hematopoietic stem cell exhaustion impacted by p18ink4c and p21cip1/waf1 in opposite manners
TRANSCRIPT

doi:10.1182/blood-2005-02-0685Prepublished online October 18, 2005;
Hui Yu, Youzhong Yuan, Hongmei Shen and Tao Cheng p21Cip1/Waf1 in opposite mannersHematopoietic stem cell exhaustion impacted by p18INK4C and
(1880 articles)Transplantation � (166 articles)Stem Cells in Hematology �
(3131 articles)Hematopoiesis and Stem Cells � (231 articles)Cell Cycle �
Articles on similar topics can be found in the following Blood collections
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://bloodjournal.hematologylibrary.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
digital object identifier (DOIs) and date of initial publication. theindexed by PubMed from initial publication. Citations to Advance online articles must include
final publication). Advance online articles are citable and establish publication priority; they areappeared in the paper journal (edited, typeset versions may be posted when available prior to Advance online articles have been peer reviewed and accepted for publication but have not yet
Copyright 2011 by The American Society of Hematology; all rights reserved.20036.the American Society of Hematology, 2021 L St, NW, Suite 900, Washington DC Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

1
Hematopoietic stem cell exhaustion impacted by p18INK4C and p21Cip1/Waf1 in opposite manners
Hui Yu 1,2,3, Youzhong Yuan1,2,3, Hongmei Shen1,2, Tao Cheng1,2,4 1Department of Radiation Oncology, University of Pittsburgh School of Medicine and
2Stem Cell Biology Laboratory, University of Pittsburgh Cancer Institute, Pittsburgh PA 15213 3These authors contributed equally to this work. 4Correspondence to: Tao Cheng, MD Hillman Cancer Center Research Pavilion 5117 Center Ave, Room 2.42e Pittsburgh, PA. 15213-1863 Tel: 412-623-3249 Fax: 412-623-7778 E-mail: [email protected] Key words: hematopoietic stem cell, self-renewal, exhaustion, aging, transplantation, cell cycle regulators Running title: Stem cell exhaustion Word counts: Abstract-204; Main body (including methods)-3825; Methods-854; Display-4 figures and 1 table; Legends-1124.
Blood First Edition Paper, prepublished online October 18, 2005; DOI 10.1182/blood-2005-02-0685
Copyright © 2005 American Society of Hematology
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

2
Abstract: Transplantation-associated stress can compromise the hematopoietic potential of
hematopoietic stem cells (HSCs). As a consequence, HSCs may undergo so called
“exhaustion” in serial transplant recipients, for which the cellular and molecular bases are not
well understood. Hematopoietic exhaustion appears to be accelerated in the absence of
p21Cip1/Waf1 (p21), a cyclin-dependent kinase inhibitor (CKI) in irradiated hosts. Our recent
study demonstrated that unlike loss of p21, deletion of p18 INK4C (p18), a distinct CKI, results
in improved long-term engraftment largely owing to increased self-renewing divisions of HSCs
in vivo. We show here that HSCs deficient in p18 sustained their competitiveness to wild-type
HSCs from unmanipulated young mice, and retained multilineage differentiation potential after
multiple rounds of serial bone marrow transfer over a period of more than 3 years. Further,
p18 absence significantly decelerated hematopoietic exhaustion caused by p21 deficiency.
Such an effect was shown to occur at the stem cell level, likely by a counteracting mechanism
against the cellular senescence outcome. Our current study provides new insights into the
distinct impacts of these cell cycle regulators on HSC exhaustion and possibly HSC aging as
well under proliferative stress, thereby offering potential pharmacological targets for
sustaining the durability of stressed HSCs in transplant or elderly patients.
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

3
Introduction
Hematopoietic stem cells (HSCs) are the most important element for establishing the long-term
engraftment of hematopoietic transplants in recipients. Thus, the sustained self-renewal potential of
donor HSCs is critical for maintaining the long-term durability of the graft. While HSCs are thought to
be capable of self-regeneration in vivo over a lifetime without an apparent limit under homeostatic
conditions,1,2 it is well known that the repopulating ability of HSCs can be significantly compromised in
transplant recipients.3-8 Studies by several laboratories have demonstrated that the functional HSC
units reach only 4-10% of normal levels after each transplantation4,7-10 and as a consequence, serial
bone marrow transfer can only sustain hematopoiesis for 4-6 rounds in irradiated mouse recipients.
The limited repopulating ability of HSCs during serial transplantation has led to one view that the self-
renewal ability of HSCs is intrinsically limited.11 But extrinsic factors such as the transplantation
procedure4 and the irradiated bone marrow microenvironment7 are also likely involved. Since a
substantial portion of stem cell transplants still involve the use of total body irradiation (TBI) (though
not with an ablative dose nowadays) as a host conditioning regime, it is of clinical relevance to define
an effective approach for sustaining the self-renewal potential of transplanted HSCs in the irradiated
recipients. Despite the importance of this issue, the potential exhaustion of donor HSCs in transplant
recipients has not been resolved at the molecular level.
Given the limited success of in vitro HSC expansion to date, in vivo manipulations of HSCs appear to
be important alternatives to enhance HSC therapy. In addition to the administration of hematopoietic
growth factors2 and the alteration of homing receptors (such as CXCR4 and CD26)12,13 or micro-
environmental elements (such as the stem cell “niche”),14 direct manipulation of the intracellular self-
renewal machinery is an important strategy currently being explored. Cell cycle regulators have been
demonstrated to be intrinsically involved in the self-renewal of adult stem cells.15 In particular, cyclin-
dependent kinase inhibitors (CKIs) appear to have an important role in modulating the self-renewing
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

4
potential of stem cells. Interestingly, different CKIs appear to affect self-renewal in distinct manners.
For instance, in the absence of p21Cip1/Waf1 (p21 hereafter), the founding member of CKIs, the
hematopoietic potential of HSCs was nearly exhausted after two rounds of bone marrow
transplantation.10 In contrast, we have recently documented an engraftment advantage of the
hematopoietic cells deficient in p18INK4C (p18 hereafter), a member of the INK4 family of CKIs.16 This
advantage appears to be largely if not solely due to increased self-renewal of transplanted HSCs in
vivo16 rather than a general increase in proliferation of the donor cells (Shen et al, manuscript in
preparation).
Based on our previous results, we sought to examine whether the known exhaustion of transplanted
HSCs in irradiated hosts can be significantly overcome or minimized by suppressing p18 expression
during prolonged long-term engraftment. To this end, we have focused on the regenerative ability of
transplanted p18-/- HSCs, in comparison with that of HSCs deficient in p21 or in both p18 and p21,
following serial bone marrow transplants over an extended time period. Our current study
demonstrates that deleting p18 in HSCs is effective in sustaining the durability of transplanted HSCs
beyond the lifetime of a mouse, and this deletion is able to substantially compensate for the
deleterious effect of p21 deletion on hematopoietic repopulation, likely due to the opposite effects of
p18 and p21 on HSC senescence under stress conditions.
Materials and Methods
Mice
p18+/- mice and p18-/-p21-/- double knockout mice in C57BL/6;129/Sv background were kindly provided by David
Franklin at Purdue University (p18+/- mice were originally created by Franklin et al in Yue Xiong’s laboratory17).
p18-/- and p18+/+ littermates were produced from p18+/- breeding pairs that are maintained in our laboratory.
p21-/- mice and p21+/+ littermates were kindly provided by Andrew Stewart at University of Pittsburgh (the p21+/-
breeding mice in C57BL/6;129/Sv background were purchased from the Jackson Laboratories [Bar Harbor, ME]
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

5
and were originally created in Tyler Jacks’ laboratory18). Female C57BL/6129/SvF1 recipients at the age of 2
months were purchased from the Jackson Laboratories. All the procedures involved in the mouse work were
approved by the Institutional Animal Care and Use Committee of University of Pittsburgh.
Serial bone marrow transplantation (sBMT)
Bone marrow nucleated cells (BMNCs) (2 x 106/host) from male p18+/+ or p18-/- donor mice at the age of 2
months were transplanted into lethally irradiated (10 Gy) female wild-type recipients (2 months old). The same
procedure was applied in secondary, tertiary and quaternary recipients at intervals as indicated in Figure 1A.
Three animals in each group were sacrificed after each sBMT for measuring the cobble-stone area forming cell
(CAFC) and colony forming cell (CFC) in bone marrow as previously described.10,16 Blood was collected 12
months after the 4th round of sBMT to measure the donor contribution with the semi-quantitative PCR for Y-
chromosome.10,16
Competitive bone marrow transplantation (cBMT) coupled with serial transfer
The cBMT procedure was detailed in our previous publication16 and is further illustrated in Figure 2A. Briefly,
test cells (2 x 106 BMNCs/host) from p18-/- or p18-/-p21-/- male mice and an equal number of wild-type BMNCs
(competitor cells) from male mice (wild-type litter mates in primary cBMT and then wild-type C57BL/6129/SvF1
mice of 2 months old in subsequent rounds of cBMT) were co-transplanted into lethally-irradiated (10 Gy)
female recipients. The competitor cells from unmanipulated marrow were repetitively used in each sequential
cBMT as a standard measure for determining the repopulation ability of varied types of the original input cells
over the course of the serial transfer. Blood was collected after each cBMT and the relative contribution of test
cells (-/-) to the competitor cells (+/+) in the reconstituted host was quantified by the semi-quantitative PCR as
previously described.16 Some mice were sacrificed after each cBMT for the genotypic analyses in bulk BMNCs,
different cell lineages, and different hematopoietic compartments by the PCR described below.
Flow cytometric analysis and cell sorting
One of the well-characterized phenotypes for murine HSC, CD34-Lin-c-Kit+Sca-1+(CD34-LKS)19, was applied in
our current study. The method used to quantify the frequency of CD34-LKS in bone marrow was previously
described.16 To isolate the cells, Sca-1+ cells were enriched with the EasySep Kit (StemCell
Technologies,Vancouver, BC, Canada) according to the manufacturer’s protocol. The cells were stained with a
mixture of biotinylated antibodies against mouse CD3, CD4, CD8, B220, Gr-1, Mac-1 and TER-119 (Caltag,
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

6
Burlingame, CA), then co-stained with streptavidin-PE-Cy7, anti-Sca-1-PE, anti-c-Kit-APC and anti-CD34-FITC
(BD Pharmingen, San Diego, CA). Propidium iodide was used for dead cell exclusion. CD34-LKS cells were
sorted into 96-well plates (Becton Dickinson, Franklin Lakes, NJ) at one cell per well using the MoFlo High-
Speed Cell Sorter with subsystems of Cyclone Automated Cloner and Sort Master Droplet Control. For sorting
the mature cells in different lineages, BMNCs were stained with a mixture of anti-CD3-PE, anti-Gr-1-PE-Cy7,
anti-Mac-1-APC and anti-B220-PE-Texas Red (BD Pharmingen). Cells of different lineages including T (CD3+),
B (B220+), and myeloid (Mac-1+) were sorted into separate tubes and then lysed for genotyping by PCR
analysis.
Clonal cell culture
Single CD34-LKS cells were deposited into 96-well plates (one cell per well). Each well contained 100µl of
IMDM supplemented with 50 ng/ml of Flt3 ligand (Flt3-L), 50ng/ml of SCF, 10ng/ml of TPO and 10ng/ml of IL-3
(PeproTech, Rocky Hill, NJ). The colony forming cell (CFC) assay with bulk BMNCs was performed according
to the manufacturer’s instructions (Stem Cell Technologies). 7~10 days after initiation of these clonal cultures,
sufficient colonies were harvested under a microscope and lysed for subsequent PCR reaction.
PCR for tracing donor contribution
Primer sequences for amplifying the p18 and p21 mutant/wild-type alleles were described in our previous
publications.10,16 In addition to the genotyping PCR for p18, the following primer pair was used to confirm the
absence of p21 in p18/p21 double mutant cells: p21null-F (5’-GCG AGG ATC TCG TCG TGA C-3’), and p21
null-R (5’-TCA TCA ATT TAT GCA GAC-3’). In the PCR for quantifying the Y-chromosome DNA (Sry), the
following primers were used: Sry-F (5’-TGG GAC TGG TGA CAA TTG TC-3’), Sry-R (5’-GAG TAC AGG TGT
GCA GCT CT), Myo-F (5’-TTA CGT CCA TCG TGG ACA GC-3’) and Myo-R (5’-TGG GCT GGG TGT TAG
TCT TA-3’). Nucleated blood cells were lysed in 1X PCR buffer containing 2.5mM MgCl2 and 20µg/ml
Proteinase K for 1 hour at 60°C, followed by an inactivation of the reaction for 20 min at 95°C. The parameters
for thermal cycling of PCR were as follows: 30s at 94°C, 30s at 57°C and 1 min at 72°C (30 cycles for the semi-
quantitative analysis with bulk blood cells and 35 cycles for the single colony analysis).
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

7
Results
In vitro activity of p18-/- hematopoietic cells from the serially-transplanted recipients We first examined the exhausting effect of irradiated hosts on transplanted HSCs with the classical
serial bone marrow transplantation (sBMT), in which no competitor cells were co-transplanted (Figure
1A). Bone marrow nucleated cells (BMNCs) from male p18+/+ or p18-/- mice were transplanted into
lethally-irradiated female mice and sBMT was sequentially performed for 4 rounds with an interval of
3-4 months. During each sBMT, the in vitro activity of both HSCs and hematopoietic progenitor cells
(HPCs) from p18-/- recipient mice was significantly higher than that from p18+/+ transplanted mice, as
examined by the in vitro surrogate assays for HSCs and HPCs, namely the cobblestone-area forming
cell (CAFC) and colony forming cell (CFC), respectively (Figure 1B). Of note, CAFC frequency at day
35 in p18-/- transplanted mice was five times higher than that in p18+/+ transplanted mice after
secondary sBMT. CFC frequency was maintained at a normal level (that of unmanipulated bone
marrow) in p18-/- transplanted mice while it gradually declined in p18+/+ transplanted mice during 3
rounds of sBMT. These results contrast with the accelerated hematopoietic exhaustion in the
absence of p21 during sBMT, as shown in a previous study.10
Sustained in vivo hematopoiesis of p18-/- HSCs after multiple BMTs
Our next step was to determine the in vivo long-term repopulating ability (LTRA) of HSCs 12 months
after quaternary sBMT, by quantifying the presence of HSC progeny in blood (Figure 1A). PCR-
based semi-quantitative analysis for the Y chromosome-specific sequence (Sry) was used to track
cellular origin.10 Based on the standardization simultaneously generated under identical PCR
conditions, we calculated the percentage of donor cell contribution to recipient hematopoiesis. As we
expected, most recipients (4 of 6) of p18+/+ HSCs had undetectable levels of donor cells in their blood
(Figure 1C). In contrast, the original p18-/- male donor-derived cells accounted for 66.7% on average
(60-73%) of whole mature blood cells in all recipients (N=7). By an analysis with the single limiting
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

8
dose, we estimated that there was only one HSC unit (being able to give rise to detectable donor cells
in the blood of a recipient animal) per 5 x 106 BMNCs in p18+/+ transplanted recipients after 4 rounds
of sBMT in contrast to one HSC per 1-2 x 105 BMNCs in normal unmanipulated marrow.20 The 100%
engraftment rate in the p18-/- transplanted group precluded us from directly estimating the frequency
of functional units of HSCs in the p18-/- recipients. However, p18-/- HSCs at this later point could be
estimated following serial competitive bone marrow transplantation (cBMT) for three times over 30
months, subsequent to our previous study (Figure 2A).16
In the cBMT experiment, equal numbers of p18-/- BMNCs and competitor cells (wild-type BMNCs
freshly isolated from untransplanted mice) were co-transplanted into lethally-irradiated recipients in
each cBMT. Because LTRA and HSC frequency in mouse wild-type BMNCs have been well
documented, the fixed number (2 x 106) of unmanipulated competitor cells can serve as a standard
measure for varied types of input cells to be compared (Figure 2A). Blood was collected 6-14 months
after each cBMT and the relative contribution to hematopoiesis from each genotype (p18-/- versus
wild-type) was quantified by the semi-quantitative PCR assay.16 After 3 rounds of cBMT in 30
months, LTRA of p18-/- HSCs was still equivalent (54.9%) to that of the competitor cells (Figure 2B
and Figure 2C), which indicates there is no apparent exhaustion of hematopoietic potential as
compared to that of the wild-type transplanted cells at this time point.
An opposing effect of p18 absence over p21 absence on HSC exhaustion
To test whether p18 absence can counteract the known accelerated hematopoietic exhaustion
caused by p21 absence,10 we directly assessed the long-term engraftment of hematopoietic cells
from mice deficient in both p21 and p18 (p18-/-p21-/-, hereafter, abbreviated for the double-
knockout).21 LTRA of HSCs from p18-/-p21-/- mice was compared to that from p18-/- mice using the
cBMT assay (Figure 2A).16 In contrast to a previous study showing early exhaustion of hematopoietic
reconstitution in the absence of p21 after sBMT,10 p18-null was able to overcome the accelerated
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

9
hematopoietic exhaustion caused by p21 deletion as demonstrated by 51.1% and 34.1% of
reconstitution level in secondary and tertiary recipients of the p18-/-p21-/- cells, respectively (Figure 2B
and Figure 2C). As a confirmatory result to the previous report10, we were also able to observe an
accelerated hematopoietic exhaustion in a different experiment where p21-/- BMNCs were co-
transplanted with the same competitor cells in the current study (Figure 2D). The fact that p18-/-p21-/-
HSCs were able to sustain their LTRA after 3 rounds of cBMT suggests that the absence of p18 can
significantly mitigate the negative impact of p21 deletion on the LTRA of HSCs. Therefore, these
data provide direct evidence for the opposite effects of p18 and p21 on the hematopoietic potential of
HSCs in the context of serial transplantation.
Multilineage differentiation of p18-/- or p18-/-p21-/- HSCs
Multilineage analysis of bone marrow cells from tertiary recipients was performed to determine
whether p18-/- or p18-/-p21-/- HSCs maintained their ability to give rise to both myeloid and lymphoid
lineages after multiple cBMTs performed over a prolonged period of time. The
immunophenotypically-defined myeloid (granulocytes and monocytes) and lymphoid (T and B
lymphocytes) cells were sorted for the determination of p18 genotype (p18-/- vs. p18+/+). As shown in
Figure 3, substantial engraftment levels of p18-/- cells were seen in both myeloid and lymphoid
lineages, indicating the multilineage potential of transplanted p18-/- or p18-/-p21-/- HSCs for 30 months
over multiple cBMTs (at the cellular age of 36 months). Notably, the relative contribution of p18-/-
donor cells to competitor cells in myeloid lineage was higher than that observed in lymphoid lineage
(Figure 3B). This result is in agreement with previous studies by others that showed favored
myelopoiesis over lymphopoiesis in aged mice, which is indicative of ongoing aging of hematopoietic
cells in vivo.22-24 However, the relative decrease of T-lymphopoiesis was significantly improved in the
p18-/- group in comparison with that in the p18-/-p21-/- group (p < 0.05), suggesting distinct roles of p18
and p21 in T-lymphopoiesis in addition to their direct effects on HSCs.
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

10
Representation of p18-/-, p18-/-p21-/- or p21-/- genotype in the HSC compartment
To further distinguish the contribution of test cells versus competitor cells in the stem cell
compartment, we isolated HSCs with the primitive immunophenotype for murine HSCs, CD34-LKS,19
from the recipients of cBMT and determined their genotype at the single cell level. It should be noted
that this immunophenotype with LTRA does not seem to change in aged or transplanted mice based
on the data by others24 and our own unpublished observation. Single HSC cells were sorted into 96-
well plates (one cell per well) and cultured in medium with appropriate cytokine stimuli to form
colonies (more than 30 cells/colony) in 7-10 days, which allowed us to efficiently track the origin of
individual HSCs that were initially isolated from the marrow. Sufficient numbers of colonies were
randomly picked up for the genotyping PCR analysis on the marrow harvested at varied time points
after cBMT (Table 1). Data demonstrated that the majority of CD34-LKS cells were of p18-/- or p18-/-
p21-/- origin after tertiary or quaternary cBMT. Interestingly, the dominance of p18-/-p21-/- genotype
was higher than that of p18-/- alone. In the p21-/- control group, p21-/- HSC progeny were barely
detectable in blood after primary cBMT (Figure 2D) while 63.3% of CD34-LKS cells and 18.9% of
CFCs remained of the p21-/- origin. When normalized to the bone marrow cellularity at the time of
sacrifice, the absolute yield of CD34-LKS cells was in fact significantly lower in the p21-/- transplant
group than in p18-/- recipients.
To further validate the different impacts of p21 and p18 on a per-cell basis, we cultured the CD34-LKS
mixed populations isolated from either the p21-/- or p18-/-p21-/- cBMT group for 14 days and then
checked the ratio of p21-/- or p18-/-p21-/- differentiated cells versus wild-type competitor progeny by the
semi-quantitative PCR. The CD34-LKS cells in both groups were able to differentiate into myeloid
cells during the culture. Consistent with our in vivo findings, the initial higher representation (63.3%)
at HSC level dropped to 25% of HSC progeny in the p21-/- group, whereas the representation
remained (78.9% to 83%) in the p18-/-p21-/- group (Figure 4). Together, these data suggest that a
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

11
progressive decrease of function per HSC is ultimately followed by a possible depletion of the HSC
pool due to the absence of p21, and that such a process can be substantially overcome by deleting
p18 at the stem cell level regardless of p21 absence.
Discussion
Our current study demonstrates that transplanted p18-/- HSCs sustain their self-renewing ability at a
comparable level to unmanipulated wild-type HSCs after multiple serial transfers in both competitive
and non-competitive repopulation models. Furthermore, p18 absence is able to significantly
counteract the accelerated exhaustion of hematopoietic repopulation resulting from p21 deficiency.
Therefore, we have defined for the first time that hematopoietic exhaustion in serial transplant models
can be overcome by targeting a specific cell cycle inhibitor and different cell cycle inhibitors can have
opposite impacts on the outcome.
Both extrinsic and intrinsic factors may account for HSC exhaustion in transplant recipients.
Regarding the extrinsic factors, at least two considerations have been given from previous studies by
others. One is the failure to achieve a physiological concentration of HSCs due to insufficient
mitogens for their regeneration upon the dilution of original donor HSCs in new recipients.2 Another
one is the physical disruption due to transplantation procedure or/and radiation damage (if TBI is used)
which can result in homing defect25 or permanent loss of self-renewal potential6,7,26 of the transplanted
HSCs. However, it is unclear how these extrinsic effects are transmitted onto the intrinsic properties
of transplanted HSCs. In fact, the intrinsic mechanism may also affect HSC exhaustion in an
autonomous manner. Because serial bone marrow transfer usually spans the lifetime of a mouse or
longer beside its imposed transplantation stress, the resultant stem cell exhaustion has been thought
to be attributable to the replicative senescence of the transplanted HSCs, resembling aging of HSC.11
This link is supported by the similarity of limited HSC self-replication in both serial transplant
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

12
recipients and aged mice.27,28 While the aging effect might be cooperative with the extrinsic factors in
leading to hematopoietic exhaustion, the hematopoietic exhaustion was distinguishable from the aged
phenotype to a certain extent in the absence of p18. On one hand, we observed the disappearance
of hematopoietic exhaustion in p18-/- groups as compared to the wild-type cells after serial
transplantation over 3 years, while on the other hand we observed a favored myeloid differentiation of
HSCs that has been similarly observed in old mice and is now considered as a sign for HSC aging.22-
24 Notably, however, the LTRA after each cBMT decreased as compared to the previous one and this
reduction coincided with a relative increase in HSC frequency as defined by the immunophenotype
(Table 1). We observed a dominant p18-/- phenotype in the HSC compartment, but the representation
of p18-/- phenotype in blood did not correlate with the ratio in the stem cell pool after tertiary cBMT.
The reverse relationship was more overt in the p18-/-p21-/- group and the most dramatic in the p21-/-
group (Table 1 and Figure 2D). This suggests a decline in repopulation potential per HSC rather than
an actual loss of the phenotypic HSCs after transplantation. This observation is consistent with
previous studies in both transplanted and untransplanted aged mice.29,30 The similarity between
HSCs in transplant recipients and aged mice suggests that extensive cell divisions may pose
replicative stress on HSCs and ultimately lead to premature cellular senescence. Interestingly, p18
absence is able to significantly counteract the process through a distinct mechanism, yet to be
defined.
The molecular mechanisms that underlie HSC exhaustion are poorly understood. Given that
replicative cellular senescence is likely involved, telomere length was hypothesized to be a critical
cellular timer in mediating the senescent outcome. This hypothesis was supported by the observation
of shortened telomere length after serial transplantation31 and early exhaustion of transplanted HSCs
deficient in telomerase.32 However, overexpression of telomerase in HSCs was not sufficient to
overcome hematopoietic exhaustion, although it could prevent telomere shortening during serial
transplantation.33 In our study, we did not observe a difference in telomere length between p18+/+ and
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

13
p18-/- sBMT groups (data not shown). Therefore, these studies suggest that telomere erosion is not a
sole or indispensable mechanism for stem cell exhaustion.
The p53 and Rb pathways are considered to be the two major activating mechanisms for cellular
senescence in many other cell types.34 p21 is a primary transcriptional target of p53 in mediating cell
cycle arrest, and p16 is a key INK4 protein inhibiting the activity of CDK4/6 in the Rb pathway. Both
cell cycle inhibitors were recently indicated to be involved in radiation-induced HSC senescence.35
However, because p21 is expressed at a higher level in quiescent HSCs,36,37 it is unlikely to play an
indispensable role in HSC senescence. Instead, the Rb pathway might be a more dominant
mechanism for HSC senescence but not HSC quiescence as p16 and its alternative reading frame,
p19ARF were up-regulated in the HSCs from serially transplanted mice (unpublished data in our
laboratory). We therefore hypothesize that under the replicative stress caused by transplantation,
HSCs may undergo senescence, and such a process is accelerated in the absence of p21 due to a
higher cycling status of HSC. In fact, given the increased number of HSCs and normal hematopoietic
potential in p21-/- under homeostatic conditions10 and the unperturbed function of HSCs after ex vivo
targeting of p2138,39, we suggest that the senesent state of p21-/- HSCs in the serial recipients reflect
a secondary effect of p21 absence on HSCs under the repetitive proliferative stress. In the absence
of p18, the senescent outcome can be substantially overcome or delayed due to the antagonistic
effect of p18 absence on the cellular senescence. But it remains to be determined with regard to how
the effect of p18 deletion is mediated by other molecules to compensate the consequence of p21 loss
on hematopoietic exhaustion and more importantly, how p18 interacts with p21 physiologically in
HSCs. Nevertheless, our current study sets the stage regarding our current understanding of the
distinct roles that p18 and p21 play in stem cell regulation. An understanding of the different impacts
that these two CKIs have in HSC kinetics offers molecular targets for possible pharmacological
intervention to improve stem cell functioning in transplant or elderly patients.
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

14
Acknowledgments
This work was supported by NIH grants (DK02761, HL70561) and the Scholar Award from American
Society of Hematology (2003). We specially thank Drs David Franklin and Andrew Stewart for
providing the knockout mouse strains.
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

15
References
1. Harrison DE. Proliferative capacity of erythropoietic stem cell lines and aging: an overview. Mech Ageing Dev. 1979;9:409-426
2. Iscove NN, Nawa K. Hematopoietic stem cells expand during serial transplantation in vivo without apparent exhaustion. Curr Biol. 1997;7:805-808
3. Siminovitch L, Till JE, McCulloch EA. Decline in Colony-Forming Ability of Marrow Cells Subjected to Serial Transplantation into Irradiated Mice. J Cell Physiol. 1964;64:23-31
4. Harrison DE, Astle CM, Delaittre JA. Loss of proliferative capacity in immunohemopoietic stem cells caused by serial transplantation rather than aging. J Exp Med. 1978;147:1526-1531
5. Hellman S, Botnick LE, Hannon EC, Vigneulle RM. Proliferative capacity of murine hematopoietic stem cells. Proc Natl Acad Sci U S A. 1978;75:490-494
6. Harrison DE, Astle CM. Loss of stem cell repopulating ability upon transplantation. Effects of donor age, cell number, and transplantation procedure. J Exp Med. 1982;156:1767-1779
7. Mauch P, Rosenblatt M, Hellman S. Permanent loss in stem cell self renewal capacity following stress to the marrow. Blood. 1988;72:1193-1196
8. Harrison DE, Stone M, Astle CM. Effects of transplantation on the primitive immunohematopoietic stem cell. J Exp Med. 1990;172:431-437
9. Pawliuk R, Eaves C, Humphries RK. Evidence of both ontogeny and transplant dose-regulated expansion of hematopoietic stem cells in vivo. Blood. 1996;88:2852-2858
10. Cheng T, Rodrigues N, Shen H, Yang Y, Dombkowski D, Sykes M, Scadden DT. Hematopoietic stem cell quiescence maintained by p21(cip1/waf1). Science. 2000;287:1804-1808
11. Bell DR, Van Zant G. Stem cells, aging, and cancer: inevitabilities and outcomes. Oncogene. 2004;23:7290-7296
12. Lapidot T. Mechanism of human stem cell migration and repopulation of NOD/SCID and B2mnull NOD/SCID mice. The role of SDF-1/CXCR4 interactions. Ann N Y Acad Sci. 2001;938:83-95
13. Christopherson KW, 2nd, Hangoc G, Mantel CR, Broxmeyer HE. Modulation of hematopoietic stem cell homing and engraftment by CD26. Science. 2004;305:1000-1003
14. Calvi LM, Adams GB, Weibrecht KW, Weber JM, Olson DP, Knight MC, Martin RP, Schipani E, Divieti P, Bringhurst FR, Milner LA, Kronenberg HM, Scadden DT. Osteoblastic cells regulate the haematopoietic stem cell niche. Nature. 2003;425:841-846
15. Cheng T, Scadden DT. Cell cycle entry of hematopoietic stem and progenitor cells controlled by distinct cyclin-dependent kinase inhibitors. Int J Hematol. 2002;75:460-465
16. Yuan Y, Shen H, Franklin DS, Scadden DT, Cheng T. In vivo self-renewing divisions of haematopoietic stem cells are increased in the absence of the early G1-phase inhibitor, p18INK4C. Nat Cell Biol. 2004;6:436-442
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

16
17. Franklin DS, Godfrey VL, Lee H, Kovalev GI, Schoonhoven R, Chen-Kiang S, Su L, Xiong Y. CDK inhibitors p18(INK4c) and p27(Kip1) mediate two separate pathways to collaboratively suppress pituitary tumorigenesis. Genes Dev. 1998;12:2899-2911
18. Brugarolas J, Chandrasekaran C, Gordon JI, Beach D, Jacks T, Hannon GJ. Radiation-induced cell cycle arrest compromised by p21 deficiency. Nature. 1995;377:552-557
19. Osawa M, Hanada K, Hamada H, Nakauchi H. Long-term lymphohematopoietic reconstitution by a single CD34- low/negative hematopoietic stem cell. Science. 1996;273:242-245
20. Harrison DE, Astle CM, Lerner C. Number and continuous proliferative pattern of transplanted primitive immunohematopoietic stem cells. Proc Natl Acad Sci U S A. 1988;85:822-826
21. Franklin DS, Godfrey VL, O'Brien DA, Deng C, Xiong Y. Functional collaboration between different cyclin-dependent kinase inhibitors suppresses tumor growth with distinct tissue specificity. Mol Cell Biol. 2000;20:6147-6158
22. Sudo K, Ema H, Morita Y, Nakauchi H. Age-associated characteristics of murine hematopoietic stem cells. J Exp Med. 2000;192:1273-1280
23. Geiger H, Rennebeck G, Van Zant G. Regulation of hematopoietic stem cell aging in vivo by a distinct genetic element. Proc Natl Acad Sci U S A. 2005;102:5102-5107
24. Rossi DJ, Bryder D, Zahn JM, Ahlenius H, Sonu R, Wagers AJ, Weissman IL. Cell intrinsic alterations underlie hematopoietic stem cell aging. Proc Natl Acad Sci U S A. 2005;102:9194-9199
25. Plett PA, Frankovitz SM, Orschell-Traycoff CM. In vivo trafficking, cell cycle activity, and engraftment potential of phenotypically defined primitive hematopoietic cells after transplantation into irradiated or nonirradiated recipients. Blood. 2002;100:3545-3552
26. Ross EA, Anderson N, Micklem HS. Serial depletion and regeneration of the murine hematopoietic system. Implications for hematopoietic organization and the study of cellular aging. J Exp Med. 1982;155:432-444
27. Ogden DA, Mickliem HS. The fate of serially transplanted bone marrow cell populations from young and old donors. Transplantation. 1976;22:287-293
28. Kamminga LM, van Os R, Ausema A, Noach EJ, Weersing E, Dontje B, Vellenga E, de Haan G. Impaired hematopoietic stem cell functioning after serial transplantation and during normal aging. Stem Cells. 2005;23:82-92
29. Spangrude GJ, Brooks DM, Tumas DB. Long-term repopulation of irradiated mice with limiting numbers of purified hematopoietic stem cells: in vivo expansion of stem cell phenotype but not function. Blood. 1995;85:1006-1016
30. Morrison SJ, Wandycz AM, Akashi K, Globerson A, Weissman IL. The aging of hematopoietic stem cells [see comments]. Nat Med. 1996;2:1011-1016
31. Allsopp RC, Cheshier S, Weissman IL. Telomere shortening accompanies increased cell cycle activity during serial transplantation of hematopoietic stem cells. J Exp Med. 2001;193:917-924
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

17
32. Allsopp RC, Morin GB, DePinho R, Harley CB, Weissman IL. Telomerase is required to slow telomere shortening and extend replicative lifespan of HSCs during serial transplantation. Blood. 2003;102:517-520
33. Allsopp RC, Morin GB, Horner JW, DePinho R, Harley CB, Weissman IL. Effect of TERT over-expression on the long-term transplantation capacity of hematopoietic stem cells. Nat Med. 2003;9:369-371
34. Ben-Porath I, Weinberg RA. The signals and pathways activating cellular senescence. Int J Biochem Cell Biol. 2005;37:961-976
35. Wang Y, Schulte BA, Larue AC, Ogawa M, Zhou D. Total body irradiation selectively induces murine hematopoietic stem cell senescence. Blood. 2005
36. Cheng T, Shen H, Rodrigues N, Stier S, Scadden DT. Transforming growth factor beta 1 mediates cell-cycle arrest of primitive hematopoietic cells independent of p21(Cip1/Waf1) or p27(Kip1). Blood. 2001;98:3643-3649
37. Walkley CR, McArthur GA, Purton LE. Cell Division and Hematopoietic Stem Cells: Not Always Exhausting. Cell Cycle. 2005;4
38. Stier S, Cheng T, Forkert R, Lutz C, Dombkowski DM, Zhang JL, Scadden DT. Ex vivo targeting of p21Cip1/Waf1 permits relative expansion of human hematopoietic stem cells. Blood. 2003
39. Zhang J, Attar E, Cohen K, Crumpacker C, Scadden D. Silencing p21(Waf1/Cip1/Sdi1) expression increases gene transduction efficiency in primitive human hematopoietic cells. Gene Ther. 2005;12:1444-1452
40. Harrison DE, Doubleday JW. Normal function of immunologic stem cells from aged mice. J Immunol. 1975;114:1314-1317
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

18
Legends
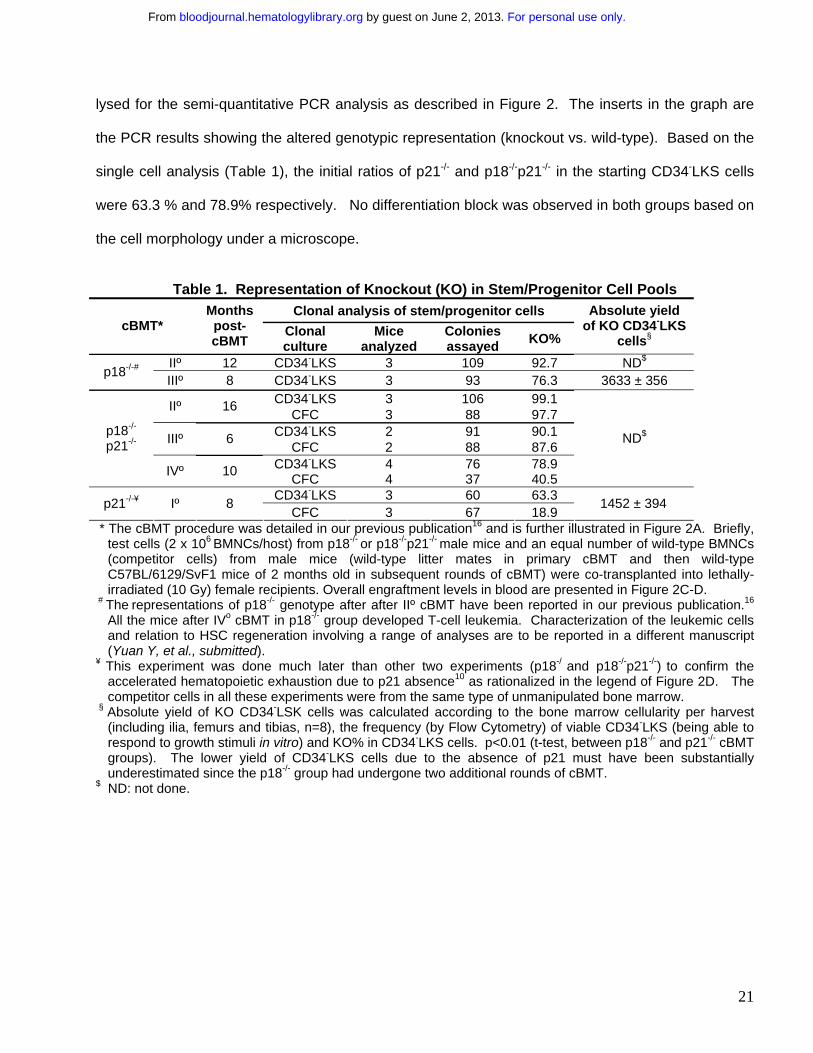
Figure 1. Continued presence of p18-/- HSC clones without apparent exhaustion during sBMT.
BMNCs were collected from p18-/- or p18+/+ male mice at 2 months of age and transplanted into
lethally irradiated (10 Gy) female recipients (2 x 106 cells/recipient). After long-term engraftment was
established in the primary recipients, the same dose of BMNCs were pooled from the primary
recipients and re-transplanted into lethally irradiated female recipients. The same procedure was
repeated for additional 3 times (A). “0o, I o, II o, III o and IVo” refer to untransplanted, 1st, 2nd, 3rd and
4th sBMT, respectively. “M” and “F” in parentheses indicate male and female respectively. Numbers
under the arrows indicate the months after each sBMT, and numbers in the parentheses are the
cellular age of original transplanted HSCs. In vitro hematopoietic activity was assayed with the
cobble-stone area forming assay (CAFC) and colony forming assay (CFC) as previously described.10
The frequencies of day 35 CAFC (left) and CFC (right) normalized to the untransplanted wild-type
control after different sBMTs are shown in the graphs (B). “*” indicates p < 0.05 (student’s t-test, n=3).
Open and filled bars indicate p18+/+ and p18-/- groups respectively. To further assess the exhaustion
of hematopoietic regeneration, peripheral blood was collected from the recipients 12 months after IVo
sBMT and the PCR-based semi-quantitative analysis for mouse Y chromosome-specific sequence
(Sry) normalized to the housekeeping gene myogenin (Myo) was applied to calculate the contribution
of the original male donor cells (C). The spleen or blood nucleated cells from male and female mice
were mixed at different ratios to acquire the standardization for the semi-quantitative analysis.
According to the standardization generated simultaneously as shown in the bottom panel, the
normalized percentages of donor cells in the blood are indicated under the PCR images in the top
and middle panels. Numbers above the gel images indicate individual p18-/- or p18+/+ transplant
recipients, and engraftment level (%) in each recipient is indicated under the gel image.
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

19
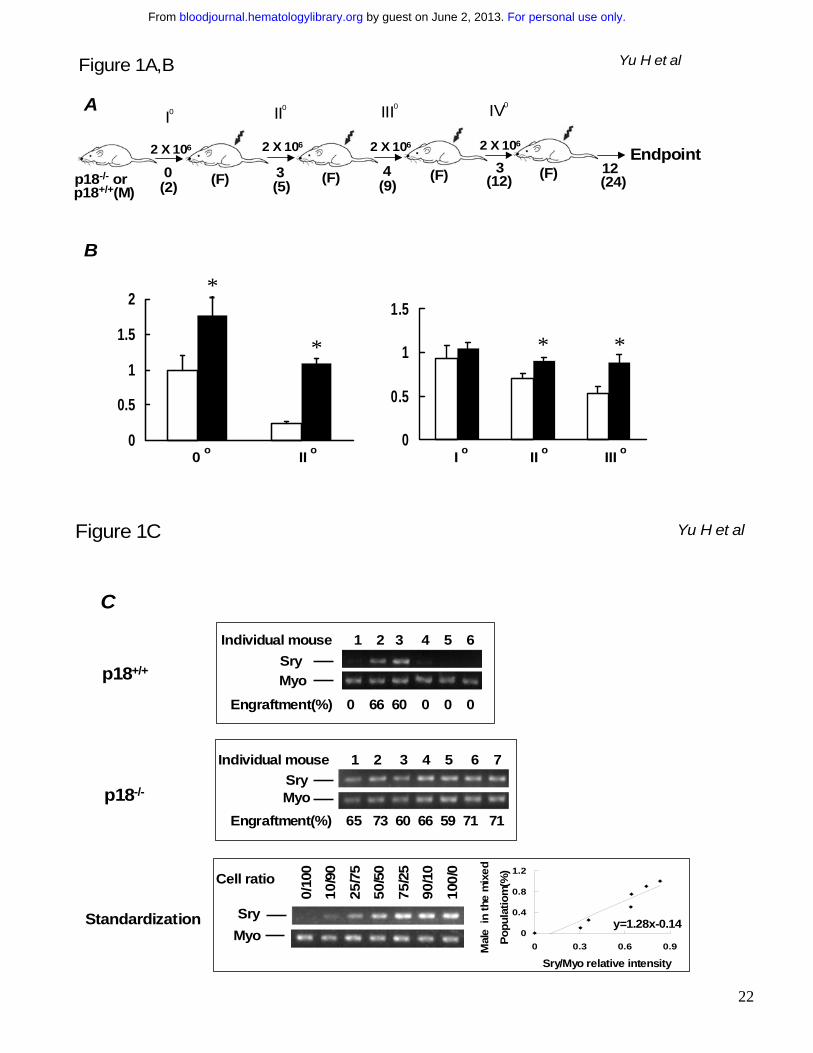
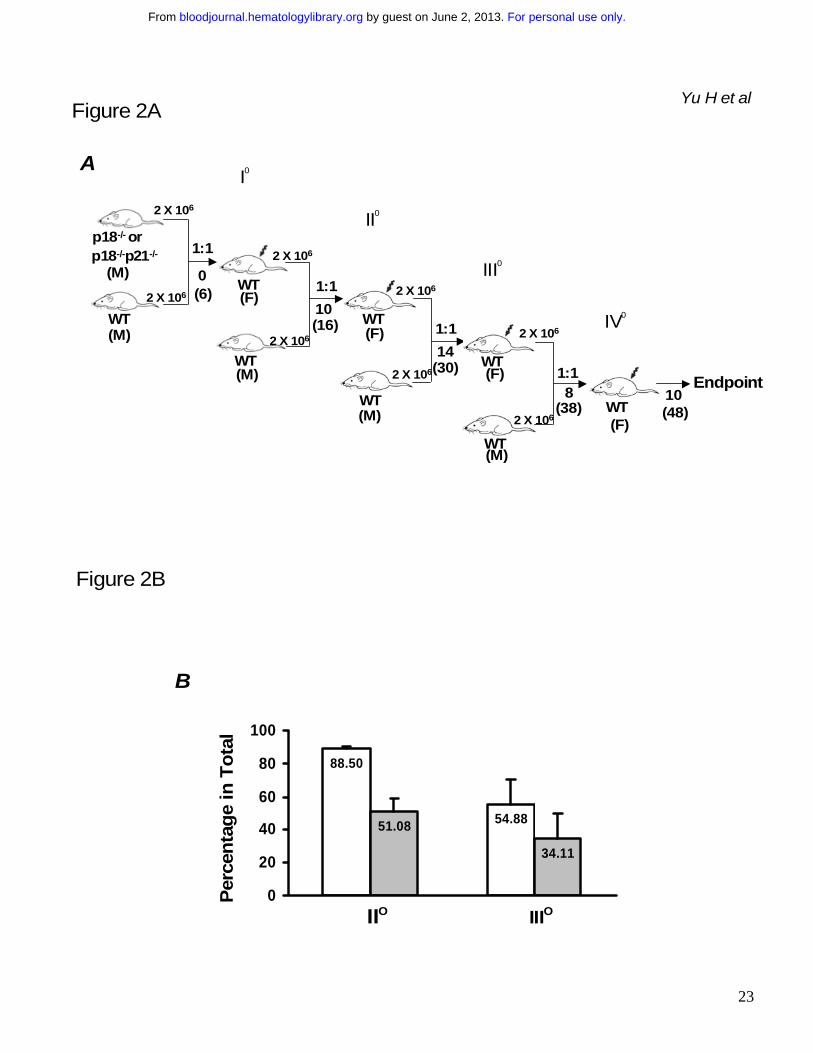
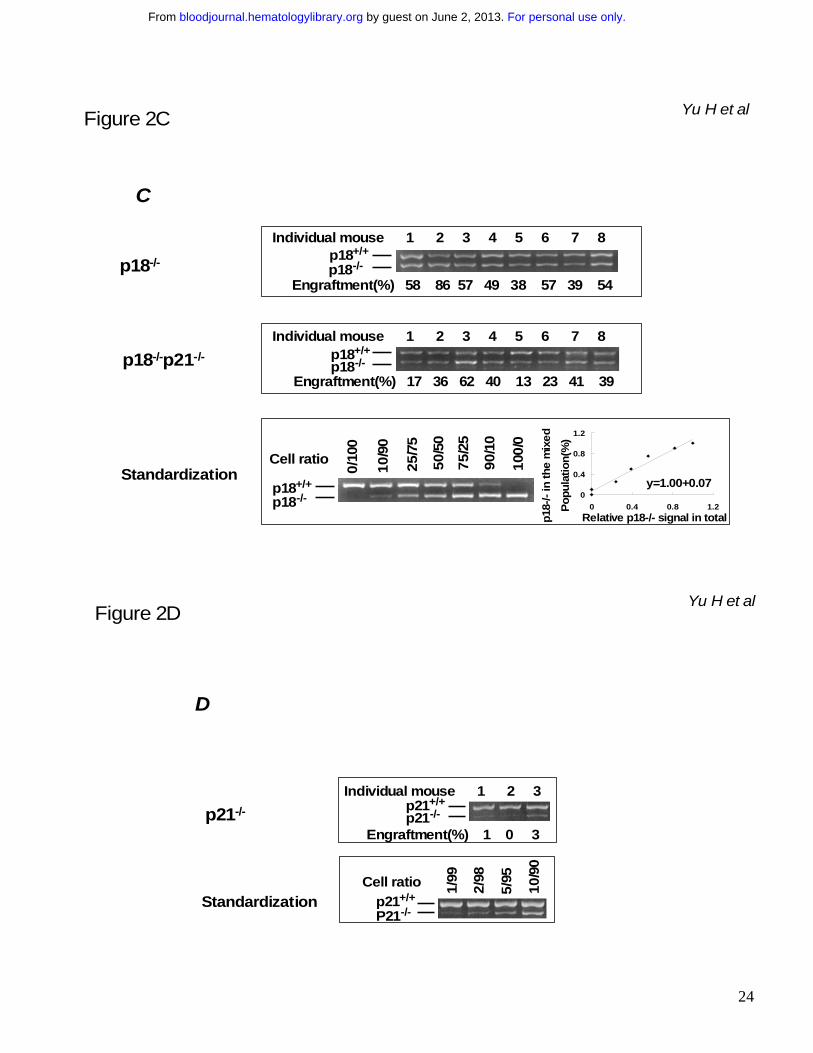
Figure 2. Sustained competitiveness of p18-/- HSCs after multiple rounds of cBMT. BMNCs
from p18-/- or p18-/-p21-/- knockout mice were mixed with an equal number (2x106/each) of competitor
cells (BMNCs from wild-type mice of 2 months old) and injected into lethally irradiated (10 Gy)
recipient mice. 10 months after primary cBMT, 2 x 106 BMNCs from the recipients were re-challenged
with an equal amount of the competitor cells and secondarily transplanted into lethally irradiated
recipients. This procedure was sequentially repeated twice (donor cells were pooled from 3-5
animals and there were 10 recipients/each group for each cBMT). The actual interval between each
cBMT is shown in the illustration (A). Some mice were kept longer than the interval “I o, II o and III o”
refer to 1st, 2nd and 3rd cBMT respectively. “1:1” means that an equal amount of test cells and
competitor cells were co-transplanted into the recipients. “M” and “F” in parentheses indicate male
and female respectively. Numbers under the arrows indicate the months after each cBMT, and
numbers in the parentheses are the cellular age of original transplanted HSCs. Peripheral blood was
collected from the recipients after long-term engraftment during each interval between sequential
cBMTs, and PCR-based semi-quantitative analysis as described in our previous study16 was
performed to determine the contribution of p18-/- or p18-/-p21-/- to the overall hematopoietic
reconstitution. The spleen or blood nucleated cells from wild-type and mutant mice were mixed at
different ratios to obtain a standardization curve for the semi-quantitative analysis. The average
levels of engraftment in blood from p18-/- or p18-/-p21-/- origin 12 months after secondary and 6months
after tertiary cBMT are summarized in the graph (B). Open and filled bars indicate p18-/-and p18-/-
p21-/- transplant groups respectively. Each group includes 8 animals and the difference between p18-
/- and p18-/-p21-/- is significant (p < 0.05) based on the student’s t-test. The representative results after
tertiary cBMT are shown (C) along with the standardization generated simultaneously (bottom panel).
Numbers above the gel images indicate individual animals, and the normalized percentages of test
cells of the total in blood are shown below the images. As a control to confirm the accelerated
exhaustion of hematopoietic repopulation in the absence of p21 as reported previously,10 the identical
cBMT procedure (A) was used to test the repopulating ability of BMNCs from p21-/- mice at the age of
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom

20
12 months (D). 8-12 months after the 1st cBMT, there was virtually no detectable p21-/- band in blood
after 30 cycles of the PCR, and there were two blood samples at a barely detectable level (1-3%) of
the engraftment after 35 cycles of the PCR as representatively shown here. This experiment was
intended to confirm a previous claim of HSC exhaustion due to p21 absence10. It was not performed
at the same time as other two groups but the competitor cells were from the same type of donor mice
as described in the Methods. Therefore, the results between p21-/- group and other groups though
with imperfection are still comparable since all these different test cells were directly assessed with
the same type of competitor cells in the irradiated hosts. The singular effect of age would not
substantially contribute to the exhaustion based on earlier studies by others showing no disadvantage
of aged bone marrow cells in the competitive repopulation model.4,27,40
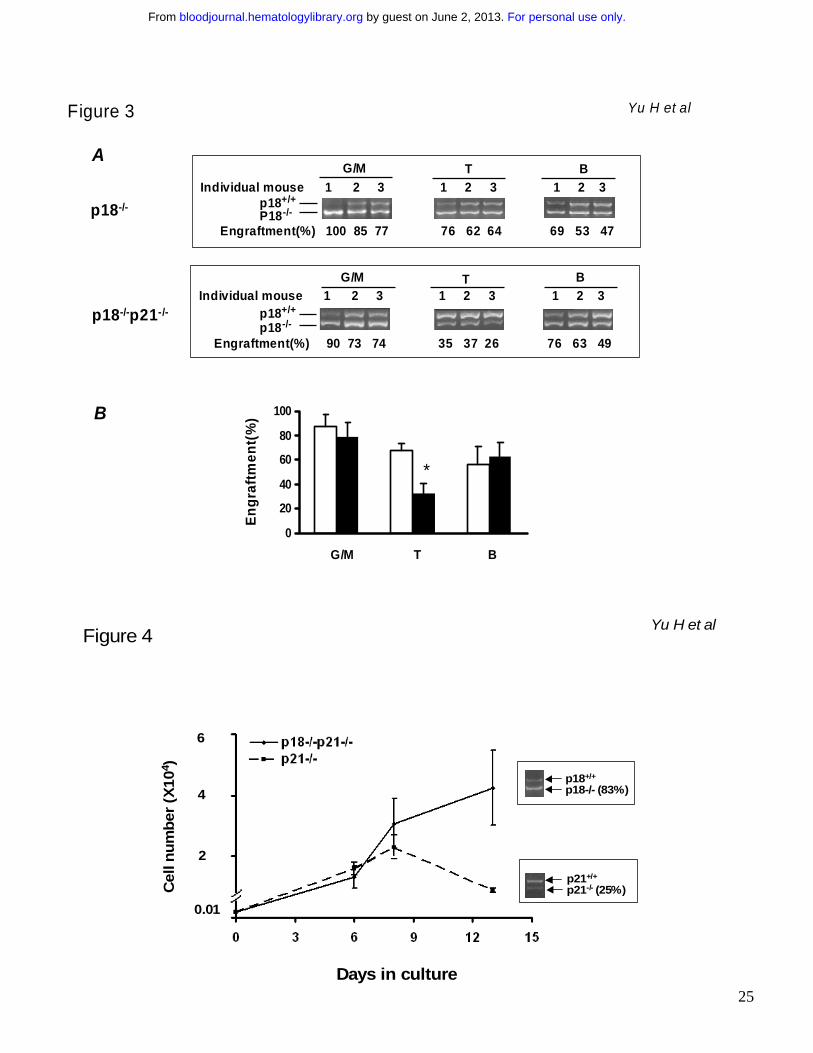
Figure 3. Multilineage differentiation that favors myelopoiesis after an extended long-term
engraftment. The recipients were sacrificed after tertiary cBMT and their BMNCs were collected and
stained with lineage markers. Myeloid (Mac-1+), T (CD3+) and B (B220+) cell lineages were sorted for
semi-quantitative PCR analysis as described in Figure 2C. The actual results are shown in “A”.
Numbers above the gel images indicate individual animals, and the calculated percentages of donor
cells are indicated below the PCR images. The graph in “B” summarizes the contribution of p18-/- or
p18-/-p21-/- donor cells to different lineages in tertiary recipients (6 months after transplantation). Open
and filled bars indicate p18-/-and p18-/-p21-/- transplant groups respectively. “*” indicates p < 0.05 (n=3,
student’s t test).
Figure 4. In vitro growth of LKS cells. CD34-LKS cells were isolated from the recipients of p21-/-
primary cBMT (n=3) or quaternary p18-/-p21-/- cBMT (n=4). 100 CD34-LKS cells were sorted into 96-
well plates and cultured in the Iscove’s MDM supplemented with 100ng/ml SCF, 50ng/ml Flt-3 and
25ng/ml TPO. Cell numbers were counted at 3 time points during the culture. The growth curves
were plotted based on the Mean ± SD of triplicate cultures. An aliquot of the differentiated cells were
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom
21
lysed for the semi-quantitative PCR analysis as described in Figure 2. The inserts in the graph are
the PCR results showing the altered genotypic representation (knockout vs. wild-type). Based on the
single cell analysis (Table 1), the initial ratios of p21-/- and p18-/-p21-/- in the starting CD34-LKS cells
were 63.3 % and 78.9% respectively. No differentiation block was observed in both groups based on
the cell morphology under a microscope.
Table 1. Representation of Knockout (KO) in Stem/Progenitor Cell Pools
Clonal analysis of stem/progenitor cells cBMT*
Months post-cBMT
Clonal culture
Mice analyzed
Colonies assayed KO%
Absolute yield of KO CD34-LKS
cells§ IIº 12 CD34-LKS 3 109 92.7 ND$ p18-/-# IIIº 8 CD34-LKS 3 93 76.3 3633 ± 356
CD34-LKS 3 106 99.1 IIº 16 CFC 3 88 97.7 CD34-LKS 2 91 90.1 IIIº 6 CFC 2 88 87.6 CD34-LKS 4 76 78.9
p18-/-
p21-/-
IVº 10 CFC 4 37 40.5
ND$
CD34-LKS 3 60 63.3 p21-/-¥ Iº 8 CFC 3 67 18.9
1452 ± 394
* The cBMT procedure was detailed in our previous publication16 and is further illustrated in Figure 2A. Briefly, test cells (2 x 106 BMNCs/host) from p18-/- or p18-/-p21-/- male mice and an equal number of wild-type BMNCs (competitor cells) from male mice (wild-type litter mates in primary cBMT and then wild-type C57BL/6129/SvF1 mice of 2 months old in subsequent rounds of cBMT) were co-transplanted into lethally-irradiated (10 Gy) female recipients. Overall engraftment levels in blood are presented in Figure 2C-D.
# The representations of p18-/- genotype after after IIº cBMT have been reported in our previous publication.16 All the mice after IVo cBMT in p18-/- group developed T-cell leukemia. Characterization of the leukemic cells and relation to HSC regeneration involving a range of analyses are to be reported in a different manuscript (Yuan Y, et al., submitted).
¥ This experiment was done much later than other two experiments (p18-/ and p18-/-p21-/-) to confirm the accelerated hematopoietic exhaustion due to p21 absence10 as rationalized in the legend of Figure 2D. The competitor cells in all these experiments were from the same type of unmanipulated bone marrow.
§ Absolute yield of KO CD34-LSK cells was calculated according to the bone marrow cellularity per harvest (including ilia, femurs and tibias, n=8), the frequency (by Flow Cytometry) of viable CD34-LKS (being able to respond to growth stimuli in vitro) and KO% in CD34-LKS cells. p<0.01 (t-test, between p18-/- and p21-/- cBMT groups). The lower yield of CD34-LKS cells due to the absence of p21 must have been substantially underestimated since the p18-/- group had undergone two additional rounds of cBMT.
$ ND: not done.
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom
22
Figure 1A,B
A
p18-/- orp18+/+(M)
2 X 106
0(2)
I0
(F)
2 X 106
II0
3(5)
2 X 106
III0
(F) 4(9)
(F)
2 X 106
3(12) (F)
IV0
12(24)
Endpoint
Yu H et al
B
0
0.5
1
1.5
2
0
0.5
1
1.5*
* * *
0 o II o I o II o III o
Figure 1C Yu H et al
C
p18+/+SryMyo
Individual mouse 1 2 3 4 5 6
Engraftment(%) 0 66 60 0 0 0
p18-/-SryMyo
Individual mouse 1 2 3 4 5 6 7
Engraftment(%) 65 73 60 66 59 71 71
SryMyo
Cell ratio
0/10
0
10/9
0
25/7
5
50/5
0
75/2
5
90/1
0
100/
0
0
0.4
0.8
1.2
0 0.3 0.6 0.9
y=1.28x-0.14
Mal
e in
the
mix
ed
Popu
latio
m(%
)
Sry/Myo relative intensity
Standardization
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom
23
Figure 2AYu H et al
A
2 X 106
2 X 106
0(6)
I0
II0
III02 X 106
2 X 106
2 X 106
2 X 106
2 X 106
2 X 106
1:1
1:1
1:1
1:1
IV0
p18-/- orp18-/-p21-/-
(M)
WT(M)
WT
WT
WT
WT
WT
WT
WT
(M)
(M)
(M)
(F)
(F)
(F)
(F)10
14
8 10
(16)
(30)
(38) (48)
Endpoint
Figure 2B
B
88.50
54.8851.08
34.11
0
20
40
60
80
100
IIO IIIO
Perc
enta
ge in
Tot
al
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom
24
Figure 2C Yu H et al
C
p18-/-p18+/+
Individual mouse 1 2 3 4 5 6 7 8
Engraftment(%) 58 86 57 49 38 57 39 54p18-/-
Individual mouse 1 2 3 4 5 6 7 8p18+/+p18-/-
Engraftment(%) 17 36 62 40 13 23 41 39
Cell ratio
0/10
0
10/9
0
25/7
5
50/5
0
75/2
5
90/1
0
100/
0
p18-/-p21-/-
p18+/+
p18-/- 0
0.4
0.8
1.2
0 0.4 0.8 1.2
y=1.00+0.07
Relative p18-/- signal in totalp18-
/-in
the
mix
ed
Popu
latio
n(%
)
Standardization
Figure 2DYu H et al
D
Individual mouse 1 2 3p21+/+p21-/-
Engraftment(%) 1 0 3p21-/-
1/99
2/98
5/95
10/9
0
Cell ratiop21+/+
P21-/-Standardization
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom
25
Figure 4Yu H et al
p18+/+
p18-/- (83%)
p21+/+
p21-/- (25%)
Days in culture
Cel
l num
ber (
X104
)
0.01
2
4
6
Figure 3 Yu H et al
AIndividual mouse 1 2 3 1 2 3 1 2 3
p18+/+
P18-/-
Engraftment(%) 100 85 77 76 62 64 69 53 47
G/M T B
Individual mouse 1 2 3 1 2 3 1 2 3p18+/+
p18-/-
Engraftment(%) 90 73 74 35 37 26 76 63 49
G/M T B
p18-/-
p18-/-p21-/-
0
20
40
60
80
100
*
G/M T B
Engr
aftm
ent(
%)B
For personal use only. by guest on June 2, 2013. bloodjournal.hematologylibrary.orgFrom