function of isoamylase-type starch debranching enzymes isa1 and isa2 in the zea mays leaf
TRANSCRIPT

Function of isoamylase-type starch debranching enzymes ISA1and ISA2 in the Zea mays leaf
Qiaohui Lin1, Maud Facon2, Jean-Luc Putaux3, Jason R. Dinges1, Fabrice Wattebled2, Christophe D’Hulst2,
Tracie A. Hennen-Bierwagen1 and Alan M. Myers1
1Roy J. Carver Department of Biochemistry, Biophysics, and Molecular Biology, Iowa State University, Ames, IA 50011, USA; 2Unit�e de Glycobiologie Structurale et Fonctionelle, UMR8576
CNRS/Universit�e Lille 1, F-59655, Villeneuve d’Ascq, France; 3Centre de Recherches sur les Macromol�ecules V�eg�etales (CERMAV-CNRS), BP 53, F-38041 Grenoble, Cedex 9, France
Author for correspondence:Tracie A. Hennen-Bierwagen
Tel: +1 515 294 8202Email: [email protected]
Received: 5 June 2013
Accepted: 8 July 2013
New Phytologist (2013)doi: 10.1111/nph.12446
Key words: isoamylase-type starchdebranching enzyme (ISA), leaf starchbiosynthesis, starch metabolism, starchstructure, Zea mays.
Summary
� Conserved isoamylase-type starch debranching enzymes (ISAs), including the catalytic ISA1
and noncatalytic ISA2, are major starch biosynthesis determinants. Arabidopsis thaliana
leaves require ISA1 and ISA2 for physiological function, whereas endosperm starch is near
normal with only ISA1. ISA functions were characterized in maize (Zea mays) leaves to deter-
mine whether species-specific distinctions in ISA1 primary structure, or metabolic differences
in tissues, are responsible for the differing ISA2 requirement.� Genetic methods provided lines lacking ISA1 or ISA2. Biochemical analyses characterized
ISA activities in mutant tissues. Starch content, granule morphology, and amylopectin fine
structure were determined.� Three ISA activity forms were observed in leaves, two ISA1/ISA2 heteromultimers and one
ISA1 homomultimer. ISA1 homomultimer activity existed in mutants lacking ISA2. Mutants
without ISA2 differed in leaf starch content, granule morphology, and amylopectin structure
compared with nonmutants or lines lacking both ISA1 and ISA2. The data imply that both the
ISA1 homomultimer and ISA1/ISA2 heteromultimer function in the maize leaf.� The ISA1 homomultimer is present and functions in the maize leaf. Evolutionary divergence
between monocots and dicots probably explains the ability of ISA1 to function as a homomul-
timer in maize leaves, in contrast to other species where the ISA1/ISA2 heteromultimer is the
only active form.
Introduction
Isoamylase-type starch debranching enzymes (ISAs) provide func-tions in starch biosynthesis that are subject to evolutionary conser-vation, as indicated by their high degree of primary sequenceidentity in chloroplast-containing organisms (i.e. the Chloroplast-ida; for recent reviews, see Hennen-Bierwagen et al., 2012; Streb& Zeeman, 2012). ISAs catalyze hydrolysis of a(1?6) glycosidebonds in branched glucose homopolymers, and are thought to beinvolved in determining a clustered arrangement of branched link-ages in precursor glucans. This structure is proposed to favor crys-tallization of unbranched regions of a(1?4)-linked linear chainsand subsequent assembly into higher order structures culminatingin insoluble starch granules. In support of this hypothesis, muta-tions in genes encoding certain ISA proteins typically causedecreased starch content and accumulation of water-soluble poly-saccharides (WSPs) that are not present in nonmutant plants.Such changes have been observed in the endosperm of severalmonocots, in dicot leaves and tubers, and in the unicellular greenalga Chlamydomonas reinhardtii. In all instances a gene encodingan ISA protein was mutated, and this distinctive phenotype hasnot been observed in the absence of altered ISA function.
Three separate genes encoding ISA proteins are conserved inthe genomes of all Chloroplastida species characterized to date(Deschamps et al., 2008). ISA3 functions primarily in starchcatabolism (Wattebled et al., 2005, 2008; Delatte et al., 2006),whereas ISA1 and ISA2 are involved in starch biosynthesis. ISA1provides the catalytic activity for glucan debranching, and in allspecies tested is required for normal contents of crystalline starchand to repress accumulation of the plant WSP termed phytogly-cogen, which is similar in structure to animal glycogen. ISA2 is anoncatalytic protein as a result of conserved substitutions inamino acid residues required for activity in members of thewell-characterized glycosyl hydrolase family 13 (Henrissat, 1991;Hussain et al., 2003; Hennen-Bierwagen et al., 2012). In dicottissues, specifically the Arabidopsis thaliana leaf and potato (Sola-num tuberosum) tuber, ISA1 and ISA2 are involved at the samestep in starch metabolism and both proteins are needed toexecute that function (Zeeman et al., 1998; Bustos et al., 2004;Delatte et al., 2005; Wattebled et al., 2005). In the rice (Oryzasativa) or maize (Zea mays) endosperm, by contrast, loss of ISA2is tolerated without a significant reduction in starch content orthe appearance of phytoglycogen (Kubo et al., 2010; Utsumiet al., 2011). Chlamydomonas reinhardtii is intermediate in this
No claim to original US goverment works
New Phytologist� 2013 New Phytologist TrustNew Phytologist (2013) 1
www.newphytologist.com
Research

regard, with loss of either ISA1 or ISA2 causing a decrease instarch content and the accumulation of phytoglycogen, but withless severe effects when ISA2 is affected (Mouille et al., 1996;Dauvillee et al., 2001). These observations raise the question ofwhy noncatalytic ISA2 would be strictly conserved in all chloro-plast-containing species even though in some plant tissues thebiosynthetic ISA function(s) can be provided by ISA1 alone.
To address this question, the roles of ISA1 and ISA2 in starchmetabolism in maize leaves were characterized in this study. ISAfunction in the leaf has been characterized genetically only inA. thaliana, where essentially identical metabolic and biochemicalphenotypes result from loss of ISA1, ISA2, or both together(Zeeman et al., 1998; Delatte et al., 2005; Wattebled et al.,2005). Leaf starch content is reduced in either single or doublemutants by c. 80%, the structure of the amylopectin componentof starch is characteristically altered, and phytoglycogen accumu-lates to a level equivalent to c. 40–50% of the starch present inwild type. A single ISA activity is detected by in-gel enzyme assays(i.e. zymograms) of leaf soluble extracts, and this is absent in nullmutants lacking ISA1 or ISA2 and in double mutants. From thesedata, ISA1 and ISA2 appear to function together in theA. thaliana leaf in a heteromultimeric enzyme complex, and bothproteins are necessary for activity and physiological function.
Loss of ISA1 in the cereal endosperm causes a phenotype simi-lar to that seen in the A. thaliana leaf, specifically a starch contentreduced by 80% or more, accumulation of high concentrationsof phytoglycogen, and altered amylopectin structure (James et al.,1995; Nakamura et al., 1996; Dinges et al., 2001; Burton et al.,2002). Genetic alterations that cause loss of ISA2 are known inrice and maize (Kubo et al., 2010; Utsumi et al., 2011). In con-trast to the A. thaliana leaf, these did not cause a substantialreduction in endosperm starch or accumulation of phytoglyco-gen. Endosperm tissues from rice and maize were shown by bio-chemical purification to possess a heteromultimeric enzymecomprising both ISA1 and ISA2 (Utsumi & Nakamura, 2006;Kubo et al., 2010), consistent with the genetic data fromA. thaliana. These tissues also possess a homomultimeric enzymethat contains only ISA1, which was not observed in theA. thaliana leaf.
Two potential explanations can be suggested for the differencesbetween ISA functions, that is, whether or not ISA2 is requiredfor normal starch metabolism, in dicot tissues and the monocotendosperm. One possibile explanation is distinctions in carbohy-drate metabolism between tissues. In particular, accumulation oftransitory starch in the leaf might require the ISA1/ISA2 hetero-multimeric enzyme, whereas ISA1 alone might be sufficient forstorage starch in the endosperm, which is not degraded during thediurnal cycle. Thus, mutations that affect ISA2 would affect leafstarch metabolism but not affect the endosperm. Alternatively,evolutionary divergence between dicots and some or all cereal spe-cies may have resulted in different ISA1 primary structures. TheISA1 sequence in cereals may possess the intrinsic ability to func-tion as a homomultimer, whereas the protein from dicots may befunctional only in heteromultimeric complexes that contain ISA2.According to this hypothesis, mutation of ISA2 should havespecies-specific rather than tissue-specific effects.
Characterization of the effects of mutations eliminating ISA1or ISA2 on starch metabolism in leaves of a monocot species willhelp to distinguish these possibilities. The results presented hereshow that ISA functions differently in maize leaves than inA. thaliana leaves. In contrast to A. thaliana leaves, the effects ofeliminating either ISA1 or ISA2 are distinct from each other inmaize. The data are consistent with the hypothesis that evolution-ary divergence has resulted in an ISA1 protein in monocots thatis enzymatically competent in the absence of ISA2. This enzymedoes not entirely supply the physiological role of ISA in leafstarch biosynthesis, however, because ISA2 mutations had dis-cernible effects on starch content and granule size.
Materials and Methods
Plant material
Maize (Zea mays L.) lines used in this study were essentially con-genic in theW64A inbred genetic background as a result of a min-imum of five backcrosses to the nonmutant standard. Molecularcharacterization was described previously for mutations of thegene sugary1 (su1) that encodes ISA1, specifically su1-4582 andsu1-Ref (James et al., 1995; Dinges et al., 2001), or the isa2-339allele of the isa2 locus that encodes ISA2 (Kubo et al., 2010).Seedlings were grown in the glasshouse until emergence of thefourth leaf. Late in the light phase of the diurnal cycle, the outerhalf of the third leaves was harvested and used fresh for enzymeactivity gels or immediately frozen in liquid nitrogen and stored at�80°C until use for carbohydrate analyses. Seedlings for analysisof gene expression in leaves were grown in a growth chamberunder a diurnal cycle with alternating periods of 8-h darkness at25°C and 16-h light at 28°C with c. 280 lmol quanta m�2 s�1 ofmixed fluorescent and incandescent light. Tissues were harvestedat various times throughout the diurnal cycle, immediately frozenin liquid nitrogen, and stored at�80°C until use.
Carbohydrate analyses
Leaf starch was isolated and quantified as previously described(Yandeau-Nelson et al., 2011) after lysis of c. 200 mg of tissue in0.7% ice-cold perchloric acid, with biological replicate analysesperformed on individual plants of the same genotype. WSP wascollected from the soluble phase of the same extracts, and quanti-fied after neutralization to approximately pH 7 by dropwise addi-tion of 2M KOH, 0.4M 2-(N-morpholino) ethanesulfonic acidand 0.4M KCl. Both starch and WSP were quantified after con-version to free glucose by treatment with thermostable a-amylaseand amyloglucosidase using commercial reagents (K-TSTA;Megazyme, Bray, Co. Wicklow, Ireland). The amylopectin linearchain length distribution was determined by fluorophore-assistedcarbohydrate electrophoresis (FACE) using a P/ACE capillaryelectrophoresis instrument (Beckman Coulter Inc., Indianapolis,IN, USA) to separate glucans (O’Shea et al., 1998; Lin et al.,2011). Glucans for amylopectin chain length distribution analy-sis were extracted from the insoluble leaf fraction by boiling inDMSO before FACE analysis. The frequency of chains of each
New Phytologist (2013) No claim to original US goverment works
New Phytologist� 2013 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist2

degree of polymerization (DP) was calculated as the percentageof the total chains within a given DP range. Glucans from a mini-mum of three biological replicates were analyzed, and the fre-quency values for each chain length were averaged amongindividuals of the same genotype. Difference plots were calcu-lated by subtracting the wild-type average value from the mutantaverage value of the same DP.
qPCR
The relative abundance of ISA1 and ISA2 mRNAs standardizedto the 18S rRNA abundance was measured using quantitativereal-time PCR (qPCR) amplification of leaf cDNA. Methods formRNA isolation, cDNA preparation, and qPCR analysis of thesetwo transcripts were described previously (Kubo et al., 2010).
Immunoblot and zymogram analyses of total solubleextracts
Fresh leaf tissue (c. 1 g) was ground to a powder under liquidnitrogen in a mortar and pestle. The powder was ground furtherin 4 ml of ice-cold extraction buffer (50 mM Tris-acetate, pH 7.5,10 mM DTT, and 1X protease inhibitor cocktail (SigmaP-2714)), and then the lysate was transferred to a 15-ml polypro-pylene tube and centrifuged at 10 000 rpm for 10 min at 4°C.The supernatant was filtered through a 0.45-lm syringe filter(431220; Corning Inc., Tewksbury, MA, USA) and then concen-trated by centrifugal filtration to c. 0.3 ml (UFC801024; EMDMillipore, Darmstadt, Germany). The protein concentration inthe total soluble lysate was determined by the Bradford method(500-0006; Bio-Rad, Hercules, CA, USA). For immunoblot anal-ysis, 180 lg of extract was fractionated by SDS-PAGE andprobed with affinity-purified immunoglobulin G (IgG) fractionsspecific for maize ISA1 or ISA2, as previously described (Kuboet al., 2010). For zymogram analysis of total leaf extracts, 100–400 lg of soluble extract was fractionated by native-PAGE usingprecast 7.5% acrylamide gels (Bio-Rad 345-0010). Conditionsfor electrophoresis and processing for detection of in-gel ISAactivity were described previously (Kubo et al., 2010). Endospermextracts used as controls in the zymograms were prepared asdescribed previously from kernels harvested 20 d after pollination(DAP; Kubo et al., 2010).
Fractionation of ISA activities
Leaf extracts were prepared as described in the previous section,except that 5 g of leaf tissue was extracted in 12 ml of buffer. Thelysate was clarified by centrifugation at 17 000 g for 10 min at4°C, and the supernatant was passed twice through 0.45-lmsyringe filters. Eight milliliters of lysate was passed through a 1-mlbed volume HiTrap Q HP anion exchange column (17-1153-01;GE Healthcare Biosciences). Proteins were eluted in a lineargradient of 0–1M NaCl in 50 mM Tris-acetate, pH 7.5, with10 mM DTT added immediately before use, and 1-ml fractionswere collected. ISA1 and ISA2 were identified by immunoblotanalyses, and both proteins were found to elute in the same three
fractions (data not shown). These fractions were pooled and con-centrated to c. 1 ml by centrifugal filtration and DTT was addedto a final concentration of 20 mM. The pooled AEC fractions(500 ll) were applied to a Superdex 200 10/300 GL gel perme-ation column (17-5175-01; GE Healthcare Life Sciences,Pittsburgh, PA, USA) and eluted in 50 mM Tris-acetate, pH 7.5,150 mMNaCl and 10 mMDTT, while collecting 1-ml fractions.Samples of each fraction were tested by SDS-PAGE and immun-blot analysis for the presence of ISA1 and ISA2. Fractionscontaining those proteins (25 ll) were applied to native-PAGEgels and subjected to zymogram analysis. Duplicate native-PAGEgels were analyzed for ISA1 and ISA2 proteins by immunoblotanalysis. Endosperm extracts were analyzed similarly, beginningwith 5 g of tissue from kernels harvested 20 DAP.
Transmission electron microscopy and scanning electronmicroscopy
The outermost regions of maize leaves from glasshouse-grownseedlings were harvested at the end of the light period of the diur-nal cycle. Tissue was cut with a fresh razor blade into smallpieces, then immersed in fixative 0.1 M cacodylate, pH 7.2, 2%paraformaldehyde and 2% glutaraldehyde. Vacuum was appliedfor 30 min. Leaf samples were embedded in Spurr’s resin, thenpostfixed with 1% osmium tetroxide, sectioned to a thickness of80 nm, affixed to grids and stained with uranyl acetate and leadcitrate. Transmission electron microscopy (TEM) observationwas performed with a Jeol 2100 microscope operating at 200 kV.
Starch granules to be analyzed by scanning electron micros-copy (SEM) were purified from insoluble leaf material as previ-ously described (Yandeau-Nelson et al., 2011). The pellet fromthe leaf crude lysate was resuspended in sterile deionized waterand filtered through two layers of Miracloth (Calbiochem,Darmstadt, Germany). The starch pellet was further purified byisopicnic centrifugation in Percoll (GE Healthcare) at 10 000 gfor 1 h at 4°C. The pellet was rinsed twice with sterile deionizedwater and stored in 20% ethanol at 4°C until use. Droplets ofstarch granule suspensions were allowed to dry on freshly cleavedmica. After coating with gold/palladium, they were observed insecondary electron mode using a Jeol JSM6300 microscope (Jeol,Peabody, MA, USA) operating at 8 kV. Size-distribution histo-grams were determined by measuring the apparent diameter of atleast 300 particles per sample from the SEM images.
Results
Transcript and protein abundance
The relative steady-state abundances of ISA1 and ISA2 mRNAsin seedling leaves over the course of a 16-h light : 8-h dark diurnalcycle were measured by qPCR. The abundances of these tran-scripts were reported previously for developing endosperm tissue,where ISA1 was c. 2.5-fold more abundant than ISA2 (Kuboet al., 2010). In seedlings leaves, the relative abundances of ISA1and ISA2 mRNAs were approximately equal at the end of thenight. ISA2 mRNA abundance was elevated within the first 3 h
No claim to original US goverment works
New Phytologist� 2013 New Phytologist TrustNew Phytologist (2013)
www.newphytologist.com
NewPhytologist Research 3

of the light phase to approximately five-fold higher than ISA1mRNA, and remained more abundant by about the same factorthroughout the day (Fig. 1a). The qPCR results are in agreementwith publicly available whole-transcriptome sequencing data(RNAseq) showing ISA2 to be two- to three-fold more abundant
than ISA1 in leaf tissue harvested during the day (Fig. 1b). Thus,the relative abundances of ISA1 and ISA2 transcripts are reversedin the maize endosperm and leaf, with ISA1 more abundant inthe nonphotosynthetic tissue and ISA2 present at higher abun-dances in leaves throughout the light phase of the diurnal cycle.
ISA1 and ISA2 proteins were both detected by immunoblotanalysis in soluble extracts of seedling leaves harvested during theday (Fig. 1c). The specificity of the IgG fractions used for thisanalysis was demonstrated using leaf extracts from plants homo-zygous for the null mutations su1-4582, in the gene sugary1 (su1)that codes for ISA1 (James et al., 1995), or isa2-339, in the genethat codes for ISA2 (Kubo et al., 2010). Leaves homozygous forsu1-4582 and thus lacking ISA1 also were completely deficientfor ISA2 as a secondary effect (Fig. 1c), as has been observed pre-viously in the endosperm (Kubo et al., 2010). This result is con-sistent with the conclusion that ISA1 is required for the stabilityof ISA2. By contrast, ISA1 continued to accumulate in isa2-339mutant leaves lacking ISA2, although at a reduced concentrationcompared with nonmutant tissue (Fig. 1c). An analogous second-ary effect of an isa2- null mutation was seen in A. thaliana leaves(Delatte et al., 2005); however, in the dicot only a trace amountof ISA1 remained, whereas in maize substantial amounts of ISA1accumulated in the absence of ISA2.
Leaf ISA activity
The ISA enzymes in maize leaves were first characterized byzymogram analyses of total soluble extracts. Proteins were sepa-rated by native-PAGE, and then electrophoretically transferred toa gel impregnated with 0.3% solubilized potato starch. The starchgel was stained with iodine-potassium iodide solution (I2/KI) sothat starch-modifying enzymes could be detected by changes incolor. In such analyses, ISAs are known to generate light-bluebands, whereas starch branching enzymes generate red or pinkbands and amylases produce white bands (Colleoni et al., 2003).Endosperm tissue subjected to such analyses revealed three formsof ISA activity, including a faster-migrating form of the ISA1homomultimer (form I) and two slower-migrating forms of theISA1/ISA2 heteromultimeric enzyme (forms II and III; Kuboet al., 2010). Leaf extracts generated two bands of ISA activity,which migrated slightly faster in native-PAGE than the twoheteromultimeric forms from the endosperm (Fig. 2). Both ofthese activities were missing in isa2-339 or su1-4582 leaves, con-sistent with their identification as the ISA1/ISA2 heteromultimer-ic enzyme. A third activity band corresponding to form I was notobvious in leaf extracts, although a diffuse, faster-migrating light-staining area was present near the position of the endosperm ISA1homomultimer. This diffuse activity was missing in extracts defi-cient for ISA1 but was not affected by loss of ISA2 (Fig. 2).
To better resolve ISA activities, enzymes from leaf and endo-sperm crude extracts were partially purified by anion-exchangechromatography (AEC) on HiTrapQ HP columns, concentrated,and then separated by gel permeation chromatography (GPC) ona Superdex 10/300 GL column. Fractions were then analyzed byzymogram and also by immunoblot of duplicate lanes to deter-mine whether the ISA1 and/or ISA2 protein co-eluted in GPC
(a)
(b)
(c)
Fig. 1 mRNA and protein abundances in Zea mays leaves. (a) QuantitativePCR (qPCR). Relative transcript abundances were quantified in wild-typeleaves harvested during a 16-h light : 8-h dark diurnal cycle. Results shownare the mean� SE of three biological replicates. When absent, the errorbars are smaller than the symbols. (b) Whole-transcriptome sequencingdata (RNAseq). Data are available at www.maizegdb.org, from Li et al.(2010) (9-d seedlings) and Davidson et al. (2011) (5-wk plants). Immature9-d-old seedling tissue was from the leaf base, where C4 metabolism hasnot yet initiated. Error bars indicate the 95% confidence interval for theexpression of a given gene as determined in the original publications. (c)Immunoblot detection of isoamylase-type starch debranching enzyme 1(ISA1) and ISA2 in seedling leaves. The same amount of protein in totalsoluble extracts from wild-type or mutant leaves (180 lg) was separatedby SDS-PAGE and probed with the indicated immunoglobulin G (IgG)fractions.
New Phytologist (2013) No claim to original US goverment works
New Phytologist� 2013 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist4

and co-migrated in native-PAGE with specific ISA activities.Endosperm extracts provided standards for the presence of theISA1 homomultimer and ISA1/ISA2 heteromultimers in specificGPC fractions (Fig. 3a). As seen previously (Kubo et al., 2010),both ISA1 and ISA2 co-migrated with enzyme forms II and III,whereas ISA1 co-migrated with form I but ISA2 clearly was notpresent in that activity band (Fig. 3a).
GPC fractions from the leaf also contained three forms of ISA.These activities eluted in the same fractions as ISA forms I, II andIII from the endosperm, and co-migrated with those enzymes innative-PAGE (Fig. 3b). ISA form I in the pooled AEC fractionsfrom nonmutant leaves co-migrated in native-PAGE with ISA1but did not contain any ISA2 (Fig. 3b). SDS-PAGE immuno-blots revealed that the ISA2 protein in leaf extracts co-eluted inGPC fractions B6 to B4 with activity forms II and III, but wasnot detected in fraction B3 which was enriched for enzyme formI (Fig. 3b). These data indicate the presence of the ISA1 homo-multimeric enzyme in the maize leaf. ISA form II in the pooledAEC fractions from the leaf co-migrated with both ISA1 andISA2, indicating the presence of an ISA1/ISA2 heteromultimerin that tissue. ISA form III in the leaf AEC pool co-migrated withISA2. The fact that ISA1 was not detected in leaf form III
probably is attributable to the low sensitivity of the native-PAGEimmunoblots and the relatively low abundance of that form ofthe enzyme. ISA1 is expected to be present in that band becauseform III is absent from whole-cell extracts of either the endo-sperm or the leaf from su1-4582 mutants. Taken together, thedata indicate that both the ISA1 homomeric enzyme andthe ISA1/ISA2 heteromultimer exist in nonmutant maize leaves.The leaf differs from the endosperm, however, in the relative pro-portions of the different ISA activities, as far as can be judgedfrom these semiquantitative assays. The ISA1 homomultimerappears as the most prevalent activity in endosperm extracts,whereas there is proportionally less of this form of the enzymein leaf extracts compared with the heteromultimeric form II(Fig. 3a,b).
ISA activity was also partially purified from leaves of an isa2-339 mutant. In this instance, a single ISA activity band was pres-ent, indicated by its blue color, that co-migrated in native-PAGEwith the homomeric enzyme from the endosperm, that is, form I(Fig. 3c). This is in contrast to the extracts from nonmutantleaves that exhibited three blue bands of ISA activity (Fig. 3b).Immunoblot analysis of duplicate fractions showed that ISA1co-eluted with the activity band in GPC and co-migrated withthat enzyme in native-PAGE. These data confirm that an ISAactivity exists in maize leaves that does not utilize ISA2, becausethis protein is entirely lacking in isa2-339 mutants (Fig. 1c).
Granule morphology in leaves lacking ISA1 or ISA2
The appearance of starch granules in seedling leaf chloroplastswas compared between wild type and mutant lines homozygousfor a defect in the gene encoding ISA1 or ISA2. Leaf sampleswere collected from glasshouse-grown seedlings at the end of thelight period of the diurnal cycle and immediately fixed for TEM.Genotypes analyzed were the wild-type inbred W64A, and con-genic mutants homozygous for isa2-339, su1-4582, or su1-Ref.The su1-Ref allele is a missense mutation that results in an enzy-matically inactive form of ISA1 present at a low level comparedwith wild type (Kubo et al., 2010), whereas isa2-339 andsu1-4582 are null alleles that result in complete losss of ISA2 orISA1, respectively. Bundle sheath chloroplasts were examinedbecause these are the site of starch granule accumulation in themature maize leaf as the result of C4 metabolism. As a control,palisade mesophyll cells from wild-type and homozygous isa1-1A. thaliana leaves were analyzed similarly. The A. thalianamutantplastids exhibited loss or a reduced number of starch granules,and a distended stromal volume apparently resulting from phyto-glycogen accumulation, as previously reported (Fig. 4; Delatteet al., 2005).
Plastids from maize leaves lacking ISA2 exhibited a nearly nor-mal appearance with the exception that starch granules appearedto be slightly smaller than in wild type (Fig. 4). This is in contrastto isa2-1 mutants of A. thaliana, in which mesophyll or bundlesheath chloroplasts typically lack recognizable starch granules andcontain amorphous glucan particles with a range of structures(Zeeman et al., 1998; Delatte et al., 2005). Such abnormal glucanparticles were never observed in maize isa2-339 mutants. Maize
Fig. 2 Isoamylase-type starch debranching enzyme (ISA) activities in Zeamays leaf extracts. Proteins in leaf or endosperm extracts of the indicatedlines were analyzed by zymogram. Roman numerals indicate homomulti-meric (form I) and heteromultimeric (forms II and III) ISA activitiespreviously characterized in the maize endosperm (Kubo et al., 2010).White arrows and lines indicate the positions of potential ISA activitiesfrom the leaf. Unmarked starch-modifying activities are starch branchingenzymes or amylases. Protein loads were 50 lg for endosperm extractsand 400 lg for leaf extracts.
No claim to original US goverment works
New Phytologist� 2013 New Phytologist TrustNew Phytologist (2013)
www.newphytologist.com
NewPhytologist Research 5

bundle sheath chloroplasts from either su1-4582 or su1-Ref mutants contained starch granules that were noticeablysmaller and more irregularly shaped than in wild type, althoughdistended organelles and apparent phytoglycogen accumulationwere not obvious (Fig. 4). The granule morphology within bun-dle sheath plastids was distinctly different between leaves mutatedfor either ISA1 or ISA2.
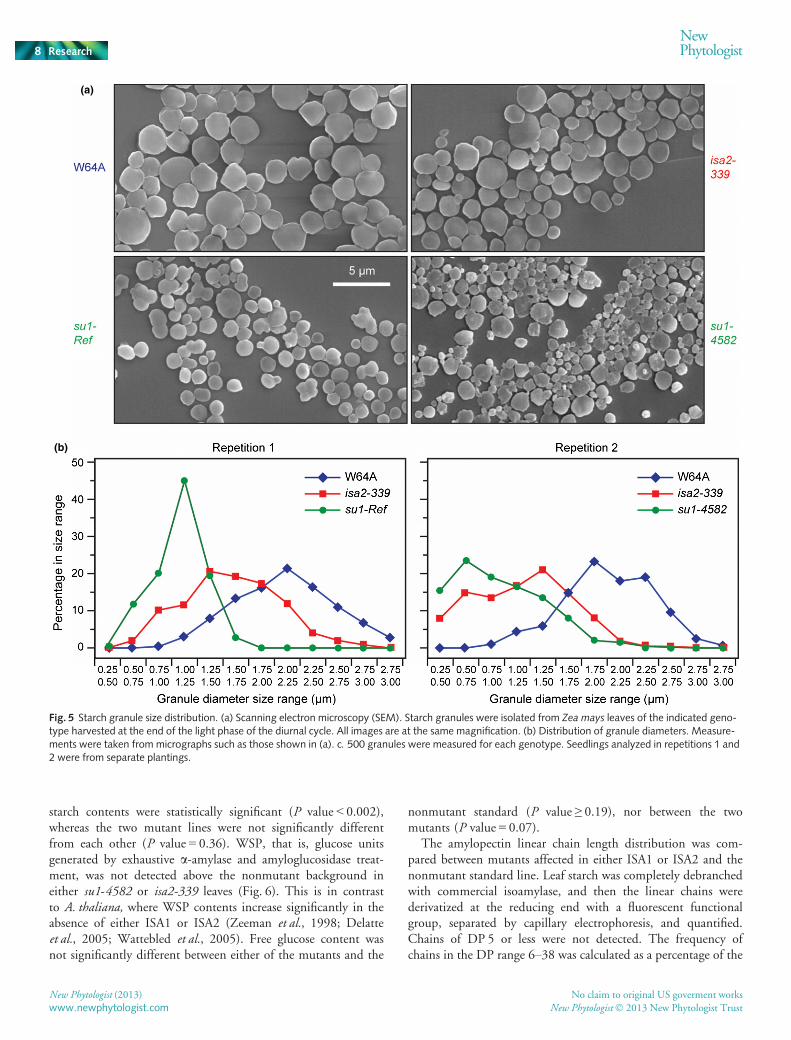
The size ranges of seedling leaf starch granules were quantifiedby SEM. Granules were extracted from W64A, isa2-339, or su1-
Ref seedlings at the end of the light phase and imaged by SEM(Fig. 5a), and then individual particle diameters were measured.The distributions of granule diameters differed among all geno-types (Fig. 5b), and the average diameters differed between anytwo lines with a high degree of statistical significance(P value < 0.0001). The results demonstrated that starch granulesfrom su1-Ref mutant leaves were smaller than those from nonmu-tants, and that granules generated in the absence of ISA2 wereintermediate in diameter between the particles from nonmutant
(a) (b)
(c)
Fig. 3 Isoamylase-type starch debranching enzyme (ISA) activities in gel permeation chromatography (GPC) fractions. (a) Nonmutant Zea mays
endosperm. ISA activities were partially purified by anion exchange chromatography (AEC), concentrated, and separated by GPC. Fractions were analyzedby native-PAGE zymograms as in Fig. 2. Duplicate native-PAGE gels, run simultaneously in a dual apparatus, were analyzed by immunoblot using theindicated immunoglobulin G (IgG) fractions so that co-migration of ISA1 and/or ISA2 with specific ISA mobility forms could be determined. Vertical arrowsindicate the peak of the elution volumes of protein standards with known molecular mass. ‘AEC’ indicates the pooled AEC fractions that were loaded ontothe GPC column and ‘ECE’ indicates endosperm crude extract. (b) Nonmutant Zea mays leaf extracts analyzed as in (a). Samples of the GPC fractions werealso analyzed by SDS-PAGE immunoblot. (c) isa2-339 leaf extracts analyzed as in (a).
New Phytologist (2013) No claim to original US goverment works
New Phytologist� 2013 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist6

and ISA1-defective leaves. As a biological replication, the experi-ment was repeated with W64A, isa2-339, and su1-4582 seedlingsfrom a separate planting. The diameter distributions again dif-fered among gentoypes (Fig. 5b) and the average values were sig-nificantly different (P value < 0.0001). The replicate analyseswere consistent in the sense that starch granules lacking func-tional ISA1 were smaller than those present in either nonmutantleaves or mutants lacking only ISA2. These data confirm theresults of the TEM analysis of granule morphology within chlo-roplasts, and indicate that in maize, unlike A. thaliana, the effectsof ISA2 mutation on starch granule morphology are differentfrom those resulting from loss of ISA1 function.
Storage glucan content and structure in leaves lacking ISA1or ISA2
Starch, WSP, and free glucose were quantified in glasshouse-grown seedling leaves of wild-type, su1-4582, and isa2-339maize plants harvested at the end of the light phase. Loss ofeither ISA1 or ISA2 caused a reduced starch content to c. 36–46% of the wild-type content (Fig. 6). Despite this defect therewas no obvious difference in plant growth as judged by theappearance of su1-4582 or isa2-339 mutants compared with thenonmutant standard over the course of the plant life cycle (datanot shown). Differences between the mutant and wild-type
Fig. 4 Plastids imaged by transmissionelectronmicroscopy (TEM). Zeamaysseedling leaf tissue was harvested and fixed atthe end of the light phase of the diurnal cycle.Two images are shown for each genotype.A. thaliana non-mutant and isa1-1mutantchloroplasts were included for comparison asa plant known to accumulate phytoglycogen.All images are at the samemagnification.
No claim to original US goverment works
New Phytologist� 2013 New Phytologist TrustNew Phytologist (2013)
www.newphytologist.com
NewPhytologist Research 7
starch contents were statistically significant (P value < 0.002),whereas the two mutant lines were not significantly differentfrom each other (P value = 0.36). WSP, that is, glucose unitsgenerated by exhaustive a-amylase and amyloglucosidase treat-ment, was not detected above the nonmutant background ineither su1-4582 or isa2-339 leaves (Fig. 6). This is in contrastto A. thaliana, where WSP contents increase significantly in theabsence of either ISA1 or ISA2 (Zeeman et al., 1998; Delatteet al., 2005; Wattebled et al., 2005). Free glucose content wasnot significantly different between either of the mutants and the
nonmutant standard (P value ≥ 0.19), nor between the twomutants (P value = 0.07).
The amylopectin linear chain length distribution was com-pared between mutants affected in either ISA1 or ISA2 and thenonmutant standard line. Leaf starch was completely debranchedwith commercial isoamylase, and then the linear chains werederivatized at the reducing end with a fluorescent functionalgroup, separated by capillary electrophoresis, and quantified.Chains of DP 5 or less were not detected. The frequency ofchains in the DP range 6–38 was calculated as a percentage of the
(a)
(b)
Fig. 5 Starch granule size distribution. (a) Scanning electron microscopy (SEM). Starch granules were isolated from Zea mays leaves of the indicated geno-type harvested at the end of the light phase of the diurnal cycle. All images are at the same magnification. (b) Distribution of granule diameters. Measure-ments were taken from micrographs such as those shown in (a). c. 500 granules were measured for each genotype. Seedlings analyzed in repetitions 1 and2 were from separate plantings.
New Phytologist (2013) No claim to original US goverment works
New Phytologist� 2013 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist8

total, and for each DP the value in nonmutant starch was sub-tracted from the mutant value to reveal structural differences(Fig. 7). Mutants affected in ISA1 as a result of carrying eitherthe su1-Ref or su1-4582 allele exhibited an increased frequency ofshort chains in the range of DP 6–12 compared with normal.The effect was stonger for the null allele than the point mutationfor the shortest chains of DP 6–8. This structural pattern is simi-lar to amylopectin from other plant sources carrying mutationsof ISA1, including the A. thaliana leaf (Delatte et al., 2005;Wattebled et al., 2005) and maize endosperm (Dinges et al.,2001). As previously mentioned, in the A. thaliana leaf thechanges observed in isa2- mutants were essentially identical tothose of isa1- mutants (Delatte et al., 2005; Wattebled et al.,2005). In maize leaf starch, the effects of isa2- mutations weresimilar but not identical to those of the isa1- mutants. Thechanges were quantitatively less severe and the range of chainlengths affected was narrower in isa2- leaf starch compared withthe isa1- chain length profiles (Fig. 7).
Fig. 6 Leaf glucan content. Glasshouse-grown Zea mays seedling leaveswere harvested at the end of the light phase of the diurnal cycle. Valuesindicated are averages + SE from four biological replicates of each mutantand three nonmutant replicates. Starch, black bars; water-solublepolysaccharides (WSPs), white bars; glucose, gray bars.
Fig. 7 Linear chain length distributions in Zea mays leaf starch amylopectin. For each genotype the frequency distribution� SE is shown as an average of atleast three biological replicates. Difference plots are shown in which the frequency value for each degree of polymerization (DP) in the nonmutant lineW64A was subtracted from that of the indicated mutant line.
No claim to original US goverment works
New Phytologist� 2013 New Phytologist TrustNew Phytologist (2013)
www.newphytologist.com
NewPhytologist Research 9

Distinctions in ISA1 primary structure conserved inmonocots and dicots
Multiple amino acid sequence alignments of ISA1 from fivemonocot species and six dicots were obtained using the programCLUSTALW (Thompson et al., 1994). In general, the alignmentbetween monocot and dicot ISA1s extends throughout the pro-teins with a high degree of sequence identity. Amino acids thatare identical in all the monocots but vary by the presence of non-conserved residues at those positions in the dicots were noted(Supporting Information Fig. S1). Two extended blocks of suchmonocot/dicot divergence were identified (Fig. 8). A span of resi-dues that differs entirely between monocots and dicots, beginningat residue 287 of full-length maize ISA1, is intercalated betweenregions of extremely high sequence conservation in both theN- and C-terminal directions (Fig. 8). All of the dicots have fiveadditional amino acids in this region compared with the mono-cots. The monocot sequences contain three successive Ser resi-dues, or in one instance a Cys and two Ser residues, none ofwhich are present in the dicots. The dicots possess a differentsequence of 10 residues, six of which are identical or essentiallyconserved in all six species examined.
Prediction of the maize ISA1 three-dimensional structurebased on comparison to the known structure of isoamylase fromPseudomonas amyloderamosa (Katsuya et al., 1998) suggests thatthis region of the protein is accessible on the surface (Fig. S2).
The second structural feature divergent between monocot anddicot ISA1s is located close to the N terminus (Fig. 8) of themature proteins. All five monocot proteins analyzed contain acontigous sequence close to their mature amino terminus of 8–10residues that are entirely Asp or Glu acidic groups or contain asingle neutral residue within the span of acidic amino acids. Noneof the dicot ISA1 proteins possess such a sequence. Furthermore,
adjacent to the acid-rich sequence is a span of 14 residues inwhich ten amino acids are identical in all the monocots and diver-gent in all the dicots. Thus, the amino termini of ISA1 frommonocots and dicots diverge greatly in character, whereas theremainder of the proteins are highly conserved (Fig. S1).
Discussion
ISA1 homomultimer function in the maize leaf
Biochemical characterization and genetic analysis both demon-strated that ISA1 functions as a homomeric enzyme, in theabsence of ISA2, in maize leaves. ISA enzymatic activity wasdetected by zymogram in isa2-339 null mutant leaves. Thisenzyme eluted as a multimeric complex in GPC analysis, atapproximately the same volume as the ISA1 homomultimer fromthe endosperm. ISA1 protein detected by immunoblot analysisco-eluted in GPC and co-migrated in native-PAGE with this ISAactivity. An ISA activity with the same purification properties waspresent in nonmutant leaves and did not contain any ISA2, asshown by immunoblot. The function of the ISA1 homomultimerwas implied by the finding that leaf starch granules generated inthe presence of ISA1 alone, that is, in isa2-339 mutants, differedin size and morphology from those of su1- mutants lacking ISA1activity. Also, the amylopectin chain length distribution was dif-ferent in starches generated in the presence of ISA1 alone com-pared with su1- mutants. Taken together, the data provide strongevidence that ISA1 functions enzymatically and physiologicallyin the maize leaf, in the absence of ISA2.
The ISA1 homomer, however, does not provide the full physi-ological function of ISA activity in maize leaves. This is in con-trast to the endosperm, where loss of ISA2, and thus the ISA1/ISA2 heteromeric enzyme, can be tolerated without major defects
Fig. 8 Alignment of isoamylase-type starch debranching enzyme 1 (ISA1) primary sequences from monocots and dicots. GenBank references are asfollows: maize (Zea mays), ACG43008; rice (Oryza sativa), ACY56095; wheat (Triticum aestivum), CAC82925; barley (Hordeum vulgare), AAM46866;Brachypodium distachyon, XP_003572394; Arabidopsis thaliana, AEC09752; poplar (Populus trichocarpa), EEF03224; Phaseolus vulgaris, BAF52941;grape (Vitis vinifera), XP_002265964; potato (Solanum tuberosum), AAN15317; pea (Pisum sativum), AAZ81835; sweet potato (Ipomoea batatas),AAY84833. The position of the first amino acid in each block in the full-length maize sequence is indicated. Asterisks indicate conservation in all 12 speciesand colons indicate conservative substitutions. The first block begins at the known mature N terminus or the teminus predicted by the ChloroP algorithm.
New Phytologist (2013) No claim to original US goverment works
New Phytologist� 2013 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist10

in starch content or structure (Kubo et al., 2010; Utsumi et al.,2011). The need for ISA2 in leaves is evident from the fact thatstarch content is reduced, starch structure is altered, and granulesare smaller on average in mutants lacking the protein comparedwith nonmutants. Thus, the ISA1 homomultimer cannot supportstarch metabolism that is normal in all of these aspects. Takentogether, the data indicate that both homomeric and heteromericISA forms are capable of function in maize leaf starch biogenesis,raising the possibility that they each play specific roles necessaryfor normal metabolism. The ISA1/ISA2 heteromultimer appearsto affect granule growth because the starch particles were smallerin the mutant lacking ISA2 than in the nonmutant standard.
The fact that the ISA1 homomultimer functions in the maizeleaf but not in A. thaliana is most simply explained by inherentdifferences in protein structure between the two species. TheA. thaliana leaf possesses only a single electrophoretic mobilityform of ISA activity, whereas one ISA1 homomultimer and twoISA1/ISA2 heteromeric forms are present in the maize leaf as theyare in the endosperm. Specific structural differences betweenISA1 proteins clearly exist that are monocot- or dicot-specific,within the context of very close sequence homology throughoutthe protein in all vascular plants examined. The positions of thelocalized regions of sequence divergence between monocot anddicot, based on the structure of Pseudomonas amyloderamosa iso-amylase (Katsuya et al., 1998), are consistent with this suggestion.The Ser-rich sequence specific to monocots aligns with a loop onthe external surface of the isoamylase protein (Fig. S2). Thisregion could thus influence interactions with other subunits inthe quaternary structure of the plant enzyme and allow monocotISA1 to function without ISA2. The acid-rich region at the Nterminus of the monocot ISA1 is not represented in theP. amyloderamosa protein, so its position within the three-dimen-sional structure cannot be predicted. The high degree of conser-vation specifically among the monocots, however, suggests thatthis region is functionally significant.
Contrary to this view, however, is the fact that the ISA1 homo-multimer was not detected in the leaves of rice, another monocotspecies (Utsumi et al., 2011). From this observation, the possibil-ity must be considered that particular aspects of leaf metabolismdetermine whether or not the ISA1 homomultimer forms and/orfunctions physiologically, and that these aspects differ in maizecompared with both rice and A. thaliana. For example, A. thalianaand rice are C3 plants that fix CO2 directly using RUBISCO,whereas maize is a C4 plant that initially fixes CO2 using pyruvateas the acceptor. Transfer of maize ISA1 to A. thaliana plants lack-ing both endogenous ISA1 and ISA2, however, showed that theC4 monocot enzyme could support starch biosynthesis in the C3
dicot (M. Facon & F. Wattebled, unpublished). This result sup-ports the interpretation that the maize ISA1 structure inherentlyprovides the ability for homomultimer function in the absence ofISA2. The reason that the ISA1 homomultimer was not detectedin rice leaves, despite its conserved primary structure comparedwith maize, is not obvious. The different results could relate to thestability of the enzyme, variation in the relative abundances ofISA1 and ISA2 in the leaves of the two species, and/or regulationof the enzymes by post-translational mechanisms.
Physiological aspects of the mutant phenotypes
Allele-specific distinctions were observed in the effects of su1-Ref and su1-4582 on leaf starch, although qualitatively the twomutations caused similar changes. For example, although bothmutations resulted in granules that were smaller than those pres-ent in nonmutant or isa2-339 mutant leaves, su1-4582 seemed tohave more severe effects than su1-Ref (Fig. 5). Quantitativelydifferent effects of the two alleles on amylopectin chain lengthdistribution were also observed (Fig. 7), and the appearance ofgranules visualized by TEM also differed slightly (Fig. 4). Asnoted, su1-Ref mutants contain reduced abundances of aninactive form of ISA1, whereas su1-4582 plants completely lackISA1. Furthermore, ISA2 accumulates to normal levels in thesu1-Ref endosperm, whereas it is completely missing from eitherthe leaf or the endosperm of su1-4582 mutants (Fig. 1; Kuboet al., 2010). Thus, the allele-specific effects on leaf starch poten-tially could result from the differences in ISA1 or ISA2 proteinabundance in the different mutants, even though enzymatic activ-ity is missing in both instances. Alternatively, the differencescould result from the growth environments of the two biologicalreplicates, which utilized seedlings grown in the glasshouse at dif-ferent times of the year. The latter explanation is supported forthe granule-size phenotype by the fact that the average diametersof all three genotypes were smaller in the second biological repli-cation of the experiment (Fig. 5).
A clear difference between maize and A. thaliana leaves was theabsence of phytoglycogen in the monocot when ISA1 or ISA2 wasmissing. This result is unusual in comparison to other geneticanalyses of ISA function, where in general phytoglycogenaccumulation is a hallmark of the mutant phenotype (Hennen-Bierwagen et al., 2012). Tissue specificity is part of the explana-tion, because ISA1 mutants of maize exhibit high concentrationsof phytoglycogen in the endosperm. The requirement for ISAfunction in the A. thaliana leaf varies by cell type, as shown by thepresence of starch granules in some cells and their apparentreplacement by phytoglycogen in others (Delatte et al., 2005).Bundle sheath and epidermal cell chloroplasts, for example, con-tain abnormally shaped starch granules in isa1- or isa2- mutants,whereas adjacent palisade or spongy mesophyll cells lack granules.Thus, ISA activity is not entirely responsible for whether or notglucans assemble into granules, but rather it appears to be a con-tributing factor that influences the equilibrium between crystalli-zation and solubility. Other enzymes are known to affect thedistribution between soluble and insoluble glucans, as shown bymultiple mutant combinations where ISA is compromised, forexample starch synthase III in the maize endosperm (Lin et al.,2011) and the a-amylase AMY3 in A. thaliana (Streb et al.,2008). Mesophyll chloroplasts in mature maize leaves do notaccumulate starch granules during normal vegetative growthbecause the Calvin cycle is not operating in those cells as a conse-quence of C4 metabolism (Majeran et al., 2010). Starch in maizeleaves accumulates for the most part in bundle sheath cells, whereRUBISCO and subsequent photosynthetic carbon metabolismreactions occur. Potentially, the metabolic nature of maize bundlesheath cells allows crystallization of precursor glucans into starch
No claim to original US goverment works
New Phytologist� 2013 New Phytologist TrustNew Phytologist (2013)
www.newphytologist.com
NewPhytologist Research 11

granules in the absence of ISA1, as it does in certain cell types inA. thaliana leaves, and this could explain why phytoglycogen wasnot observed in su1- or isa2- mutant maize leaves. Despite theabsence of phytoglycogen, the formation of starch in the maizeleaf in the absence of ISA1 or ISA2 is abnormal. This was shownby reduced contents of total granular material, abnormal granulesize and shape, and changes in amylopectin fine structure.Another consideration regarding the lack of measurable WSP inmutant maize leaves is the fact that in A. thaliana phytoglycogenis degraded to some extent during the day, as shown by accumula-tion of the breakdown product maltose (Delatte et al., 2005; Strebet al., 2008). Whether or not such degradation occurs in maizesu1- or isa2-mutant leaves and contributes to the lack of observedWSP and the reduced total glucan contents remains to be deter-mined.
The decrease in total glucan polymer in the maize su1- or isa2-mutants had no obvious effect on plant health or growth rate,consistent with the fact that plants with su1-Ref and other allelesare widely grown as commercial sweetcorn varieties (Tracy et al.,2006). This might be explained by the fact that the amount ofstarch that accumulates in the mutants, c. 36–46% of normal, issufficient to meet the physiological demands of the plant duringthe dark phase of the diurnal cycle. Some amount of leaf starch isrequired for normal maize growth, as shown by analysis of anear-starchless mutant affected in leaf ADPglucose pyrophos-phorylase (Schlosser et al., 2012). In these mutants grown in fieldconditions, slight effects were observed on plant height, floweringtime, and seed yield; however, plant health was not severely com-promised. Thus, there appears to be some degree of physiologicalplasticity regarding how maize plants utilize photosynthate andtransitory starch.
In summary, homomeric ISA1 was detected in maize leavesand shown to partially supply the in vivo requirement for ISAactivity in starch biosynthesis. This is the first demonstration thathomomeric ISA1 functions in leaf tissue. The ISA1/ISA2 hetero-multimer was also shown to function in the maize leaf, althoughwhether or not this form by itself could supply full physiologicalactivity remains to be determined. The functional differencesbetween maize and A. thaliana leaves, where ISA1 alone appar-ently is not active, are probably explained by structural differencesthat arose by evolutionary divergence in the monocot lineage.
Acknowledgements
The authors acknowledge T. Pepper and the Iowa State Univer-sity Microscopy and NanoImaging Facility for performing theTEM analyses and C. Lancelon-Pin (CERMAV) for the SEMobservation of purified starch granules.
References
Burton RA, Jenner H, Carrangis L, Fahy B, Fincher GB, Hylton C, Laurie DA,
Parker M, Waite D, van Wegen S et al. 2002. Starch granule initiation and
growth are altered in barley mutants that lack isoamylase activity. Plant Journal31: 97–112.
Bustos R, Fahy B, Hylton CM, Seale R, Nebane NM, Edwards A, Martin C,
Smith AM. 2004. Starch granule initiation is controlled by a heteromultimeric
isoamylase in potato tubers. Proceedings of the National Academy of Sciences,USA 101: 2215–2220.
Colleoni C, Myers AM, James MG. 2003.One- and two-dimensional native
PAGE activity gel analyses of maize endosperm proteins reveal functional
interactions between specific starch metabolizing enzymes. Journal of AppliedGlycoscience 50: 207–212.
Dauvillee D, Colleoni C, Mouille G, Buleon A, Gallant DJ, Bouchet B, Morell
MK, d’Hulst C, Myers AM, Ball SG. 2001. Two loci control phytoglycogen
production in the monocellular green alga Chlamydomonas reinhardtii. PlantPhysiology 125: 1710–1722.
Davidson RM, Hansey CN, Gowda M, Childs KL, Lin H, Vaillancourt B,
Sekhon RS, de Leon N, Kaeppler SM, Jiang N et al. 2011. Utility of RNA
sequencing for analysis of maize reproductive transcriptomes. Plant Genome 4:191–203.
Delatte T, Trevisan M, Parker ML, Zeeman SC. 2005. Arabidopsis mutants
Atisa1 and Atisa2 have identical phenotypes and lack the same multimeric
isoamylase, which influences the branch point distribution of amylopectin
during starch synthesis. Plant Journal 41: 815–830.Delatte T, Umhang M, Trevisan M, Eicke S, Thorneycroft D, Smith SM,
Zeeman SC. 2006. Evidence for distinct mechanisms of starch granule
breakdown in plants. Journal of Biological Chemistry 281: 12050–12059.Deschamps P, Moreau H, Worden AZ, Dauvillee D, Ball SG. 2008. Early gene
duplication within chloroplastida and its correspondence with relocation of
starch metabolism to chloroplasts. Genetics 178: 2373–2387.Dinges JR, Colleoni C, Myers AM, James MG. 2001.Molecular structure of
three mutations at the maize sugary1 locus and their allele-specific phenotypic
effects. Plant Physiology 125: 1406–1418.Hennen-Bierwagen TA, James MG, Myers AM. 2012. Involvement of
debranching enzymes in starch biosynthesis. In: Tetlow IJ, ed. Essential reviewsin experimental biology, volume 5. The synthesis and breakdown of starch.London, UK: Society For Experimental Biology, 179–215.
Henrissat B. 1991. A classification of glycosyl hydrolases based on amino-acid
sequence similarities. Biochemistry Journal 280: 309–316.Hussain H, Mant A, Seale R, Zeeman S, Hinchliffe E, Edwards A, Hylton C,
Bornemann S, Smith AM, Martin C et al. 2003. Three isoforms of isoamylase
contribute different catalytic properties for the debranching of potato glucans.
The Plant Cell 15: 133–149.James MG, Robertson DS, Myers AM. 1995. Characterization of the maize gene
sugary1, a determinant of starch composition in kernels. The Plant Cell 7:417–429.
Katsuya Y, Mezaki Y, Kubota M, Matsuura Y. 1998. Three-dimensional
structure of Pseudomonas isoamylase at 2.2 A resolution. Journal of MolecularBiology 281: 885–897.
Kubo A, Colleoni C, Dinges JR, Lin Q, Lappe RR, Rivenbark JG, Meyer AJ,
Ball SG, James MG, Hennen-Bierwagen TA et al. 2010. Functions ofheteromeric and homomeric isoamylase-type starch debranching enzymes in
developing maize endosperm. Plant Physiology 153: 956–969.Li P, Ponnala L, Gandotra N, Wang L, Si Y, Tausta SL, Kebrom TH, Provart
N, Patel R, Myers CR et al. 2010. The developmental dynamics of the maize
leaf transcriptome. Nature Genetics 42: 1060–1067.Lin Q, Huang B, Zhang M, Rivenbark JG, Zhang X, James MG, Myers AM,
Hennen-Bierwagen TA. 2011. Functional interactions between starch synthase
III and isoamylase-type starch debranching enzyme in maize endosperm. PlantPhysiology 158: 679–692.
Majeran W, Friso G, Ponnala L, Connolly B, Huang M, Reidel E, Zhang C,
Asakura Y, Bhuiyan NH, Sun Q et al. 2010. Structural and metabolic
transitions of C4 leaf development and differentiation defined by
micro-scopy and quantitative proteomics in maize. The Plant Cell 22:3509–3542.
Mouille G, Maddelein ML, Libessart N, Talaga P, Decq A, Delrue B, Ball S.
1996. Preamylopectin processing: a mandatory step for starch biosynthesis in
plants. The Plant Cell 8: 1353–1366.Nakamura Y, Umemoto T, Takahata T, Komae K, Amano E, Satoh H. 1996.
Changes in structure of starch and enzyme activities affected by sugary
mutations in developing rice endosperm: possible role of starch debranching
enzyme (R-enzyme) in amylopectin biosynthesis. Physiologia Plantarum 97:
491–498.
New Phytologist (2013) No claim to original US goverment works
New Phytologist� 2013 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist12

O’Shea MG, Samuel MS, Konik CM, Morell MK. 1998. Fluorophore-assisted
carbohydrate electrophoresis (FACE) of oligosaccharides: efficiency of labelling
and high-resolution separation. Carbohydrate Research 307: 1–12.Schlosser AJ, Martin JM, Hannah LC, Giroux MJ. 2012. The maize leaf starch
mutation has diminished field growth and productivity. Crop Science 52:700–706.
Streb S, Delatte T, Umhang M, Eicke S, Schorderet M, Reinhardt D, Zeeman
SC. 2008. Starch granule biosynthesis in Arabidopsis is abolished by removal of
all debranching enzymes but restored by the subsequent removal of an
endoamylase. The Plant Cell 20: 3448–3466.Streb S, Zeeman S. 2012. Starch metabolism in Arabidopsis. The ArabidopsisBook 10: e0160.
Thompson JD, Higgins DG, Gibson TJ. 1994. CLUSTALW: improving the
sensitivity of progressive multiple sequence alignment through sequence
weighting, position specific gap penalties and weight matrix choice. NucleicAcids Research 22: 4673–4680.
Tracy WF, Whitt SR, Buckler ES. 2006. Recurrent mutation in genomic
evolution: example of Sugary1 and the origin of sweet maize. Crop Science 46:S49–S54.
Utsumi Y, Nakamura Y. 2006. Structural and enzymatic characterization of the
isoamylase1 homo-oligomer and the isoamylase1-isoamylase2 hetero-oligomer
from rice endosperm. Planta 225: 75–87.Utsumi Y, Utsumi C, Sawada T, Fujita N, Nakamura Y. 2011. Functional
diversity of isoamylase oligomers: the ISA1 homo-oligomer Is essential for
amylopectin biosynthesis in rice endosperm. Plant Physiology 156: 61–77.Wattebled F, Dong Y, Dumez S, Delvalle D, Planchot V, Berbezy P, Vyas D,
Colonna P, Chatterjee M, Ball S et al. 2005.Mutants of Arabidopsis lacking achloroplastic isoamylase accumulate phytoglycogen and an abnormal form of
amylopectin. Plant Physiology 138: 184–195.Wattebled F, Planchot V, Dong Y, Szydlowski N, Pontoire B, Devin A, Ball S,
D’Hulst C. 2008. Further evidence for the mandatory nature of polysaccharide
debranching for the aggregation of semicrystalline starch and for overlapping
functions of debranching enzymes in Arabidopsis leaves. Plant Physiology 148:1309–1323.
Yandeau-Nelson MD, Laurens L, Shi Z, Xia H, Smith AM, Guiltinan MJ.
2011. Starch-branching enzyme IIa is required for proper diurnal cycling of
starch in leaves of maize. Plant Physiology 156: 479–490.Zeeman SC, Umemoto T, Lue W-L, Au-Yeung P, Martin C, Smith AM, Chen
J. 1998. A mutant of Arabidopsis lacking a chloroplastic isoamylase accumulates
both starch and phytoglycogen. The Plant Cell 10: 1699–1711.
Supporting Information
Additional supporting information may be found in the onlineversion of this article.
Fig. S1 Multiple sequence alignment of the primary structures ofmonocot and dicot ISA1 proteins.
Fig. S2 Structure of P. amyloderamosa isoamylase and predictedlocation of maize ISA1 residues based on primary sequence align-ment.
Please note: Wiley Blackwell are not responsible for the contentor functionality of any supporting information supplied by theauthors. Any queries (other than missing material) should bedirected to the New Phytologist Central Office.
New Phytologist is an electronic (online-only) journal owned by the New Phytologist Trust, a not-for-profit organization dedicatedto the promotion of plant science, facilitating projects from symposia to free access for our Tansley reviews.
Regular papers, Letters, Research reviews, Rapid reports and both Modelling/Theory and Methods papers are encouraged. We are committed to rapid processing, from online submission through to publication ‘as ready’ via Early View – our average timeto decision is <25 days. There are no page or colour charges and a PDF version will be provided for each article.
The journal is available online at Wiley Online Library. Visit www.newphytologist.com to search the articles and register for tableof contents email alerts.
If you have any questions, do get in touch with Central Office ([email protected]) or, if it is more convenient,our USA Office ([email protected])
For submission instructions, subscription and all the latest information visit www.newphytologist.com
No claim to original US goverment works
New Phytologist� 2013 New Phytologist TrustNew Phytologist (2013)
www.newphytologist.com
NewPhytologist Research 13