gibberellin biosynthesis and signal transduction is essential for internode elongation in deepwater...
TRANSCRIPT

Original Article
Gibberellin biosynthesis and signal transduction isessential for internode elongation in deepwater rice
Madoka Ayano1, Takahiro Kani1, Mikiko Kojima2, Hitoshi Sakakibara2, Takuya Kitaoka1, Takeshi Kuroha1,
Rosalyn B. Angeles-Shim1, Hidemi Kitano1, Keisuke Nagai1 & Motoyuki Ashikari1
1Bioscience and Biotechnology Center, Nagoya University, Nagoya, Aichi 464-8601 and 2RIKEN Center for SustainableResource Science, Yokohama, Kanagawa 230-0045, Japan
ABSTRACT
Under flooded conditions, the leaves and internodes of deep-water rice can elongate above the water surface to captureoxygen and prevent drowning. Our previous studies showedthat three major quantitative trait loci (QTL) regulatedeepwater-dependent internode elongation in deepwaterrice. In this study, we investigated the age-dependent inter-node elongation in deepwater rice. We also investigated therelationship between deepwater-dependent internode elon-gation and the phytohormone gibberellin (GA) by physiologi-cal and genetic approach using a QTL pyramiding line(NIL-1 + 3 + 12). Deepwater rice did not show internodeelongation before the sixth leaf stage under deepwater condi-tion. Additionally, deepwater-dependent internode elonga-tion occurred on the sixth and seventh internodes during thesixth leaf stage. These results indicate that deepwater ricecould not start internode elongation until the sixth leaf stage.Ultra-performance liquid chromatography tandem mass-spectrometry (UPLC-MS/MS) method for the phytohormonecontents showed a deepwater-dependent GA1 and GA4 accu-mulation in deepwater rice. Additionally, a GA inhibitorabolished deepwater-dependent internode elongation indeepwater rice. On the contrary, GA feeding mimicked inter-node elongation under ordinary growth conditions. However,mutations in GA biosynthesis and signal transduction genesblocked deepwater-dependent internode elongation. Thesedata suggested that GA biosynthesis and signal transductionare essential for deepwater-dependent internode elongationin deepwater rice.
Key-word: gibberellin.
INTRODUCTION
Flooding occurs during the rainy seasons in South Asia andSouth East Asia. In general, the leaves and internodes ofgeneral cultivated rice do not elongate during the vegetativestage, and when subjected to deepwater (DW) conditions, theplant dies due to oxygen starvation (Fig. 1a,b). Deepwaterrice shows a similar gross morphology (i.e. leaves and inter-nodes are not elongated) as the general rice cultivar under
shallow water (SW) conditions. However, during flooding,the leaves and internodes of deepwater rice elongate to avoidoxygen deficiency under rising water levels (Fig. 1c,d). Theelongated internodes keep the top leaves above the watersurface and aerenchyma formation in the internode suppliesoxygen to the rest of the plant that is underwater. Internodescan elongate by up to 20–25 cm in a period of days (Catling1992).This characteristic has allowed deepwater rice to adaptto flood-prone areas.
Physiological and genetic analysis has shown that thephytohormones ethylene and gibberellin (GA) are involvedin internode elongation in deepwater rice. Under DW condi-tion, enhanced ethylene biosynthesis and low diffusion ofethylene in water result in ethylene accumulation in the plantbody. This ethylene accumulation triggers internode elonga-tion in deepwater rice (Sauter & Kende 1992; Hattori et al.2009).
The internode elongation ability of deepwater rice hasbeen characterized based upon three parameters: (1) totalelongated internode length (TIL); (2) number of elongatedinternodes (NEI); and (3) lowest elongated internode (LEI)(Inouye & Mogami 1980; Nemoto et al. 2004; Hattori et al.2007; Kawano et al. 2008). Among these, LEI is a key param-eter of internode elongation initiation in deepwater rice(Inouye & Mogami 1980) because LEI is based upon the leafstage that first shows internode elongation.
In this report, we explored the developmental stage inwhich the internode of deepwater rice could first elongate.Atpresent, it remains unknown whether internode elongationcan occur at any leaf stage during the vegetative phase andwhether elongation occurs in any of the internodes. In thisreport, we determined whether specific internodes in deep-water rice elongate under DW conditions. Previous studiesrevealed that three major QTLs regulate DW responses(Nemoto et al. 2004; Tang et al. 2005; Hattori et al. 2007;Kawano et al. 2008). In this study, we used a QTL pyramidingline (NIL-1 + 3 + 12) carrying C9285 (deepwater rice)genomic fragments possessing three major QTLs on chromo-somes 1, 3 and 12 in the T65 (general cultivated rice) geneticbackground to examine internode elongation patterns andinitiation under DW conditions.
GAs are a family of diterpenoids, and more than 100 GAshave been identified (MacMillan 2002). Among them, only aCorrespondence: M. Ashikari. e-mail: [email protected]
Plant, Cell and Environment (2014) 37, 2313–2324 doi: 10.1111/pce.12377
bs_bs_banner
© 2014 The Authors. Plant, Cell & Environment published by John Wiley & Sons Ltd.This is an open access article under the terms of the Creative Commons Attribution-NonCommercial-NoDerivs License, which permits use anddistribution in any medium, provided the original work is properly cited, the use is non-commercial and no modifications or adaptations aremade.
2313

small number, such as GA1 and GA4, are thought to bebioactive in plants. Bioactive GA regulates plant growth anddevelopment, including seed germination, stem elongation,leaf expansion, and flower and seed development(Yamaguchi 2008). GA-deficient and GA signal-deficientmutants (due to the lack of a GA biosynthetic gene and a GAsignal transduction gene) in rice show a dwarf phenotype(Ross et al. 1997; Ueguchi-Tanaka et al. 2000; Itoh et al.2002a,b; Sakamoto et al. 2004). Exogenous GA treatmentinduces internode elongation of deepwater rice (Raskin &Kende 1983; Suge 1985; Hattori et al. 2009), suggesting thatGA plays an important role in internode elongation.However, the mechanism of internode elongation in deepwa-ter rice remains poorly understood.
We also investigated the physiological and genetic interac-tions between GA and internode elongation in deepwaterrice. We treated NIL-1 + 3 + 12 with active GA3 and devel-oped mutant pyramiding (MP) lines possessing mutations inGA biosynthesis or signal transduction genes to determinewhether GA is essential for internode elongation during theDW response. Moreover, we measured bioactive GAs andprecursor concentrations and determined the expressionlevels of GA biosynthetic and signal transduction genes. Inthis report, we show the internode elongation pattern duringdifferent leaf stages under DW conditions, as well as thephysiological and genetic relationship between internodeelongation and GA in deepwater rice.
MATERIALS AND METHODS
Plant materials and growth conditions
Deepwater rice C9285 (Oryza sativa L.) was kindly providedby the National Institute of Genetics in Japan (http://www.shigen.nig.ac.jp/rice/oryzabase/), whereas T65 (O. sativaL. ssp. japonica) was a cultivar maintained at Nagoya Univer-sity.The QTL pyramiding line 1 + 3 + 12 (NIL-1 + 3 + 12) waspreviously produced at Nagoya University (Hattori et al.2008, 2009). Plant materials were grown in the greenhouse,growth chamber or paddy field in Nagoya University.
In all experiments, seeds were pre-germinated at 29 °C inwater for 3 d, and then sown in soil mixture (Mikawa baido)in plastic pots. At the 3–8 leaf stage (LS), seedlings wereprepared for each experiment (Supporting InformationFig. S1). In this study, complete submergence conditions wereapplied to simulate DW conditions for 6 h to 14 d (Support-ing Information Fig. S1). SW condition was simulated bywater levels less than 2 cm under the plant base (ordinarygrowth conditions). DW condition represented completesubmergence, which means that the entire plant body includ-ing the leaf tips remained underwater.
Submergence, GA and uniconazole treatments
All submergence, GA and GA biosynthesis inhibitor[uniconazole (Uni); inhibitor of ent-Kaurene oxidase] treat-ments were replicated in at least three independent biologi-cal experiments. The period of DW treatment varied. The
(f)
T65SW DW
NIL-1+3+12SW DW
C9285SW DW
T65 NIL-1+3+12 C9285SW DW SW DW SW DW
(e)
L2L3L4
L5L6
L7 L8
SW DW
T65
SW DW
SW
DW
L2
L4
L6
L3
L5
L7
L8
inte
rnod
e
node
node
C9285
SW DW
(a)
(c)
(b)
(d)
1
312
T65
C9285
Figure 1. Diagram and gross morphology of general cultivatedrice, NIL-1 + 3 + 12 and deepwater rice before and aftersubmergence. Diagram of the general cultivated rice T65 (a) anddeepwater rice C9285(c) under shallow water (SW) and deepwater(DW) conditions. Arrowhead represents water level. (b) Node andinternode components in general cultivated rice. Internodeelongation was not induced in general cultivated rice under DWconditions. (d) Node and internode components in deepwater rice.Internode elongation induced in deepwater rice under DWconditions. (e) Gross morphology showing increased plant heightbefore (SW) and after 7 d of DW treatment in T65, NIL-1 + 3 + 12and C9285. Graphical genotypes are shown below the plant. Greensolid rectangle indicates the T65 fragment and red solid rectangleindicates C9285 fragments. Yellow open rectangle shows the baseof the internode. (f) Magnified base of internode shown in (e).Longitudinal sections showing only the basal part of the plantcomposed of the nodes and internodes. Pith cavities (*) are presentin NIL-1 + 3 + 12 under DW and C9285 under SW and DWconditions. Scale bars: 5 mm in (f), 5 cm in (e).
2314 M. Ayano et al.
© 2014 The Authors. Plant, Cell & Environment published by John Wiley & Sons Ltd, 37, 2313–2324

12–24 h of DW treatment was performed for gene expres-sion analysis (Fig. 7a,b and Supporting Information Fig. S3);the 7 d treatment was carried out for gross morphologyanalysis (Fig. 1e,f), internode length analysis (Fig. 2) andgenetic analysis based upon MP lines (Fig. 4); and the 14 dtreatment was performed for GA treatment analysis(Fig. 4). A time course of submergence was performed forGA content analysis (Fig. 6). The GA feeding experiments(GA3 in Fig. 4) was carried out in the growth chamber byexogenous application of GA3 at 10−5 m on the plants for14 d. SW and DW treatments were carried out by placingthe plants in SW or DW conditions for 14 d. For theDW + Uni treatment, plants were pre-treated with 10−6 mUni for 3 d before submerging them for 14 d (Fig. 4).Experiments using MP line (see below) were performed ina greenhouse at Togo field of the Nagoya University. Otherexperiments were performed in a greenhouse and growthchamber of Nagoya University.
Genetic analysis
d18-dy [Waito-C, genetic background is ssp. Japonica(cultivar name is unknown)] contains a mutation in the riceGA biosynthesis gene, OsGA3ox2, which catalyses GA9 toGA4 and GA20 to GA1, and shows a weak dwarf phenotype(Itoh et al. 2002a; Sakamoto et al. 2004). gid1-7 (geneticbackground is ssp. japonica cv Nipponbare) and gid1-8[genetic background is ssp. japonica cv Taichung 65 (T65)]contain mutations in the rice GA receptor GID1. gid1-7and gid1-8 are weak alleles of gid1 mutants that produceflowers (Ueguchi-Tanaka et al. 2007b). slr1-d1 [geneticbackground is ssp. japonica cv Taichung 65 (T65] is a semi-dominant dwarf mutant that has mutation in the DELLAdomain of the SLR protein. A mutation in the DELLAdomain results in inefficient GA-dependent degradation ofthe SLR1 protein due to a reduced interaction with GID1(Asano et al. 2009). slr1-d1 mutants of rice exhibit a reduc-tion in the length of all internodes but are reported to befertile. gid2-2 and gid2-5 [genetic background is ssp.japonica cv Taichung 65 (T65)] contain mutations in theGA signalling factor, GID2, which is the F-box protein (E3ligase) that targets SLR protein (Sasaki et al. 2003;Ueguchi-Tanaka et al. 2007a). We obtained the MP linesthat have the 3 DW QTLs and GA homozygous mutationsfrom an F2 population derived from crosses between therice GA mutants (GA biosynthesis mutant and GA signaltransduction mutant) and NIL-1 + 3 + 12. The MP lineswere genotyped to confirm the presence of QTLs 1, 3, 12using the molecular markers used by Hattori et al. (2008)(Supporting Information Fig. S2). No background selectionwas carried out since all the GA mutants have a japonicabackground (T65 or Nipponbare except for d18-dy).The GA-related mutants were selected from the F1 and F2
generations based upon the dwarf phenotype and bysequence analysis. The MP lines were submerged for 7 d atthe 8 LS, after which plant height and TIL were measured.TIL was scored as the total length of internodes longer than5 mm.
Measurement of phytohormone contents
To measure endogenous phytohormone contents, ∼200 mg ofall aerial tissues, except the developed leaf blade, was col-lected before and after submergence. Each sample was col-lected into a 2.0 mL Master-Tube hard (Qiagen, TurnberryLane, Valencia, CA, USA), frozen and crushed with four ironbeads in liquid nitrogen. The concentration of endogenousGA was measured using UPLC-MS/MS (UPLC-Xevo TQ-S;Waters, Maple Street, Milford, MA, USA) at the Institute ofPlant Productivity Systems Research Group RIKEN Centerfor Sustainable Resource Science. Each compound wasmeasured as described previously (Kojima et al. 2009).
RNA isolation and expression analysis
For semi-quantitative real-time RT-PCR (semi-qRT-PCR)and qRT-PCR analyses, samples were collected from the 7 LSof the whole plants. Frozen samples were crushed and col-lected into liquid nitrogen. Total RNA was extracted withRNA solution using the RNeasy Plant mini kit (Qiagen).Total cDNA were obtained using TOYOBO-reverse tran-scription Ace (TOYOBO, Dojima Hama, Kita-ku, Osaka,Japan). For semi-qRT-PCR analysis, gene expression levelswere analysed using the Gene Amp PCR system 9700(Thermo Fisher Scientific Inc., Wyman Street, Waltham, MA,USA). The quantile normalization method was employedusing OsActin1 as an internal control. For qRT-PCR, analysisof the expression of the GA biosynthesis genes in T65 andC9285 at submergence was carried out as described previ-ously (Chomczynski & Sacchi 1987). For the analysis, gene-specific primer sets (Li et al. 2011) and SsoAdvanced SYBRGreen Supermix (Thermo Fisher Scientific Inc., WymanStreet, Waltham, MA, USA) were used. The refined geneexpression was analysed by Step One Plus (AppliedBiosystems). Sequencing of regions flanking the primers usedin the study confirmed that the primers can anneal to bothT65 and C9285 genomic and cDNA. The gene-specificprimers used for amplification in semi-qRT-PCR and qRT-PCR are described in Supporting Information Table S2.
RESULTS
Gross morphology and internode elongationpatterns in deepwater rice
We first compared the gross morphology among three ricelines,namely the general cultivated riceTaichung 65 (T65), theQTL pyramiding line (NIL-1 + 3 + 12) and deepwater ricevariety, C9285 under SW and DW conditions (Fig. 1e,f). Dif-ferences in the gross morphology of T65, NIL-1 + 3 + 12 andC9285 were not observed before and after submergence(Fig. 1e). However, there were clear differences in internodeelongation among the three lines (Fig. 1f). Internode(enclosed open yellow rectangle in Fig. 1e) elongation was notobserved in T65 under SW and DW conditions. On the con-trary,we observed significant internode elongation under DWcondition in NIL-1 + 3 + 12 and C9285. In C9285, internodeelongation with pith cavity formation was observed under SW
GA in deepwater rice 2315
© 2014 The Authors. Plant, Cell & Environment published by John Wiley & Sons Ltd, 37, 2313–2324

0.0
3.0
6.0
9.0
12.0
1 2 3 4 5 6 7 8 9 10
T65 NIL-1+3+12 C9285
0.0
0.2
0.4
0.6T65 NIL-1+3+12 C9285
0.0
0.2
0.4
0.6
0.0
0.2
0.4
0.6
0.0
2.0
4.0
6.0
8.0
10.0
0.0
0.2
0.4
0.6T65 NIL-1+3+12 C9285
0.0
2.0
4.0
6.0
8.0
10.0
0.0
10.0
20.0
30.0
40.0
50.0
60.0
1 2 3 4 5 6 7 8 9 10
SW DW
1 2 3 4 5 6 7
3LS
1 2 3 4 5 6 7
4LS
1 2 3 4 5 6 7
5LS
Inte
rnode l
ength
(cm
)
Internode Internode
(b)
L2
L4
L6
L8(a)
8th
7th
6th
Inte
rnode
1 2 3 4 5 6 7 8 9 10
6LS
1 2 3 4 5 6 7 8 9 10
7LS
1 2 3 4 5 6 7 8 9 10
8LS
Inte
rnode l
ength
(cm
)
Internode Internode
SW DW
0.0
0.2
0.4
0.6
1 2 3 4 5 6 7
T65 NIL-1+3+12 C9285
0.0
0.2
0.4
0.6
1 2 3 4 5 6 7
0.0
0.2
0.4
0.6
1 2 3 4 5 6 7
0.0
10.0
20.0
30.0
40.0
50.0
60.0
1 2 3 4 5 6 7 8 9 10
Figure 2. Leaf age-dependent internode elongation during submergence in T65, NIL-1 + 3 + 12 and C9285. (a) Diagram represents thelongitudinal section of C9285 to show the internode position. (b) Deepwater response of T65 (green rectangle), NIL-1 + 3 + 12 (bluerectangle) and C9285 (red rectangle) was represented based upon comparison of the internode length during the three- to eight-leaf stage(3–8 LS) of the plant under shallow water (SW) and 7 d of deepwater (DW) treatment. X-axis indicates the position of the internodecounted from the base to top, as shown in (a). Vertical bars indicate SD of the mean (n = 5–10) of three independent experiments.
2316 M. Ayano et al.
© 2014 The Authors. Plant, Cell & Environment published by John Wiley & Sons Ltd, 37, 2313–2324

conditions, but elongated internode length was enhanced sig-nificantly under DW conditions (Fig. 1f). Based upon theseresults, we examined the developmental stage at which inter-nodes of deepwater rice could elongate. To accomplish this,deepwater rice at various leaf stages was monitored for inter-node elongation under DW conditions and the specific inter-node that first elongated under deepwater was identified.
To evaluate deepwater rice traits, it is important tomeasure the TIL, NEI and LEI (Vergara & Mazaredo 1979;Inouye & Mogami 1980; Nemoto et al. 2004; Tang et al. 2005;Hattori et al. 2007; Kawano et al. 2008). However, thesemeasurements do not quantify each internode elongation.Here, we measured each internode length at different leafstages before and after submergence to evaluate the devel-opmental point (starting time) of the DW response. T65,NIL-1 + 3 + 12 and C9285 at the 3–8 LS were submerged for7 d (Supporting Information Fig. S1). We measured thelength of all internodes under SW and DW conditions. Usingthis approach, we determined when internode elongationwas initiated (Fig. 2a). Internode elongation was notobserved under any condition from the 3–8 LS in T65(Fig. 2b). The C9285 internode did not elongate (over 5 mmin length) from the 3–5 LS under any conditions, but the sixthand seventh internode showed elongation under DW condi-tion at 6 LS (Fig. 2b). At the 7 LS of C9285, we also observedslight internode elongation at the seventh internode underSW conditions (Fig. 2b). Under SW conditions for C9285, theseventh internode length was 2.5 cm at the 7 LS, and theseventh and eighth internode lengths were 7.9 and 2.1 cm,respectively, at the 8 LS. However, more significant internodeelongation was observed under DW conditions in C9285.Theseventh and eighth internode lengths were 18.9 and 17.6 cmat the 7 LS, and the seventh, eighth and ninth internodelengths were 15.6, 52.0 and 8.2 cm at the 8 LS under DWconditions in C9285, respectively. In NIL-1 + 3 + 12, theeighth internode elongated by 9.1 cm at the 7 LS, and theeighth and ninth internode elongated by 23.1 and 9.4 cm atthe 8 LS under DW conditions, respectively. Internode elon-gation patterns of C9285 under SW and DW conditions weresummarized in Fig. 3a–c.
C9285 showed internode elongation at the 6 LS under DWconditions (Fig. 3c), indicating that internode elongationoccurs at 6 LS in C9285 under DW conditions. Under SWconditions, C9285 internode elongated slightly from the 7 LS(Fig. 3b). However, the elongated length and number ofinternodes were higher under DW condition than SW condi-tion after the 6 LS. Under DW condition, C9285 elongatedtwo internodes at the 6 LS (the sixth and seventh internode)and 7 LS (seventh and eighth internode), and three inter-nodes at the 8 LS (seventh, eighth and ninth internodes).Thesixth internode of C9285 at the 7 LS and 8 LS did not elon-gate. Internode elongation under DW conditions showed agood correlation with LS. Deepwater rice could initiate elon-gation at the sixth internode of C9285 at the 6 LS, but not atlater leaf stages. This shows that the sixth internode has thepotential to elongate under DW condition at the 6 LS, butmay lose its internode elongation ability in later leaf stages.Thus, plants may use younger and upper internodes to
achieve rapid internode elongation during emergency condi-tions, such as rapid flooding.
Physiological relationship between GAs andinternode elongation
Previously, Raskin & Kende (1984) reported that 10 μm GA3
treatment of stem sections of deepwater rice induced inter-node elongation (Raskin & Kende 1984). Suge (1985) alsoreported that 0.1, 1.0 and 10.0 ppm GA3 treatment of stemsections of three deepwater rice varieties [the methodslightly modified by Raskin & Kende (1984)] induced inter-node elongation. Recently, exogenous ethylene for stemsection induced not only stem elongation but also genes of
(a)
3456
7
SW
DW
7
8
3456
7
345
6
345
3LS 4LS 5LS 6LS 7LS 8LS
3 34
345
7
9
L3 L4L5
L6
L7
L8
L2
L1
L3 L4
L1
L2
L3
L2
L4L3
L5
L1
L4
L6
L3
L5
L1
3
L3
34
L4L5
L6
L7
L8
3456
8
L4
L6L5
L7 L8
L4
L6
L3
L5
L7
3LS 4LS 5LS 6LS 7LS 8LS
3LS 4LS 5LS 6LS 7LS 8LS
Leaf stage
Gross morphology of C9285
3456
7
8
(b)
(c)
L3L2
L1L2
L1L2
3456
Figure 3. Elongation pattern of internode and leaves in C9285after submergence. (a–c) Diagram of the three- to eight-leaf stage(3–8 LS) of C9285 under shallow water (SW) and deepwater (DW)conditions, showing gross morphology of the plant (a) andanatomical structure of the node and internode (b, c). (a) Grossmorphology in the 3–8 LS of C9285 grown under SW conditions.(b, c) Position of internodes and leaves under SW (b) and DW(c) conditions. Only one leaf (marked in green L3-L8) representsthe leaf stage. LS; leaf stage. L; leaf.
GA in deepwater rice 2317
© 2014 The Authors. Plant, Cell & Environment published by John Wiley & Sons Ltd, 37, 2313–2324
GA metabolism enzymes (Choi 2007). This suggests that GAplays a key role in internode elongation in deepwater rice.However, these experiments were performed using stem sec-tions, so we investigated the role of GA in whole deepwaterrice plants.
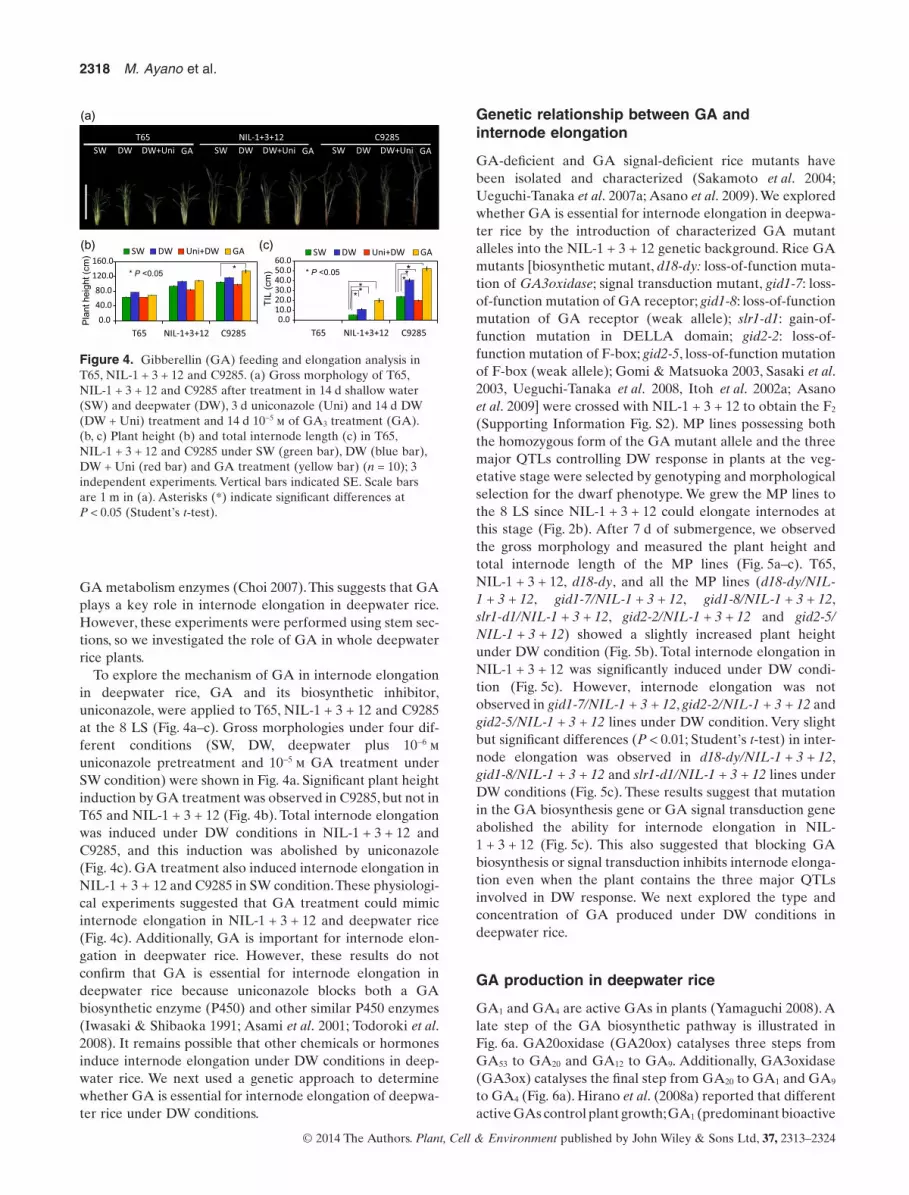
To explore the mechanism of GA in internode elongationin deepwater rice, GA and its biosynthetic inhibitor,uniconazole, were applied to T65, NIL-1 + 3 + 12 and C9285at the 8 LS (Fig. 4a–c). Gross morphologies under four dif-ferent conditions (SW, DW, deepwater plus 10−6 municonazole pretreatment and 10−5 m GA treatment underSW condition) were shown in Fig. 4a. Significant plant heightinduction by GA treatment was observed in C9285, but not inT65 and NIL-1 + 3 + 12 (Fig. 4b). Total internode elongationwas induced under DW conditions in NIL-1 + 3 + 12 andC9285, and this induction was abolished by uniconazole(Fig. 4c). GA treatment also induced internode elongation inNIL-1 + 3 + 12 and C9285 in SW condition.These physiologi-cal experiments suggested that GA treatment could mimicinternode elongation in NIL-1 + 3 + 12 and deepwater rice(Fig. 4c). Additionally, GA is important for internode elon-gation in deepwater rice. However, these results do notconfirm that GA is essential for internode elongation indeepwater rice because uniconazole blocks both a GAbiosynthetic enzyme (P450) and other similar P450 enzymes(Iwasaki & Shibaoka 1991; Asami et al. 2001; Todoroki et al.2008). It remains possible that other chemicals or hormonesinduce internode elongation under DW conditions in deep-water rice. We next used a genetic approach to determinewhether GA is essential for internode elongation of deepwa-ter rice under DW conditions.
Genetic relationship between GA andinternode elongation
GA-deficient and GA signal-deficient rice mutants havebeen isolated and characterized (Sakamoto et al. 2004;Ueguchi-Tanaka et al. 2007a; Asano et al. 2009). We exploredwhether GA is essential for internode elongation in deepwa-ter rice by the introduction of characterized GA mutantalleles into the NIL-1 + 3 + 12 genetic background. Rice GAmutants [biosynthetic mutant, d18-dy: loss-of-function muta-tion of GA3oxidase; signal transduction mutant, gid1-7: loss-of-function mutation of GA receptor; gid1-8: loss-of-functionmutation of GA receptor (weak allele); slr1-d1: gain-of-function mutation in DELLA domain; gid2-2: loss-of-function mutation of F-box; gid2-5, loss-of-function mutationof F-box (weak allele); Gomi & Matsuoka 2003, Sasaki et al.2003, Ueguchi-Tanaka et al. 2008, Itoh et al. 2002a; Asanoet al. 2009] were crossed with NIL-1 + 3 + 12 to obtain the F2
(Supporting Information Fig. S2). MP lines possessing boththe homozygous form of the GA mutant allele and the threemajor QTLs controlling DW response in plants at the veg-etative stage were selected by genotyping and morphologicalselection for the dwarf phenotype. We grew the MP lines tothe 8 LS since NIL-1 + 3 + 12 could elongate internodes atthis stage (Fig. 2b). After 7 d of submergence, we observedthe gross morphology and measured the plant height andtotal internode length of the MP lines (Fig. 5a–c). T65,NIL-1 + 3 + 12, d18-dy, and all the MP lines (d18-dy/NIL-1 + 3 + 12, gid1-7/NIL-1 + 3 + 12, gid1-8/NIL-1 + 3 + 12,slr1-d1/NIL-1 + 3 + 12, gid2-2/NIL-1 + 3 + 12 and gid2-5/NIL-1 + 3 + 12) showed a slightly increased plant heightunder DW condition (Fig. 5b). Total internode elongation inNIL-1 + 3 + 12 was significantly induced under DW condi-tion (Fig. 5c). However, internode elongation was notobserved in gid1-7/NIL-1 + 3 + 12, gid2-2/NIL-1 + 3 + 12 andgid2-5/NIL-1 + 3 + 12 lines under DW condition. Very slightbut significant differences (P < 0.01; Student’s t-test) in inter-node elongation was observed in d18-dy/NIL-1 + 3 + 12,gid1-8/NIL-1 + 3 + 12 and slr1-d1/NIL-1 + 3 + 12 lines underDW conditions (Fig. 5c). These results suggest that mutationin the GA biosynthesis gene or GA signal transduction geneabolished the ability for internode elongation in NIL-1 + 3 + 12 (Fig. 5c). This also suggested that blocking GAbiosynthesis or signal transduction inhibits internode elonga-tion even when the plant contains the three major QTLsinvolved in DW response. We next explored the type andconcentration of GA produced under DW conditions indeepwater rice.
GA production in deepwater rice
GA1 and GA4 are active GAs in plants (Yamaguchi 2008). Alate step of the GA biosynthetic pathway is illustrated inFig. 6a. GA20oxidase (GA20ox) catalyses three steps fromGA53 to GA20 and GA12 to GA9. Additionally, GA3oxidase(GA3ox) catalyses the final step from GA20 to GA1 and GA9
to GA4 (Fig. 6a). Hirano et al. (2008a) reported that differentactive GAs control plant growth;GA1 (predominant bioactive
TIL
(cm
)
*
*
(a)
(b) (c)
Pla
nt h
eigh
t (cm
)
* P <0.05 *
**
**
* P <0.05
Figure 4. Gibberellin (GA) feeding and elongation analysis inT65, NIL-1 + 3 + 12 and C9285. (a) Gross morphology of T65,NIL-1 + 3 + 12 and C9285 after treatment in 14 d shallow water(SW) and deepwater (DW), 3 d uniconazole (Uni) and 14 d DW(DW + Uni) treatment and 14 d 10−5 m of GA3 treatment (GA).(b, c) Plant height (b) and total internode length (c) in T65,NIL-1 + 3 + 12 and C9285 under SW (green bar), DW (blue bar),DW + Uni (red bar) and GA treatment (yellow bar) (n = 10); 3independent experiments. Vertical bars indicated SE. Scale barsare 1 m in (a). Asterisks (*) indicate significant differences atP < 0.05 (Student’s t-test).
2318 M. Ayano et al.
© 2014 The Authors. Plant, Cell & Environment published by John Wiley & Sons Ltd, 37, 2313–2324

Figure 5. Gross morphology and deepwater response of mutantsand mutant pyramiding lines after 7 d of deepwater treatment.(a) Gross morphology of GA biosynthesis (d18-dy), GA signaltransduction (gid1-7, gid1-8), DELLA (slr1-dy), and F-box (gid2-2,gid2-5) mutants and mutant pyramiding (MP) lines after 7 d ofshallow water (SW) and deepwater (DW) treatments. (b, c) Plantheight (b) and total internode length (c) after 7 d of SW and DWconditions were determined (n = 8–10). Vertical bars indicate SE.Asterisks represents significant differences; *P < 0.05 and**P < 0.01 (Student’s t-test). ND in (c) means not detected. Scalebar is 1 m in (a).
(b)
(c)
(d)
(a)
0.0
40.0
80.0
120.0
160.0
200.0
240.0
0 12 24 36 48 60
T65 NIL-1+3+12 C9285
0.0
4.0
8.0
12.0
16.0
20.0
0 12 24 36 48 60
T65 NIL-1+3+12 C9285
0.0
4.0
8.0
12.0
0 12 24 36 48 60
T65 NIL-1+3+12 C9285
0.0
5.0
10.0
15.0
20.0
0 12 24 36 48 60
T65 NIL-1+3+12 C9285
0.0
15.0
30.0
45.0
0 12 24 36 48 60
T65 NIL-1+3+12 C9285
0.0
2.0
4.0
6.0
8.0
10.0
12.0
0 12 24 36 48 60
T65 NIL-1+3+12 C9285
GA12GA53
GA9GA20
GA1 GA4
GA12-aldehyde
KAO
GA20ox
GA3ox
GA13ox
(e)
(f)
(g)
Time of DW treatment (h) Time of DW treatment (h)
Time of DW treatment (h)Time of DW treatment (h)
Time of DW treatment (h) Time of DW treatment (h)
pm
ol g
−1 F
W
pm
ol g
−1 F
Wp
mo
l g
−1 F
W
pm
ol g
−1 F
Wp
mo
l g
−1 F
W
pm
ol g
−1 F
W
GA53 GA12
GA9
GA4
GA20
GA1
Figure 6. Measurement of endogenous hormone concentrationsunder deepwater treatment at 7 LS in T65, NIL-1 + 3 + 12 andC9285. (a) Diagram represents the late step of GA1 and GA4
biosynthesis pathway to show enzymes and parallel relationships.(b–g) Endogenous levels of gibberellin (GA1) (d) and itsprecursors GA53 (b) and GA20 (c), as well as GA4 (g) and itsprecursors GA12 (e) and GA9 (f), were analysed after 0, 12, 24 and48 h of deepwater treatment in T65 (green closed circle),NIL-1 + 3 + 12 (blue closed triangle) and C9285 (red closedrectangle) using UPLC-MS/MS (n = 4–7). Vertical bars indicateSD.
GA in deepwater rice 2319
© 2014 The Authors. Plant, Cell & Environment published by John Wiley & Sons Ltd, 37, 2313–2324

molecule) controls vegetative organ growth, whereas GA4
(dominant bioactive molecule) controls the reproductive riceorgans (Hirano et al. 2008a).Kojima et al. (2009) reported thatendogenous GA4 accumulates at the flower and paniclebranch and not at the internode. It has also been reported thataccumulation of GA4 induces an uppermost internode elon-gation during the heading stage of general cultivated rice, butnot in vegetative organs (Magome et al. 2013). Hattori et al.(2009) reported that endogenous GA1 accumulates at the baseof the stem in deepwater rice C9285 after submergence.However, the type and concentration of GA that inducedinternode elongation in deepwater rice at submerged condi-tion are still unknown.
To examine changes in the GA level and the types of GAsthat accumulate after submergence in deepwater rice, weemployed mass spectrometry (MS) analysis. Both GA1 andGA4 accumulated in the aerial parts except in the developedleaves of NIL-1 + 3 + 12 and C9285 at after submergence(Fig. 6d,g and Supporting Information Table S1). Minor GA1
and GA4 accumulation was observed in T65, but GA produc-tion in C9285 was higher than in T65 (Fig. 6d,g and Support-ing Information Table S1). The maximum GA1 and GA4
levels in C9285 after 24 h of submergence were similar(7–8 pmol g−1 FW). Additionally, time-dependent accumula-tion patterns of GA1 and GA4 were similar in C9285(Fig. 6d,g).The time-dependent production of GA20 and GA9
(precursors of GA1 and GA4) were also similar under DWconditions in C9285 (Fig. 6c,f).These results suggest that GA1
and GA4 production under DW conditions is C9285-specific,and both GA1 and GA4 play a role in internode elongation indeepwater rice.
The time-dependent production of GA1 and GA4 and theirprecursors GA20 and GA9 in NIL-1 + 3 + 12 were similar toC9285 (Fig. 6c,f,d,g). This suggests that internode elongationin NIL-1 + 3 + 12 and C9285 is due to production of activeGA1 and GA4. It also suggests that the accumulated GA1 andGA4 is caused by at least one of the QTL loci (originatingfrom C9285), since NIL-1 + 3 + 12 and C9285 shared threechromosome fragments around the three major QTLs con-trolling the DW response (Fig. 1e). Based upon these results,we explored why active GA1 and GA4 accumulated underDW conditions in deepwater rice. To accomplish this, weexamined the expression of GA biosynthetic and signaltransduction genes in deepwater rice.
The expression profile of GA biosynthetic genesin deepwater rice
Bioactive GAs are synthesized through sequential stepsby specific enzymes from trans-geranylgeranyl diphosphate(GGDP) (Yamaguchi 2008). We examined the steady-state levels of transcripts from GA biosynthesis genes(OsCPS1, OsCPS2, OsKS2, OsKS5, OsKO2, OsKAO,Os13ox, OsGA20ox1, OsGA20ox2, OsGA20ox3, OsGA3ox1,OsGA3ox2) (Magome et al. 2013), GA metabolic genes(OsGA2ox1, OsGA2ox3, OsGA2ox6, EUI, EUIL4) (Zhuet al. 2006) and signal transduction genes (OsGID1, OsGID2,OsSPY, OsSEC, OsGAMYB) (Tsuji et al. 2006; Ueguchi-
Tanaka et al. 2007a; Hirano et al. 2008b; Phanchaisri et al.2012) in whole plants of T65 and C9285 before and aftersubmergence (Fig. 7 and Supporting Information Fig. S3).
Comparison of most of GA-associated gene transcriptlevels revealed no significant differences in whole T65 andC9285 plants (Fig. 7a). However, differences in the transcript
0.0
0.5
1.0
1.5
2.0
2.5
3.0
0 12 24
T65 C9285
(a)
(b)
Rela
tive
expre
ssio
n level of
OsG
A20
ox2
Time of DW treatment (h)
0 h
T65
24 h12 h 0 h
C9285
24 h12 hgDNA gDNA
OsCPS1
OsCPS2
OsKAO
OsKS2
OsKS5
OsKO2
OsGA20ox2
OsGA20ox1
OsGA3ox1
OsGA20ox3
OsGA3ox2
OsActin1
OsActin1
Cycle
× 35
× 35
× 35
× 35
× 35
× 30
× 30
× 30
× 30
× 30
× 35
× 30
× 35
Os13ox × 35
Figure 7. Semi-qRT-PCR and qRT-PCR analysis of GAbiosynthesis gene expression analysis. (a) mRNA was obtainedfrom the 7 LS of T65 and C9285 after 0, 12 and 24 h ofsubmergence. PCR was performed for 30 and 35 cycles for GAbiosynthesis and signal transduction genes and also 30 and 35cycles for OsActin1. (b) qRT-PCR was performed using theSsoAdvanced SYBR Green Supermix with Step One Plus PCRsystem. Representation of the relative expression levels ofGA20ox2 gene was normalized to the expression level of ubiquitin.The relative value of OsGA20ox2 expression in T65 at 0 h was 1.Vertical bars indicate SD of the mean of four replicates.
2320 M. Ayano et al.
© 2014 The Authors. Plant, Cell & Environment published by John Wiley & Sons Ltd, 37, 2313–2324

accumulation of the GA biosynthesis gene OsGA20ox2 wereobserved between T65 and C9285. The expression ofOsGA20ox2 in C9285 was highly up-regulated and prolongedafter submergence. However, the transcript accumulation ofOsGA20ox2 in T65 is only slightly up-regulated (Fig. 7a).qRT-PCR analysis showed a higher transcript accumulation ofOsGA20ox2 in C9285 compared with T65 in DW condition(Fig. 7b). OsGA20ox2 catalyses GA53 to GA20 and GA12 toGA9 (Fig. 6a). The amount of GA20 and GA9 increased aftersubmergence in C9285 (Fig. 6c,f), which corresponded to theinduction of GA20ox gene expression in C9285 (Fig. 7b).Expression of GA metabolic genes and signal transductiongenes were also observed, although gene transcript accumu-lation in T65 and C9285 did not differ when subjected to SWand submerged conditions (Supporting Information Fig. S3).Internode elongation in C9285 under DW conditions may becaused by GA1 and GA4 accumulation after induction of theGA20ox gene in plants under deepwater.
DISCUSSION
To characterize the internode elongation mechanism in deep-water rice, we determined the developmental stage at whichinternode elongation occurs in deepwater rice under DWconditions. Internode elongation under DW conditions wasfirst detected at the sixth internode at 6 LS and not beforethat (Figs 2 & 3). Based upon this result, an anatomical phasechange in internodes or GA biosynthesis (or GA-associatedprocesses) may occur between the 5 and 6 LS in C9285. Forexample, intercalary meristem (IM) located in the nodescould be activated, which can induce cell division and subse-quently produce new cells. IM activation, which induces celldivision and expansion, is a driving force in internode elon-gation.We propose three hypotheses to describe the relation-ship between IM and internode elongation (Fig. 8a–d). Forthese results, the longitudinal section was anatomically illus-trated (Fig. 8a).
The first hypothesis is that C9285 does not contain IMsbefore the 6 LS, but produces IM at the 6 LS under naturalconditions which can then be activated by DW condition(Fig. 8b). The second hypothesis is that C9285 does notcontain IMs at the nodes under SW conditions, but submer-gence induces IM production at the 6 LS (Fig. 8c). The lasthypothesis is that C9285 establishes IMs at every node duringan early leaf stage, but the IM remains dormant until the 6 LSuntil such time that it is activated by submergence (Fig. 8d).
Ikeda et al. (2001) reported that the GA constitutive signaltransduction mutant (slr1-1: Nipponbare background)induced internode elongation from the third node.This singlerecessive mutant contains a single base deletion in the SLR1gene causing a loss of function of the gene (Ikeda et al. 2001).This report suggests that IM could be established at least inthe third node in T65. So based upon the result, hypothesis 1,in which deepwater rice establishes IM at the 6LS, is unlikelysince deepwater rice requires rapid internode elongation toavoid flooding. Additionally, if the deepwater rice initiatesIM after submergence (hypothesis 2), there will be a delaybefore internode elongation, which would put the plant at a
high risk of drowning. Thus, hypothesis 2 is also unlikely. Inthe context of emergency conditions, hypothesis 3, in whichdeepwater rice establishes the IM at every node in an earlyleaf stage but remains dormant until submergence, is themost likely scenario.
We next explored how IM becomes established and acti-vated at the sixth internode in deepwater rice. Plants becomehypoxic under DW conditions, and it is known that ethylenegas accumulates in plants underwater (Kende et al. 1998;Hattori et al. 2009). Hypoxia, accumulation of ethylene andGA, or changes in the role of GA during submergence inC9285 may establish IM or activate dormant IM at the sixthnode. In this study, GA was shown to be essential for inter-node elongation in deepwater rice. We also showed that GAaccumulated in response to induction of the GA20ox gene.GA accumulation during submergence in C9285 may alsoactivate dormant IM at the sixth node. Cell division andelongation would then induce internode elongation. Basedupon these results, hypothesis 3 seems most likely, althoughhypotheses 1 and 2 remain possible. We do not clearly
(b) (c)
(a)DWSW
Dormant or inactiveActive
IM status
C9285 may produce IM at the 6 leaf stage in nature. This IM respond at submergence.
C9285 may not have IMs at every nodes in shallow water condition, but submergence may induce establishment of IM at the 6 leaf stage.
C9285 establishes IMs at every node from the early leaf stage, but IM may be dormant and it is not activated until the 6 leaf stage. Submergence mayactivate IM at 6 leaf stage.
(d)
Longitudinal section
Figure 8. Schematic diagram representing three hypotheses ofthe IM activation mechanism. (a) The image shows a longitudinalsection of deepwater rice under shallow water (SW) anddeepwater (DW) conditions. (b) Image represents hypothesis 1,where IM formation of deepwater rice started at the sixthinternode in the 6 LS. (c) Image represents hypothesis 2, where IMformation initiated upon submergence. (d) Image representshypothesis 3, where IM formed at every internode but remaineddormant until submergence. The old internode disappeared inresponse to DW. Active (elongation ability) IM is indicated inpink. The dormant or non-elongated (absent elongation ability) IMis indicated in orange.
GA in deepwater rice 2321
© 2014 The Authors. Plant, Cell & Environment published by John Wiley & Sons Ltd, 37, 2313–2324

understand why deepwater rice can initiate internode elon-gation at the 6 LS. Additionally, the genesis of IM in deep-water rice remains poorly understood.Thus, the identificationof IM specific markers and the study of IM in deepwater riceare required.
In the previous study, the QTL on chromosome 3 isdetected based upon the LEI (Nemoto et al. 2004; Hattoriet al. 2008; Kawano et al. 2008). The QTL on chromosome 3determines the leaf stage that can initiate internode elonga-tion. At this time, the QTL that regulates IM at the 6 LS hasnot been identified, but the QTL on chromosome 3 may beinvolved in activating or repressing IM.
C9285 showed slight internode elongation even under Swcondition (Figs 2 & 3). Under SW condition, C9285 wouldstill produce ethylene or GA, which is induced by physicaltouch or wind. IM may have a high sensitivity to ethylene orGA in C9285, and could respond to low concentrations ofethylene or GA to induce internode elongation.The majorityof elongated internodes under DW conditions correlatedwith the leaf stage (Figs 2 & 3). The sixth internode washighly elongated at the 6 LS, and the seventh internode washighly elongated at the 7 LS. However, at later leaf stages, thesixth and seventh internode of deepwater rice can no longerelongate. We observed that deepwater rice uses (activates)younger IMs for rapid internode elongation. Additionally,the internode elongation ability moves up the internodeaccording to leaf stage, and old nodes may lack meristematicactivity. The use of younger IMs for rapid internode elonga-tion may be important to survive DW conditions.
UPLC-MS/MS analysis revealed that GA1 and GA4 accu-mulate not only in the leaf but also in the internode underDW conditions in C9285, but not in T65 (Fig. 6). Our analysisrevealed that both GAs (GA1 or GA4) accumulate in theaerial parts including the internode, establishing the role ofGAs in internode elongation. In case of GA1 production,GA53 in T65 is higher than that in C9285 under DW condi-tion. On the contrary, the GA20 amount in C9285 is higherthan that in T65 in deepwater (Fig. 6b,c). The higher expres-sion of GA20ox2 in C9285 compared with T65 under DWcondition may account for the observed GA53 accumulationin T65 and GA20 accumulation in C9285 (Fig. 7). The higherexpression of GA20ox2 was consistent with the elevation ofits product, GA1. GA9, GA4 and GA1 were detected in C9285by MS (Fig. 6) but not GA12, probably because that theamount of GA12 produced was too low for detection byUPLC-MS/MS. Both GA1 and GA4 biosynthesis pathway isregulated by 13oxidase activity (Fig. 6a). However, theexpression of GA13ox is not different in C9285 before andafter DW condition. This stable expression may lead to bothGA1 and GA4 accumulation. The expression of the GAmetabolite gene, GA2ox6, was reduced in C9285 at 12 h. In aprevious report, OsGA2ox6 has been identified to contributeto the inhibition of the vegetative stage of the plant, with itsbasal expression in T65 being very low (Lo et al. 2008). Thisindicates that metabolism of bioactive GAs (GA1 and GA4)in C9285 was also inhibited, thereby inducing internode elon-gation. In this study, the expression of GA3ox1 and GA3ox2,which are important in the final steps of GA biosynthesis, did
not change before and after submergence in C9285 (Fig. 7a).These data suggest that the up-regulation of the GA20oxgene is an important step in generating active GA1 and GA4
in C9285.An eightfold increase in active GA accumulation issufficient to enhance internode elongation. The currentresults on the transcript levels of GA metabolism genes werebased upon the semi-qRT-PCR analysis and therefore arestill preliminary. In the future, mRNA-sequence analysismight be able to shed additional light on the regulation of keygenes in GA metabolism. For example, recent report fromvan Veen et al. (2014) revealed that two species of the eudicotgenus Rumex, which inhabit different elevations of the RhineRiver floodplain, display opposite submergence responsestrategies similar to what is observed in rice. Petiole elonga-tion is exhibited in Rumex palustris, which endures prolongeddeep floods, whereas this elongation is inhibited in Rumexacetosa, which only occasionally experiences short-termflooding. Unlike the scenario in rice, the petiole elongationwas not evidently regulated by Group VII Ethylene responsefactors as in rice. Rather, R. palustris induced a distinct genetranscript profile as compared with rice that intersects withshade avoidance mediated petiole elongation. Interestingly,strong induction of the EIN3 BINDING F-BOX (EBF)OMCL gene family during submergence was observed onlyin R. palustris. The putative ortholog of rice is likely involvedin ethylene-induced growth stimulation by preventing nega-tive regulation of GA synthesis by ethylene (Kim et al. 2012;van Veen et al. 2013, 2014). Therefore, ethylene-regulatedelongation may have diverged between eudicots andmonocots. In monocots, IM is common; therefore, elongationof aerial part by cell elongation in IM had been reported inAvena (Kaufman 1965; Kaufman et al. 1965), Cyperus (Fisher1970), Eleocharis (Evans 1965, 1969) and Zea mays (Sharman1942). GA-induced internode elongation was also reportedin Avena (Kaufman 1965). Recently, we detected thatBrachypodium (Brachypodium distachyon) had also pos-sessed GA responses and internode elongation (data notshown).Thereby, GA regulated elongation mechanism is alsodiverged among glass plants we thought. Future mRNA-sequence analysis of IMs of deepwater rice and other grassspecies may help to clarify ethylene- and GA-regulated elon-gation mechanisms.
In Fig. 4c, internode length in NIL-1 + 3 + 12 and C9285 inthe presence of 10−5 m GA3 increased under DW conditions.This may be explained by the fact that 10−5 M GA3 is at ahigher concentration than GA1 and GA4 under DW condi-tions in NIL-1 + 3 + 12 and C9285. The internode length ofC9285 is greater than that of NIL-1 + 3 + 12 under DW con-ditions (Fig. 2). NIL-1 + 3 + 12 carry the three major QTLsthat regulate internode elongation of C9285 in a T65 geneticbackground. However, an additional QTL is required toregulate internode elongation of DW condition in C9285.Nagai et al. (2012) detected two minor QTLs associated withinternode elongation in C9285 (Nagai et al. 2012). Thus, theidentification of other QTLs involved in internode elonga-tion in deepwater rice is required.
Enhancement of leaf length by GA was similar in T65,NIL-1 + 3 + 12 and C9285 (Fig. 4b). GA treatment induced
2322 M. Ayano et al.
© 2014 The Authors. Plant, Cell & Environment published by John Wiley & Sons Ltd, 37, 2313–2324

internode elongation in NIL-1 + 3 + 12 and C9285, but not inT65 (Fig. 4c). Both leaf and internode in NIL-1 + 3 + 12 andC9285 responded to GA. Leaves in T65 responded to GA,but internodes in T65 did not. These results suggest thatorgan-specific GA sensitivity differed between leaf (leafsheath) and internode in T65, or that GA transport to the leafand internode may differ between T65 and C9285.
Internode elongation was induced in NIL-1 + 3 + 12 byGA, suggesting that the QTL encodes genes responsible forGA sensitivity or transport. These could be important differ-ences between non-deepwater rice and deepwater rice.Cloning and functional analysis of the QTLs could addressthe differences in leaf expansion and internode elongation byGA in T65.
ACKNOWLEDGMENTS
We thank Dr Nori Kurata at the National Institute of Genet-ics (Mishima, Japan) and the NBRP Project for providingdeepwater rice (C9285). This study was supported by Grant-in-Aid 22119001 (to M.A.) from the Ministry of Education,Culture, Sports, and Science and by grants from the Ministryof Agriculture, Forestry and Fisheries of Japan (Genomics forAgricultural Innovation, QTL-4002).
REFERENCES
Asami T., Mizutani M., Fujioka S., Goda H., Min Y.K., Shimada Y. & YoshidaS. (2001) Selective interaction of triazole derivatives with DWF4, acytochrome P450 monooxygenase of the brassinosteroid biosyntheticpathway, correlates with brassinosteroid deficiency in planta. The Journal ofBiological Chemistry 276, 25687–25691.
Asano K., Hirano K., Ueguchi-Tanaka M., Angeles-Shim R.B., Komura T.,Satoh H. & Ashikari M. (2009) Isolation and characterization of dominantdwarf mutants, Slr1-d, in rice. Molecular Genetics and Genomics 281, 223–231.
Catling H.D. (1992) Rice in Deep Water. Macmillan [for the] International RiceResearch Institute, London.
Choi D. (2007) Ethylene-induced stem growth of deepwater rice is correlatedwith expression of gibberellin- and abscisic acid-biosynthetic genes. Journalof Plant Biology 50, 595–599.
Chomczynski P. & Sacchi N. (1987) Single-step method of RNA isolation byacid guanidinium thiocyanate-phenol-chloroform extraction. Analytical Bio-chemistry 162, 156–159.
Evans P.S. (1965) Intercalary growth in aerial shoot of Eleocharis acuta R BrProdr. I. Structure of growing zone. Annals of Botany 29, 205–217.
Evans P.S. (1969) Intercalary growth in the aerial shoot of Eleocharis acuta R.BRII. Development of the main internode. New Zealand Journal of Botany7, 36–42.
Fisher J.B. (1970) Control of internodal intercalary meristem of Cyperusalternifolius. American Journal of Botany 57, 1017–1026.
Gomi K. & Matsuoka M. (2003) Gibberellin signalling pathway. CurrentOpinion in Plant Biology 6, 489–493.
Hattori Y., Miura K., Asano K., Yamamoto E., Mori H., Kitano H., . . . AshikariM. (2007) A major QTL confers rapid internode elongation in response towater rise in deepwater rice. Breeding Science 57, 305–314.
Hattori Y., Nagai K., Furukawa S., Song X.J., Kawano R., Sakakibara H. &Ashikari M. (2009) The ethylene response factors SNORKEL1 andSNORKEL2 allow rice to adapt to deep water. Nature 460, 1026–1030.
Hattori Y.N., Nagai K., Mori H., Kitano H., Matsuoka M. & Ashikari M. (2008)Mapping of three QTLs that regulate internode elongation in deepwaterrice. Breeding Science 58, 39–46.
Hirano K., Aya K., Hobo T., Sakakibara H., Kojima M., Shim R.A. & MatsuokaM. (2008a) Comprehensive transcriptome analysis of phytohormonebiosynthesis and signaling genes in microspore/pollen and tapetum of rice.Plant & Cell Physiology 49, 1429–1450.
Hirano K., Ueguchi-Tanaka M. & Matsuoka M. (2008b) GID1-mediatedgibberellin signaling in plants. Trends in Plant Science 13, 192–199.
Ikeda A., Ueguchi-Tanaka M., Sonoda Y., Kitano H., Koshioka M., FutsuharaY. & Yamaguchi J. (2001) Slender rice, a constitutive gibberellin responsemutant, is caused by a null mutation of the SLR1 gene, an ortholog of theheight-regulating gene GAI/RGA/RHT/D8. The Plant Cell 13, 999–1010.
Inouye J. & Mogami Y. (1980) Position of the lowest elongated internode offloating rices originated in different countries. Japanese Journal of TropicalAgriculture 24, 13–17.
Itoh H., Ueguchi-Tanaka M., Sakamoto T., Kayano T., Tanaka H., Ashikari M.& Matsuoka M. (2002a) Modification of rice plant height by suppressing theheight-controlling gene, D18, in rice. Breeding Science 52, 215–218.
Itoh H., Ueguchi-Tanaka M., Sato Y., Ashikari M. & Matsuoka M. (2002b) Thegibberellin signaling pathway is regulated by the appearance and disappear-ance of SLENDER RICE1 in nuclei. The Plant Cell 14, 57–70.
Iwasaki T. & Shibaoka H. (1991) Brassinosteroids act as regulators oftracheary-element differentiation in isolated Zinnia Mesophyll-Cells. Plant& Cell Physiology 32, 1007–1014.
Kaufman P.B. (1965) Effects of growth substances on intercalary growth andcellular differentiation in developing internodes of Avena sativa. 2. Effects ofgibberellic acid. Physiologia Plantarum 18, 703–724.
Kaufman P.B., Cassell S.J. & Adams P.A. (1965) On nature of intercalarygrowth and cellular differentiation in internodes of Avena sativa. BotanicalGazette 126, 1–13.
Kawano R., Doi K., Yasui H., Mochizuki T. & Yoshimura A. (2008) Mappingof QTLs for floating ability in rice. Breeding Science 58, 47–53.
Kende H., van der Knaap E. & Cho H.T. (1998) Deepwater rice: a model plantto study stem elongation. Plant Physiology 118, 1105–1110.
Kim J., Wilson R.L., Case J.B. & Binder B.M. (2012) A comparative study ofethylene growth response kinetics in eudicots and monocots reveals a rolefor gibberellin in growth inhibition and recovery. Plant Physiology 160,1567–1580.
Kojima M., Kamada-Nobusada T., Komatsu H., Takei K., Kuroha T., MizutaniM., & Sakakibara H. (2009) Highly sensitive and high-throughput analysis ofplant hormones using MS-probe modification and liquid chromatography-tandem mass spectrometry: an application for hormone profiling in Oryzasativa. Plant & Cell Physiology 50, 1201–1214.
Li J., Jiang J., Qian Q., Xu Y., Zhang C., Xiao J., . . . Chong K. (2011) Mutationof rice BC12/GDD1, which encodes a kinesin-like protein that binds to aGA biosynthesis gene promoter, leads to dwarfism with impaired cell elon-gation. The Plant Cell 23, 628–640.
Lo S.F., Yang S.Y., Chen K.T., Hsing Y.I., Zeevaart J.A., Chen L.J. & Yu S.M.(2008) A novel class of gibberellin 2-oxidases control semidwarfism,tillering, and root development in rice. The Plant Cell 20, 2603–2618.
MacMillan J. (2002) Occurrence of gibberellins in vascular plants, fungi, andbacteria (vol 20, pg 387, 2002). Journal of Plant Growth Regulation 21,242–243.
Magome H., Nomura T., Hanada A., Takeda-Kamiya N., Ohnishi T., Shinma Y.& Yamaguchi S. (2013) CYP714B1 and CYP714B2 encode gibberellin13-oxidases that reduce gibberellin activity in rice. Proceedings of theNational Academy of Sciences of the United States of America 110, 1947–1952.
Nagai K., Kuroha T., Ayano M., Kurokawa Y., Angeles-Shim R.B., Shim J.H. &Ashikari M. (2012) Two novel QTLs regulate internode elongation in deep-water rice during the early vegetative stage. Breeding Science 62, 178–185.
Nemoto K., Ukai Y., Tang D.Q., Kasai Y. & Morita M. (2004) Inheritance ofearly elongation ability in floating rice revealed by diallel and QTL analyses.Theoretical and Applied Genetics 109, 42–47.
Phanchaisri B., Samsang N., Yu L., Singkarat S. & Anuntalabhochai S. (2012)Expression of OsSPY and 14-3-3 genes involved in plant height variations ofion-beam-induced KDML 105 rice mutants. Mutation Research 734, 56–61.
Raskin I. & Kende H. (1983) How does deep-water rice solve its aerationproblem. Plant Physiology 72, 447–454.
Raskin I. & Kende H. (1984) Role of gibberellin in the growth-response ofsubmerged deep-water rice. Plant Physiology 76, 947–950.
Ross J.J., Murfet I.C. & Reid J.B. (1997) Gibberellin mutants. PhysiolPlantarum 100, 550–560.
Sakamoto T., Miura K., Itoh H., Tatsumi T., Ueguchi-Tanaka M., Ishiyama K.& Matsuoka M. (2004) An overview of gibberellin metabolism enzymegenes and their related mutants in rice. Plant Physiology 134, 1642–1653.
Sasaki A., Itoh H., Gomi K., Ueguchi-Tanaka M., Ishiyama K., Kobayashi M. &Matsuoka M. (2003) Accumulation of phosphorylated repressor forgibberellin signaling in an F-box mutant. Science 299, 1896–1898.
GA in deepwater rice 2323
© 2014 The Authors. Plant, Cell & Environment published by John Wiley & Sons Ltd, 37, 2313–2324

Sauter M. & Kende H. (1992) Gibberellin-induced growth and regulation ofthe cell-division cycle in deep-water rice. Planta 188, 362–368.
Sharman B.C. (1942) Developmental anatomy of the shoot of Zea mays L.Annals of Botany 6, 245–282.
Suge H. (1985) Ethylene and gibberellin: regulation of internodal elongationand nodal root development in floating rice. Plant & Cell Physiology 26,607–614.
Tang D.Q., Kasai Y., Miyamoto N., Ukai Y. & Nemoto K. (2005) Comparisonof QTLs for early elongation ability between two floating rice cultivars witha different phylogenetic origin. Breeding Science 55, 1–5.
Todoroki Y., Kobayashi K., Yoneyama H., Hiramatsu S., Jin M.H., Watanabe B.& Hirai N. (2008) Structure-activity relationship of uniconazole, a potentinhibitor of ABA 8′-hydroxylase, with a focus on hydrophilic functionalgroups and conformation. Bioorganic & Medicinal Chemistry 16, 3141–3152.
Tsuji H., Aya K., Ueguchi-Tanaka M., Shimada Y., Nakazono M., Watanabe R.,. . . Matsuoka M. (2006) GAMYB controls different sets of genes and isdifferentially regulated by microRNA in aleurone cells and anthers. ThePlant Journal 47, 427–444.
Ueguchi-Tanaka M., Fujisawa Y., Kobayashi M., Ashikari M., Iwasaki Y.,Kitano H. & Matsuoka M. (2000) Rice dwarf mutant d1, which is defectivein the alpha subunit of the heterotrimeric G protein, affects gibberellinsignal transduction. Proceedings of the National Academy of Sciences of theUnited States of America 97, 11638–11643.
Ueguchi-Tanaka M., Nakajima M., Katoh E., Ohmiya H., Asano K., Saji S. &Matsuoka M. (2007a) Molecular interactions of a soluble gibberellin recep-tor, GID1, with a rice DELLA protein, SLR1, and gibberellin. The Plant Cell19, 2140–2155.
Ueguchi-Tanaka M., Nakajima M., Motoyuki A. & Matsuoka M. (2007b)Gibberellin receptor and its role in gibberellin signaling in plants. AnnualReview of Plant Biology 58, 183–198.
Ueguchi-Tanaka M., Hirano K., Hasegawa Y., Kitano H. & Matsuoka M.(2008) Release of the repressive activity of rice DELLA protein SLR1 bygibberellin does not require SLR1 degradation in the gid2 mutant. The PlantCell 20, 2437–2446.
van Veen H., Mustroph A., Barding G.A., Vergeer-Van Eijk M.,Welschen-Evertman R.A., Pedersen O. & Sasidharan R. (2013) Two Rumexspecies from contrasting hydrological niches regulate flooding tolerancethrough distinct mechanisms. The Plant Cell 25, 4691–4707.
van Veen H., Akman M., Jamar D.C., Vreugdenhil D., Kooiker M., vanTienderen P., . . . Sasidharan R. (2014) Group VII Ethylene Response Factordiversification and regulation in four species from flood-prone environ-ments. Plant, Cell & Environment 37, 2421–2432.
Vergara B. S. & Mazaredo A. (1979) Using the new standard evaluation systemto measuring elongation ability. Proceedings of the 1978 International Deep-water Rice Workshop. International Rice Research Institute, Manila.pp. 139–142.
Yamaguchi S. (2008) Gibberellin metabolism and its regulation. AnnualReview of Plant Biology 59, 225–251.
Zhu Y., Nomura T., Xu Y., Zhang Y., Peng Y., Mao B. & He Z. (2006) ELON-GATED UPPERMOST INTERNODE encodes a cytochrome P450monooxygenase that epoxidizes gibberellins in a novel deactivation reactionin rice. The Plant Cell 18, 442–456.
Received 21 December 2013; received in revised form 15 May 2014;accepted for publication 21 May 2014
SUPPORTING INFORMATION
Additional Supporting Information may be found in theonline version of this article at the publisher’s web-site:
Figure S1. Diagrams of methods of cultivation and deepwa-ter treatment and experiments. (a) Deepwater treatment.Details of plant growth conditions were described in theMaterials and Methods section. The three- to eight-leafstages (3–8 LS) of plants were treated under shallow (SW)and complete submergence (DW). Plant height was meas-ured to determine internode and leaf elongation, which rep-resent the length from the base of the aerial part of the plantto the leaf tip.Total internode length represents the sum of allinternode lengths in each plant. For gene expression analysis,stem sections containing young internode, shoot apicalmeristem and leaf sheath were collected.Figure S2. Crossing method for the mutant pyramiding line.Parental line of GA biosynthesis and signal transductionmutants and NIL-1 + 3 + 12 were shown in the upper graphi-cal genotype. The yellow closed rectangle represents the T65genes, the red closed rectangle represents QTLs from C9285and the blue closed rectangle represents GA-related genes.The F1 generations were obtained by crossing (representedas X). Backcrossing was performed several times. Thehomozygote plants of the MP line were finally selected basedupon genotyping using molecular markers. The mutant pyra-miding (MP) line possessed the three QTLs controllingthe deepwater response and the GA-related genes ashomozygotes.Figure S3. Semi-qRT-PCR analysis of GA metabolic andsignal transduction gene expression. mRNA was obtainedfrom the 7 LS of T65 and C9285 after 0, 12 and 24 h ofsubmergence. PCR was performed for 30 and 35 amplifica-tion cycles for GA metabolic and signal transduction genesexpression and compared with that of OsActin1 amplified for30 and 35 cycles.Table S1. Adjusted P-values for the comparative GA con-tents in T65 and C9285 using the Holm’s method. Significantdata (P-value under 0.05; Student’s t-test) was obtained byHolm’s method. The data show GA1 and GA4 contentsbetween T65 versus C9285 at 0, 6, 12, 24 and 48 h points andT65 versus NIL1 + 3 + 12 at 0, 6, 12, 24 and 48 h points.Table S2. Sequence of primers used for the GA-related geneexpression analysis using semi-qRT-PCR and qRT-PCR. Spe-cific primers were designed for semi-qRT-PCR. Biosynthesisand signal transduction of GA were analysed as describedpreviously.
2324 M. Ayano et al.
© 2014 The Authors. Plant, Cell & Environment published by John Wiley & Sons Ltd, 37, 2313–2324