fossil remains of macaca sylvanus florentina (cocchi, 1872) (primates, cercopithecidae) from the...
TRANSCRIPT

Author's personal copy
Fossil remains of Macaca sylvanus !orentina (Cocchi, 1872) (Primates,Cercopithecidae) from the Early Pleistocene of Quibas (Murcia, Spain)
David M. Alba a,*, Juan Abel Carlos Calero b, Miguel Ángel Mancheño b, Plini Montoya c,Jorge Morales d, Lorenzo Rook e
a Institut Català de Paleontologia, Universitat Autònoma de Barcelona, Edi"ci ICP, Campus de la UAB s/n, 08193 Cerdanyola del Vallès, Barcelona, SpainbDepartamento de Química Agrícola, Geología y Edafología, Universidad de Murcia Campus de Espinardo, 30100 Murcia, SpaincÀrea de Paleontologia, Departament de Geologia, Universitat de València, c/ Doctor Moliner 50, 46100 Burjassot, València, SpaindDepartamento de Paleobiología, Museo Nacional de Ciencias Naturales, CSIC, c/ José Gutiérrez Abascal 2, 28006 Madrid, SpaineDipartimento di Scienze della Terra, Università degli Studi di Firenze, via G. La Pira 4, 50121 Firenze, Italy
a r t i c l e i n f o
Article history:Received 22 December 2010Accepted 13 September 2011
Keywords:Barbary macaqueDentitionTaxonomyCercopithecoideaIberian PeninsulaPlio-Pleistocene
a b s t r a c t
The macaque material from the Early Pleistocene site of Quibas (Albanilla, Murcia, Spain), includingdentognathic remains, isolated teeth and some postcranial bone fragments, is described. Both metricallyand morphologically, this sample must be attributed to Macaca sylvanus (the Barbary macaque). Thisspecies is currently distributed through North Africa and Gibraltar, but was much morewidely distributedduring the Plio-Pleistocene, being represented by several European fossil subspecies. Metrical compari-sons of dental size and proportions between extantM. s. sylvanus and fossilMacaca sylvanus !orentina fromthe type locality and other Italian sites are undertaken, in order to classify the remains from Quibas at thesubspecies level. The results show that the Quibas sample not only !ts the range of variation of M. s.!orentina from the type locality, but also differs from the extant Barbary macaque condition in severalregards. This permits us to formally attribute the material from Quibas to M. s. !orentina. The materialdescribed in this paper therefore signi!cantly improves the knowledge of this fossil taxon, particularlyregarding the upper dentition, and further con!rms the taxonomic distinctiveness of this extinct taxonat the subspecies rank. Taken as a whole, M. s. !orentina largely overlaps in dental dimensions withM. s. sylvanus, but differs from the latter by displaying (on average): (1) absolutely longer upper molars(especially M1 and M3); (2) relatively wider upper molars (especially M1 and M2); (3) longer M3 ascompared with the M2; (4) absolutely longer M1 and M3; and (5) relatively narrower M3.
! 2011 Elsevier Ltd. All rights reserved.
Introduction
European fossil macaques
The macaque lineage, which apparently diverged from otherPapionini by about 7Ma (millions of years ago) (Stewart and Disotell,1998), is !rst recorded in Europe in the latest Miocene (Köhler et al.,2000), and persists through the Pliocene and Pleistocene (Fooden,2007). There is consensus that fossil European macaques belong tothe Macaca sylvanus (Linnaeus, 1758) lineage, of African origin.According tomolecular studies,M. sylvanus is the sister taxonof otherextant species of this genus (Morales and Melnick, 1998) otherwiseknown exclusively from Asia. Current knowledge of the geographic
variation of the Barbary macaque is insuf!cient to warrant therecognition of extant subspecies (Fooden, 2007). On the contrary,with the exception ofMacacamajori, Plio-Pleistocenemacaques fromEurope are customarily referred to several subspecies ofM. sylvanus.The latter is currently represented by the nominotypical subspeciesinNorthAfrica, where they display a disjunct distribution in the Atlasand Rif Mountain ranges in Algeria and Morocco (Fooden, 2007), aswell as in Gibraltar. The latter population is undoubtedly of anthro-pochorous origin (Groves, 2001; Fooden, 2007; Masseti and Bruner,2009), apparently resulting from repeated introductions in histor-ical times. Romans occasionally imported thesemonkeys into Europeas pets or curiosities (Masseti and Bruner, 2009), but the presentGibraltar population is more likely descended from individualsof mixed Algerian-Moroccan origin that were introduced duringthe 20th century (Modolo et al., 2005).
With regard to fossil European macaques, three extinct Euro-pean subspecies are customarily recognized, following Szalay andDelson (1979) and Delson (1980): Macaca sylvanus prisca Gervais,
* Corresponding author.E-mail addresses: [email protected] (D.M. Alba), [email protected] (J.A. Carlos
Calero), [email protected] (M.Á. Mancheño), [email protected] (P. Montoya),[email protected] (J. Morales), lorenzo.rook@uni!.it (L. Rook).
Contents lists available at SciVerse ScienceDirect
Journal of Human Evolution
journal homepage: www.elsevier .com/locate/ jhevol
0047-2484/$ e see front matter ! 2011 Elsevier Ltd. All rights reserved.doi:10.1016/j.jhevol.2011.09.003
Journal of Human Evolution 61 (2011) 703e718

Author's personal copy
1859, from the Pliocene (earliest Ruscinian to early Villafranchian)of Europe (type locality: Montpellier, France); Macaca sylvanus!orentina Cocchi, 1872, from the late Pliocene to early Pleistocene(middle through !nal Villafranchian) of southern and centralEurope (type locality: Upper Valdarno, Italy); and Macaca sylvanuspliocena Owen, 1846, from the middle Pleistocene of Europe, theCaucasus and Israel (type locality: Grays Thurrock, England). Szalayand Delson (1979) and Delson (1980) provisionally considered,with some doubts, that M. majori from Sardinia was also a subspe-cies ofM. sylvanus, although noting some differences with the latterand considering that a distinct species status was likely for theformer. The latter opinion has been subsequently favored by mostscholars (e.g., Gentili et al., 1998; Zanaga, 1998; Delson et al., 2000;Köhler et al., 2000; Rook and O’Higgins, 2005; but see; Mazza et al.,2005). In fact, M. majori can be readily distinguished fromM. s. !orentina and other M. sylvanus subspecies on the basis ofdentognathic features, including the smaller dental size (withtypically very short P3), the less sharply-angled cusps, the more"ared and in"ated crowns, and the shorter palate of the former, aswell as on the basis of lower molar proportions (Zanaga, 1998;Mazza et al., 2005). It has been also shown that M. majoridisplays a derived cranial morphology, as shown among others byzygomatic morphology, probably as a result of dietary adaptations(Rook and O’Higgins, 2005; Smith et al., 2008). Given thatM. majoriis best interpreted as an insular, endemic species (Rook andO’Higgins, 2005), and that the aim of this paper is to describe andtaxonomically attribute the macaque material from Quibas,comparisons of the latter are restricted to extant M. s. sylvanus aswell as fossil M. s. !orentina, the latter being the subspecies towhich the Quibas material could a priori be attributed on chrono-logical grounds (e.g., Szalay and Delson, 1979).
The macaque from Quibas
In this paper, we describe a sample of dentognathic and post-cranial remains of M. sylvanus from the Early Pleistocene site ofQuibas (Albanilla, Murcia, Spain). Some of these remains were re-ported by Montoya et al. (1999, 2001), who !gured some of thespecimens and attributed them to M. sylvanus ssp., albeit notingthat they overlapped in size with Italian remains attributed to M. s.!orentina. Alba et al. (2008b) preliminarily attributed this sample tothe latter subspecies. However, neither Montoya et al. (1999) norAlba et al. (2008a) provided detailed descriptions or measurementsto support their taxonomic attributions. In this paper, we !gure anddescribe all of the available macaque remains from Quibas andprovide metric comparisons with other fossil and living macaques,which allows us to attribute the fossils to M. s. !orentina.
Age and geological background
The paleontological site of Quibas (Montoya et al., 1999, 2001;Mancheño et al., 2003; Carlos Calero et al., 2004) is located in theSE portion of the Sierra de Quibas, at the intersection of threedistinct faults (Rodríguez Estrella et al., 2004) (Fig. 1). The Sierra deQuibas is a 6 km long and 2.5 km wide carbonate massif of NEeSEdirection, which is mainly composed of Jurassic limestones anddolomites from the Subbetic region (Rodríguez Estrella et al.,2004). This massif was affected by considerable karstic activityduring the Plio-Pleistocene, resulting in numerous exokarstic andendokarstic structures (Durán et al., 2004), which are in!lled bychemical precipitates and/or by mixed detritic sediments. Thefossiliferous sediments from the Quibas site constitute the Pleis-tocene karstic in!lling of several cavities situated inside thedolomites of the lower Lias. Most of the fossils come from a 5 mwide and 9 m high gallery (known as Ec on maps of the cavesystem) that extends for more than 30 m in length, and displaysseveral collateral branches separated by calcitic speleothems(Durán et al., 2004; Rodríguez Estrella et al., 2004). One of these isa 20 m deep and 2 m wide chasm (known as feature S), whereseveral preliminary excavations have been carried out. In thepaleontological site, a 6 m thick stratigraphic sequence of reddishdetritic sediments outcrops in the wall crevices of a small aban-doned limestone quarry (see Fumanal and Blázquez in Montoyaet al., 1999 for further details on the stratigraphy). Six detriticlayers have been distinguished, being mainly composed of clays,silts and more or less cemented breccias. Magnetostratigraphiccorrelation indicates that the fossiliferous sediments correspondto the Late Matuyama subchron of reversed polarity (Carlos Caleroet al., 2004, 2006a,b), i.e., between 1.770 and 1.070 Ma. This datingagrees with the age of 1.3e1.0 Ma (probably closer to 1.3 Ma) thathad been previously suggested on biostratigraphic grounds(Montoya et al., 1999, 2001).
Material and methods
The fossil sample from Quibas
Montoya et al. (1999) !gured some of themacaque remains thenavailable from Quibas, which come from the debris of gallery Ec.After the 2002e2005 !eld campaigns, however, the macaquesample from Quibas has considerably increased, mainly includingseveral maxillary and mandibular fragments, as well as isolatedteeth and some postcranial remains (see Table 1), which are here!gured and described for the !rst time.
Figure 1. Location map of the site of Quibas (Albanilla, Murcia, Spain).
D.M. Alba et al. / Journal of Human Evolution 61 (2011) 703e718704

Author's personal copy
Comparative sample
Given the lack of de!nite morphologic criteria on which thesubspeci!c attribution of theMacaca remains from Quibas could bebased, we employed standard dental measurements of bothM. s. sylvanus and M. s. !orentina from the type locality (Upper
Valdarno) and from additional sites in central Italy (Lower Valdarnoand Mugello), in order to perform metrical comparisons of dentalsize and proportions. Measurements for M. s. !orentina were takenfrom original specimens and good quality casts housed at theMuseo di Storia Naturale (Sezione di Paleontologia e Geologia) inFlorence (Italy) by one of the authors (DMA). This collectionincludes IGF7510V and IGF7509V, respectively, a right M2 (cast) andleft M3 (cast) from Barberino di Mugello (the originals are housed inthe collections of theMuseo di Storia Naturale e del Territorio of theUniversity of Pisa, without numbers); IGF13088 right P4 and otherdental fragments from Ligniti del Pollaio near Barberino di Mugello;IGF7519V, right M3 from Castel!orentino (Lower Valdarno); andthe following specimens from several Upper Valdarno localities:IGF10034 (holotype), partial mandible with left I2eM3 and rightP3eM3 from Le Forre; IGF10035, right lower canine and P4eM2 andleft lower canine to M3 of a male individual from Le Mignaie;IGF13084, left mandibular fragment withM1eM3 from Le Ville nearTerranova; IGF13085 left M3 from Terranova; IGF13086, right I2
from Il Tasso; IGF13087, several isolated teeth (fragment of cheektooth, left P4, right M2, left M2 and left M3) from Il Tasso; IGF13083,M1eM3 (cast) from Upper Valdarno indeterminate (originalshoused at the Museo di Storia Naturale, Milan, without numbers);IGF13569, partial mandible with left P3eM3 (cast) from UpperValdarno indeterminate (original housed at the Museo Paleon-tologico dell’Accademia del Poggio, Montevarchi, withoutnumbers); IGF13570, partial mandible with complete dentition(cast) from Upper Valdarno indeterminate (original housed at theMuseo Geologico, Rome, without number); IGF14286, right and leftlower canine to M3 (cast) from Le Strette (original NMB VA 352,housed at the Basel Naturhistorisches Museum); IGF14287, leftP3eM3 (cast) from Inferno di Sotto (original NMB VA 2058);IGF14288, symphyseal fragment with incisor alveoli, both caninesand left P3eP4 (cast) from Inferno (original NMB VA 1415);IGF14289, left P4eM2 (cast) from Inferno (original NMB VA 1088);IGF14290, right P4eM1 (cast) from Tassinaia (original NMB VA2075); and IGF14291, right M1 (cast) from Inferno (original NMBVA1415). Additional measurements of European Pleistocenemacaqueswere taken from Zapfe (2001) and Alba et al. (2008a), whereasindividual measurements for extant M. s. sylvanus were taken fromZanaga (1998). Sex-speci!c average values for dental measure-ments of both upper and lower fourth premolars and molars inextant M. s. sylvanus are also available from Mottura and Gentili(2006).
Measurements and variables
Measurements of maximummesiodistal crown length (MD) andbuccolingual crown breadth (BL) were employed. In the case ofmolars, the latter measurement was taken separately at the mesialand the distal lobes. In the case of canines, MD was taken as themaximum crown diameter, whereas BL was taken perpendicular toit. An index re"ecting the particular proportions of each tooth, theBLI (breadth/length index), was computed on the basis of theabove-mentioned measurements as follows: BLI ! BL (maximum)/MD " 100.
Statistical methods
Bivariate plots of BL vsMD for upper and lower fourth premolarsand !rst, second and third molars were employed to visualize thescatter of points of the two comparative samples, as well as theproportions of fossil macaque material from Quibas and other fossillocalities. Graphics depicting the mean values of MD and BLI forboth upper and lower molar series were also employed to comparethe Quibas sample with extant M. s. sylvanus and M. s. !orentina
Table 1List of the fossil remains of Macaca sylvanus !orentina (Cocchi, 1872) from Quibas(Murcia, Spain).
Figure Record No. Description
3C0eD0 GCP-CV4052 Left upper canine (male); advanceddegree of wear
3E0eF0 GCP-CV4053 Left upper canine (male); crown verydamaged, root complete
3MeO GCP-CV4054 Right maxillary fragment with M1eM2
3PeQ GCP-CV4055 Right maxillary fragment with I2 alveolusand upper canine to P4 series (female)
3A0eB0 GCP-CV4056 Right upper canine (male); very worn(pulp cavity exposed), and apex of rootslightly damaged
3UeV GCP-CV4057 Right I2
3WeX GCP-CV4058 right deciduous upper caninee GCP-CV4059 Right (?) Lower canine distal crown and
root fragment2NeP GCP-CV4060 Right mandibular fragment with
M1eM3 series3ReT GCP-CV4061 Right P3 nearly complete (only tips
of roots missing)2QeS GCP-CV4062 Left M2 nearly complete (only apex
of distal root missing)2TeV GCP-CV4063 Left M3 with a slight degree of wear2ZeZ0 Q03-E-1 Mandibular fragment with no teeth2AeC Q03-E-4a Right mandibular fragment with P4eM2
series and M3 (in crypt)2DeG Q03-E-4b Left mandibular fragment with lower
canine germ (crown completely formed,and emerging), P3 (crown formed withroots, the mesial one still open, inside thecrypt just beginning to emerge)
2HeK Q03-E-4c Right lower canine (crown with open root)2LeM Q03-E-4d Right P3 (crown with roots, the mesial
one still open)e Q03-E-4e Small mandibular fragments with no teeth4GeJ Q08-E-165 Partial left proximal humeral fragment4CeD Q04-Ec-90 Proximal radial head3GeI Q03-E-155a Right maxillary fragment with distal crown
fragment of M1 and M2eM3 series; preservingpart of the palate and an anterior portionof the zygomatic
3JeL Q03-E-155b Left maxillary fragment with M2 (slightlydamaged) and M3; preserving the lowerzygomatic root
e Q03-E-155c Small maxillary fragments with no teethe Q03-E-155d Small maxillary fragments with no teeth4EeF Q04-Ei-C4-16 Proximal phalanx2WeY Q03-Ec-1 Left M3 unworn crown3AeC Q05-Ec-87 Right maxillary fragment with P3eM3 series;
preserving the lower root of the zygomatic3DeF Q05-Ec-88 Right maxilla and premaxilla with I1, I2 and
canine alveoli, and P3eM3 series (female);including premaxillary suture and most of thelateral rim of nasal aperture
4AeB Q05-Ec-89 Left distal humeral fragment3YeZ Q05-S10-21 Left maxillary fragment; very damaged with
no crowns, but including the alveoli and partof the roots of I1, I2, C and M2, and the rootsof P3, P4, M1 and M3, and also the lowermostportion of the zygomatic root
Notes: Fossil remains with the acronym GCP-CV (Grup Cultural Paleontològicd’ElxeComunitat Valenciana) are housed at the Museu Paleontològic d’Elx-MUPE(Elche, Spain), whereas the rest of the fossils are housed at the collections of theDepartamento de Química Agrícola, Geología y Edafología of the Universidad deMurcia (Spain). The Murcia numbers are indicated by the acronym Q (Quibas), fol-lowed by the year of collection, the locality provenance within the site (Ec, S10, etc.),and the !eld collection number within each locality.
D.M. Alba et al. / Journal of Human Evolution 61 (2011) 703e718 705
Author's personal copy
from Italy. Boxplots were used in the case of lower molars tovisualize the degree of overlap between both comparative samples,and to determine the sample to which the Quibas remains !t best.Finally, analysis of variance (ANOVA) was employed to statisticallytest the differences in mean MD, BL and BLI between bothcomparative samples. These analyses were performed with SPSS v.16.0, and were restricted to the lower molars because the sample ofupper molars of M. s. !orentina from Italy is too small.
Systematic paleontology
Order Primates Linnaeus, 1758Infraorder Catarrhini É. Geoffroy Saint-Hilaire, 1812Superfamily Cercopithecoidea Gray, 1821Family Cercopithecidae Gray, 1821
Subfamily Cercopithecinae Gray, 1821Tribe Papionini Burnett, 1828Subtribe Macacina Owen, 1843Genus Macaca Lacépède, 1799Macaca sylvanus (Linnaeus, 1758)Macaca sylvanus !orentina (Cocchi, 1872)(Figs. 2e4)
Material and measurements
Studied material See Table 1. The described material is currentlyhoused at the Museu Paleontològic d’Elx-MUPE (Elche, Spain) andat the collections of the Universidad de Murcia (Spain).Measurements (Tables 2 and 3).
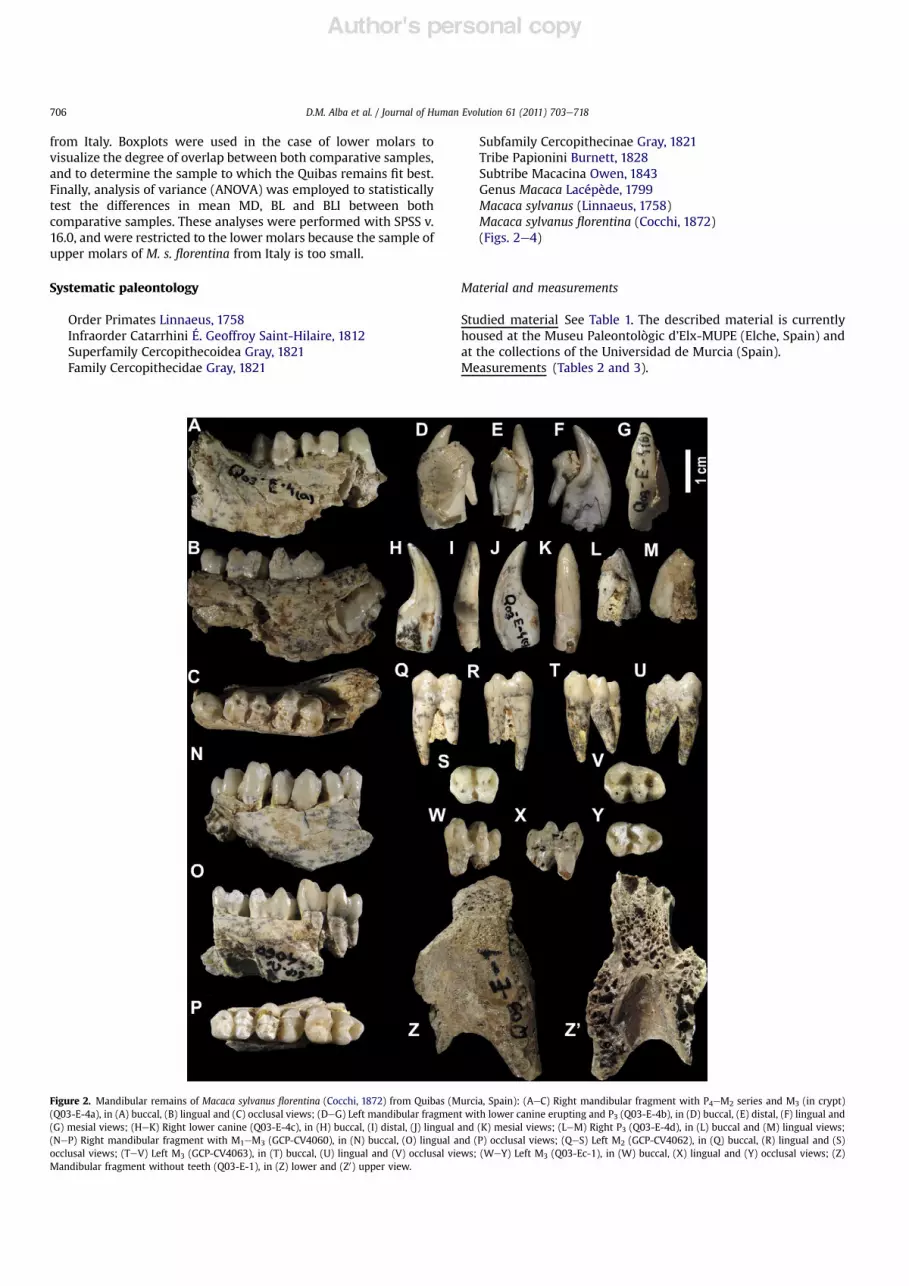
Figure 2. Mandibular remains of Macaca sylvanus !orentina (Cocchi, 1872) from Quibas (Murcia, Spain): (AeC) Right mandibular fragment with P4eM2 series and M3 (in crypt)(Q03-E-4a), in (A) buccal, (B) lingual and (C) occlusal views; (DeG) Left mandibular fragment with lower canine erupting and P3 (Q03-E-4b), in (D) buccal, (E) distal, (F) lingual and(G) mesial views; (HeK) Right lower canine (Q03-E-4c), in (H) buccal, (I) distal, (J) lingual and (K) mesial views; (LeM) Right P3 (Q03-E-4d), in (L) buccal and (M) lingual views;(NeP) Right mandibular fragment with M1eM3 (GCP-CV4060), in (N) buccal, (O) lingual and (P) occlusal views; (QeS) Left M2 (GCP-CV4062), in (Q) buccal, (R) lingual and (S)occlusal views; (TeV) Left M3 (GCP-CV4063), in (T) buccal, (U) lingual and (V) occlusal views; (WeY) Left M3 (Q03-Ec-1), in (W) buccal, (X) lingual and (Y) occlusal views; (Z)Mandibular fragment without teeth (Q03-E-1), in (Z) lower and (Z0) upper view.
D.M. Alba et al. / Journal of Human Evolution 61 (2011) 703e718706
Author's personal copy
Preservation and description
Mandible Besides isolated teeth, there are three availablemandibular specimens. Q03-E-1 (Fig. 2ZeZ0) is an edentulousmandibular fragment preserving the symphysis, the basal portionof the alveoli of the four incisors and the two canines, anda small basal portion of the left corpus. GCP-CV4060 (Fig. 2NeP)is a right mandibular fragment with the molar series. Dentalwear on this specimen ranges from almost absent in the M3 tomoderate in M1, and although there is some damage on thecrowns of M2 and M3, it does not preclude taking reliablemeasurements. Finally, Q03-E-4 (Fig. 2AeM) includes severalmandibular fragments and isolated teeth from a single individual,which taken together preserve the left lower canine to P3 seriesand the right lower canine to M3 series. This specimencorresponds to a juvenile individual, in which the third molar isstill inside the crypt. The third premolars were beginning toemerge but had not !nished the formation of the mesial root. Thecanines, albeit already emerging, had only completed theformation of the crown and the basal-most portion of the root.This individual displays only a slight degree of wear on the M2and a moderate degree of wear on the P4 and M1, although it
should be taken into account that part of the M1 and most of theM2 occlusal surfaces are abraded.
The fragmentary nature of all of the available mandibularspecimens does not provide much morphologic informationbesides that of the teeth. The single exception is Q03-E-1(Fig. 2ZeZ0), which displays a deep symphysis (>32 mm) witha very marked and thick inferior torus on its lowermost portion.When all of the available specimens are taken into account, all ofthe permanent teeth except for the incisors are preserved, whereasno lower deciduous teeth are available.Lower canines Mandible Q03-E-4 preserves the tooth germs ofboth the right (Q03-E-4c; Fig. 2HeK) and left (Q03-E-4b;Fig. 2DeG) canines, which include the unworn crowns but onlythe most basal portion of the roots. Crown height is 17.5 mm onthe left side and around 17.1 mm on the right side. The crown isbuccolingually compressed with an elliptical basal outline and nocingulids. In occlusal view, the crown walls are convex except bythe "atter lingual wall, where there is a moderately developedsulcus that progressively disappears without reaching the crownapex.Lower premolars Only two P3 (Q03-E-4d right, and Q03-E-4b left;Fig. 2DeG and LeM, respectively) and a single right P4 (Q03-E-4a;
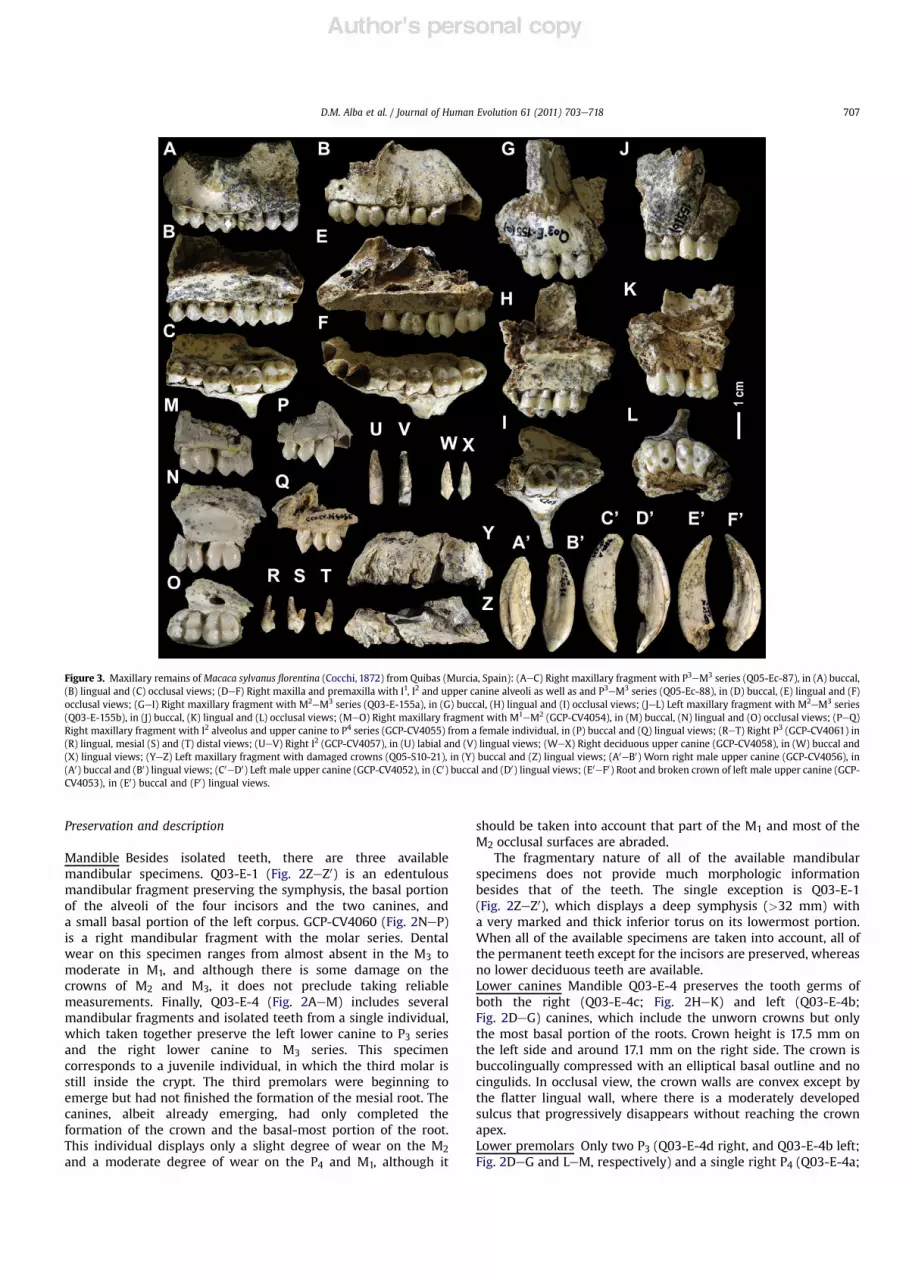
Figure 3. Maxillary remains of Macaca sylvanus !orentina (Cocchi, 1872) from Quibas (Murcia, Spain): (AeC) Right maxillary fragment with P3eM3 series (Q05-Ec-87), in (A) buccal,(B) lingual and (C) occlusal views; (DeF) Right maxilla and premaxilla with I1, I2 and upper canine alveoli as well as and P3eM3 series (Q05-Ec-88), in (D) buccal, (E) lingual and (F)occlusal views; (GeI) Right maxillary fragment with M2eM3 series (Q03-E-155a), in (G) buccal, (H) lingual and (I) occlusal views; (JeL) Left maxillary fragment with M2eM3 series(Q03-E-155b), in (J) buccal, (K) lingual and (L) occlusal views; (MeO) Right maxillary fragment with M1eM2 (GCP-CV4054), in (M) buccal, (N) lingual and (O) occlusal views; (PeQ)Right maxillary fragment with I2 alveolus and upper canine to P4 series (GCP-CV4055) from a female individual, in (P) buccal and (Q) lingual views; (ReT) Right P3 (GCP-CV4061) in(R) lingual, mesial (S) and (T) distal views; (UeV) Right I2 (GCP-CV4057), in (U) labial and (V) lingual views; (WeX) Right deciduous upper canine (GCP-CV4058), in (W) buccal and(X) lingual views; (YeZ) Left maxillary fragment with damaged crowns (Q05-S10-21), in (Y) buccal and (Z) lingual views; (A0eB0) Worn right male upper canine (GCP-CV4056), in(A0) buccal and (B0) lingual views; (C0eD0) Left male upper canine (GCP-CV4052), in (C0) buccal and (D0) lingual views; (E0eF0) Root and broken crown of left male upper canine (GCP-CV4053), in (E0) buccal and (F0) lingual views.
D.M. Alba et al. / Journal of Human Evolution 61 (2011) 703e718 707

Author's personal copy
Fig. 2AeC), all from the same individual, are available. The thirdpremolars correspond to tooth germs that display intact crowns(which were inside the crypt, just beginning to emerge, as shownby Q03-E-4b) and roots (with the mesial one still open). The P3 isa unicuspid and markedly sectorial tooth, with a gracile and shortdistal root, and a much stouter single mesial root. On themesiobuccal aspect of the mesial root, there is a considerableextension of the enamel, which nearly attains the apex of the root(which is not yet closed). In occlusal view, the crown displaysa suboval contour, with its main axis somewhat exorotated. Thebuccal wall of the crown is markedly convex, whereas the lingualwall is "at to very slightly concave. The single main cuspid isquite centrally situated. There is a sharp and long mesial cristidthat descends from the crown axis by slightly curving inmesiolingual direction until reaching the mesial end, close to thecrown base. Two similarly sharp but shorter (distal anddistolingual) cristids steeply descend from close to the crownapex until about mid-crown height. The distolingual cristid endsin a secondary cuspulid that is situated on the distolingual cornerof the crown, whereas the distal cristid merges with the !nedistal marginal ridge, thus completely enclosing a shallow andrestricted distal fovea, which is somewhat longer than wide.There is a subtle lingual cingulid subdivided into two distinct(mesial and distal) portions.
Themorphology of the P4 can be only ascertained on the basis ofQ03-E-4a (Fig. 2AeC). It is a bicuspid tooth that, in occlusal view,displays a suboval contour (slightlywider on the distalmoiety of thecrown) and is slightly exorotated with respect to the main mesio-distal axis of themolar series. The twomain cuspids, being subequalin size (the protoconid being only slightly larger than the meta-conid), are situated on the mesial moiety of the crown and con-nected by a transverse cristid. The short preprotocristid andpremetacristid merge with the mesial marginal ridge, thus delim-iting a restricted mesial fovea, which is longer than wide. Thepostprotocristid and postmetacristid are longer than their corre-sponding mesial cristids, thus delimiting an extensive and deepdistal fovea, which is subquadrangular in shape. These distal cristidsend at two very slight thickenings on the distobuccal and
distolingual corners of the crown,where the subtle andcurveddistalmarginal ridge that encloses the distal fovea further converges.There are no cingulids.Lower molars The morphology of the lower molars can bedescribed on the basis of two M1eM3 series, GCP-CV4060(Fig. 2NeP) and Q03-E4a (Fig. 2AeC)dalthough in the latter, M3morphology can be only partially ascertained, this tooth being stillinside the cryptd, as well as several isolated molars: a single M2,GCP-CV4062 (Fig. 2QeS), and two M3, GCP-CV4063 (Fig. 2TeV)and Q03-Ec-1 (Fig. 2WeY). On the basis of size and contact facetcongruence, GCP-CV4062 and 4063 might belong to the sameadult individual. This would also be consistent with GCP-CV4062displaying a degree of wear slightly more advanced than GCP-CV4063. Q03-Ec-1, in turn, does not completely preserve thecrown, but as far as it can be ascertained, it was not completelyformed, which together with the lack of tooth wear suggests thatthis tooth is a germ that had not yet erupted or was just beginningto erupt. In all the worn specimens, dental wear is moreaccentuated on the buccal than on the lingual portion of the crown.
There is a marked size increase from M1 to M3, with the formertooth being both shorter and narrower than the remaining molars,and theM3 being longer than theM2. Themolars display the typicalpapionin morphology, with a bilophodont occlusal pattern. The M1and M2 are subquadrangular in shape, being clearly longer than
Figure 4. Postcranial remains of Macaca sylvanus !orentina (Cocchi, 1872) from Quibas(Murcia, Spain): (AeB) Distal fragment of left humerus (Q05-Ec-89), in (A) posteriorand (B) anterior views; (CeD) Distal fragment of radius (Q04-Ec-90), in (C) distal and(D) lateral views; (EeF) Proximal phalanx (Q04-Ei-C4-16), in (E) ventral and (F) dorsalviews; (GeJ) Partial right humeral head (Q08-E-165), in anterior (G), medial (H),posterior (I) and proximal (J) views.
Table 2Dental measurements (in mm) of Macaca sylvanus !orentina (Cocchi, 1872) fromQuibas (Murcia, Spain).
Record No. Site Teeth MD BL(m) BLd BLI (%)
GCP-CV4063 Quibas M3 L 12.1 8.8 7.6 72.7GCP-CV4057 Quibas I2 R 3.9 5.3GCP-CV4056 Quibas Upper canine R >12.0 >7.8GCP-CV4058 Quibas D upper canine R 5.2 3.3 63.5GCP-CV4061 Quibas P3 R 5.2 5.9 113.5GCP-CV4062 Quibas M2 L 10.2 8.5 7.6 83.3GCP-CV4060 Quibas M1 R 8.1 5.8 5.8 71.6GCP-CV4060 Quibas M2 R 9.4 (7.6) 7.2 80.9GCP-CV4060 Quibas M3 R (12.5) 7.8 6.7 62.4GCP-CV4052 Quibas Upper canine M L 11.7 8.1 69.2GCP-CV4053 Quibas Upper canine M L (12.5)GCP-CV4055 Quibas Upper canine F R 6.9 5.2 75.4GCP-CV4055 Quibas P3 F R 5.5 6.3 114.5GCP-CV4055 Quibas P4 F R 5.3 6.8 128.3GCP-CV4054 Quibas M1 R 8.9 8.5 8.3 95.5GCP-CV4054 Quibas M2 R 9.1 8.9 7.7 97.8Q03-E-4a Quibas P4 R 6.3 4.9 77.8Q03-E-4a Quibas M1 R 8.5 6.5 6.3 76.5Q03-E-4a Quibas M2 R 10 8.2 6.8 82.0Q03-E-4b Quibas Lower canine L 10.4Q03-E-4c Quibas Lower canine R 10.3 5.4 52.4Q03-E-4d Quibas P3 R 6.9 4.6 66.7Q03-Ec-1 Quibas M3 L 12 7.5 6.6 62.5Q03-E-155a Quibas M2 R 9.6 9.3 8.3 96.9Q03-E-155a Quibas M3 R 9.8 9.4 8.6 95.9Q03-E-155b Quibas M2 L (9.8) 9.6 8.5 98.0Q03-E-155b Quibas M3 L 10.6 9.9 8.9 93.4Q05-Ec-87 Quibas P3 R 5.2 6.3 121.2Q05-Ec-87 Quibas P4 R 5.1 6.7 131.4Q05-Ec-87 Quibas M1 R 8.4 8 95.2Q05-Ec-87 Quibas M2 R 9.5 (9.1) 8.5 95.8Q05-Ec-87 Quibas M3 R 9.9 9.3 8.2 93.9Q05-Ec-88 Quibas P3 F L 5.6 6.9 123.2Q05-Ec-88 Quibas P4 F L 5 7.2 144.0Q05-Ec-88 Quibas M1 F L 8 7.9 7.1 98.8Q05-Ec-88 Quibas M2 F L 9.2 9.5 8.6 103.3Q05-Ec-88 Quibas M3 F L 9.3 9.1 7.6 97.8
Abbreviations: MD!mesiodistal length; BL! buccolingual breadth; D! deciduous;F ! female; M ! male; m ! mesial; d ! distal; L ! left; R ! right; BLI ! breadth/length index (%).Notes: Parentheses indicate that the measurement cannot be reliably taken due tosome damage.
D.M. Alba et al. / Journal of Human Evolution 61 (2011) 703e718708

Author's personal copy
wide, and display a fused single vertical and stout root under eachlobe. These molars display two distinct lobes, separated byamarked cleft (with no ectostylid) on the buccal wall, whereas onlya short vertical groove can be found on the lingual side. Accord-ingly, in occlusal view the molars display a rather straight pro!le inall sides except the buccal one, which is clearly biconvex. In the M1,the two lobes are subequal in width, whereas in the M2, the distallobe is somewhat narrower than the mesial one. The M3 is moreelongated than the preceding molars, and displays a more ovoidocclusal contour. This is due to a more marked distal tapering ascompared with the M2, as well as to the presence of a distinct andwell-developed third lobe. The latter is separated from the centrallobe by a shallower and narrower (more groove-like) buccal cleftthan the one separating the mesial and central lobes, and is alsosomewhat buccally twisted with respect to the main axis of thecrown. In the M3, the roots of the mesial lobe are fused like in thepreceding molars, whereas the roots of the central lobe are distallyinclined toward the root of the third lobe, to which they are fused.The M1 and M2 display four cuspids that are similar in size,although the trigonid is slightly higher than the talonid, and themetaconid is generally somewhat more extensive than theremaining cuspids. Unlike the preceding molars, the M3 furtherdisplays a well-developed hypoconulid, which is quite mediallysituated on the distobuccal corner of the crown and less protrudingthan the remaining cusps. All of the available M3s except for Q03-Ec-1 also display a much smaller, albeit distinct, tuberculum sex-tum, which is situated on the distolingual corner of the crown, andseparated from the concomitant cuspids by subtle and shortvertical grooves on the lingual wall of the crown. In all of the lowermolars, the buccal cuspids are slightly less mesially situated than
the lingual ones, and also less peripherally positioned, thusresulting in some degree of buccal "are, which increases fromM1 toM3. The shallow and restricted distal and mesial foveae are clearlywider than long andmainly positioned toward the buccal moiety ofthe crown (especially the mesial ones), except by the distal fovea ofthe M3, which is deeper, more extensive and further displaysa subtriangular shape. In all instances, the central fovea (talonidbasin) is much deeper and more extensive, further displayinga subquadrangular (in M1 and M2) to subrectangular shape (in M3,where it tends to be somewhat longer than wide). Particularly inthe M2 and M3, the short preprotocristid ends in an enamel thick-ening that can be interpreted as a vestigial paraconulid, which isseparated from the protoconid by a subtle vertical groove on themesiobuccal aspect of the crown wall. Together with the mesialmarginal ridge, this secondary cuspulid constitutes a ledge-likeprojection of the crown toward a concomitant distal extension ofthe crown of the preceding molar, thus constituting a continuous,basin-like occlusal structure. There are no appreciable cingulids.Several specimens show some development of secondary enamelfolds, grooves and tubercle-like thickenings on the distal fovea andmarginal ridge, particularly of the M3.Maxilla Besides several isolated upper teeth (GCP-CV4052, 4053,4056, 4057 and 4058; Fig. 3C0eD0, E0eF0, A0eB0, UeV and WeX,respectively), there are several maxillary specimens with upperteeth in the sample from Quibas, representing at least fourdifferent individuals. GCP-CV4054 (Fig. 3MeO) is a rightmaxillary fragment that preserves the M1eM2 series and part ofthe palate, whereas GCP-CV4055 (Fig. 3PeQ) is a right maxillaryfragment that preserves the external portion of the I1 alveolus,the complete I2 alveolus, and the dental series from the upper
Table 3Descriptive statistics of dental measurements of cheek teeth of Macaca sylvanus !orentina (Cocchi, 1872) from Quibas (Murcia, Spain), compared to M. s. !orentina from Italy(Upper and Lower Valdarno, as well as Mugello), and extant M. s. sylvanus.
Tooth Mesiodistal length (in mm) Buccolingual breadth (in mm) Breadth/length index (%)
N Mean SD 95% CI Range N Mean SD 95% CI Range N Mean SD 95% CI Range
Macaca sylvanus !orentina from QuibasP3 1 6.90 1 4.60 1 66.70P4 1 6.30 1 4.90 1 77.80M1 2 8.30 0.28 5.76 10.84 8.1 8.5 2 6.15 0.49 1.70 10.60 5.8 6.5 2 74.05 3.46 42.92 105.18 71.6 76.5M2 3 9.87 0.42 8.83 10.90 9.4 10.2 3 8.10 0.46 6.96 9.24 7.6 8.5 3 82.07 1.20 79.08 85.05 80.9 83.3M3 3 12.20 0.26 11.54 12.86 12.0 12.5 3 8.03 0.68 6.34 9.72 7.5 8.8 3 65.87 5.92 51.17 80.57 62.4 72.7P3 4 5.38 0.21 5.05 5.70 5.2 5.6 4 6.35 0.41 5.69 7.01 5.9 6.9 4 118.10 4.82 110.43 125.77 113.5 123.2P4 3 5.13 0.15 4.75 5.51 5.0 5.3 3 6.90 0.26 6.24 7.56 6.7 7.2 3 134.57 8.32 113.91 155.22 128.3 144.0M1 3 8.43 0.45 7.31 9.55 8.0 8.9 3 8.13 0.32 7.33 8.93 7.9 8.5 3 96.50 2.00 91.54 101.46 95.2 98.8M2 5 9.44 0.29 9.08 9.80 9.1 9.8 5 9.28 0.29 8.92 9.64 8.9 9.6 5 98.36 2.90 94.77 101.96 95.8 103.3M3 4 9.90 0.54 9.05 10.75 9.3 10.6 4 9.43 0.34 8.88 9.97 9.1 9.9 4 95.25 2.01 92.05 98.45 93.4 97.8
Macaca sylvanus !orentina from Italy (Upper and Lower Valdarno, and Mugello)a
P3 10 8.39 0.68 7.90 8.88 7.7 9.9 10 5.57 0.65 5.11 6.03 4.9 6.6 10 66.42 6.17 62.01 70.84 61.0 76.8P4 14 6.29 0.36 6.08 6.49 5.7 6.9 13 5.35 0.40 5.10 5.59 4.8 6.0 13 85.08 4.15 82.57 87.59 78.1 93.1M1 13 8.19 0.36 7.97 8.41 7.8 9.0 13 6.42 0.30 6.23 6.60 5.9 6.9 13 78.36 3.26 76.39 80.33 71.8 84.0M2 14 9.55 0.35 9.35 9.75 9.1 10.2 15 7.91 0.50 7.63 8.19 7.0 8.7 14 82.44 4.56 79.81 85.07 75.3 90.6M3 12 13.11 0.93 12.52 13.70 11.7 15.2 12 8.24 0.35 8.02 8.46 7.4 8.7 12 63.06 3.61 60.76 65.35 55.9 68.3P4 1 5.10 1 6.90 1 135.29M1 1 8.50 1 8.80 1 103.53M2 1 9.40 1 10.40 1 110.64M3 3 10.63 0.76 8.74 12.53 9.8 11.3 3 9.60 0.35 8.74 10.46 9.4 10.0 3 90.57 6.60 74.16 106.97 83.2 95.9
Extant Macaca sylvanus sylvanusb
P4 33 6.16 0.48 5.99 6.33 5.0 7.2 33 5.27 0.45 5.11 5.44 4.5 6.2 33 85.91 7.71 83.18 88.65 74.6 103.3M1 32 7.72 0.36 7.59 7.85 7.0 8.5 32 6.04 0.35 5.91 6.17 5.3 7.0 32 78.25 3.66 76.93 79.57 70.6 86.4M2 35 9.53 0.65 9.31 9.76 8.4 11.1 35 7.79 0.66 7.57 8.02 6.3 9.3 35 81.82 5.78 79.84 83.81 72.2 101.1M3 26 11.76 1.29 11.24 12.28 7.6 14.6 26 8.16 0.62 7.91 8.41 7.1 9.5 26 70.24 10.22 66.11 74.36 59.7 113.2P4 37 5.61 0.32 5.51 5.72 5.0 6.1 37 6.86 0.60 6.66 7.06 5.4 7.9 37 119.69 20.60 112.82 126.56 11.0 145.0M1 35 7.81 0.41 7.67 7.96 7.1 8.8 35 7.09 0.41 6.95 7.23 6.0 7.8 35 90.88 5.39 89.03 92.73 78.8 102.8M2 40 9.50 0.71 9.27 9.72 8.0 11.4 40 8.79 0.69 8.56 9.01 7.0 10.1 40 92.61 5.02 91.00 94.21 78.6 100.0M3 32 9.29 0.80 9.01 9.58 7.6 11.0 32 8.61 0.66 8.37 8.84 7.1 10.0 32 92.85 6.33 90.57 95.14 81.1 109.8
Abbreviations: N ! sample size; SD ! standard deviation; CI ! con!dence interval.a No P3 available for M. s. !orentina from Italian localities.b P3 and P3 measurements not available for extant M. s. sylvanus.
D.M. Alba et al. / Journal of Human Evolution 61 (2011) 703e718 709

Author's personal copy
canine to the P4. Both specimens might belong to a singleindividual, although this cannot be securely ascertained. GCP-CV4055, in any case, can be attributed to a female individual onthe basis of canine size and shape. Q03-E-155 includes severalfragments that correspond to the same individual. The twolargest fragments are Q03-E-155a (Fig. 3GeI) and Q03-E-155b(Fig. 3JeL), which respectively correspond to the right and leftmaxillae. Q03-E-155a, in particular, preserves a distal crownfragment of M1 and the M2eM3 series, as well as part of thepalate "oor and the palatine suture, and the basal-most portionof the zygomatic. Q03-E-155b, in its turn, similarly preserves theM2eM3 series (although the M2 is slightly damaged at its mesialportion), as well as a smaller portion of the zygomatic root. Theteeth of this individual display a relatively advanced degree ofwear, with dentine exposure on the four main cusps of themolars. Another individual, Q05-Ec-88 (Fig. 3DeF), preserves theright premaxilla and maxilla, including most of the lateral rim ofthe nasal aperture, the alveoli of the incisors and canine, and theP3eM3 series. This specimen shows only a moderate degree ofwear on the premolars and M1, and a slight degree on the M2;the M3 is unworn, had not yet completed eruption, and its lingualroot is neither closed, thus indicating that this specimencorresponds to a subadult or very young adult individual. Thesmall size of the upper canine alveolus indicates that thespecimen corresponds to a female individual. Finally, Q05-Ec-87(Fig. 3AeC) is a right maxillary fragment, preserving the lowerportion of the zygomatic root, the P3eM3 series (although thecrowns of M1 and M2 are slightly damaged on the buccal side),and a portion of the "oor of the palate (with the palatine suturepreserved from P3 to the mesial level of M2). This specimenshows a moderate degree of wear on the premolars and M1, anda slight degree of wear on the M2, while the M3 is unworn. Thereis an additional maxillary fragment, Q05-S10-21 (Fig. 3YeZ),which is very damaged and preserves no tooth crown (butmerely the alveoli and the partial roots of the incisors, canine andcheek teeth), and which might represent another individual orcorrespond to the left side of the individual represented by eitherQ05-Ec-87 or Q05-Ec-88.
Most of the above-mentioned specimens provide a good sourceof metrical and morphologic information regarding the upperdentition (particularly, the postcanine one), but provide onlya limited amount of information on cranial morphology. Themaxillary sinus is large, as shown by Q05-Ec-87 and, to a lesserextent, Q03-E-155 (the latter specimen showing that it extendsuntil about themesial level of theM3). As shown by Q03-E-155 and,to a lesser extent, Q05-Ec-88 (Fig. 3DeF) and Q03-Ec-87(Fig. 3AeC), the zygomatic must have been quite "at and anteriorlydirected, with the anterior zygomatic root originating from thelevel of distal M2 to M2eM3. Q05-Ec-88 (Fig. 3DeF) further showsa short and thick premaxilla that tapers somewhat distally and isslightly inclined with respect to the palate (with whom it musthave delimited a wide palatine fenestra), and that maximum nasalwidth was attained at about mid-height of the nasal aperture.Upper deciduous teeth There is a single deciduous toothwithin thesample, GCP-CV4058 (Fig. 3WeX), which corresponds toa deciduous right canine. The crown is quite low (crown height>6.0 mm) as compared with the length of the root (root height9.8 mm, length 5.2 mm, width 3.3 mm). It displays a slightlyasymmetric elliptical occlusal pro!le, with the apex of the singlecusp being somewhat mesially positioned. The buccal sidedisplays a convex occlusal contour, whereas the lingual side isalso convex at the base, otherwise displaying a biconcave contourdue to the presence of a broad but ill-de!ned crista that descendsfrom the apex of the crown in lingual direction. There is alsoa subtle lingual cingulum along the lingual side of the crown, as
well as an elongated wear facet situated on the distolingualaspect of the crown. In buccal or lingual view, the crown displaysa rather straight mesial pro!le and a slightly concave distal one,being de!ned by the two !ne cristae that descend from thecrown apex in mesial and distal directions, the latter beingsomewhat longer than the mesial one.Upper incisors The only available upper incisor is the I2 GCP-CV4057 (Fig. 3UeV), although two I2 alveoli are preserved in themaxillary fragments Q-05-Ec-88 (Fig. 3DeF) and GCP-CV4055(Fig. 3PeQ). The available incisor, which preserves the crown andmost of the root, is a relatively high-crowned (labial height9.5 mm), moderately spatulate and non-waisted tooth. The mesialand distal pro!les of the crown are approximately parallel to oneanother, and the crown is labiolingually broadest mesially. Theroot is slightly mesiodistally compressed and becomes narrowerfrom base to tip. It is slightly tilted mesially, and more markedlycurved in lingual direction. On the mesial side, thecementoenamel junction is V-shaped, and there is a small contactfacet with the I1 on the enamel, situated close to the occlusalsurface. There is also a longer apical wear facet exposing thedentine all along the occlusal surface, which further extends alongthe distolingual portion of the crown, indicating wear against theupper canine. Besides these facets, the lingual aspect of the crownis almost entirely devoid of enamel (except for the basal-mostportion), so that it cannot be ascertained whether a lingual sulcuswas originally present. No basal enamel thickening is present onthe lingual side.Upper canines The three available isolated canines (GCP-CV4056,GCP-CV4052 and GCP-CV4053; Fig. 3A0eB0, C0eD0 and E0eF0,respectively) can be attributed to male individuals on the basis ofboth size and shape. In these regards, these teeth considerablydiffer from the much smaller and less dagger-like female caninepreserved in the maxillary specimen GCP-CV4055 (Fig. 3PeQ),which is much more similar in morphology to (albeit larger than)the deciduous specimen GCP-CV4058 (Fig. 3WeX). As such, it canbe concluded that the macaque from Quibas displayed a highdegree of canine sexual dimorphism, as in extant macaques.
The morphology of the male canine crown can be only ascer-tained in GCP-CV4052 (Fig. 3C0eD0), in spite of the relativelyadvanced degree of wear (preserved buccal height 18.9 mm),because in GCP-CV4056 (Fig. 3A0eB0) the crown is almostcompletely worn down (thereby exposing the pulp cavity) and inGCP-CV4053 (Fig. 3E0eF0) the crown is broken away. The root wasprobably higher than the crown (root height 35.8 mm in GCP-CV4053, which preserves the complete root, and >26.1 mm inGCP-CV4052, where it is incomplete). Overall, the male canine isa dagger-like, buccolingually compressed tooth that displaysa subtriangular occlusal outline, being broader on the mesial thanon the distal moiety of the crown. The maximum diameters of thecrown and the root are attained at about the cementoenameljunction, both progressively narrowing from base to apex. In buccalor lingual views, both the root and the crown display a convexmesial pro!le and a concave distal pro!le, although the degree ofcurvature is more accentuated on the mesial side. In occlusal view,the buccal contour of the male canine is slightly biconvex, due tothe presence of an ill-de!ned and shallow groove on the buccalcrown wall from base to apex. On the lingual side, the male caninedisplays a strongly biconvex contour. On its mesiolingual aspect,a deep and well-de!ned sulcus extends along the root, becomingnarrower and deeper toward the crown, whereas on the dis-tolingual aspect of the root there is a much more diffuse andshallower depression. On the buccal side, the cementoenameljunction is V-shaped. Regarding dental wear, in GCP-CV4052(Fig. 3C0eD0), the crown is partially broken on its distobuccalapical aspect, and further displays two large wear facets:
D.M. Alba et al. / Journal of Human Evolution 61 (2011) 703e718710

Author's personal copy
a mesiolingual facet that occupies about two-thirds of the crownheight, due to shearing against the lower canine, and another facetthat occupies most of the distolingual aspect of the crown (from theapicalmost preserved portion of the crown up to the cementoe-namel junction), due to shearing against the P3. Both wear facetsconverge on the lingual side of the crown, giving rise to a ridge-likestructure that occupies the apical half of the crown, whereas thedistolingual wear facet further generates a sharp distobuccalcutting edge that runs from base to tip. On the much worn spec-imen GCP-CV4056 (Fig. 3A0eB0), there is a small mesiolingual facetand a much more extensive, "at but obliquely inclined triangularfacet that extends over the cementoenamel junction and furtherexposes the pulp cavity.
As stated above, the female canine is much lower-crowned thanthe male canine (crown height >7.8 mm), although it clearlyprotrudes from the occlusal level de!ned by the premolars. Inocclusal view, it displays a buccolingually compressed ellipticalpro!le, with a convex buccal contour and a rather biconcave lingualone. The latter is divided into two distinct portions as a result ofa curved crest that runs from the apex toward the base of the crownin a somewhat mesiolingual direction. The distolingual aspect ofthe crown is slightly concave, whereas the mesiolingual onedisplays a relatively shallow and ill-de!ned groove, which origi-nates close to the crown apex and progressively widens slightlytoward the base. Two additional well-de!ned and sharper crestsrun in mesial and distal directions from crown apex to base. Thedistal crest is somewhat worn as a result of a triangular wear facetsituated on the distolingual portion of the crown.Upper premolars The female P3 from themaxillary fragments GCP-CV4055 (Fig. 3PeQ) and Q05-Ec-88 (Fig. 3DeF) are comparable insize and shape to the unsexed isolated specimen GCP-CV4061(Fig. 3ReT) and to that from the unsexed maxillary fragmentQ05-Ec-87 (Fig. 3AeC). The P4 is only known from the samemaxillary specimens (no isolated P4 is known). The upperpremolars have three roots, with the lingual root being stouterand longer than the distobuccal one, and the mesiobuccal rootbeing also apparently stouter than the distobuccal one (althoughthis cannot be completely ascertained because the former isbroken away in the only isolated available specimen). There issome extension of the enamel onto the basal-most aspects of themesiobuccal and lingual roots. All available specimens displaya relatively advanced degree of wear (with dentine exposure inboth the lingual and buccal cusps) except for GCP-CV4061(Fig. 3ReT), which shows only a moderate degree of wear (moreappreciable on its distal moiety, from the apices of the two maincusps until the distal marginal ridge).
Both the P3 and P4 are bicuspid and display a basally in"atedcrown (particularly the P3). Both premolars are wider than long.However, whereas the P4 displays an elliptical to suboval occlusaloutline and is mesiodistally longer along the crown midline, the P3
displays a subtriangular outline, being clearly longer on the buccalthan at the lingual moiety of the crown, and further displayinga style-like mesial extension on the mesiobuccal corner of thecrown. The buccal and especially the lingual basal contours areconvex, whereas the distal one is only moderately so, and themesial contour is rather straight. The paracone is more protrudingand slightly more extensive than the protocone, which in its turn isslightly more peripherally and mesially situated than the paracone.These two main cusps are united by a sharp but relatively !netransverse crista, which separates the subtriangular mesial foveafrom the longer and more extensive, but equally deep, distal fovea(trigon basin). The mesial fovea is further enclosed by the prepar-acrista and preprotocrista, which converge toward the biconvexmesial marginal ridge at about crown midline. The short, relativelysharp and straight postparacrista, and themore curved, blunter and
somewhat longer postprotocrista run in distal direction from thetwomain cusps until merging with the distal marginal ridge, whichis considerably worn in all the available specimens.Upper molars All of the available maxillary fragments preserve atleast two upper molars, except for GCP-CV4055 (Fig. 3PeQ). Thewhole upper molar series is preserved in both Q05-Ec-87(Fig. 3AeC) and Q05-Ec-88 (Fig. 3DeF), whereas GCP-CV4054(Fig. 3MeO) preserves the M1eM2 series, and Q03-E-155a(Fig. 3GeI) and Q03-E-155b (Fig. 3JeL) preserve the M2eM3 series.The occlusal morphology of the !rst and second upper molars canbe best appreciated in GCP-CV4054 (Fig. 3MeO) due to the veryslight degree of wear, whereas that the M3 can be well ascertainedin both Q05-Ec-87 (Fig. 3AeC) and Q05-Ec-88 (Fig. 3DeF).
All of these molars display a bilophodont occlusal pattern andare slightly longer than wide. The M1 is morphologically similar to,but slightly smaller than, the M2, whereas the M3 differs from thepreceding molars by being somewhat larger and by displayinga suboval instead of subrectangular occlusal pro!le (due to themarkedly convex distal contour of the M3, which tapers distally). Inall of the upper molars, the crown is slightly longer on the buccalthan on the lingual side. In occlusal view, the upper molars alldisplay markedly biconvex lingual and buccal pro!les, with themesial lobe being wider than the distal lobe (particularly in the M2
and M3), both being separated from one another by a relativelywell-developed lingual cleft (although with no interconule) anda shallower buccal cleft. The four main cusps are not particularlybuccolingually compressed. The buccal cusps (paracone and met-acone) are only slightly more mesially positioned than the corre-sponding lingual cups (protocone and hypocone). The paracone isslightly larger than the remaining cusps. The buccal cusps are moreprotruding and alsomore peripheral than the buccal ones (less so inthe M3), so that the foveae (particularly the mesial and the centralones) are somewhat displaced toward (although not restricted to)the buccal moiety of the crown. Due to the distal tapering of theM3,the distal cusps of this molar are closer to each other than those ofthe preceding molars.
The relatively deep but short mesial fovea, de!ned by the shortpreprotocrista and preparacrista as well as the marked marginalridge, is shorter than wide and somewhat crescent-like, partiallycurving along the mesial aspect of the paracone base. The centralfovea (trigon basin) is deeper and more extensive than the mesialfovea. The former is mesially and distally delimited by the twoconspicuous transverse lophs, although the latter are partiallyinterrupted at the crown midline by two corresponding subtle dis-tolingual grooves. The distal fovea is smaller than the two precedingones, being delimited by the short postmetacrista and post-hypocrista, which merge with the relatively sharp and somewhatcurveddistalmarginal ridge. Thedistal fovea is particularly restrictedin the M1 and M2, and less so in the M3, although in all molars it iswider than longer as well as shallower than the trigon basin.
No cingula are present, except for a small, verticalized andgroove-like style on the mesiolingual corner of the crown of allmolars. It is adjacent to a small, tubercle-like thickening that ispresent at the end of the preparacrista. This basic thickening,together with the very thick marginal ridge, constitutes a smallledge-like extension of the enamel toward the preceding tooth. Thedistal marginal ridge is thick in the M1 andM2, whereas in the M3 itshows some development of secondary grooves and tubercles. Inboth Q05-Ec-88 and Q05-Ec-87, a secondary cuspule (distoconule)that occupies most of the distal ridge and part of the distal foveacan be clearly appreciated, thanks to the slight dental wear. Thiscuspule is most well-developed in Q05-Ec-87, where it is prolongedby vertical grooves along the distal crown wall, but to a lesserdegree it can be also discerned in Q03-E-155b, whereas in Q03-E-155a it was probably originally present but it has been worn away.
D.M. Alba et al. / Journal of Human Evolution 61 (2011) 703e718 711

Author's personal copy
Postcranial Only a few postcranial bones of Macaca are availablefrom the Quibas sample: a distal humeral fragment (Q04-Ec-89;Fig. 4AeB), a proximal radial fragment (Q04-Ec-90; Fig. 4CeD),a proximal humeral fragment (Q08-E-165; Fig. 4GeJ), anda proximal phalanx (Q04-Ei-C4-16; Fig. 4EeF). These remains arenot signi!cantly different from those of extant M. sylvanus, so nometrical comparisons are provided.
The proximal humeral fragment (Fig. 4GeJ) only partiallypreserves the humeral head, since the articular surface is damagedon its distal and medial portions. The lesser tubercle is furtherdamaged. The greater tubercle protrudes slightly from the head,and the bicipital groove (intertubercular sulcus) is not surroundedby marked crests running from either the lesser or the greatertubercles. The distal humeral fragment (Fig. 4AeB) preserves thewhole distal epiphysis and the distal-most portion of the diaphysis(more completely preserved on the anterior than on the posteriorside). The preserved distal portion of the diaphysis is somewhatanteroposteriorly "attened, being quite "at on the posterior sidebut showing a convex pro!le on the anterior side. There is a markedlateral supracondylar crest and a less de!ned medial supracondylarcrest. Themedial epicondyle is small and not very protruding, beingposteromedially directed (i.e., rather retro"exed), whereas thelateral epicondyle is proximodistally longer but less protrudingthan the medial epicondyle. On the posterior side, the olecranonfossa is wide, deep and subtriangular, with a vertical and ridge-likelateral margin (continued along the distal portion of the epiphysistoward the capitulum), and a more oblique and blunter medialmargin. The portion of the shaft medial to the olecranon fossa is
narrower than the lateral portion, although the latter is veryobliquely inclined. On the anterior side, the radial fossa is quiteindistinct, whereas the coronoid fossa is relatively well delimited,although not very deep. The capitulum is quite globular and ratherdistally protruding, although not very well de!ned. The trochlea isnot spool-shaped and markedly asymmetric, with a veryprotruding and well developed medial trochlear keel, but a very ill-de!ned and almost "at lateral one. The latter is separated from thecapitulum by a relatively wide but very shallow zona conoidea.
The radial fragment (Fig. 4CeD) only preserves the proximalhead and the proximal-most portion of the diaphysis. The latterdisplays a subelliptical to subtriangular cross-section, whereas theproximal articular portion is subcircular and somewhat concave.This articular surface is not fully perpendicular, but slightlyobliquely oriented relative to the transverse plane of the proximalpreserved portion of the diaphysis.
Finally, the proximal phalanx (Fig. 4EeF) is a relatively slenderand only moderately curved, with a proportionately stout basalportion as compared to the diaphysis and the trochlea. Thediaphysis is higher on its proximal portion, markedly "atteningtoward distal. On this basis, and given the slightly convex andsomewhat protruding ventral portion, this specimen might corre-spond to the foot. However, given the morphologic similaritiesbetween manual and pedal phalanges in papionins, as well as theirgreat overlap in morphometric measurements (S. Almécija, pers.com. to DMA), it is not possible to unambiguously assert whether itcorresponds to a pedal or manual phalanx. The ventral aspect of thediaphysis displays faint insertions for the "exors on the middle and
Figure 5. Specimens of Macaca sylvanus !orentina (Cocchi, 1872) from Upper Valdarno, housed at the Museo di Storia Naturale at Firenze (Italy): (AeC) Holotype mandible(IGF10034), in occlusal (A), right buccal (B) and left buccal (C) views; (DeF) Left mandibular fragment with molar series (IGF13084), in lingual (D), buccal (E) and occlusal (F) views.
D.M. Alba et al. / Journal of Human Evolution 61 (2011) 703e718712

Author's personal copy
Figure 6. Bivariate plots showing the proportions (buccolingual breadth vs. mesiodistal length) of individual fourth upper premolar and upper molars of Macaca sylvanus !orentina(Cocchi, 1872) from Quibas (Murcia, Spain), as compared with extant M. s. sylvanus (data taken from Zanaga, 1998), M. s. !orentina from Italy (measured on original specimens orgood quality casts), and M. sylvanus cf. !orentina from Terrassa (data taken from Alba et al., 2008b). The solid line is the least-squares best-!t line for the M. s. sylvanus sample.Abbreviations: BL ! buccolingual breadth (mesial); MD ! mesiodistal length.
Figure 7. Bivariate plots showing the proportions (buccolingual breadth vs. mesiodistal length) of individual fourth lower premolar and lower molars of Macaca sylvanus !orentina(Cocchi, 1872) from Quibas (Murcia, Spain), as compared with extant M. s. sylvanus (data taken from Zanaga, 1998), M. s. !orentina from Italy (measured on original specimens orgood quality casts), M. sylvanus ssp. indet. from Untermassfeld (data taken from Zapfe, 2001), and M. sylvanus cf. !orentina from Terrassa (data taken from Alba et al., 2008b). Thesolid line is the least-squares best-!t line for the M. s. sylvanus sample. Abbreviations: BL ! buccolingual breadth (mesial); MD ! mesiodistal length.
D.M. Alba et al. / Journal of Human Evolution 61 (2011) 703e718 713

Author's personal copy
distal portion of the shaft, which do not reach the trochlear region.The distal trochlea does not protrude laterally beyond the diaph-ysis, and in dorsal/ventral views it shows a biconvex pro!le. Thedistal articular surface is dorsally restricted but it is proximodistallyquite long on the ventral side. With regard to ray assignment, onthe basis of the pronounced basal asymmetry and the moderatetrochlear asymmetry, we favor the hypothesis that this specimencorresponds to the right fourth ray, although alternatively it mightbe interpreted as a left specimen from the second ray. Assuming ourpreferred assignment, the trochlea is only slightly more protrudingon the medial side, further being slightly tilted toward lateral,whereas a second phalanx would display a more marked trochlearasymmetry. Moreover, the basal region of the phalanx is moremarkedly asymmetrical, being much more proximally and laterallyprotruding on the presumably lateral than on the medial side, as itis typical of fourth proximal phalanges, for the insertion of thefourth dorsal interosseous. However, second phalanges also displaya medially protruding basal region for the insertion of the !rstdorsal interosseous, so that the alternative hypothesis cannot bediscarded that the specimen corresponds to a left second proximalphalanx. The ventral tubercles are relatively well developed(particularly the presumably lateral one), being separated from oneanother by a conspicuous depression. In proximal view, the artic-ular surface for the metapodial head displays an elliptical (widerthan high) contour, being laterally well buttressed by bonyprojections on both sides (also more marked on the presumablylateral side). This articular surface is not perpendicular, but obliqueto the main axis of the phalanx. Basally it does not reach the ventraltubercles, whereas dorsally it somewhat extends onto the dorso-proximal aspect of the diaphysis, as in other quadrupedal taxa.
Morphologic and metrical comparisons
The dentognathic material from Quibas reported in this paperdoes not show any substantial morphological distinction ascompared with the material from Upper Valdarno (Italy), fromwhere the holotype (and most of the available material) ofM. s. !orentina comes (compare the mandibular material fromQuibas in Fig. 2 with the material of M. s. !orentina from Italy inFig. 5). Possible comparisons are limited, becausemost of the Italianmaterial comprises mandibles and lower teeth, whereas a greaterproportion of the material from Quibas comprises maxillary spec-imens. In this sense, however, the newly reported material fromQuibas (together with the material from Terrassa recentlydescribed by Alba et al., 2008a) permits us to better characterize theupper dentition of this taxon.
The various subspecies of the Barbary macaque recognized fromthe European Plio-Pleistocene fossil record are far from being wellcharacterized. Their differences with respect to the extant,nominotypical subspecies are subtle and mostly consist in metricaldifferences in size and proportions, with a large degree of overlap.In Figs. 6e10, we report metrical comparisons of the sample fromQuibas with a sample of M. s. !orentina (mainly from Upper Val-darno localities, as well as Mugello and Lower Valdarno: see Cocchi,1872; Ristori, 1890; Szalay and Delson, 1979; Delson, 1980; Rook,1997) and a sample of extant M. s. sylvanus. The remains of M. s.!orentina from Pietra!tta (Gentili et al., 1998) have not beenincluded in the comparative fossil sample because no measure-ments of these specimens have been thus far published. Thematerial of M. sylvanus cf. !orentina from Cal Guardiola and Vall-paradís in Terrassa, Spain (Alba et al., 2008a) and ofM. sylvanus ssp.indet. from Untermassfeld, Germany (Zapfe, 2001) has been alsoincluded in some of the comparisons (Figs. 6 and 7).
Regarding the upper dentition, the sample of M. s. !orentinafrom the Italian sites (Upper and Lower Valdarno, and Mugello) is
very limited, so that comparisons must be cautious. All of the fossilspecimens show a considerable degree of overlap with the extantBarbary macaque with regard to mesiodistal dimensions, althoughthe uppermolars (particularly theM1 andM3) tend to be somewhat
Figure 8. Mean mesiodistal length (in mm) of upper (A) and lower (B) molars inMacaca sylvanus !orentina (Cocchi, 1872) from Quibas (Murcia, Spain), as comparedwith M. sylvanus !orentina from Italy and extant M. s. sylvanus. The discontinuous lineindicates the 95% con!dence interval for the mean of M. s. sylvanus. Measurements forM. s. sylvanus were taken from Zanaga (1998: Figs. 47e49 and 51e53), while those forM. s. !orentina were taken for this study on original specimens or good quality casts.
Figure 9. Mean breadth/length index (in %) of upper (A) and lower molars (B) inMacaca sylvanus !orentina (Cocchi, 1872) from Quibas (Murcia, Spain), as comparedwith M. sylvanus !orentina from Italy and extant M. s. sylvanus. The discontinuous lineindicates the 95% con!dence interval for the mean of M. s. sylvanus. Measurements forM. s. sylvanus were taken from Zanaga (1998: Figs. 47e49 and 51e53), while those forM. s. !orentina were taken for this study on original specimens or good quality casts.
D.M. Alba et al. / Journal of Human Evolution 61 (2011) 703e718714
Author's personal copy
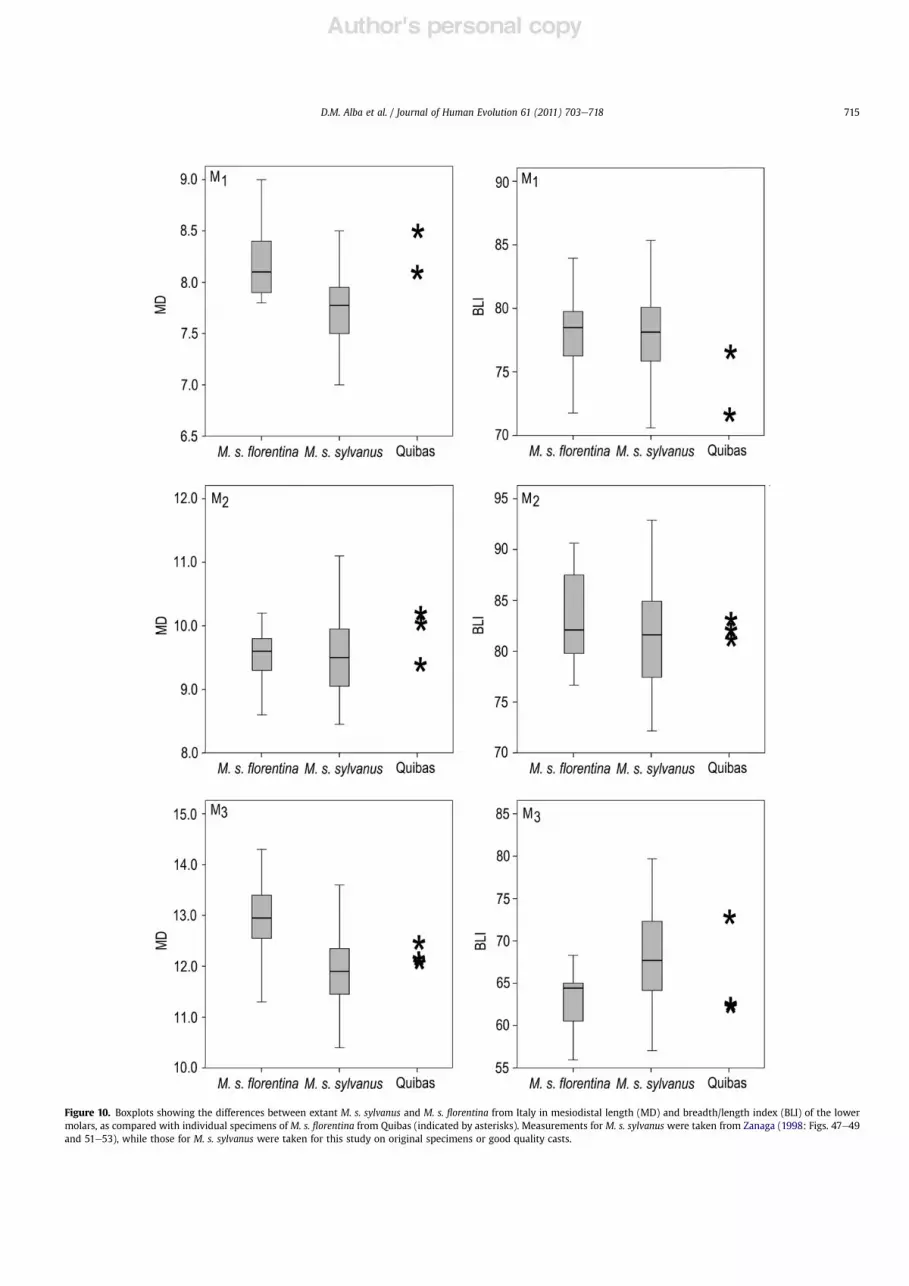
Figure 10. Boxplots showing the differences between extant M. s. sylvanus and M. s. !orentina from Italy in mesiodistal length (MD) and breadth/length index (BLI) of the lowermolars, as compared with individual specimens of M. s. !orentina from Quibas (indicated by asterisks). Measurements for M. s. sylvanus were taken from Zanaga (1998: Figs. 47e49and 51e53), while those for M. s. sylvanus were taken for this study on original specimens or good quality casts.
D.M. Alba et al. / Journal of Human Evolution 61 (2011) 703e718 715

Author's personal copy
longer in the fossils (Figs. 6 and 8A). As a result, in M. s. !orentinafrom both the Italian localities and Quibas, the third upper molartends to be longer the second one, whereas extant M. s. sylvanusshows the opposite condition (Fig. 8A). Finally, the fossil specimensincluded in the study tend to display larger buccolingual widths atequal mesiodistal lengths than the extant Barbary macaque (Fig. 6).This is most clear regarding the !rst and second upper molars,which display breadth/length indices above the 95% con!denceinterval for the mean of M. s. sylvanus (Fig. 9A).
With regard to the lower dentition, given the relatively largersample of M. s. !orentina from the type locality, it is possible toundertake several statistical comparisons. In the case of the lowerdentition, however, the degree of overlap between the fossilspecimens and the extant Barbary macaque is greater than in theupper dentition (Fig. 7). Both the M1 and the M3, however, tend tobe absolutely longer (Fig. 8B) in the fossil taxon, while the M3 tendsto be relatively narrower (Fig. 9B). These differences are con!rmedby ANOVA comparisons between M. s. sylvanus and M. s. !orentina.Thus, differences in lower molar length are signi!cant at p < 0.001in the case of the M1 (F ! 16.229), and at p < 0.01 in the case of theM3 (F ! 9.365) (see also Fig. 10). The M1 ofM. s. !orentina, however,is also absolutely narrower (p< 0.01, F! 10.875), so that differencesin breadth/length index regarding this tooth are not statisticallysigni!cant (F ! 0.000) (see also Fig. 10). On the contrary, there areno differences in absolute width of the M3 between both taxa(F ! 0.186), so that differences in breadth/length index are signi!-cant at p < 0.05 (F ! 4.774) (see also Fig. 10). To sum up, the sampleof M. s. !orentina from Upper and Lower Valdarno and Mugellodiffers from the extant Barbary macaque by displaying longer M1and M3, and a relatively narrower M3. The Spanish material showssome peculiarities, such as displaying relatively narrower M1 thanboth the Italian M. s. !orentina and the extant M. s. sylvanussamples, and M3 somewhat intermediate in length between thetwo samples. The signi!cance of these differences cannot becurrently evaluated, due to the relatively small sample size of lowermolars from Quibas, together with the large degree of overlapbetween both subspecies. Nevertheless, the Quibas material best!ts the condition displayed byM. s. !orentina from the type localityregarding both M1 length and M3 breadth/length index in two outof three individuals.
Discussion and conclusions
Taxonomy
Anatomically, extant M. sylvanus is characterized by an extremereduction of the tail, but this organ was apparently independentlyreduced several times during the evolution of this genus (Fooden,2007). Fooden (2007) has argued that the ancestral macaquestock that dispersed from Africa to Eurasia was long-tailed, andthat, if so, it should not be probably regarded as conspeci!c withextant M. sylvanus. Nevertheless, most of the available material ofEuropean fossil macaques consists of mandibular and maxillaryremains, which are very similar to the extant North Africansubspecies. As a result, the classi!cation of fossil Europeanmacaques as temporal-geographic subspecies of M. sylvanus,following the proposal by Szalay and Delson (1979) and Delson(1980), is currently used by most investigators (e.g., Rook et al.,2001; Jablonski, 2002; Alba et al., 2008a; Jablonski and Frost,2010). However, this is problematic in several regards, because asalready noted by Szalay and Delson (1979: 359), these fossilmacaques are not “clearly or signi!cantly distinguishable from theliving northwest African M. sylvanus”, so that “pending a morecomplete revision, all are referred to subspecies of this taxon,de!ned in part by biochronology”. The distinction of subspecies
requires the correlation of geographic and/or temporal differenceswith morphologic/metrical variation, even if there is a large degreeof overlap between the several subspecies. Szalay and Delson(1979) did not provide a satisfactory operational diagnosis of theseveral extinct subspecies. Thus, although these authors (see alsoDelson, 1980) noted that M. s. priscawas smaller, and M. s. pliocenahad relatively broader molars, compared with M. s. sylvanus,M. s. !orentina was not distinguished from the extant form. Thislack of adequate diagnostic morphologic criteria has complicatedthe subspeci!c attribution of several samples of fossil Europeanmacaques (e.g., Montoya et al., 1999; Zapfe, 2001). Moreover, sizedoes not appear to be a reliable criterion, since there is a largedegree of overlap between the extant Barbary macaque and theseveral extinct subspecies (e.g., Rook et al., 2001). Thus, althoughM. s. prisca has been claimed to be somewhat smaller than theliving form (Szalay and Delson, 1979; Jablonski, 2002; Rosendahlet al., 2011), body mass estimates indicate that among the fossilEuropean forms, only M. majori signi!cantly departs from extantM. sylvanus (by being about 15% smaller; see Delson et al., 2000).
The sample from Quibas described in this paper signi!cantlycontributes to the knowledge of Early Pleistocene macaques fromEurope, particularly by enlarging the available sample of upperteeth, since most of the Italian remains from Upper Valdarnocorrespond tomandibular fragments and lower teeth. Both samplesare morphologically very similar and surely correspond to anextinct form of the Barbarymacaque, but the question remains as towhether both samples can be attributed to the same taxon at thesubspecies level. Metrical comparisons indicate that, althoughthere is a large degree of overlap with extant M. s. sylvanus, thedental sample from Quibas !ts well with the range of variationof the M. s. !orentina sample from Italian localities. Overall,M. s. !orentina differs on average from M. s. sylvanus by displayingabsolutely longer upper molars (especially M1 and M3), relativelywider upper molars (especially M1 andM2), longer M3 as comparedto theM2, absolutely longer M1 andM3, and relatively narrowerM3.The material from Quibas shows some peculiarities as comparedwith the Italianmaterial fromUpper Valdarno andMugello (such asrelatively narrower M1 and less elongated M3), although thesigni!cance of these minor differences is dif!cult to evaluate due torelatively small sample size. To sum up, the Early Pleistocenemacaque material from Quibas best !ts the proportions ofM. s. !orentina from the type locality, and can be therefore formallyattributed to the same extinct subspecies. Thematerial fromQuibasdescribed in this paper, hence, reinforces the view that M. s. !or-entina can be distinguished from extant M. s. sylvanus at thesubspecies rank, and it might contribute in the future to providingan emended diagnosis of the former subspecies. However, this isoutside the scope of this paper, because a revision of the availablematerial from the other two extinct subspecies (M. s. prisca andM. s. pliocena) would be also required to con!rm their taxonomicvalidity on the basis of morphologic or morphometric criteria.
Paleoenvironmental implications
On the basis of the faunal list from Quibas (Montoya et al., 1999,2001; Carlos Calero et al., 2004, 2006a,b), which includes severalgastropod mollusks and arthropods, amphibians, reptiles, birds,and eighteen mammalian species, xerophilous climatic conditionsand a calcareous rocky paleoenvironment, mainly consisting of anopen bushland with some nearby woodlands and wet areas, havebeen inferred for this locality (Montoya et al., 1999, 2001). This isconsistent with the presence ofM. sylvanus. During the Pleistocene,European representatives of this macaque species mainly displayeda Mediterranean distribution, being probably constrained by bothclimatic and latitudinal factors (Elton and O’Regan, 2008). Thus,
D.M. Alba et al. / Journal of Human Evolution 61 (2011) 703e718716

Author's personal copy
European macaques extended their geographical ranges muchfarther northwards during the interglacials, and retreated into theirMediterranean refugia during glacials (Fooden, 2007; Elton andO’Regan, 2008). In Italy, they survived at least until the Late Pleis-tocene (OIS5e3) (Mazza et al., 2005), whereas in the IberianPeninsula they are last recorded at ca. 87e79 ka (late OIS5)(Castaños et al., 2011) and in Central Europe during the Würmian(OIS4) (Rosendahl et al., 2011). Apparently, European macaquesultimately became locally extinct because of their inability totolerate the most severe stage of the last glaciation (Fooden, 2007).Leaving cold climates aside, however, extant Barbary macaques arenot very sensitive to temperature, and their current distributionrange is characterized by a Mediterranean climate, with warm drysummers and relatively wet winters, even tolerating winter snowat high elevations (Delson, 2004; Fooden, 2007). Moreover, habitatsoccupied by extant M. sylvanus are quite varied, including cedar, !rand evergreen and deciduous oak forests, scrub, grasslands, androcky ridges dominated by herbaceous vegetation (Fooden, 2007,and references therein). Evergreen forests of cedar and oak appearto be their optimal biotope, whereas rocky slopes without arborealvegetation are more unfavorable because they need to be nearwater (Masseti and Bruner, 2009). Although extant Barbarymacaques are mainly terrestrial during the day (Fooden, 2007),they require trees for sleeping and, frequently, feeding andescaping from predators (Delson, 2004; Fooden, 2007).When theseconsiderations are taken together, the presence of macaques inQuibas is not very informative from a paleoecological viewpoint,other than con!rming that wooded and wet areas must have beenpresent near to the site, further being compatible with the exis-tence of a rocky and relatively xerophilous environment (Montoyaet al., 1999, 2001).
Acknowledgments
We thank Elisabetta Cioppi (Museo di Storia Naturale, Firenze),Francisco Pastor (Museo de Anatomía, Universidad de Valladolid),José Manuel Marín (Museu Paleontològic d’Elx-MUPE) and JoséMiguel Noguera (Director General de Cultura de la ComunidadAutónoma de Murcia) for giving us permission to study materialunder their care. We also acknowledge the comments andsuggestions by Eric Delson and two anonymous reviewers, whichsigni!cantly helped to improve a previous version of this paper.DMA also thanks Sergio Almécija for helpful discussion on thepostcranial material and Salvador Moyà-Solà for various support.This study has been possible thanks to the Spanish Ministerio deCiencia e Innovación (CGL2008-00325/BTE, and RYC-2009-04533to DMA) and the Departament d’Innovació, Universitats i Empresaof the Generalitat de Catalunya (2009 SGR 754, GRC). Fieldwork atQuibas was funded by the Government of the Region of Murcia(Séneca PI/718/02 and PI/676/04). We also acknowledge thecollaboration of the Ayuntamiento de Albanilla, the DirecciónGeneral de Cultura de la CARM, and Cajamurcia. Part of theanalytical work background for this manuscript was developed bythe senior author (DMA) in 2008e2009, while being at the EarthSciences Department of the University of Florence as a visitingresearcher.
References
Alba, D.M., Carlos Calero, J.A., Montoya, P., Moyà-Solà, S., Madurell, J., Aurell, J.,Morales, J., Mancheño, M.A., 2008a. Fossil remains of Macaca (Primates: Cer-copithecidae) from the Early Pleistocene of Spain [Abstract]. In: Giornate diPaleontologia VIII, Simposio della Società Paleontologica Italiana. Workshop suiPrimati Fossili Europei. Riassunti dei Lavori, Accademia dei Fisiocritici, Siena9e13 Settembre 2008, pp. 115e116.
Alba, D.M., Moyà-Solà, S., Madurell, J., Aurell, J., 2008b. Dentognathic remains ofMacaca (Primates, Cercopithecidae) from the late early Pleistocene of Terrassa(Catalonia, Spain). J. Hum. Evol. 55, 1160e1163.
Burnett, G.T., 1828. Illustrations of the Manupeda or apes and their allies: being thearrangements of the Quadrumana or Anthropomorphous beasts indicated inthe outline. Q. J. Sci. Lit. Art 26, 300e307.
Carlos Calero, J.A., Mancheño, M.A., Montoya, P., Ruiz Bustos, A., 2004. El yacimientopleistoceno de la Sierra de Quibas (Albanilla, Murcia). Nuevas aportaciones. In:Calonge, A., Gonzalo, R., López Carrillo, M.D., Pardo, M.V. (Eds.), Libro deResúmenes de las XX Jornadas de la Sociedad Española de Paleontología. Uni-versidad de Alcalá de Henares, pp. 40e41.
Carlos Calero, J.A., Montoya, P., Mancheño, M.A., Morales, J., 2006a. Presencia deVulpes praeglacialis (Kormos, 1932) en el yacimiento pleistoceno de la Sierra deQuibas (Abanilla, Murcia). Est. Geol. 62, 395e400.
Carlos Calero, J.A., van der Made, J., Mancheño, M.A., Montoya, P., Romero, G., 2006b.Capra alba Moyà-Solà, 1987 del Pleistoceno Inferior de la Sierra de Quibas(Abanilla, Murcia, España). Est. Geol. 62, 571e578.
Castaños, P., Murelaga, X., Arrizabalaga, A., Iriarte, M.-J., 2011. First evidence ofMacaca sylvanus (Primates, Cercopithecidae) from the Late Pleistocene ofLezetxiki II cave (Basque Country, Spain). J. Hum. Evol. 60, 816e820.
Cocchi, I., 1872. Su di due Scimmie fossili italiane. Boll. R. Com. Geol. It. 3, 59e71.Delson, E., 1980. Fossil macaques, phyletic relationships and a scenario of deploy-
ment. In: Lindburg, D.E. (Ed.), The Macaques. Studies in Ecology, Behavior andEvolution. Van Nostrand, New York, pp. 10e30.
Delson, E., 2004. Cercopithecidae from the Pliocene of Saint-Vallier. Geobios 37,S318eS322.
Delson, E., Terranova, C.J., Jungers, W.L., Sargis, E.J., Jablonski, N.G., Dechow, P.C.,2000. Body mass in Cercopithecidae (Primates, Mammalia): estimation andscaling in extinct and extant taxa. Anthropol. Pap. Am. Mus. Nat. Hist. 83, 1e159.
Durán, J.J., López-Martínez, J., Mancheño, M.A., 2004. Dos registros de espeleotemaspleistocenos de gran potencia en la Península Ibérica: primeros resultados. Bol.Geol. Min. 115, 265e270.
Elton, S., O’Regan, H.J., 2008. The biogeography and behavioural ecology of Macacain Europe [Abstract]. In: Giornate di Paleontologia VIII, Simposio della SocietàPaleontologica Italiana. Workshop sui Primati Fossili Europei. Riassunti deiLavori, Accademia dei Fisiocritici, Siena 9e13 Settembre 2008, p. 125.
Fooden, J., 2007. Systematic review of the Barbary macaque, Macaca sylvanus(Linnaeus, 1758). Fieldiana Zool. 113, 1e60.
Gentili, S., Mottura, A., Rook, L., 1998. The Italian fossil primate record: recent !ndsand their geological context. Géobios 31, 675e686.
Geoffroy Saint-Hilaire, É, 1812. Tableau des quadrumanes, 1. Ord. Quadrumanes.Ann. Mus. Hist. Nat. Paris 19, 85e122.
Gervais, P., 1859. Zoologie et Paléontologie Françaises, second ed.. Bertrand, Paris.Gray, J.E., 1821. On the natural arrangement of vertebrose animals. London Med.
Repository Rec. 15, 296e310.Groves, C., 2001. Primate Taxonomy. Smithsonian Institution Press, Washington.Jablonski, N.G., 2002. Fossil Old World monkeys: the late Neogene radiation. In:
Hartwig, W.C. (Ed.), The Primate Fossil Record. Cambridge University Press,Cambridge, pp. 255e299.
Jablonski, N., Frost, S., 2010. Cercopithecoidea. In: Werdelin, L., Sanders, W.J. (Eds.),CenozoicMammals ofAfrica.Universityof California Press, Berkeley, pp. 393e428.
Köhler, M., Moyà-Solà, S., Alba, D.M., 2000. Macaca (Primates, Cercopithecidae)from the Late Miocene of Spain. J. Hum. Evol. 38, 447e452.
Lacépède, B.G.È.de la V., 1799. Tableaux des divisions, sous-divisions, ordres etgenres des mammifères. In: Discours d’Ouverture et de Clôture du Coursd’Histoire Naturelle, l’An VII de la République; et Tableaux Méthodiques desMammifères et des Oiseaux. Plassan, Paris.
Linnaeus, C., 1758. Systema Naturae per Regna Tria Naturae, Secundum Classes,Ordines, Genera, Species, cum Characteribus, Differentiis, Synonymis, Locis.Vol. I, Regnum Animale. Editio Decima, Reformata. Laurentii Salvii, Stockholm.
Mancheño, M.A., Rodríguez Estrella, T., Romero, G., Hernández, J.M., Montoya, P.,Morales, J., Robles, F., Ruiz Bustos, A., 2003. The early Pleistocene palae-ontological site of the Sierra de Quibas (Albanilla, Murcia, SE Spain). In:Agustí, J., Oms, O., Toro, I. (Eds.), The Pleistocene: Climatic Change, DispersalDynamics and Archaeology, Meeting of the International Quaternary Associa-tion, Granada, pp. 35e36.
Masseti, M., Bruner, E., 2009. The primates of the western Palaearctic: a biogeo-graphical, historical, and archaeozoological review. J. Anthropol. Sci. 87, 33e91.
Mazza, P., Rustioni, M., Agostini, S., Rossi, A., 2005. An unexpected Late Pleistocenemacaque remain from Grotta degli Orsi Volanti (Rapino, Chieti, central Italy).Geobios 38, 211e217.
Modolo, L., Salzburger, W., Martin, R.D., 2005. Phylogeography of Barbary macaques(Macaca sylvanus) and the origin of the Gibraltar colony. Proc. Natl. Acad. Sci.102, 7392e7397.
Montoya, P., Alberdi, M.T., Blázquez, A.M., Barbadillo, L.J., Fumanal, M.P., van derMade, J.,Marín, J.M.,Molina, A.,Morales, J.,Murelaga,X., Peñalver, E., Robles, F., RuizBustos, A., Sánchez, A., Sanchiz, B., Soria, D., Szyndlar, Z., 1999. La fauna del Pleis-toceno Inferior de la Sierra de Quibas (Abanilla, Murcia). Est. Geol. 55, 127e161.
Montoya, P., Alberdi, M.T., Barbadillo, L.J., van der Made, J., Morales, J., Murelaga, X.,Peñalver, E., Robles, F., Ruiz Bustos, A., Sánchez, A., Sanchiz, B., Soria, D.,Szyndlar, Z., 2001. Une faune très diversi!ée du Pléistocène inférieur de la Sierrade Quibas (province duMurcia, Espagne), vol. 332. C. R. Acad. Sci. Paris. 387e393.
Morales, J.C., Melnick, D.J., 1998. Phylogenetic relationships of the macaques (Cer-copithecidae: Macaca), as revealed by high resolution restriction site mappingof mitochondrial ribosomal genes. J. Hum. Evol. 34, 1e23.
D.M. Alba et al. / Journal of Human Evolution 61 (2011) 703e718 717

Author's personal copy
Mottura, A., Gentili, S., 2006. Cranio-mandibular biometrics and skull maturation ofMacaca sylvanus L., 1758. Hum. Evol. 21, 223e239.
Owen, R., 1843. Report on the British fossil Mammalia. Part I: Unguiculata andCetacea. In: Report on the 12th Meeting of the British Association for theAdvancement of Science for 1842, pp. 54e74.
Owen, R., 1846. A History of British Fossil Mammals and Birds. John Van Voorst,London.
Ristori, G., 1890. Le Scimmie fossili italiane. Boll. R. Com. Geol. It. ser. III (1),178e196. 225e237.
Rodríguez Estrella, T., Mancheño, M.A., Romero, G., Hernández, J.M., 2004. Car-acterísticas geológicas de la Sierra de Quibas (Albanilla, Murcia). Su relación conun yacimiento paleontológico pleistoceno. Geogaceta 35, 115e118.
Rook, L., 1997. Macaca sylvanus !orentina (Cocchi, 1872) from the PleistoceneMugello basin (Northern Apennines, Italy). Boll. Soc. Paleontol. Ital 35,357e359.
Rook, L., Mottura, A., Gentili, S., 2001. Fossil Macaca remains from RDB quarry(Villafranca d’Asti, Italy): new data and overview. J. Hum. Evol. 40, 187e202.
Rook, L., O’Higgins, P., 2005. A comparative study of adult facial morphology and itsontogeny in the fossil macaque Macaca majori from Capo Figari, Sardinia, Italy.Folia Primatol. 76, 151e171.
Rosendahl, W., Ambros, D., Hilpert, B., Hambach, U., Alt, K.W., Knipping, M., Reisch, L.,Kaulich, B., 2011. Neanderthals and monkeys in the Würmian of Central Europe:theMiddle Paleolithic site ofHunas, SouthernGermany. In: Conard,N.J., Richter, J.(Eds.), Neanderthal Lifeways, Subsistence and Technology: One Hundred FiftyYears of Neanderthal Study. Springer, Dordrecht, pp. 15e23.
Smith, A.L., Kupzik, K., Fagan, M.J., Rook, L., Strait, D.S., O’Higgins, P., 2008. Changingfaces: an examination of robust craniofacial features in Macaca majori andimplications for the hominid fossil record [Abstract]. In: Giornate di Paleon-tologia VIII, Simposio della Società Paleontologica Italiana. Workshop sui Pri-mati Fossili Europei. Riassunti dei Lavori, Accademia dei Fisiocritici, Siena 9e13Settembre 2008, p. 146.
Stewart, C.-B., Disotell, T.R., 1998. Primate evolutiondin and out of Africa. Curr. Biol.8, R582eR588.
Szalay, F., Delson, E., 1979. Evolutionary History of the Primates. Academic Press,New York.
Zanaga, M., 1998. Macaca majori Azzaroli 1946, Primate endemico del Pleistocenedella Sardegna. Unpublished graduate thesis, Università degli Studi di Firenze.
Zapfe, H., 2001. Zähne von Macaca aus dem Unterpleistozän von Untermassfeld. In:Kahlke, R.-D. (Ed.), Das Pleistozän von Untermassfeld bei Meiningen (Thür-ingen). Teil 3. Dr. Rudolf Habelt GMBH, Bonn, pp. 889e895.
D.M. Alba et al. / Journal of Human Evolution 61 (2011) 703e718718