food habits of the indian fox (vulpes bengalensis) in kutch, gujarat, india
TRANSCRIPT

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
ORIGINAL INVESTIGATION
Food habits of the Indian fox (Vulpes bengalensis)in Kutch, Gujarat, India
Chandrima Homea,b,�, Yadvendradev V. Jhalaa
aDepartment of Animal Ecology and Conservation Biology, Wildlife Institute of India,
P.O. Box no. 18, Chandrabani, Dehradun 248001, Uttaranchal, IndiabNational Centre for Biological Sciences, GKVK Campus, Bellary Road, Bangalore 560 065, India
Received 27 February 2008; accepted 30 May 2009
Abstract
We characterized the diet of the Indian fox (Vulpes bengalensis) during the breeding season in a semi-desert region ofWestern India. Diet was estimated using scat analysis. We used Index of Relative Importance (IRI) to determine thecontribution of prey items in the diet of the Indian fox.
Indian foxes were observed to feed on a wide variety of prey items. Arthropods were the most frequently occurringprey in their diet. IRI scores were highest for the group Coleoptera and Orthoptera followed by rodents, termites,Ziziphus fruits and spiny tailed lizards (Uromastyx hardwickii). IRI scores for rodents were higher for pups, differingsignificantly from proportions present in adult diet, thus indicating that they are crucial food items for the young ones.Prey proportions in the fox diet differed between the two habitats in the study area (grassland and scrubland).
Our data suggest that the Indian fox is essentially an omnivore showing similar diet (in terms of high incidence ofarthropods) to foxes inhabiting arid and semi-arid regions. The opportunistic and generalist strategy has probablyhelped the species to survive in varied habitats across the Indian subcontinent.r 2009 Deutsche Gesellschaft fur Saugetierkunde. Published by Elsevier GmbH. All rights reserved.
Keywords: Canidae; Food habits; Generalist forager; Index of relative importance; Semi-arid
Introduction
Food utilization is an important aspect in the study ofcarnivore ecology, since trophic resources dominateseveral aspects of their biology (Macdonald 1983;Bekoff et al. 1984). Much of the data on the diet offoxes comes from studies on Red fox (Vulpes vulpes)from varied habitats (Ciampalini and Lovari 1985;Jedrzejewski and Jedrzejewska 1992; Paltridge 2002;
Contesse et al. 2004; Basouny et al. 2005). These studiessuggest that it is a generalist forager, although differingin the proportion of various food items consumedacross geographical areas. Arthropods have beenreported to constitute large proportions in the diet ofRed foxes occurring in arid and semi-arid environmentsin comparison to their diets seen at higher latitudes(Macdonald et al. 1999; Paltridge 2002; Dell’Arte andLeonardi 2005). A similar dietary spectrum has alsobeen observed for arid inhabitants like Blandford’s fox(Vulpes cana) and Ruppell’s fox (Vulpes rupelli) (Geffenet al. 1992; Lenain et al. 2004) where seasonal fruits areconsumed in copious amounts along with arthropods. Ingeneral, semi-arid and arid habitats impose challenging
ARTICLE IN PRESS
www.elsevier.de/mambio
1616-5047/$ - see front matter r 2009 Deutsche Gesellschaft fur Saugetierkunde. Published by Elsevier GmbH. All rights reserved.
doi:10.1016/j.mambio.2009.05.011 Mamm. biol. 74 (2009) 403–411
�Corresponding author. National Centre for Biological Sciences,
GKVK Campus, Bellary Road, Bangalore 560 065, India. Mobile:
+91 9632201140.
E-mail address: [email protected] (C. Home).

Author's personal copy
conditions to survival of its inhabitants, since resourceavailability varies largely owing to environmentalfluctuations (Louw and Seely 1982). As food availabilityfunctions both as a proximate and ultimate cause forvariation in reproductive output (Bronson 1989), areliable knowledge of diet during this crucial periodis of paramount interest in understanding the species’biology.
The Indian fox (Vulpes bengalensis) is the smallest(average weight is about 2.5 kg) of the five species ofcanids found in India. It is endemic to the Indiansubcontinent (Johnsingh 1986), occupying semi-arid,flat to undulating terrain, scrub and grassland habitatsthroughout India, Pakistan and lowlands of Nepal(Johnsingh and Jhala 2004; Gompper and Vanak2006). Although the Indian fox is afforded legalprotection under the Indian Wildlife Protection Act1972 (amended 2002) (Anon 2002), its survival atpresent is threatened by persistent habitat loss due tolarge-scale land use changes, particularly in WesternIndia. Being a common species, it has received negligiblescientific attention and little information exists regard-ing aspects of its ecology (Johnsingh 1978; Manakadanand Rahmani 2000; Vanak 2005; Vanak and Gompper2007).
The present study examined the diet of the Indian foxduring a crucial period, the breeding season in a semi-desert region of Kutch, Gujarat. Den use in the Indianfox is restricted to the breeding and pup-rearing season(Manakadan and Rahmani 2000). Foxes being centralplace foragers (Orions and Pearson 1979) would preferto carry heavier prey back to the dens during the puprearing season. Hence we predicted a largely mamma-lian diet in pup scats with a greater proportion of largeprey (Lindstrom 1994). This is likely to be true for theIndian fox since parents would be bringing food forpups that are still confined to the immediate surround-ings of den sites. To test this prediction we analysed thediet of pups and compared it with that of adults.Sampling two distinct habitats within the study areaallowed us to investigate dietary differences at a localscale.
Material and methods
Study area
The study was carried out from November 2004 toApril 2005 in Abdasa taluka (681270E 221560N to691160E 231310N), in the Kutch district of Gujarat,Western India. The study spanned the breeding and thepup-rearing season of Indian fox in Kutch. Matingoccurs during December-January and pups are bornduring March-April (Johnsingh and Jhala 2004).
The area is a part of the semi desert region of India(Biogeographic Zone 3B) (Rodgers et al. 2000), char-acterized by scanty and erratic rainfall (average annualprecipitation of 332mm [1980-2002]) as well as tem-perature extremes, resulting in high evapo-transpirationrates (Tripathi and Pandey 2005). Classification ofIndian forest flora by Champion and Seth (1968)describes the vegetation as Northern Tropical ThornForest (6B) further sub classified as Desert Thorn Forest(6B/C1). The area has an undulating terrain with thelow hillocks dominated by tree and shrub species such asAcacia nilotica, A. senegal, Prosopis juliflora, Salvadora
persica, S. oleoides and Euphorbia nudiflora. Thegrassland areas are dominated by Cymbopogon sp.,Chrysopogon sp., Aristida sp. and Dicanthium sp. andsmall shrubs of Ziziphus nummularia and Capparis
decidua. The study area though primarily semi-aridencompassed two major habitats-grassland and scrub-land. The study site was not within the domain of legalprotection, as a result of which uncontrolled distur-bances like grazing, poaching, quarrying and extractionof Prosopis occurred at varying magnitudes. About21 km2 was sampled in the scrubland and 30 km2 in thegrassland. These were spatially separated and comprisedthe denning territories of foxes (minimum of five pairs inthe scrubland and three pairs in the grassland). Theconfirmation of distinct pairs was based on priorinformation collected by researchers engaged in theproject titled ‘‘Threatened and Endangered Fauna ofKutch- An Integrated Approach’’, followed by personalobservations.
Estimating food habits
We determined diet of the Indian foxes by analyzingfood remains in scats (Korschgen 1980). Scats werecollected every month from the surrounds of active foxden sites to avoid confusion with similar sized scats ofjungle cat (Felis chaus), desert cat (F. sylvestris) and thecaracal (Caracal caracal). Scats of pups (confined to theimmediate surroundings of the den during the studyperiod) could be easily distinguished from those ofadults on the basis of size. Pup scats were collected fromthree den sites (belonging to three different pairs) in thescrubland and two den sites (two pairs) in the grassland.Pups were sighted in one of the three den sites inscrubland (litter size of two) and in one den site in thegrassland (litter size of three).
Scats were sun-dried in the field and stored inpolythene ziplock bags with wire fold over closures(Custom Poly Packaging, USA). For each scat, infor-mation regarding date of collection and den identity wasrecorded. In the laboratory, scats were oven dried at60 1C for 48 h and then carefully teased apart usingforceps and a needle to separate the indigestible
ARTICLE IN PRESSC. Home, Y.V. Jhala / Mamm. biol. 74 (2009) 403–411404
Author's personal copyARTIC
LEIN
PRES
S
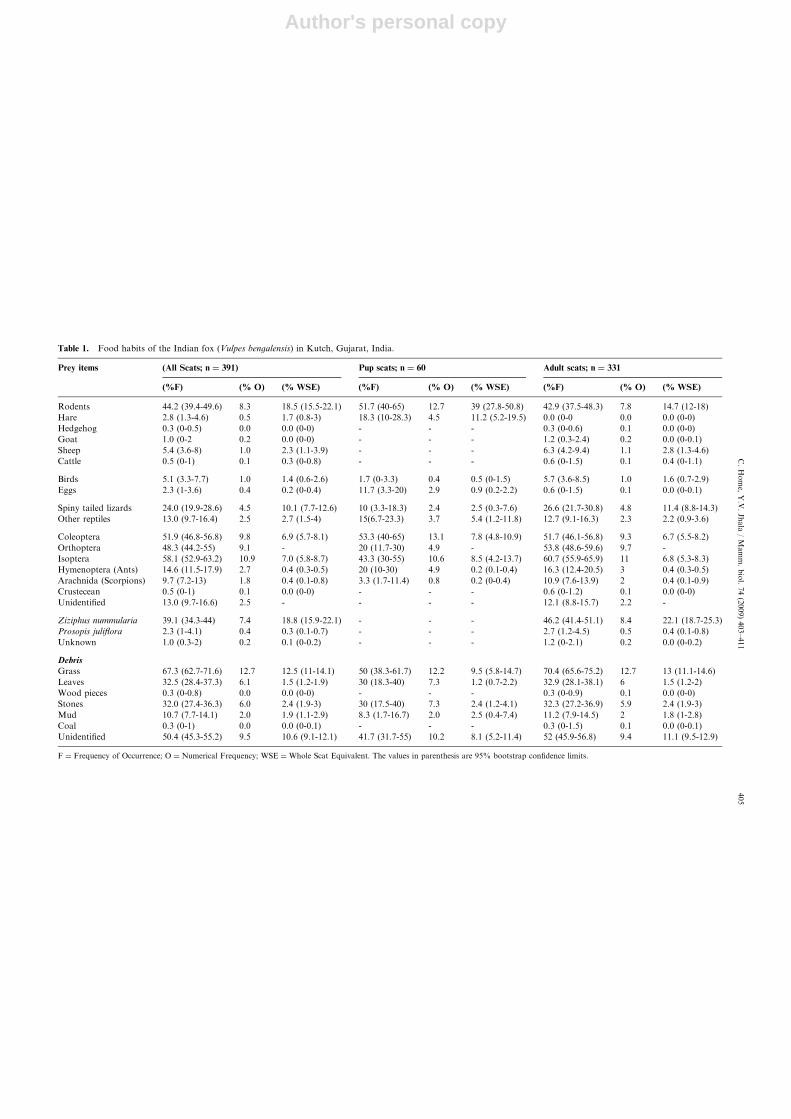
Table 1. Food habits of the Indian fox (Vulpes bengalensis) in Kutch, Gujarat, India.
Prey items (All Scats; n ¼ 391) Pup scats; n ¼ 60 Adult scats; n ¼ 331
(%F) (% O) (% WSE) (%F) (% O) (% WSE) (%F) (% O) (% WSE)
Rodents 44.2 (39.4-49.6) 8.3 18.5 (15.5-22.1) 51.7 (40-65) 12.7 39 (27.8-50.8) 42.9 (37.5-48.3) 7.8 14.7 (12-18)
Hare 2.8 (1.3-4.6) 0.5 1.7 (0.8-3) 18.3 (10-28.3) 4.5 11.2 (5.2-19.5) 0.0 (0-0 0.0 0.0 (0-0)
Hedgehog 0.3 (0-0.5) 0.0 0.0 (0-0) - - - 0.3 (0-0.6) 0.1 0.0 (0-0)
Goat 1.0 (0-2 0.2 0.0 (0-0) - - - 1.2 (0.3-2.4) 0.2 0.0 (0-0.1)
Sheep 5.4 (3.6-8) 1.0 2.3 (1.1-3.9) - - - 6.3 (4.2-9.4) 1.1 2.8 (1.3-4.6)
Cattle 0.5 (0-1) 0.1 0.3 (0-0.8) - - - 0.6 (0-1.5) 0.1 0.4 (0-1.1)
Birds 5.1 (3.3-7.7) 1.0 1.4 (0.6-2.6) 1.7 (0-3.3) 0.4 0.5 (0-1.5) 5.7 (3.6-8.5) 1.0 1.6 (0.7-2.9)
Eggs 2.3 (1-3.6) 0.4 0.2 (0-0.4) 11.7 (3.3-20) 2.9 0.9 (0.2-2.2) 0.6 (0-1.5) 0.1 0.0 (0-0.1)
Spiny tailed lizards 24.0 (19.9-28.6) 4.5 10.1 (7.7-12.6) 10 (3.3-18.3) 2.4 2.5 (0.3-7.6) 26.6 (21.7-30.8) 4.8 11.4 (8.8-14.3)
Other reptiles 13.0 (9.7-16.4) 2.5 2.7 (1.5-4) 15(6.7-23.3) 3.7 5.4 (1.2-11.8) 12.7 (9.1-16.3) 2.3 2.2 (0.9-3.6)
Coleoptera 51.9 (46.8-56.8) 9.8 6.9 (5.7-8.1) 53.3 (40-65) 13.1 7.8 (4.8-10.9) 51.7 (46.1-56.8) 9.3 6.7 (5.5-8.2)
Orthoptera 48.3 (44.2-55) 9.1 - 20 (11.7-30) 4.9 - 53.8 (48.6-59.6) 9.7 -
Isoptera 58.1 (52.9-63.2) 10.9 7.0 (5.8-8.7) 43.3 (30-55) 10.6 8.5 (4.2-13.7) 60.7 (55.9-65.9) 11 6.8 (5.3-8.3)
Hymenoptera (Ants) 14.6 (11.5-17.9) 2.7 0.4 (0.3-0.5) 20 (10-30) 4.9 0.2 (0.1-0.4) 16.3 (12.4-20.5) 3 0.4 (0.3-0.5)
Arachnida (Scorpions) 9.7 (7.2-13) 1.8 0.4 (0.1-0.8) 3.3 (1.7-11.4) 0.8 0.2 (0-0.4) 10.9 (7.6-13.9) 2 0.4 (0.1-0.9)
Crustecean 0.5 (0-1) 0.1 0.0 (0-0) - - - 0.6 (0-1.2) 0.1 0.0 (0-0)
Unidentified 13.0 (9.7-16.6) 2.5 - - - - 12.1 (8.8-15.7) 2.2 -
Ziziphus nummularia 39.1 (34.3-44) 7.4 18.8 (15.9-22.1) - - - 46.2 (41.4-51.1) 8.4 22.1 (18.7-25.3)
Prosopis juliflora 2.3 (1-4.1) 0.4 0.3 (0.1-0.7) - - - 2.7 (1.2-4.5) 0.5 0.4 (0.1-0.8)
Unknown 1.0 (0.3-2) 0.2 0.1 (0-0.2) - - - 1.2 (0-2.1) 0.2 0.0 (0-0.2)
DebrisGrass 67.3 (62.7-71.6) 12.7 12.5 (11-14.1) 50 (38.3-61.7) 12.2 9.5 (5.8-14.7) 70.4 (65.6-75.2) 12.7 13 (11.1-14.6)
Leaves 32.5 (28.4-37.3) 6.1 1.5 (1.2-1.9) 30 (18.3-40) 7.3 1.2 (0.7-2.2) 32.9 (28.1-38.1) 6 1.5 (1.2-2)
Wood pieces 0.3 (0-0.8) 0.0 0.0 (0-0) - - - 0.3 (0-0.9) 0.1 0.0 (0-0)
Stones 32.0 (27.4-36.3) 6.0 2.4 (1.9-3) 30 (17.5-40) 7.3 2.4 (1.2-4.1) 32.3 (27.2-36.9) 5.9 2.4 (1.9-3)
Mud 10.7 (7.7-14.1) 2.0 1.9 (1.1-2.9) 8.3 (1.7-16.7) 2.0 2.5 (0.4-7.4) 11.2 (7.9-14.5) 2 1.8 (1-2.8)
Coal 0.3 (0-1) 0.0 0.0 (0-0.1) - - - 0.3 (0-1.5) 0.1 0.0 (0-0.1)
Unidentified 50.4 (45.3-55.2) 9.5 10.6 (9.1-12.1) 41.7 (31.7-55) 10.2 8.1 (5.2-11.4) 52 (45.9-56.8) 9.4 11.1 (9.5-12.9)
F ¼ Frequency of Occurrence; O ¼ Numerical Frequency; WSE ¼Whole Scat Equivalent. The values in parenthesis are 95% bootstrap confidence limits.
C.Home,
Y.V.Jhala
/Mamm.biol.74(2009)403–411
405

Author's personal copy
components such as fruit seeds, hairs, claws, scales,feathers, bones and insect chitin. Seeds were identifiedby comparing them to known samples and othervegetable matter collected during field surveys. Inverte-brates were identified to the nearest order throughcomparison of exoskeleton fragments (legs, elytra andcapsule heads) with the reference collection. Identifica-tion of mammals was based on cuticular and medullarycharacteristics of fur excreted in the scats (Litvaitis et al.1996). Rodents were considered as one broad taxa sinceindividual species identification was not possible.
The best illustration of the food habits of a species isrendered by a combination of both frequency-based andvolumetric-based methods (Zabala and Zuberogoitia2003) since each method used alone has its own biases.In the current study, as there was no conversion factoravailable for computing prey biomass from scats, weassumed that the proportion of remains of a prey speciesin the scat gives a fair representation of its proportion inthe diet. We estimated the minimum number of scatsrequired to give a representation of diet composition byplotting the proportion of prey occurrences calculatedfor cumulative increments of ten scats against the totalnumber of scats. The cumulative number of scats atwhich the proportion of prey categories stabilizedindicates the approximate number of scats required toaccurately depict the food habits (Jethva and Jhala2003).
The diet of the Indian fox was estimated by usingthree different methods based on both frequencyand volumetric analysis; viz. frequency of occurrence(expressed as percentage of the total number of scats),numerical frequency (expressed as percentage of thetotal number of occurrences of all food items) (Corbett1989; Reynolds and Aebischer 1991; Paltridge 2002) and‘whole scat equivalents’ (WSE was derived as percentdry volume of prey in scats through visual estimation)(Angerbjorn et al. 1999, Elmhagen et al. 2000).Coleoptera and Orthoptera remains in scats were largelyfragmented and hence a combined estimate was madevisually. We subjected the results of the scat analysis tore-sampling with bootstrapping using SIMSTAT 2.0(Peladeau 2000). Sub-samples equalling the originalsample size of scats for each prey item were iterated 1000times, to generate means and bias-corrected 95%confidence intervals for frequency of occurrence andpercent WSE (Krebs 1989).
In order to minimise the individual biases of the threemethods used to estimate the diet of the Indian fox, weused Index of Relative Importance (IRI) (Pinkas et al.1971; Short et al. 1999; Paltridge 2002; Medina et al.2008) to determine the importance of different preyitems in the diet of the Indian fox [where IRI ¼(Numerical Frequency+Whole Scat Equivalent) �Frequency of occurrence]. The IRI scores for thedifferent prey items obtained were also subjected to re-
sampling with 1000 iterations using R 2.7.1 (R Devel-opment Core Team 2008) to compare the diet of adultsand pups. For the comparison of diet between the twohabitats, we considered the diet of adult animals sincethey would be actively foraging.
Results
Diet composition
We collected 391 scats (193 from grassland and 198from scrubland inclusive of fox pup scats) from activedens (mean ¼ seven occasions per month) and identified20 different prey items (Table 1). Ninety percent of scatscontained more than one food type (mean number offood types per scat: 3.3771.55 SD). Proportion of preyoccurrences for the five principal food categories(mammals, reptiles, avian remains, arthropods andfruits) versus cumulative number of scats analyzed,stabilized between 100 and110 scats suggesting ade-quacy of sampling.
Arthropods contributed the major proportion of foxdiet, (both Frequency of Occurrence and NumericalFrequency), followed by rodents, fruits (mainlyZiziphus) and reptiles. However, in terms of percentWSE, both rodents and fruits comprised 19% each ofthe whole scats followed by arthropods (15%) andreptiles (13%) (Table 1). Overall IRI scores were highestfor the group Coleoptera and Orthoptera followed byrodents, termites, fruits of Z. nummularia and Spiny
ARTICLE IN PRESS
Table 2. Importance of prey categories in the diet of the
Indian fox (Vulpes bengalensis) in Kutch, based on the Index of
Relative Importance (IRI).
Prey IRI
Coleoptera & Othroptera 1540.2
Rodents 1204.9
Isoptera (Termites) 1078.9
Ziziphus nummularia 1037.7
Spiny tailed lizards 356.4
Other reptiles 67.5
Hymenoptera (Ants) 47.7
Arachnids (Scorpions) 22.3
Avian remains (Feathers & Eggshells) 21.3
Sheep 18.3
Hare 6.4
Prosopis juliflora 1.8
Other Fruits 0.3
Goat 0.2
Cattle 0.2
Crustacean 0.1
IRI scores of prey items in the diet of the Indian fox (n ¼ 391).
IRI (Index of Relative Importance) ¼ (Numerical Frequency +Vo-
lume Frequency) � Frequency of Occurrence (Pinkas et al. 1971).
C. Home, Y.V. Jhala / Mamm. biol. 74 (2009) 403–411406
Author's personal copy
tailed lizards (Uromastyx hardwickii) (Table 2).Although grass was of frequent occurrence in scats, itwas assumed that most of the ingestion was incidental,and hence it was included as non-dietary item.
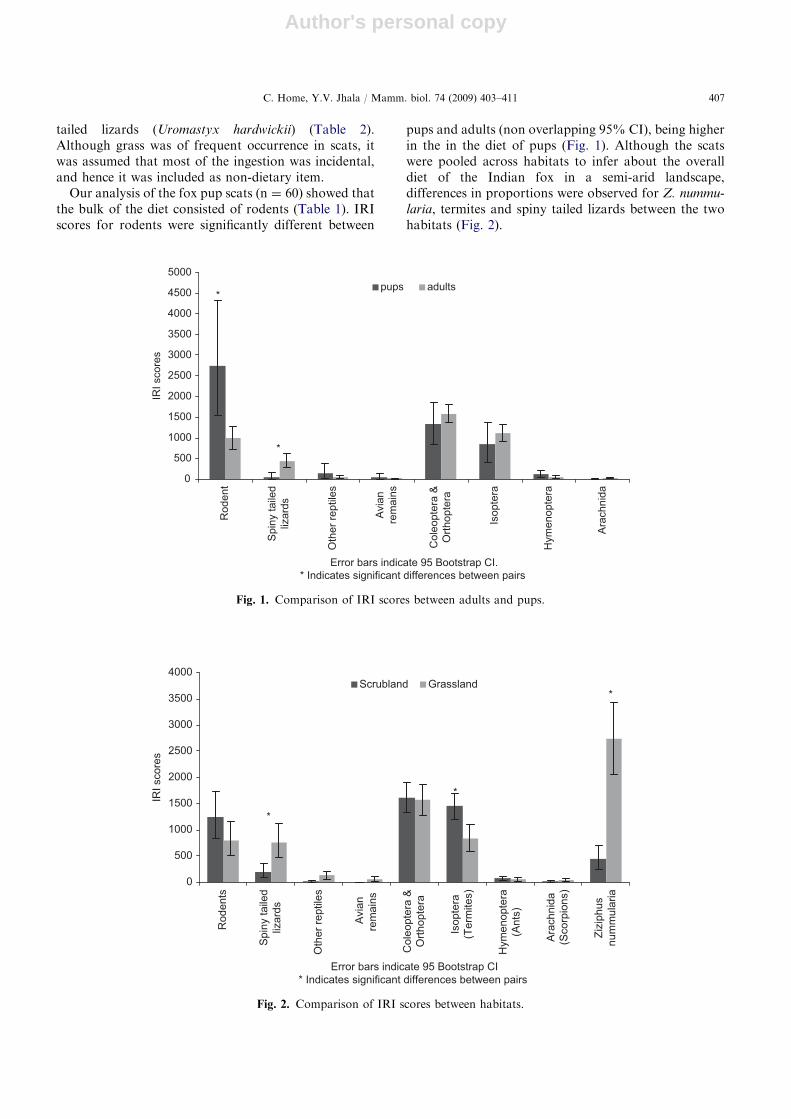
Our analysis of the fox pup scats (n ¼ 60) showed thatthe bulk of the diet consisted of rodents (Table 1). IRIscores for rodents were significantly different between
pups and adults (non overlapping 95% CI), being higherin the in the diet of pups (Fig. 1). Although the scatswere pooled across habitats to infer about the overalldiet of the Indian fox in a semi-arid landscape,differences in proportions were observed for Z. nummu-
laria, termites and spiny tailed lizards between the twohabitats (Fig. 2).
ARTICLE IN PRESS
0
500
1000
1500
2000
2500
3000
3500
4000
4500
5000
Rod
ent
Spi
ny ta
iled
lizar
ds
Oth
er re
ptile
s
Avi
anre
mai
ns
Col
eopt
era
&O
rthop
tera
Isop
tera
Hym
enop
tera
Ara
chni
da
IRI s
core
s
pups adults
Error bars indicate 95 Bootstrap CI.* Indicates significant differences between pairs
*
*
Fig. 1. Comparison of IRI scores between adults and pups.
0
500
1000
1500
2000
2500
3000
3500
4000
Rod
ents
Spi
ny ta
iled
lizar
ds
Oth
er re
ptile
s
Avi
anre
mai
ns
Col
eopt
era
&O
rthop
tera
Isop
tera
(Ter
mite
s)
Hym
enop
tera
(Ant
s)
Ara
chni
da(S
corp
ions
)
Zizi
phus
num
mul
aria
IRI s
core
s
Scrubland Grassland
Error bars indicate 95 Bootstrap CI* Indicates significant differences between pairs
*
*
*
Fig. 2. Comparison of IRI scores between habitats.
C. Home, Y.V. Jhala / Mamm. biol. 74 (2009) 403–411 407

Author's personal copy
Discussion
The Indian fox is essentially an omnivore with a widerange of food items signifying its adaptability in thissemi-arid habitat of Kutch. Much of the diet consistedof small sized prey (o0.25 kg). Arthropods were themost frequent prey item in the diet of the Indian fox,being encountered in 84% of the scats analysed.Amongst arthropods, IRI score was highest for thegroup Coleoptera and Orthoptera comparable withfrequent incidence of insect remains seen in the dietsof foxes inhabiting arid habitats elsewhere (Geffen et al.1992; Lenain et al. 2004; Dell’Arte and Leonardi 2005).Studies have indicated that arthropods have a higherproportion of water per unit nitrogen as well as a higherproportion of fat per gram of body mass thanvertebrates (Konecny 1987). This means that a highproportion in the diet would help in harnessing essentialnutrients and much required water in a semi-aridenvironment. Most termite species in Kutch hadsubterranean termitaria (pers. obs) and the fact thatthey were found frequently in scats (%F ¼ 58 and thirdhighest IRI score) indicate that the Indian foxesefficiently tapped these concentrated nutritive sources.Small mammals are high energy food resources(Mukherjee et al. 2004) and also a crucial source ofpreformed water for numerous small carnivores inhabit-ing arid habitats (Williams et al. 2002). In the presentstudy, rodents (scored second in terms of their relativeimportance) constituted 83% of the total mammaliandiet of the Indian fox.
Consumption of alternative food sources such asfruits, which are associated with low foraging costs, ischaracteristic of generalist predators (Lucherini et al.1995; Martinoli et al. 2001). Fruits of Z. nummularia
form an important seasonal resource in Kutch withfruiting occurring between November and January. Z.
nummularia comprised the bulk of the fruit diet in theIndian fox (92% of overall fruits) being represented in39% of the scats with an average of 10 seeds/scat. HighIRI scores of Z. nummularia indicate that the Indian foxactively exploits such sessile seasonal resources, andtherefore could be playing an important role in seeddispersal (Herrera 1989; Silva et al. 2005) in this semi-desert region of Kutch. We also found pods of Prosopis
juliflora in scats though it was present in minimalamounts (%F ¼ 2.3).
Spiny tailed lizards (U. hardwickii) dominated thereptilian diet in the Indian fox. Uromastyx spp. havealso been recorded in the scats of Ruppell’s foxes andRed foxes in Saudi Arabia particularly during winter, asthe foxes are able to dig them out from their hibernatingburrows (Macdonald et al. 1999; Lenain et al. 2004).During hibernation, Uromastyx have considerable fatcontent (Afroz et al. 1979) which must be important forthe Indian fox in the breeding season. Other reptiles
occurring in the diet of the Indian fox, included snakes(mainly Saw-scaled vipers Echis carinatus, Red sand boaEryx johnii and Common sand boa Eryx conicus) andlizards such as geckos, skinks and short tailed agamids(Brachysaura minor). Avian remains (birds and eggs)contributed much less to the diet of the Indian fox interms of percent frequency in scats (5.1% & 2.3%) aswell as WSE (1.4% & 0.2%). However, the contributionof ground nesting birds is likely to increase duringmonsoon when most of the birds breed in this semi-aridregion. Presence of livestock (sheep, goat and cattle) inscats suggests scavenging of carcasses that had diednaturally or were predated by wolves (Canis lupus
pallipes) or jackals (C. aureus).A comparison of the diet between adults and pups
emphasized the importance of rodents in the diet ofpups. IRI score of rodents for pups was significantlyhigher than that of adults. This finding can be comparedto that of Johnsingh (1978) and Manakadan andRahmani (2000), where they also reported the solepresence of rodent fur in pup excrements. In our study,hare remains (Lepus nigricollis ruficaudata) were foundexclusively in pup scats. Insects such as beetles, grass-hoppers and termites also frequently occurred in pupscats, implying that they might be honing their foragingskills in the vicinity of dens, as the adults are unlikely toregurgitate or bring such food items to the lair. A similarkind of mixed diet in pups has also been reported forRuppell’s fox in Saudi Arabia (Lenain et al. 2004). Wedid not record any Ziziphus fruits in pup scats. Thus, theIndian fox behaves as a typical central place forager,with fruits being consumed on the spot and rodents andhares being brought to the den for the pups. Priorstudies have shown the presence of large sized prey inscats of fox pups (Lindstrom 1994); however, in ourstudy, larger sized prey mainly- hare, spiny tailed lizardsand birds occurred in much lower proportions thanrodents, differing from what we had predicted.
Small-scale variation in diet not only gives an insightinto food availability in habitats where resourcesfluctuate, but also on how foxes utilize these foodresources. Local dietary differentiation is largely sug-gestive of differential availability of prey attributed toenvironmental heterogeneity by natural (Dalerum andAngerbjorn 2000; Dell’Arte et al. 2007) or human-induced alterations (Farias and Kittlein 2008). In thecurrent study, we observed a variation in the propor-tions of certain prey in the diets of foxes in grasslandand scrubland. IRI scores were significantly different forZ. nummularia fruits and spiny tailed lizards, which wereconsumed in higher proportions in grassland, andtermites that were consumed more in scrubland.
Preliminary data collected on the encounter ratesof certain prey species (spiny tailed lizard burrows,rodent burrows and Ziziphus shrubs) supports thedifferential availability of food items in the two habitats
ARTICLE IN PRESSC. Home, Y.V. Jhala / Mamm. biol. 74 (2009) 403–411408

Author's personal copy
(Home 2005). Encounter rates were higher for spinytailed lizard burrows (5.2 burrows/km) and Ziziphus
shrubs (19.4 shrubs/km) in grassland as compared tothat of scrubland (2.6 burrows/km and 1.1 shrubs/km).Although more rodent burrows were encountered in thescrubland (46.2 burrows/km) compared to the grassland(9.8 burrows/km), IRI scores for rodents in the foxdiet showed no significant difference between habitats.This information should be treated with a certaindegree of caution, since in this study rodents wereclassified as broad taxa due to lack of genus levelidentification. There are about 15 species of rodents thatare known to occur in Kutch (Prakash 1992; Menon2003) and no prior information exists for the rodentcommunities occurring in the study area. Except for theburrows of the Indian desert jird (Meriones hurrianae),which can be easily identified from their expansivecolonies, other burrows may not be indicative of thespecies, and can hence be used only as an index ofrodent presence in a habitat. Livestock remains werepresent only in the scats collected from grassland.Nonetheless, it cannot be inferred that this foodresource, largely determined by anthropogenic influence,occurred in only one habitat. Both the habitats weresubject to grazing and it is probably by chance thatthe samples captured the scavenging events in only onehabitat.
This is essentially the first intensive study on the dietof the Indian fox in a semi-arid landscape. Earlierstudies (Johnsingh 1978; Manakadan and Rahmani2000) have emphasized the generalist strategy ofthe species in terms of food procurement. Our dataprovides support to these studies through a much moredetailed investigation of their food habits. The dietaryspectrum of the Indian fox can also be compared tothat of foxes that inhabit other arid and semi-aridlandscapes where arthropods contribute significantlyin terms of prey proportions (Geffen et al. 1992;Macdonald et al. 1999; Lenain et al. 2004). Arthropodsare an extremely successful group of arid inhabitants(Edney 1967), and their temporal activity patternbeing synchronous with that of the Indian fox mightincrease their availability resulting in high proportionsin the diet of the predator. Populations of Indian foxhave been reported to have wide local fluctuations(Johnsingh and Jhala 2004) which can be attributed tochanges in prey populations in response to rainfall(Dickman et al. 1999; Shenbrot and Krasnov 2001).In a semi-arid landscape like Kutch, where resourcefluctuations would largely be regulating predatorpopulations and subsequently their dietary spectrum,an opportunistic strategy would definitely favour thespecies. Thus being a generalist predator subsisting onmany prey groups, the Indian fox has been able toexploit a wider niche as well as survive in variedhabitats.
Acknowledgements
We would like to thank the Director, WildlifeInstitute of India for logistic support under the auspicesof the ongoing WII grant-in aid project (Threatened andEndangered Fauna of Kutch: An Integrated Approach),the Gujarat Forest Department (Kutch Circle) for theirhelp and cooperation and the field crew at Tera FieldStation. Special thanks go to Qamar Qureshi andKulbhushansingh Suryavanshi for their assistance indata analysis. We thank Shomita Mukherjee, NimaManjrekar, Brian Cypher, Upamanyu Hore, RishiKumar Sharma, M.O.Anand, Vidyadhar Atkore, VidyaAthreya, Hari Sridhar, Amit Kurien, Umesh Srinivasan,Raman Kumar and Bibek Yumnam for their valuableinputs during manuscript preparation. We also thankthe anonymous referees who reviewed the manuscriptfor their critical comments. C.H. would personally liketo thank I.P. Bopanna, Paulamee Sinha for assistance infield work, Vinod Thakur, Ishan Agarwal and Upama-nyu Hore for facilitating identification of fox prey andNature Conservation Foundation for infrastructuralsupport during manuscript preparation.
References
Afroz, H., Ishaq, M., Ali, S.S., 1979. Seasonal changes in the
plasma lipid distribution of hibernating lizard Uromastyx
hardwickii. Pak. J. Sci. Ind. Res. 22 (3), 138–142.
Angerbjorn, A., Tannerfeldt, M., Erlinge, S., 1999. Predator
prey relationships: arctic foxes and lemmings. J. Anim.
Ecol. 68, 34–49.
Anon, 2002. The Wildlife (Protection) Act, 1972 (with
amendments). In: Wildlife Protection Society of India
(Eds.), A Hand Guide with Case Laws and Commentaries.
Natraj Publishers, India.
Basouny, M., Saleh, M., Riad, A., Fathy, W., 2005. Food
composition and feeding ecology of the Red Fox Vulpes
vulpes (Linnaeus 1758) in Egypt. Egypt. J. Biol. 7, 96–102.
Bekoff, M., Daniels, T.J., Gittleman, J.L., 1984. Life history
patterns and comparative social ecology of carnivores.
Ann. Rev. Ecol. Sys. 15, 191–232.
Bronson, F.H., 1989. Mammalian Reproductive Biology.
Chicago University Press, Chicago.
Champion, H.G., Seth, S.K., 1968. The Forest Types of India.
The Manager of Publications, New Delhi.
Ciampalini, B., Lovari, S., 1985. Food habits and trophic
niche overlap of the badger (Meles meles L.) and the red fox
(Vulpes vulpes L.) in a Mediterranean coastal area. Mamm.
Biol. 50, 226–234.
Contesse, P., Hegglin, D., Gloor, S., Bontadina, F., Deplazes,
P., 2004. The diet of urban foxes (Vulpes vulpes) and the
availability of anthropogenic food in the city of Zurich,
Switzerland. Mamm. Biol. 69 (2), 81–95.
Corbett, L.K., 1989. Assessing the diet of dingoes from feces:
a comparison of 3 methods. J. Wildlife Manage. 53 (2),
343–346.
ARTICLE IN PRESSC. Home, Y.V. Jhala / Mamm. biol. 74 (2009) 403–411 409

Author's personal copy
Dalerum, F., Angerbjorn, A., 2000. Arctic fox (Alopex
lagopus) diet in Karupelv Valley, East Greenland, during
a summer with low lemming density. Arctic 53 (1), 1–8.
Dell’Arte, G., Leonardi, G., 2005. Effects of habitat composi-
tion on the use of resources by the red fox in a semi arid
environment of North Africa. Acta Oecol. 28, 77–85.
Dell’Arte, G.L., Laaksonen, T., Norrdahl, K., Korpimaki, E.,2007. Variation in the diet composition of a generalist
predator, the red fox, in relation to season and density of
main prey. Acta Oecol. 31, 276–281.
Dickman, C.R., Mahon, P.S., Masters, P., Gibson, D.F., 1999.
Long-term dynamics of rodent populations in arid Aus-
tralia: the influence of rainfall. Wildl. Res. 26, 389–403.
Edney, E.B., 1967. Water balance in desert arthropods. Science
156 (3778), 1059–1066.
Elmhagen, B., Tannerfeldt, M., Verucci, P., Angerbjorn, A.,
2000. The arctic fox (Alopex lagopus): an opportunistic
specialist. J. Zool. (London) 251, 139–149.
Farias, A.A., Kittlein, M.J., 2008. Small-scale spatial varia-
bility in the diet of pampas foxes (Pseudalopex gymnocer-
cus) and human-induced changes in prey base. Ecol. Res.
23, 543–550.
Geffen, E., Hefner, R., Macdonald, D.W., Ucko, M., 1992.
Diet and foraging behaviour of Blandford’s foxes, Vulpes
cana in Israel. J. Mamm. 73 (2), 395–402.
Gompper, M.E., Vanak, A.T., 2006. Vulpes bengalensis.
Mamm. Species 795, 1–5.
Herrera, C.M., 1989. Frugivory and seed dispersal by
carnivorous mammals, and associated food characteristics,
in undisturbed Mediterranean habitats. Oikos 55, 250–262.
Home, C., 2005. Resource utilization by the Indian fox (Vulpes
bengalensis) in Kutch, Gujarat. M.S. Thesis, Saurashtra
University, Rajkot, India.
Jedrzejewski, W., Jedrzejewska, B., 1992. Foraging and diet of
the red fox Vulpes vulpes in relation to variable food
resources in Bialoweiza National Park. Ecography 15,
212–220.
Jethva, B.D., Jhala, Y.V., 2003. Sample size considerations of
food habit studies of wolves from scats. Mammalia 68 (4),
589–591.
Johnsingh, A.J.T., 1978. Some aspects of the ecology and
behaviour of the Indian fox—Vulpes bengalensis. (Shaw). J.
Bombay Nat. Hist. Soc. 75, 397–405.
Johnsingh, A.J.T., 1986. Diversity and conservation of
carnivorous mammals in India. Proc. Ind. Acad. Sci.
(Anim. Sci./Plant Sci.) Suppl. 73–89.
Johnsingh, A.J.T., Jhala, Y.V., 2004. Vulpes bengalnesis (Shaw
1800). In: Sillero-Zubiri, C., et al. (Eds.), Canids: Foxes,
Wolves, Jackals and Dogs. Status Survey and Conservation
Action Plan, IUCN/SSC Canid Specialist Group. IUCN,
Gland, pp. 219–222.
Konecny, M.J., 1987. Food habits and energetics of feral
house cats in the Galapagos Islands. Oikos 50, 24–32.
Korschgen, L.K., 1980. Procedures for food habit analysis. In:
Schemnitz, S.D. (Ed.), Wildlife Management Techniques
Manual. The Wildlife Society, Bethesda, MD, pp. 113–128.
Krebs, C.J., 1989. Ecological Methodology, second ed.
Addison Wesley Longman, California.
Lenain, D.M., Olfernmann, E., Warrington, S., 2004. Ecology,
diet and behaviour of two fox species in a large, fenced
protected area in central Saudi Arabia. J. Arid. Environ.
57, 45–60.
Lindstrom, E.R., 1994. Large prey for small cubs: on crucial
resources of a boreal red fox population. Ecography 17 (1),
17–22.
Litvaitis, J.A., Kimberly, T., Anderson, E.M., 1996. Measur-
ing vertebrate use of terrestrial habitats and foods. In:
Bookhout, T.A. (Ed.), Research and Management Techni-
ques for Wildlife and Habitats. The Wildlife Society,
Bethesda, MD, pp. 254–274.
Louw, G.N., Seely, M.K., 1982. Ecology of Desert Organisms.
Longman, London, 184pp.
Lucherini, M., Lovari, S., Crema, G., 1995. Habitat use and
ranging behaviour of the red fox (Vulpes vulpes) in a
Mediterranean rural area: is shelter availability a key
factor? J. Zool. (London) 237, 57–591.
Macdonald, D.W., 1983. The ecology of carnivore social
behaviour. Nature 301, 379–384.
Macdonald, D.W., Courtenay, O., Forbes, S., Mathews, F.,
1999. The red fox (Vulpes vulpes) in Saudi Arabia: loose
knit groupings in the absence of territoriality. J. Zool.
(London) 249, 383–391.
Manakadan, R., Rahmani, A.R., 2000. Population and
ecology of the Indian fox Vulpes bengalensis at the
Rollapadu Wildlife Sanctuary, Andhra Pradesh, India. J.
Bombay Nat. Hist. Soc. 97, 3–14.
Martinoli, A., Preatoni, D.G., Chiarenzi, B., Wauters, L.A.,
Tosi, G., 2001. Diet of stoats (Mustela erminea) in an
Alpine habitat: the importance of fruit consumption in
summer. Acta Oecol. 22, 45–53.
Medina, F.M., Lopez-Darias, M., Nogales, M., Garchia, R.,
2008. Food habits of feral cats (Felis sylvestris catus L) in
insular semiarid environments. Wildl. Res. 35, 162–169.
Menon, V., 2003. A Field Guide to Indian Mammals. Dorling
Kindersley, Delhi.
Mukherjee, S., Goyal, S.P., Johnsingh, A.J.T., Leite Pitman,
M.R.P., 2004. The importance of rodents in the diet of
jungle cat (Felis chaus), caracal (Caracal caracal) and
golden jackal (Canis aureus) in Sariska Tiger Reserve,
Rajasthan, India. J. Zool. (London) 262, 405–411.
Orions, G.H., Pearson, N.E., 1979. On the theory of central
place foraging. In: Horn, D.J., Stairs, G.R., Mitchell, R.D.
(Eds.), Analysis of Ecological Systems. Ohio State Uni-
versity Press, Columbus, pp. 155–177.
Paltridge, R., 2002. The diets of cats, foxes and dingoes in
relation to prey availability in the Tanami Desert, Northern
Territory. Wildl. Res. 29, 389–403.
Peladeau, N., 2000. Simstat v 2.0 (software). Provalis
Research, Montreal, Canada.
Pinkas, L., Oliphant, M.S., Iverson, I.L.K., 1971. Food habits
of Albacore, Bluefin Tuna and Bonito in California waters.
California Department of Fish and Game, Fish Bulletin
152, 1–83.
Prakash, I., 1992. In: Prakash, I., Ghosh, P.K. (Eds.), Rodents
in Indian Agriculture, Volume 1. Scientific Publishers,
Jodhpur.
R Development Core Team, 2008. R: A Language and
Enviroment for Statistical Computing. R Foundation for
Statistical Computing, Vienna, Austria /http://www.
R-project.orgS.
ARTICLE IN PRESSC. Home, Y.V. Jhala / Mamm. biol. 74 (2009) 403–411410

Author's personal copy
Reynolds, J.C., Aebischer, N.J., 1991. Comparison and
quantification of carnivore diet by faecal analysis: a
critique, with recommendations, based on a study of the
red fox Vulpes vulpes. Mamm. Rev. 21, 97–122.
Rodgers, W.A., Panwar, H.S., Mathur, V.B., 2000. Wildlife
Protected Area Network in India: A Review (Executive
Summary). Wildlife Institute of India, Dehradun.
Shenbrot, G., Krasnov, B., 2001. Rodents in desert environ-
ment: Is density dynamics really correlated with annual
rainfall fluctuations? In: Prakash, I. (Ed.), Ecology of
Desert Environments. Scientific Publishers (India), Jodh-
pur, pp. 405–421.
Short, J., Calver, M.C., Risbey, D.A., 1999. The impact of cats
and foxes on the small vertebrate fauna of Heirisson Prong,
Western Australia; exploring potential impact using diet
analysis. Wildl. Res. 26 (5), 621–630.
Silva, S.I., Bozinovic, F., Jaksic, F.M., 2005. Frugivory and
seed dispersal by foxes in relation to mammalian prey
abundance in a semiarid thornscrub. Aus. Ecol. 30,
739–746.
Tripathi, A.K., Pandey, U.K., 2005. Study of rainwater
harvesting potential of Zura village of Kutch District of
Gujarat. J. Hum. Ecol. 18 (1), 63–67.
Vanak, A.T., 2005. Distribution and status of the Indian fox
(Vulpes bengalensis) in southern India. Canis News 8 (1)
[online].
Vanak, A.T., Gompper, M.E., 2007. Effectiveness of non-
invasive techniques for surveying activity and habitat use of
the Indian fox in southern India. Wild. Biol. 13, 219–224.
Williams, J.B., Lenain, D., Ostrowski, S., Tieleman, B.I., Seddon,
P.J., 2002. Energy expenditure and water flux of Ruppell’sfoxes in Saudi Arabia. Physiol. Biochem. Zool. 75 (5), 479–488.
Zabala, Z., Zuberogoitia, I., 2003. Badger, Meles meles
(Mustelidae, Carnivora), diet assessed through scat analy-
sis: a comparison and critique of different methods. Folia
Zool. 52 (1), 23–30.
ARTICLE IN PRESSC. Home, Y.V. Jhala / Mamm. biol. 74 (2009) 403–411 411