genetic characterization of captive cuban crocodiles (crocodylus rhombifer) and evidence of...
TRANSCRIPT

A Journal of Integrative Biology
Genetic Characterization of Captive Cuban Crocodiles(Crocodylus rhombifer) and Evidence of HybridizationWith the American Crocodile (Crocodylus acutus)
JEREMY P. WEAVER1�, DAVID RODRIGUEZ1, MIRYAM VENEGAS-ANAYA1,JOSE ROGELIO CEDENO-VAZQUEZ2, MICHAEL R.J. FORSTNER3, AND
LLEWELLYN D. DENSMORE III1
1Department of Biological Sciences, Texas Tech University, Lubbock, Texas2El Colegio de la Frontera Sur, Unidad Chetumal, Chetumal, Q. Roo, Mexico3Department of Biology, Texas State University-San Marcos, San Marcos, Texas
ABSTRACT There is a surprising lack of genetic data for the Cuban crocodile (Crocodylusrhombifer), especially given its status as a critically endangered species. Samples from captiveindividuals were used to genetically characterize this species in comparison with other New Worldcrocodilians. Partial mitochondrial sequence data were generated from cyt-b (843 bp) and thetRNAPro- tRNAPhe-D-loop region (442 bp). Phylogenetic analyses were performed by generatingmaximum parsimony, maximum likelihood, and Bayesian-based topologies. In addition, in an effortto identify species-specific alleles, ten polymorphic microsatellite loci were genotyped. Distance andmodel-based clustering analyses were performed on microsatellite data, in addition to a model-basedassignment of hybrid types. Both mitochondrial and nuclear markers identified two distinctC. rhombifer genetic sub-clades (a and b); and microsatellite analyses revealed that most admixedindividuals were F2 hybrids between C. rhombifer-a and the American crocodile (C. acutus). Allindividuals in the C. rhombifer-b group were morphologically identified as C. acutus and formed adistinct genetic assemblage. J. Exp. Zool. 309A, 2008. r 2008 Wiley-Liss, Inc.
How to cite this article: Weaver JP, Rodriguez D, Venegas-Anaya M, Cedeno-Vazquez JR,Forstner MRJ, Densmore III LD. 2008. Genetic characterization of captive Cubancrocodiles (Crocodylus rhombifer) and evidence of hybridization with the Americancrocodile (Crocodylus acutus). J. Exp. Zool. 309A:[page range].
Cuban crocodiles (Crocodylus rhombifer) areconsidered endangered (CITES Appendix, IUCNRed List-EN) due to their limited distribution,habitat loss, and the introduction of exotic animalsinto their environment (Ross, ’98). The recentencroachment of humans into C. rhombifer’sterritory has limited its distributional range toabout 186 square miles (300 km2) within Cienagade Zapata and Cienaga de Lanier in southwesternCuba (Fig. 1). Sub-fossils of C. rhombifer found inCuba are dated to the Pleistocene (Varona, ’66;’84); whereas several sub-fossils found on GrandCayman (Morgan et al., ’93) and the Bahamas(Franz et al., ’95) are from the Holocene, whichsuggests it was historically found throughout mostof the Caribbean. It has been suggested that apossible contributing factor to the declineC. rhombifer over the past 25 years may havebeen the introduction of the Brown caiman
(Caiman crocodilus fuscus) into Cienaga de Lanier(Ross, ’98). Although recent reports by R. Soberon(personal communication) have indicated that thisis not possible because the C. rhombifer populationon that island was already extirpated beforeC. c. fuscus was introduced. C. rhombifer is alsonaturally sympatric with the American crocodile(C. acutus), but C. acutus has a more extensivedistribution that extends from North America intoSouth America and the Caribbean (Ramos et al.,’94; Thorbjarnarson et al., 2006).
Published online in Wiley InterScience (www.interscience.wiley.com). DOI: 10.1002/jez.471
Received 16 July 2007; Revised 14 February 2008; Accepted 4 May2008
Grant sponsor: Howard Hughes Medical Institute Grant.�Correspondence to: Jeremy P. Weaver, Department of Biological
Sciences, Texas Tech University, Lubbock, TX 79409.E-mail: [email protected]
r 2008 WILEY-LISS, INC.
JOURNAL OF EXPERIMENTAL ZOOLOGY 309A (2008)

The Cuban crocodile’s breeding season overlapswith that of C. acutus by a few days in the monthof January (Varona, ’66). The variation in lengthbetween these two animals is approximately 1.5 mwith adult C. acutus males reaching 5 m andC. rhombifer reaching 3.5 m (Varona, ’66); thus,making hybridization physically possible. Therehave been several documented cases of hybridiza-tion between crocodiles in captive populations;most pertinently between C. acutus and C.rhombifer at the Laguna del Tesoro farm in Cuba(Ross, ’98), and consequently these hybrids couldhave been distributed to US zoological parks andprivate collectors.
The ability to detect hybrids is essential inidentifying pure breeding populations for reintro-ductions into extirpated areas (Allendorf et al.,2001; FitzSimmons et al., 2002). Despite thecritical status of wild C. rhombifer populationsthere has been little or no genetic data publishedor reported on this species; although, there is anongoing ecological study that will include somegenetic analyses (R. Ramos and O. Sanjur,personal communication). Hybrid introgressionhas been detected in some New World crocodilians(Hekkala, 2004; Ray et al., 2004; Rodriguez, 2007;Cedeno-Vazquez et al., 2008; Rodriguez et al.,2008), and owing to C. rhombifer’s smallerpopulation numbers and its frequent sympatrywith C. acutus, the genetic integrity of this speciesis at risk. It has been suggested that detectinghybrids is less exhaustive when the two parentalcrocodiles possess different karyotypes, but detec-tion of hybridization between individuals with
similar karyotypes requires more in-depth ana-lyses (Chavananikul et al., ’94; FitzSimmons et al.,2002). Due to the chromosomal and biochemicalsimilarity (Cohen and Gans, ’70; Densmore, ’83)and the relatively recent divergence (Brochu,2000) between C. rhombifer and C. acutus, detect-ing hybrids based on morphological charactersalone may be problematic. In this specific case, theuse of molecular markers is warranted.
Molecular markers have been used routinely tocharacterize threatened species and populations(Frankham et al., 2002), but genetic studies firstrequire a point of reference to accurately assessspecies assignments. Mitochondrial DNA(mtDNA) is useful in constructing phylogeniesand haplotype networks. However, the maternalinheritance of mtDNA limits our ability to detecthybridization to cases where there is disagreementbetween morphology and mtDNA assignments.Therefore, nuclear DNA (nDNA) markers mustalso be utilized to more accurately evaluate speciesdesignations and determine hybrid types. Devel-oping a pure breeding stock of C. rhombifer will beessential in maintaining the genetic integrity ofthe species, which is why any potential hybridiza-tion with other species can be a problem in captivepopulations. The purpose of our study was togenetically characterize all available samples ofcaptive Cuban crocodiles from US zoologicalinstitutions and to describe C. rhombifer-likehaplotypes found in the Caribbean, Florida, andMexico, which may present a threat to the geneticpurity of other endemic crocodile species. Thiswork will provide the foundation for future genetic
Fig. 1. Map of Cuba with localities of C. rhombifer-like haplotypes (a and b) found outside their present range (verticalstripes).
WEAVER ET AL.2
J. Exp. Zool.

treatments of wild Cuban crocodile populations,assist in further efforts to identify hybrids outsideof Cuba, and assess the utility of genetic methodsin detecting inter-specific admixture within othercaptive populations.
MATERIALS AND METHODS
Samples
Wild C. rhombifer populations from Cuba couldnot be sampled, but zoological specimens werereadily available. Whole blood or skin clips werecollected from captive C. rhombifer, wild caughtC. acutus, wild caught C. moreletii (Morelet’scrocodile) and a captive C. intermedius (Orinococrocodile) (see Appendix). Blood was collected viathe caudal or dorsal sinus and stored in cell lysisbuffer (Gorzula et al., ’76; Bayliss, ’87), whereasskin clips were stored in 95% ethanol. Both sets oftissue were stored at �201C prior to DNAisolation. Total genomic DNA was extracted usingthe PureGene isolation kit (Gentra Systems,Minneapolis, MN), electrophoresed on a 2%agarose gel, and visualized with ethidium bromideunder UV light.
Genetic markers
Mitochondrial DNA
A partial cytochrome-b (cyt-b) fragment (843 bp)was amplified from C. rhombifer, C. acutus,C. moreletii, and C. intermedius using primerscrCYTBfor and crCYTBrev. Primers drL15459(modified from Glenn et al., 2002) and CR2HA(modified from Ray and Densmore, 2002) wereused to amplify the tRNAPro-tRNAPhe-D-loopregion (442 bp; Table 1) for only C. rhombiferand C. acutus. Polymerase chain reactions (PCR)were performed in 50mL volumes using 0.50 mL oftotal genomic DNA (tDNA) (50 ng/mL), 36.25 mL ofddH20, 10 mL of buffer (0.3 M TRIS, 0.0175 MMgCl2, and 0.075 M (NH4)2SO4), 2.0mL of 2.5 mMdNTPs, 0.50 mL (10 mM) of forward primer,0.50 mL (10 mM) reverse of primer and 0.25 mL(1.25 U) of Promega Taq polymerase (PromegaCorp., Madison, WI). Thermocycling conditions forall primers consisted of an initial denaturationstep of 2 min at 941C, then 33 cycles of 30 sec at941C, 1 min at 581C, and 45 sec at 721C; with afinal extension of 7 min at 721C. Unincorporateddinucleotides and primers were removed fromPCR products using the Qiagen PCR purificationkit (Qiagen, Inc., Valencia, CA). Products werecycle sequenced using Big Dye v3.1 dye terminator
(Applied Biosystems, Inc., Foster City, CA). Cyclesequence products were purified by passingthrough a G-50 Sephadex column (0.5 gm ofSephadex/800 mL ddH2O), which was incubatedat room temperature for 30 min and centrifuged at3,000 rpm for 2 min to construct the column. Driedcycle sequence product was denatured in forma-mide and electrophoresed on an ABI 3100-Avantgenetic analyzer (Applied Biosystems, Inc., FosterCity, CA). Chromatograms were viewed andtrimmed using Sequencher 4.1.4 (Gene CodesCorp. Ann Arbor, MI), and then aligned usingClustal X (Thompson et al., ’97) and BioEdit 5.0.6(Hall, ’99). All newly generated sequences wereaccessioned into the GenBank database(EU034541-EU034627). Three sequences obtainedfrom NCBI were used for comparison with ourcyt-b sequence data, the Estuarine crocodile(C. porosus; AJ810453), the Siamese crocodile(C. siamensis; DQ353946) and the Nile crocodile(C. niloticus; AJ810452).
Microsatellites
Ten polymorphic microsatellite loci (Dever andDensmore, 2001; FitzSimmons et al., 2001) werealso amplified for each individual (Table 2).Primers were fluorescently labeled with WellReddyes (Beckman Coulter, Inc., Fullerton, CA) andamplified using 12.5 mL PCR reactions, whichincluded 0.125 mL of tDNA, 9.06 mL of ddH2O,2.50 mL buffer (0.3 M TRIS, 0.0175 M MgCl2, and0.075 M (NH4)2SO4), 0.50 mL of 2.5mM dNTPs(10 mM), 0.13 mL forward primer (10 mM),0.13 mL reverse primer (10 mM), and 0.0625 mL(0.31 U) Promega Taq polymerase. PCR conditionsfor all primers consisted of an initial denaturationstep of 2 min at 941C, then 33 cycles of 30 sec at941C, 1 min at 581C or 621C (Table 2), and 45 sec at721C; with a final extension of 5 min at 721C.Fragments were sized based on a 400 bp size
TABLE 1. Primer sequences used to generate mtDNA frag-ments
Primer Sequence
crCYTBfor 50 ATGACCCACCAACTACGAAAATC 30
crCYTBrev 50 CGAAGGGGTTTGATTAATAGGTT 30
CrCYTBintfor1 50 TAGCAACTGCCTTCATAGGCTAC 30
drL15459 50 AGGAAAGCGCTGGCCTTGTAA 30
CR2HA 50 GGGGCCACTAAAAACTGGGGGGA 30
1Used only for sequencing.
GENETIC CHARACTERIZATION OF THE CUBAN CROCODILE 3
J. Exp. Zool.
standard, using a CEQ8800 genetic analyzer andsoftware (Beckman Coulter, Inc., Fullerton, CA).Using identical scoring methods, genotypes fromall C. rhombifer samples were compared with 14pure C. acutus (see Appendix) identified byRodriguez (2007) and Rodriguez et al. (2008).
Data analysis
Sequence data
MODELTEST (Posada and Crandall, ’98) wasused to obtain the best-fit model of nucleotidesubstitution for maximum likelihood (ML), andMRMODELTEST (Nylander, 2004) was used toobtain the best-fit model of evolution for Bayesianinference (BI). Phylogenetic analyses of cyt-bsequence data, under the ML and maximumparsimony criteria, were performed in PAUP�4.0b10(Swofford, 2002). A starting tree was generated bystepwise addition of taxa, with swapping performedby utilizing the tree bisection reconnection algo-rithm. Node support was determined by bootstrap-ping topologies for 1,000 replications. Additionally, aBI tree, with posterior probabilities, was constructedusing MrBayes v3.1.2 (Huelsenbeck and Ronquist,2001). Four Markov chains were implemented for1,000,000 iterations after an initial burn-in of100,000 iterations. To obtain finer haplotype resolu-tion, a neighbor-joining tree was constructed inPAUP using uncorrected pairwise genetic distancesbased on tRNAPro-tRNAPhe-D-loop sequence data.
Node support was determined by bootstrapping theresulting topology for 1,000 iterations.
Microsatellite data
The program POPULATIONS v1.2.28 (Langel-la, ’99) was employed to estimate Dc pairwisegenetic distances (Cavalli-Sforza and Edwards,’67) to construct an exploratory neighbor-joiningtree for all C. rhombifer and C. acutus individuals,which was visualized in TREEVIEW (Page, ’96).The program STRUCTURE (Pritchard et al.,2000) was used to determine assignment prob-abilities to specific genetic clusters by constrainingK to the number of clades suggested by theneighbor-joining topology. We assumed that pure-bred individuals will have high assignment prob-abilities (40.97) to species genetic clusters,whereas hybrids will have intermediate assign-ment probabilities (o0.97). CERVUS 3.0 (Mar-shall et al., ’98) was used to estimate measures ofmicrosatellite diversity. If hybrids were detected,then NEWHYBRIDS (Anderson and Thompson,2002) was used to implement a Bayesian-basedalgorithm, which assigns individuals into sixgenotypic classes. Genotype classes consist of twoparental groups (C. rhombifer and C. acutus), firstgeneration hybrids (F1), second generation hy-brids (F2), F1 backcrosses to C. rhombifer and F2
backcrosses to C. acutus.
TABLE 2. Microsatellite diversity values for each genetic cluster inferred using STRUCTURE (without admixed individuals)
C. rhombifer-I C. rhombifer-II C. acutus
Locus AT (C1)1 N2 A3 HO4 HE
5 N2 A3 HO4 HE
5 N2 A3 HO4 HE
5
C391 58 19 1 0.00 0.00 7 5 1.00 0.78 14 5 0.57 0.68Cj16 62 22 4 0.82 0.65 7 3 0.57 0.58 14 7 0.86 0.78Cj18 58 21 3 0.57 0.64 7 3 0.57 0.65 14 4 0.64 0.60Cj20 62 22 4 0.64 0.66 7 5 0.71 0.73 14 4 0.50 0.56Cj109 62 20 3 0.80 0.63 7 4 0.29 0.50 14 6 0.50 0.82Cj119 58 21 4 0.43 0.50 7 3 0.43 0.69 14 3 0.50 0.62Cj131 58 21 6 0.91 0.66 7 2 0.86 0.53 14 2 0.14 0.25Cu5-123 58 22 3 0.32 0.35 7 5 0.71 0.81 14 5 0.50 0.48Cud68 58 18 3 0.67 0.48 7 3 0.86 0.67 14 2 0.36 0.52Cuj131 58 20 2 0.05 0.05 7 4 0.43 0.71 14 2 0.29 0.35mean 3.3 0.52 0.46 3.7 0.64 0.66 4.0 0.49 0.57
1Annealing temperature.2Number of individuals sampled per locus.3Number of average alleles per locus.4Observed heterozygosity.5Expected heterozygosity.
WEAVER ET AL.4
J. Exp. Zool.

RESULTS
Sequence-based analyses
For cyt-b sequence data, MODELTEST andMRMODELTEST indicated the best model of
nucleotide substitution was GTR1G for both MLand BI. The C. rhombifer species group fell withinthe New World crocodilian clade that includedC. niloticus (Fig. 2). Only two haplotypes werefound using cyt-b sequence data (a and b), whichalso corresponded to two haplotypes detectedusing tRNAPro-tRNAPhe-D-loop sequences. Mea-sures of percent uncorrected distances based oncyt-b sequences (Table 3), between the C. rhombi-fer-a haplotype and C. acutus, and between theC. rhombifer-b haplotype and C. acutus were bothestimated at 5.3%. Percent divergence betweenthe two C. rhombifer cyt-b haplotypes was0.9%. Similarly, estimated divergences fortRNAPro-tRNAPhe-D-loop sequences were 3.4%between C. rhombifer-a haplotypes and C. acutus,and 4.1% between C. rhombifer-b haplotypes andC. acutus, respectively; whereas the genetic dis-tance between C. rhombifer-a and C. rhombifer-bhaplotypes was estimated at 1.6% (data notshown).
Microsatellite analyses
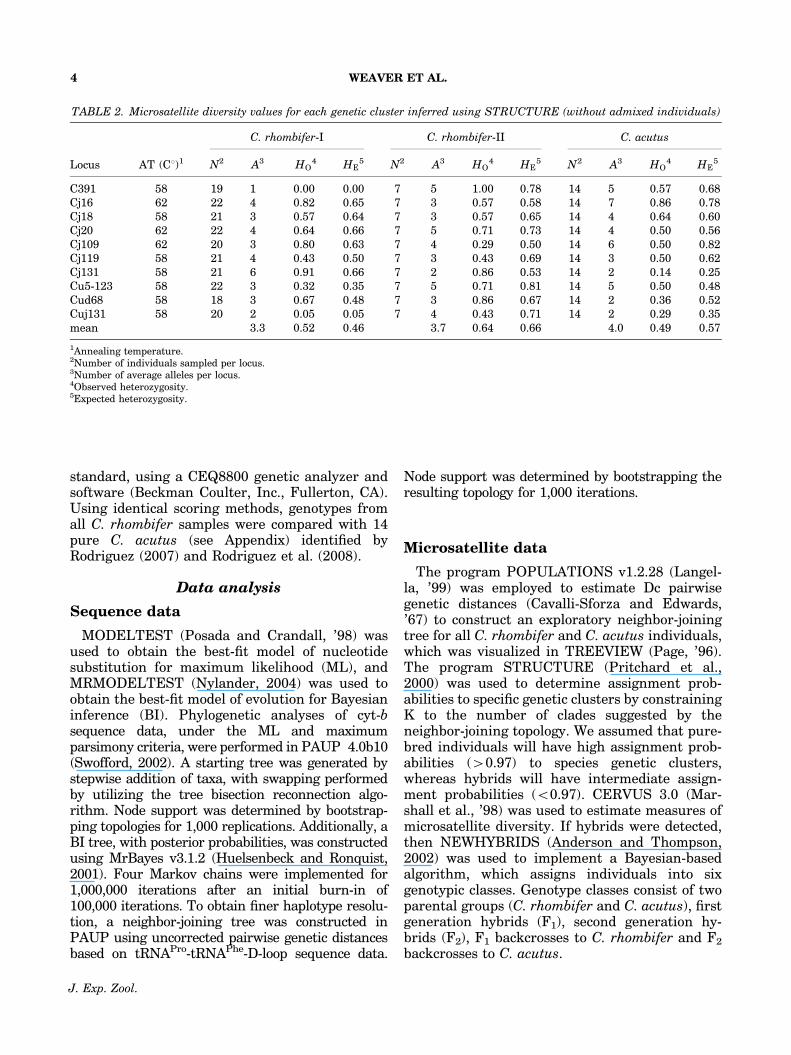
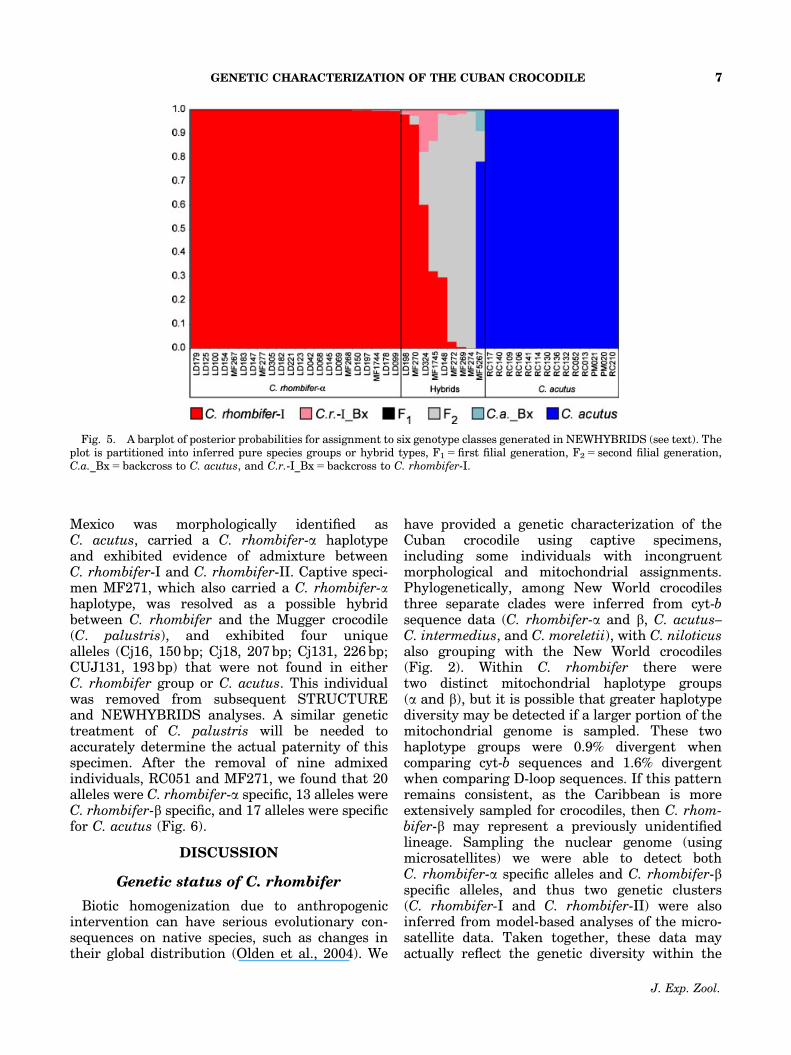
When all the samples were pooled, a matrix ofDc distances returned three distinct clades on aneighbor-joining tree (C. rhombifer-I, C. rhombi-fer-II, and C. acutus) with several individualsclustering between clades (Fig. 3). Using theneighbor-joining tree as a guide, K was con-strained to three for model-based clusteringmethods implemented in STRUCTURE; thesegenetic clusters were named C. rhombifer-,C. rhombifer-2, and C. acutus. Posterior assign-ment probabilities suggested that admixture wasprimarily occurring between C. rhombifer-I andC. acutus (Fig. 4), therefore, individuals withintermediate probabilities were designated ashybrids (Appendix). After the STRUCTURE re-sults, NEWHYBRIDS was used to classify hybridtypes between C. rhombifer-I and C. acutus,these individuals were mostly F2 hybrids (Fig. 5).One individual, RC051, collected in Cancun,
Fig. 2. (A) Bayesian consensus tree for cyt-b sequences,showing values (BI/MP/ML) where �5 (1.00/100/100). (B)Neighbor-joining tree based on tRNAPro-tRNAPhe-D-loopsequences with bootstrap support values. Two distinct sub-clades were inferred using both mitochondrial sequences.
TABLE 3. Uncorrected pairwise genetic distance values for cyt-b
(1) RC051 (2) MF348 (3) RC210 (4) LD175 (5) RC206 (6) AJ810452 (7) AJ810453 (8) DQ353946
(1) C. rhombifer-a –(2) C. rhombifer-b 0.009 –(3) C. acutus 0.053 0.053 –(4) C. intermedius 0.057 0.055 0.015 –(5) C. moreletii 0.052 0.052 0.045 0.046 –(6) C. niloticus 0.079 0.077 0.056 0.059 0.061 –(7) C. porosus 0.104 0.107 0.093 0.093 0.098 0.114 –(8) C. siamensis 0.121 0.121 0.107 0.107 0.110 0.123 0.021 –
GENETIC CHARACTERIZATION OF THE CUBAN CROCODILE 5
J. Exp. Zool.
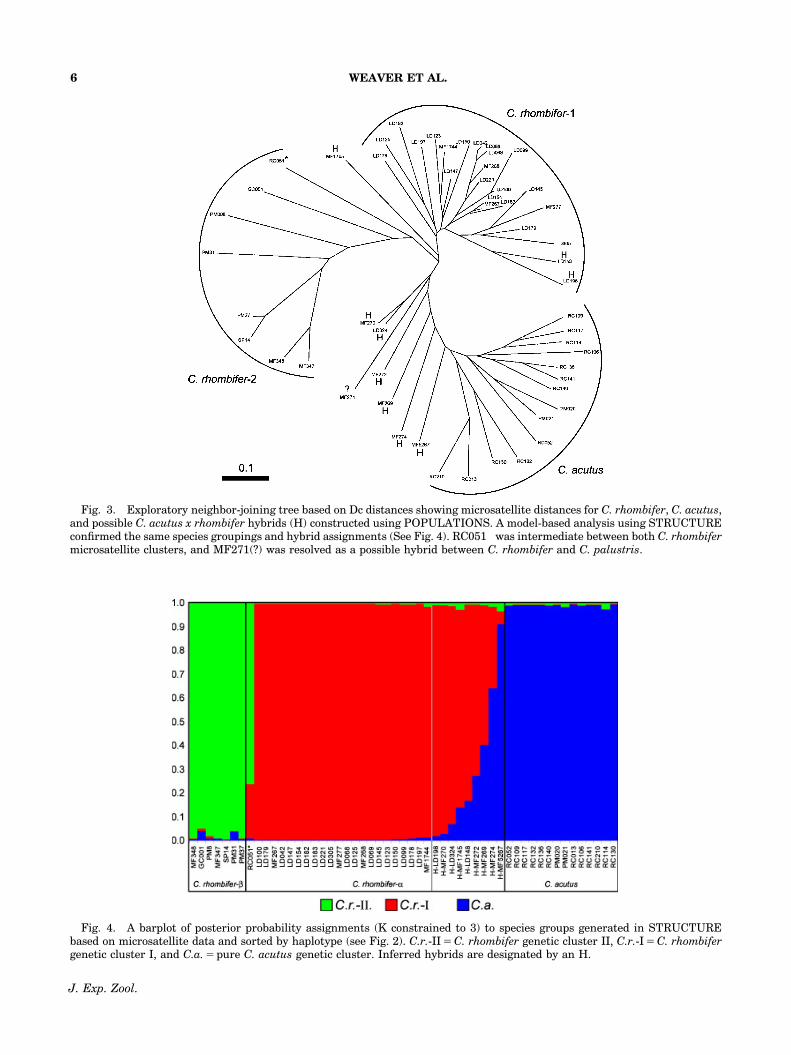
Fig. 4. A barplot of posterior probability assignments (K constrained to 3) to species groups generated in STRUCTUREbased on microsatellite data and sorted by haplotype (see Fig. 2). C.r.-II 5 C. rhombifer genetic cluster II, C.r.-I 5 C. rhombifergenetic cluster I, and C.a. 5 pure C. acutus genetic cluster. Inferred hybrids are designated by an H.
Fig. 3. Exploratory neighbor-joining tree based on Dc distances showing microsatellite distances for C. rhombifer, C. acutus,and possible C. acutus x rhombifer hybrids (H) constructed using POPULATIONS. A model-based analysis using STRUCTUREconfirmed the same species groupings and hybrid assignments (See Fig. 4). RC051� was intermediate between both C. rhombifermicrosatellite clusters, and MF271(?) was resolved as a possible hybrid between C. rhombifer and C. palustris.
WEAVER ET AL.6
J. Exp. Zool.
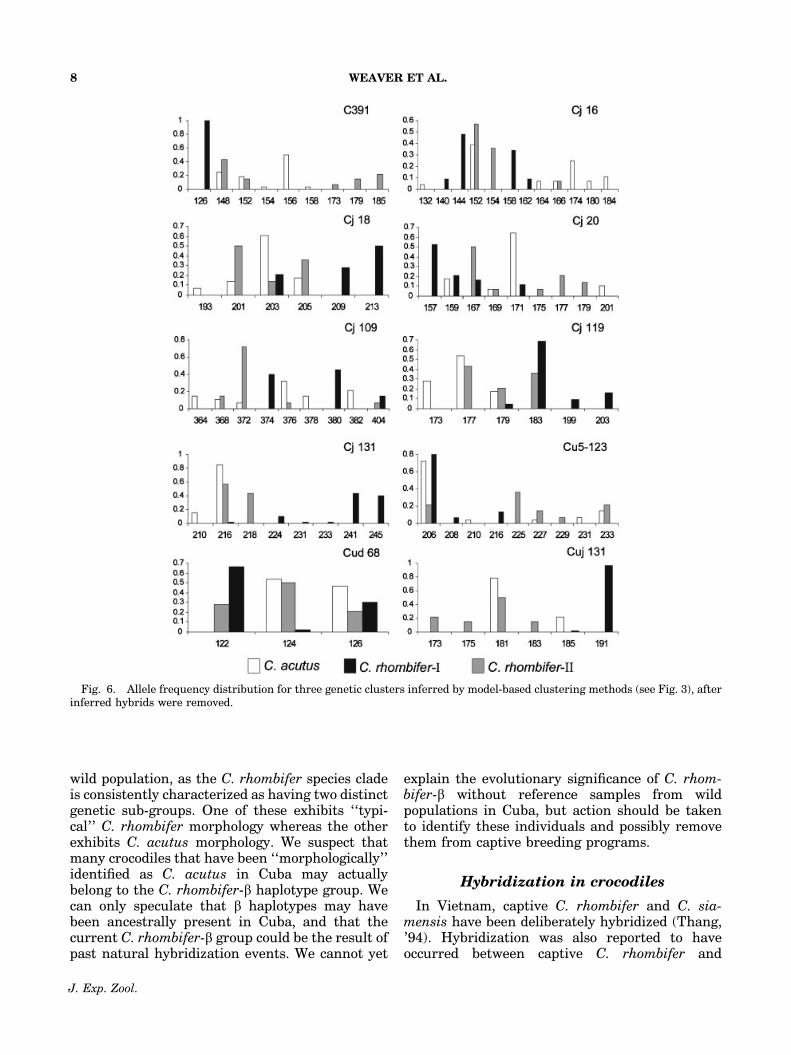
Mexico was morphologically identified asC. acutus, carried a C. rhombifer-a haplotypeand exhibited evidence of admixture betweenC. rhombifer-I and C. rhombifer-II. Captive speci-men MF271, which also carried a C. rhombifer-ahaplotype, was resolved as a possible hybridbetween C. rhombifer and the Mugger crocodile(C. palustris), and exhibited four uniquealleles (Cj16, 150 bp; Cj18, 207 bp; Cj131, 226 bp;CUJ131, 193 bp) that were not found in eitherC. rhombifer group or C. acutus. This individualwas removed from subsequent STRUCTUREand NEWHYBRIDS analyses. A similar genetictreatment of C. palustris will be needed toaccurately determine the actual paternity of thisspecimen. After the removal of nine admixedindividuals, RC051 and MF271, we found that 20alleles were C. rhombifer-a specific, 13 alleles wereC. rhombifer-b specific, and 17 alleles were specificfor C. acutus (Fig. 6).
DISCUSSION
Genetic status of C. rhombifer
Biotic homogenization due to anthropogenicintervention can have serious evolutionary con-sequences on native species, such as changes intheir global distribution (Olden et al., 2004). We
have provided a genetic characterization of theCuban crocodile using captive specimens,including some individuals with incongruentmorphological and mitochondrial assignments.Phylogenetically, among New World crocodilesthree separate clades were inferred from cyt-bsequence data (C. rhombifer-a and b, C. acutus–C. intermedius, and C. moreletii), with C. niloticusalso grouping with the New World crocodiles(Fig. 2). Within C. rhombifer there weretwo distinct mitochondrial haplotype groups(a and b), but it is possible that greater haplotypediversity may be detected if a larger portion of themitochondrial genome is sampled. These twohaplotype groups were 0.9% divergent whencomparing cyt-b sequences and 1.6% divergentwhen comparing D-loop sequences. If this patternremains consistent, as the Caribbean is moreextensively sampled for crocodiles, then C. rhom-bifer-b may represent a previously unidentifiedlineage. Sampling the nuclear genome (usingmicrosatellites) we were able to detect bothC. rhombifer-a specific alleles and C. rhombifer-bspecific alleles, and thus two genetic clusters(C. rhombifer-I and C. rhombifer-II) were alsoinferred from model-based analyses of the micro-satellite data. Taken together, these data mayactually reflect the genetic diversity within the
Fig. 5. A barplot of posterior probabilities for assignment to six genotype classes generated in NEWHYBRIDS (see text). Theplot is partitioned into inferred pure species groups or hybrid types, F1 5 first filial generation, F2 5 second filial generation,C.a._Bx 5 backcross to C. acutus, and C.r.-I_Bx 5 backcross to C. rhombifer-I.
GENETIC CHARACTERIZATION OF THE CUBAN CROCODILE 7
J. Exp. Zool.
wild population, as the C. rhombifer species cladeis consistently characterized as having two distinctgenetic sub-groups. One of these exhibits ‘‘typi-cal’’ C. rhombifer morphology whereas the otherexhibits C. acutus morphology. We suspect thatmany crocodiles that have been ‘‘morphologically’’identified as C. acutus in Cuba may actuallybelong to the C. rhombifer-b haplotype group. Wecan only speculate that b haplotypes may havebeen ancestrally present in Cuba, and that thecurrent C. rhombifer-b group could be the result ofpast natural hybridization events. We cannot yet
explain the evolutionary significance of C. rhom-bifer-b without reference samples from wildpopulations in Cuba, but action should be takento identify these individuals and possibly removethem from captive breeding programs.
Hybridization in crocodiles
In Vietnam, captive C. rhombifer and C. sia-mensis have been deliberately hybridized (Thang,’94). Hybridization was also reported to haveoccurred between captive C. rhombifer and
Fig. 6. Allele frequency distribution for three genetic clusters inferred by model-based clustering methods (see Fig. 3), afterinferred hybrids were removed.
WEAVER ET AL.8
J. Exp. Zool.

C. acutus in breeding pens of the Laguna delTesoro farm in Cuba (Ross, ’98), as well as in thewild (Ramos et al., ’94). Varona (’66) suggestedthat in Cuba at least some admixture was takingplace between C. acutus and C. rhombifer, becauseseveral specimens exhibited morphological char-acters typical of both species, but clearly outsidethe normal range of variation found in C. acutus.However, among our samples we found that onlyMF271 and MF5267 exhibited anomalous mor-phology and that all F2 hybrids appeared to exhibitC. rhombifer morphology. We found that hybridi-zation events involving captive C. rhombifer wereinvariably between the C. rhombifer-a group andC. acutus. All hybrid individuals had C. rhombifer-like mtDNA, which suggests that in captivityhybridization is typically between a femaleC. rhombifer and a male C. acutus. These resultsare consistent and congruent with those ofFitzSimmons et al. (2002).
Anthropogenic perturbation and natural migra-tion events may pose a potential threat to thegenetic integrity of C. acutus populations in theCaribbean, Florida, and Mesoamerica. Further,given the large proportion of admixture betweenC. acutus and C. moreletii (Cedeno-Vazquez et al.,2008; Rodriguez et al., 2008), populations of truecrocodiles in Mexico may be threatened by anadditional source of hybrid introgression fromC. rhombifer. For example, RC051, which wasfound in waterways near Cancun, Mexico, carrieda C. rhombifer-a haplotype and was morphologi-cally identified as C. acutus; however, it wasultimately designated as an admixture betweenC. rhombifer-I and C. rhombifer-II by the nucleardata.
Conservation and managementimplications
Hybridization ultimately presents a manage-ment problem for New World crocodilians andcomplicates the identification of species based onmorphology alone. A genetic evaluation (usingboth mtDNA and nDNA) in conjunction with amorphometric characterization can provide amore accurate view of an individual’s ancestrythan either method alone. As an example, acrocodile marked for reintroduction into Cat TienNational Park (Vietnam) was genetically identi-fied as a C. rhombifer� siamensis hybrid and wassubsequently prevented from being released intothe wild population (FitzSimmons et al., 2002). Inorder for reintroductions of native species to be
successful, only purebred individuals should bereleased back into their native habitats (Allendorfet al., 2001). Our work has provided an initialgenetic assessment of the critically endangeredCuban crocodile. We hope these data can be usedto identify pure individuals for breeding stock,which should be considered if repopulation ofextirpated areas (e.g. Cienaga de Lanier) is to takeplace.
An evaluation of current Cuban crocodile stocksis warranted to ensure a purebred captive breed-ing line, especially considering that out of sevenUS captive stocks surveyed five had some level ofgenetic admixture. Identification of hybrids usingmorphology may be problematic given that onlytwo individuals studied were morphologicallyanomalous. Additionally, thorough genetic assess-ments of wild Cuban and American crocodilepopulations in the Caribbean are needed toprovide a better genetic reference for assignmenttests, to quantify the amount of potential geneticadmixture between genetically differentiatedgroups, and to help clarify the evolutionaryimplications of the C. rhombifer-b sub-clade. Theincrease in ease and the concomitant decrease incost of generating and analyzing molecular geneticdata with statistical model-based analyses can bepractical and informative for both managementand conservation efforts. These methods areespecially important for the conservation ofendangered fauna with limited distributions, suchas the Cuban crocodile.
ACKNOWLEDGMENTS
We thank R. Bradley and J. Hanson forassistance with sequencing, the TTU Core Labfor assistance with genotyping, and O. Sanjur andS. Mahecha at the Smithsonian Tropical ResearchInstitute for laboratory assistance. We thank S. K.Davis and T. Guerra. In addition, we thank J.McVay for his input on data analyses, S. McCrack-en for curatorial assistance, M. A. Mullen, D.Hamilton, M. Vandewege, and P. Larsen foreditorial assistance; and J. Isom, D. Fabing, andL. Durham for administrative assistance. Thisresearch was supported in part by a HowardHughes Medical Institute grant through theUndergraduate Science Education Program toTexas Tech University. This research wasdeveloped under the following permits: SIM/A-4-07, 2007/KY/000107, MX31441 (NRA:CFSTB2300411) and PRT 2-2996.
GENETIC CHARACTERIZATION OF THE CUBAN CROCODILE 9
J. Exp. Zool.
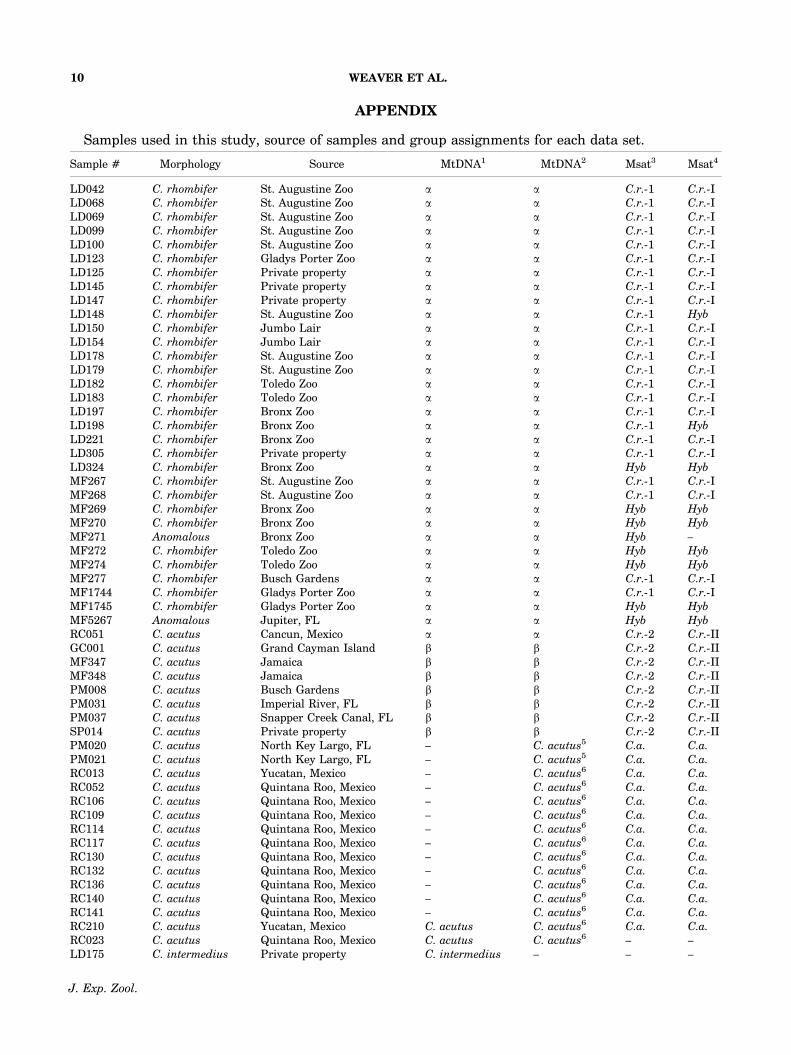
APPENDIX
Samples used in this study, source of samples and group assignments for each data set.
Sample # Morphology Source MtDNA1 MtDNA2 Msat3 Msat4
LD042 C. rhombifer St. Augustine Zoo a a C.r.-1 C.r.-ILD068 C. rhombifer St. Augustine Zoo a a C.r.-1 C.r.-ILD069 C. rhombifer St. Augustine Zoo a a C.r.-1 C.r.-ILD099 C. rhombifer St. Augustine Zoo a a C.r.-1 C.r.-ILD100 C. rhombifer St. Augustine Zoo a a C.r.-1 C.r.-ILD123 C. rhombifer Gladys Porter Zoo a a C.r.-1 C.r.-ILD125 C. rhombifer Private property a a C.r.-1 C.r.-ILD145 C. rhombifer Private property a a C.r.-1 C.r.-ILD147 C. rhombifer Private property a a C.r.-1 C.r.-ILD148 C. rhombifer St. Augustine Zoo a a C.r.-1 Hyb
LD150 C. rhombifer Jumbo Lair a a C.r.-1 C.r.-ILD154 C. rhombifer Jumbo Lair a a C.r.-1 C.r.-ILD178 C. rhombifer St. Augustine Zoo a a C.r.-1 C.r.-ILD179 C. rhombifer St. Augustine Zoo a a C.r.-1 C.r.-ILD182 C. rhombifer Toledo Zoo a a C.r.-1 C.r.-ILD183 C. rhombifer Toledo Zoo a a C.r.-1 C.r.-ILD197 C. rhombifer Bronx Zoo a a C.r.-1 C.r.-ILD198 C. rhombifer Bronx Zoo a a C.r.-1 Hyb
LD221 C. rhombifer Bronx Zoo a a C.r.-1 C.r.-ILD305 C. rhombifer Private property a a C.r.-1 C.r.-ILD324 C. rhombifer Bronx Zoo a a Hyb HybMF267 C. rhombifer St. Augustine Zoo a a C.r.-1 C.r.-IMF268 C. rhombifer St. Augustine Zoo a a C.r.-1 C.r.-IMF269 C. rhombifer Bronx Zoo a a Hyb Hyb
MF270 C. rhombifer Bronx Zoo a a Hyb Hyb
MF271 Anomalous Bronx Zoo a a Hyb –MF272 C. rhombifer Toledo Zoo a a Hyb Hyb
MF274 C. rhombifer Toledo Zoo a a Hyb HybMF277 C. rhombifer Busch Gardens a a C.r.-1 C.r.-IMF1744 C. rhombifer Gladys Porter Zoo a a C.r.-1 C.r.-IMF1745 C. rhombifer Gladys Porter Zoo a a Hyb Hyb
MF5267� Anomalous Jupiter, FL a a Hyb HybRC051� C. acutus Cancun, Mexico a a C.r.-2 C.r.-IIGC001� C. acutus Grand Cayman Island b b C.r.-2 C.r.-IIMF347� C. acutus Jamaica b b C.r.-2 C.r.-IIMF348� C. acutus Jamaica b b C.r.-2 C.r.-IIPM008 C. acutus Busch Gardens b b C.r.-2 C.r.-IIPM031� C. acutus Imperial River, FL b b C.r.-2 C.r.-IIPM037� C. acutus Snapper Creek Canal, FL b b C.r.-2 C.r.-IISP014 C. acutus Private property b b C.r.-2 C.r.-IIPM020� C. acutus North Key Largo, FL – C. acutus5 C.a. C.a.
PM021� C. acutus North Key Largo, FL – C. acutus5 C.a. C.a.RC013� C. acutus Yucatan, Mexico – C. acutus6 C.a. C.a.
RC052� C. acutus Quintana Roo, Mexico – C. acutus6 C.a. C.a.RC106� C. acutus Quintana Roo, Mexico – C. acutus6 C.a. C.a.
RC109� C. acutus Quintana Roo, Mexico – C. acutus6 C.a. C.a.RC114� C. acutus Quintana Roo, Mexico – C. acutus6 C.a. C.a.
RC117� C. acutus Quintana Roo, Mexico – C. acutus6 C.a. C.a.RC130� C. acutus Quintana Roo, Mexico – C. acutus6 C.a. C.a.
RC132� C. acutus Quintana Roo, Mexico – C. acutus6 C.a. C.a.RC136� C. acutus Quintana Roo, Mexico – C. acutus6 C.a. C.a.
RC140� C. acutus Quintana Roo, Mexico – C. acutus6 C.a. C.a.RC141� C. acutus Quintana Roo, Mexico – C. acutus6 C.a. C.a.
RC210� C. acutus Yucatan, Mexico C. acutus C. acutus6 C.a. C.a.RC023� C. acutus Quintana Roo, Mexico C. acutus C. acutus6 – –LD175 C. intermedius Private property C. intermedius – – –
WEAVER ET AL.10
J. Exp. Zool.

RC206� C. moreletii Quintana Roo, Mexico C. moreletii C. moreletii – –RC207� C. moreletii Yucatan, Mexico C. moreletii C. moreletii – –
�Wild caught individuals.1Based cyt-b sequence data (see Fig. 2A).2Based on tRNAPro- tRNAPhe-Dloop sequence data (see Fig. 2B).3Distance-based analysis of microsatellite data (see Fig. 3).4Model-based analysis of microsatellite data (see Fig. 4).5Rodriguez (2007).6Cedeno-Vazquez et al. (2008).
LITERATURE CITED
Allendorf FW, Leary RF, Spruell P, Wenburg JK. 2001. Theproblems with hybrids: setting conservation guidelines.Trends Ecol Evol 16:613–622.
Anderson EC, Thompson EA. 2002. A model-based method foridentifying species hybrids using multilocus genetic data.Genetics 160:1217–1229.
Bayliss P. 1987. Survey methods and monitoring withincrocodile management programmes. In: Webb GJ, ManolisSC, Whithead PJ, editors. Wildlife management: crocodilesand alligators. Sydney, Australia: Surrey Beatty and Sons.p 157–175.
Brochu CA. 2000. Phylogenetic relationships and divergencetiming of Crocodylus based on morphology and the fossilrecord. Copeia 2000:657–673.
Cavalli-Sforza LL, Edwards AWF. 1967. Phylogenetic analy-sis: models and estimation procedures. Am J Hum Genet19:233–257.
Cedeno-Vazquez JR, Rodriguez D, Calme S, Ross JP,Densmore III LD, Thorbjarnarson JB. 2008. Hybridizationbetween Crocodylus acutus and Crocodylus moreletii in theYucatan Peninsula: I. Evidence from mitochondrial DNAand Morphology. J Exp Zool 309A.
Chavananikul V, Wattanodorn S, Youngprapakorn P. 1994.Karyotypes of 5 species of crocodile kept in Samutprakancrocodile farm and zoo. In: Crocodiles, Proceedings of the12th Working Meeting of the Crocodile Specialist Group,IUCN—The World Conservation Union, Gland, Switzer-land. p 58–62.
Cohen MM, Gans C. 1970. The chromosomes of the ordercrocodilia. Cytogenetics 9:81–105.
Densmore LD. 1983. Biochemical and immunological systema-tics of the order crocodilia. In: Hecht MK, Wallace B,Prance GT, editors. Evolutionary biology. Vol. 16. NewYork: Plenum Publ Co. p 397–465.
Dever JA, Densmore III LD 2001. Microsatellites in Morelet’scrocodile (Crocodylus moreletii) and their utility in addres-sing crocodilian population genetics questions. J Herpetol35:541–544.
FitzSimmons NN, Tanksley S, Forstner MRJ, Louis EE,Daglish R, Gratten J, Davis S. 2001. Microsatellite markersfor Crocodylus: new genetic tools for population genetics,mating system studies and forensics. In: Grigg GC,Seebacher F, Franklin CE, editors. Crocodilian biologyand evolution. Chipping Norton: Surrey Beatty & Sons.
FitzSimmons NN, Buchan JC, Lam PV, Polet G, Hung TT,Thang NQ, Gratten J. 2002. Identification of purebredCrocodylus siamensis for reintroduction in Vietnam. J ExpZool 294:373–381.
Frankham R, Ballou JD, Briscoe DA. 2002. Introduction toconservation genetics. Cambridge: Cambridge UniversityPress.
Franz R, Morgan GS, Albury N, Buckner SD. 1995.Fossil skeleton of a cuban crocodile (Crocodylus rhombifer)from a blue hole on Abaco Bahamas. Caribbean J Sci31:149–152.
Glenn TC, Staton JL, Vu AT, Davis LM, Bremer JRA,Rhodes WE, Brisbin Jr. IL, Sawyer RH. 2002. Lowmitochondrial DNA variation among American alligatorsand a novel non-coding region in crocodilians. J Exp Zool294:312–324.
Gorzula S, Arocha-Pinango CL, Salazar C. 1976. A method ofobtaining blood by vein puncture from large reptiles.Copeia 1976:838–839.
Hall TA. 1999. BioEdit: a user-friendly biological sequencealignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp Ser 41:95–98.
Hekkala ER. 2004. Conservation genetics at the speciesboundary: case studies from African and Caribbeancrocodiles (Genus: Crocodylus). PhD Dissertation. Colum-bia University, New York, NY.
Huelsenbeck JP, Ronquist FR. 2001. MRBAYES: Bayesianinference of phylogenetic trees. Bioinformatics 17:754–755.
Langella. 1999. Populations v1.2.28: Available at http://www.cnrs-gif.fr/pge/bioinfo/populations.
Marshall TC, Slate J, Kruuk LEB, Pemberton JM. 1998.Statistical confidence for likelihood-based paternity infer-ence in natural populations. Mol Ecol 7:639–655.
Morgan GS, Richard F, Crombie RI. 1993. The Cubancrocodile, Crocodylus rhombifer, from late quaternaryfossil deposits on Grand Cayman. Caribbean J Sci29:153–156.
Nylander JAA. 2004. MrModeltest 2.1: Program distributed bythe author. Evolutionary Biology Centre, Uppsala.
Olden JD, Poff NL, Douglas MR, Douglas ME, Fausch KD.2004. Ecological and evolutionary consequences of biotichomogenization. Trends Ecol Evol 19:18–24.
Page RDM. 1996. TreeView: an application to displayphylogenetic trees on personal computers. Comput ApplBiosci 12:357–358.
Posada D, Crandall KA. 1998. MODELTEST: testingthe model of DNA substitution. Bioinformatics 14:817–818.
Pritchard JK, Stephens M, Donnelly P. 2000. Inference ofpopulation structure using multilocus genotype data.Genetics 155:945–959.
Ramos R, de Buffrenil V, Ross JP. 1994. Current status of theCuban crocodile, Crocodylus rhombifer, in the wild. In:Crocodiles, Proceedings of the 12th Working Meeting of the
Sample # Morphology Source MtDNA1 MtDNA2 Msat3 Msat4
GENETIC CHARACTERIZATION OF THE CUBAN CROCODILE 11
J. Exp. Zool.

Crocodile Specialist Group, IUCN—The World Conserva-tion Union, Gland, Switzerland. p 113–140.
Ray DA, Densmore L. 2002. The crocodilian mitochondrialcontrol region: general structure, conserved sequences, andevolutionary implications. J Exp Zool 294:334–345.
Ray DA, Dever JA, Platt SG, Rainwater TR, Finger AG,McMurry ST, Batzer MA, Barr B, Stafford PJ,McKnight J, Densmore LD. 2004. Low levels ofnucleotide diversity in Crocodylus moreletii and evidenceof hybridization with C. acutus. Conserv Genet 5:449–462.
Rodriguez D. 2007. Crocodilian evolution, systematics andpopulation genetics: recovery and ecological interactions ofthe American crocodile (Crocodylus acutus). PhD Disserta-tion. Texas Tech University, Lubbock, Texas.
Rodriguez D, Cedeno-Vazquez JR, Forstner MRJ, DensmoreIII LD. 2008. Hybridization between Crocodylus acutus andCrocodylus moreletii in the Yucatan Peninsula: II. Evi-dence from microsatellites. J Exp Zool 309A.
Ross JP. 1998. Crocodiles status survey and conservationaction plan. Gainesville, FL: Crocodile Specialist Group.96p.
Swofford DL. 2002. PAUP� v4.0b10: phylogenetic analysisusing parsimony (� and other methods). Version 4.0b10.Sunderland, MA: Sinauer.
Thang NQ. 1994. The status of Crocodylus rhombifer in theSocialist Republic of Vietnam. In: Crocodiles, Proceedingsof the 12th Working Meeting of the Crocodile SpecialistGroup, IUCN—World Conservation Union. Gland, Swit-zerland. p 141–151.
Thompson J, Gibson T, Plewniak F, Jeanmougin F, Higgins D.1997. The CLUSTAL_X windows interface: flexible strate-gies for multiple sequence alignment aided by qualityanalysis tools. Nucleic Acids Res 25:4876–4882.
Thorbjarnarson J, Mazzotti F, Sanderson E, Buitrago F,Lazcano M, Minkowski K, Muniz M, Ponce P, Sigler L,Soberon R, Trelancia AM, Velasco A. 2006. Regionalhabitat conservation priorities for the American crocodile.Biol Conserv 128:25–36.
Varona LS. 1966. Notas sobre los crocodılidos de Cuba ydescripcion de una nueva especie del Pleistoceno. Poeyana16:1–21.
Varona LS. 1984. Los cocodrilos fosiles De Cuba (reptilia:crocodylidae). Caribbean J Sci 20:13–18.
WEAVER ET AL.12
J. Exp. Zool.