freeze-fracturing for low-temperature scanning electron microscopy of hartig net in synthesized...
TRANSCRIPT

\'ew Phytol. (1993), 123, 123-132
Freeze-fracturing for low-temperaturescanning electron microscopy of Hartig netin synthesized Picea abies - Hebelomacrustuliniforme and - Tricholoma vaccinumectomycorrhizas"^
BY C H R I S T O P H S C H E I D E G G E R AND I V A N O B R U N N E R
Swiss Federal Institute for Forest, Snow and Landscape Research {WSL),CH-8903 Birmensdorf, Switzerland
{Received 5 February 1992; accepted 21 August 1992)
SUMMARY
Rapidly frozen ectomycorrhizal roots were freeze-fractured under high vacuum in a dedicated cryo-preparationunit and investigated in the frozen, partially freeze-dried, or fully hydrated state. Tangential, radial and occasionaltransverse surface views of Hartig net were obtained after freeze-fracturing. Protoplasmic and exoplasmic fracturefaces of the fungal plasmalemma and occasional fractures hetween the fungal cell wall and the extracellular matrixwere seen. Primary pit-fields with plasmodesmata occurred in non-ectomycorrhizal parts of the root as well as insmall gaps of Hartig net. Septa with dolipores were observed in the finger-like complexes of the Hartig net.
Key words: Synthesized ectomycorrhizas, freeze-fracturing, low-temperature scanning electron microscopy,Hartig net.
INTRODUCTION
One of the original features used to confirm theectomycorrhizal status of a lateral root is the presenceof a Hartig net in the epidermis alone or in epidermisand cortex (Marks & Foster, 1973; Harley & Smith,1983). The Hartig net is so called because intransverse sections of ectomycorrhizal roots hyphaebetween epidermal and cortical cells have a net-likeappearance. In fact, hyphae of the Hartig netcompletely envelope the host cells to provide maxi-mum contact between host and fungus. The natureof such plectenchyma and how these hyphae areformed has been the subject of controversy. TheHartig net exhibits a complex labyrinthine growthmode with finger-like structures termed palmettesand with rare hyphal septations (Blasius et al., 1986).This labyrinthine effect has been explained as aninfolding due to compression and anastomoses(Duddridge & Read, 1984a), incomplete septationsor wall ingrowth (Massicotte et al., 1986), and walloutgrowth (Nylund & Unestam, 1982; Blasius et al..
* Picea abies (L.) Karst; Hebeloma crustuliniforme (Bull, ex St.Amans) Quel.; Tricholoma vaccinum (Pers. ex Fr.) Kummer.
1986). It is now accepted that such fingerlikecomplexes are highly organized structures whichenable an active bidirectional transfer of nutrients(Kottke & Oberwinkler, 1989). The morphology ofsuch cells is best understood as highly modifiedtransfer cell-like hyphae (Kottke & Oberwinkler,1987).
Similar morphological alterations of the hyphalgrowth form can also be observed when hyphaecontact root surfaces (Jacobs, Peterson & Massicotte,1989; Moore, Massicotte & Peterson, 1989; Brunner& Scheidegger, 1992). Hyphae then change frombeing elongated and rarely branched to short andextensively branched, with a somewhat swollen andfused appearance. The three-dimensional architec-ture of the Hartig net is difficult to demonstrate.Three-dimensional models have been proposedbased on light and transmission electron micro-graphs from transverse and longitudinal sections ofectomycorrhizal roots (Blasius et al., 1986; Melvilleet al., 1987). Others have used the scanning electronmicroscope after physical detachment of the mantleand upper cortical regions (StruUu, 1979) or afterfreeze fracturing of ectomycorrhizal roots (Nylundet al., 1982). Micrographs of cryo-fixed ectomy-

124 C. Scheidegger and I. Brunner
... fU . •4
—LI—J
T
Figures 1-3. For legend see opposite.

Lozv temperature SEM of eetomycorrhizas 125
corrhizas have been published (Alexander, Jones &McHardy, 1987; Massicotte, Melville & Peterson,1987; Muller et al., 1988; Jones & McHardy 1991)but only Alexander et al. (1987) discussed theirresults and compared them with critical point-driedmaterial. From these studies it became evident thatfrozen, fully hydrated or partially freeze driedectomycorrhizal specimens showed better structuralpreservation than critical point-dried material.
The present study applies the freeze fracturingtechnique of cryofixed material to synthesized Piceaabies (L.) K^arst.-Hebeloma crustuliniforme (Bull, exSt. Amans) Quel. (Brunner, 1991; Brunner, Amiet &Schneider, 1991) and P. abies-Tricholoma vaccinum(Pers. ex Fr.) Kummer (Agerer, 1987 a, 6) eeto-mycorrhizas. Surface features of Hartig net hyphaeand root cells, their three-dimensional architecture,and the advantages and limitations of the techniqueare considered.
MATERIALS AND METHODS
Seeds of P. abies were collected in 1977 fromDielsdorf (Ct. Zurich, Switzerland), surfacesterilized for 30 min in 30% H.jOg, rinsed, andgerminated on water agar plates. After 2—3 wkseedlings were used for synthesis experiments.
Mycelia of H. crustuliniforme and T. vaccinumwere obtained from cultures isolated from fruitbodies collected in September 1990 in a 100-yr-oldP. abies forest in Moosboden (Ct. Freiburg,Switzerland).
Autoclaved polyethylene pouches 13x16 cm(Cellpack, Switzerland) including an activated char-coal filter paper (Schleicher & Schuell, Switzerland)were moistened with 10 ml modified Melin-Norkrans nutrient solution (MNN: Marx & Bryan,1975) without glucose and malt at pH 6-5. Oneseedling was inserted into each pouch, and after 2-3months, when the root systems had developed, 4—6inoculum discs were placed within 3 mm of thelateral roots and another 10 ml MNN (including5 g r^ glucose, no malt) was added. Fungal discswere grown prior to inoculation on solid MNN-media including glucose (10 g T ) and malt (3 g 1 )for 5 d. At inoculation two strips of foam wereintroduced into the pouches to provide air space.Sterile, distilled water was added frequently.The synthesis experiments were carried out in a
growth chamber with a 16 h photoperiod (PAR:100 fimol m-' s-') at 20 °C and 70% humidity.
Short roots were harvested after 40 d and fixed in2-5 % glutaraldehyde, dehydrated in alcohol, em-bedded in glycol-methacrylate, longitudinallysectioned (1-5//m), and stained for chitin withGiemsa. Microphotographs were taken with a LeitzAristoplan photomicroscope.
For low-temperature scanning electron micro-scopy (LTSEM) ectomycorrhizal lateral rootswere harvested after 75 d (Tricholoma) and 172 d(Hebeloma). They were embedded in a small dropletof water on an aluminum stub, and immediatelyfrozen in liquid nitrogen (LN2). The specimens werethen transferred to the cold stage in the preparationchamber of the Balzers SCU 020 scanning cryo-preparation unit (Muller et al., 1991; Scheidegger etal., 1991). If not otherwise stated the specimens werefractured with a microtome at — 90 °C and partiallyfreeze-dried in a high vacuum (P < 2x 10"* Pa) for1 min. Platinum sputter coating was then carried outafter raising the pressure to 2-2 Pa. The coating was10 nm thick, measured by a quartz thin film monitor.After coating, the specimens were transferred with amanipulator through the sliding vacuum valve ontothe SEM cold stage in a Philips SEM 515. Thetemperature on the cold stage in the microscopewas kept below — 120°C. After observation, thespecimens were withdrawn onto the cold stage inthe preparation chamber where the procedure offracturing and coating could be repeated up to fivetimes on the same preparation.
Nomenclature for freeze-fractured membranesfollows Branton et al. (1975).
RESULTS
Development of eetomycorrhizas
P. abies seedlings in growth pouches formed a well-developed root system with numerous first-orderlateral roots (Fig. 1). Hyphae emanating from fungalplugs contacted lateral roots and induced eeto-mycorrhizas. Eetomycorrhizas of H. crustuliniformedeveloped a white, thin and loose to dense mantle(Fig. 2) and a Hartig net (Fig. 3) reaching radially tothe endodermis and apically to the meristematiczone. Eetomycorrhizas of T. vaccinum developed ayellow, thick and dense mantle and a Hartig net
Figures 1-3. Seedlings of Picea abies inoculated with Hebeloma crustuliniforme.
Fig 1 Seedling in a growth pouch, 47 d after inoculation with mycelial plugs (*) of H. crustuliniforme.Numerous ectomycorrhizal lateral roots (arrowheads) have formed. Scale bar, 1 cm. Figure 2. Scanningelectron micrograph of a mature ectomycorrhizal lateral root 71 d after inoculation. The root is not fractured.A compact mantle (arrowhead) has formed. Scale bar, 01 mm. Figure 3. Light micrograph of matureectomycorrhizal lateral root 40 d after inoculation. Outer (OM) and mner (IM) mantle and a Hartig net (H)have developed between epidermal (E) and cortical (C) cells. Scale bar, :>0 fim.

126 C. Scheidegger and I. Brunner
Figures 4—7. For legend see opposite.

Low temperature SEM of ectomycorrhizas 127
reaching radially to the endodermis and apically tothe meristematic zone.
Ereeze fracturing
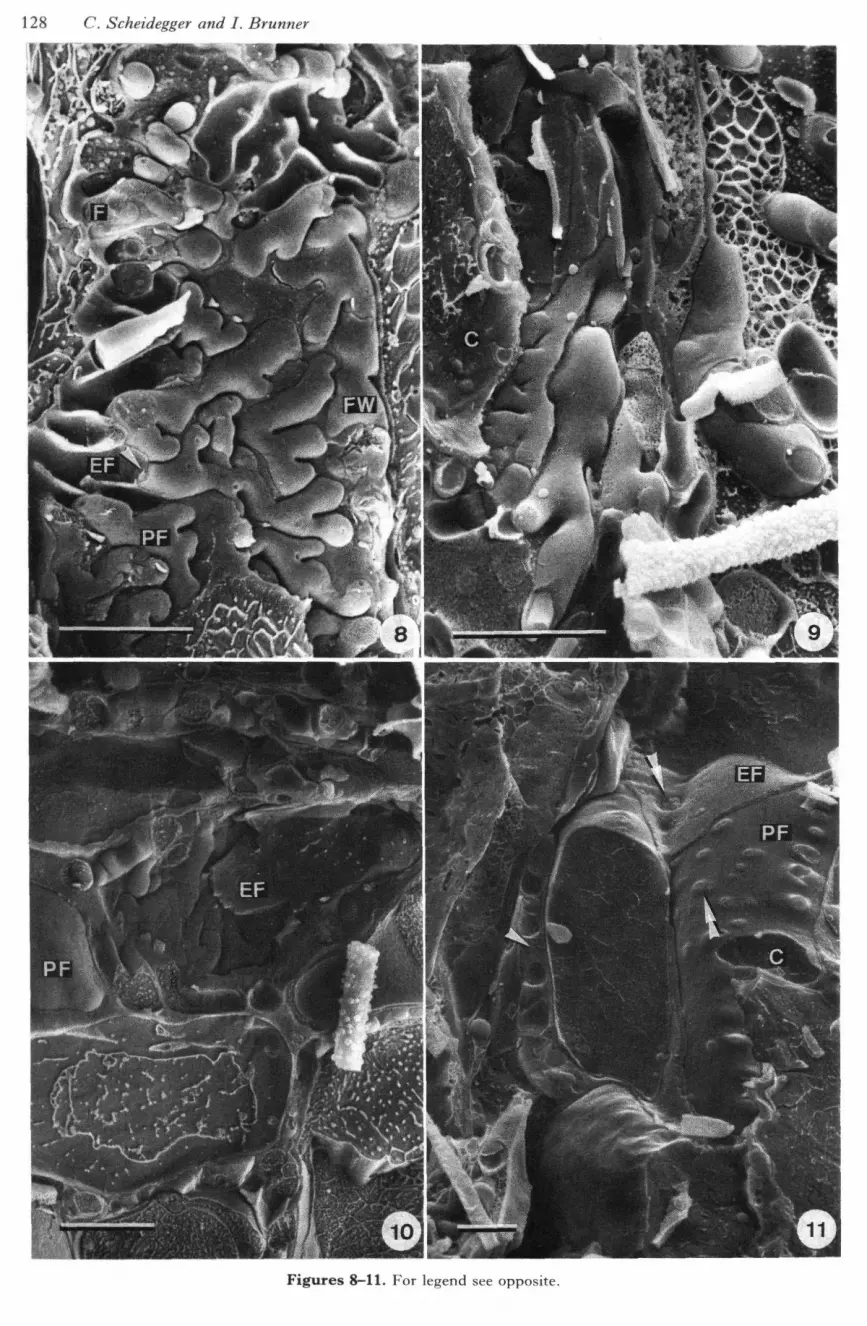
Transverse fractures of frozen, hydrated mycorrhizalroots were easy to obtain when the roots weremounted perpendicular to the fracturing plane. Withthis observation, however, most fracture planes werewithin root cells and surface views of Hartig net wereonly very rarely found. Better results were obtainedwhen ectomycorrhizal roots were mounted parallelor slightly inclined to the fracturing plane of themicrotome. With this orientation, tangential (Figs7, 8, 14, 15), radial (Figs 4, 12) and, rarely, alsotransverse (Fig. 9) surface views of the Hartig netwere obtained, although fractured parts withoutknife marks were always limited to minor areas of thepreparation. In most fractures through root cells orhyphae a typical pattern caused by crystallization ofpure water during the freezing process and sub-sequent etching of the pure water (Read & Jeffree,1988; Read, 1991) was visible (Fig. 8). Structureswith only minor amounts of water such as cell wallsand plasmalemmas were better preserved and nofreezing artifacts were visible. Root cells and fungalhyphae always had a turgid appearance and plasma-lemmas were always tightly adpressed to the cellwalls (e.g. Fig. 11).
Major evidence in the interpretation of structuressuch as fungal cell walls and fungal plasmalemmacame from hyphae where septa were fractured.Figures 7, 8 and 14 show fracture planes where septawere clearly visible. The convex fracture planes canbe interpreted as protoplasmic fracture faces (PF) ofthe fungal plasmalemma because the fracture step ofthe fungal walls can be seen. Therefore, the concavefracture planes are the exoplasmic fracture faces(EF). In these fractures the double walls between thefingerlike structures were clearly distinguishablefrom the much thinner septa. A fracture planebetween the fungal wall and the extracellular matrixcan be seen in Figure 8, where a fracture step fromthe fungal PF of the plasmalemma and the fungalwall was resolved.
Although no complementary fractures were avail-able by this method, the convex structures on the EF
of the fungal plasmalemma of T. vaccinum, seen aswhite dots, are probably particles sticking to theplasmalemma (Fig. 15). Impressions of similarparticles can be seen on the PF of the same hyphae.
Hebeloma crustuliniforme
In H. crustuliniforme ectomycorrhizas the hyphae ofthe outer mantle consisted of elongated cylindricalcells and were markedly verrucose. Clamps wereregularly found (see Brunner & Scheidegger, 1992)and branching was rarely observed (Fig. 2). Theinner mantle consisted only of one to a few hyphallayers. Its hyphae were tightly adpressed to the rootsurface or penetrated between damaged root capcells and epidermal cells. The surface of the hyphaewas smooth (Fig. 4).
Hyphae of the Hartig net were firmly attached tothe root cells and gaps between hyphae were filledwith an extracellular matrix (Figs 7, 11). Hyphalwalls were about 0-1 /im thick and the transition tothe extracellular matrix was often not clearly visible.
Fungal hyphae of the Hartig net of presumablyincompletely developed ectomycorrhizas often dis-played apical dominance and they were rarelybranched. Growth direction was centripetal andlongitudinal (Fig. 4). Such hyphae began to branchirregularly in their proximal parts and apical domi-nance was probably lost (Fig. 7). Fungal hyphae ofthe Hartig net of mature ectomycorrhizas had a non-polar growth and a completely changed hyphalmorphology. They lost their apical dominance,became multibranched and their original growthdirection became obscured by the fingerlike lateralhyphae (Fig. 8). Cortical root cells were slightlyindented by the Hartig net (Fig. 5) and formedinternal wall thickenings (Fig. 5). A few root cellwalls also had a different type of internal wallthickening with hemispherical inclusions (Fig. 6).
Tangential views of Hartig net revealed multi-branched hyphae which enveloped the cortical cellsover large areas (Figs 7, 8). Branches were mostlyparallel and were predominantly oriented per-pendicular to the root axis. In fully developedectomycorrhizas only small gaps between hyphalbranches were present. Only in such gaps wereprimary pit-fields with plasmodesmata found (Fig.
Figures 4-7. LTSEM micrographs of fractured Picea abies-Hebeloma crustuliniforme ectomycorrhizal roots.
Figure 4 Radial fracture of an ectomycorrhizal root. An outer (OM) and a monolayered inner mantle (IM) aswell as fractured hyphae of the Hartig net (arrowhead) between epidermal (E) and cortical (C) cells are present.Hyphae of the outer mantle show a microrugulate ornamentation which is not seen in the inner mantle^ Scalebar 10 am Figure 5 Transverse fracture of Hartig net and cortical cells (C). Cell walls are slightly mdentedby the Hartig net (arrowhead) and form internal wall thickenings (double arrowhead). Scale barl() /^m. Figure6. Radial fracture of Hartig net and cortical cells. Cortical cell (C) with marked cell wall thickemngs withinclusions (arrowhead). Scale bar, 10/.m. Figure 7. Tangential fracture of Hartig net. Exoplasm.c v.ew of theplasmalemma of a cortical cell showing primary pit-field (arrowhead) with plasmodesmata in a gap betweenhyphae of the Hartig net. Septa with dolipores are present (double arrowheads). Scale bar, 5 /xm
128 C. Scheidegger and I. Brunner
Figures 8-11. P'or legend see opposite.

Low temperature SEM of ectomycorrhizas 129
7). They also were more frequent in incompletelydeveloped ectomycorrhizas (Fig. 11).
Transverse (Fig. 9) and radial (Fig. 10) views ofthe Hartig net clearly showed that hyphae penetratedcentripetally between the cortical cells.
Tricholoma vaccinum
In T. vaccinum ectomycorrhizas the hyphae of theouter mantle were smooth but had an arachnoidincrustation. Clamps were not found. The innermantle was multilayered and penetrated betweenroot cap cells (Fig. 12). The hyphae were smooth,highly branched and strongly glued together. Tan-gential views of Hartig net showed clusters offingerlike hyphae in both protoplasmic andexoplasmic fractures of the plasmalemma (Figs 14,15). Fingerlike hyphae of one branch were mostlyparallel and completely covered the surface of thecortical cells. Septa with dolipores were found infingerlike clusters but were probably relatively rareand did not occur in every cluster. The fungal cellwall was about 0-1 /im thick. Cell wall thickenings ofcortical cells were never found in ectomycorrhizasof T. vaccinum (Fig. 13).
DISCUSSION
During Hartig net formation primary hyphae withapical dominance colonized intercellular spacesbetween cortical cells. Growth was predominantlylongitudinal and centripetal. These hyphae wererelatively elongate and rarely branched, and clampswere never observed, although they regularly occurin the outer and inner mantle of H. crustuliniformeectomycorrhizas (Brunner & Scheidegger, 1992).Intensive formation of outgrowths from primaryhyphae probably started soon after and led to analmost complete ensheathment of the cortical cellsby a hyphal monolayer. Growth of such fingerlikehyphae was either stopped or the growth directionwas changed when such outgrowths touched eachother. During this process the cortical cells becamealmost separated. The few gaps in the fungalenvelope were filled with an extracellular matrixand often showed primary pit-fields with plasmo-desmata, probably indicating symplastic continuity
(Nylund, 1980; Nylund et al., 1982; Nylund &Unestam, 1982; Massicotte e a/., 1987). Outgrowthswhich originate from closely related parts of theHartig net grew almost parallel and with a broadlobed front (Blasius et aL, 1986).
The difference in thickness of the double fungalwalls in the fingerlike structures and hyphal septawas obvious and supports the interpretation ofmultiple branching (Kottke & Oberwinkler, 1987),in contrast to lab^^rinthine growth of hyphae (Nylund& Unestam, 1982), or anastomosing hyphae andcomplex infoldings of fungal walls (Duddridge &Read, 1984 a).
Fractures of septa of fingerlike hyphae wereregularly found in both ectomycorrhizas studiedhere. Such septa have previously been reported byDebaud, Pepin & Bruchet (1981), Melville et al.(1988), and Massicotte et al. (1990). Light micro-scopy investigations of H. crustuliniforme-P. abiesectomycorrhizas indicate that septation of the finger-like complexes of Hartig net occur only relativelylate in ectomycorrhizal development. Septa arerarely found before 36 d after inoculation and occuronly in about one fifth of the fingerlike structures.After 72 d about one half of the fingerlike structurescontain septa (Scheidegger & Brunner, un-published). This contrasts with the observationsmade by Kottke & Oberwinkler (1987), who foundonly a few septa in Amanita muscaria (Pers. ex Fries)Hook.-P. abies ectomycorrhiza 3 months afterinoculation. They interpreted the coenocyticorganization of the fingerlike structures as anadaptation to active bidirectional transport. Ourresults suggest that septum formation in thefingerlike structures is considerably retarded but notcompletely stopped. Whether the coenocytic state ofsuch hyphae is temporary or permanent, and whatconsequences for the bidirectional transport this has,are questions for further studies.
Although many common characteristics werefound between H. crustuliniforme and T. vaccinumectomycorrhizas, some striking differences wereobvious and illustrate the value of freeze fracturingfor the general morphological characterization ofectomycorrhizas. Hyphal ornamentations or incrus-tations and the degree of clamp formation are
Figures 8-11. LTSEM micrographs of fractured Picea abies-Hebeloma crustuliniforme ectomycorrhizal roots.Scale bar, 10 //m.
Figure 8. Tangential fracture of Hartig net with many branched fingerlike hyphae. Fracture planes showexoplasmic (EF) and protoplasmic (PF) fracture faces of the fungal plasmalemma, fractures through the fungalcytoplasm (F) with cell organelles, and the fungal cell wall (FW). Note dolipores and septa where hyphae aretransversely fractured (arrowhead). Figure 9. Transverse fracture of Hartig net with centripetal orientation ofhyphae. (C), cortical cell. Figure 10. Radial fracture of Hartig net with centripetal orientation of hyphae.Protoplasmic (PF) and exoplasmic (EF) fracture faces of plasmalemma of cortical root cells are evident. Figure11. Radial fracture with incomplete ectomycorrhiza. Numerous primary pit-fields (double arrowheads) withplasmodesmata can be seen on protoplasmic (PF) and exoplasmic (EF) fracture faces of plasmalemma ofcortical cells (C) in regions where Hartig net has not developed. Tangential Hartig net is transversely fractured.Gaps between Hartig net hyphae are filled with extracellular matrix (arrowhead).
123

130 C. Scheidegger and I. Brunner
Figures 12—15. For legend see opposite.

Low temperature SEM of ectomycorrhizas 131
widely accepted characteristics in the description ofpure cultures of some fungi (Stalpers, 1978) and alsoshow their value in the two ectomycorrhizas dis-cussed here. Specimen preparation for conventionalSFM involves the use of organic solvents for de-hydration of samples and consequently hyphalincrustations of organic composition are often de-stroyed during preparation or are damaged underthe electron beam of the microscope (Scheidegger,unpublished). Thickness of the inner mantle is anadditional morphological difference between the twoectomycorrhizas. H. crustuliniforme induces shortwall ingrowths of cortical root cells, in contrast to T.vaccinum where such ingrowths were never observed.Wall ingrowths induced by Hartig net hyphae havepreviously been reported in ectomycorrhizas ofAlnus crispa (Ait.) Pursh (Massicotte et aL, 1986),Picea abies (Haug, 1987; Kottke & Oberwinkler,1988;), Pinus sylvestris L. (Duddridge & Read,19846: Warmbrodt & Fschrich, 1985) and Pisoniagrandis R. Br. (Ashford & Allaway, 1982). Thedifferent reactions of two alder species to Alpovadiplophloeus (Zeller & Dodge) Trappe & Smithwere discussed by Massicotte, Ackerley & Peterson(1989).
Surprisingly, freeze-fracture (transmission) elec-tron microscopy (Chapman & Staehelin, 1986) ofHartig net structures has not been described,probably because of the difficulties in the preparationof platinum-carbon replicas of plant material(Robards & Parish, 1971; Platt-Aloia & Thomson,1982). With low temperature scanning electronmicroscopy, direct observation of frozen, fullyhydrated or partially freeze-dried biological samplesis possible and the technical problems with replicapreparation are avoided (Beckett & Read, 1986;Sargent, 1986; 1988; Bastacky et al. 1987, 1988).With the dedicated SCU 020 cryo system (Mulleret aL, 1991) it is possible repeatedly to fracture, etchand sputter coat a given specimen.
Although many fractures of fungal hyphae androot cells could be interpreted satisfactorily, somedetails could not be unambiguously described. Thisis mainly due to the relatively restricted resolutionof a conventional SEM. Future developments willmainly be in two directions: first, field emission
SEM known for its higher resolution power thanconventional SEM, and which has resolved intra-membrane particles in frozen, partially freeze-driedyeast cells (Walther et aL, 1990) could be used inectomycorrhizal research; secondly, the preparationof complementary fractures (Read & Jeffree, 1991)could help in the interpretation of interfaces andthree-dimensional structures especially in complexsystems such as ectomycorrhizas.
ACKNOWLEDGEMENTS
We thank Mrs B. Schneider for sectioning and staining theectomycorrhizas, Mrs M. Zollinger for assistance insynthesizing the material, Mr P. Hatvani for assistance atthe LTSEM and Mrs M.J. Sieber for correcting theEnglish text. We gratefully acknowledge Professor 1. J.Alexander and an anonymous reviewer for their valuablesuggestions on the manuscript.
REFERENCES
Agerer R. 1987o. Studies on ectomycorrhizae IX. Mycorrhizaeformed by Tricholoma sulphureum and T. vaccinum on spruce.Mycotaxon 28: 327-360.
Agerer R. 19876. Tricholoma vaccinum. In: .'\gerer R, ed. Colouratlas of ectomycorrhizae. Schwabisch Gmiind : Einhorn-Verlag,plate 10.
Alexander C, Jones D, McHardy WJ. 1987. Scanning electronmicroscopy of cryofixed mycorrhizas of sitka spruce, Piceasitchensis (Bong.) Carr.: A comparison with critical point-driedmaterial. Nezv Phytologist 105: 613-717.
Ashford AE, Allaway WG. 1982. A sheathing mycorrhiza onPisonia grandis R. BR. (Nyctaginaceae) with development oftransfer cells rather than a Hartig net. New Phvtologist 90:511-519.
Bastacky J, Wodley C, La Brie R, Backus C. 1987. Ahihliography of low-temperature scanning electron microscopy(LTSEM, Cryo-SEM) and scanning electron microscopy offrozen hydrated biological systems. Scanni?ig 9: 219-225.
Bastacky J, Wodley C, La Brie R, Backus C. 1988. Abibliography of low-temperature scanning electron microscopy(LTSEM, Cryo-SEM) and scanning electron microscopy offrozen hydrated biological systems. Scanning 10: 37-38.
Beckett A, Read ND. 1986. Low temperature scanning electronmicroscopy. In; Aldrich HC, Todd WJ, eds. Ultrastructiiraltechniques for microorganisms. New York: Plenum Press, 45-86.
Blasius D, Feil W, Kottke I, Oberwinkler F. 1986. Hartig netformation in fully ensheated ectomycorrhizas. Nordic Journal ofBotany 6: 837-842.
Branton D, Bullivant S, Gilula NB, Karnovsky MJ, Moor H,Muhlethaler K, Northcote DH, Packer L, Satir B, Satir P,Speth V, Staehelin LA, Steere RL, Weinstein RS. 1975.Freeze-etching nomenclature. Science 190: 54—56.
Brunner 1. 1991. Comparative studies on ectomycorrhizae
Figures 12-15. LTSEM micrographs of fractured Picea abies-Tricholoma vaccinum ectomycorrhizal roots.
Figure 12. Radial fracture of ectomycorrhizal root with outer (OM) and multilayered inner mantle (IM). Innermantle penetrates between damaged root cap cells (RC) and between epidermal (E) and cortical (C) cellsmonolayered Hartig net is developed. Scale bar, \0 fim. Figure 13. Tangential fracture of cortical cells (C) witbuniformly thick walls. Clusters of parallel hyphae (arrowheads) have diflFerent growth directions. Scale bar,10//m. Figure 14. Tangential fracture of Hartig net. The Hartig net with protoplasmic (PF) and exoplasmic(EE) fracture faces of the plasmalemma is septated (arrowhead). Apices of hyphae are covered by a cortical cell(C). The difference between double walls and septa are clearly visible. Note bright structures in the exoplasmic- and the dark dots in the protoplasmic fracture face of the fungal plasmalemma. Scale bar, 5 /im. Figure 15.Tangential view of Hartig net with protoplasmic (PE) and exoplasmic (EE) fracture faces of the fungalplasmalemma. White dots on the protoplasmic fracture face of Hartig net are contammations. Scale bar, 5 /im.
9-2

132 C. Scheidegger and I. Brunner
synthesized with various in vitro techniques using Picea abiesand two Hebeloma species. Trees 5: 90-94.
Brunner I, Antiiet R, Schneider B. 1991. Characterization ofnaturally grown and in vitro synthesized ectomycorrhizas ofHebeloma crustuliniforme and Picea abies. Mycological Research95: 1407-1413.
Brunner I, Scheidegger C. 1992. Ontogeny of synthesized Piceaabies (L.) Karst.-Hebeloma crustuliniforme (Bull, ex St. Amans)Quel. ectomycorrhizas. New Phytologist 120: 359-369.
Chapman RL, Staehelin LA. 1986. Freeze-fracture (-etch)electron microscopy. In; Aldrich MC, Todd WJ, eds.Ultrastructure techniques for microorganisms. New York: PlenumPress, 213-240.
Debaud JC, Pepin R, Bruchet G. 1981. Ultrastructure desectomycorhizes synthetiques a Hebeloma alpinum et Hebelomamarginatulum de Dryas octopetala. Canadian Journal of Botany59: 2160-2166.
Duddridge JA, Read DJ. 1984a. The development andultrastructure of ectomycorrhizas. I. Ectomycorrhizal devel-opment on pine in the field. New Phytologist 96: 565-573.
Duddridge JA, Read DJ. 19846. Modification of the host-fungusinterface in mycorrhizas synthesized between Suillus bovinus(Fr.) O. Kuntze and Pinus svlvestris L. New Phytologist 96:583-588.
Harley JL, Smith SE. 1983. Mycorrhizal symbiosis. AcademicPress.
Haug I. 1987. Licht- und elektronenmikroskopische Untersuchungenan Mykorrhizen von Fichtenbestdnden im Schwarzwald. Ph.D.thesis, Eberhard-Karls-Universitat Tubingen.
Jacobs PF, Peterson RL, Massicotte HB. 1989. Altered fungalmorphogenesis during early stages of ectomycorrhiza formationin Eucalyptus pilularis. Scanning Microscopy 3 : 249-255.
Jones D, McHardy WJ. 1991. Why is it necessary to examinemycorrhizas in a scanning electron microscope ? ProceedingsRMS 26: 7\-75.
Kottke I, Oberwinkler F. 1987. The cellular structure of theHartig net: coenocytic and transfer cell-like organization.Nordic Journal of Botany 2: 85-95.
Kottke I, Oberwinkler F. 1988. Comparative studies on themycorrhization of Larix decidua and Picea abies by Suillusgrevillei. Trees 2: 115-128.
Kottke I, Oberwinkler F. 1989. Amplification of root-fungusinterface in ectomycorrhizae hy Hartig net architecture. Annalesdes Sciences Forestieres. 46 suppL, 737-740.
Marks GC, Foster RC. 1973. Structure, morphogensis andultrastructure of ectomycorrhizae. In: Marks GC, KozlowskiT T , eds. Ectomvcorrhizae : their ecology and physiology. NewYork: Academic Press, 1-41.
Marx DH, Bryan WC. 1975. Growth and ectomycorrhizaldevelopment of loholly pine seedlings in fumigated soil infestedwith the fungal symhiont Pisolithus tinctorius. Forest Science 21 :245-254.
Massicotte HB, Ackerley CA, Peterson RL. 1989. Ontogeny ofAlnus rubra-Alpova diplophloeus ectomycorrhizae. II trans-mission electron microscopy. Canadian Journal of Botany 67:201-210.
Massicotte HB, Melville LH, Peterson RL. 1987. Scanningelectron microscopy of ectomycorrhizae; potential andlimitations. Scanning Microscopy 1: 1439-1454.
Massicotte HB, Peterson RL, Ackerley CA, Ashford AE.1987. Ontogeny of Eucalyptus pilularis-Pisolithus tinctoriusectomycorrhizae II. Transmission electron microscopy.Canadian Journal of Botany 65: 1940-1947.
Massicotte HB, Peterson RL, Ackerley CA, Melville LH.1990. Structure and ontogeny of Betula alleghaniensis-Pisolithustinctorius ectomycorrhizae. Canadian Journal of Botany 68:579-593.
Massicotte HB, Peterson RL, Ackerley CA, Piche, Y. 1986.Structure and ontogeny of Alnus crispa-Alpova diplophloeusectomycorrhizae. Canadian Journal of Botany 64: 177-192.
Melville LH, Ackerley CA, Massicotte HB, Peterson RL.1987. Morphogenesis and model of Hartig net in two ecto-mycorrhizal systems. In: Sylvia DM, Hung LL, Graham JH.
eds. Mycorrhizae in the next decade. University of Florida,Gainesville: University of Florida, 210.
Melville LH, Massicotte HB, Ackerley CA, Peterson RL.1988. An ultrastructural study of modifications in Dryasintegrifolia and Hebeloma cylindrosporum during ecto-mycorrhiza formation. Botanical Gazette 149: 408-418.
Moore AEP, Massicotte HB, Peterson RL. 1989.Ectomycorrhiza formation between Eucalyptus pilularis Sm.and Hydnangium carneum Wallr. in Dietr. New Phytologist112: 193-204.
Muller T, Guggenheim R, Dtiggelin M, Mestres P, VanAelst AC, Heyser W, Kumpfer W. 1988. Freeze etching andcryo scanning microscopy (CSEM) of plant and animal tissuewith SCU 020. In: Dickinson HG, Goodhew PJ, eds. EUREM88; Proc. of the 9th European Congress on Electron Microscopyheld in York, England, 4-9 Sept, 1988, Vol. 3 : Biology. Bristoland Philadelphia: Institute of Physics Conference Series No 93,15-16.
Muller T, Guggenheim R, Diiggelin M, Scheidegger C.1991. Freeze-fracturing for conventional and field emissionlow-temperature scanning electron microscopy: the scanningcryo unit SCU 020. Journal of Microscopy 161: 73-83.
Nylund JE. 1980. Symplastic continuity during Hartig netformation in Norway spruce ectomycorrhizae. New Phytologist86: 373-378.
Nylund JE, Kasimir A, Strandberg Arveby A, Unestam T.1982. Simple diagnosis of ectomycorrhiza formation anddemonstration of the architecture of the Hartig net by means ofclearing technique. European Journal of Forest Pathology 12:103-107.
Nylund J, Unestam T. 1982. Structure and physiology ofectomycorrhizae. I. The process of mycorrhiza formation inNorway spruce in vitro. New Phytologist 91: 63-79.
Platt-Aloia KA, Thomson W. 1982. Freeze fracture of intactplant tissues. Stain Technology 57: 323—334.
Read N. 1991. Low-temperature scanning electron microscopyof fungi and fungus-plant interactions. In: Mendgen K,Lesemann DE. eds. Electron microscopy of plant pathogens.Berlin: Springer-Verlag, 17-29.
Read ND, Jeflfree CE. 1988. Common artifacts associated withbiological material examined hy low-temperature scanningelectron microscopy. In: Dickinson HG, Goodhew PJ, eds.EUREM 88 ; Proc. of the 9th European Congress on ElectronMicroscopy held in York, England, 4-9 Sept, 1988, Vol. 3:Biology. Bristol: Institute of Physics Conference Series Number93, 17-18.
Read ND, Jeffree CE. 1991. Low temperature SEM in biology.Journal of Microscopy 161: 59-72.
Robards AW, Parish GR. 1971. Preparation and mechanicalrequirements for freeze-etching thick-walled plant tissues.Journal of Microscopy 93: 61-66.
Sargent JA. 1986. Cryo-preservation of roots for scanningelectron microscopy. Annals of Botany 58: 183-185.
Sargent JA. 1988. Low temperature scanning electron mi-croscopy: advances and applications. Scanning Microscopy 2:835-849.
Scheidegger C, Giinthardt-Goerg M, Matyssek R, HatvaniP. 1991. Low-temperature scanning electron microscopy ofbirch leaves after exposure to ozone. Journal of Microscopy 161:85-95.
Stalpers JA. 1978. Identification of wood-inhabiting Aphyllo-phorales in pure culture. Studies in Mycology 16: 1-248.
Strullu DG. 1979. Ultrastructure et representation spatiale dumanteau fongique des ectomycorhizes. Canadian Journal ofBo/am'57: 2319-2324.
Walther P, Hentschel J, Herter P, Muller T, Zierold K. 1990.Imaging of intramemhranous particles in frozen-hydrated cells(Saccharomyces cerevisiae) hy high resolution cryo SEM.Scanning 12: 300-307.
Warmbrodt RD, Eschrich W. 1985. Studies on the mycorrhizasof Pinus sylvestris L. produced in vitro with the hasidiomyceteSuillus variegatus (SW. ex Fr.) O. Kuntze; I. Ultrastructure ofthe mycorrhizal rootlets. New Phytologist 100: 215-223.
