foraging by experienced and inexperienced cory’s shearwater along a 3-year period of ameliorating...
TRANSCRIPT

1 23
Marine BiologyInternational Journal on Life in Oceansand Coastal Waters ISSN 0025-3162 Mar BiolDOI 10.1007/s00227-015-2612-1
Foraging by experienced and inexperiencedCory’s shearwater along a 3-year period ofameliorating foraging conditions
Fredrik D. Haug, Vítor H. Paiva, AntjeC. Werner & Jaime A. Ramos

1 23
Your article is protected by copyright and
all rights are held exclusively by Springer-
Verlag Berlin Heidelberg. This e-offprint is
for personal use only and shall not be self-
archived in electronic repositories. If you wish
to self-archive your article, please use the
accepted manuscript version for posting on
your own website. You may further deposit
the accepted manuscript version in any
repository, provided it is only made publicly
available 12 months after official publication
or later and provided acknowledgement is
given to the original source of publication
and a link is inserted to the published article
on Springer's website. The link must be
accompanied by the following text: "The final
publication is available at link.springer.com”.

1 3
Mar BiolDOI 10.1007/s00227-015-2612-1
ORIGINAL PAPER
Foraging by experienced and inexperienced Cory’s shearwater along a 3‑year period of ameliorating foraging conditions
Fredrik D. Haug · Vítor H. Paiva · Antje C. Werner · Jaime A. Ramos
Received: 18 September 2014 / Accepted: 7 January 2015 © Springer-Verlag Berlin Heidelberg 2015
inexperienced individuals are enhanced by food scarcity. In fact, there was a higher spatial, trophic and behavioural segregation between both groups when environmental con-ditions were poor, which progressively diminished with improving environmental conditions. Still, we cannot rule out the fact that inexperienced birds might be gaining expe-rience with each breeding season and thus honing their for-aging skills towards those of experienced individuals.
Introduction
In vertebrates, survival and reproductive performance typi-cally increase as individuals age and get more experience (Forslund and Pärt 1995; Zimmer et al. 2011). In studies with mammals and birds, young and inexperienced breed-ers have been shown to have lower breeding success (For-slund and Pärt 1995), lower post-breeding survival (Clut-ton-Brock et al. 1996) and higher probability of skipping breeding the following season (Weimerskirch 1992), when compared to experienced individuals (Nevoux et al. 2007). There are a number of hypotheses that are not mutually exclusive which could explain these experience-related dif-ferences in breeding performance (Curio 1983). Individu-als performing poorly should likely disperse or die off until the next breeding season and, could thereby allow the mean performance of younger cohorts to improve (Smith 1981; Nol and Smith 1987), individuals could be increasing their reproductive effort with age (Williams 1966) or improving their foraging skills as they get older and gain more expe-rience (Péron and Grémillet 2013; Gutowsky et al. 2014). Changes in foraging abilities is thought to be an important variable explaining why individuals improve their breeding success as they age and get more experience (Daunt et al. 2007). Foraging performance can be considered a function
Abstract Experience is believed to be an important fac-tor determining the foraging success of animals, but there is limited knowledge on how foraging tactics differ among individuals, and on how individuals develop efficient forag-ing strategies. Pelagic seabirds are some of the longest liv-ing organisms, and in several species, breeding is deferred far beyond their physical maturity. The complex foraging skills needed to successfully rear a young is considered the most likely explanation for this life trait, making sea-birds particularly interesting for the investigation of how foraging skills differ and develop through their life span. In our study, the spatial distribution and foraging tactics of experienced and inexperienced males of a Procellari-iform seabird species, the Cory’s shearwater (Calonectris borealis) breeding on the Portuguese continental shelf, were compared along three consecutive breeding seasons with ameliorating environmental conditions (from 2010 towards 2012). Kernel overlaps of foraging areas and habi-tat modelling demonstrated that while experienced males showed high fidelity to shallow feeding grounds, inexpe-rienced birds were more explorative and relied more on less-productive pelagic areas. Our results seem to support the prediction that differences between experienced and
Communicated by S. Garthe.
F. D. Haug · V. H. Paiva (*) · A. C. Werner · J. A. Ramos MARE – Marine and Environmental Sciences Centre, Departamento de Ciências da Vida, Faculdade de Ciências e Tecnologia, Universidade de Coimbra, Coimbra, Portugale-mail: [email protected]
A. C. Werner Departamento de Ornitología, Museo de Historia Natural “Javier Prado”, Universidad Nacional Mayor de San Marcos, Av. Arenales 1256 Jesús María, Apartado 14-0434, Lima 14, Peru
Author's personal copy

Mar Biol
1 3
of foraging efficiency—the energy gained from localizing, catching and handling food items, and foraging effort—the time, distance or energy spent to catch the food item (Daunt et al. 2007). Increase in foraging performance is likely to occur as inexperienced individuals improve their knowledge of their surrounding habitat (Pärt 2001; Åkes-son and Weimerskirch 2014), expand their niche to make use of more diet items (Pütz and Cherel 2005) and develop their repertoire of foraging tactics (Daunt et al. 2007). A number of avian studies have found results in support of the idea that foraging efficiency develops with age and expe-rience (Zimmer et al. 2011; Le Vaillant et al. 2013), but there are also studies that found no emerging pattern (e.g. Weimerskirch et al. 2005). In interpreting such conflicting results, it has been suggested that some species have such an extended immaturity that they are efficient foragers by the time they start breeding (Weimerskirch et al. 2005). On the other hand, it is believed that the acquisition of efficient foraging skills is one of the key processes determining the length of an animal’s immature phase (Ashmole 1963), and according to life history theory, one would then expect individuals to commence breeding at the point when they are efficient foragers enough, and not wait until their forag-ing abilities peak. Furthermore, it has been predicted that there is an interaction between experience and environ-mental conditions (Sydeman et al. 1991; Pardo et al. 2013), suggesting that actual experience-based differences in for-aging abilities may be masked by environmental variability and only be apparent during food scarcity (Laaksonen et al. 2002; Bunce et al. 2005).
The long immature period of pelagic seabirds suggests that the foraging skills they need to attain in order to suc-cessfully rear young are very complex. These features make pelagic seabirds suitable models and interesting subjects to study ageing processes (Zimmer et al. 2011), and the sophistication of telemetry technologies are allowing bet-ter assessment of foraging behaviour at-sea. In this study, we tracked male Cory’s shearwaters (Calonectris borealis) breeding on Berlenga Island in the North Atlantic, through three consecutive pre-breeding seasons (2010–2012) with increasingly improving oceanographic conditions, and ana-lysed how at-sea distribution and behaviour of experienced males differed from those of inexperienced individuals. The oceanographic conditions around Berlenga, measured in terms of surrogates of climatic conditions (North Atlan-tic Oscillation, the NAO index; Hurrell 1995) and marine productivity [chlorophyll a concentration and sea surface temperature (SST)], improved strongly from 2010 to 2012, with a presumed positive effect on the abundance of pelagic prey fish, the main prey of Cory’s shearwater (Paiva et al. 2013b; Ceia et al. 2014).
We tested the following predictions: (1) experienced male breeders exploit different at-sea areas from those
exploited by inexperienced individuals and track environ-mental variables more efficiently than inexperienced breed-ers, which should be particularly evident during years of poor oceanographic conditions. This is expected because inexperienced individuals should have less knowledge of predictable foraging grounds, and perhaps also less ability to track environmental and biological cues (e.g. MacLean 1986). (2) Inexperienced and experienced male breeders should demonstrate increasingly similar foraging patterns and trophic ecology during years of good foraging condi-tions (Bunce et al. 2005). More specifically, we defined home ranges and key foraging areas (FAs) through kernel density estimation, assessed the degree of overlap between experienced and inexperienced birds and evaluated the similitude of their trophic ecology (stable isotopic signa-tures). The affinity for environmental variables was esti-mated through an habitat suitability modelling approach, interpreting habitat preferences of experienced and inexpe-rienced individuals over time (from 2010 to 2012).
Materials and methods
Fieldwork was conducted on Berlenga Island (39°12′40″49N, 009°30′29″W), Portugal, during the pre-breeding period of Cory’s shearwaters (April–May) along 2010, 2011 and 2012. There are about 300 breeding pairs of Cory’s shearwaters on the island (Lecoq et al. 2011) along with immature birds and failed breeders. Upon arrival at the colony (from their wintering grounds), Cory’s shearwaters require time to assess the condition of their nest site and mate, and defend that site from conspecifics before egg-laying (Warham 1990). Following these activi-ties, pairs often lack sufficient nutrient reserves to form an egg (female) or initiate incubation (male), and thus, they depart the colony for 1–2 weeks to meet these needs (Warham 1990; Paiva et al. 2013a). As in many animals reproductive success typically increases with breeding age and experience in seabirds (e.g. Forslund and Pärt 1995), so here we utilized breeding success as a proxy for expe-rience. This study benefited from a long-term database of the breeding population, such that minimum age, sex and previous breeding experience (i.e. fledging success) were known for most of the individuals (Lecoq et al. 2011). Only the at-sea activity of males was investigated, because their foraging behaviour is not dictated by egg-laying, but by the requisites of nest defence (Werner et al. 2014) and building of their own energetic reserves in order to under-take the upcoming incubation shifts. Thus, it was expected that their foraging choices and success should be largely influenced by their experience. Birds included in this study (N = 39 male Cory’s shearwaters tracked with GPS log-gers) were categorized as inexperienced (≤2 years of
Author's personal copy

Mar Biol
1 3
breeding success) and experienced (>2 successful years) individuals, based on the frequency distribution of the his-torical breeding success records for Berlenga. The breed-ing scores extended back to 2005 for our study site, and we considered the 6-year breeding success of the individuals, prior to our study (2005–2010). During our study period (2010–2012), no inexperienced individual changed to the experienced category (i.e. inexperienced kept ≤2 success-ful breeding attempts). There are caveats by assuming a correlation between cumulative breeding success and experience, but the assumption is supported by a posi-tive significant correlation (Spearman rS = 0.72, n = 39, P < 0.001) between estimated age of adult males and chicks that fledged by each individual per breeding attempt since 2005. Many individuals in the colony were not ringed as chicks between 1986 and 2005, but in 1986 every incu-bating adult was ringed and, thereafter, every unringed bird found during the breeding season was also ringed. All breeding adults were ringed by 2005, and thereafter only young breeders recruited (personal observations from birds ringed as chicks), which by 2010 had a minimum age of 7–11 years, because Cory’s shearwaters began breeding with 7–10 years of age (Mougin et al. 2000). Based on this information, in 2010 we selected inexperienced birds with 7–14 years of age and experienced birds with >20 years of age, eliminating the possibility of overlap between the two groups. Male birds used within this study were paired with the same females throughout the 3 years of study so that the overall breeding success score can be attributed to the pair. Moreover, we also found a positive significant correlation between estimated age of those females and chicks fledged per breeding attempt since 2005 (Spearman rS = 0.77, n = 39, P < 0.001).
Loggers: deployment and specifications
Cory’s shearwaters were equipped with mini-GPS log-gers (15 g; see Steiner et al. 2000 for original design), pro-grammed to collect one location (±4 m) every 5 min. The devices were attached to the birds’ back feathers, using a small piece of Tesa® tape (Fig. 1). Attachments of tags took less than 10 min, and birds were returned immediately to their nest, in order to minimize handling stress. Several studies have reported that there are no deleterious effects for birds carrying loggers if they do not represent more than 3 % of the bird’s body weight (Phillips et al. 2003), includ-ing the study species during short-term deployments (Igual et al. 2005). Assessment of logger effect was performed on the study species at this colony in 2007 (Paiva et al. 2010a), and no difference in body mass change or hatching suc-cess was found between birds that were deployed with log-gers and a randomly selected subset of birds without log-gers. In this study, the birds were weighed after retrieval
of the devices (median tracking duration for the 2010–2012 period = 14 days) to continue this evaluation of possible deleterious logger effects. The body mass change is crucial, as the birds should be building up their fat reserves for the incubation period at this time. Birds were weighed in a bag using a Pesola® balance (±20 g). Other biometric measures (especially bill measurements) were collected during field-work in previous years (e.g. Paiva et al. 2010b) to deter-mine the sex of birds that did not vocalize by a discriminant function developed by Granadeiro (1993).
Environmental data
The extended winter North Atlantic Oscillation (NAO) index was used as a large-scale environmental predictor for the North Atlantic area, and specifically for the West-ern Iberia Upwelling Ecosystem (WIUE). The NAO index refers to a north–south alternation in atmospheric mass between the subtropical Atlantic and the Arctic, and thus involves out-of-phase behaviour between the climatologi-cal low-pressure centre near Iceland and the high-pressure centre near the Azores (https://climatedataguide.ucar.edu/climate-data/hurrell-north-atlantic-oscillation-nao-index-station-based). We also used some small-scale environmen-tal predictors, such as chlorophyll a concentration (CHL) and SST data, downloaded from http://oceanocolor.gsfc.nasa.gov/, as daily night-time products with a resolution of 0.04° (approx. 4 km) in the SMI-HDF format (Table 1). Bathymetric data (BAT), taken as water depth, was down-loaded from the ETOPO2v2 database at a spatial resolu-tion of 0.03° (approximately 3 km), and 8-day composites of primary productivity (PP) at a resolution of 0.04° data were both downloaded as ASC files from the Bloom-watch website (http://coastwatch.pfel.noaa.gov/coastwatch/CWBrowserWW180.jsp). HDF files were converted to ras-ter using the Marine Geospatial Ecology Tools in ArcGIS 9.2 (Roberts et al. 2010), and then to ASCII to create com-posites. All composites were constructed using the free-ware R environment (version 2.14, R Core Team 2011) and mosaic function of the raster package. Spatial gradients of SST, CHL, PP and BAT were obtained by estimating the proportional change (PC) within a surrounding 3 × 3 cell grid using a moving window as follows: PC = [(maximum value − minimum value) × 100/maximum value] (Lou-zao et al. 2006). Gradients of SST, CHL and PP (SSTG, CHLG, PPG) are believed to be good indicators of oceanic fronts, while the BAT gradient (BATG) was used as a proxy for slope. Additionally, three more static variables were generated. Distance to colony (DCOL) and distance to land (DLAN) were calculated using the distance tool in Arc-GIS 9.2 spatial analyst, while chlorophyll a peak (CHPK) was calculated from binominal layers of CHL concentra-tion in the study period (Apr–May) during the 11 last years
Author's personal copy

Mar Biol
1 3
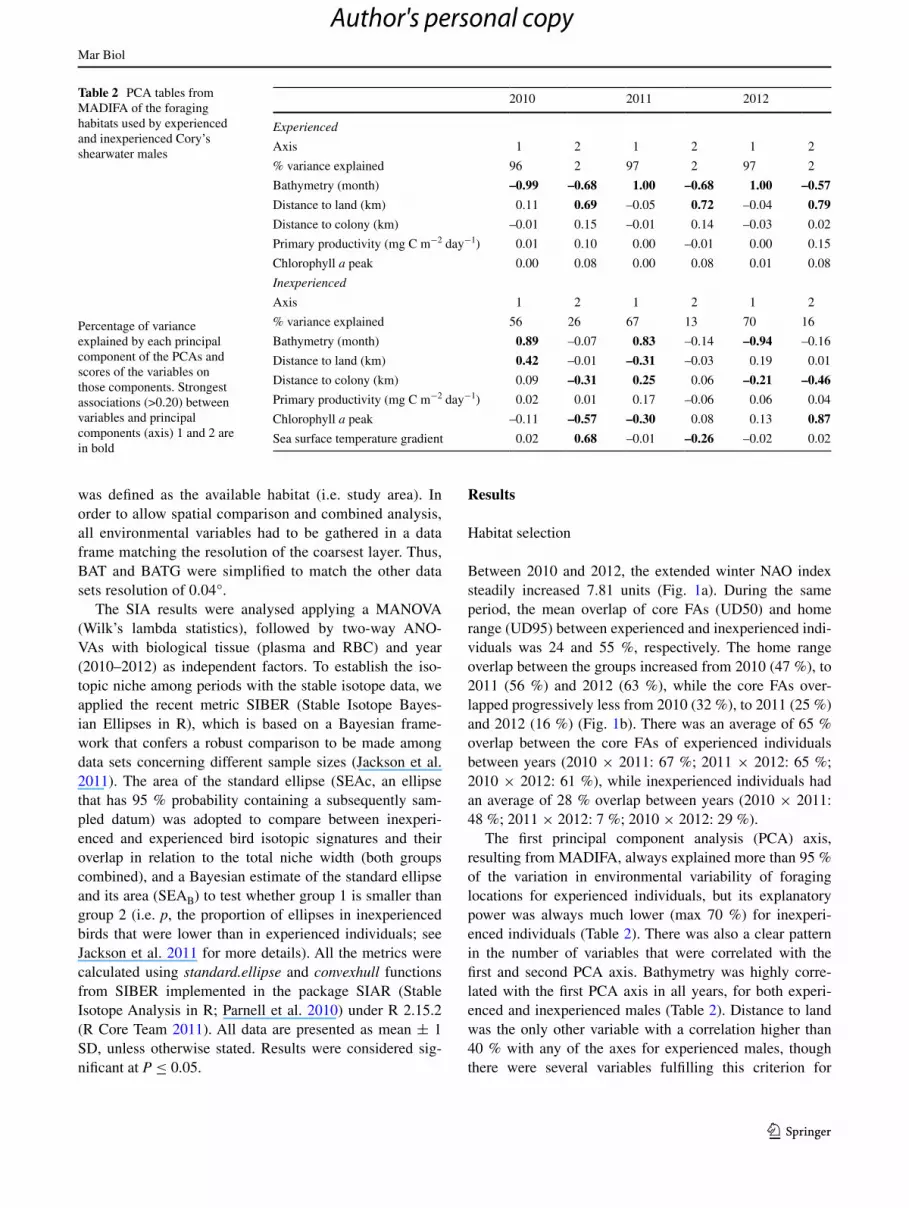
Fig. 1 a Values of the extended winter (December–March) NAO index (https://climatedataguide.ucar.edu/climate-data/hurrell-north-atlantic-oscillation-nao-index-station-based). b Home range (UD95; lines) and core FAs (UD50; filled polygons) of experienced (blue) and inexperienced (red) Cory’s shearwaters from Berlenga Island (white star), estimated for each year (2010–2012) and plotted over bathym-
etry (month) of the study area. c Isotopic niche area based on stable isotope ratios (δ13C and δ15N) in RBCs and plasma of experienced (blue dots) and inexperienced (red dots) tracked Cory’s shearwaters. The Standard ellipses areas (SEAc) are represented by the solid bold lines (see Jackson et al. 2011 for more details on these metrics of iso-topic niche width)
Author's personal copy

Mar Biol
1 3
(2002–2012). Cells with a CHL concentration >1 mg m−3 were assigned a value of 1 and lower values assigned zeros (Louzao et al. 2012; Suryan et al. 2012; Afán et al. 2014).
Trip filtering
GPS data were divided into individual foraging excur-sions by calculating the running distance to colony from GPS position and colony location. Trip duration was calcu-lated, and the data were divided in short (≤1 day) and long (>1 day) trips by inspecting a frequency histogram of trip duration. For analysis of at-sea habitat use by experienced and inexperienced birds, we selected only the short excur-sions (=1 day of duration) of male individuals because: (1) the short excursions are more representative of the male Cory’s shearwater foraging strategy during the pre-breed-ing season, as most individuals return every night to defend their burrow site (i.e. foraging movements are rather local; see Werner et al. 2014). (2) As a result, the long trips are under-represented and the sample sizes were too small to analyse and interpret differences between experienced and inexperienced individuals. Thus, long foraging excursions of experienced and inexperienced males were not consid-ered for further analysis (2010, n = 9 and 8; 2011, n = 3 and 4 and 2012, n = 4 and 4, respectively). A random sub-set of the 2010 data set of short trips was selected in order to have a comparable sample size between individuals of both groups (i.e. experienced and inexperienced males) within each year and between years. Then, geolocations (between consecutive tracking points) were filtered on running flight speed. In attempt to exclude periods where
the birds were resting and drifting on the water surface, a lower threshold of 9 km h−1 (i.e. based on the frequency distribution of speed records; Guilford et al. 2008) was set for the flight data set that was used for estimation of home ranges (HR). We realize that the bird might still be foraging at lower speeds (instead of landing or leaving the sea surface) or feeding while drifting through a “sit and wait” technique, but because we cannot confirm a foraging behaviour in those situations, those dubious situations (i.e. locations with speed records <9 km h−1) were excluded from the analysis. In addition to the filtered flight data set, a data set of foraging locations was created to estimate FA. This was done by calculating path sinuosity for all the loca-tions, defined as the ratio of the actual flight speed given by the GPS receiver to the velocity between every third fix (geographical location). Birds that are circling an area will display a lower calculated speed than the actual GPS speed, and therefore have a higher sinuosity index (Grémillet et al. 2004). A histogram of the sinuosity distribution was used to determine the break-off value, which was set at sinuos-ity index = 1.7. Finally, following the same approach, a distance-to-colony filter of 2 km was applied in order to remove sinuosity that may be associated with social inter-action and particularly while flying in the colony surround-ings before landing.
Kernel estimation
Utilization distribution (UD) kernels were used to char-acterize the distribution of experienced and inexperi-enced birds. Home ranges (UD95) and FAs (UD50) were
Table 1 Overview of the environmental variables used in the analysis of habitat selection and associated oceanographic processes
Explanatory variables Satellite Spatial resolution Temporal resolution Range (min–max) Oceanographic process (descrip-tion)
Dynamic
Chlorophyll a (CHL, mg m−3) Aqua-MODIS 0.04° Daily 0.06–31.7 Ocean productivity
Primary productivity (PP, mg C m−2 day−1)
BLENDED 0.04° Daily 293–7,150 Ocean productivity
Sea surface temperature (SST, °C)
Aqua-MODIS 0.04° Daily 13.7–22.7 Water mass distribution
CHL gradient (CHLG) Aqua-MODIS 0.04° Daily 0.1–98.4 Oceanic fronts
PP gradient (PPG) Aqua-MODIS 0.04° Daily 0.1–81.2 Oceanic fronts
SST gradient (SSTG) Aqua-MODIS 0.04° Daily 0.05–78.3 Oceanic fronts
Static
Bathymetry (BAT, month) ETOPO 0.04° – 1–5,215 Neritic versus pelagic domains
BAT gradient (BATG) ETOPO 0.04° – 0.1–99.3 Slope
Distance to colony (DCOL, km) – 0.03° – 0–276.5 Neritic versus pelagic domains
Distance to land (DLAN, km) – 0.03° – 0–283.8 Neritic versus pelagic domains
Chlorophyll peak (CHPK, CHL >1 mg month−3)
Aqua-MODIS 0.04° – 0–10 Productivity persistence
Author's personal copy

Mar Biol
1 3
calculated from the flight-filtered and foraging-filtered data sets, respectively, using UD kernel methods (Wor-ton 1989). All kernels were estimated using adehabitat and adehabitatHR R packages (Calenge 2006). To allow comparison between the two groups and all years, the ad hoc method was used to find a reasonable smoothing fac-tor (h) and the mean h value of individual kernels for the study sample was computed. A fixed smoothing factor of 3,100 m was then used to compute all kernels. Trips that were too short and fine-scaled to be estimated at this level were discarded along with those trips with less than 5 loca-tions (i.e. the minimum number of locations to estimate a kernel) of either flight or foraging after the filtering process in the previous section (n = 5 trips for all study years). The overlap between kernels of experienced and inexperienced males each year was calculated to determine whether there was any spatial segregation between the groups. Overlaps were also estimated between each year within the groups to assess the consistency of habitat selection in each group. Both calculations were performed in R with the kernelover-lap function of adehabitat and using the VI method.
Trip filtering and kernel estimation resulted in a total sample of 152 trips used for further analysis. We sampled 10, 5 and 6 experienced males (corresponding to 26, 32 and 18 foraging trips), and 8, 5 and 5 inexperienced males (cor-responding to 26, 29 and 21 foraging excursions), respec-tively, for 2010, 2011 and 2012. The same individuals were tracked during the three consecutive years (though some individuals of 2010 were tracked only that year) and sam-pled for stable isotope analysis (SIA, see below). During pre-laying, birds attend the breeding colony less frequently than during the incubation and chick provisioning periods, so we could not track all experienced (n = 10) and inexpe-rienced (n = 8) males of 2010 in the two following years.
Stable isotope analysis (SIA)
Stable-nitrogen isotope ratios (15N:14N, expressed as δ15N) and stable-carbon isotope ratios (13C:12C, expressed as δ13C) on plasma and red blood cells (RBCs) of Cory’s shearwater were assessed to investigate the effect of expe-rience on their foraging ecology and prey selection. The ratios represent the prey ingested in the past few weeks before the blood sample. The δ15N is mainly used to define the trophic position of the consumer, while δ13C reflects the foraging habitat of the consumer (Inger and Bearhop 2008; Ceia et al. 2012). It is assumed that potential intraspecific differences in assimilation efficiency or physiology do not impact the blood isotope values significantly (Votier et al. 2011). There is a gradient of high to low values of δ13C from coast to offshore due to the organic enrichment at the coast that is gradually diluted. RBCs are regenerated every
12–22 days, while plasma has a turnover rate of about 7 days; therefore, they represent prey ingestion in different time scopes: RBC reflects the trophic ecology the last few weeks, and plasma reflects choices made in the last trips before sampling (i.e. around 7 days; Inger and Bearhop 2008).
Each of the tracked birds was sampled upon return from a foraging trip. Blood samples (1 ml) were collected from the tarsal or brachial vein using 25-gauge needles under licence. Blood samples were then separated into plasma and red blood cells (hereafter termed RBCs) by a centrifu-gation at 12,000 rpm for 15 min, within 2–4 h of sampling and stored frozen at −20 °C until preparation for analysis. Successive rinses with a 2:1 chloroform/methanol solution were performed on the plasma samples in order to deplete it for lipids, which may disturb the results (Cherel et al. 2005b). As the lipid content of RBCs is typically low, the lipid extraction is not required for that tissue (Cherel et al. 2005a). Samples were dried in an aspirating hood for 48 h at 60 °C for the ethanol to evaporate. Isotope ratios of car-bon and nitrogen were then determined through standard methodology (Bearhop et al. 2006; Phillips et al. 2009) by continuous-flow isotope ratio mass spectrometry, using an EA-IRMS (Isoprime, Micromass, UK). The analytical pre-cision for the measurement was 0.2 ‰ for both carbon and nitrogen. All values presented are means ± 1 SD unless otherwise stated.
Data analysis
To understand whether there was an effect of experience on the habitat use, a Mahalanobis distance factor analy-sis (MADIFA, Calenge et al. 2008) comparing the envi-ronmental characteristics of chosen foraging grounds with those of the available environment was applied. MADIFA is a multivariate modelling technique developed to make habitat suitability maps from presence-only data, such as tracking locations (e.g. Thiebot et al. 2011). The MADIFA was a good option for this study because it con-siders the environmental variability of the area where the niche was sampled, and may therefore be more accurate than other methods (Tsoar et al. 2007). It is also comple-mentary to the commonly applied ecological-niche factor analysis (ENFA) and runs on the freely available R envi-ronment (R Core Team 2011). The available environment had to be defined for the niche and MADIFA analyses. As only short trips of 1 day or less were kept for the analy-sis, the maximum daily range was estimated by the fol-lowing formula: ([mean trip duration − mean time spent foraging per trip] × mean flight speed × 1/2). Thus, a daily range of about 240 km was estimated, approximat-ing the 2° in each direction off and along the shore, which
Author's personal copy
Mar Biol
1 3
was defined as the available habitat (i.e. study area). In order to allow spatial comparison and combined analysis, all environmental variables had to be gathered in a data frame matching the resolution of the coarsest layer. Thus, BAT and BATG were simplified to match the other data sets resolution of 0.04°.
The SIA results were analysed applying a MANOVA (Wilk’s lambda statistics), followed by two-way ANO-VAs with biological tissue (plasma and RBC) and year (2010–2012) as independent factors. To establish the iso-topic niche among periods with the stable isotope data, we applied the recent metric SIBER (Stable Isotope Bayes-ian Ellipses in R), which is based on a Bayesian frame-work that confers a robust comparison to be made among data sets concerning different sample sizes (Jackson et al. 2011). The area of the standard ellipse (SEAc, an ellipse that has 95 % probability containing a subsequently sam-pled datum) was adopted to compare between inexperi-enced and experienced bird isotopic signatures and their overlap in relation to the total niche width (both groups combined), and a Bayesian estimate of the standard ellipse and its area (SEAB) to test whether group 1 is smaller than group 2 (i.e. p, the proportion of ellipses in inexperienced birds that were lower than in experienced individuals; see Jackson et al. 2011 for more details). All the metrics were calculated using standard.ellipse and convexhull functions from SIBER implemented in the package SIAR (Stable Isotope Analysis in R; Parnell et al. 2010) under R 2.15.2 (R Core Team 2011). All data are presented as mean ± 1 SD, unless otherwise stated. Results were considered sig-nificant at P ≤ 0.05.
Results
Habitat selection
Between 2010 and 2012, the extended winter NAO index steadily increased 7.81 units (Fig. 1a). During the same period, the mean overlap of core FAs (UD50) and home range (UD95) between experienced and inexperienced indi-viduals was 24 and 55 %, respectively. The home range overlap between the groups increased from 2010 (47 %), to 2011 (56 %) and 2012 (63 %), while the core FAs over-lapped progressively less from 2010 (32 %), to 2011 (25 %) and 2012 (16 %) (Fig. 1b). There was an average of 65 % overlap between the core FAs of experienced individuals between years (2010 × 2011: 67 %; 2011 × 2012: 65 %; 2010 × 2012: 61 %), while inexperienced individuals had an average of 28 % overlap between years (2010 × 2011: 48 %; 2011 × 2012: 7 %; 2010 × 2012: 29 %).
The first principal component analysis (PCA) axis, resulting from MADIFA, always explained more than 95 % of the variation in environmental variability of foraging locations for experienced individuals, but its explanatory power was always much lower (max 70 %) for inexperi-enced individuals (Table 2). There was also a clear pattern in the number of variables that were correlated with the first and second PCA axis. Bathymetry was highly corre-lated with the first PCA axis in all years, for both experi-enced and inexperienced males (Table 2). Distance to land was the only other variable with a correlation higher than 40 % with any of the axes for experienced males, though there were several variables fulfilling this criterion for
Table 2 PCA tables from MADIFA of the foraging habitats used by experienced and inexperienced Cory’s shearwater males
Percentage of variance explained by each principal component of the PCAs and scores of the variables on those components. Strongest associations (>0.20) between variables and principal components (axis) 1 and 2 are in bold
2010 2011 2012
Experienced
Axis 1 2 1 2 1 2
% variance explained 96 2 97 2 97 2
Bathymetry (month) –0.99 –0.68 1.00 –0.68 1.00 –0.57
Distance to land (km) 0.11 0.69 –0.05 0.72 –0.04 0.79
Distance to colony (km) –0.01 0.15 –0.01 0.14 –0.03 0.02
Primary productivity (mg C m−2 day−1) 0.01 0.10 0.00 –0.01 0.00 0.15
Chlorophyll a peak 0.00 0.08 0.00 0.08 0.01 0.08
Inexperienced
Axis 1 2 1 2 1 2
% variance explained 56 26 67 13 70 16
Bathymetry (month) 0.89 –0.07 0.83 –0.14 –0.94 –0.16
Distance to land (km) 0.42 –0.01 –0.31 –0.03 0.19 0.01
Distance to colony (km) 0.09 –0.31 0.25 0.06 –0.21 –0.46
Primary productivity (mg C m−2 day−1) 0.02 0.01 0.17 –0.06 0.06 0.04
Chlorophyll a peak –0.11 –0.57 –0.30 0.08 0.13 0.87
Sea surface temperature gradient 0.02 0.68 –0.01 –0.26 –0.02 0.02
Author's personal copy
Mar Biol
1 3
inexperienced males, including SST, SST gradient and dis-tance to colony (Table 2).
Stable isotope analysis (SIA)
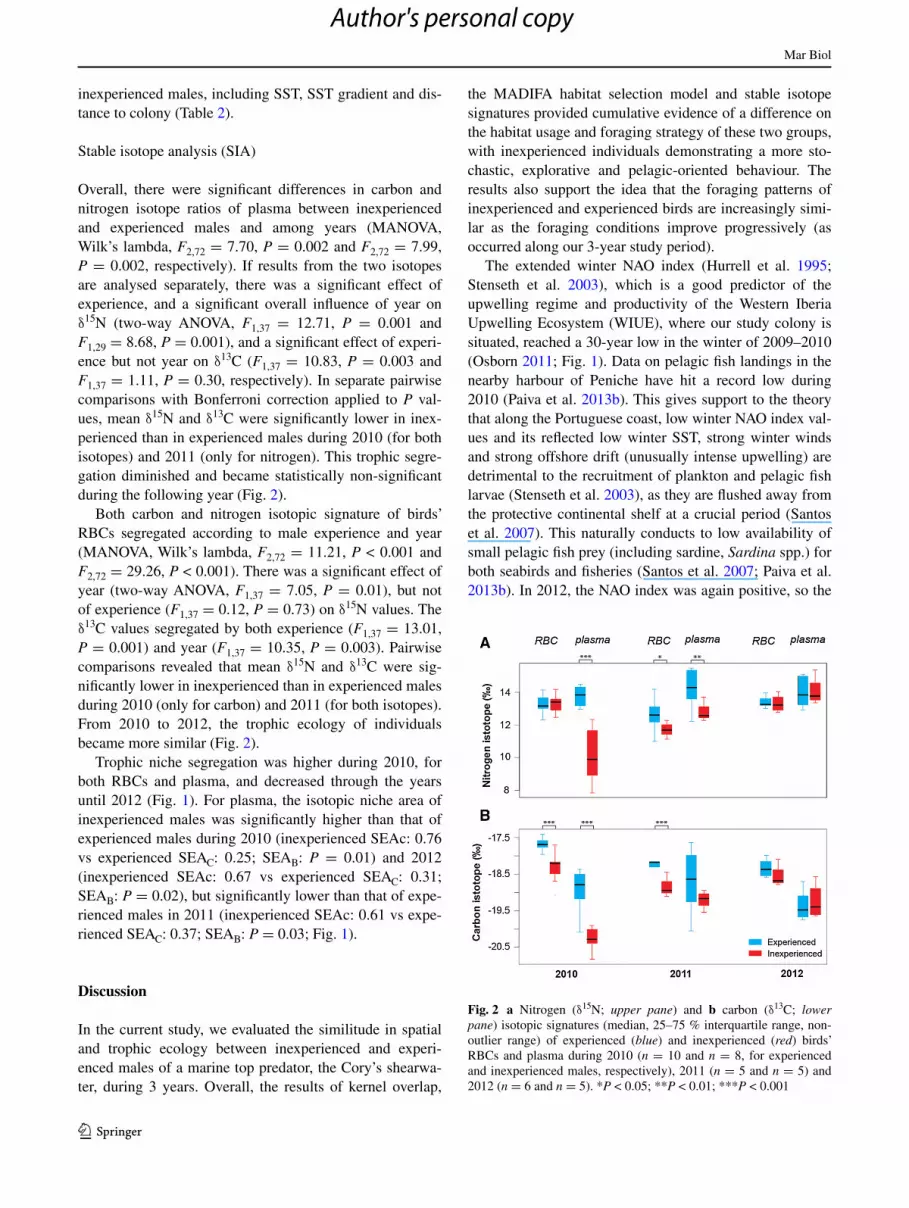
Overall, there were significant differences in carbon and nitrogen isotope ratios of plasma between inexperienced and experienced males and among years (MANOVA, Wilk’s lambda, F2,72 = 7.70, P = 0.002 and F2,72 = 7.99, P = 0.002, respectively). If results from the two isotopes are analysed separately, there was a significant effect of experience, and a significant overall influence of year on δ15N (two-way ANOVA, F1,37 = 12.71, P = 0.001 and F1,29 = 8.68, P = 0.001), and a significant effect of experi-ence but not year on δ13C (F1,37 = 10.83, P = 0.003 and F1,37 = 1.11, P = 0.30, respectively). In separate pairwise comparisons with Bonferroni correction applied to P val-ues, mean δ15N and δ13C were significantly lower in inex-perienced than in experienced males during 2010 (for both isotopes) and 2011 (only for nitrogen). This trophic segre-gation diminished and became statistically non-significant during the following year (Fig. 2).
Both carbon and nitrogen isotopic signature of birds’ RBCs segregated according to male experience and year (MANOVA, Wilk’s lambda, F2,72 = 11.21, P < 0.001 and F2,72 = 29.26, P < 0.001). There was a significant effect of year (two-way ANOVA, F1,37 = 7.05, P = 0.01), but not of experience (F1,37 = 0.12, P = 0.73) on δ15N values. The δ13C values segregated by both experience (F1,37 = 13.01, P = 0.001) and year (F1,37 = 10.35, P = 0.003). Pairwise comparisons revealed that mean δ15N and δ13C were sig-nificantly lower in inexperienced than in experienced males during 2010 (only for carbon) and 2011 (for both isotopes). From 2010 to 2012, the trophic ecology of individuals became more similar (Fig. 2).
Trophic niche segregation was higher during 2010, for both RBCs and plasma, and decreased through the years until 2012 (Fig. 1). For plasma, the isotopic niche area of inexperienced males was significantly higher than that of experienced males during 2010 (inexperienced SEAc: 0.76 vs experienced SEAC: 0.25; SEAB: P = 0.01) and 2012 (inexperienced SEAc: 0.67 vs experienced SEAC: 0.31; SEAB: P = 0.02), but significantly lower than that of expe-rienced males in 2011 (inexperienced SEAc: 0.61 vs expe-rienced SEAC: 0.37; SEAB: P = 0.03; Fig. 1).
Discussion
In the current study, we evaluated the similitude in spatial and trophic ecology between inexperienced and experi-enced males of a marine top predator, the Cory’s shearwa-ter, during 3 years. Overall, the results of kernel overlap,
the MADIFA habitat selection model and stable isotope signatures provided cumulative evidence of a difference on the habitat usage and foraging strategy of these two groups, with inexperienced individuals demonstrating a more sto-chastic, explorative and pelagic-oriented behaviour. The results also support the idea that the foraging patterns of inexperienced and experienced birds are increasingly simi-lar as the foraging conditions improve progressively (as occurred along our 3-year study period).
The extended winter NAO index (Hurrell et al. 1995; Stenseth et al. 2003), which is a good predictor of the upwelling regime and productivity of the Western Iberia Upwelling Ecosystem (WIUE), where our study colony is situated, reached a 30-year low in the winter of 2009–2010 (Osborn 2011; Fig. 1). Data on pelagic fish landings in the nearby harbour of Peniche have hit a record low during 2010 (Paiva et al. 2013b). This gives support to the theory that along the Portuguese coast, low winter NAO index val-ues and its reflected low winter SST, strong winter winds and strong offshore drift (unusually intense upwelling) are detrimental to the recruitment of plankton and pelagic fish larvae (Stenseth et al. 2003), as they are flushed away from the protective continental shelf at a crucial period (Santos et al. 2007). This naturally conducts to low availability of small pelagic fish prey (including sardine, Sardina spp.) for both seabirds and fisheries (Santos et al. 2007; Paiva et al. 2013b). In 2012, the NAO index was again positive, so the
Fig. 2 a Nitrogen (δ15N; upper pane) and b carbon (δ13C; lower pane) isotopic signatures (median, 25–75 % interquartile range, non-outlier range) of experienced (blue) and inexperienced (red) birds’ RBCs and plasma during 2010 (n = 10 and n = 8, for experienced and inexperienced males, respectively), 2011 (n = 5 and n = 5) and 2012 (n = 6 and n = 5). *P < 0.05; **P < 0.01; ***P < 0.001
Author's personal copy

Mar Biol
1 3
reverse effect of what we described before should occur, and the study of Ceia et al. (2014) on Yellow-legged gulls Larus michahellis breeding on Berlengas strongly indicates that the abundance of small pelagic fish was considerably higher in 2012 than in 2011.
There were some evidences of possible niche spatial segregation between experienced and inexperienced males: (1) The kernel utilization distribution analysis (kernel UD) showed that the FA’s of the two groups had a mean overlap of 24 % among years, which suggests some spatial segrega-tion between the two groups considering the relatively dis-closed habitat males make use of during the pre-breeding season, when compared to the vast distribution they can also have during the same season (Werner et al. 2014) and during the remaining phases of their breeding period (Paiva et al. 2013b). (2) The mean overlap between experienced and inexperienced males was also considerably lower than the mean overlap of experienced males among the 3 years (65 %), and about the same as the mean for the inexperi-enced males among the same 3-year period (28 %), sug-gesting a more stochastic and explorative foraging pattern for the inexperienced group of birds. (3) Overall, inexpe-rienced males had larger home ranges and relied more on FAs farther from the mainland coast and the breeding col-ony. Although there were few FAs used exclusively by one of the groups, experienced males highly used low depth habitats in all the three study years, suggesting a consist-ent use of potentially productive shallow areas along the coast and seamounts outside Berlenga archipelago, which is giving a clear signal of interannual foraging consistency for experienced males. Bathymetry was the most impor-tant explanatory variable for the inexperienced males too, but here the signal was weaker and there were a number of other variables with explanatory power changing from year to year. Experience had a significant effect on the carbon stable isotope signatures, with inexperienced males hav-ing a more pelagic signature (i.e. lower-carbon signature; Paiva et al. 2010c). This more pelagic distribution was also corroborated by their association with oceanographic prox-ies of frontal systems, like gradients of SST (Paiva et al. 2010a, 2013b). More explorative strategies among younger members of a population have been reported by several studies, comparing immatures, juveniles and adults (Thie-bot et al. 2011; Votier et al. 2011), but we are not aware of any study demonstrating such a clear separation within a breeding population. Another factor, which may be contrib-uting to the spatial segregation of Cory’s shearwater breed-ing on Berlengas, is that the population has been increas-ing by 10 % each year in the last two decades (Lecoq et al. 2011).
Another aim of our study was to assess whether expe-rienced and inexperienced males approached each other in habitat use and trophic niche as foraging conditions
improved progressively. From 2010 to 2012, the trophic ecology of both groups approached year after year, either in terms of the trophic level of the prey that birds were consuming (nitrogen signature) or in terms of the loca-tion where they were feeding (carbon signature), in spite of a slight reduction in the overlap of FAs (but increasing home range overlap). Inexperienced males seem to have been preying on low trophic level prey when compared to experienced males, because they presented a lower nitro-gen signature mostly during 2010 and 2011. There might be at least two reasons for this: (1) the inexperienced birds were more explorative in 2010 and 2011 (i.e. years of poor environmental conditions, thus probably with lower food availability) preying on low trophic level prey, such as cephalopod or less nitrogen-rich small pelagic fish spe-cies (e.g. Scomber sp.; Paiva et al. 2010c) or (2) in years of food scarcity, birds tend to attend more to fishery discards (Bicknell et al. 2013), and when doing so, experienced individuals might gain access to higher trophic level prey in competition with inexperienced males (Navarro et al. 2010). Although both hypotheses can be true, only the col-lection of more tracking and blood data along with vessel monitoring system data during subsequent years will disen-tangle the major driver of this pattern between experienced and inexperienced males. Nevertheless, both groups were isotopically segregated even in years where experienced and inexperienced males were foraging on the same regions (i.e. a higher FA overlap in 2010 and 2011 than in 2012), which certainly results from individuals feeding on isotopi-cally different prey (Votier et al. 2011).
Comparable studies on other seabird species have reported on (1) inexperienced individuals differing in their foraging strategy, but not in their efficiency, from experi-enced individuals (Weimerskirch et al. 2005), (2) a signifi-cant effect of experience on breeding success, but not on foraging efficiency (Nevoux et al. 2007), (3) differences in area usage and foraging performance between juveniles and adults, but not between immatures and adults, while other studies show differences in either reproductive (Sydeman et al. 1991) or foraging performance (Bunce et al. 2005) between experienced and inexperienced birds, particularly during years of reduced food availability. A better forag-ing performance by experienced birds during years of food scarcity could be a result of traits being canalized against environmental variation in more experienced individuals (Gaillard and Yoccoz 2003; Pardo et al. 2013). Daunt et al. (2007) found in European Shags that experienced individu-als coped better with environmental variability, because they adjusted their foraging effort during periods of food scarcity. Also in our study, in years of food scarcity (mostly during 2010, but also in 2011) experienced birds foraged on a more confined area (i.e. less foraging effort) and in higher trophic level prey when compared to inexperienced
Author's personal copy

Mar Biol
1 3
individuals, thus suggesting that they performed better under environmental stochasticity. Experienced and inexpe-rienced individuals may also approach each other in habitat selection and foraging performance as a result of the devel-opment of foraging abilities by the inexperienced indi-viduals. A recent study focusing on learning processes of Wandering Albatrosses (Riotte-Lambert and Weimerskirch 2013) found evidence of differences in area usage and for-aging performance between juveniles and adults. Consid-ering the relatively short time frame of our study and the great variation in environmental conditions, we suggest that inexperienced males approached foraging abilities of expe-rienced males mainly due to improved foraging conditions (although we cannot exclude the “learning hypothesis”).
In future studies, it would be interesting to collect simi-lar data on birds of known age from their first breeding attempt, and using longer time series, to separate the role of environmental conditions and learning, in explaining the foraging patterns and breeding success as individuals age and acquire breeding and foraging experience. The follow-up of foraging movements throughout the year would ena-ble to assess whether there is continuous evidence of the inexperienced breeders being more naive foragers, and to model the response of the animals based on experience. Finally, it would also be interesting to look into senes-cence-related questions, and perhaps determine until what age physiological deterioration is no longer balanced by foraging experience (Zimmer et al. 2011).
In summary, our study found at-sea spatial segrega-tion between experienced and inexperienced male Cory’s shearwaters during the pre-breeding periods of 2010–2012. The MADIFA habitat use model and stable isotope signa-tures of carbon demonstrated that while experienced males showed high fidelity to shallow feeding grounds, inexpe-rienced birds were more explorative and relied more on less-productive pelagic areas. Plus, in years of poor envi-ronmental conditions (2010–2011) both groups seem to segregate more in their foraging tactics and trophic ecology (i.e. higher nitrogen and carbon signatures for experienced birds). With ameliorating foraging conditions (especially during 2012), both groups resembled more in their forag-ing tactics and trophic ecology. In the current context of unprecedented rapid global changes and loss of biodiver-sity, it is valuable to appreciate such intraspecific differ-ences in area usage and foraging abilities to better model future area usage and population trajectories (Pardo et al. 2013).
Acknowledgments We would like to thank the Instituto da Con-servação da Natureza e Florestas (ICNF) for their logistical support (lodging), especially the wardens of the Reserva Natural das Ber-lengas, Paulo Crisóstomo and Eduardo Mourato, for their compan-ionship. Also a special thanks to Filipe Ceia and Margarida Soares for their help during fieldwork. GPS loggers were financed by the
EU INTERREG project FAME: The Future of the Atlantic Marine Environment. F.D.H. acknowledges the study grant given by the EMMC-EMAE consortium and the European Commission. V.H.P. acknowledges the postdoctoral grants given by Fundação para a Ciência e Tecnologia (FCT; SFRH/BPD/63825/2009 and SFRH/BPD/85024/2012). The experimental approach was conducted with permission from the Portuguese Government (ICNF). All methods used in this study comply with the current laws of Portugal.
References
Afán I, Navarro J, Cardador L, Ramírez F, Kato A, Rodríguez B, Ropert-Coudert Y, Forero MG (2014) Foraging movements and habitat niche of two closely related seabirds breeding in sympa-try. Mar Biol 161:657–668
Åkesson S, Weimerskirch H (2014) Evidence for sex-segregated ocean distributions first-winter wandering albatrosses at Crozet Islands. PLoS One 9(2):e86779
Ashmole NP (1963) The regulation of numbers of tropical oceanic birds. Ibis 103b:458–473
Bearhop S, Phillips R, McGill R, Cherel Y, Dawson D, Croxall J (2006) Stable isotopes indicate sex-specific and long-term indi-vidual foraging specialisation in diving seabirds. Mar Ecol Prog Ser 311:157–164
Bicknell A, Oro D, Camphuysen K (2013) Potential consequences of discard reform for seabird communities. J Appl Ecol 50:649–658
Bunce A, Ward SJ, Norman FI (2005) Are age-related variations in breeding performance greatest when food availability is limited? J Zool 266:163–169
Calenge C (2006) The package “adehabitat” for the R software: a tool for the analysis of space and habitat use by animals. Ecol Model 197:516–519
Calenge C, Darmon G, Basille M, Loison A, Jullien J-M (2008) The factorial decomposition of the Mahalanobis distances in habitat selection studies. Ecology 89:555–566
Ceia FR, Phillips RA, Ramos JA, Cherel Y, Vieira RP, Richard P, Xavier JC (2012) Short- and long-term consistency in the forag-ing niche of wandering albatrosses. Mar Biol 159:1581–1591
Ceia FR, Paiva VH, Fidalgo V, Morais L, Beata A, Crisóstomo P, Mourato E, Garthe S, Marques JC, Ramos JA (2014) Annual and seasonal consistency in the feeding ecology of an opportunistic species, the yellow-legged gull (Larus michahellis). Mar Ecol Prog Ser 497:273–284
Cherel Y, Hobson KA, Hassani S (2005a) Isotopic discrimination between food and blood and feathers of captive penguins: impli-cations for dietary studies in the wild. Physiol Biochem Zool 78:106–115
Cherel Y, Hobson KA, Weimerskirch H (2005b) Using stable isotopes to study resource acquisition and allocation in procellariform sea-birds. Oecologia 145:533–540
Clutton-Brock TH, Stevenson IR, Marrow P, MacColl AD, Houston AI, McNamara JM (1996) Population fluctuations, reproductive costs and life-history tactics in female Soay sheep. J Anim Ecol 65:675–689
Curio E (1983) Why do young birds reproduce less well? Ibis 125:400–404
Daunt F, Wanless S, Harris MP, Money L, Monaghan P (2007) Older and wiser: improvements in breeding success are linked to better foraging performance in European shags. Funct Ecol 21:561–567
Forslund P, Pärt T (1995) Age and reproduction in birds—hypotheses and tests. Trends Ecol Evol 10:364–378
Gaillard JM, Yoccoz NG (2003) Temporal variation in survival of mammals: a case of environmental canalization? Ecology 84:3294–3306
Author's personal copy

Mar Biol
1 3
Granadeiro JP (1993) Variation in measurements of Cory’s shearwater between populations and sexing by discriminant analysis. Ring Migr 14:103–112
Grémillet D, Omo GD, Ryan PG, Peters G, Ropert-Coudert Y, Weeks SJ (2004) Offshore diplomacy, or how seabirds mitigate intra-specific competition : a case study based on GPS tracking of Cape gannets from neighbouring colonies. Mar Ecol Prog Ser 268:265–279
Guilford TC, Meade J, Freeman R, Biro D, Evans T, Bonadonna F, Boyle D, Roberts S, Perrins CM (2008) GPS tracking of the for-aging movements of Manx Shearwaters Puffinus puffinus breed-ing on Skomer Island, Wales. Ibis 150:462–473
Gutowsky SE, Tremblay Y, Kappes MA, Flint EN, Klavitter J, Lani-awe L, Shaffer SA (2014) Divergent post-breeding distribution and habitat associations of fledgling and adult Black-footed Alba-trosses Phoebastria nigripes in the North Pacific. Ibis 156:60–72
Hurrell JW (1995) Decadal trends in the North Atlantic Oscillation: regional temperatures and precipitation. Science 269:676–679
Igual JM, Forero MG, Tavecchia G, González-Solís J et al (2005) Short-term effects of data-loggers on Cory’s shearwater (Calonectris diomedea). Mar Biol 146:619–624
Inger R, Bearhop S (2008) Applications of stable isotope analyses to avian ecology. Ibis 150:447–461
Jackson AL, Inger R, Parnell AC, Bearhop S (2011) Comparing iso-topic niche widths among and within communities: SIBER—Sta-ble Isotope Bayesian Ellipses in R. J Anim Ecol 80:595–602
Laaksonen T, Korpimäki E, Hakkarainen H (2002) Interactive effects of parental age and environmental variation on the breeding per-formance of Tengmalm’s owl. J Anim Ecol 71:23–31
Le Vaillant M, Le Bohec C, Prud’Homme O, Wienecke B, Le Maho Y, Kato A, Ropert-Coudert Y (2013) How age and sex drive the foraging behaviour in the king penguin. Mar Biol 160:1147–1156
Lecoq M, Catry P, Granadeiro JP (2011) Population trends of Cory’s shearwaters Calonectris diomedea borealis breeding at Berlengas Islands, Portugal. Airo 20:36–41
Louzao M, Hyrenbach KD, Abelló P, de Sola LG, Oro D (2006) Ocean-ographic habitat of an endangered Mediterranean procellariform: implications for marine protected areas. Ecol Appl 16:1683–1695
Louzao M, Delord K, García D, Boué A, Weimerskirch H (2012) Pro-tecting persistent dynamic oceanographic features: transbound-ary conservation efforts are needed for the critically endangered balearic shearwater. PLoS One 7:1–12
MacLean AAE (1986) Age-specific foraging ability and the evolu-tion of deferred breeding in three species of gulls. Wilson Bull 98:267–279
Mougin J-L, Jouanin C, Roux F (2000) Démographie du puffin cen-dré Calonectris diomedea de Selvagem Grande. Rev Ecol (Terre Vie) 55:275–290
Navarro J, Oro D, Bertolero A, Genovart M, Delgado A, Forero MG (2010) Age and sexual differences in the exploitation of two anthropogenic food resources for an opportunistic seabird. Mar Biol 157:2453–2459
Nevoux M, Weimerskirch H, Barbraud C (2007) Environmental vari-ation and experience-related differences in the demography of the long-lived black-browed albatross. J Anim Ecol 76:159–167
Nol E, Smith JNM (1987) Effects of age and breeding experience on seasonal reproductive success in the song sparrow. J Anim Ecol 56:301–313
Osborn TJ (2011) Winter 2009/2010 temperatures and a record-break-ing North Atlantic Oscillation index. Weather 66:19–21
Paiva VH, Geraldes P, Ramírez I, Garthe S, Ramos JA (2010a) How area restricted search of a pelagic seabird changes while perform-ing a dual foraging strategy. Oikos 119:1423–1434
Paiva VH, Geraldes P, Ramírez I, Meirinho A, Garthe S, Ramos JA (2010b) Oceanographic characteristics of areas used by Cory’s shearwaters during short and long foraging trips in the North Atlantic. Mar Biol 157:1385–1399
Paiva VH, Xavier JC, Geraldes P, Ramírez I, Garthe S, Ramos JA (2010c) Foraging ecology of Cory’s shearwaters in different oceanic environments of the North Atlantic. Mar Ecol Prog Ser 410:257–268
Paiva VH, Geraldes P, Ramírez I, Werner AC, Garthe S, Ramos JA (2013a) Overcoming difficult times: the behavioural resilience of a marine predator when facing environmental stochasticity. Mar Ecol Prog Ser 486:277–288
Paiva VH, Geraldes P, Marques V, Rodrígues R, Garthe S, Ramos JA (2013b) Environmental variability affecting the distribution and abundance of different trophic levels of the North Atlantic food web. Mar Ecol Prog Ser 477:15–28
Pardo D, Barbraud C, Authier M, Weimerskirch H (2013) Evidence for an age-dependent influence of environmental variations on a long-lived seabird’s life-history traits. Ecology 94:208–220
Parnell AC, Inger R, Bearhop S, Jackson AL (2010) Source partition-ing using stable isotopes: coping with too much variation. PLoS One 5:e9672
Pärt T (2001) The effects of territory quality on age-dependent reproductive performance in the northern wheatear, Oenanthe oenanthe. Anim Behav 62:379–388
Péron C, Grémillet D (2013) Tracking through life stages: adult, immature and juvenile autumn migration in a long-lived seabird. PLoS One 8:e72713
Phillips RA, Xavier JC, Croxall JP (2003) Effects of satellite trans-mitters on albatrosses and petrels. Auk 120:1082–1090
Phillips RA, Bearhop S, McGill RAR, Dawson DA (2009) Stable isotopes reveal individual variation in migration strategies and habitat preferences in a suite of seabirds during the nonbreeding period. Oecologia 160:795–806
Pütz K, Cherel Y (2005) The diving behaviour of brooding king penguins (Aptenodytes patagonicus) from the Falkland Islands: variation in dive profiles and synchronous underwater swimming provide new insights into their foraging strategies. Mar Biol 147:281–290
Riotte-Lambert L, Weimerskirch H (2013) Do naive juvenile seabirds forage differently from adults? Proc R Soc B 280:20131434
Roberts JJ, Best BD, Dunn DC, Treml EA, Halpin PN (2010) Marine geospatial ecology tools: an integrated framework for ecological geoprocessing with ArcGIS, Python, R, MATLAB, and C++. Environ Model Soft 25:1197–1207
Santos AMP, Chícharo A, Dos Santos A, Moita T, Oliveira PB, Peliz Á, Ré P (2007) Physical–biological interactions in the life history of small pelagic fish in the Western Iberia Upwelling Ecosystem. Prog Oceanogr 74:192–209
Smith JNM (1981) Does high fecundity reduce survival in song spar-rows? Evolution 35:1142–1148
Steiner I, Bürgi C, Werffeli S, Dell’Omo G, Valenti P, Tröster G, Wolfer DP, Lipp H-P (2000) A GPS logger and software for anal-ysis of homing in pigeons and small mammals. Physiol Behav 71:589–596
Stenseth NC, Ottersen G, Hurrell JW, Mysterud A et al (2003) Study-ing climate effects on ecology through the use of climate indices: the North Atlantic Oscillation, El Niño Southern Oscillation and beyond. Proc R Soc B 270:2087–2096
Suryan R, Santora J, Sydeman W (2012) New approach for using remotely sensed chlorophyll a to identify seabird hotspots. Mar Ecol Prog Ser 451:213–225
Sydeman WJ, Penniman JF, Penniman TM, Pyle P, Ainley DG (1991) Breeding performance in the Western gull—effects of parental age, timing of breeding and year in relation to food availability. J Anim Ecol 60:135–149
R Core Team (2011) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Aus-tria. http://www.R-project.org/
Thiebot JB, Lescroël A, Pinaud D, Trathan PN, Bost C-A (2011) Larger foraging range but similar habitat selection in
Author's personal copy

Mar Biol
1 3
non-breeding versus breeding sub-Antarctic penguins. Antarct Sci 23:117–126
Tsoar A, Allouche O, Steinitz O, Rotem D, Kadmon R (2007) A com-parative evaluation of presence-only methods for modelling spe-cies distribution. Divers Distrib 13:397–405
Votier SC, Grecian WJ, Patrick S, Newton J (2011) Inter-colony movements, at-sea behaviour and foraging in an immature sea-bird: results from GPS-PPT tracking, radio-tracking and stable isotope analysis. Mar Biol 158:355–362
Warham J (1990) The petrels: their ecology and breeding systems. Academic Press, London
Weimerskirch H (1992) Reproductive effort in long-lived birds: age-specific patterns of condition, reproduction and survival in the wandering albatross. Oikos 64:464–473
Weimerskirch H, Gault A, Cherel Y (2005) Prey distribution and patchiness: factors in foraging success and efficiency of Wander-ing Albatrosses. Ecology 86:2611–2622
Werner AC, Paiva VH, Ramos JA (2014) On the “real estate market”, individual quality and the foraging ecology of male Cory’s shear-waters. Auk 131:265–274
Williams GC (1966) Natural selection, the costs of reproduction, and a refinement of Lack’s principle. Am Nat 100:687
Worton BJ (1989) Kernel methods for estimating the utilization distri-bution in home-range studies. Ecology 70:164–168
Zimmer I, Ropert-Coudert Y, Kato A, Ancel A, Chiaradia A (2011) Does foraging performance change with age in female little pen-guins (Eudyptula minor)? PLoS One 6(1):e16098
Author's personal copy