genetic and metabolic diversity of streptomycetes in pulp and paper mill effluent treated crop...
TRANSCRIPT

ORIGINAL PAPER
Genetic and metabolic diversity of streptomycetes in pulpand paper mill effluent treated crop fields
Binu Mani Tripathi • Rajeev Kaushik •
Priyanka Kumari • Anil Kumar Saxena •
Dilip Kumar Arora
Received: 8 May 2010 / Accepted: 15 November 2010 / Published online: 27 November 2010
� Springer Science+Business Media B.V. 2010
Abstract Irrigation of farm field with water mixed
with pulp and paper mill effluent from Century pulp and
paper mill in Uttrakhand state of India for over last
25 years in succession increased streptomycetes population
(120 9 105) compared to the fresh water irrigated fields
(48 9 103 in WIF). Denaturing gradient gel electrophore-
sis, amplified ribosomal DNA restriction analysis, 16S
rRNA gene sequencing, BIOLOGTM substrate usage, pro-
duction of extracellular enzymes (xylanase and cellulase)
and plant growth promoting attributes were applied to
monitor changes in genetic and metabolic diversity of
streptomycetes. Significant variation was observed for
production of extracellular enzymes, Indolic compounds,
siderophore and P-solubilisation among isolates. Metabolic
substrate usage of Streptomyces isolates was evaluated
using the BIOLOGTM GP2 plates and unique carbon sub-
strate usage profiles were observed. Based on 16S rRNA
gene sequencing, the isolates were identified as Strepto-
myces variabilis, Streptomyces spp. S. glaucescens,
S. viridochromogenes, S. cinnabarinus, S. aburaviensis,
S. viridis, S. xylophagus, S. macrosporeus, S. thermocarb-
oxydus, and S. albogriseolus. The diversity index parame-
ters like Shannon index, reciprocal of Simpson’s index
(1/D), and Pielou index of evenness based on ARDRA
revealed that streptomycetes community in effluent
irrigated field (EIF) was more diverse. DGGE profiles
of Streptomyces specific 16S rRNA gene fragments
(16S-DGGE) amplified directly from soil samples were
highly similar in both soils.
Keywords ARDRA � BIOLOGTM � DGGE � PGP �Pulp and paper mill effluent � Streptomyces
Introduction
Pulp and paper mill is the major industry in India. The
heavy demand for the paper helps in steady expansion of
paper industries. Since early fifties, the number of paper
pulp mills in India has increased from 17 to more than
406 in 2008, with simultaneous increase in paper pro-
duction from 0.13 to 1.9 million tons per annum (Singh
and Thakur 2006). Pulp and paper mills are utilizing huge
amount of lignocellulosic plant components along with
various chemicals and thus regarded as major polluting
industries. Irrigating crops with pulp and paper mill
effluent is a cheap and attractive alternative for discharge
of effluent to natural waterways. Pulp and paper mill
effluent contains several elements including important
plant nutrients such as nitrogen (N), phosphorus (P) and
potassium (K), which contribute to higher crop yields
when applied to nutrient deficient soils (Udayasoorian
et al. 2004). Other elements (magnesium, sodium, chlo-
rides, sulfur) and organic compounds (chlorinated lignins,
phenolic derivatives) that are common in pulp and paper
mill effluent can cause toxicities and nutrient imbalance
in plants. The tendency of certain elements (especially
Electronic supplementary material The online version of thisarticle (doi:10.1007/s11274-010-0614-1) contains supplementarymaterial, which is available to authorized users.
B. M. Tripathi � R. Kaushik (&) � P. Kumari � D. K. Arora
National Bureau of Agriculturally Important Microorganisms,
Mau, Uttar Pradesh 275101, India
e-mail: [email protected]
A. K. Saxena
Division of Microbiology, Indian Agricultural Research
Institute, New Delhi, India
123
World J Microbiol Biotechnol (2011) 27:1603–1613
DOI 10.1007/s11274-010-0614-1
Na) to accumulate in pulp and paper mill effluent irri-
gated soils affects soil structure, increases soil salinity,
resistance to root expansion and reduces water percolation
and soil aeration (Howe and Wagner 1996). Furthermore,
the addition of such a ‘‘mixed bag’’ of compounds may
induce changes in physiochemical properties of the soil
and also the significant shifts in structure and function of
the associated microbial community, which in turn may
ultimately affect the soil viability for agriculture purposes
(Oved et al. 2001).
Pulp and paper mill effluent application to soil is known
to increase the population of bacteria, actinomycetes, fungi
and yeast (Kannan and Oblisami 1990a). Gram-positive,
filamentous bacteria of the genus Streptomyces within the
class Actinobacteria (Stackebrandt et al. 1997) are regarded
as common saprophytic soil bacteria with a complex life
cycle. They typically occur in soil as spores, which germi-
nate and produce substrate mycelium under favourable
nutritional conditions. Streptomyces appears to be most vital
ecologically and accounts for approximately 90% of the soil
actinomycetes community. It is also well known that
Streptomyces play an important role in the transformation of
lignocelluloses (Ramachandra et al. 1987). These microor-
ganisms are able to degrade cellulose and hemicelluloses as
they oxidize and solubilize the lignin component. Several
works reported dominance of streptomycetes population in
wetlands constructed for industrial waste water treatment
(Shatoury et al. 2004), however no systematic study has been
carried out to account for the diversity of Streptomyces
specifically in arable soils facing paper mill effluents over a
long period. Therefore, the aim of present study was to
determine the effect of irrigating sugarcane crop with the
water from local channel mixed with paper mill effluent
from Century paper mill (CPM), Lal Kuan, Uttrakhand,
India, on culturable as well as unculturable streptomycetes
community.
Materials and methods
Field sites and collection of soil samples
The effluent from the factory Century Pulp and Paper mill
(CPM) at Lal Kuan, Uttrakhand, India (79�100E longitude
and 29�30N latitude), which is discharged in local water-
way containing fresh water is being used as source of
irrigation to the sugarcane fields since last 25 years. A field
was selected from this site for sampling and was designated
as Effluent irrigated field (EIF). Samples were also col-
lected from a tube well-water irrigated field (WIF) from the
same area for comparison. The chemical composition of
effluent from CPM, effluent mixed with water and tube
well water is given in Table 1. Samples were collected
during March (2008) using a soil auger along zigzag paths
(Zigzag sampling) to account for the randomness from a
depth of approximately 15 cm. The samples were trans-
ported to the laboratory in the insulated container at 4�C.
The soil was sandy loam texture with pH 8.16; electrical
conductivity (EC), 0.73 dSm-1; organic carbon (OC),
0.95%; available N, 64.85 kg ha-1; Olsen P, 16.86 kg ha-1
and extractable K, 130.62 kg ha-1. The soil of control site
also had sandy loam texture, pH, 7.30; electrical conduc-
tivity (EC), 0.30 dSm-1; organic carbon (OC), 0.81%;
available N, 63.09 kg ha-1; Olsen P, 12.57 kg ha-1 and
extractable K, 168.56 kg ha-1.
Selective isolation and enumeration of presumptive
streptomycetes
One gram of each composite sample was suspended in
9 ml of � Ringer’s solution (Oxoid) in a universal bottle.
The resultant 10-1 preparations were agitated on a shaker
at 150 revolutions per minute (rpm) for 10 min at room
temperature, heated at 55�C for 6 min in a water bath, and
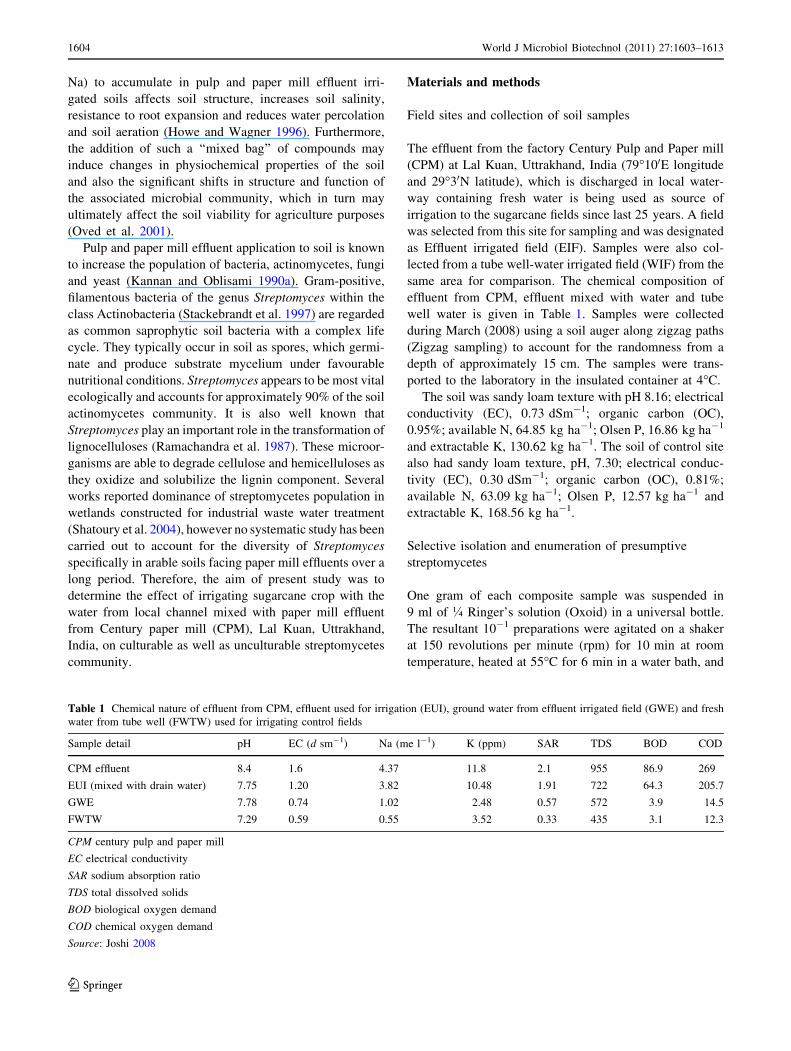
Table 1 Chemical nature of effluent from CPM, effluent used for irrigation (EUI), ground water from effluent irrigated field (GWE) and fresh
water from tube well (FWTW) used for irrigating control fields
Sample detail pH EC (d sm-1) Na (me l-1) K (ppm) SAR TDS BOD COD
CPM effluent 8.4 1.6 4.37 11.8 2.1 955 86.9 269
EUI (mixed with drain water) 7.75 1.20 3.82 10.48 1.91 722 64.3 205.7
GWE 7.78 0.74 1.02 2.48 0.57 572 3.9 14.5
FWTW 7.29 0.59 0.55 3.52 0.33 435 3.1 12.3
CPM century pulp and paper mill
EC electrical conductivity
SAR sodium absorption ratio
TDS total dissolved solids
BOD biological oxygen demand
COD chemical oxygen demand
Source: Joshi 2008
1604 World J Microbiol Biotechnol (2011) 27:1603–1613
123

cooled at room temperature. The resultant preparations
were serially diluted down to 10-6, using � strength
Ringer’s solution, and aliquots of each dilution (100 ll)
spread over the surfaces of dried starch casein agar plates
amended with cycloheximide (50 lg ml-1). The pre-
sumptive streptomycetes were ascertained from the leath-
ery colonies and an aerial spore mass, counted and
expressed as the number of colony forming units (CFUs)
per gram dry soil. The isolates were examined for the
presence of isomers of diaminopimelic acid (A2pm) in
whole organism hydrolysates using the procedure descri-
bed by (Staneck and Roberts 1974). A standard solution
(10 lM) of A2pm (Sigma) containing a mixture of DL-,
LL- and meso-A2pm isomers was used as a reference.
Purified isolates were maintained on oatmeal agar slopes
while spores and hyphal fragments stored in glycerol (20%,
v/v) at -20�C. Isolates obtained from EIF and WIF are
designated by prefixing NBE and NBC before isolate
number, respectively.
Assay of xylanase and cellulase and plant growth
promoting activity
All the isolates were tested qualitatively for production of
xylanase and cellulase. Based on initial screening, selected
isolates were also screened on quantitative basis. The
production of xylanase and cellulase was estimated fol-
lowing the method as described by Sanghi et al. (2007) and
Zvereva et al. (2006). Indolic compounds production was
estimated colorimetrically (Gordon and Weber 1951). All
the isolates were also screened for phosphate solubilization
on Pikovskaya’s agar plates (Mehta and Nautiyal 2001).
Siderophore production was examined on chrome-Azurol-
S agar medium as described (Schwyn and Neilands 1987).
Carbon substrate usage
Carbon substrate usage by isolates was measured using
the BIOLOGTM GP2 automated identification system
(Hayward, California). The strains were grown on oatmeal
agar plates for 7 days at 32�C when spores were harvested
by scrapping them from agar surfaces. The resultant spore
preparations were suspended in sterile distilled water and
washed by centrifugation at 14,000 rpm for 10 min and the
supernatants discarded (Kieser et al. 2000). This procedure
was repeated five times and the resultant washed spore
suspensions (20 ll) pipetted into two 1.5 ml microfuge
tubes each of which contained 500 ll of peptone-yeast
extract broth (Kieser et al. 2000) and incubated at 32�C in
an horizontal shaker at 220 rpm. Tubes were removed from
the shaker after 72 h and resultant preparations were cen-
trifuged, and the pelleted cells suspended in a sterile
solution. Cell density was adjusted to an OD590 of
between 0.34 and 0.39. The BIOLOGTM GP2 microplates
were immediately inoculated with 125 ll of cell suspen-
sion per well. Triplicate plates were used for each isolate
and were incubated at 32�C in sealed plastic bags for 96 h.
Substrate oxidation was measured with a microplate reader
at 590 nm. Clustering was based on binary data (usage/
nonusage) for each of the 95 substrates. Data were sub-
mitted to cluster analysis using a simple matching coeffi-
cient (SSM) (Sokal and Michener 1958) and clustering was
achieved by the unweighted-pair-group method of associ-
ation (UPGMA) (Sneath and Sokal 1973).
Genomic DNA extraction from isolates and soil
samples
Genomic DNA extraction from all the isolates was carried
out as described by Pospiech and Neumann (1995). The
total microbial community DNA was extracted directly
from WIF and EIF soil samples (0.25 g of each sample in
duplicate) using power soil DNA kit (MO BIO, USA). Soil
DNA was further purified by using Wizard DNA clean up
system (Promega, USA) to remove humic acid contamina-
tion. DNA preparations were visualized after electropho-
resis in a 0.8% agarose gel in 19 TBE buffer to assess their
integrity and then stored at 4�C prior to PCR amplification.
Amplified ribosomal DNA restriction analysis
(ARDRA)
The gene encoding 16S rRNA from selected isolates was
amplified by PCR using the pair of universal primers pA
and pH and the conditions described in Massol-Deya et al.
(1995). The PCR products were monitored through gel
electrophoresis (1% agarose, w/v), followed by ethidium
bromide staining and UV transillumination. Approximately
1 lg of PCR-amplified 16S rRNA gene fragments were
restricted with endonucleases Dde I, Mbo I and Taq I
(Fermentas, USA) separately at 37�C for overnight and
resolved by electrophoresis in 2.5% agarose. Banding
pattern was visualized by ethidium bromide staining and
documented in gel documentation and analysis system
(Alphaimager, USA). Strong and clear bands were scored
for similarity and clustering analysis using the software,
NTSYS-PC2 package (Numerical taxonomy analysis pro-
gram package, Exeter software, USA). Similarity among
the strains was calculated by Jaccard’s coefficient, and
dendrogram constructed using UPGMA method (Nei and
Li 1979).
16S rRNA gene sequencing and phylogenetic analysis
Purified 16S rRNA gene of representative isolates from
each cluster was used as a template in cycle sequencing
World J Microbiol Biotechnol (2011) 27:1603–1613 1605
123
reactions with fluorescent dye-labeled terminators (Big
Dye, Applied Biosystems). Both primers pA and pH were
used for sequencing and run in 3130 9 l ABI prism auto-
mated DNA sequencer. All the sequences were compared
with 16S rRNA gene sequences available in the GenBank
databases by BLASTn search. Identification to the species
level was determined based on 16S rRNA gene sequence
similarity ([97%) with that of a prototype strain sequence.
Multiple sequence alignment of approx 1,500-bp sequences
was performed using CLUSTAL W, version 1.8 (Thomp-
son et al. 1994). A phylogenetic tree was constructed using
the neighbor-joining method (Saitou and Nei 1987). Tree
topologies were evaluated through bootstrap analysis of
1,000 data sets by MEGA 4.0 package (Tamura et al.
2007). The 16S rRNA gene sequences were submitted to
NCBI GenBank database under accession numbers
GQ268015 to GQ268026 and GU136398 to GU136399.
Diversity indices
The Shannon index (H) (Shannon and Weaver 1949),
reciprocal of Simpson’s index (1/D) (Magurran 1988) and
Pielou index (E) (Pielou 1969) were chosen to characterize
the Streptomyces communities based on the ARDRA pro-
file in WIF and EIF soils. The use of 1/D instead of the
original formulation of Simpson’s index ensures that an
increase in the reciprocal index reflects an increase in
diversity (Magurran 1988).
PCR amplification of 16S rRNA coding genes
for DGGE
Fragments of 16S rRNA genes were amplified using DNA
from both WIF and EIF soils by first PCR with primers
specific to streptomycetes StrepB 50-ACA AGC CCT GGA
AAC GGG GT-30 and StrepF 50-ACG TGT GCA GCC
CAA GAC A-30 described by Rintala et al. (2001). Second
PCR was performed by using first PCR product as template
with primers F243 50-GGA TGA GCC CGC GGC CTA
and R513-GC 50-CGC CCG GGG CGC GCC CCG GGC
GGG GCG GGG GCA CGG GGG GCG GCC GCG GCT
GCT GGC ACG TA-30 Heuer et al. (1997). The PCR
products were visualized by agarose gel electrophoresis
(1.4% gel) followed by staining with ethidium bromide.
Amplicons were stored at -20�C until DGGE analysis.
DGGEs were carried out using a Bio-Rad DCode Universal
Mutation Detection System (Bio-Rad Laboratories,
Germany). PCR products (approximately 150 ng) were
applied directly onto 8% (w/v) polyacrylamide gels in
19 TAE buffer (40 mM Tris-acetate; pH 7.4, 20 mM
sodium acetate, 1 mM disodium EDTA) containing a
denaturing gradient of urea and formamide varying from
40 to 60%. The gels were run for 6 h at 60�C and 150 V.
Statistical analysis
Data was subjected to analysis of variance (ANOVA) using
software SPSS ver. 10 for enzyme assays and Indolic
compounds production in the respective isolates and least
significant difference (LSD) at P = 0.01 among means
compared using standard error.
Results
Selective isolation and enumeration
The streptomycetes population was significantly higher
(120 9 105 cfu g-1 dw soil) in EIF soils compared to WIF
counterpart (48 9 103 cfu g-1 dw soil) (Table 2). The 55
isolates (29 from WIF and 26 from EIF soils) gave whole-
organism hydrolysates rich in LL-A2pm thereby confirming
that they were members of the genus Streptomyces, were
selected for further analysis.
Enzyme assays and PGP attributes
Significant variation was observed among the isolates
(from WIF or EIF soils) with respect to production of
xylanase and cellulase. Significant variation was rule with
regard to numerical dominance of producer of these
Table 2 CFU count and percentage of Streptomyces isolates from WIF and EIF with production of extracellular enzymes and plant growth
promoting activity
Site CFU gm-1 soil Morphotypes % of positive isolates
Xylanase Cellulase Siderophore IAA P-solubilizers Siderophore
WIF 48 9 103 29 13.79 10.34 13.79 13.79 24.13 13.79
EIF 120 9 105 26 46.15 42.30 38.46 50.00 7.69 38.46
WIF water irrigated field
EIF effluent irrigated field
1606 World J Microbiol Biotechnol (2011) 27:1603–1613
123
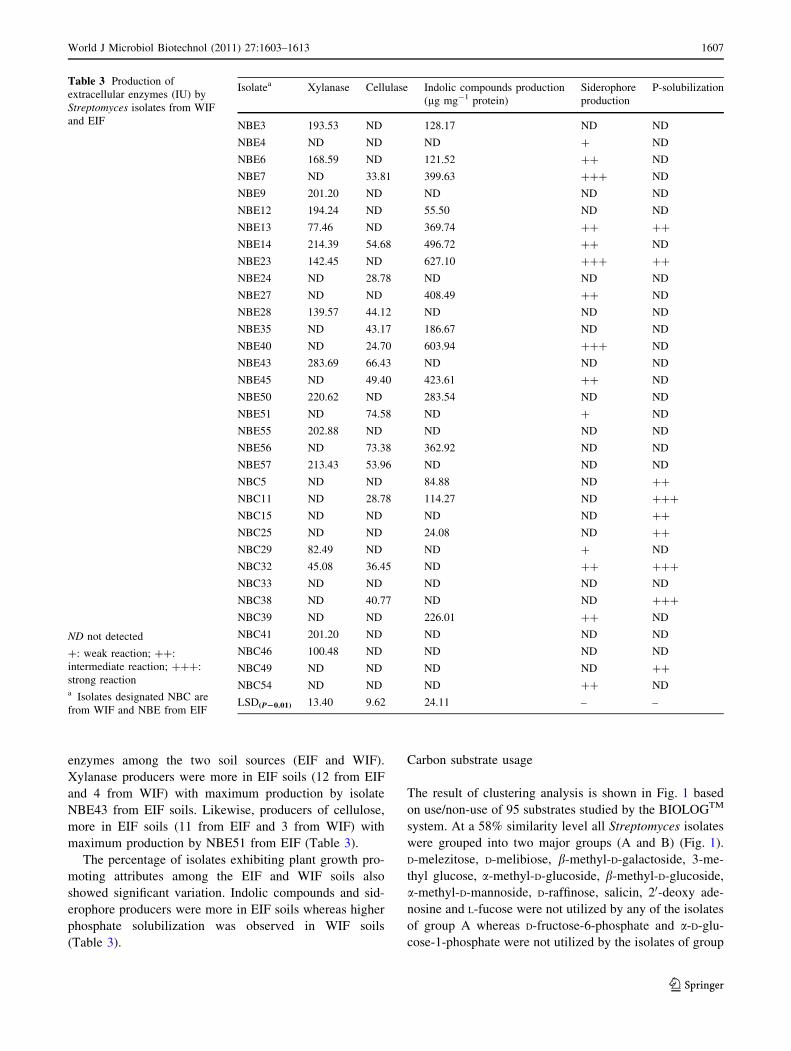
enzymes among the two soil sources (EIF and WIF).
Xylanase producers were more in EIF soils (12 from EIF
and 4 from WIF) with maximum production by isolate
NBE43 from EIF soils. Likewise, producers of cellulose,
more in EIF soils (11 from EIF and 3 from WIF) with
maximum production by NBE51 from EIF (Table 3).
The percentage of isolates exhibiting plant growth pro-
moting attributes among the EIF and WIF soils also
showed significant variation. Indolic compounds and sid-
erophore producers were more in EIF soils whereas higher
phosphate solubilization was observed in WIF soils
(Table 3).
Carbon substrate usage
The result of clustering analysis is shown in Fig. 1 based
on use/non-use of 95 substrates studied by the BIOLOGTM
system. At a 58% similarity level all Streptomyces isolates
were grouped into two major groups (A and B) (Fig. 1).
D-melezitose, D-melibiose, b-methyl-D-galactoside, 3-me-
thyl glucose, a-methyl-D-glucoside, b-methyl-D-glucoside,
a-methyl-D-mannoside, D-raffinose, salicin, 20-deoxy ade-
nosine and L-fucose were not utilized by any of the isolates
of group A whereas D-fructose-6-phosphate and a-D-glu-
cose-1-phosphate were not utilized by the isolates of group
Table 3 Production of
extracellular enzymes (IU) by
Streptomyces isolates from WIF
and EIF
ND not detected
?: weak reaction; ??:
intermediate reaction; ???:
strong reactiona Isolates designated NBC are
from WIF and NBE from EIF
Isolatea Xylanase Cellulase Indolic compounds production
(lg mg-1 protein)
Siderophore
production
P-solubilization
NBE3 193.53 ND 128.17 ND ND
NBE4 ND ND ND ? ND
NBE6 168.59 ND 121.52 ?? ND
NBE7 ND 33.81 399.63 ??? ND
NBE9 201.20 ND ND ND ND
NBE12 194.24 ND 55.50 ND ND
NBE13 77.46 ND 369.74 ?? ??
NBE14 214.39 54.68 496.72 ?? ND
NBE23 142.45 ND 627.10 ??? ??
NBE24 ND 28.78 ND ND ND
NBE27 ND ND 408.49 ?? ND
NBE28 139.57 44.12 ND ND ND
NBE35 ND 43.17 186.67 ND ND
NBE40 ND 24.70 603.94 ??? ND
NBE43 283.69 66.43 ND ND ND
NBE45 ND 49.40 423.61 ?? ND
NBE50 220.62 ND 283.54 ND ND
NBE51 ND 74.58 ND ? ND
NBE55 202.88 ND ND ND ND
NBE56 ND 73.38 362.92 ND ND
NBE57 213.43 53.96 ND ND ND
NBC5 ND ND 84.88 ND ??
NBC11 ND 28.78 114.27 ND ???
NBC15 ND ND ND ND ??
NBC25 ND ND 24.08 ND ??
NBC29 82.49 ND ND ? ND
NBC32 45.08 36.45 ND ?? ???
NBC33 ND ND ND ND ND
NBC38 ND 40.77 ND ND ???
NBC39 ND ND 226.01 ?? ND
NBC41 201.20 ND ND ND ND
NBC46 100.48 ND ND ND ND
NBC49 ND ND ND ND ??
NBC54 ND ND ND ?? ND
LSD(P20.01) 13.40 9.62 24.11 – –
World J Microbiol Biotechnol (2011) 27:1603–1613 1607
123

B. Only one carbon source, D-ribose, was used universally
by all the isolates and sedoheptulosan was the only sub-
strate not utilized by any isolate from both groups.
The substrate usage patterns for the isolates tested
revealed a broad variability, isolate NBE13 from group A
used the fewest substrates, 10/95, about 9.5% of the total
no. of substrates tested. Whereas two isolates NBE7 and
NBE35 from group B used the most, 78/95, about 78.94%
of the substrates.
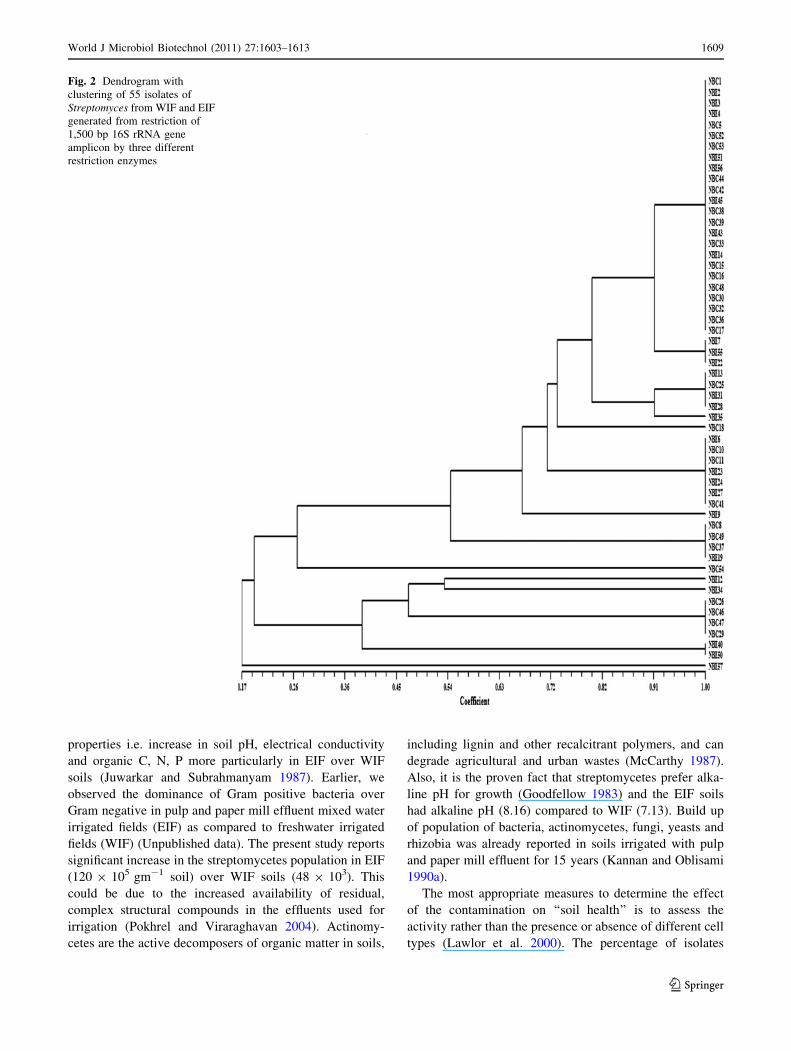
Amplified ribosomal DNA restriction
Restriction digestion of 16S rRNA gene using three
endonucleases (Dde I, Mbo I and Taq I) yielded 7–9 dis-
tinct restriction patterns for each enzyme. About 2–8
restricted fragments of varying sizes were common to each
of the restriction patterns. Cluster analysis of combined
16S rRNA gene restriction pattern based on Jaccard’s
similarity index, grouped all the 55 isolates under 14 dis-
tinct groups with similarity percentage ranging from 17 to
100% (Fig. 2). Majority of the isolates were under group I
(43% of the total number), while the remaining 57% iso-
lates shared rest of groups. ARDRA cluster I, III, VI and
VIII had isolates from both WIF and EIF. ARDRA cluster
II, IV, VII, X, XI, XIII and XIV had isolates only from EIF,
where as cluster V, IX and XII had isolates only from WIF.
16S rRNA gene sequence
For sequence analysis, a total 14 isolates was chosen in
such a way that each of the isolates represents a group with
similar ARDRA pattern as generated by three different
restriction enzymes. The representative isolates from all 14
clusters were identified as S. variabilis, Streptomyces spp.
S. glaucescens, Streptomyces spp., S. viridochromogenes,
S. cinnabarinus, S. aburaviensis, S. viridis, Streptomyces
spp., S. xylophagus, S. macrosporeus, S. thermocarboxy-
dus, Streptomyces spp., and S. albogriseolus (Fig. 3;
Table 4).
Diversity index of streptomycetes
It is evident from different diversity index that EIF
harboured high streptomycetes diversity (H = 2.098 and
1/D = 6.259) relative to WIF (H = 1.419 and 1/D =
2.870).The evenness of the community structure was
higher in EIF (E = 0.874) compared to WIF (E = 0.729)
soils (Table 5).
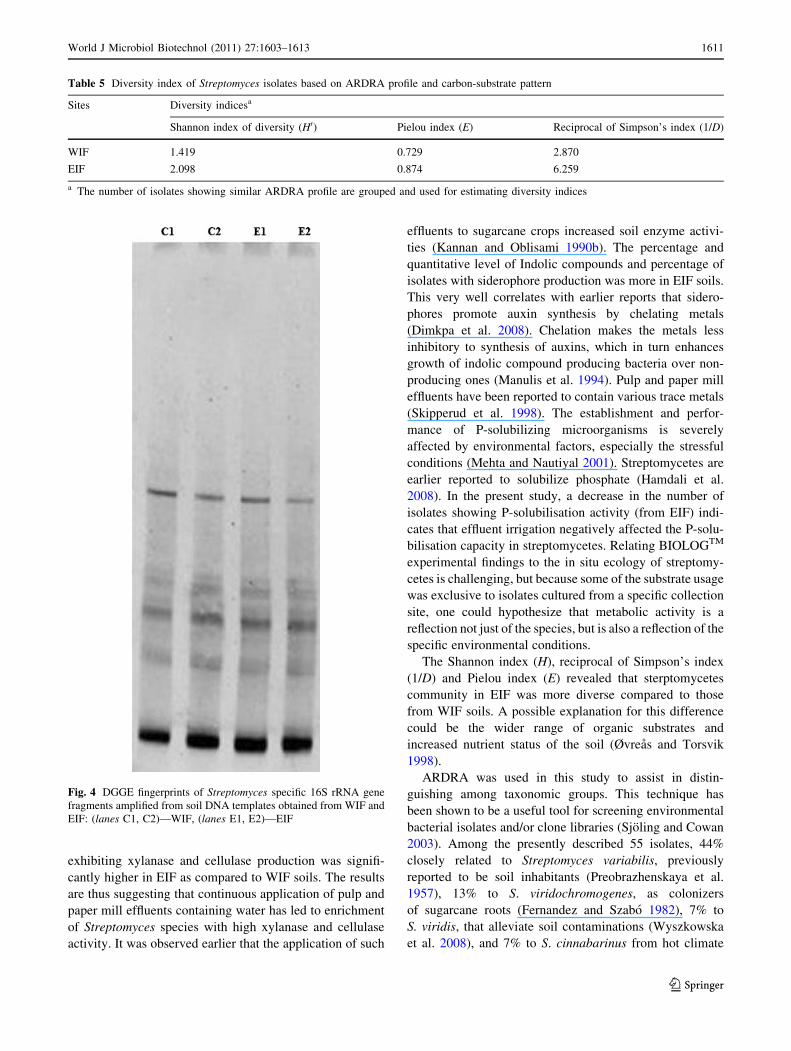
Analysis of streptomycetes DGGE fingerprints
DNA was recovered from both WIF and EIF soil samples.
Streptomycetes community fingerprints were generated
from two replications of each of the sample, and repro-
ducible DGGE profiles of 16S rRNA gene were obtained.
The DGGE profiles were found to be very similar among
the both soils (WIF and EIF). No difference (presence or
absence of bands) could be observed visually when the two
patterns were compared (Fig. 4), indicating that the pre-
dominant unculturable streptomycetes community found in
these soils does not vary regarding the effluent treatment.
Discussion
Pulp and paper mill effluent is usually discharged in local
channel in many parts of India. Sugarcane, rice and wheat
growing regions of Northern Indo-Gangetic plains often
face water scarcity during critical stages of crop growth. In
Lal Kuan, Uttrakhand, India, farmers use water from
channel containing effluent of CPM for irrigating their
sugarcane fields since last 25 years. It is a common practice
near many other paper mills in irrigated agro-ecosystems of
Northern Indo-Gangetic plains, in terms of being the
readily available and inexpensive option to water. How-
ever, irrigation with such type of effluent mixed water also
has the possible environmental side effects; this transports
a wide variety of elements into the environment (Juwarkar
and Subrahmanyam 1987) and in long run this practice
has led to significant changes in soil physicochemical
Fig. 1 Dendrogram showing the clustering of 14 Streptomyces on the
basis of use/non-use of 95 substrates studied by the BIOLOGTM
system). The simple matching (SSM) coefficient and unweighted-pair
group method with average (UPGMA) clustering were used
1608 World J Microbiol Biotechnol (2011) 27:1603–1613
123
properties i.e. increase in soil pH, electrical conductivity
and organic C, N, P more particularly in EIF over WIF
soils (Juwarkar and Subrahmanyam 1987). Earlier, we
observed the dominance of Gram positive bacteria over
Gram negative in pulp and paper mill effluent mixed water
irrigated fields (EIF) as compared to freshwater irrigated
fields (WIF) (Unpublished data). The present study reports
significant increase in the streptomycetes population in EIF
(120 9 105 gm-1 soil) over WIF soils (48 9 103). This
could be due to the increased availability of residual,
complex structural compounds in the effluents used for
irrigation (Pokhrel and Viraraghavan 2004). Actinomy-
cetes are the active decomposers of organic matter in soils,
including lignin and other recalcitrant polymers, and can
degrade agricultural and urban wastes (McCarthy 1987).
Also, it is the proven fact that streptomycetes prefer alka-
line pH for growth (Goodfellow 1983) and the EIF soils
had alkaline pH (8.16) compared to WIF (7.13). Build up
of population of bacteria, actinomycetes, fungi, yeasts and
rhizobia was already reported in soils irrigated with pulp
and paper mill effluent for 15 years (Kannan and Oblisami
1990a).
The most appropriate measures to determine the effect
of the contamination on ‘‘soil health’’ is to assess the
activity rather than the presence or absence of different cell
types (Lawlor et al. 2000). The percentage of isolates
Fig. 2 Dendrogram with
clustering of 55 isolates of
Streptomyces from WIF and EIF
generated from restriction of
1,500 bp 16S rRNA gene
amplicon by three different
restriction enzymes
World J Microbiol Biotechnol (2011) 27:1603–1613 1609
123

NBE57 (GU136398)
Streptomyces xylophagus AB184526
Streptomyces thermocarboxydus EU593727
NBE34 (GQ268015)
NBC47 (GQ268021)
Streptomyces spp. EU368776
NBE6 (GQ268025)
Streptomyces viridochromogenes EU812168
NBE9 (GQ268020)
Streptomyces spp. FJ842682
NBC5 (GQ268022)
Streptomyces variabilis EU841659
NBE40 (GQ268024)
Streptomyces albogriseolus FJ486402
NBC8 (GQ268026)
Streptomyces viridis AB184361
NBE12 (GQ268016)
Streptomyces macrosporeus EF063494
NBC54 (GU136399)
Streptomyces spp. DQ663155
NBC18 (GQ268017)
Streptomyces glaucescens X79322
NBE7 (GQ268018)
Streptomyces spp. EF063473
NBE13 (GQ268019)
Streptomyces cinnabarinus AJ399487
NBE35 (GQ268023)
Streptomyces aburaviensis AB184178
Escherichia coli U00096
100
100
62
100
99
97
67
67
99
96
80
9399
49
81
92
99
63
72
56
33
66
4453
74
71
0.02
Fig. 3 Phylogenetic tree based
on the 16S rRNA gene
sequences of Streptomycesisolates and their closest
phylogenetic relatives. The tree
was created by the neighbour-
joining method. The numbers on
the tree indicate the percentage
of bootstrap sampling derived
from 1,000 replicates
Table 4 Phylogenetic
affiliations of Streptomycesisolates from WIF and EIF
a Species identified based on
16S rRNA gene sequence
similarity ([97%) of
representative isolateb Percent similarity of the
sequence in BLAST result
Site Representative isolate 16S rRNA sequence homology
Species identified NCBI accession numbera % homologyb
WIF NBC5 Streptomyces variabilis GQ 268022 99
NBC8 Streptomyces viridis GQ 268026 99
NBC18 Streptomyces glaucescens GQ 268017 99
NBC47 Streptomyces spp. GQ 268021 99
NBC54 Streptomyces spp. GU 136399 99
EIF NBE6 Streptomyces viridochromogenes GQ 268025 97
NBE7 Streptomyces spp. GQ 268018 97
NBE9 Streptomyces spp. GQ 268020 100
NBE12 Streptomyces macrosporeus GQ 268016 99
NBE13 Streptomyces cinnabarinus GQ 268019 97
NBE34 Streptomyces thermocarboxydus GQ 268015 99
NBE35 Streptomyces aburaviensis GQ 268023 98
NBE40 Streptomyces albogriseolus GQ 268024 99
NBE57 Streptomyces xylophagus GU 136398 99
1610 World J Microbiol Biotechnol (2011) 27:1603–1613
123
exhibiting xylanase and cellulase production was signifi-
cantly higher in EIF as compared to WIF soils. The results
are thus suggesting that continuous application of pulp and
paper mill effluents containing water has led to enrichment
of Streptomyces species with high xylanase and cellulase
activity. It was observed earlier that the application of such
effluents to sugarcane crops increased soil enzyme activi-
ties (Kannan and Oblisami 1990b). The percentage and
quantitative level of Indolic compounds and percentage of
isolates with siderophore production was more in EIF soils.
This very well correlates with earlier reports that sidero-
phores promote auxin synthesis by chelating metals
(Dimkpa et al. 2008). Chelation makes the metals less
inhibitory to synthesis of auxins, which in turn enhances
growth of indolic compound producing bacteria over non-
producing ones (Manulis et al. 1994). Pulp and paper mill
effluents have been reported to contain various trace metals
(Skipperud et al. 1998). The establishment and perfor-
mance of P-solubilizing microorganisms is severely
affected by environmental factors, especially the stressful
conditions (Mehta and Nautiyal 2001). Streptomycetes are
earlier reported to solubilize phosphate (Hamdali et al.
2008). In the present study, a decrease in the number of
isolates showing P-solubilisation activity (from EIF) indi-
cates that effluent irrigation negatively affected the P-solu-
bilisation capacity in streptomycetes. Relating BIOLOGTM
experimental findings to the in situ ecology of streptomy-
cetes is challenging, but because some of the substrate usage
was exclusive to isolates cultured from a specific collection
site, one could hypothesize that metabolic activity is a
reflection not just of the species, but is also a reflection of the
specific environmental conditions.
The Shannon index (H), reciprocal of Simpson’s index
(1/D) and Pielou index (E) revealed that sterptomycetes
community in EIF was more diverse compared to those
from WIF soils. A possible explanation for this difference
could be the wider range of organic substrates and
increased nutrient status of the soil (Øvreas and Torsvik
1998).
ARDRA was used in this study to assist in distin-
guishing among taxonomic groups. This technique has
been shown to be a useful tool for screening environmental
bacterial isolates and/or clone libraries (Sjoling and Cowan
2003). Among the presently described 55 isolates, 44%
closely related to Streptomyces variabilis, previously
reported to be soil inhabitants (Preobrazhenskaya et al.
1957), 13% to S. viridochromogenes, as colonizers
of sugarcane roots (Fernandez and Szabo 1982), 7% to
S. viridis, that alleviate soil contaminations (Wyszkowska
et al. 2008), and 7% to S. cinnabarinus from hot climate
Table 5 Diversity index of Streptomyces isolates based on ARDRA profile and carbon-substrate pattern
Sites Diversity indicesa
Shannon index of diversity (H0) Pielou index (E) Reciprocal of Simpson’s index (1/D)
WIF 1.419 0.729 2.870
EIF 2.098 0.874 6.259
a The number of isolates showing similar ARDRA profile are grouped and used for estimating diversity indices
Fig. 4 DGGE fingerprints of Streptomyces specific 16S rRNA gene
fragments amplified from soil DNA templates obtained from WIF and
EIF: (lanes C1, C2)—WIF, (lanes E1, E2)—EIF
World J Microbiol Biotechnol (2011) 27:1603–1613 1611
123

soils (Preobrazhenskaya et al. 1957). Streptomyces ther-
mocarboxydus, S. aburaviensis, S. macrosporeus, S. xyl-
ophagus and S. albogriseolus were specifically prevalent in
EIF soils. Earlier workers reported S. thermocarboxydus
from composts, soils and sewage (O’Donnell et al. 1993)
whereas S. xylophagus is a well studied xylanase producing
actinomycetes (Tangnu et al. 1981). One isolate was found
similar to S. glaucescens from WIF, reported to be the soil
inhabitant by Preobrazhenskaya et al. (1957). These
observations collectively suggest that irrigation of agri-
cultural fields with pulp and paper mill effluent exceeding
20 years in succession has altered the genetic and meta-
bolic diversity of streptomycetes. Oved et al. (2001) also
observed the significant and consistent shift in population
composition of ammonia oxidizing bacteria as a result of
effluent irrigation with the enrichment of Nitorosmonas-
like population. The DGGE fingerprints obtained for the
streptomycetes communities analyzed here hardly varied,
regardless of effluent treatment (Fig. 4). It indicates that
the effect of effluent irrigation on the dominant bands
observed in DGGE analysis of DNA extracted directly
from soil was minimal compared to the effects on the
culturable portion of the community. This observation
supports the previous studies that readily culturable bac-
teria are probably the largest, most active prokaryotes in a
given sample (Bakken 1997) and so provide a useful, rapid
assessment of biological responses to determining the
impact of anthropogenic activity.
This study indicated that pulp and paper mill effluent
contamination did not have a significant effect on the total
genetic diversity of streptomycetes but affected physio-
logical and metabolic status, so that the number of strep-
tomycetes isolates capable of responding to laboratory
culture and their taxonomic distribution were altered. Thus,
it appears that plate counts may be a more appropriate
method for determining the effect of effluent contamination
on streptomycetes than culture-independent approaches.
Streptomyces isolates in this study could be further
exploited for commercial production of enzymes of bio-
mass degradation as well as promoting plant growth in
such soils.
Acknowledgments This research was conducted with funds pro-
vided by the Indian Council of Agricultural Research under Network
Project on Application of Microorganisms in Agriculture and Allied
Sectors.
References
Bakken LR (1997) Culturable and nonculturable bacteria in soil. In:
van Elsas JD, Trevors JT, Wellington EMH (eds) Modern soil
microbiology. Marcel Dekker, New York, pp 47–61
Dimkpa CO, Svatos A, Dabrowska P, Schmidt A, Boland W, Kothe E
(2008) Involvement of siderophores in the reduction of metal
induced inhibition of auxin synthesis in Streptomyces spp.
Chemosphere 74:19–25
Fernandez C, Szabo IM (1982) Studies on the selection of strepto-
mycetes in the rhizoplane of sugar-cane. Folia Microbiol
27:423–427
Goodfellow M (1983) Ecology of actinomycetes. Annu Rev Micro-
biol 37:189–216
Gordon AS, Weber RP (1951) Colorimetric estimation of indole
acetic acid. Plant Physiol 26:192–195
Hamdali H, Bouizgarne B, Hafidi M, Lebrihi A, Virolle MJ,
Ouhdouch Y (2008) Screening for rock phosphate solubilizing
actinomycetes from Moroccan phosphate mines. Appl Soil Ecol
38:12–19
Heuer H, Krsek M, Baker P, Smalla K, Wellington EMH (1997)
Analysis of actinomycetes community by specific amplification
of genes encoding 16S rRNA and gel electrophoresis separation
in denaturing gradient. Appl Environ Microbiol 63:3233–3241
Howe J, Wagner MR (1996) The effect of paper mill wastewater
irrigation and gypsum soil amendments on sodium accumulation
by cottonwood and soil. Agric Water Manage 31:295–306
Joshi HC (2008) Utilization of treated effluent from paper mills for
crop irrigation and its impact on crop productivity and soil
health. Project Final Report submitted from Division of Envi-
ronmental Sciences, Indian Agricultural Research Institute, New
Delhi, India to Central Pollution Control Board, New Delhi,
India
Juwarkar AS, Subrahmanyam PVR (1987) Impact of pulp and paper
mill wastewater on crop and soil. Water Sci Technol 19:693–700
Kannan K, Oblisami G (1990a) Influence of irrigation with pulp and
paper mill effluent on soil chemical and microbiological
properties. Biol Fertil Soils 10:197–201
Kannan K, Oblisami G (1990b) Influence of paper mill effluent
irrigation on soil enzyme activities. Soil Biol Biochem 22:
923–926
Kieser T, Bibb MJ, Buttner M, Chater KF, Hopwood DA (2000)
Practical Streptomyces genetics. The John Innes Foundation,
Norwich, pp 162–170
Lawlor K, Knight BP, Barbosa-Jefferson VL, Lane PW, Lilley AK,
Paton GI, McGrath SP, O’Flaherty SM, Hirsch PR (2000)
Comparison of methods to investigate microbial populations in
soils under different agricultural management. FEMS Microbiol
Ecol 33:129–137
Magurran E (1988) Ecological diversity and its measurement.
Princeton University Press, Princeton
Manulis S, Shafrir H, Epstein E, Lichterl A, Barash I (1994)
Biosynthesis of indole-3-acetic acid via the indole-3-acetarnide
pathway in Streptomyces spp. Microbiology 140:1045–1050
Massol-Deya AA, Odelson DA, Hickey RF, Tiedje JM (1995)
Bacterial community fingerprinting of amplified 16S and 16–23S
ribosomal DNA gene sequences and restriction endonuclease
analysis (ARDRA). In: Akkermans ADL, van Elsas JD, de
Bruijn FJ (eds) Molecular microbial ecology. Kluwer Academic
Publishers, Dordrecht
McCarthy AJ (1987) Lignocellulose-degrading actinomycetes. FEMS
Microbiol Rev 46:145–163
Mehta S, Nautiyal CS (2001) An efficient method for qualitative
screening of phosphate solubilizing bacteria. Curr Microbiol
43:51–56
Nei M, Li WH (1979) Mathematical model for studying genetic
variation in terms of restriction endonucleases. Proc Natl Acad
Sci USA 76:5269–5273
O’Donnell AG, Falconer C, Goodfellow M, Ward AC, Williams E
(1993) Biosystematics and diversity amongst novel carboxydo-
trophic actinomycetes. Antonie van Leeuwenhoek 64:325–340
1612 World J Microbiol Biotechnol (2011) 27:1603–1613
123

Oved T, Shaviv A, Goldrath T, Mandelbaum RT, Minz D (2001)
Influence of effluent irrigation on community composition and
function of ammonia-oxidizing bacteria in soil. Appl Environ
Microbiol 67:3426–3433
Øvreas L, Torsvik V (1998) Microbial diversity and community
structure in two different agricultural soil communities. Micro-
bial Ecol 36:303–315
Pielou EC (1969) An introduction to mathematical ecology. Wiley,
London
Pokhrel D, Viraraghavan T (2004) Treatment of pulp and paper mill
wastewater—a review. Sci Total Environ 333:37–58
Pospiech A, Neumann B (1995) A versatile quick-prep of genomic
DNA from gram-positive bacteria. Trends Genet 11:217–218
Preobrazhenskaya TP, Ryabova ID, Blinov NO (1957) Problems of
classification of actinomycetes-antagonists. In: Gauze GF,
Preobrazhenskaya TP, Kudrina ES, Blinov NO, Ryabova ID,
Sveshnikova MA (eds) Government publishing house of medical
literature. Medgiz, Moscow, pp 1–398
Ramachandra M, Crawford DL, Pometto AL III (1987) Extracellular
enzyme activities during lignocellulose degradation by Strepto-myces spp.: a comparative study of wild-type and genetically
manipulated strains. Appl Environ Microbiol 53:2754–2760
Rintala H, Nevalainen A, Ronka E, Suutari M (2001) PCR primers
targeting the 16S rRNA gene for the specific detection of
streptomycetes. Mol Cell Probes 15:337–347
Saitou N, Nei M (1987) The neighbour joining method a new method
for reconstructing phylogenetic trees. Mol Biol Evol 4:406–425
Sanghi A, Garg N, Sharma J, Kuhar K, Kuhad RC, Gupta VK (2007)
Optimization of xylanase production using inexpensive agro-
residues by alkaliphilic Bacillus subtilis ASH in solid state
fermentation. World J Microbiol Biotechnol 24:633–640
Schwyn B, Neilands JB (1987) Universal chemical assay for the
detection and determination of siderophores. Anal Biochem
160:47–56
Shannon CE, Weaver W (1949) The mathematical theory of
communication. University of Illinois Press, Urbana
Shatoury SE, Mitchell J, Bahgat M, Dewedar A (2004) Biodiversity
of actinomycetes in a constructed wetland for industrial effluent
treatment. Actinomycetologica 18:1–7
Singh P, Thakur IS (2006) Colour removal of anaerobically treated
pulp and paper mill effluent by microorganisms in two steps
bioreactor. Bioresour Technol 97:218–223
Sjoling S, Cowan DA (2003) High 16S rDNA bacterial diversity in
glacial meltwater lake sediment, Bratina Island, Antarctica.
Extremophiles 7:275–282
Skipperud L, Salbu B, Hagebo E (1998) Speciation of trace elements
in discharges from the pulp industry. Sci Total Environ 217:
251–256
Sneath PHA, Sokal RR (1973) Numerical taxonomy. The principles
and practice of numerical classification. Freeman, Williams and
Wilkins Co, San Francisco
Sokal RR, Michener CD (1958) A statistical method for evaluating
systematic relationships. Univ Kansas Sci Bull 38:1409–1438
Stackebrandt E, Rainey FA, Ward-Rainey NL (1997) Proposal for a
new hierarchic classification system, Actinobacteria classis nov.
Int J Syst Bacteriol 47:479–491
Staneck JL, Roberts GD (1974) Simplified approach to identification
of aerobic actinomycetes by thin-layer chromatography. Appl
Environ Microbiol 28:226–231
Tamura K, Dudley J, Nei M, Kumar S (2007) MEGA4: molecular
evolutionary genetics analysis (MEGA) software version 4.0.
Mol Biol Evol 24:1596–1599
Tangnu SK, Blanch HW, Wilke CR (1981) Production of xylanase by
Streptomyces xylophagus nov. sp. Acta Biotechnol 1:31–40
Thompson JD, Higgins DG, Gibson TJ (1994) CLUSTAL W:
improving sensitivity of progressive multiple sequence align-
ments through sequence weighing, position-specific gap penal-
ties and weight matrix choice. Nucleic Acids Res 22:4673–7680
Udayasoorian C, Prabu PC, Mini K (2004) Influence of composted
bagasse pith and treated paper mill effluent irrigation on
groundnut. Madras Agric J 91:126–129
Wyszkowska J, Kucharski M, Kucharski J (2008) Role of actinomy-
ces of the genus Streptomyces in alleviating the effects of soil
contamination with diesel oil. Pol J Nat Sci 23(3):709–717
Zvereva EA, Fedorova TV, Kevbrin VV, Zhilina TN, Rabinovich ML
(2006) Cellulase activity of a haloalkaliphilic anaerobic bacte-
rium, strain Z-7026. Extremophiles 10:53–60
World J Microbiol Biotechnol (2011) 27:1603–1613 1613
123