hays et al., 2013
TRANSCRIPT

lable at ScienceDirect
Quaternary Science Reviews 82 (2013) 78e92
Contents lists avai
Quaternary Science Reviews
journal homepage: www.elsevier .com/locate/quascirev
Biological and climatic consequences of a cold, stratified, high latitudeocean
James D. Hays a,*, Douglas G. Martinson b, Joseph J. Morley c
aColumbia University, Lamont Doherty Earth Observatory, P.O. Box 1000 Palisades, New York 10964, USAb Lamont Doherty Earth Observatory, Dept of Earth and Environmental Sciences, Columbia University, USAc Lamont Doherty Earth Observatory, USA
a r t i c l e i n f o
Article history:Received 16 August 2012Received in revised form16 September 2013Accepted 24 September 2013Available online
Keywords:Biological pumpRadiolariaNorth PacificDeep sea coresOcean carbon sequestrationFluxGlacial oceanStratificationPleistoceneHoloceneSea of Okhotsk
* Corresponding author. Tel.: þ1 845 351 2451.E-mail address: [email protected] (J.D. H
0277-3791/$ e see front matter � 2013 Elsevier Ltd.http://dx.doi.org/10.1016/j.quascirev.2013.09.022
a b s t r a c t
The flux from deep- and shallow-living radiolarian assemblages provides evidence of a glacial, highlatitude, cold ocean stratification that increased biological pump efficiency and promoted ocean carbonsequestration. Greater deep (>200 m) than shallow-living (<200 m) radiolarian assemblage flux char-acterizes glacial North Pacific (>45� N) sediments with the deep-living Cycladophora davisiana dominant(>24%). By contrast modern radiolarian flux consists primarily of shallow-living species (C. davisiana<10%). Clues to the cause of this unusual glacial radiolarian flux come from the presently, stronglystratified Sea of Okhotsk. Here beneath a thin nutrient depleted mixed layer radiolarian and zooplanktonfaunas conform to the sea’s physical stratification with lower concentrations of both in a Cold (�1.5 to1 �C) Intermediate Layer (CIL) (20e125 m) and higher concentrations in waters between 200 and 500 m(Nimmergut and Abelmann, 2002). This biological stratification generates a radiolarian flux echoing thatof the glacial northwest Pacific with C. davisiana 26% of total flux. Widespread C. davisiana percentages(>20%) in high latitude (>45�) glacial sediments of both hemispheres is evidence that these oceans werecapped with an Okhotsk-Like Stratification (O-LS). O-LS provides mechanisms to (1) strip nutrients fromsurface waters depriving the deep-ocean of preformed nutrients, increasing biological pump efficiencyand (2) deepen carbon re-mineralization increasing deep-ocean alkalinity. Both may have contributed tolower glacial atmospheric CO2 concentrations. O-LS would also have amplified glacial climatic cycles bypromoting the spread of high latitude sea ice in winter as occurs in the Sea of Okhotsk today, andreducing gas exchange between ocean and atmosphere in summer.
� 2013 Elsevier Ltd. All rights reserved.
1. Introduction
Positive correlations between temperature and CO2 concentra-tion in ice cores have led to a consensus that atmospheric CO2variations amplified ice age climate change (Berner et al., 1979;Petit et al., 1999) but the cause of these variations are poorly un-derstood (Sigman et al., 2010). The partial pressure of atmosphericCO2 (pCO2) is controlled by the average steady state of ocean sur-face water pCO2 weighted by area and gas exchange kinetics(Archer et al., 2000), which in turn varies with temperature andmajor nutrient concentrations. Much of the present-day ocean’smixed layer is stripped of major nutrients by an efficient biologicalpump; however, large areas of the Antarctic, equatorial Pacific andNorth Pacific are not. Increased glacial biological pump efficiency inthese areas could cause more complete nutrient utilization and
ays).
All rights reserved.
consequent atmospheric CO2 drawdown. The Antarctic Ocean isespecially important, for today its unused nutrients are convectedto the deep-ocean as so called preformed nutrients. Thus atmo-spheric pCO2 depends on both surface ocean nutrient and deep-ocean preformed nutrient concentrations. Deeper glacial carbonre-mineralization could also have lowered atmospheric pCO2 bytriggering deep-ocean alkalinity changes (Broecker and Peng, 1987;Boyle, 1988).
Increased glacial ocean carbon sequestration is suggested bylower glacial than Holocene deep-ocean oxygen levels (Thompsonet al., 1990; Francois et al., 1997; Gebhardt et al., 2008; Jaccard et al.,2009) and more depleted glacial d13C below 2000 m than above(Herguera, 1992; Keigwin, 1998; Curry and Oppo, 2005). Proxies forproductivity and export, e.g. opal and barium flux, to subarcticNorth Pacific sediments do not support increased glacial relative toHolocene primary productivity (Narita et al., 2002; Kienast et al.,2004; Haug et al., 2005; Jaccard et al., 2005; Shigemitsu et al.,2007). So lower North Pacific diatom d15N (increased surface wa-ter nitrogen utilization) values in glacial relative to interglacial

Fig. 1. Recent distribution of northern he misphere permafrost (Washburn, 1980 and references therein) and percent C. davisiana in ocean floor sediments (Morley and Hays, 1983;Morley, 1983; and Pisias personal communication). Holocene high percentages of this species occur only within the Sea of Okhotsk and adjacent Pacific.
J.D. Hays et al. / Quaternary Science Reviews 82 (2013) 78e92 79
sediments, have been attributed to lower upward nutrient fluxcaused by increased stratification, rather than higher primaryproductivity (Sigman et al., 1999; Galbraith et al., 2008). Glacialstratification has also been invoked in the Antarctic (Francois et al.,1997), however the biological consequences of this stratificationare poorly understood in either the Antarctic or North Pacific.
Although organic carbon export is often linked to primaryproductivity (Eppley and Peterson, 1979), it can also vary with littleor no primary production change (Boyd and Newton,1995) becausethe quantity of sinking organic carbon is strongly affected by con-sumer community structure (Michaels and Silver, 1988) that can beinfluenced by physical water column properties (Hargrave, 1975;Gardner et al., 1993). Glacial Ocean cooling has been suggested asa water column change that could reduce heterotrophic con-sumption rates and thereby increase biological pump efficiencyrelative to today (Matsumoto, 2007).
The remains of shallow (<200 m) and deep-living (>200 m)organisms can provide information about past depths of organiccarbon respiration but fewmembers of the deep-living communityleave a fossil record. Of those that do, polycystine radiolarians arethe most abundant and diverse group (Kling, 1979; Gowing, 1986;
Takahashi, 1991; Abelmann and Gowing, 1997; Itaki, 2003;Okazaki et al., 2004; Abelmann and Nimmergut, 2005; Tanakaand Takahashi, 2005). Radiolarians, as trophic generalists, feedingon sinking organic detritus and associated microorganismsincluding bacteria (Anderson, 1983; Gowing, 1986), respond tochanging organic flux. Because their shells suffer less from disso-lution than diatoms (Shemesh et al., 1989; Morley et al., in press)fossil radiolarians should record production changes at differentlevels within the water column.
Shallow-living species (0e200 m) dominate radiolarianscaptured by plankton tows (Kling and Boltovskoy,1995; Tanaka andTakahashi, 2008) but a deep-living species, Cycladophora davisiana,dominates Last Glacial Maximum (LGM) high latitude (>45�)radiolarian faunas of both hemispheres (Hays et al., 1976; Morleyet al., 1982; Morley, 1983; Gersonde et al., 2003). In Holocenesediments only those of the Sea of Okhotsk approach the highC. davisiana percentages found in high latitude glacial ocean sedi-ments (Fig. 1). In the Okhotsk water column radiolarian concen-trations below 200 m exceed those above, reaching a maximumbetween 200 and 500 m, where C. davisiana dominates(Nimmergut and Abelmann, 2002; Okazaki et al., 2004). This

J.D. Hays et al. / Quaternary Science Reviews 82 (2013) 78e9280
radiolarian concentration profile is well documented, but its causeremains controversial. Some attribute high C. davisiana productiv-ity between 200 and 500 m to organic matter advected fromneighboring shelves (Okazaki et al., 2003a) or transported by seaice (Okazaki et al., 2003b). Others appeal to favorable intermediatewater properties to enhance production and a possible influencefrom overlying stratification (Abelmann and Nimmergut, 2005).Deep-living radiolarians (>200 m) may also benefit from enhancedcarbon export resulting from reduced respiration in the cold watersbetween 20 and 125 m (Hays and Morley, 2004).
Views also diverge on the cause of high C. davisiana percentages(>20%) in high-latitude glacial sediments. A popular proposal isthat they are a response to well-ventilated IntermediateWater flow(Nimmergut and Abelmann, 2002; Ohkushi et al., 2003; Abelmannand Nimmergut, 2005; Matul, 2011). Alternatively it has beenargued they are a product of a cold, highly stratified water column,similar to that of the modern Sea of Okhotsk (Morley and Hays,1983; Hays and Morley, 2004). These proposals are testablebecause a glacial North Pacific water column, similar to that of themodern Sea of Okhotsk, would have produced lower shallow-livingflux than its Holocene counterpart while C. davisiana production,promoted solely by increased intermediate water flow, should nothave influenced shallow-living radiolarian production. This paperuses a set of radiolarian species that today live dominantly eitherabove or below 200 m to test these hypotheses. It also documentsfor the first time glacial-interglacial changes in the flux of shallow-and deep-living radiolarian assemblages and uses this informationto explore the biological and climatic consequences of glacial NorthPacific stratification.
Table 1
2. Selection of material
Three northwest Pacific cores are selected that have nearlyconstant late Pleistocene sedimentation rates (Ninkovich andRobertson, 1975; Keigwin, 1995) and those rates are sufficientlyhigh to assure good opal preservation (Broecker and Peng, 1982).They include a hydraulic piston core, raised from atop DetroitSeamount (ODP site 883D) and two piston cores from the PacificOcean floor (V20-122, V20-124). Holocene sediments from a Sea of
60°N
55°N
50°N
45°N
40°N
140°E 150°E 160°E
Sea of
Japa
n NorthPacific
V20-122
883D
Sea ofOkhotsk
V20-124
V32-161
Fig. 2. Location of four sediment cores used in this study, Sea of Okhotsk V32-161,48.238�N, 149.066�E, 1600 m; deep northwest Pacific cores V20-122, 46.566�N,161.683�E, 5563 m; V20-124, 45.833�N, 154.5�E, 5534 m; and Detroit Sea Mount 883D,51.198�N, 167.678�E, 2396 m.
Okhotsk core (V32-161) are also included (Fig. 2). No evidence ofdissolution was observed except in Pleistocene sediments of theSea of Okhotsk core. The three Pacific cores are sampled from latePleistocene (about 70 ka BP) through Holocene but only the Holo-cene of the Sea of Okhotsk core (V32-161) because of poor Pleis-tocene radiolarian preservation (Morley et al., 1991). Speciesabundances, the degree to which species are restricted to depthsabove or below 200 m and their occurrence in both Sea of Okhotskand northwest Pacific sediments, guided the selection of six speciesthat are grouped into Shallow (<200 m) and Deep (>200 m) As-semblages. The morphological variations of these species areillustrated in references listed in Table 1. All other species are placedin an Other Assemblage.
The Shallow Assemblage consists of Spongotrochus glacialis(Popofsky), Stylochlamydium venustum (Bailey) and their probablejuveniles (Spongodiscidae spp). Stratified plankton tow studiesshow that these species live dominantly above 50 m in the Sea ofOkhotsk (Nimmergut and Abelmann, 2002) and above 100m in theAntarctic Ocean (Abelmann and Gowing, 1997). They compose>75% of all radiolarians separated from Recent Bering Sea surfacesediments, raised from<150 m (Blueford, 1983; Wang et al., 2006).Blueford (1983) refers to these species as “groups” indicatingconsiderable morphological variation, we also recognize widemorphological variation (Table 1). In stratified plankton towsS. venustum is found mostly above 100 m and S. glacialis mostlyabove 250 m in the North Pacific (Kling, 1979; Tanaka andTakahashi, 2008) and other high-latitude seas including the ArcticOcean (Hülsemann, 1963; Tibbs, 1967; Kling, 1979; Morley andStepien, 1985; Abelmann and Gowing, 1997; Itaki et al., 2003) butcan occur in lower numbers down to 3000 m (Itaki et al., 2003;Tanaka and Takahashi, 2008). S. venustum is the most abundantradiolarian in Bering Sea and northwest Pacific Holocene sedimentsnorth of 45� (Ling et al., 1971; Robertson, 1975; Wang et al., 2006).Tanaka and Takahashi (2005) showed that its flux to Holocenesediments is greater than its flux to glacial sediments in both theBering Sea and northwest Pacific. Both S. venustum and S. glacialisare residents of polar and sub-polar waters of both hemispheres
This table presents a list of references that together illustrates the morphologicalvariability of the species included in this study. S. venustum, S. glacialis and theirprobable juveniles the Spongodiscidae spp. dominate the Shallow Assemblage andare well illustrated in numerous studies of North Pacific and Antarctic radiolarians.Similarly C. davisiana and D. hirundo dominate the Deep Assemblage and are like-wise widely illustrated. A. micropora and Spongurus sp. are distinctive but much lessabundant and if omitted would not change the conclusions of this paper.
Acanthodesmia micropora (Popofsky), Kruglikova, 1975, p. 84, Fig. 2, (19);Nimmergut and Abelmann, 2002, p. 470, pl. 2, (7); Hays and Morley, 2004, p.601, Fig. 7, (c).
Cycladophora davisiana Ehrenberg, Petrushevskaya, 1967, p. 122, Fig. 69, (IeVII);Tanaka and Takahashi, 2008, p. 69, pl. 3, (7, 8); Hays andMorley, 2004, p. 601,Fig. 7, (g, h, i); Nimmergut and Abelmann, 2002, p. 469, pl. 1, (9, 10, 11).
Dictyophimus hirundo (Haeckel), Petrushevskaya, 1967, p. 114, Fig. 67, (IeV);Tanaka and Takahashi, 2008, p. 71, pl. 5 (5, 6, 7); Ling et al., 1971, p. 729, pl. 2,(8, 9, 10); Nimmergut and Abelmann, 2002, p. 471, pl. 2 (5a, 5b).
Spongodiscidae spp., Tanaka and Takahashi, 2008, p. 68, pl. 2, (5, 6, 7); Okazakiet al., 2003, p. 204, pl. I (18, 20).
Spongotrochus glacialis (Popofsky), Nigrini and Moore, 1979, p. S117, pl. 15, (2);Tanaka and Takahashi, 2008, p. 68, pl. 2 (1, 2); Okazaki et al., 2003b, p.204,plate 1, (21).
Spongurus (?) sp. Petrushevskaya, 1968, p. 49, Fig. 26, (I); Nigrini and Moore,1979, p. S67, pl. 8, (4); Tanaka and Takahashi, 2008, p. 69, pl. 3 (3); Okazakiet al., 2003b, p. 204, pl. 1 (23); Ling et al., 1971, p. 727, pl. 1 (6).
Stylochlamydium venustum (Bailey), Boltovskoy and Riedel, 1980, p. 141, pl. 4,(3); Kruglikova, 1975, p. 84, Fig. 2, (1, 2); Tanaka and Takahashi, 2008, p. 68,pl. 2 (3);
Ling et al., 1971, p.727, pl. 1 (7, 8); Nimmergut and Abelmann, 2002, p. 469,pl. 1 (1).
J.D. Hays et al. / Quaternary Science Reviews 82 (2013) 78e92 81
(Kling, 1979; Morley and Stepien, 1985; Abelmann and Gowing,1997; Tanaka and Takahashi, 2008) but similar forms have beenreported from low latitudes (Renz, 1976; McMillen and Casey,1978).
The Deep Assemblage consists of four species that stratifiedplankton tow collections indicate have depth preferences below200 m. The cosmopolitan C. davisiana Ehrenberg, is by far the mostabundant member of this assemblage in glacial and Recent sedi-ments. It has been found to be abundant below 150 m and mostabundant between 200 and 500 m in Sea of Okhotsk stratifiedplankton tows (Nimmergut and Abelmann, 2002; Okazaki et al.,2004). Globally it lives primarily below 400 m (Abelmann andGowing, 1997; Itaki, 2003; Itaki et al., 2003). Although some re-searchers recognize several subspecies of C. davisiana, we havechosen for this study to include all those described byPetrushevskaya (1967, Fig. 69) as C. davisiana and these have beenshown to be dominantly deep dwellers (>200 m) in the Sea ofOkhotsk (Nimmergut and Abelmann, 2002; Abelmann andNimmergut, 2005). Dictyophimus hirundo (Haeckel) is cosmopol-itan (Boltovskoy and Riedel, 1980) and most abundant in Okhotsk’swater column below 400m (Nimmergut and Abelmann, 2002), andbelow 250 m in the subpolar North Pacific (Tanaka and Takahashi,2008). Acanthodesmia micropora (Popofsky) was caught in deep1061 m but rarely in shallow 258 m sediment traps in the centralSea of Okhotsk (Hays and Morley, 2004). Spongurus (?) sp. Pet-rushevskaya, a cosmopolitan species, occurs primarily below 400min Antarctic waters (Abelmann and Gowing, 1996) and predomi-nantly between 1000 and 3000 m in the North Pacific (Tanaka andTakahashi, 2008). It has not been reported from the Sea of Okhotskbut is included here because of its deep-living Antarctic and NorthPacific habitats.
0
20
40
60
80
100
120
140
160
0 100 200 300 400 500 600 700 800
V20-122 883D V20-124
Depth in cm.
Age
x 10
3 yea
rs
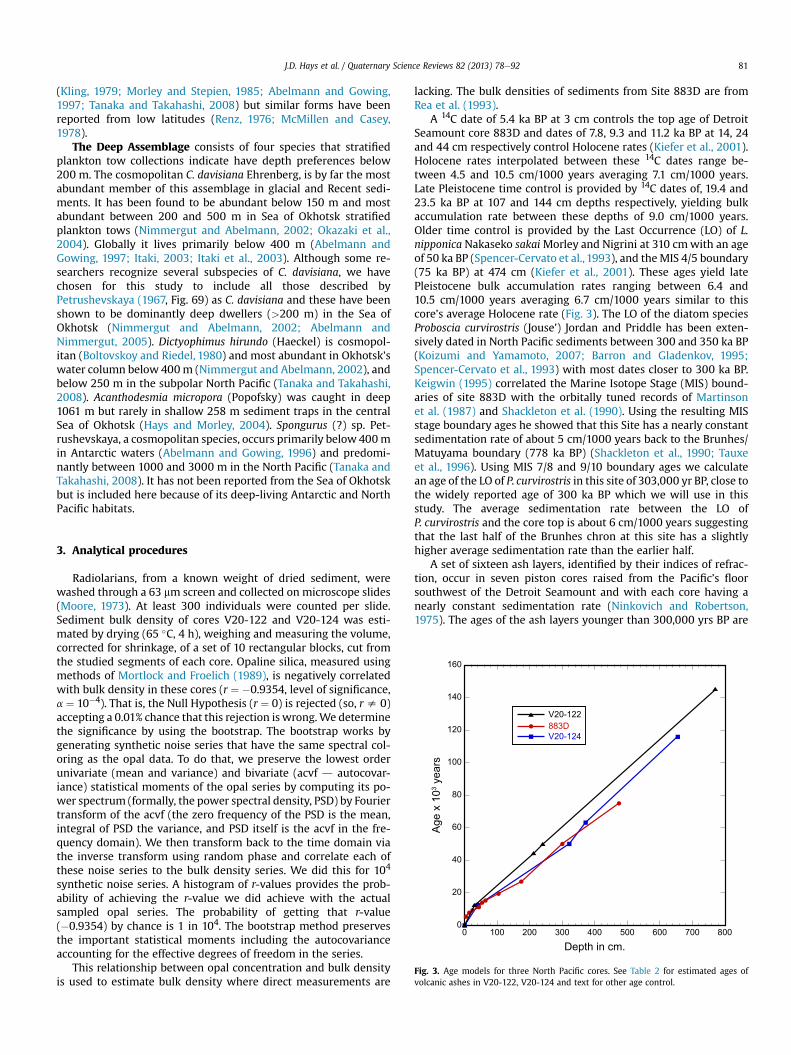
Fig. 3. Age models for three North Pacific cores. See Table 2 for estimated ages ofvolcanic ashes in V20-122, V20-124 and text for other age control.
3. Analytical procedures
Radiolarians, from a known weight of dried sediment, werewashed through a 63 mm screen and collected on microscope slides(Moore, 1973). At least 300 individuals were counted per slide.Sediment bulk density of cores V20-122 and V20-124 was esti-mated by drying (65 �C, 4 h), weighing and measuring the volume,corrected for shrinkage, of a set of 10 rectangular blocks, cut fromthe studied segments of each core. Opaline silica, measured usingmethods of Mortlock and Froelich (1989), is negatively correlatedwith bulk density in these cores (r ¼ �0.9354, level of significance,a ¼ 10�4). That is, the Null Hypothesis (r ¼ 0) is rejected (so, r s 0)accepting a 0.01% chance that this rejection is wrong.We determinethe significance by using the bootstrap. The bootstrap works bygenerating synthetic noise series that have the same spectral col-oring as the opal data. To do that, we preserve the lowest orderunivariate (mean and variance) and bivariate (acvf d autocovar-iance) statistical moments of the opal series by computing its po-wer spectrum (formally, the power spectral density, PSD) by Fouriertransform of the acvf (the zero frequency of the PSD is the mean,integral of PSD the variance, and PSD itself is the acvf in the fre-quency domain). We then transform back to the time domain viathe inverse transform using random phase and correlate each ofthese noise series to the bulk density series. We did this for 104
synthetic noise series. A histogram of r-values provides the prob-ability of achieving the r-value we did achieve with the actualsampled opal series. The probability of getting that r-value(�0.9354) by chance is 1 in 104. The bootstrap method preservesthe important statistical moments including the autocovarianceaccounting for the effective degrees of freedom in the series.
This relationship between opal concentration and bulk densityis used to estimate bulk density where direct measurements are
lacking. The bulk densities of sediments from Site 883D are fromRea et al. (1993).
A 14C date of 5.4 ka BP at 3 cm controls the top age of DetroitSeamount core 883D and dates of 7.8, 9.3 and 11.2 ka BP at 14, 24and 44 cm respectively control Holocene rates (Kiefer et al., 2001).Holocene rates interpolated between these 14C dates range be-tween 4.5 and 10.5 cm/1000 years averaging 7.1 cm/1000 years.Late Pleistocene time control is provided by 14C dates of, 19.4 and23.5 ka BP at 107 and 144 cm depths respectively, yielding bulkaccumulation rate between these depths of 9.0 cm/1000 years.Older time control is provided by the Last Occurrence (LO) of L.nipponica Nakaseko sakaiMorley and Nigrini at 310 cmwith an ageof 50 ka BP (Spencer-Cervato et al., 1993), and theMIS 4/5 boundary(75 ka BP) at 474 cm (Kiefer et al., 2001). These ages yield latePleistocene bulk accumulation rates ranging between 6.4 and10.5 cm/1000 years averaging 6.7 cm/1000 years similar to thiscore’s average Holocene rate (Fig. 3). The LO of the diatom speciesProboscia curvirostris (Jouse’) Jordan and Priddle has been exten-sively dated in North Pacific sediments between 300 and 350 ka BP(Koizumi and Yamamoto, 2007; Barron and Gladenkov, 1995;Spencer-Cervato et al., 1993) with most dates closer to 300 ka BP.Keigwin (1995) correlated the Marine Isotope Stage (MIS) bound-aries of site 883D with the orbitally tuned records of Martinsonet al. (1987) and Shackleton et al. (1990). Using the resulting MISstage boundary ages he showed that this Site has a nearly constantsedimentation rate of about 5 cm/1000 years back to the Brunhes/Matuyama boundary (778 ka BP) (Shackleton et al., 1990; Tauxeet al., 1996). Using MIS 7/8 and 9/10 boundary ages we calculatean age of the LO of P. curvirostris in this site of 303,000 yr BP, close tothe widely reported age of 300 ka BP which we will use in thisstudy. The average sedimentation rate between the LO ofP. curvirostris and the core top is about 6 cm/1000 years suggestingthat the last half of the Brunhes chron at this site has a slightlyhigher average sedimentation rate than the earlier half.
A set of sixteen ash layers, identified by their indices of refrac-tion, occur in seven piston cores raised from the Pacific’s floorsouthwest of the Detroit Seamount and with each core having anearly constant sedimentation rate (Ninkovich and Robertson,1975). The ages of the ash layers younger than 300,000 yrs BP are

J.D. Hays et al. / Quaternary Science Reviews 82 (2013) 78e9282
determined by interpolation between the core tops and the LO of P.curvirostris. For cores that do not reach the LO of P. curvirostris weuse the interpolated age of the oldest ash layer for control.
Two cores from this set (V20-122 and V20-124), that containfour of the 16 ash layers, are chosen to supplement data from core883D. The similarity of the interpolated ages of each of these ashlayers in the selected cores (bold faced type) and five nearby cores(Table 2) can result only if these cores have nearly constant sedi-mentation rates. The mean ages of the four ash layers (Table 2)together with the LO of L. nipponica sakaii (50 ka BP) (V20-122,240 cm; V20-124, 320 cm) are used to control their Pleistocenesedimentation rates. This age control generates Pleistocene bulksedimentation rates ranging between 5.0 and 6.2 cm/1000 yrs,averaging 5.5 cm/1000 yrs in V20-122 and between 3.8 and 7.5 cm/1000 yrs averaging 5.9 cm/1000 yrs in V20-124 (Fig. 3).
The depth to the major deglacial Assemblage’ changes in thetrigger cores of V20-122 and V20-124 is no greater than in theirrespective piston cores suggesting that little sediment is missingfrom the piston core tops but it is unlikely that these tops have zeroage. Therefore Holocene Assemblage flux is calculated using zerocore top ages and the midpoints of the Holocene/Pleistocenetransitions of C. davisiana percentages (12.5 ka BP) (Fig. 5a and b)and by extrapolating late Pleistocene rates to the core tops. Theformer method yields Holocene bulk sedimentation rates of 2.4(V20-122) and 3.2 cm/1000 yrs (V20-124) the latter 5.5 and 5.9 cm/1000 yrs respectively, similar to the Holocene rates of core 883D.These different estimated Holocene bulk accumulation rates alterthe magnitude but not the direction of Holocene/PleistoceneShallow and Deep Assemblage flux changes in both cores.
3.1. Analysis of flux errors
The flux (F) is a nonlinear multivariate product of 3 randomvariables (Xk, variables whose exact value we do not know):
FðXkÞ ¼ FðX1;X2;X3Þ ¼ X1X2X3 (1)
where X1 ¼ rads./gm.; X2 ¼ bulk density; X3 ¼ sed. rate.The uncertainty in the individual variables is computed as
follows:
Table 2Depth to and estimated ages of, four ash layers (AL) in seven northwest Pacific cores.Cores used in this study are in boldfaced type. Ash layer ages are determined bylinear interpolation, using core tops (zero age) and last occurrence (LO) of thediatom P. curvirostris at 300,000 yrs. BP (Barron and Gladenkov, 1995; Koitzumi andYamamoto, 2007) for time control. For cores that do not reach the LO of P. curvirostristhe interpolated age of the oldest ash layer is used instead. The similarity of indi-vidual ash layer ages in multiple cores is evidence these cores have nearly constantbulk accumulation rates.
Ash layer(AL)
Core Depth to ALbase (cm.)
Estimatedage ka.
Mean age
1 V20-122 212 44.5 44 � 1.3V20-123 215 42.7RC14-106
204 45.3
2 V20-124 372 63.6 63 � 0.6RC14-106
287 63.8
V20-123 315 62.65 V20-124 653 111.7 116 � 4
RC14-106
539 119.8
8 V20-119 119 132 141 � 9V20-120 490 136V20-121 520 137V20-122 770 145V20-123 844 150
3.1.1. Radiolarian counts (X1)Errors associated with how representative a single sample of
sediment is, as well as counting errors. Three counts were made oneach half of a split sample giving 6 estimates that are averaged togive a single estimate of uncertainty.
3.1.2. Bulk density (X2)This variable is estimated by regressing grams of sediment per
cm3 (bulk density) against grams of opal per grams of sediment(concentration) of ten samples. For this it is not the error in theslope we use, but rather the error in predicting the bulk densityfrom opal concentration. This error is a function of opal concen-tration, so we estimate a single “representative” error in this pre-diction by using the error associated with an opal concentrationthat is two thirds of the way from the mean concentration to thehighest. The highest opal concentration is large, so this estimate isconservative.
3.1.3. Sedimentation rate (X3)Here the deptheage plot of the ash layers in cores V20-122, V20-
124 and the 14C dates of 883D is fit with a least squares straight line(consistent with the assumption of a constant sedimentation rateas justified above). The uncertainty in the slope (the sedimentationrate) is estimated from calculating the standard error of the slope(Draper and Smith, 1988).
Because of the nonlinearity in F, we make a first order linearapproximation to its uncertainty (variance) by truncating a Taylorseries expansion to first order. Applying the expectance operator, inthe form of variance, gives:
s2FzX3
i¼1
X3
j¼1
vFðXiÞvXi
����mi
vF�Xj�
vXj
�����mj
Cov�XiXj
�(2)
We assume that the 3 random variables are independent (thenumber of radiolarians is so small relative to the other sedimentarycomponents (diatoms, clay and volcanic ash) that any relationshipbetween X1 and the other variables is considered trivial). Thus thecovariance term (Cov[]) in (2) is 0 whenever, i s j; it only surviveswhen i ¼ j, reducing covariance to variance giving:
s2FzX3
i¼1
vFðXiÞvXi
����mi
vFðXiÞvXi
����mi
Var½Xi� (3)
or, expanded:
s2Fz½X2ðmÞX3ðmÞ�2Var½X1� þ ½X1ðmÞX3ðmÞ�2Var½X2�þ ½X1ðmÞX2ðmÞ�2Var½X3� (4)
The uncertainties of each variable (as variance) are combined, asdictated by Formula (4) to estimate the standard error for each fluxestimate yielding the mean fluxes, � their standard error (to firstorder) and percent error of the mean flux, all in units of rads/(cm2 ka) (Table 3).
4. Results
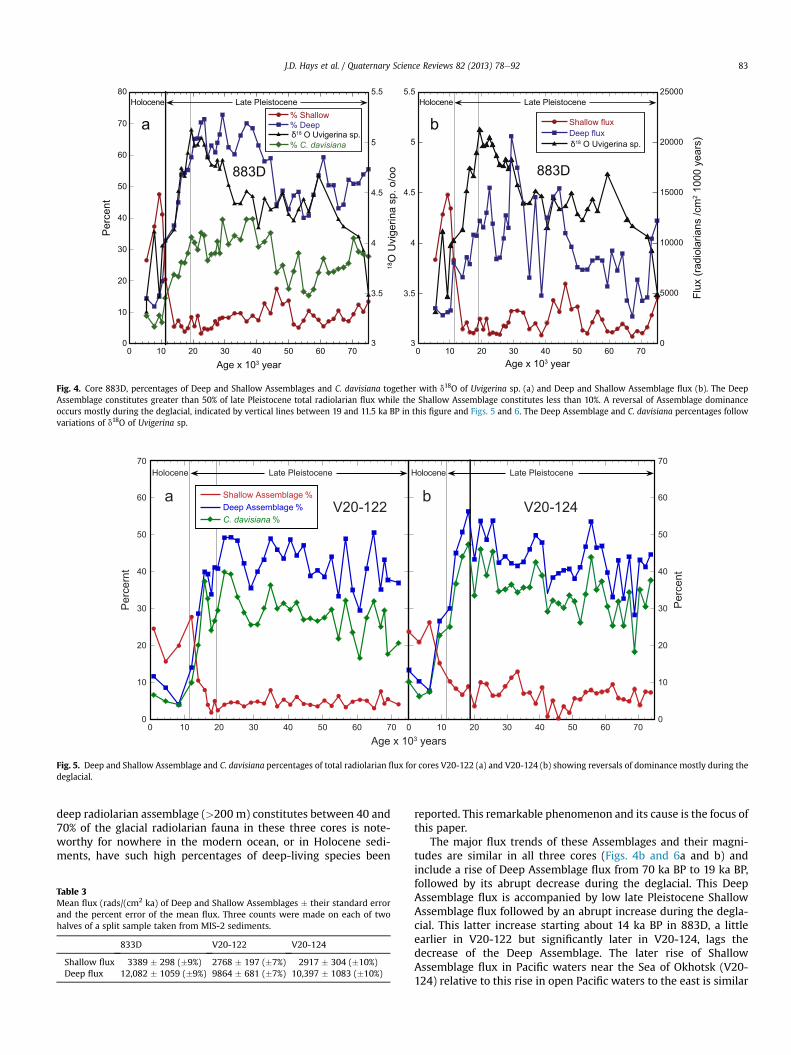
In all three northwest Pacific cores the Deep Assemblage’s per-centage of total radiolarians declines during the deglacial (19 ka BPto 11.5 ka BP) from Pleistocene values of as much as 70% to Holo-cene values of about 10%. This decline parallels declines ofC. davisiana percentages and d18O of Uvigerina sp. in core 883D(Figs. 4a and 5a and b). Nearly simultaneously the ShallowAssemblage rises from about 10% in the late Pleistocene to between20 and 50% in the Holocene (Figs. 4a and 5a and b). The fact that a
3
3.5
4
4.5
5
5.5
0
5000
10000
15000
20000
25000
883D 883D
18O
Uvi
gerin
a sp
. o/o
o
Flux
(rad
iola
rians
/cm
2 100
0 ye
ars)
0 10 20 30 40 50 60 70Age x 103 year
Holocene Late Pleistocene
b
0 10 20 30 40 50 60 70Age x 103 year
Holocene Late Pleistocene
a
3
3.5
4
4.5
5
5.5
0
10
20
30
40
50
60
70
80
% Shallow% Deep
% C. davisiana
δ O Uvigerina sp.
Perc
ent
Shallow fluxDeep fluxδ O Uvigerina sp.
Fig. 4. Core 883D, percentages of Deep and Shallow Assemblages and C. davisiana together with d18O of Uvigerina sp. (a) and Deep and Shallow Assemblage flux (b). The DeepAssemblage constitutes greater than 50% of late Pleistocene total radiolarian flux while the Shallow Assemblage constitutes less than 10%. A reversal of Assemblage dominanceoccurs mostly during the deglacial, indicated by vertical lines between 19 and 11.5 ka BP in this figure and Figs. 5 and 6. The Deep Assemblage and C. davisiana percentages followvariations of d18O of Uvigerina sp.
0
10
20
30
40
50
60
70
0
10
20
30
40
50
60
70
V20-122
Perc
ernt
Age x 103 years
Holocene Late Pleistocene
aDeep Assemblage %Shallow Assemblage %
C. davisiana %
0 10 20 30 40 50 60 70 0 10 20 30 40 50 60 70
V20-124
Perc
ent
Holocene Late Pleistocene
b
Fig. 5. Deep and Shallow Assemblage and C. davisiana percentages of total radiolarian flux for cores V20-122 (a) and V20-124 (b) showing reversals of dominance mostly during thedeglacial.
J.D. Hays et al. / Quaternary Science Reviews 82 (2013) 78e92 83
deep radiolarian assemblage (>200 m) constitutes between 40 and70% of the glacial radiolarian fauna in these three cores is note-worthy for nowhere in the modern ocean, or in Holocene sedi-ments, have such high percentages of deep-living species been
Table 3Mean flux (rads/(cm2 ka) of Deep and Shallow Assemblages � their standard errorand the percent error of the mean flux. Three counts were made on each of twohalves of a split sample taken from MIS-2 sediments.
833D V20-122 V20-124
Shallow flux 3389 � 298 (�9%) 2768 � 197 (�7%) 2917 � 304 (�10%)Deep flux 12,082 � 1059 (�9%) 9864 � 681 (�7%) 10,397 � 1083 (�10%)
reported. This remarkable phenomenon and its cause is the focus ofthis paper.
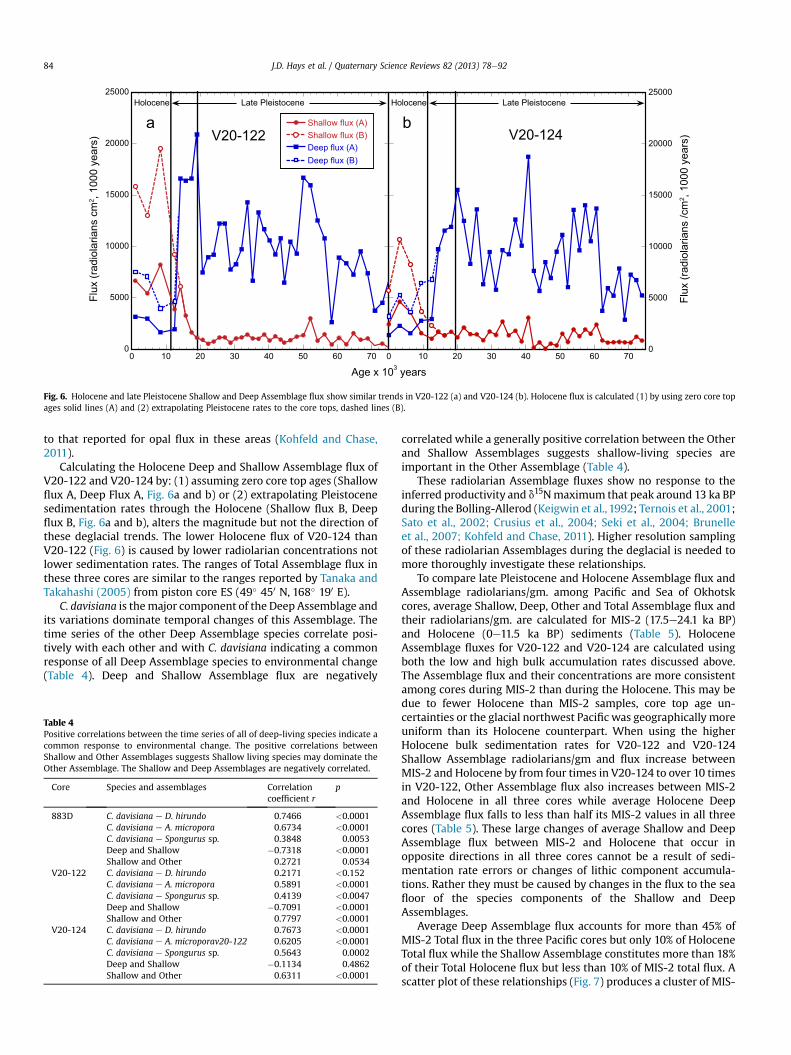
The major flux trends of these Assemblages and their magni-tudes are similar in all three cores (Figs. 4b and 6a and b) andinclude a rise of Deep Assemblage flux from 70 ka BP to 19 ka BP,followed by its abrupt decrease during the deglacial. This DeepAssemblage flux is accompanied by low late Pleistocene ShallowAssemblage flux followed by an abrupt increase during the degla-cial. This latter increase starting about 14 ka BP in 883D, a littleearlier in V20-122 but significantly later in V20-124, lags thedecrease of the Deep Assemblage. The later rise of ShallowAssemblage flux in Pacific waters near the Sea of Okhotsk (V20-124) relative to this rise in open Pacific waters to the east is similar
0
5000
10000
15000
20000
25000
0 10 20 30 40 50 60 70
Flux
(rad
iola
rians
/cm
2 , 10
00 y
ears
)V20-124
Holocene Late Pleistocene
V20-122
0
5000
10000
15000
20000
25000
0 10 20 30 40 50 60 70
Age x 103 years
a bHolocene Late Pleistocene
Flux
(rad
iola
rians
cm
2 , 10
00 y
ears
)
Shallow flux (A)Shallow flux (B)
Deep flux (B)Deep flux (A)
Fig. 6. Holocene and late Pleistocene Shallow and Deep Assemblage flux show similar trends in V20-122 (a) and V20-124 (b). Holocene flux is calculated (1) by using zero core topages solid lines (A) and (2) extrapolating Pleistocene rates to the core tops, dashed lines (B).
J.D. Hays et al. / Quaternary Science Reviews 82 (2013) 78e9284
to that reported for opal flux in these areas (Kohfeld and Chase,2011).
Calculating the Holocene Deep and Shallow Assemblage flux ofV20-122 and V20-124 by: (1) assuming zero core top ages (Shallowflux A, Deep Flux A, Fig. 6a and b) or (2) extrapolating Pleistocenesedimentation rates through the Holocene (Shallow flux B, Deepflux B, Fig. 6a and b), alters the magnitude but not the direction ofthese deglacial trends. The lower Holocene flux of V20-124 thanV20-122 (Fig. 6) is caused by lower radiolarian concentrations notlower sedimentation rates. The ranges of Total Assemblage flux inthese three cores are similar to the ranges reported by Tanaka andTakahashi (2005) from piston core ES (49� 450 N, 168� 190 E).
C. davisiana is themajor component of the Deep Assemblage andits variations dominate temporal changes of this Assemblage. Thetime series of the other Deep Assemblage species correlate posi-tively with each other and with C. davisiana indicating a commonresponse of all Deep Assemblage species to environmental change(Table 4). Deep and Shallow Assemblage flux are negatively
Table 4Positive correlations between the time series of all of deep-living species indicate acommon response to environmental change. The positive correlations betweenShallow and Other Assemblages suggests Shallow living species may dominate theOther Assemblage. The Shallow and Deep Assemblages are negatively correlated.
Core Species and assemblages Correlationcoefficient r
p
883D C. davisiana e D. hirundo 0.7466 <0.0001C. davisiana e A. micropora 0.6734 <0.0001C. davisiana e Spongurus sp. 0.3848 0.0053Deep and Shallow �0.7318 <0.0001Shallow and Other 0.2721 0.0534
V20-122 C. davisiana e D. hirundo 0.2171 <0.152C. davisiana e A. micropora 0.5891 <0.0001C. davisiana e Spongurus sp. 0.4139 <0.0047Deep and Shallow �0.7091 <0.0001Shallow and Other 0.7797 <0.0001
V20-124 C. davisiana e D. hirundo 0.7673 <0.0001C. davisiana e A. microporav20-122 0.6205 <0.0001C. davisiana e Spongurus sp. 0.5643 0.0002Deep and Shallow �0.1134 0.4862Shallow and Other 0.6311 <0.0001
correlated while a generally positive correlation between the Otherand Shallow Assemblages suggests shallow-living species areimportant in the Other Assemblage (Table 4).
These radiolarian Assemblage fluxes show no response to theinferred productivity and d15Nmaximum that peak around 13 ka BPduring the Bolling-Allerod (Keigwin et al., 1992; Ternois et al., 2001;Sato et al., 2002; Crusius et al., 2004; Seki et al., 2004; Brunelleet al., 2007; Kohfeld and Chase, 2011). Higher resolution samplingof these radiolarian Assemblages during the deglacial is needed tomore thoroughly investigate these relationships.
To compare late Pleistocene and Holocene Assemblage flux andAssemblage radiolarians/gm. among Pacific and Sea of Okhotskcores, average Shallow, Deep, Other and Total Assemblage flux andtheir radiolarians/gm. are calculated for MIS-2 (17.5e24.1 ka BP)and Holocene (0e11.5 ka BP) sediments (Table 5). HoloceneAssemblage fluxes for V20-122 and V20-124 are calculated usingboth the low and high bulk accumulation rates discussed above.The Assemblage flux and their concentrations are more consistentamong cores during MIS-2 than during the Holocene. This may bedue to fewer Holocene than MIS-2 samples, core top age un-certainties or the glacial northwest Pacific was geographically moreuniform than its Holocene counterpart. When using the higherHolocene bulk sedimentation rates for V20-122 and V20-124Shallow Assemblage radiolarians/gm and flux increase betweenMIS-2 and Holocene by from four times in V20-124 to over 10 timesin V20-122, Other Assemblage flux also increases between MIS-2and Holocene in all three cores while average Holocene DeepAssemblage flux falls to less than half its MIS-2 values in all threecores (Table 5). These large changes of average Shallow and DeepAssemblage flux between MIS-2 and Holocene that occur inopposite directions in all three cores cannot be a result of sedi-mentation rate errors or changes of lithic component accumula-tions. Rather they must be caused by changes in the flux to the seafloor of the species components of the Shallow and DeepAssemblages.
Average Deep Assemblage flux accounts for more than 45% ofMIS-2 Total flux in the three Pacific cores but only 10% of HoloceneTotal flux while the Shallow Assemblage constitutes more than 18%of their Total Holocene flux but less than 10% of MIS-2 total flux. Ascatter plot of these relationships (Fig. 7) produces a cluster of MIS-
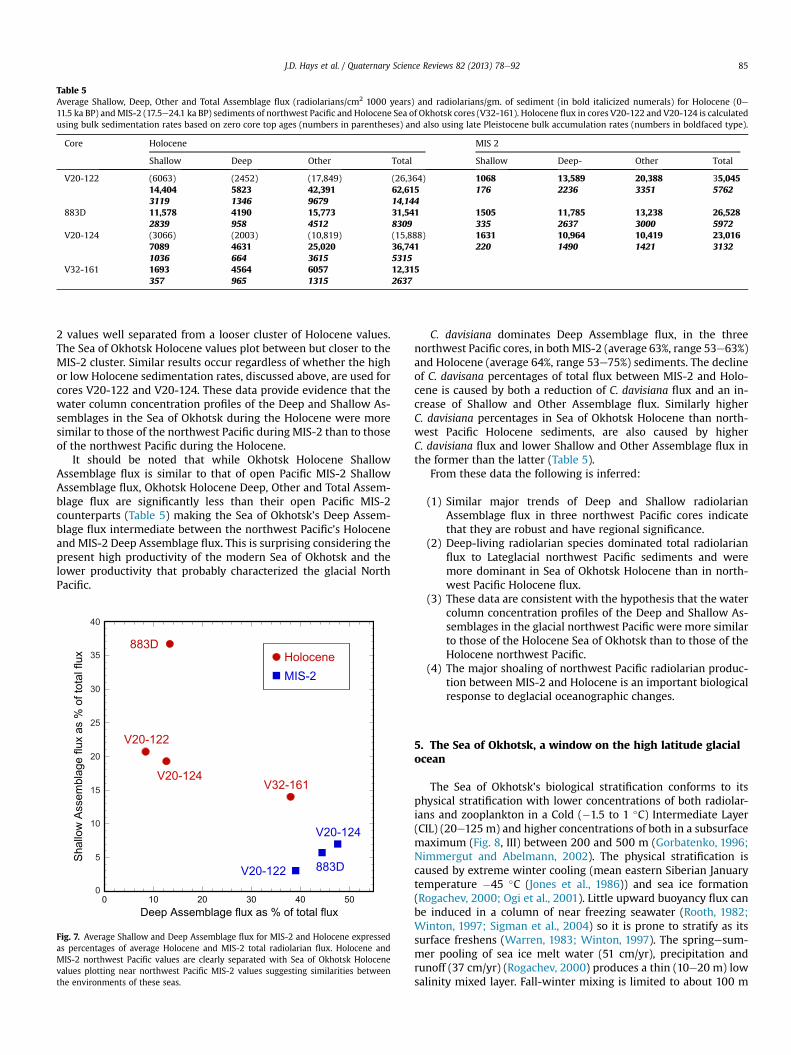
Table 5Average Shallow, Deep, Other and Total Assemblage flux (radiolarians/cm2 1000 years) and radiolarians/gm. of sediment (in bold italicized numerals) for Holocene (0e11.5 ka BP) andMIS-2 (17.5e24.1 ka BP) sediments of northwest Pacific and Holocene Sea of Okhotsk cores (V32-161). Holocene flux in cores V20-122 and V20-124 is calculatedusing bulk sedimentation rates based on zero core top ages (numbers in parentheses) and also using late Pleistocene bulk accumulation rates (numbers in boldfaced type).
Core Holocene MIS 2
Shallow Deep Other Total Shallow Deep- Other Total
V20-122 (6063)14,4043119
(2452)58231346
(17,849)42,3919679
(26,364)62,61514,144
1068176
13,5892236
20,3883351
35,0455762
883D 11,5782839
4190958
15,7734512
31,5418309
1505335
11,7852637
13,2383000
26,5285972
V20-124 (3066)70891036
(2003)4631664
(10,819)25,0203615
(15,888)36,7415315
1631220
10,9641490
10,4191421
23,0163132
V32-161 1693357
4564965
60571315
12,3152637
J.D. Hays et al. / Quaternary Science Reviews 82 (2013) 78e92 85
2 values well separated from a looser cluster of Holocene values.The Sea of Okhotsk Holocene values plot between but closer to theMIS-2 cluster. Similar results occur regardless of whether the highor low Holocene sedimentation rates, discussed above, are used forcores V20-122 and V20-124. These data provide evidence that thewater column concentration profiles of the Deep and Shallow As-semblages in the Sea of Okhotsk during the Holocene were moresimilar to those of the northwest Pacific during MIS-2 than to thoseof the northwest Pacific during the Holocene.
It should be noted that while Okhotsk Holocene ShallowAssemblage flux is similar to that of open Pacific MIS-2 ShallowAssemblage flux, Okhotsk Holocene Deep, Other and Total Assem-blage flux are significantly less than their open Pacific MIS-2counterparts (Table 5) making the Sea of Okhotsk’s Deep Assem-blage flux intermediate between the northwest Pacific’s Holoceneand MIS-2 Deep Assemblage flux. This is surprising considering thepresent high productivity of the modern Sea of Okhotsk and thelower productivity that probably characterized the glacial NorthPacific.
0
5
10
15
20
25
30
35
40
0 10 20 30 40 50
Holocene MIS-2
Shal
low
Ass
embl
age
flux
as %
of t
otal
flux
Deep Assemblage flux as % of total flux
V20-122
V20-124
883D
883D
V32-161
V20-124
V20-122
Fig. 7. Average Shallow and Deep Assemblage flux for MIS-2 and Holocene expressedas percentages of average Holocene and MIS-2 total radiolarian flux. Holocene andMIS-2 northwest Pacific values are clearly separated with Sea of Okhotsk Holocenevalues plotting near northwest Pacific MIS-2 values suggesting similarities betweenthe environments of these seas.
C. davisiana dominates Deep Assemblage flux, in the threenorthwest Pacific cores, in bothMIS-2 (average 63%, range 53e63%)and Holocene (average 64%, range 53e75%) sediments. The declineof C. davisana percentages of total flux between MIS-2 and Holo-cene is caused by both a reduction of C. davisiana flux and an in-crease of Shallow and Other Assemblage flux. Similarly higherC. davisiana percentages in Sea of Okhotsk Holocene than north-west Pacific Holocene sediments, are also caused by higherC. davisiana flux and lower Shallow and Other Assemblage flux inthe former than the latter (Table 5).
From these data the following is inferred:
(1) Similar major trends of Deep and Shallow radiolarianAssemblage flux in three northwest Pacific cores indicatethat they are robust and have regional significance.
(2) Deep-living radiolarian species dominated total radiolarianflux to Lateglacial northwest Pacific sediments and weremore dominant in Sea of Okhotsk Holocene than in north-west Pacific Holocene flux.
(3) These data are consistent with the hypothesis that the watercolumn concentration profiles of the Deep and Shallow As-semblages in the glacial northwest Pacific were more similarto those of the Holocene Sea of Okhotsk than to those of theHolocene northwest Pacific.
(4) The major shoaling of northwest Pacific radiolarian produc-tion between MIS-2 and Holocene is an important biologicalresponse to deglacial oceanographic changes.
5. The Sea of Okhotsk, a window on the high latitude glacialocean
The Sea of Okhotsk’s biological stratification conforms to itsphysical stratification with lower concentrations of both radiolar-ians and zooplankton in a Cold (�1.5 to 1 �C) Intermediate Layer(CIL) (20e125 m) and higher concentrations of both in a subsurfacemaximum (Fig. 8, III) between 200 and 500 m (Gorbatenko, 1996;Nimmergut and Abelmann, 2002). The physical stratification iscaused by extreme winter cooling (mean eastern Siberian Januarytemperature �45 �C (Jones et al., 1986)) and sea ice formation(Rogachev, 2000; Ogi et al., 2001). Little upward buoyancy flux canbe induced in a column of near freezing seawater (Rooth, 1982;Winton, 1997; Sigman et al., 2004) so it is prone to stratify as itssurface freshens (Warren, 1983; Winton, 1997). The springesum-mer pooling of sea ice melt water (51 cm/yr), precipitation andrunoff (37 cm/yr) (Rogachev, 2000) produces a thin (10e20 m) lowsalinity mixed layer. Fall-winter mixing is limited to about 100 m
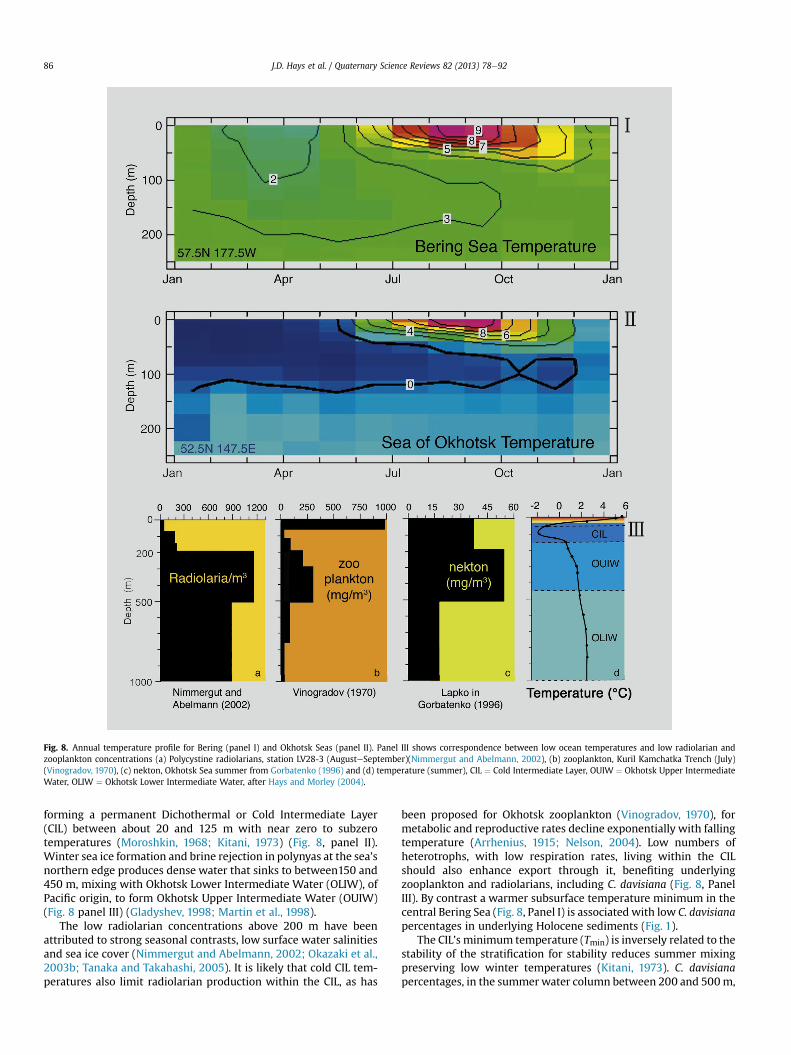
Fig. 8. Annual temperature profile for Bering (panel I) and Okhotsk Seas (panel II). Panel III shows correspondence between low ocean temperatures and low radiolarian andzooplankton concentrations (a) Polycystine radiolarians, station LV28-3 (AugusteSeptember)(Nimmergut and Abelmann, 2002), (b) zooplankton, Kuril Kamchatka Trench (July)(Vinogradov, 1970), (c) nekton, Okhotsk Sea summer from Gorbatenko (1996) and (d) temperature (summer), CIL ¼ Cold Intermediate Layer, OUIW ¼ Okhotsk Upper IntermediateWater, OLIW ¼ Okhotsk Lower Intermediate Water, after Hays and Morley (2004).
J.D. Hays et al. / Quaternary Science Reviews 82 (2013) 78e9286
forming a permanent Dichothermal or Cold Intermediate Layer(CIL) between about 20 and 125 m with near zero to subzerotemperatures (Moroshkin, 1968; Kitani, 1973) (Fig. 8, panel II).Winter sea ice formation and brine rejection in polynyas at the sea’snorthern edge produces dense water that sinks to between150 and450 m, mixing with Okhotsk Lower Intermediate Water (OLIW), ofPacific origin, to form Okhotsk Upper Intermediate Water (OUIW)(Fig. 8 panel III) (Gladyshev, 1998; Martin et al., 1998).
The low radiolarian concentrations above 200 m have beenattributed to strong seasonal contrasts, low surface water salinitiesand sea ice cover (Nimmergut and Abelmann, 2002; Okazaki et al.,2003b; Tanaka and Takahashi, 2005). It is likely that cold CIL tem-peratures also limit radiolarian production within the CIL, as has
been proposed for Okhotsk zooplankton (Vinogradov, 1970), formetabolic and reproductive rates decline exponentially with fallingtemperature (Arrhenius, 1915; Nelson, 2004). Low numbers ofheterotrophs, with low respiration rates, living within the CILshould also enhance export through it, benefiting underlyingzooplankton and radiolarians, including C. davisiana (Fig. 8, PanelIII). By contrast a warmer subsurface temperature minimum in thecentral Bering Sea (Fig. 8, Panel I) is associated with low C. davisianapercentages in underlying Holocene sediments (Fig. 1).
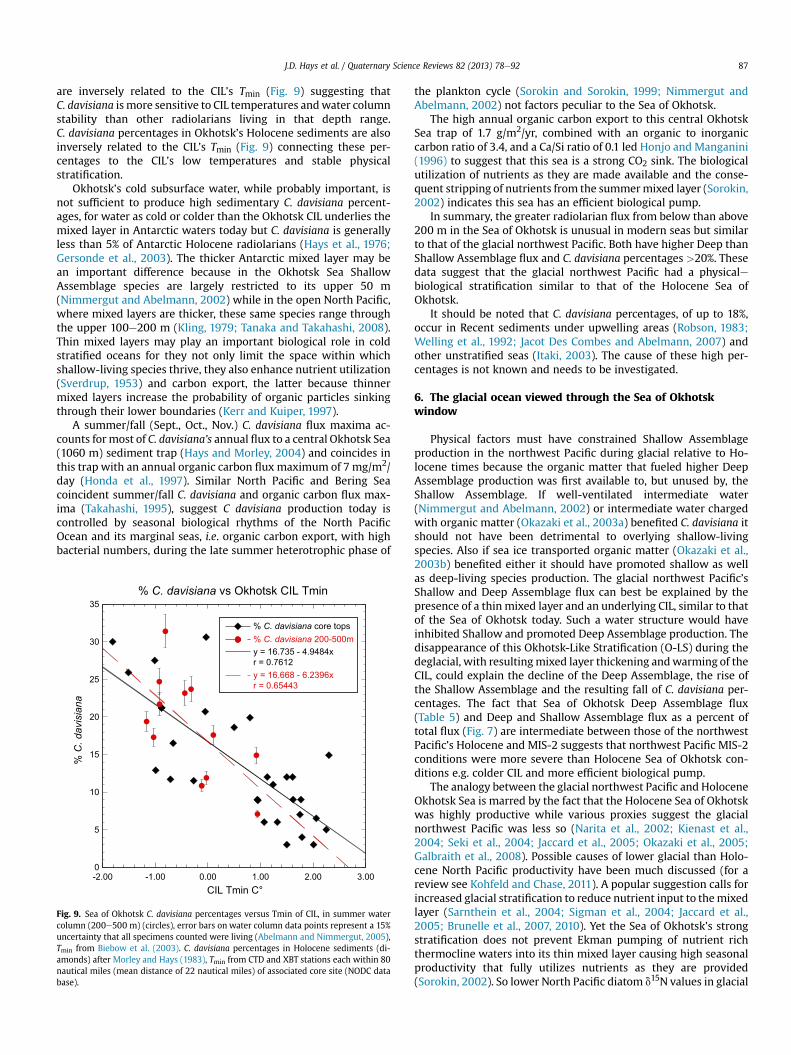
The CIL’s minimum temperature (Tmin) is inversely related to thestability of the stratification for stability reduces summer mixingpreserving low winter temperatures (Kitani, 1973). C. davisianapercentages, in the summerwater column between 200 and 500m,
J.D. Hays et al. / Quaternary Science Reviews 82 (2013) 78e92 87
are inversely related to the CIL’s Tmin (Fig. 9) suggesting thatC. davisiana is more sensitive to CIL temperatures andwater columnstability than other radiolarians living in that depth range.C. davisiana percentages in Okhotsk’s Holocene sediments are alsoinversely related to the CIL’s Tmin (Fig. 9) connecting these per-centages to the CIL’s low temperatures and stable physicalstratification.
Okhotsk’s cold subsurface water, while probably important, isnot sufficient to produce high sedimentary C. davisiana percent-ages, for water as cold or colder than the Okhotsk CIL underlies themixed layer in Antarctic waters today but C. davisiana is generallyless than 5% of Antarctic Holocene radiolarians (Hays et al., 1976;Gersonde et al., 2003). The thicker Antarctic mixed layer may bean important difference because in the Okhotsk Sea ShallowAssemblage species are largely restricted to its upper 50 m(Nimmergut and Abelmann, 2002) while in the open North Pacific,where mixed layers are thicker, these same species range throughthe upper 100e200 m (Kling, 1979; Tanaka and Takahashi, 2008).Thin mixed layers may play an important biological role in coldstratified oceans for they not only limit the space within whichshallow-living species thrive, they also enhance nutrient utilization(Sverdrup, 1953) and carbon export, the latter because thinnermixed layers increase the probability of organic particles sinkingthrough their lower boundaries (Kerr and Kuiper, 1997).
A summer/fall (Sept., Oct., Nov.) C. davisiana flux maxima ac-counts formost of C. davisiana’s annual flux to a central Okhotsk Sea(1060 m) sediment trap (Hays and Morley, 2004) and coincides inthis trap with an annual organic carbon flux maximum of 7 mg/m2/day (Honda et al., 1997). Similar North Pacific and Bering Seacoincident summer/fall C. davisiana and organic carbon flux max-ima (Takahashi, 1995), suggest C davisiana production today iscontrolled by seasonal biological rhythms of the North PacificOcean and its marginal seas, i.e. organic carbon export, with highbacterial numbers, during the late summer heterotrophic phase of
0
5
10
15
20
25
30
35
-2.00 -1.00 0.00 1.00 2.00 3.00
% C. davisiana vs Okhotsk CIL Tmin
% C. davisiana core tops% C. davisiana 200-500my = 16.735 - 4.9484xr = 0.7612 y = 16.668 - 6.2396xr = 0.65443
% C
. d
avis
ian
a
CIL Tmin C°
Fig. 9. Sea of Okhotsk C. davisiana percentages versus Tmin of CIL, in summer watercolumn (200e500 m) (circles), error bars on water column data points represent a 15%uncertainty that all specimens counted were living (Abelmann and Nimmergut, 2005),Tmin from Biebow et al. (2003). C. davisiana percentages in Holocene sediments (di-amonds) after Morley and Hays (1983), Tmin from CTD and XBT stations each within 80nautical miles (mean distance of 22 nautical miles) of associated core site (NODC database).
the plankton cycle (Sorokin and Sorokin, 1999; Nimmergut andAbelmann, 2002) not factors peculiar to the Sea of Okhotsk.
The high annual organic carbon export to this central OkhotskSea trap of 1.7 g/m2/yr, combined with an organic to inorganiccarbon ratio of 3.4, and a Ca/Si ratio of 0.1 led Honjo and Manganini(1996) to suggest that this sea is a strong CO2 sink. The biologicalutilization of nutrients as they are made available and the conse-quent stripping of nutrients from the summermixed layer (Sorokin,2002) indicates this sea has an efficient biological pump.
In summary, the greater radiolarian flux from below than above200 m in the Sea of Okhotsk is unusual in modern seas but similarto that of the glacial northwest Pacific. Both have higher Deep thanShallow Assemblage flux and C. davisiana percentages >20%. Thesedata suggest that the glacial northwest Pacific had a physicalebiological stratification similar to that of the Holocene Sea ofOkhotsk.
It should be noted that C. davisiana percentages, of up to 18%,occur in Recent sediments under upwelling areas (Robson, 1983;Welling et al., 1992; Jacot Des Combes and Abelmann, 2007) andother unstratified seas (Itaki, 2003). The cause of these high per-centages is not known and needs to be investigated.
6. The glacial ocean viewed through the Sea of Okhotskwindow
Physical factors must have constrained Shallow Assemblageproduction in the northwest Pacific during glacial relative to Ho-locene times because the organic matter that fueled higher DeepAssemblage production was first available to, but unused by, theShallow Assemblage. If well-ventilated intermediate water(Nimmergut and Abelmann, 2002) or intermediate water chargedwith organic matter (Okazaki et al., 2003a) benefited C. davisiana itshould not have been detrimental to overlying shallow-livingspecies. Also if sea ice transported organic matter (Okazaki et al.,2003b) benefited either it should have promoted shallow as wellas deep-living species production. The glacial northwest Pacific’sShallow and Deep Assemblage flux can best be explained by thepresence of a thin mixed layer and an underlying CIL, similar to thatof the Sea of Okhotsk today. Such a water structure would haveinhibited Shallow and promoted Deep Assemblage production. Thedisappearance of this Okhotsk-Like Stratification (O-LS) during thedeglacial, with resultingmixed layer thickening andwarming of theCIL, could explain the decline of the Deep Assemblage, the rise ofthe Shallow Assemblage and the resulting fall of C. davisiana per-centages. The fact that Sea of Okhotsk Deep Assemblage flux(Table 5) and Deep and Shallow Assemblage flux as a percent oftotal flux (Fig. 7) are intermediate between those of the northwestPacific’s Holocene and MIS-2 suggests that northwest Pacific MIS-2conditions were more severe than Holocene Sea of Okhotsk con-ditions e.g. colder CIL and more efficient biological pump.
The analogy between the glacial northwest Pacific and HoloceneOkhotsk Sea is marred by the fact that the Holocene Sea of Okhotskwas highly productive while various proxies suggest the glacialnorthwest Pacific was less so (Narita et al., 2002; Kienast et al.,2004; Seki et al., 2004; Jaccard et al., 2005; Okazaki et al., 2005;Galbraith et al., 2008). Possible causes of lower glacial than Holo-cene North Pacific productivity have been much discussed (for areview see Kohfeld and Chase, 2011). A popular suggestion calls forincreased glacial stratification to reduce nutrient input to themixedlayer (Sarnthein et al., 2004; Sigman et al., 2004; Jaccard et al.,2005; Brunelle et al., 2007, 2010). Yet the Sea of Okhotsk’s strongstratification does not prevent Ekman pumping of nutrient richthermocline waters into its thin mixed layer causing high seasonalproductivity that fully utilizes nutrients as they are provided(Sorokin, 2002). So lower North Pacific diatom d15N values in glacial

Fig. 10. LGM C. davisiana percentages suggest Okhotsk-Like Stratification existed at that time north of the present position of the Pacific Polar Front (Belkin et al., 2002). Thegeographical continuity between permafrost and O-LS suggests both were responses to cold winter air. Glacial permafrost distribution (Svendsen et al., 2004; Washburn, 1980 andreferences there in) and C. davisiana percentages; Atlantic data (Morley, 1983), Pacific data (Sachs, 1973; Robertson, 1975; Morley and Hays, 1983; Morley and Robinson, 1986;Morley et al., 1982; Ohkushi et al., 2003; Tanaka and Takahashi, 2005; N. Pisias personal communication and Morley personal communication).
J.D. Hays et al. / Quaternary Science Reviews 82 (2013) 78e9288
relative to interglacial sediments (Sigman et al., 1999; Galbraithet al., 2008) may not have been caused only by stratificationlimiting upward nutrient flux. Alternatively the glacial relative toHolocene supply of silica to the glacial North Pacific and Sea ofOkhotsk thermocline may have been reduced (Gorbarenko et al.,2004; Seki et al., 2004; Okazaki et al., 2005; Kohfeld and Chase,2011). Today mixing in the Kurile Island region brings nutrientrich water from the deep Pacific to and above North Pacific Inter-mediate Water (NPIW) contributing to Sea of Okhotsk and NorthPacific productivity (Tsunogai, 2002; Sarmiento et al., 2004).Because this mixing occurs well below the surface, nutrient ratiosare not reset by sea surface biological processes and retain high Si/Nratios. In glacial times the apparently nutrient depleted water massabove 2000 m (Herguera et al., 1992; Keigwin, 1998; Matsumotoet al., 2002) was probably ventilated in the far North Pacific
(Duplessy et al., 1988; Keigwin, 1998) or Bering Sea (Matsumotoet al., 2002; Ohkushi et al., 2003; Gorbarenko et al., 2010;Horikawa et al., 2010). Because cooling the North Pacific todaywould further stabilize it (Warren, 1983), it is likely that in glacialtime such ventilation occurred through brine rejection in highlatitude polynyas as in the Sea of Okhotsk today (Gorbarenko et al.,2010). During ventilation sea surface biological activity would havereset upwelled nutrient ratios, depleting silica relative to nitrate asoccurs in the Antarctic today (Sarmiento et al., 2004). Glacial NorthPacific Intermediate water depleted of silica in a similar way couldhave lowered glacial relative to Holocene opal flux in the northwestPacific and Sea of Okhotsk (Okazaki et al., 2005; Brunelle et al.,2007; Kohfeld and Chase, 2011). The deglacial initiation of deepmixing in the northwest Pacific should have caused a simultaneousHolocene increase in diatom production throughout the North

J.D. Hays et al. / Quaternary Science Reviews 82 (2013) 78e92 89
Pacific not just in the northwest Pacific and Sea of Okhotsk, yet thismay not have been the case for the northeast Pacific (Kohfeld andChase, 2011). Because the northwest Pacific and Sea of Okhotskare beneficiaries of nutrients supplied directly by upwelled NorthPacific Deep Water while the northeast Pacific receives nutrientsvia the NPIW, the latter’s history may be different (Tsunogai, 2002).The strong dissolution of diatom and radiolarian tests in the glacialsediments of core V32-161 indicates the deep waters of this seawere more under saturated in silica in glacial than Holocene times(Van Cappellen and Qui, 1997; Rickert et al., 2002). A similar degreeof dissolution does not occur in northwest Pacific glacial sediments.In any event it is possible that the glacial northwest Pacific andHolocene Sea of Okhotsk could have had similar physical biologicalstratifications but differing silica supply mechanisms and conse-quent opal flux values.
Cold winter air masses and resultant extensive sea ice causeOkhotsk’s physical stratification today. There is ample evidence ofair colder than today’s eastern Siberian winter air across northerncontinents at the LGM (Atkinson et al., 1987; Cuffey et al., 1995;Isarin et al., 1998; Severinghaus et al., 1998; Isarin and Bohncke,1999; Denton et al., 2005). North Pacific sea ice rafted detritusfrom this time, a consequence of cold winter air, extends to about45�N (Conolly and Ewing, 1970; Kent et al., 1971). Conditionstherefore were in place for O-LS to form and the C. davisiana per-centages north of the 20% line in Fig.10 are evidence that it did. Thisline follows the trend of today’s North Pacific Polar Front thatseparates water with a summer subsurface minimum to the northfrom water without or with a weakly developed subsurface mini-mum to the south (Belkin et al., 2002). For O-LS to develop north ofthe 20% line today would only require a thinning of the mixed layerand cooling the subsurface minimum (CIL).
While summer temperatures play a major role in ice sheetgrowth and decay (Denton et al., 2005) cold winter temperaturesand their duration control the development of permafrost (Nelsonand Outcalt, 1987) and O-LS. The geographical continuity of glacialpermafrost and O-LS (Fig. 10) is consistent with their mutualdependence on long cold winters. Modeling experiments suggestthat reduced northern summer insolation, in response to orbitalvariations, cooled northern continents producing vegetative al-bedo feed-backs that promoted further cooling and associatedcold winter air masses (Harrison et al., 1995; Gallimore andKutzbach, 1996; Ganopolski et al., 1998; Notoro and Liu, 2007,2008). This cold winter air could have engendered permafroston land as well as O-LS on northern seas and the climatic feed-backs that accompany them. If so, O-LS formation and destructioncould have been as abrupt as changes of the seasonal winter airthat controls it.
Widespread high C. davisiana percentages in Antarctic OceanLGM sediments south of the Polar Front (Hays et al., 1976; Gersondeet al., 2003) result from a combination of higher C. davisiana fluxand lower flux of other radiolarians (Hays et al., 1976) likelysignaling the presence of O-LS in this region as well. Radiolarianassemblage data therefore supports inferences of North Pacific andAntarctic stratification based on increased glacial relative to inter-glacial surface water nitrate utilization (lower d15N) (Francois et al.,1997; Sigman et al., 1999; Brunelle et al., 2007, 2010; Galbraithet al., 2008). Phosphate (Elderfield and Rickaby, 2000) and silicadata (De La Rocha et al., 1998) however suggest that the glacialAntarctic was not highly stratified. Low nitrate concentrations alsocharacterize Okhotsk’s thin mixed layer (Sapozhnikov et al., 1999;Arzhanova and Naletova, 1999; Sorokin, 2002), so high latitudeglacial O-LS obviates the need for iron fertilization (Martin, 1990) tolower glacial, relative to Holocene, surface water nitrate concen-trations. Relative to the Holocene this glacial stratification wouldhave deepened organic carbon re-mineralization by radiolarians
and other plankton (Fig. 8, panel III). This potential for O-LS toreduce shallow carbon re-mineralization combined with a generalglacial ocean cooling (Matsumoto, 2007) provides a mechanism forhigh latitude oceans to efficiently transfer carbon to the deep ocean.
Glacial high latitude oceans capped with O-LS had the potentialto amplify glacial cycles by: 1) promoting the spread of sea ice, asoccurs in the Sea of Okhotsk today where the CIL insulates sea icefrom underlying warmer water allowing its spread to the shores ofHokkaido (45�N) (Bulgakov, 1965), a low-latitude extreme formodern sea ice, and a latitude similar to that of the southern limitof glacial North Pacific sea ice rafted detritus (Conolly and Ewing,1970; Kent et al., 1971). Glacial sea ice expansion combined withstrong summer stratification (Fig. 8, panel II) could have reducedglacial CO2 transfer from ocean to atmosphere (Stephens andKeeling, 2000) relative to today; 2) increasing high latitude bio-logical pump efficiency through full utilization of surface waternutrients, depriving the deep-ocean of preformed nutrients andlowering atmospheric CO2 concentrations (Sigman et al., 2010); 3)triggering ocean alkalinity changes by deepening carbon re-mineralization further lowering atmospheric pCO2 (Broecker andPeng, 1987; Boyle, 1988). The deglacial transition from domi-nantly deep to dominantly shallow radiolarian production mayreflect the change from deep to shallow organic carbon re-mineralization envisaged by Boyle (1988). If so it should leadalkalinity induced changes in atmospheric pCO2 by about 2500years (Broecker and Peng, 1987).
This model of the high latitude glacial ocean makes severaltestable predictions. The causal relationship between O-LS and seaice predicts a consistent relationship betweenmaximumwinter seaice extent and high sedimentary C. davisiana percentages, althoughtheir low latitude limits need not be the same. Opal flux peaks inSouthern Ocean cores (Anderson et al., 2002) should be accompa-nied by shallow-living radiolarian flux peaks signaling reducedSouthern Ocean stratification. The deglacial decline of C. davisianapercentages should coincide with or lead the rise of atmosphericpCO2.
7. Conclusions
1. The deepening of northwest Pacific glacial relative to Holoceneradiolarian production records an important change of thisoceans consumer community structure. A similar preference fordeep-living radiolarians and zooplankton occurs in today’s Seaof Okhotsk where cold ocean stratification (O-LS) reducesshallow-relative to deep-living radiolarian production givingrise to the dominance of the deep-living C. davisiana. C. davisianapercentages (>20%) in high latitude (>45�) LGM sediments ofboth hemispheres suggest these seas were capped with O-LS.
2. The physical and biological structure of glacial high latitudeocean stratification presented here has several potential climaticfeedbacks; a) it promotes winter sea ice spread by insulating seaice from warmer underlying water; b) its thin mixed layer fullyutilizes surface water nutrients enhancing biological pump ef-ficiency, and low respiration rates within the CIL deepen carbonre-mineralization, both of which could aid CO2 transfer fromatmosphere to ocean in summer; c) the summer stratificationmay have reduced CO2 return to the atmosphere;
3. O-LS and its biological consequences rest on two fundamentalproperties of Earth’s climate system; a) the non-linearity ofseawater’s equation of state that promotes cold ocean stratifi-cation; b) the exponential relationship between temperatureand heterotrophic metabolic and reproductive rates that limitthe rate of organic carbon re-mineralization within cold nearsurface waters (CILs).

J.D. Hays et al. / Quaternary Science Reviews 82 (2013) 78e9290
Acknowledgements
We are grateful to Aaron Putnam, Dorothy Peteet, O. RogerAnderson, Michael Sarnthein, Taro Takahashi, Maureen Raymoand Wallace Broecker for reading the manuscript and offeringuseful suggestions. We are especially grateful to Ray Sambrotto,and Arnold Gordon for helpful discussions during manuscriptpreparation and for useful comments on multiple drafts of thispaper. We are also grateful for the constructive suggestions of fouranonymous reviewers. We appreciate the expert assistance ofMaureen McAuliffe Anders during final manuscript preparationand are grateful to Nick Pisias for unpublished C. davisiana data.We thank the Ocean Drilling Program and its successor IODP forcollecting and archiving the drill core used. The piston cores arearchived at the Lamont-Doherty Earth Observatory of ColumbiaUniversity whose core repository is supported by funds fromNational Science Foundation Grant OCE09-62010. This work wassupported with funds through the Cooperative Institute forClimate Applications and Research (CICAR) under award numberNA030AR4320179 from the National Oceanic and AtmosphericAdministration, U.S. Department of Commerce. The statements,findings, conclusions, and recommendations are those of the au-thors and do not necessarily reflect the views of the NationalOceanic and Atmospheric Administration or the Department ofCommerce. This paper bears Lamont Doherty Earth Observatorycontribution number 7735.
References
Abelmann, A., Gowing, M.M., 1996. Horizontal and vertical distribution pattern ofliving radiolarians along a transect from the Southern Ocean to the SouthAtlantic subtropical region. Deep-Sea Res. Part I 43, 361e382.
Abelmann, A., Gowing, M.M., 1997. Spatial distribution pattern of living polycystineradiolarian taxa e baseline study for paleoenvironmental reconstructions in theSouthern Ocean (Atlantic sector). Mar. Micropaleontol. 30, 3e28.
Abelmann, A., Nimmergut, A., 2005. Radiolarians in the Sea of Okhotsk and theirecological implication for paleoenvironmental reconstructions. Deep-Sea Res.Part II 52, 2302e2331.
Anderson, O.R., 1983. Radiolaria. Springer-Verlag, New York, p. 355.Anderson, R.F., Chase, Z., Fleisher, M.Q., Sachs, J., 2002. The southern Ocean’s bio-
logical pump during the last glacial maximum. Deep-Sea Res. Part II 49, 1909e1938.
Archer, D.E., Eshel, G., Winguth, A., Broecker, W., Pierrehumbert, R., Tobis, M.,Jacob, R., 2000. Atmospheric pCO2 sensitivity to the biological pump in theocean. Glob. Biogeochem. Cycles 14, 1219e1230.
Arrhenius, S., 1915. Quantitative Laws in Biological Chemistry. G. Bell and Sons Ltd.,London, p. 164.
Arzhanova, N.V., Naletova, I.A., 1999. Hydrochemical structure, mesoscale eddies,and primary production in the northern part of the Sea of Okhotsk. Okeanology39, 741e749.
Atkinson, T.C., Briffa, K.R., Coope, G.R., 1987. Seasonal temperature in Britainduring the last 22,000 years, reconstructed using beetle remains. Nature 325,587e592.
Barron, J.A., Gladenkov, Y.A., 1995. Early MioceneePleistocene diatom stratigraphyof leg 145. In: Rea, D.K., Basov, I.A., Scholl, D.W., Allan, J.F. (Eds.), Proc. OceanDrilling Prog. Sci. Res. 145, pp. 3e19.
Belkin, I., Krishfield, R., Honjo, S., 2002. Decadal variability of the North Pacific PolarFront: subsurface warming versus surface cooling. Geophys. Res. Lett. 29, 65-1e65-4.
Berner, W.H., Stouffer, B., Oeschger, H., 1979. Past atmospheric composition andclimate, gas parameters measured on ice cores. Nature 275, 53e55.
Biebow, N., Kulinich, R., Baranov, B.V., 2003. Cruise Reports: RV Akademik M.A.Lavrentyev Cruise 29, Leg 1 and Leg 2. GEOMAR Rept. 110, p. 190.
Blueford, J.R., 1983. Distribution of quaternary radiolaria in the Navarin Basingeologic province, Bering Sea. Deep-Sea Res. Part A 30, 763e781.
Boltovskoy, D., Riedel, W.R., 1980. Polycystine radiolaria from the southwesternAtlantic Ocean plankton. Rev. Esp. Micropaleontol. 12, 99e146.
Boyd, P.W., Newton, P., 1995. Evidence of the potential influence of planktoniccommunity structure on the interannual variability of particulate organic car-bon flux. Deep-Sea Res. Part I 42, 619e639.
Boyle, E.A., 1988. Vertical oceanic nutrient fractionation and glacial/interglacial CO2cycles. Nature 331, 55e56.
Broecker, W.S., Peng, T.-H., 1982. Tracers in the Sea. Eldigio Press, Lamont-DohertyGeological Observatory, Columbia University, Palisades, N. Y., p. 690
Broecker, W.S., Peng, T.-H., 1987. The role of CaCO3 compensation in the glacial tointerglacial atmospheric CO2 change. Glob. Biogeochem. Cycles 1, 15e29.
Brunelle, B.G., Sigman, D.M., Cook, M.S., Keigwin, L.D., Haug, G.H., Plessen, B.,Schettler, G., Jaccard, S.L., 2007. Evidence from diatom-bound nitrogen isotopesfor subarctic Pacific stratification during the last ice age and a link to NorthPacific denitrification changes. Paleoceanography 22, PA1215. http://dx.doi.org/10.1029/2005PA001205.
Brunelle, B.G., Sigman, D.M., Jaccard, S.L., Keigwin, L.D., Plessen, B., Schettler, G.,Cook, M.S., Haug, G.H., 2010. Glacial/interglacial changes in nutrient supply andstratification in the western subarctic North Pacific since the penultimateglacial maximum. Quat. Sci. Rev. 29, 2579e2590.
Bulgakov, N.P., 1965. The extreme winter boundary of ice in far easternseas ¼ Predel’naya zimniaya granitsa l’dov v dal’nevostochnykh moryakh. Nav.Oceanogr. Off. Transl. 292 (13), 66e76 (in Russian).
Conolly, J.R., Ewing, M., 1970. Ice-rafted detritus in northwest Pacific deep-seasediments. In: Hays, J.D. (Ed.), Geological Investigations of the North Pacific,Geol. Soc. Am. Mem. 126, Boulder, Colorado, pp. 219e232.
Crusius, J., Pedersen, T.F., Kienast, S., Keigwin, L., Labeyrie, L., 2004. Influence of north-west Pacific productivity on North Pacific intermediate water oxygen concentra-tions during the Boiling-Allerød interval (14.7e12.9 ka). Geology 32, 633e636.
Cuffey, K.M., Clow, G.D., Alley, R.B., Stuiver, M., Waddington, E.D., Saltus, R.W., 1995.Large Arctic temperature change at the Wisconsin-Holocene transition. Science270, 455e458.
Curry, W.B., Oppo, D.W., 2005. Glacial water mass geometry and the distribution ofdelta 13C of total CO2 in the western Atlantic Ocean. Paleoceanography 20,PA1017. http://dx.doi.org/10.1029/2004PA001021.
De La Rocha, C.L., Brzezinski, M.A., DeNiro, M.J., Shemesh, A., 1998. Silicon-isotopecomposition of diatoms as an indicator of past oceanic change. Nature 395,680e683.
Denton, G.H., Alley, R.B., Comer, G.C., Broecker, W.S., 2005. The role of seasonality inabrupt climate change. Quat. Sci. Rev. 24, 1159e1182.
Draper, N.R., Smith, H., 1988. Applied Regression Analysis, third ed. Wiley-Inter-science, p. 736.
Duplessy, J.C., Arnold, M., Shackleton, N.J., Kallel, N., Labeyrie, L., Juillet-Leclerc, A.,1988. Changes in the rate of ventilation of intermediate and deep-water massesin the Pacific Ocean during the last deglaciation. Chem. Geol. 70, 108.
Elderfield, H., Rickaby, R., 2000. Oceanic Cd/P ratio and nutrient utilization in theglacial Southern Ocean. Nature 405, 305e310.
Eppley, R.W., Peterson, B.J., 1979. Particulate organic matter flux and planktonic newproduction in the deep ocean. Nature 282, 677e680.
Francois, R., Altabet, M.A., Yu, E.-F., Sigman, D.M., Bacon, M.P., Frank, M.,Bohrmann, G., Bareille, G., Labeyrie, L.D., 1997. Contribution of Southern Oceansurface-water stratification to low atmospheric CO2 concentrations during thelast glacial period. Nature 389, 929e935.
Galbraith, E.D., Kienast, M., Jaccard, S.L., Pedersen, T.F., Brunelle, B.G., Sigman, D.M.,Kiefer, T., 2008. Consistent relationship between global climate and surfacenitrate utilization in the western subarctic Pacific throughout the last 500 ka.Paleoceanography 23, PA2212. http://dx.doi.org/10.1029/2007PA001518.
Gallimore, R.G., Kutzbach, J.E., 1996. Role of orbitally induced changes in tundra areain the onset of glaciation. Nature 381, 503e505.
Ganopolski, A., Kubatzki, C., Clausen, M., Brovkin, V., Petoukhov, V., 1998. The in-fluence of vegetationeatmosphereeocean interaction on climate during theMid-Holocene. Science 280, 1916e1919.
Gardner, W.D., Walsh, I.D., Richardson, M.J., 1993. Biophysical forcing of particleproduction and distribution during a spring bloom in the North Atlantic. Deep-Sea Res. Part II 40, 171e195.
Gebhardt, H., Sarnthein, M., Grootes, P.M., Kiefer, T., Kuehn, H., Schmieder, F.,Röhl, U., 2008. Paleonutrient and productivity records from the subarctic NorthPacific for Pleistocene glacial terminations I to V. Paleoceanography 23, PA4212.http://dx.doi.org/10.1029/2007PA001513.
Gersonde, R., Abelmann, A., Brathauer, U., Becquey, S., Bianchi, C., Cortese, G.,Grobe, H., Kuhn, G., Niebler, H.-S., Segl, M., Sieger, R., Zielinski, U., Fuetterer, D.K.,2003. Last glacial sea surface temperatures and sea-ice extent in the SouthernOcean (AtlanticeIndian sector): a multiproxy approach. Paleoceanography 18,PA1061. http://dx.doi.org/10.1029/2002PA000809.
Gladyshev, S., 1998. Estimation of dense cold water formation on the northern shelfof the Sea of Okhotsk. Russ. Meteorol. Hydrol. 4, 53e59.
Gorbarenko, S.A., Southon, J., Keigwin, L., Cherepanova, M.V., Gvozdeva, I.G., 2004.Late PleistoceneeHolocene oceanographic variability in the Okhotsk Sea:geochemical, lithological and paleontological evidence. Palaeogeogr. Palae-oclimatol. Palaeoecol. 209, 281e301.
Gorbarenko, S.A., Psheneva, O. Yu, Artemova, A.V., Matul, A.G., Tiedemann, R.,Nürnberg, D., 2010. Paleoenvironment changes in the NW Okhotsk Sea for thelast 18 ka determined with micropaleontological, geochemical, and lithologicaldata. Deep-Sea Res. Part I 57, 797e811.
Gorbatenko, K.M., 1996. Seasonal aspects of the vertical distribution of zooplanktonin the Sea of Okhotsk. Izvestya Ticookeanskovo Nauchno-IsledovatelskvoRibocoziastvenovo Centra 119, 88e119 (in Russian).
Gowing, M.M., 1986. Trophic biology of phaeodarian radiolarians and flux of livingradiolarians in the upper 2000 m of the North Pacific central gyre. Deep-SeaRes. Part A 33, 655e674.
Hargrave, B.T., 1975. The importance of total and mixed layer depth in the supply oforganic material to bottom communities. Symp. Biol. Hung. 15, 157e165.
Harrison, S.P., Kutzbach, J.E., Prentice, I.C., Behling, P.J., Sykes, M.T., 1995. Theresponse of the Northern Hemisphere extratropical climate and vegetation toorbitally induced changes in insolation during the last interglaciation. Quat. Res.43, 174e184.

J.D. Hays et al. / Quaternary Science Reviews 82 (2013) 78e92 91
Haug, G.H., Ganopolski, A., Sigman, D.M., Rosell-Mele, A., Swann, G.E.A.,Tiedemann, R., Jaccard, S.L., Bollmann, J., Maslin, M.A., Leng, M.J., Eglinton, G.,2005. North Pacific seasonality and the glaciation of North America 2.7 millionyears ago. Nature 433, 821e825.
Hays, J.D., Lozano, J.A., Shackleton, N.J., Irving, G., 1976. Reconstructions of theAtlantic and western Indian Ocean sectors of the 18,000 B.P. Antarctic Ocean. In:Cline, R.M., Hays, J.D. (Eds.), Investigations of Late Quaternary Paleoceanog-raphy and Paleoclimatology, Geol. Soc. Am. Mem. 145, Boulder, Colorado,pp. 337e372.
Hays, J.D., Morley, J.J., 2004. The Sea of Okhotsk: a window on the ice age ocean.Deep-Sea Res. Part I 51, 593e618.
Herguera, J.C., 1992. Deep-sea benthic foraminifera and biogenic opal: glacial topostglacial productivity changes in the western equatorial Pacific. Mar. Micro-paleontol. 19, 79e98.
Herguera, J.C., Jansen, C., Berger, W.H., 1992. Evidence of a bathyl front at 2000 mdepth in the glacial Pacific, based on a depth transect on Ontong Java Plateau.Paleoceanography 7, 273e288.
Honda, M.C., Kusakabe, M., Nakabayashi, S., Manganini, S.J., Honjo, S., 1997. Changein pCO2 through biological activity in the marginal seas of the western NorthPacific: the efficiency of the biological pump estimated by a sediment trapexperiment. J. Oceanogr. 53, 645e662.
Honjo, S., Manganini, S.J., 1996. Dichothermal Layer and Biological Export Produc-tion in the Sea of Okhotsk, International Workshop on the Okhotsk Sea andArctic; the Physics and Biogeochemistry Implied to the Global Cycles (Influenceof Sea Ice on Climate and Marine Ecosystems). Japan Marine Science andTechnology Center, Yokosuka, Japan, pp. 103e110.
Horikawa, K., Asahara, Y., Yamamoto, K., Okazaki, Y., 2010. Intermediate water for-mation in the Bering Sea during glacial periods: evidence from neodymiumisotope ratios. Geology 38, 435e438.
Hülsemann, K., 1963. Radiolaria in Plankton from the Arctic Drifting Station T-3,Including the Description of Three New Species. Arctic Institute of NorthAmerica Technical Papers 13, pp. 1e52.
Isarin, R.F.B., Bohncke, S.J.P., 1999. Mean july temperatures during the YoungerDryas in northwestern and central Europe as inferred from climate indicatorplant species. Quat. Res. 51, 158e173.
Isarin, R.F.B., Renssen, H., Vandenberghe, J., 1998. The impact of the North AtlanticOcean on the Younger Dryas climate of northwestern and central Europe.J. Quat. Sci. 13, 447e453.
Itaki, T., 2003. Depth-related radiolarian assemblage in the water-column andsurface sediments of the Japan Sea. Mar. Micropaleontol. 47, 253e270.
Itaki, T., Ito, M., Narita, H., Ahagon, N., Sakai, H., 2003. Depth distribution of radi-olarians from the Chukchi and Beaufort Seas, western Arctic. Deep-Sea Res. PartI 50, 1507e1522.
Jaccard, S.L., Haug, G.H., Sigman, D.M., Pedersen, T.F., Thierstein, H.R., Röhl, U., 2005.Glacial/interglacial changes in subarctic North Pacific stratification. Science 308,1003e1006.
Jaccard, S.L., Galbraith, E.D., Sigman, D.M., Haug, G.H., Francois, R., Pedersen, T.F.,Dulski, P., Thierstein, H.R., 2009. Subarctic Pacific evidence for a glacial deep-ening of the oceanic respired carbon pool. Earth Planet. Sci. Lett. 277, 156e165.
Jacot Des Combes, H., Abelmann, A., 2007. A 350-ky radiolarian record off Lüderitz,Namibiaeevidence for changes in the upwelling regime. Mar. Micropaleontol.62, 194e210.
Jones, P.D., Raper, S.C.B., Bradley, R.S., Diaz, H.F., Kelly, P.M., Wigley, T.M.L., 1986.Northern hemisphere surface air temperature variations: 1851e1984. J. Clim.25, 161e179.
Keigwin, L., Jones, G.A., Froelich, P.N., 1992. A 15,000 year paleoenvironmental re-cord from Meiji Seamount, far northwestern Pacific. Earth Planet. Sci. Lett. 111,425e440.
Keigwin, L.D., 1995. Stable isotope stratigraphy and chronology of he upper Quater-nary section of site 883 Detroit Seamount. In: Rea, D.K., Basov, I.A., Scholl, D.W.,Allan, J.F. (Eds.), Proc. Ocean Drilling Prog. Sci. Res. 145, pp. 257e264.
Keigwin, L.D., 1998. Glacial-age hydrography of the far northwest Pacific Ocean.Paleoceanography 13, 323e339.
Kent, D.V., Opdyke, N.D., Ewing, M., 1971. Climate change in the North Pacific usingice-rafted detritus as a climatic indicator. Geol. Soc. Am. Bull. 82, 2741e2754.
Kerr, R.C., Kuiper, G.S., 1997. Particle settling through a diffusive-type thermohalinestaircase in the ocean. Deep-Sea Res. Part I 44, 399e412.
Kiefer, T., Sarnthein, M., Erlenkeuser, H., Grootes, P.M., Roberts, A.P., 2001. NorthPacific response to millennial-scale changes in ocean circulation over the last60 ka. Paleoceanography 16, 179e189.
Kienast, S.S., Hendy, I.L., Crusius, J., Pedersen, T.F., Calvert, S.E., 2004. Export pro-duction in the subarctic North Pacific over the last 800 kyrs: no evidence foriron fertilization? J. Oceanogr. 60, 189e203.
Kitani, K., 1973. An oceanographic study of the Okhotsk Sea e particularly in regardto cold waters. Bull. Far Seas Fish. Res. Lab. 9, 45e76.
Kling, S.A., 1979. Vertical distribution of polycystine radiolarians in the centralNorth Pacific. Mar. Micropaleontol. 4, 295e318.
Kling, S.A., Boltovskoy, D., 1995. Radiolarian vertical distribution patterns across theSouthern California current. Deep-Sea Res. Part I 42, 191e231.
Kohfeld, K., Chase, Z., 2011. Controls on deglacial changes in biogenic fluxes in theNorth Pacific Ocean. Quat. Sci. Rev. 30, 3350e3363.
Koizumi, I., Yamamoto, H., 2007. Paleohydrography of the Kuroshio-Kuroshioextension off Sanriku coast based on fossil diatoms. JAMSTEC Rep. Res.Develop. 5, 1e8.
Kruglikova, S.B., 1975. Radiolarians in the surface layer of sediments in the Sea ofOkhotsk. Oceanology 15 (1), 82e86.
Ling, H.Y., Stadum, C.J., Welch, M.L., 1971. Polycystine radiolaria from Bering Seasurface sediments. In: Farinacci, A. (Ed.), Proceedings of the 2nd PlanktonicConference, Rome, 1970, pp. 705e729. Tecnoscienza, Rome.
Martin, J.H., 1990. Glacialeinterglacial CO2 change: the iron hypothesis. Paleo-ceanography 5, 1e13.
Martin, S., Drucker, R., Yamashita, K., 1998. The production of ice and dense shelfwater in the Okhotsk Sea polynyas. J. Geophys. Res. 103, 27771e27782.
Martinson, D.G., Pisias, N.G., Hays, J.D., Imbrie, J., Moore Jr., T.C., Shackleton, N.J.,1987. Age dating and the orbital theory of the Ice Ages: development of a highresolution 0e300,000-year chronostratigraphy. Quat. Res. 1, 1e29.
Matsumoto, K., Oba, T., Lynch-Stieglitz, J., Yamamoto, H., 2002. Interior hydrographyand circulation of the glacial deep Pacific. Quat. Sci. Rev. 21, 1693e1704.
Matsumoto, K., 2007. Biology-mediated temperature control on atmospheric pCO2and ocean biogeochemistry. Geophys. Res. Lett. 34. http://dx.doi.org/10.1029/2007GL031301.
Matul, A.G., 2011. The recent and quaternary distribution of the radiolarian speciesCycladophora davisiana: a biostratigraphic and paleoceanographic tool. Ocean-ology 51, 335e346.
McMillen, K.J., Casey, R.E., 1978. Distribution of living polycystine radiolarians in theGulf of Mexico and Caribbean Sea, and comparison with the sedimentary re-cord. Mar. Micropaleontol. 3, 121e145.
Michaels, A.F., Silver, M.W., 1988. Primary production, sinking fluxes and the mi-crobial food web. Deep-Sea Res. Part A 35, 473e490.
Moore, T.C.J., 1973. Method of randomly distributing grains for microscopic exam-ination. J. Sediment. Petrol 43, 904e906.
Morley, J.J., 1983. Identification of density-stratified waters in the late PleistoceneNorth Atlantic: a faunal derivation. Quat. Res. 20, 374e386.
Morley, J.J., Hays, J.D., 1983. Oceanographic conditions associated with the highabundances of the radiolarian Cycladophora davisiana. Earth Planet. Sci. Lett. 66,63e72.
Morley, J.J., Robinson, S.W., 1986. Improved method for correlating late Pleistocene/Holocene records from the Bering Sea: application of a biosiliceous/geochem-ical stratigraphy. Deep-Sea Res. 33, 1203e1211.
Morley, J.J., Stepien, J.C., 1985. Antarctic Radiolaria in late winter/early springWeddell Sea waters. Micropaleontology 31, 365e371.
Morley, J.J., Hays, J.D., Robertson, J.H., 1982. Stratigraphic framework for the latePleistocene in the northwest Pacific Ocean. Deep-Sea Res. Part A 29, 1485e1499.
Morley, J.J., Heusser, L.E., Shackleton, N.J., 1991. Late Pleistocene/Holocene radio-larian and pollen records from sediments in the Sea of Okhotsk. Paleoceanog-raphy 6, 121e131.
Morley, J.J., Shemesh, A., Abelmann, A., 2013. Laboratoryanalysis of dissolution effectson Southern Ocean polycystine Radiolaria. Mar. Micropaleontol. (in press).
Moroshkin, K.V., 1968. Water Masses of the Sea of Okhotsk. Joint PublicationsResearch Service, Washington, D.C, p. 98.
Mortlock, R.A., Froelich, P.N., 1989. A simple method for the rapid determination ofbiogenic opal in pelagic marine sediments. Deep-Sea Res. Part A 36, 1415e1426.
Narita, H., Sato, M., Tsunogai, S., Murayama, M., Ikehara, M., Nakatsuka, T.,Wakatsuchi, M., Harada, N., Ujiié, Y., 2002. Biogenic opal indicating less pro-ductive northwestern North Pacific during the glacial ages. Geophys. Res. Lett.29, 22-1e22-4. http://dx.doi.org/10.1029/2001GL014320.
Nelson, F.E., Outcalt, S.I., 1987. A computational method for prediction andregionalization of permafrost. Arct. Alp. Res. 19, 279e288.
Nelson, P., 2004. Biological Physics: Energy, Information, Life. W. H. Freeman, NewYork, p. 598.
Nigrini, C.A., Moore, T.C., 1979. A Guide to Modern Radiolaria. In: Cushman Foun-dation for Foraminiferal Res. Spec. Publ. 16, 342 pp.
Nimmergut, A., Abelmann, A., 2002. Spatial and seasonal changes of radiolarianstanding stocks in the Sea of Okhotsk. Deep-Sea Res. Part I 49, 463e493.
Ninkovich, D., Robertson, J.H., 1975. Volcanogenic effects on rates of deposition ofsediments in northwest Pacific Ocean. Earth Planet. Sci. Lett. 27, 127e136.
Notoro, M., Liu, Z., 2007. Potential impact of the Eurasian boreal forest on NorthPacific climate variability. J. Clim. 20, 981e992.
Notoro, M., Liu, Z., 2008. Statistical and dynamical assessment of vegetation feed-backs on climate over the boreal forest. Clim. Dyn. 31, 691e712.
Ogi, M., Tachibana, Y., Nishio, F., Danchenkov, M.A., 2001. Does the fresh watersupply from the Amur River flowing into the Sea of Okhotsk affect sea iceformation? J. Meteorol. Soc. Jpn. 79, 123e129.
Ohkushi, K., Itaki, T., Nemoto, N., 2003. Last GlacialeHolocene change inintermediate-water ventilation in the Northwestern Pacific. Quat. Sci. Rev. 22,1477e1484.
Okazaki, Y., Takahashi, K., Nakatsuka, T., Honda, M.C., 2003a. The productionscheme of Cycladophora davisiana (Radiolaria) in the Okhotsk Sea and thenorthwestern North Pacific: implications for the palaeoceanographic conditionsduring the glacials in the high latitude oceans. Geophys. Res. Lett. 30. http://dx.doi.org/10.1029/2003GL018070.
Okazaki, Y., Takahashi, K., Yoshitani, H., Nakatsuka, T., Ikehara, M., Wakatsuchi, M.,2003b. Radiolarians under the seasonally sea-ice covered conditions in theOkhotsk Sea: flux and their implications for palaeoceanography. Mar. Micro-paleontol. 49, 195e230.
Okazaki, Y., Takahashi, K., Itaki, T., Kawasaki, Y., 2004. Comparison of radiolarianvertical distributions in the Okhotsk Sea near Kuril islands and the north-western North Pacific off Hokkaido Island. Mar. Micropaleontol. 51, 257e284.

J.D. Hays et al. / Quaternary Science Reviews 82 (2013) 78e9292
Okazaki, Y., Takahashi, K., Katsuki, K., Ono, A., Hori, J., Sakamoto, T., Uchida, M.,Shibata, Y., Ikehara, M., Aoki, K., 2005. Late Quaternary paleoceanographicchanges in the southwestern Okhotsk Sea: evidence from geochemical, radio-larian, and diatom records. Deep-Sea Res. Part II 52, 2332e2350.
Petit, J.R., Jouzel, J., Raynaud, D., Barkov, N.I., Barnola, J.-M., Basile, I., Bender, M.L.,Chappellaz, J., Davis, M., Delaygue, G., Delmotte, M., Kotlyokov, V.M.,Legrand, M., Lipenkov, V.Y., Lorius, C., Pepin, L., Ritz, C., Saltzman, E.,Stievenard, M., 1999. Climate and atmospheric history of the past 420,000 yearsfrom the Vostok ice core, Antarctica. Nature 399, 429e436.
Petrushevskaya, M., 1967. Radiolarian of orders Spumellaria and Nassellaria of theAntarctic region. In: Andriyashev, A.P., Ushakov, P.V. (Eds.), Biological Report ofthe Soviet Antarctic Expedition (1955e1958), vol. 3. Academy of Sciences of theUSSR, Israel Program for Scientific Translations, Jerusalem, pp. 2e186.
Rea, D.K., Basov, I.A., Janecek, T.R., Palmer-Julson, A., et al., 1993. Proc. Ocean DrillingProg., Init. Repts. Leg 145. College Station, TX. http://dx.doi.org/10.2973/odp.proc.ir.145.1993.
Renz, G.W., 1976. The distribution and ecology of Radiolaria in the central Pacific:plankton and surface sediments. Bull. Scripps Inst. Oceanogr. 22, 1e267.
Rickert, D., Schluter, M., Wallmann, K., 2002. Dissolution kinetics of biogenic silicafrom the water column to the sediments. Geochim. Cosomochim. Acta 66,439e455.
Robertson, J.H., 1975. Glacial to Interglacial Oceanographic Changes in the North-west Pacific, Including a Continuous Record of the Last 400,000 Years. Ph.D.thesis. Columbia University, New York, p. 355.
Robson, S., 1983. The distribution of recent Radiolaria in the surficial sediments ofthe continental margin off northern Namibia. J. Micropalaeontol. 2, 31e38.
Rogachev, K.A., 2000. Recent variability in the Pacific western subarctic boundarycurrents and Sea of Okhotsk. Prog. Oceanogr. 47, 299e336.
Rooth, C., 1982. Hydrology and ocean circulation. Prog. Oceanogr. 11, 131e149.Sachs, H.M., 1973. Quantitative Radiolarian-based Paleo-oceanography in Late
Pleistocene Subarcitc Pacific Sediments. Ph.D. thesis. Brown University, Provi-dence, Rhode Island, p. 208.
Sapozhnikov, V.V., Gruzevich, A.K., Arzhanova, N.V., Naletova, I.A., Zubarevich, V.L.,Sapozhnikov, M.V., 1999. Principal features of spatial distribution of organic andinorganic nutrient compounds in the Sea of Okhotsk. Okeanology 39, 198e204.
Sarmiento, J.L., Gruber, N., Brzezinski, M.A., Dunne, J.P., 2004. High-Latitude controlsof thermocline nutrients and low latitude biological productivity. Nature 427,56e60.
Sarnthein, M., Gebhardt, H., Kiefer, T., Kucera, M., Cook, M., Erlenkeuser, H., 2004.Mid Holocene origin of the sea surface salinity low in the Subarctic North Pa-cific. Quat. Sci. Rev. 23, 2089e2099.
Sato, M.M., Narita, H., Tsugonai, S., 2002. Barium increasing prior to opal during thelast termination of glacial ages in the Okhotsk Sea sediments. J. Oceanogr. 58,461e467.
Seki, O., Ikehara, M., Kawamura, K., Nakatsuka, T., Ohnishi, K., Wakatsuchi, M.,Narita, H., Sakamoto, T., 2004. Reconstruction of paleoproductivity in the Sea ofOkhotsk over the last 30 ka. Paleoceanography 19, PA1016. http://dx.doi.org/10.1029/2002PA000808.
Severinghaus, J., Sowers, T., Brook, E.J., Alley, R.B., Bender, M.L., 1998. Timing ofabrupt climate change at the end of the Younger Dryas interval from thermallyfractionated gases in polar ice. Nature 391, 141e146.
Shackleton, N.J., Berger, A., Peltier, W.R., 1990. An alternative astronomical calibra-tion of the lower Pleistocene timescale based on ODP site 677. Trans. Royal Soc.Edinb.: Earth Sci. 81, 251e261.
Shemesh, A., Burckle, L.H., Froelich, P.N., 1989. Dissolution and preservation ofAntarctic diatoms and the effect on sediment thanatocoenoses. Quat. Res. 31,288e308.
Shigemitsu, M., Narita, H., Watanabe, Y.W., Harada, N., Tsunogai, S., 2007. Ba, Si, U,Al, Sc, La, Th, C, and 13C/12C in a sediment core in the western subarctic Pacific asproxies of past biological production. Mar. Chem. 106, 442e455.
Sigman, D.M., Altabet, M.A., Francois, R., McCorkle, D.C., Gaillard, J.-F., 1999. Theisotopic composition of diatom-bound nitrogen in Southern Ocean sediments.Paleoceanography 14, 118e134.
Sigman, D.M., Jaccard, S.L., Haug, G.H., 2004. Polar ocean stratification in a coldclimate. Nature 428, 59e63.
Sigman, D.M., Hain, M.P., Haug, G.H., 2010. The polar ocean and glacial cycles inatmospheric CO2 concentration. Nature 466, 47e55.
Sorokin, Y.I., 2002. Dynamics of inorganic phosphorus in pelagic communities of theSea of Okhotsk. J. Plankton Res. 24, 1253e1263.
Sorokin, Y.I., Sorokin, P.Y., 1999. Production in the Sea of Okhotsk. J. Plankton Res. 21,201e230.
Spencer-Cervato, C., Lazarus, D.B., Beckmann, J.-P., von Salis Perch-Nielsen, K.,Biolzi, M., 1993. New calibration of Neogene radiolarian events in the NorthPacific. Mar. Micropaleontol. 21, 261e293.
Stephens, B.B., Keeling, R.F., 2000. The influence of Antarctic sea ice on glacialeinterglacial CO2 variations. Nature 404, 171e174.
Svendsen, J.I., Alexanderson, H., Astakhov, V.I., Demidov, I., Dowdeswell, J.A.,Funder, S., Gataullin, V., Henriksen, M., Hjort, C., Houmark-Nielsen, M.,Hubberten, H.W., Ingolfsson, O., Jacobsson, M., Kjaer, K.H., Larsen, E.,Lokrantz, H., Lunkka, J.P., Lysa, A., Mangerud, J., Matiouchkov, A., Murray, A.,Moller, P., Niessen, F., Nikolskaya, O., Polhyak, L., Saarnisto, M., Siegert, C.,Siegert, M., Spielhagen, R.F., Stein, R., 2004. Late Quaternary ice sheet history ofnorthern Europe. Quat. Sci. Rev. 23, 1229e1271.
Sverdrup, H.U., 1953. On conditions for the vernal blooming of phytoplankton.J. Cons. Int. Explor. Mer. 18, 287e295.
Takahashi, K., 1991. Radiolaria: flux, ecology, and taxonomy in the Pacific andAtlantic. In: Honjo, S. (Ed.), Ocean Biocoenosis Series No. 3. Woods HoleOceanographic Institution, Woods Hole, Massachusetts, p. 303.
Takahashi, K., 1995. Opal particle flux in the subarctic Pacific and Bering Sea andsidocoenosis preservation hypothesis. In: Tsunogai, S., Iseki, K., Koike, I., Oba, T.(Eds.), Global Fluxes of Carbon and its Related Substances in the Coastal SeaeOceaneAtmosphere System, Proceedings of the 1994 Sapporo IGBP Sympo-sium. M & J International, Yokohama, Japan, pp. 458e466.
Tanaka, S., Takahashi, K., 2005. Late Quaternary paleoceanographic changes in theBering Sea and the western subarctic Pacific based on radiolarian assemblages.Deep-Sea Res. Part II 52, 2131e2149.
Tanaka, S., Takahashi, K., 2008. Detailed vertical distribution of radiolarian assem-blage (0e3000 m, fifteen layers) in the central subarctic Pacific, June 2006.Mem. Fac. Sci. Kyushu Univ. Ser. D, Earth Planet. Sci. Lett. 32, 49e72.
Tauxe, L., Herbert, T., Shackleton, N.J., Kok, Y.S., 1996. Astronomical calibration of theMatuyama-Brunhes boundary: consequences for magnetic remanence acqui-sition in marine carbonates and the Asian loess sequences. Earth Planet. Sci.Lett. 140, 133e146.
Ternois, Y., Kawamura, K., Keigwin, L.D., Ohkouchi, N., Nakatsuka, T., 2001.A biomarker approach for assessing marine and terrigenous inputs to thesediments of Sea of Okhotsk for the last 27,000 years. Geochim. Cosmo-chem.Acta 65, 791e802.
Thompson, J., Wallace, H.E., Colley, S., Toole, J., 1990. Authigenic uranium in Atlanticsediments of the last glacial stageea diagenetic phenomenon. Earth Planet. Sci.Lett. 98, 222e232.
Tibbs, J.F., 1967. On some planktonic protozoa taken from the track of drift stationARLIS I, 1960e1961. Arctic Inst. N. Am. 20, 247e254.
Tsunogai, S., 2002. The western North Pacific playing a key role in global biogeo-chemical cycles. J. Oceanogr. 58, 245e257.
Van Cappellen, P., Qui, L., 1997. Biogenic silica dissolution in sediments of theSouthern Ocean. Deep-Sea Res. II 44, 1129e1149.
Vinogradov, M.E., 1970. Vertical Distribution of the Oceanic Zooplankton. Translatedfrom Russian Izdatel’stvo “Nauka” Moskva 1968 by Israel Program for ScientificTranslations, p. 339.
Wang, R., Xiao, W., Li, Q., Chen, R., 2006. Polycystine radiolarians in surface sedi-ments from the Bering Sea Green Belt area and their ecological implication forpaleoenvironmental reconstructions. Mar. Micropaleontol. 59, 135e152.
Washburn, A.L., 1980. Geocryology. John Wiley and Sons, New York, p. 406.Warren, B.A., 1983. Why is no deep water formed in the North Pacific? J. Marine Res.
41, 327e347.Welling, L.A., Pisias, N.G., Roelofs, A.K., 1992. Radiolarian Microfauna in the
Northern California Current System: Indicators of Multiple Processes Control-ling Productivity. In: Geol. Soc. London Spec. Pub. 64, pp. 177e195.
Winton, M., 1997. The effect of cold climate upon North Atlantic deep water for-mation in a simple oceaneatmosphere model. J. Clim. 10, 37e51.