1–0.5% ofthe implanted MB eventually remained as SC. Theseproportions are in the range reported for stem cells, e.g.,between 0.4% in the hematopoietic system and 10% inthe epidermis [15]. In any case, it must be clear that thepercentage of stem cells given in the present study is val-id for a proliferation time of 8–9 weeks. Note that thisfraction may vary for different times of proliferation, asstem cells could be progressively diluted out, dependingon the rate of production of the other cells in the proge-ny, for example. Other durations of the proliferation peri-od were not examined.
Future stem cells are segregatedbefore the fusion process
Our observations suggest that future stem cells and pre-differentiated MB may be predetermined early during theproliferation phase of MB. The α-SR actin-positive pre-differentiated MB are already present after 4 days of pro-liferation of the native HMSC, but may even appearsooner, which we did not investigate [2]. This predeter-mination could be the consequence of asymmetrical divi-sions, which is a characteristic of stem cells [15, 23]. Inthe rat it was noticed that following each SC division,only 50% of the progeny was able to fuse with the mus-cle [27]. Also, it appears that during myogenesis in iso-lated rat fibers, only half SC descendants express a dif-ferentiation marker, developmental myosin, suggestingthat only one daughter cell enters the phase of terminaldifferentiation [44]. This asymmetry could either be in-trinsically determined, or be induced and reinforced by
stochastic events occuring within the cell or by cell sig-nalling between sister cell [20]. For example, membranereceptors, such as the acetylcholine receptor [22] or ionicchannels [42], could appear stochastically on some cellsand then be reinforced by positive feedback mechanisms[20], triggered by the functioning of such receptors orionic channels, thus determining the fate of the cell. Withregard to ACh receptors, the release of an ACh-like com-pound by myogenic cells [18] may induce such a feed-back mechanism. In any case, it is likely that as forchicken embryonic muscle cells [31] both symmetricaland asymmetrical mitosis eventually takes place to ac-count for the entire progeny of human mucle SC, whichis made up of a small number of stem cells and a largenumber of predifferentiated cells.
Regarding cellular interaction, smaller MB either didnot divide at all or cycled very slowly, when they weremixed with the entire progeny of a HMSC. As soon asthese smaller MB were isolated, about 7% of themresulted in large colonies which were able to differenti-ate, to fuse and form MT. The smaller cells, contrary tothe larger predifferentiated MB (see above), may be un-der the negative control of the predifferentiated progeny[24, 25]. This quiescent state could then be extendedthroughout differentiation, by contact with the plasma-lemma of newly formed MB [4], thus explaining thepresence of the NFMB, which we believe are the in vitroequivalent of the new SCs.
Predictive value of the immunocytochemicalcharacteristics of MB cultures
Our results suggest that the self-renewal potential ofHMSC in vitro can be predicted from the level of predif-ferentiation. It is known that the regeneration potential ofmucle is not infinite and that it decreases with age [5, 19,37]. In vitro, the proliferation potential of HMSC is esti-mated to be 80 divisions in the fetus. This potential di-minishes to 50 divisions in the child and decreases fur-ther to 20 divisions in the 50-year-old person [41, 42].This suggests that muscle stem cells, like other stemcells, are not immortal [40], but that their self-renewalproperties are adequate to face the requirement of a nor-mal life. The regeneration potential, however, may be-come limited after repeated regeneration cycles [36]. Inparticular, the continuous regeneration process imposedby pathological states, e.g., severe myopathies, rapidlyleads to the exhaustion of SC [41].
A decrease in self-renewal potential with age and withcontinued proliferation of HMSC is apparent in our cul-tures. We noticed a tendency for the fusion index to de-crease over cohorts. This may reflect an aging process,as we observed that senescent HMSC, that have sponta-neously stopped dividing in GM after approximately 50divisions, are unable to fuse when transferred into DM,even though they were plated at confluency (unpublishedobservation). Also, we observed that the fraction of un-differentiated MB found among proliferating MB and
55

NFMB decreased over cohorts. This observation sug-gests that the pool of stem cells is progressively depletedand probably explains why the yield of the subculturingdeclined in our experiments from one cohort to the next.In this respect, it is important to mention that the cloneswith the smallest proportion of predifferentiated MBwere able to produce more cohorts of subcultures. In-deed, we found an inverse correlation between the self-renewal potential and the level of predifferentiation with-in a given clone.
Regarding the use of HMSC for transplantation andex vivo somatic gene therapy, our results raise severalquestions that will require additional investigation. First,it will be necessary to examine whether proliferating pre-differentiated MB constitute the optimal source of cellsfor obtaining the best yield of MB fusion with the pa-tient’s muscle fibers. Second, it will be worth examiningwhether the small undifferentiated cell population can bespecifically utilized as a source of muscle regeneratingcells, and whether their regenerating potential can be in-creased. The abilility of preparing both pure HMSC di-rectly from muscle biopsies [1] and a cell population en-riched in small undifferentiated MB (this study) may beuseful tools for these studies.
&p.2:Acknowledgements We are very grateful to Drs. R. Bischoff, andJ. Fisher-Lougheed for helpful comments on the manuscript. Wealso thank J.-C. Rumbeli and F. Strub for the photographic work,G. van Kaenel for graphical representation, M. Berti and A. Mar-guerat for helpful technical assistance on cell cultures and autora-diography, Dr. A. Kaelin for providing us with the muscle biop-sies, and D. Wohlwend for flow cytometry analysis and sorting.This work was supported by the Swiss National Science Founda-tion no. 31-18692-90 and by the Fondation Suisse de Recherchesur les Maladies Musculaires.
References
1. Baroffio A, Aubry JP, Kaelin A, Krause RM, Hamann M, Ba-der CR (1993) Purification of human muscle satellite cells byflow cytometry. Muscle Nerve 16:498–505
2. Baroffio A, Bochaton-Piallat ML, Gabbiani G, Bader CR(1995) Heterogeneity in the progeny of single human satellitecells. Differentiation 59:259–268
3. Bischoff R (1986) Proliferation of human satellite cells on in-tact myofibers in culture. Dev Biol 115:129–149
4. Bischoff R (1990) Interaction between satellite cells and skele-tal muscle fibers. Development 109:943–952
5. Blau HM, Webster C (1981) Isolation and characterization ofhuman mucle cells. Proc Natl Acad Sci USA 78:5623–5627
6. Bochaton-Piallat ML, Ropraz P, Gabbiani G, Santeusanio G,Palmieri G, Schiaroli S, Spagnoli LG (1992) Actin isoformand intermediate filament protein expression in human devel-oping skeletal muscle. Basic Appl Myol 2:83–87
7. Campion DR (1984) The muscle satellite cell: a review. IntRev Cytol 87:225–251
8. De Witte T, Plas A, Koekman E, Blankenborg G, Salden M,Wessels J, Haanen C (1984) Separation of human bone mar-row by counterflow centrifugation monitored by DNA-flowcy-tometry. Br J Haematol 58:249–258
9. Fleming WH, Alpern EJ, Uchida N, Ikuta K, Spangrude GJ,Weissman IL (1993) Functional heterogeneity is associatedwith the cell cycle status of murine hematopoietic stem cells. JCell Biol 122:897–902
10. Florini JR, Ewton DZ, Magri KA (1991) Hormones, growthfactors, and myogenic differentiation. Annu Rev Physiol53:201–216
11. Goldstein S (1990) Replicative senescence: the human fibro-blast comes of age. Science 249:1129–1133
12. Gordon MY (1993) Human haemopoietic stem cell assays.Blood Rev 7:190–197
13. Gulati AK (1986) Pattern of skeletal muscle regeneration afterreautotransplantation of regenerated muscle. J Embryol ExpMorphol 92:1–10
14. Gunning P, Hardeman E, Wade R, Ponte P, Bains W, BlauHM, Kedes L (1987) Differential patterns of transcript accu-mulation during human myogenesis. Mol Cell Biol 7:4100–4114
15. Hall PA, Watt FM (1989) Stem cells: the generation and main-tenance of cellular diversity. Development 106:619–633
16. Ham RG, StClair JA, Blau HM, Webster C (1989) Serum-freemedia for growth and differentiation of human muscle satellitecells. In: Kedes LE, Stockdale FE (eds) Cellular and molecularbiology of muscle development. Liss, New York, pp 905–914
17. Hamann M, Widmer H, Baroffio A, Aubry JP, Krause RM,Kaelin A, Bader CR (1994) Sodium and potassium currents infreshly isolated and in proliferating human muscle satellitecells. J Physiol 475:305–317
18. Hamann M, Chamoin MC, Portalier P, Bernheim L, BaroffioA, Widmer H, Bader CR, Ternaux JP (1995) Synthesis and re-lease of an acetylcholine-like compound by human myoblastsand myotubes. J Physiol (Lond) 489.3:791–803
19. Hauschka SD, Linkhart TA, Clegg C, Merill G (1979) Clonalstudies of human and mouse muscle. In: Mauro A (ed) Muscleregeneration. Raven Press, New York, pp 311–322
20. Horvitz HR, Herskowitz I (1992) Mechanisms of asymmetriccell division: two Bs or not two Bs, that is the question. Cell68:237–255
21. Kocher O, Skalli O, Bloom WS, Gabbiani G (1984) Cytoskel-eton of rat aortic smooth muscle cells. Normal conditions andexperimental intimal thickening. Lab Invest 50:645–652
22. Krause RM, Hamann M, Bader CR, Liu JH, Baroffio A, Bern-heim L (1995) Activation of nicotinic acetylcholine receptorsincreases the rate of fusion of cultured human myoblasts. JPhysiol (Lond) 489.3:779–790
23. Mayani H, Dragowska W, Lansdorp PM (1993) Lineage com-mitment in human hemopoiesis involves asymmetric cell divi-sion of multipotent progenitors and does not appear to be in-fluenced by cytokines. J Cell Physiol 157:579–586
24. Mezzogiorno A, Coletta M, Zani BM, Cossu G, Molinaro M(1993) Paracrine stimulation of senescent satellite cell prolif-eration by factors released by muscle or myotubes from youngmice. Mech Ageing Dev 70:35–44
25. Moore MAS (1979) Stem cell concepts. In: Mauro A (ed)Muscle regeneration. Raven Press, New York, pp 1–7
26. Morlet K, Grounds MD, McgGeachie JK (1989) Muscle pre-cursor replication after repeated regeneration of skeletal mus-cle in mice. Anat Embryol 180:471–478
27. Moss FP, Leblond CP (1971) Satellite cells as the source ofnuclei in muscles of growing rats. Anat Rec 170:421–435
28. Pavlovitch JH, Rizk Rabin M, Gervaise M, Metezeau P, Grun-wald D (1989) Cell subpopulations within proliferative anddifferentiating compartments of epidermis. Am J Physiol256:977–986
29. Quinn LS, Nameroff M, Holtzer H (1984) Age-dependentchanges in myogenic precursor cell compartment sizes. Evi-dence for the existence of a stem cell. Exp Cell Res 154:65–82
30. Quinn LS, Holtzer H, Nameroff M (1985) Generation of chickskeletal muscle cells in groups of 16 from stem cells. Nature313:692–694
31. Quinn LS, Norwood TH, Nameroff M (1988) Myogenic stemcell commitment probability remains constant as a function oforganismal and mitotic age. J Cell Physiol 134:324–336
32. Rosenblatt JD, Yong D, Parry DJ (1994) Satellite cell activityis required for hypertrophy of overloaded adult rat muscle.Muscle Nerve 17:608–613
56

33. Sassoon DA, Garner I, Buckinham M (1988) Transcripts of al-pha-cardiac and alpha-skeletal actins are early markers formyogenesis in the mouse embryo. Development 104:155–164
34. Sawtell NM, Lessard JL (1989) Cellular distribution of smoothmuscle actins during mammalian embryogenesis: expressionof the alpha-vascular but not the gamma-enteric isoform in dif-ferentiating striated myocytes. J Cell Biol 109:2929–2937
35. Schultz E (1978) Changes in the satellite cells of growingmuscle following denervation. Anat Rec 190:299–311
36. Schultz E, Jaryszak DL (1985) Effects of skeletal muscle re-generation on the proliferation potential of satellite cells.Mech Ageing Dev 30:63–72
37. Schultz E, Lipton BH (1982) Skeletal muscle satellite cells:changes in proliferation potential as a function of age. MechAgeing Dev 20:377–383
38. Skalli O, Gabbiani G, Babai F, Seemayer TA, Pizzolato G,Schurch W (1988) Intermediate filament proteins and actinisoforms as markers for soft tissue tumor differentiation andorigin. II. Rhabdomyosarcomas. Am J Pathol 130:515–531
39. StClair JA, Meyer S, Demarest SD, Ham RG (1992) Improvedmedium with EGF and BSA for differentiated human skeletalmuscle cells. Muscle Nerve 15:774–779
40. Vaziri H, Dragowska W, Allsopp RC, Thomas TE, Harley CB,Lansdorp PM (1994) Evidence for a mitotic clock in human
hematopoietic stem cells: loss of telomeric DNA with age.Proc Natl Acad Sci USA 91:9857–9860
41. Webster C, Blau HM (1990) Accelerated age-related decline inreplicative life-span of Duchenne muscular dystrophy myo-blasts – implications for cell and gene therapy. Somat CellMol Genet 16:557–565
42. Widmer H, Hamann M, Baroffio A, Bijlenga P, Bader CR(1995) Voltage-dependent potassium current precedes fusionof human muscle satellite cells (myoblasts). J Cell Physiol162:52–63
43. Yablonka-Reuveni Z, Quinn LS, Nameroff M (1987) Isolationand clonal analysis of satellite cells from chicken pectoralismuscle. Dev Biol 119:252–259
44. Yablonka-Reuveni Z, Rivera AJ (1994) Temporal expressionof regulatory and structural muscle proteins during myogene-sis of satellite cells on isolated adult rat fibers. Dev Biol164:588–603
45. Yao SN, Kurachi K (1993) Implanted myoblasts not only fusewith myofibres but also survive as muscle precursor cells. JCell Sci 105:957–963
46. Zhang M, McLennan IS (1994) Use of antibodies to identifysatellite cells with a light microscope. Muscle Nerve 17:987–994
57