galpha3 and protein kinase a represent cross-talking pathways for gene expression in dictyostelium...
TRANSCRIPT

Develop. Growth Differ.
(2002)
44
, 457–465
G
�
3 and protein kinase A represent cross-talking pathways for gene expression in
Dictyostelium discoideum
Maureen A. Brandon,
1
* Dana C. Mahadeo
2
and Gregory J. Podgorski
3
1
Department of Biological Sciences, Campus Box 8007, Idaho State University, Pocatello, ID 83209,
2
Department of Biochemistry and Molecular Biology, Wayne State University, Detroit, MI 48201 and
3
Department of Biology, Utah State University, Logan, UT 84322, USA.
Heterotrimeric G proteins and protein kinase A (PKA) are regulators of development in
Dictyosteliumdiscoideum
. It has been reported that disruption of the
Dictyostelium
G
�
3 gene (
g
�
3
–
) blocks development andexpression of several early development genes, characteristics that are reminiscent of mutants lacking thecatalytic subunit of PKA (
pkac
–
). The hypothesis that G
�
3 and PKA signaling pathways may interact to controldevelopmental gene expression was tested by comparing the regulation of seven genes expressed early indevelopment in the wild-type and in
g
�
3
–
and
pkac
–
mutants, and comparing PKA activity in the wild-type andin a
g
�
3
–
mutant. The expression patterns of six genes were affected similarly by the G
�
3 and PKA mutations,while the expression of only one gene, the cAMP receptor 1 (
cAR1
), differed between the mutants. PKA activity,measured by phosphorylation of the PKA-specific substrate Kemptide, was higher in
g
�
3
–
cells than in wild-type cells, suggesting that G
�
3 normally exerts an inhibitory effect on PKA activity. Although some earlydevelopment genes appear to require both G
�
3 and PKA for expression, the differing response of cAR1expression and the inhibitory effect of G
�
3 on PKA activity suggest that G
�
3 and PKA are members of inter-acting pathways controlling gene expression early in development.
Key words:
Dictyostelium discoideum
, gene regulation, G-protein, protein kinase A, signal transduction.
Introduction
Heterotrimeric G proteins are ubiquitous, multifacetedregulators of cell physiology and development. Onceactivated by their cognate receptors, G proteinssignal through both G
�
and G
��
subunits to a diversearray of downstream effectors that include manyisoforms of adenylyl cyclase, phospholipase C,protein kinase C, phosphodiesterases, tyrosinekinases, elements of the ras/raf/MAPK pathway,phosphatidyl inositol 3-kinases and ion channels(Hamm 1998). In mammals, the 20 known
�
, 6
�
and12
�
subunits assemble in different combinations intoan array of heterotrimeric G proteins. The web ofinteractions between the extended family of receptors,the combinatorial ensemble of heterotrimeric Gproteins and their diverse effectors has only recentlybeen appreciated.
The simple eukaryote
Dictyostelium discoideum
isan experimental model for the study of G proteinfunction (Parent & Devreotes 1996). This soil-dwelling
microorganism undergoes a developmental cycle asa protective response to stressful environments.Development is triggered by nutrient deprivation andproduces a mass of environmentally resistant sporessupported above the soil by a slender stalk. Duringdevelopment, cells respond to a host of molecularsignals, many of which are ligands of G-protein-coupled receptors, including extracellular cAMP(Devreotes 1994). G-protein coupled receptorsactivate a family of developmentally regulated hetero-trimeric G proteins in
Dictyostelium
. There are nineknown
Dictyostelim
G
�
subunits and one each of
�
and
�
subunits (Lilly
et al
. 1993; Kim
et al
. 1996;Zhang
et al
. 2001; J. Brzostowski and A. Kimmel,pers. comm. 2000). The phenotypes of mutants withdisruptions of various G-protein subunit genes, suchas G
�
2 and G
�
, clearly demonstrate that G proteinsare essential regulators of developmental geneexpression (Kim
et al
. 1996).We have previously reported that a strain of cells
containing a targeted disruption of the
Dictyostelium
G
�
3 gene (
g
�
3
–
cells) are blocked in aggregation andthe expressions of several early development genesare aberrant (Brandon & Podgorski 1997; Brandon
et al
. 1997). These characteristics are reminiscent ofmutants lacking the catalytic subunit of protein kinase
*Author to whom all correspondence should be addressed.Email: [email protected] 1 May 2002; revised 29 May 2002; accepted 20 July
2002.

458 M. A. Brandon
et al.
A (PKA; Mann & Firtel 1991; Schulkes & Schaap 1995;Mann
et al
. 1997). The
Dictyostelium
PKA is a hetero-dimer composed of a catalytic polypeptide (PKAC)that contains the kinase and an inhibitory regulatorypolypeptide (PKAR) that associates with the catalyticsubunit in the absence of cAMP. In addition to
Dictyostelium
, PKA has been shown to controldevelopment in
Drosophila
(Pan & Rubin 1995) andthe zebrafish (Hammerschmidt
et al
. 1996).The similarities between the phenotypes of the
g
�
3
–
and
pkac
–
mutants suggest that G
�
3 and PKA signal-ing pathways may be linked to control early develop-mental gene expression. In this investigation, weexplore the relationship between G
�
3 and PKA byexamining how mutations of these genes affectexpression of a more comprehensive set of earlydevelopment genes, and how loss of G
�
3 affects PKAactivity.
Methods
Strains, growth, development and RNA analysis
The strains used in this study are shown in Table 1.Cells were grown in liquid culture and starved asdescribed previously in Brandon & Podgorski (1997).HL5 medium for HPS400 and JH10 was supple-mented with 100
µ
g/mL thymidine. Total RNA wasextracted according to the method of Chomczynski &Sacchi (1987) from cells starved in non-nutrient sus-pension cultures shaken without treatment or withtreatment by 50 n
M
cAMP pulses every 6 min. cAMPpulses mimic cAMP signaling in wild-type cells. Formutant strains such as
g
�
3-
and
pkac-
that arearrested early in development due to lack of endog-enous cAMP pulses, the exogenous pulses mayinduce expression of genes necessary for later stagesof development. Gene expression was measuredusing northern blots as described by Brandon &Podgorski (1997), to examine mRNA abundance.
Assay of protein kinase A activity
Starving cells were shaken in non-nutrient buffer eitherwith or without added cAMP pulses (Brandon &
Podgorski 1997). At 3 h intervals between 0 and 6 h ofstarvation, 4
�
10
7
cells were removed, pelleted, andresuspended in 100
µ
L ice-cold lysis buffer (20 m
M
Tris-HCl pH 7.4, 10 m
M
MgCl
2
, 1 m
M
ATP, 0.5
µ
g/mLantipain, 1
µ
g/mL leupeptin, 1
µ
g/mL aprotinin). Cellsuspensions were lysed by rapidly pushing through a5
µ
m polycarbonate membrane (Poretics, Livermore,CA, USA). The cold lysates were centrifuged at13 000
g
for 15 s to remove membrane fragments andunlysed cells. Cell lysate (10
µ
L) was assayed for PKAactivity using a PepTag PKA assay kit according to themanufacturer’s directions (Promega, Madison, WI,USA). In addition to lysate, the 25
µ
L reactions con-tained 2 mg of the fluorescent-labeled PKA-specificsubstrate Kemptide diluted in lysis buffer without pro-tease inhibitors. The reaction was incubated for 2 minin a 22
�
C water bath. The reaction was terminated byrapid freezing in a dry ice/ethanol bath. Samples werestored at
–
70
�
C until analysis. Phosphorylated andunphosphorylated forms of Kemptide were separatedby electrophoresis of the reactions for 1 h at 120 V on
Fig. 1.
Independence of G
�
3 and protein kinase A (PKA) geneexpression. RNA blots were prepared using RNA obtained every2 h from cultures of HPS400,
g
�
3
–
or
pkac
–
cells which eitherreceived (+) or did not receive (–) exogenous cAMP pulsesduring 6 h of starvation. The blots were hybridized to radio-labeled coding region fragments of PKA catalytic peptide(PKAC; HPS400 and
g
�
3
–
cells) or G
�
3 (HPS400 and
pkac
–
cells) and the bands were visualized by autoradiography.
Table 1.
Summary of
Dictyostelium discoideum
strains used in the study
StrainRelevantgenotype Disrupted gene
Isogenicparent
Developmentalphenotype Reference
HPS400 tmp600 (None) HPS83 Wild-type Podgorski & Deering (1984)DH1
�pyr5–6 Uridine monophosphate synthase AX3K Wild-type Caterina et al. (1994)JH10 thy1::PYR5–6 Thymidine biosynthetic gene 1 JH8 Wild-type Hadwiger & Firtel (1992)G3T2 g�3::THY1 G�3 HPS400 agg– Brandon et al. (1997)PKAC pkacat::THY1 Protein kinase A catalytic subunit JH10 agg– Mann et al. (1997)
Interacting pathways for gene expression 459
0.8% agarose gels prepared in 50 mM Tris (pH 8.0).Fluorescent-labeled Kemptide was visualized by ultra-violet (UV) illumination (312 nm) and photographedusing a digital camera. The amount of phosphorylatedKemptide was quantified using a Kodak 1D imageanalysis system (Kodak, Rochester, NY, USA) andreported as net intensity/min per mg total cellularprotein.
Results
Independence of G�3 and PKA gene expression
The evidence that both G�3 and PKA signal transduc-tion pathways control gene expression prompted us totest whether G�3 controls expression of the PKAsubunits, and vice versa. Figure 1 shows a compari-son of the expression of the PKA catalytic subunit inthe g�3– mutant and wild-type HPS400 cells. Expres-
sion of PKAC in g�3– cells is similar to that in HPS400cells, which indicates that PKAC is expressed in cellswith a non-functional G�3 gene.
G�3 expression in the pkac– mutant is also shownin Figure 1. In HPS400 cells, G�3 expression isinduced rapidly by 2 h of starvation, then graduallydeclines. Treating the HPS400 cells with artificialcAMP pulses reduces G�3 expression to undetecta-ble levels after 4 h. This indicates that G�3 is a cAMPpulse-repressed gene. In pkac– cells that did notreceive artificial cAMP pulses, G�3 expression contin-ues to increase through 6 h of starvation, a time whenthe level of G�3 mRNA decreases in HPS400 cells. Inpkac– cells which receive cAMP pulses, G�3 mRNAexpression rises with increasing time of starvation. Incontrast, G�3 mRNA is absent at this time in HPS400cells. These results demonstrate that the signalingpathway that activates G�3 expression does not
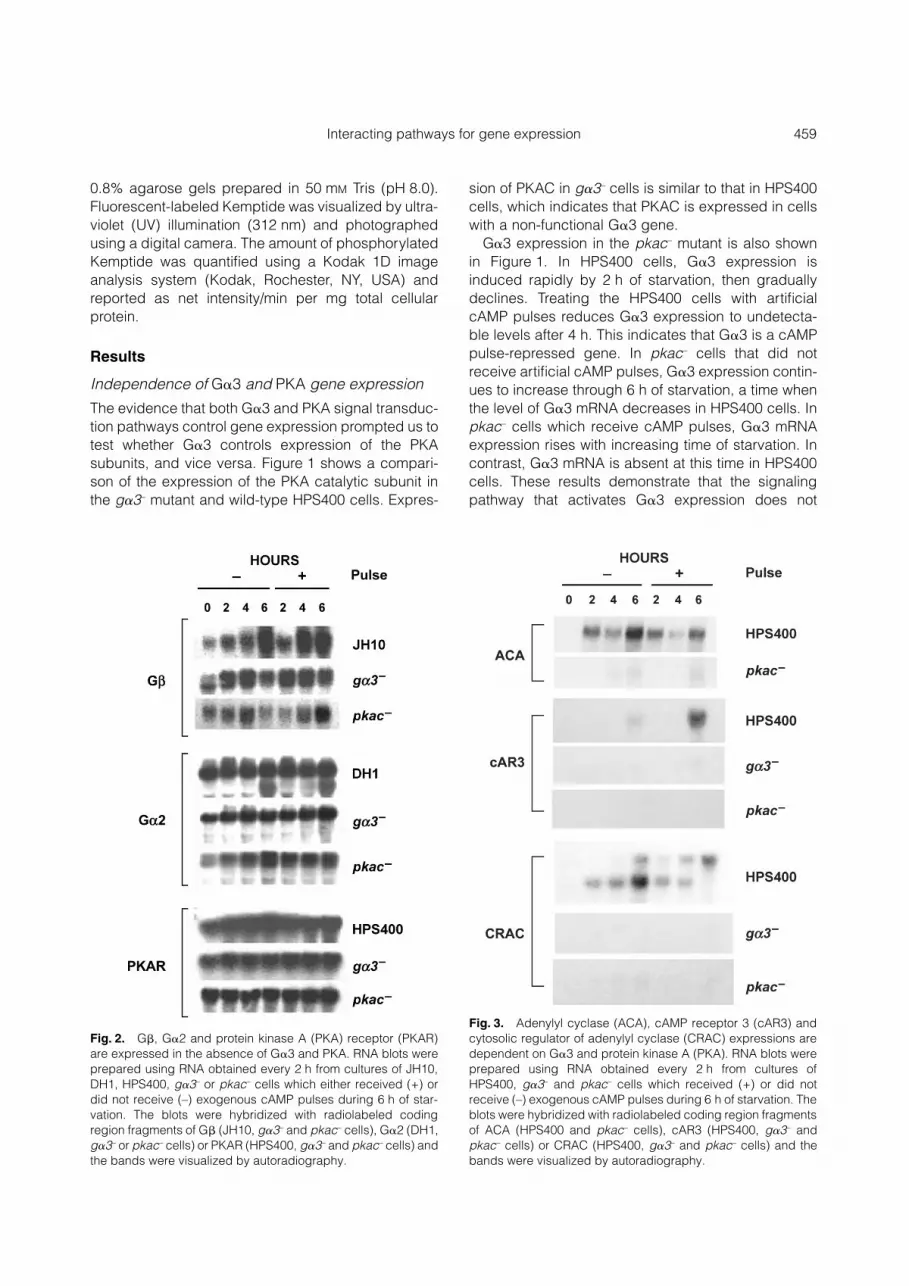
Fig. 2. G�, G�2 and protein kinase A (PKA) receptor (PKAR)are expressed in the absence of G�3 and PKA. RNA blots wereprepared using RNA obtained every 2 h from cultures of JH10,DH1, HPS400, g�3– or pkac– cells which either received (+) ordid not receive (–) exogenous cAMP pulses during 6 h of star-vation. The blots were hybridized with radiolabeled codingregion fragments of G� (JH10, g�3– and pkac– cells), G�2 (DH1,g�3– or pkac– cells) or PKAR (HPS400, g�3– and pkac– cells) andthe bands were visualized by autoradiography.
Fig. 3. Adenylyl cyclase (ACA), cAMP receptor 3 (cAR3) andcytosolic regulator of adenylyl cyclase (CRAC) expressions aredependent on G�3 and protein kinase A (PKA). RNA blots wereprepared using RNA obtained every 2 h from cultures ofHPS400, g�3– and pkac– cells which received (+) or did notreceive (–) exogenous cAMP pulses during 6 h of starvation. Theblots were hybridized with radiolabeled coding region fragmentsof ACA (HPS400 and pkac– cells), cAR3 (HPS400, g�3– andpkac– cells) or CRAC (HPS400, g�3– and pkac– cells) and thebands were visualized by autoradiography.

460 M. A. Brandon et al.
depend on PKA. However, the cAMP signal whichrepresses G�3 expression must be transmittedthrough PKA.
Effects of G�3 and PKAC mutations on developmental gene expression
The expression patterns of a set of genes that functionin early development were compared in the g�3– andpkac– mutant strains. This set includes G�2, G�, theregulatory subunit of PKA (PKAR), adenylyl cyclase(ACA), cAMP receptors 1 (cAR1) and 3 (cAR3), andthe cytosolic regulator of adenylyl cyclase (CRAC).The results suggest that G�3 and PKA regulate theexpression of these genes in three distinct ways,either: (i) genes are expressed in the absence of G�3and PKA; (ii) gene expression is dependant on G�3and PKA; or (iii) gene expression is affected differentlyby the G�3 and PKA mutations.
Genes expressed in the absence of G�3 and PKA(G�, G�2 and PKAR) Dictyostelium is reported tohave only one G-protein �-subunit gene (Lilly et al.1993). This single G� gene is involved in all G protein-regulated signal transduction pathways, including thecAMP signal relay system that establishes endo-genous cAMP pulses. We examined G� mRNA levelsin the wild-type strain JH10 and in the g�3– and pkac–
mutants. In JH10 cells that did not receive artificialcAMP pulses, G� is expressed in growing cells, andgradually increases through at least 6 h of starvation(Fig. 2). Treating the JH10 cells with artificial cAMPpulses induces maximal G� expression 2 h earlierthan without artificial cAMP pulses. G� expression ing�3– cells is similar to that in JH10 cells except that alower molecular weight transcript is present at theinitiation of starvation (0 h). G� expression is alsopresent in pkac– cells, although the level is less thanin the wild-type. These results indicate that G�
expression is not dependent on G�3 or PKA signaltransduction pathways, but the amount of expressionmay be sensitive to PKA.
The G-protein �-subunit, G�2, is required for theestablishment of the cAMP signal relay system. Wehave previously reported that G�2 is transcribed intoat least four different mRNA (Pupillo et al. 1989). In thewild-type strain DH1, G�2 mRNA is expressed consti-tutively, regardless of whether cells are treated withcAMP (Fig. 2). This supports our earlier report ofconstitutive G�2 protein expression in HPS400 cells(Brandon & Podgorski 1997). The pattern of G�2expression in the g�3– cells is not different to that seenin DH1 cells in the presence or absence of artificialcAMP pulses. However, the levels are lower overall,
and the g�3– cells fail to express a lower molecularweight form at 6 h of starvation. In the pkac– cells, G�2expression resembles that of g�3– cells both in thepresence and absence of exogenous cAMP pulses.The overall similarity in expression pattern for G�2 in
Fig. 4. cAMP receptor 1 (cAR1) mRNA expression is affecteddifferently by G�3 and protein kinase A (PKA) catalytic peptide(PKAC) mutations. (A) RNA blots were prepared from RNAobtained every 2 h from cultures of HPS400, g�3– and pkac–
cells which received (+) or did not receive (–) exogenous cAMPpulses during 6 h of starvation. The blots were probed with aradiolabeled fragment of cAR1 and the bands were visualized byautoradiography. (B) Protein blots were prepared from culturesof HPS400 and pkac– cells treated exactly as described above.The proteins were probed with an antibody to cAR1 and theywere visualized by chemiluminescence. The small molecularweight band at the bottom of each blot is a protein which reactsnon-specifically with the cAR1 antibody and is used to indicateequal protein loading.
Interacting pathways for gene expression 461
all strains indicates that G�2 expression is notdependent on G�3 or PKAC.
Similar to G� and G�2, the regulatory subunit ofPKA (PKAR) is expressed in the g�3– and pkac–
mutants (Fig. 2). In g�3– and pkac– cells, PKAR isexpressed at a constantly high level, regardless oftreatment with cAMP. Although the level of PKAR isless in pkac– cells than that seen in HPS400 cells(similar to G�), these results demonstrate that PKARexpression is not dependent on G�3 or PKAC.Moreover, all components necessary for normal PKAfunction, that is, the PKA catalytic and regulatorysubunits, are present in the g�3– mutant.
Genes with expression dependent on G�3 and PKA(ACA, cAR3 and CRAC) We have shown previouslythat ACA expression and adenylyl cyclase activity areabsent in g�3– cells. However, both gene expressionand enzyme activity can be restored to normal levelswhen the mutant is starved in the presence of artificialcAMP pulses (Brandon & Podgorski 1997). ACAexpression was examined in pkac– cells to determineif it follows a similar pattern (Fig. 3). ACA transcript isnot detected at the initiation of starvation (0 h) inHPS400 cells. It is induced by 2 h of starvation,decreased slightly at 4 h, and increases further by 6 hof starvation. cAMP pulses do not increase ACAexpression. In the pkac– cells, weak ACA expressionis detected at 4 and 6 h of starvation and cAMP pulsesdo not increase expression. Combined with our pre-vious report of ACA gene expression and enzymeactivity (described above), these results suggest thatboth G�3 and PKA are required for ACA expression.
However, unlike the g�3– mutant, ACA expressioncannot be rescued by artificial cAMP pulses in thepkac– cells.
In wild-type HPS400 cells, the cAMP receptor 3(cAR3) gene is expressed, starting at 6 h of starvation.Providing the HSP400 cells with artificial cAMP pulsescauses increased expression of cAR3 at 6 h ofstarvation (Fig. 3). In g�3– cells, cAR3 transcript isnot detectable in cells starved for 6 h withoutcAMP pulses. Treatment of g�3– cells with cAMPpulses for 6 h induces very weak expression ofcAR3. In pkac– cells, no cAR3 transcript is detectedwith or without cAMP pulses. These results demon-strate that cAR3 expression is dependent on G�3and PKA.
The cytosolic regulator of adenylyl cyclase (CRAC)is an essential component of the cAMP signal relaysystem. We found that two CRAC transcripts areexpressed in HPS400 cells (Fig. 3). The smallertranscript is first detected after 2 h of starvationwithout artificial cAMP pulses, and graduallyincreases in abundance through 6 h of starvation. Alarger transcript is detected at 6 h of starvation. cAMPpulses suppress expression of the small transcript at6 h of starvation and advance the expression of thelarge transcript. In g�3– and pkac– cells, weak expres-sion of only the small CRAC transcript is detected.These data suggest that normal CRAC expressionrequires both G�3 and PKA.
CAR1 expression is affected differently by the G�3and PKA mutations We previously reported that thecAMP receptor 1 (cAR1) protein is absent in g�3– cellsthat do not receive cAMP pulses, but present atnormal levels when the cells receive cAMP pulses.(Brandon & Podgorski 1997). In this experiment, wecompare the cAR1 expression patterns in HPS400,g�3– and pkac– strains, and the cAR1 protein levels inHPS400 and pkac– cells.
cAR1 expression in g�3– cells is similar to that seenin HPS400 cells when the cells do not receive cAMPpulses, except that expression at 4 h of starvation isless (Fig. 4a). However, cAMP pulses dramaticallyincrease cAR1 expression in g�3– cells. In contrast,cAR1 expression is greatly reduced in the pkac–
mutant, and is not rescued by cAMP pulses. Similarly,cAR1 protein in the pkac– mutant was not detectedunder any conditions tested (Fig. 4b). Unlike the othergenes we have examined in this study which showsimilar patterns of regulation in the g�3– and pkac–
mutants, cAR1 expression appears independently ofG�3 yet requires PKA. The presence of cAR1 mRNAand lack of detectable cAR1 protein (Brandon &Podgorski 1997) in g�3– cells suggests a relationship
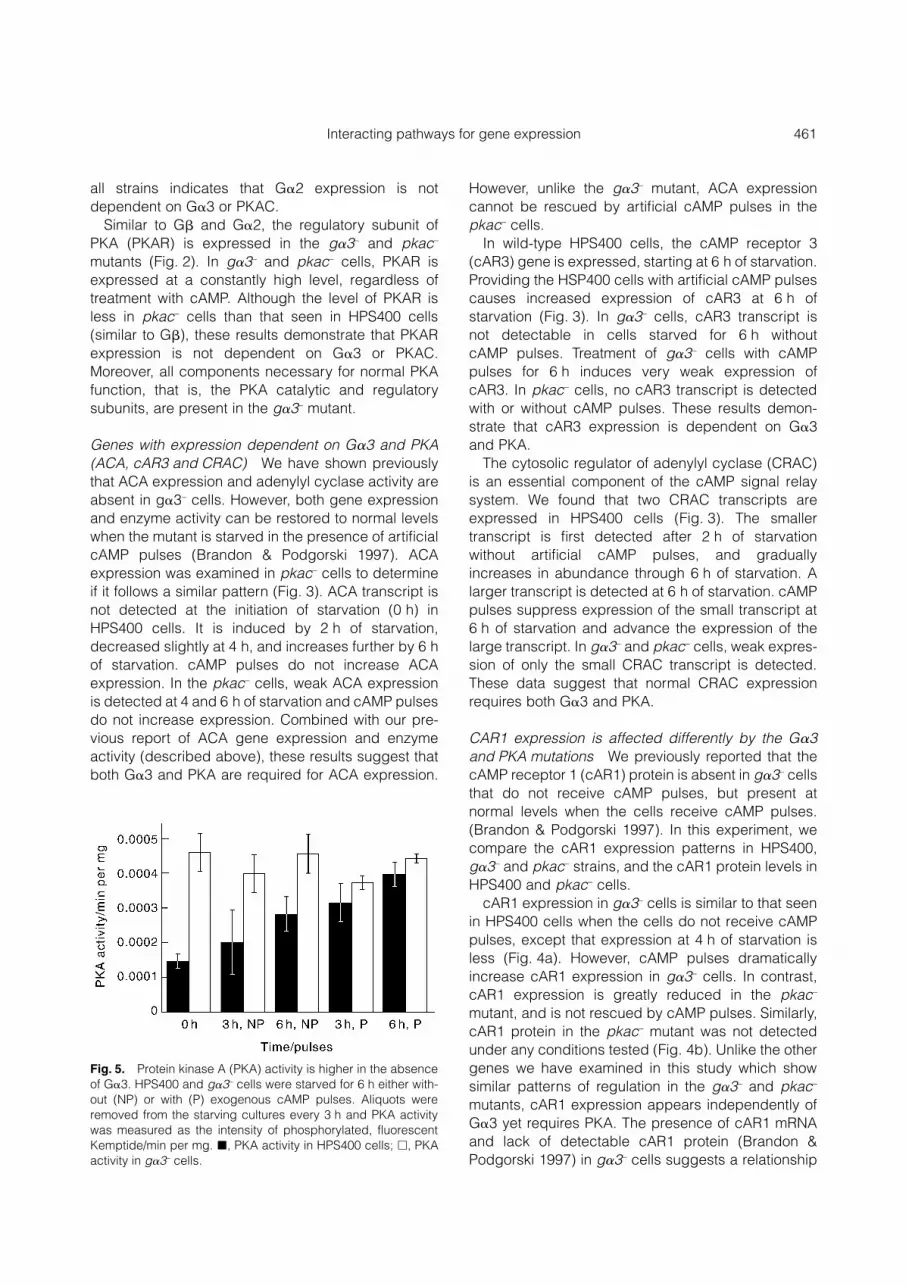
Fig. 5. Protein kinase A (PKA) activity is higher in the absenceof G�3. HPS400 and g�3– cells were starved for 6 h either with-out (NP) or with (P) exogenous cAMP pulses. Aliquots wereremoved from the starving cultures every 3 h and PKA activitywas measured as the intensity of phosphorylated, fluorescentKemptide/min per mg. �, PKA activity in HPS400 cells; �, PKAactivity in g�3– cells.

462 M. A. Brandon et al.
between G�3 and cAR1 mRNA stability or translationefficiency.
Protein kinase A activity is higher in the absence of G�3
The similarities in expression patterns of this group ofsignaling genes in g�3– and pkac– cells suggest thatthe G�3 and PKA signal transduction pathways mayinteract to control gene expression. Because PKA is awell-established regulator of transcription factoractivity, we hypothesized that G�3 functions upstreamof PKA in a signal transduction pathway. To test thishypothesis, we compared PKA activity in g�3– andHPS400 cells.
At the initiation of starvation (0 h), PKA activity inHPS400 cells is approximately 1.5 � 10–4 units/minper mg (Fig. 5, �). The activity gradually increasesthrough 6 h of starvation in the absence of exogenouscAMP pulses to approximately 2.8 � 10–4 units/minper mg (Fig. 5, 3 h and 6 h, NP). Providing theHPS400 cells with artificial cAMP pulses increasesPKA activity further so that at 3 h of starvation withpulses, PKA activity is approximately the same as at6 h without pulses (compare 6 h, NP to 3 h, P). By 6 hof starvation with cAMP pulses, PKA activity isapproximately 4 � 10–4 units/min per mg.
The PKA activity in g�3– cells at the initiation ofstarvation is approximately 4.6 � 10–4 units/min permg (Fig. 5, �). This is approximately threefold greaterthan that of HPS400 at the same developmental stage.Unlike HPS400, the PKA activity in g�3– cells remainsat approximately the same level through 6 h of star-vation and is not altered by the addition of artificialcAMP pulses.
These data suggest that G�3 controls an inhibitorysignal for PKA activity. In addition, increasing the timeof starvation or treatment with exogenous cAMPpulses does not change the PKA activity of g�3– cells,but increases the PKA activity in HPS400 cells to thelevel seen in g�3– cells. This indicates that the PKAactivity in g�3– cells is at a maximum level at thedevelopmental stages measured.
Discussion
In this report we compare the gene expressionpatterns of a group of early signaling genes in twosignal transduction mutants, g�3– and pkac–. Thegene expression patterns we report for G�, G�2,PKAR, PKAC, ACA, cAR1, cAR3, CRAC and G�3 inwild-type cells are similar to the original reports (Kleinet al. 1988; Pupillo et al. 1989; Mann & Firtel 1991; Pittet al. 1992; Simon et al. 1992; Johnson et al. 1993;
Lilly et al. 1993; Insall et al. 1994; Brandon et al.1997). Our results indicate that, with a few exceptions,these early signaling genes respond similarly to thelack of G�3 or PKAC. These results prompted us toexamine the potential connection of G�3 and PKA ina signal transduction pathway by analysis of PKAactivity in the g�3– mutant. We show that PKA activityis significantly higher in the g�3– mutant than in wild-type cells early in the developmental program, beforeendogenous cAMP pulses are established.
A summary of the relationships of G�3 and PKAC onexpression of the different signaling genes we haveexamined, including those previously reported by us,is shown in Table 2. With the exceptions of cAMP-dependent phosphodiesterase (PDE) and cAR1, thedependencies of the gene expression patterns onG�3 or PKA are very similar. The G�, G�2 and PKARgenes are expressed in the g�3– and pkac– mutants.The ACA, cAR3 and CRAC genes require both G�3and PKA for expression, although nearly normal ACAexpression can be achieved by treating the g�3– cellswith exogenous cAMP pulses (Brandon & Podgorski1997). This suggests that, for some aggregation-stagesignaling genes, signals from both G�3 and PKA arerequired for proper gene expression.
Although PDE and cAR1 respond differently to theG�3 and PKAC mutations, their responses are oppo-site. PDE does require G�3, but not PKAC, for expres-sion; cAR1 does require PKAC, but not G�3, forexpression. Apparently normal amounts of cAR1mRNA are present in g�3– cells, but no cAR1 proteinis produced unless the mutant is treated with cAMP
Table 2. Summary of dependence of gene expression patternson G�3 or PKAC
Gene G�3 PKAC
G�3 N/A –PKAC – N/AG� + ++G�2 + ++PKAR + +ACA +++ +++cAR3 +++ +++CRAC +++ +++PDI +++† +++‡
PDE +/++† –‡
cAR1 – +++
–, no dependence; +, slight dependence; ++, moderatedependence; +++, absolute dependence.
†Reported in Brandon & Podgorski (1997); ‡reported in Wuet al. (1995).
ACA, adenylyl cyclase; cAR, cAMP receptor; CRAC,cytosolic regulator of adenylyl cyclase; PDE, cAMP-dependentphosphodiesterase; PDI, phosphodiesterase inhibitor; PKAC,protein kinase A catalytic peptide; PKAR, protein kinase Aregulatory subunit.

Interacting pathways for gene expression 463
pulses (Brandon & Podgorski 1997), suggesting thatthe G�3 mutation interferes with cAR1 translation.These results support separate signaling pathways forG�3 and PKA.
The initial activation of the G�3 and PKA catalyticsubunit genes are not dependent on each other. Thismeans that some other mechanism, functioningbefore the aggregation stage genes ACA, cAR3 andCRAC, turns on the genes that control early pattern-ing. The mechanism that activates G�3 expressionhas yet to be discovered. However, as developmentprogresses, cAMP signaling through PKA is neces-sary to inhibit G�3 expression. A mechanism forcontrolling PKA expression has recently been dis-covered by Souza et al. (1998, 1999). A serine/threonine kinase, YakA, is induced by starvation andappears to control PKA activity by increasing theexpression of PKAC and inhibiting the expression of atranslational regulator of PKAC, PufA. In pufA– cells,the amount of PKAC protein is greater and the cellsaggregate faster than wild-type cells.
Our analysis of PKA activity in the g�3– mutantdemonstrates that, like PufA, G�3 exerts an inhibitoryeffect on PKA activity. However, a comparison of theexpression patterns of G�3 and PufA suggest that theinhibition by G�3 does not involve PufA. PufA isexpressed in growing cells, and decreases by 2–3 hafter starvation at the time when G�3 expressionpeaks. It seems likely that G�3 control of PKA activityfollows that of PufA temporally. Additionally, release ofG�3 inhibition causes at most a threefold increase inPKA activity rather than the fivefold increase in the
absence of PufA. If a fivefold increase is a threshholdvalue for accelerated aggregation, as hypothesizedby Souza et al. (1999), this may explain why the g�3–
mutants do not aggregate while the pufA– mutantsdisplay accelerated aggregation.
We expect that the mechanism of G�3 inhibition ofPKA activity is not direct, but depends on the actionof a component of the G�3 signaling pathway. Theclassical paradigm for G-protein regulation of PKA
Fig. 6. Temporal relationships between changes in G�3 andprotein kinase A (PKA) catalytic peptide (PKAC) expressionsand PKA activity. Starvation causes G�3 and PKAC expressionlevels to rise rapidly. G�3 and PKAC are necessary to induceexpression of cAMP signal relay genes such as cAMP receptor1 (cAR1), adenylyl cyclase (ACA) and cytosolic regulator ofadenylyl cyclase (CRAC), which establish endogenous cAMPpulses. cAMP pulses cause G�3 expression to decrease,allowing PKA activity to rise. Increased PKA activity enhancesthe expression of cAMP signal relay genes, and therefore endo-genous cAMP pulses, which promotes aggregation.
Fig. 7. Model for G�3 and protein kinase A (PKA) control of de-velopmental gene expression. Early in development, starvationinduces the expression of G�3 and PKA catalytic peptide(PKAC). In the case of PKAC, expression is controlled by theserine/threonine kinase YakA. YakA enhances PKAC expressionand inhibits the expression of the PKAC translational regulatorPufA. In the presence of YakA and absence of PufA, PKACexpression is induced. The starvation signal that induces G�3expression is not currently known. G�3 appears to have tworoles in early development: (i) to maintain PKA activity at a lowlevel; and (ii) to work with PKA to induce the expressions ofcAMP receptor 1 (cAR1), adenylyl cyclase (ACA), cytosolicregulator of adenylyl cyclase (CRAC) and cAMP receptor 3(cAR3). The aggregation stage of development requires theestablishment of cAMP pulses, which require the products of thecAR1, ACA, CRAC and cAR3 genes. The presence of cAMPpulses reduces G�3 expression and allows PKA activity to reachmaximum levels.

464 M. A. Brandon et al.
activity is via G-protein regulation of adenylyl cyclaseactivity. In mammalian and insect systems, adenylylcyclase can be stimulated or inhibited dependingon the G-protein �-subunit or the combination of��-subunits with which it interacts (Perrimon 1996;Marinissen & Gutkind 2001). The amount of adenylylcyclase activity determines the amount of cAMP avail-able for binding to the regulatory subunit of PKA andthe subsequent release of active catalytic subunit.Because ACA is the only known Dictyosteliumadenylyl cyclase that is G-protein regulated, and ACAexpression depends on G�3 early in development, itseems unlikely that inhibition of ACA activity by G�3 isthe only mechanism for G�3 inhibition of PKA activity(Söderbom et al. 1999). An alternative hypothesis isprovided by the growing body of evidence frommammalian systems which indicates that phos-phatases can be effectors in G-protein regulatedsignaling systems (Kuschel et al. 1999; Graness et al.2000; Grundker et al. 2001). If G�3 regulates a phos-phatase that removes phosphates from PKA sub-strates, PKA activity would appear to be inhibited inour assay. This potential G�3-regulated phosphatasemay represent the point of interaction between theG�3 and PKA signaling pathways, and could accountfor the divergent effects of the G�3 mutation reportedhere.
Our results also demonstrate that G�3 is a cAMPpulse repressed gene. Because its transcription ismaximal early in the developmental program, then isreduced as cAMP pulses are established, it is likelythat G�3 function is limited to the early stages ofaggregation. In the pkac– mutant, however, exo-genous cAMP pulses fail to repress G�3 mRNAexpression. This demonstrates that the signal fromcAMP pulses to the gene expression machinery istransmitted through PKA, and explains why artificialcAMP pulses do not restore expression of cAR1 orACA in the pkac– mutant.
Temporal relationships between changes in G�3and PKAC gene expression and PKA activity areshown in Figure 6. As amoeba begin to starve, G�3and PKAC expression levels rapidly rise, increasingthe expression of several signaling genes: ACA,cAR1, CRAC and cAR3. The expression of thesignaling genes establishes endogenous cAMPpulses, which reduces G�3 expression, but not PKACexpression. PKA activity is low as long as G�3 expres-sion is high, immediately after starvation. As cAMPpulses are established and G�3 levels recede, PKAactivity increases. High G�3 expression is necessaryto induce expression of the signaling genes, but mustbe reduced to allow increased PKA activity for aggre-gation. A model that depicts the potential G�3/PKA
signaling network that controls developmental geneexpression early in development is shown in Figure 7.
Our data provide further support for signalingnetworks controlling the transition from growth todifferentiation. Continued scrutiny of the mechanismof this transition should lead to identification of theG�3 signaling pathway components and their pointsof interaction with the PKA signaling pathway.
Acknowledgements
We thank Laurens Smith for critical reading of themanuscript. This work was supported in part by aNational Institutes of Health (NIH) NCRR grant(RR09765) to M. A. B.
References
Brandon, M. A. & Podgorski, G. J. 1997. G�3 regulates thecAMP signaling system in Dictyostelium. Mol. Biol. Cell 8,1677–1685.
Brandon, M. A., Voglmaier, S. & Siddiqi, A. A. 1997. Molecularcharacterization of a Dictyostelium G-protein �-subunitrequired for development. Gene 200, 99–105.
Caterina, M. J., Milne, J. L. S. & Devreotes, P. N. 1994. Mutationof the third intracellular loop of the cAMP receptor, cAR1, ofDictyostelium yields mutants impaired in multiple signalingpathways. J. Biol. Chem. 269, 1523–1532.
Chomczynski, P. & Sacchi, N. 1987. Single-step method of RNAisolation by acid guanidinuim thiocyanate-phenol-chloro-form extraction. Ann. Biochem. 162, 156–159.
Devreotes, P. N. 1994. G protein-linked signaling pathwayscontrol the developmental program of Dictyostelium.Neuron 12, 235–241.
Graness, A., Hanke, S., Boehmer, F. D., Presek, P. & Liebmann,C. 2000. Protein-tyrosine-phosphatase-mediated epidermalgrowth factor (EGF) receptor transinactivation and EGFreceptor-independent stimulation of mitogen-activatedprotein kinase by bradykinin in A431 cells. Biochem. J.347, 441–447.
Grundker, C., Volker, P. & Emons, G. 2001. Antiproliferativesignaling of luteinizing hormone-releasing hormone inhuman endometrial and ovarian cancer cells through Gprotein �1-mediated activation of phosphotyrosine phos-phatase. Endocrinology 142, 2369–2380.
Hadwiger, J. A. & Firtel, R. A. 1992. Analysis of G�4, a G-proteinsubunit required for multicellular development in Dictyo-stelium. Genes Dev. 6, 38–49.
Hamm, H. E. 1998. The many faces of G protein signaling.J. Biol. Chem. 273, 669–672.
Hammerschmidt, M., Bitdoog, M. J. & McMahon, A. P. 1996.Protein kinase A is a common negative regulator ofHedgehog signaling in the vertebrate embryo. Genes Dev.10, 647–658.
Insall, R., Kuspa, A., Lilly, P. J. et al. 1994. CRAC, a cytosolicprotein containing a pleckstrin homlolgy domain, is requiredfor receptor and G protein-mediated activation of adenylylcyclase in Dictyostelium. J. Cell Biol. 126, 1537–1545.
Johnson, R. L., Saxe, C. L. III, Gollop, R., Kimmel, A. R. &Devreotes, P. N. 1993. Identification and targeted genedisruption of cAR3, a cAMP receptor subtype expressed

Interacting pathways for gene expression 465
during multicellular stages of Dictyostelium development.Genes Dev. 7, 273–282.
Kim, J. Y., van Haastert, P. & Devreotes, P. N. 1996. Socialsenses: G-protein-coupled receptor signaling pathwaysin Dictyostelium discoideum. Chem. Biol. 3, 239–243.
Klein, P. S., Sun, T. J., Saxe, C. L. III, Kimmel, A. R., Johnson,R. L. & Devreotes, P. N. 1988. A chemoattractant receptorcontrols development in Dictyostelium discoideum. Science241, 1467–1472.
Kuschel, M., Zhou, Y.-Y., Cheng, H. et al. 1999. Gi protein-mediated functional compartmentalization of cardiac�2-adrenergic signaling. J. Biol. Chem. 274, 22 048–22 052.
Lilly, P., Wu, L. J., Welker, D. L. & Devreotes, P. N. 1993. AG-protein �-subunit is essential for Dictyostelium develop-ment. Genes Dev. 7, 986–995.
Mann, S. K. O., Brown, J. M., Briscoe, C. et al. 1997. Role ofcAMP-dependent protein kinase in controlling aggregationand postaggregative development in Dictyostelium. Dev.Biol. 183, 208–221.
Mann, S. K. O. & Firtel, R. A. 1991. A developmentally regulated,putative serine/threonine protein kinase is essential fordevelopment in Dictyostelium. Mech. Dev. 35, 89–101.
Marinissen, M. J. & Gutkind, J. S. 2001. G-protein-coupledreceptors and signaling networks: emerging paradigms.Trends Pharm. Sci. 22, 368–375.
Pan, D. & Rubin, G. M. 1995. cAMP-dependent protein kinaseand hedgehog act antagonistically in regulating decapen-taplegic transcription in Drosophila imaginal discs. Cell 80,543–552.
Parent, C. A. & Devreotes, P. N. 1996. Molecular genetics ofsignal transduction in Dictyostelium. Ann. Rev. Biochem.65, 411–440.
Perrimon, N. 1996. Serpentine proteins slither in to the Winglessand Hedgehog fields. Cell 86, 513–516.
Pitt, G. S., Milona, N., Borleis, J., Lin, K. C., Reed, R. R. &Devreotes, P. N. 1992. Structurally distinct and stage-specific adenylyl cyclase genes play different roles inDictyostelium development. Cell 69, 305–315.
Podgorski, G. & Deering, R. A. 1984. Thymidine-requiringmutants of Dictyostelium discoideum. Mol. Cell. Biol. 4,2784–2791.
Pupillo, M., Kumagai, A., Pitt, G. S., Firtel, R. A. & Devreotes,P. N. 1989. Multiple �-subunits of guanine nucleotide-binding proteins in Dictyostelium. Proc. Natl Acad. Sci. USA86, 4892–4896.
Schulkes, C. & Schaap, P. 1995. cAMP-dependent proteinkinase activity is essential for preaggregative geneexpression in Dictyostelium. FEBS Lett. 368, 381–384.
Simon, M. N., Pelegrini, O., Veron, M. & Kay, R. R. 1992. Mutationof protein kinase-A causes heterochronic development ofDictyostelium. Nature 356, 171–172.
Söderbom, F., Anjard, C., Iranfar, N., Fuller, D. & Loomis, W. F.1999. An adenylyl cyclase that functions during latedevelopment of Dictyostelium. Development 126,5463–5471.
Souza, G. M., de Silva, S. M. & Kuspa, A. 1999. Starvationpromotes Dictyostelium development by relieving PufAinhibition of PKA translation through the YakA kinasepathway. Development 126, 3263–3274.
Souza, G. M., Lu, S. J. & Kuspa, A. 1998. YakA, a protein kinaserequired for the transition from growth to development inDictyostelium. Development 125, 2291–2302.
Wu, L., Hansen, D., Franke, J., Kessin, R. H. & Podgorski, G. J.1995. Regulation of Dictyostelium early development genesin signal transduction mutants. Dev. Biol. 171, 149–158.
Zhang, N., Long, Y. & Devreotes, P. N. 2001. G� in Dictyo-stelium: Its role in localization of G�� to the membrane isrequired for chemotaxis in shallow gradients. Mol. Biol. Cell12, 3204–3213.