focal cerebral ischemia in rats induces expression of p75 neurotrophin receptor in resistant...
TRANSCRIPT

~ Pergamon Neuroscience Vol. 84, No. 4, pp. 1113 1125, 1998
Copyright ~)) 1998 IBRO. Published by Elsevier Science Ltd Printed in Great Britain. All rights reserved
PII: S0306-4522(97)00579-4 0306~[522/98 $19.00+0.00
FOCAL CEREBRAL ISCHEMIA IN RATS INDUCES EXPRESSION OF P75 NEUROTROPHIN RECEPTOR IN
RESISTANT STRIATAL CHOLINERGIC NEURONS
Z. KOKAIA,*§ G. ANDSBERG,* A. MARTINEZ-SERRANOt~ and O. LINDVALL*
Sections of *Restorative Neurology, and tNeurobiology, Wallenberg Neuroscience Center, Lund University Hospital, Lund, Sweden
~Center of Molecular Biology Severo Ochoa, Autonomous University of Madrid - C.S.I.C., Campus Cantoblanco, Madrid, Spain
Abstract--Expression of p75 neurotrophin receptor and survival of medium-sized spiny projection neurons and cholinergic interneurons in the rat striatum were studied using immunocytochemistry at different times after transient, unilateral middle cerebral artery occlusion. Thirty minutes of middle cerebral artery occlusion caused a major loss of projection neurons, identified by their immunoreactivity to dopamine- and adenosine Y:5'-monophosphate-regulated phosphoprotein with a molecular weight of 32,000, in the lateral part of the striatum, as observed at 48 h following the insult with no further change at one week. In contrast, no reduction of the number of choline acetyltransferase-positive, cholinergic interneurons, which also expressed TrkA, was detected at either time-point. At 48 h following middle cerebral artery occlusion, expression of p75 neurotrophin receptor was observed in striatal cells which, by the use of double-label immunostaining, were identified as the cholinergic interneurons. No p75 neurotrophin receptor immunoreactivity remained in cholinergic cells after one week of reperfusion.
Based on current hypotheses regarding the function of the p75 neurotrophin receptor, the transient expression of this receptor in striatal cholinergic interneurons might contribute to their high resistance to ischemic neuronal death. However, the expression of p75 neurotrophin receptor could also be a first step in a pathway leading to apoptosis, which is inhibited after the present insult due to concomitant activation of TrkA. © 1998 IBRO. Published by Elsevier Science Ltd.
Key words: neurotrophic factor, striatum, cholinergic interneurons, immunocytochemistry, neuronal death.
The biological effects of the neurotrophins, nerve growth factor (NGF), brain-derived neurotrophic factor (BDNF), neurotrophin-3 (NT-3) and neurotrophin-4/5 (NT-4/5) are primarily mediated through high-affinity receptors belonging to the trk gene family of proto-oncogenes. 6 The Trk receptors and their ligands exhibit a high degree of selectivity: NGF interacts with TrkA, BDNF and NT-4/5 with TrkB, and NT-3 activates the TrkC receptor. In addition, all neurotrophins bind to the p75 neuro- trophin receptor (p75 NTR) with similar low affinity but with different kinetics. 7t'72 The function of p75 NTR is not completely understood. It has been proposed that p75 NTR acts to enhance Trk receptor
§To whom correspondence should be addressed. Abbrev&tions: BDNF, brain-derived neurotrophic factor;
CHAT, choline acetyltransferase; CNTF, ciliary neuro- trophic factor; DARPP-32, dopamine- and adenosine Y:5'-monophosphate-regulated phosphoprotein with a molecular weight of 32,000; KPBS, potassium phosphate- buffered saline; MCAO, middle cerebral artery occlusion; NeuN, neuronal specific nuclear protein; NGF, nerve growth factor; NMDA, N-methyl-D-aspartate; NT-3, neurotrophin-3; NT-4/5, neurotrophin-4/5; p75 NTR, p75 neurotrophin receptor.
responsiveness and selectivity of neurotrophin-Trk interaction. 16 Also, that p75 NTR participates in the retrograde axonal transport of neurotrophins from their target regions. 23'79 Several recent studies have indicated that p75 NTR can induce apoptosis both during normally occurring cell death and following injury, and that TrkA and p75 NTR are able to influence each other's function. 44 If p75 NTR is highly activated in the presence of low TrkA activity, TrkA signalling is suppressed and apoptosis is induced. When co-activated with TrkA, p75 NTR enhances TrkA signalling, which leads to cell differentiation and survival whereas apoptosis is inhibited. 44
Medium-sized spiny projection neurons represent more than 90% of all neurons in the striatum (for review see Ref. 42). These neurons, which project to globus pallidus and substantia nigra, are GABAergic but, in addition, they synthesize neuropeptides including enkephalin, substance P and dynor- phin. 35'38'69 Cholinergic interneurons constitute a much smaller cell population (1.7% of all striatal neurons), 68 and are believed to act by integrating synaptic inputs over large areas of the striatum. 45 Striatal cholinergic interneurons express both TrkA and p75 NTR during development, whereas only
1113

1114 Z. Kokaia et al.
TrkA is found in detectable amounts in the adult rat striatum. 74,s3
Striatal projection neurons and cholinergic inter- neurons seem to exhibit different vulnerability to brain insults. The cholinergic neurons have been reported to be more resistant to excitotoxin-induced cell death 9'1a'25'3°'31 and to the degeneration pro-
cess in Hunt ington 's disease 61 as compared to the GABAergic projection neurons. Furthermore, Goto et al. 37 described that following transient focal cer- ebral ischemia, which caused a marked reduction in the number of medium-sized spiny neurons, the dis- tribution, frequency and morphology of cholinergic cells in the damaged striatum appeared normal. No quantitative analysis was performed. In this context, it is of interest that damage to the adult striatum has been reported to induce expression of p75 NTR in cholinergic interneurons, 34 although this finding is controversial. 8 If brain insults give rise to p75 N-rR expression in selectively spared cholinergic inter- neurons in the striatum, this implies that a high level of p75 NTR activity may contribute to the low vulner- ability of these neurons. However, this hypothesis is not supported by other data which suggest that p75 NTa, at least during development, mediates apop- tosis of striatal cholinergic neurons with low TrkA expression. Thus, in p75NTR-deficient transgenic mice, the number of striatal cholinergic neurons was found to be 23% higher as compared to control strain. 77
In the present study immunocytochemical meth- ods, in some cases in combinat ion with stereology, have been used to quantify death of projection neurons and cholinergic interneurons as well as ex- pression of p75 NTR in the striatum of rats subjected to 30 min of focal cerebral ischemia, produced by middle cerebral artery occlusion (MCAO). This ischemic insult has been reported to lead to neuronal death mainly confined to the lateral striatum. 6° Recent data indicate that focal ischemia leads to both apoptot ic and necrotic neuronal death.17"28'53'54
Necrosis predominates in the ischemic core, whereas apoptosis is primarily occurring in the penumbra area. iv Which type of neuronal death predominates probably depends on the severity of the insul t] 3 The specific objectives of this study were two-fold: first, to explore the possibility that the vulnerability of striatal cholinergic interneurons to an ischemic insult differs from that of GABAergic projection neurons. Second, to determine if transient focal ischemia leads to altered expression of p75 NTR in striatal cholinergic interneurons.
EXPERIMENTAL PROCEDURES
Animals and experimental design
Thirteen adult male Wistar rats (MMlegaard's Breeding Centre, Copenhagen, Denmark) weighing 295-320 g were used. The rats were housed under 12h light/12h dark conditions with ad libitum access to food and water. Prior to MCAO, the animals were fasted overnight, but allowed free
access to water. All animals were subjected to unilateral MCAO for 30 min and were then killed either at 48 h (n=7) or at one week (n=6) after the start of reperfusion. Brains were sectioned and processed for immunocytochemical staining using specific antibodies against dopamine- and adenosine 3':5'-monophosphate-regulated phosphoprotein with a molecular weight of 32,000 (DARPP-32, marker for GABAergic projection neurons), choline acetyltransferase (CHAT, marker for cholinergic neurons), neuronal specific nuclear protein (NeuN, general marker for postmitotic neurons), TrkA and p75 NvR. Quantification of immuno- positive neurons was performed by light microscopy, manually or using stereological procedures.
Middle cerebral artery occlusion
Anaesthesia was induced by inhalation of 3.5% halothane in N20:O 2 (70:30). Animals were intubated and then arti- ficially ventilated using a small respirator with inhalation of 1-1.5% halothane during the operation. A polyethylene catheter was inserted into the tail artery for blood sampling and blood pressure recording. Arterial blood pressure and body temperature were monitored during the entire pro- cedure, and pO2, pCO2, pH, and blood glucose concen- trations were measured just before MCAO. For the induction of focal ischemia, the middle cerebral artery was occluded by an intraluminal filament technique, as de- scribed in detail elsewhere. 48'86 In brief, the right common, internal and external carotid arteries were exposed and the external carotid artery was ligated. The common carotid artery was closed by a ligature, and the internal carotid artery was temporarily closed by a microvascular clip. A filament with a distal cylinder of silicon rubber was inserted into the internal carotid artery through the common carotid artery and advanced to close the origin of the middle cerebral artery. Thirty minutes later, the filament was withdrawn in order to allow for reperfusion. The animals were then randomly assigned to the 48 h or one week survival groups. As summarized in Table 1, physiological parameters were within the normal range at the start of the MCAO. No significant differences were observed between the two groups of rats with different reperfusion times. The efficacy of the MCAO was evaluated at 1 and 24 h after the occlusion using a modification s of the neurological assess- ment described previously] °'26 All animals showed clear neurological deficits (gait disturbances and circling or walk- ing towards the left side) and were, therefore, included for further analysis.
Immunocytochemistry
The procedures used for immunostaining have been described in detail elsewhere, s8 After 48 h or one week of reperfusion, the animals were deeply anaesthetized with chloral hydrate (400mg/kg) and transcardially perfused with 0.9% NaC1 solution followed by ice-cold 4% para- formaldehyde in 0.1 M phosphate buffer. The brains were removed, postfixed with the same fixative overnight, equili- brated in 30°/,, sucrose and then sectioned in seven to 10 series at 40 ~un thickness. Sections were stored at - 20°C in cryoprotectant solution. Immunocytochemistry was per- formed on free-floating sections in potassium phosphate- buffered saline (KPBS). Briefly, sections were rinsed and endogeneous peroxidase activity was blocked by 3% hydro- gen peroxide and 10% methanol. After pre-incubation with 5% appropriate serum in 0.25% Triton X-100 for 1 h, the sections were incubated with the primary antibody in 1% appropriate serum in 0.25% Triton X-100 overnight at room temperature (for CHAT, incubation with the primary anti- body was for 72 h at 4°C). On the next day, sections were rinsed and incubated with the appropriate biotinylated secondary antibody (from Vector, horse anti-mouse, 1:200 for DARPP-32, CHAT, NeuN and p75NTR; swine anti- rabbit, 1:200 for TrkA) in KPBS containing 0.25% Triton X-100 for 1 h. Sections were then rinsed, incubated in

p75 ~TR expression in focal ischemia
Table 1. Physiological parameters in rats subjected to middle cerebral artery occlusion
1115
Parameter
Experimental group
48 h of reperfusion (n=7) One week of reperfusion (n=6)
Blood pressure (mmHg) 121.9 4- 2.9 120.0 4- 3.7 pCO 2 (mmHg) 36.5 4- 0.9 38.0 ± 0.4 pO2 (mmHg) 110.4 4- 4.3 114.8 4- 5.2 pH 7.39 ± 0.01 7.38 4- 0.01 Blood glucose (raM) 4.2 ± 0.7 5.3 ± 0.5 Body temperature (°C) 37.1 4- 0.1 37.1 4- 0. !
Means + S.E.M. of measurements made just before insertion of the occluding filament in the middle cerebral artery. There were no significant differences in any parameter between the groups with different reperfusion times (P>0.05; Student's unpaired t-test).
avid±n-biotin complex (Elite ABC kit, Vector) for 1 h, and developed by nickel-intensified reaction with diaminobenzi- dine. The following primary antibodies were used: mouse monoclonal anti-DARPP-32 (1:20,000, a gift from Dr P. Greengard, Rockefeller University, New York, NY, U.S.A.), mouse monoclonal anti-NeuN (1:100, a gift from Dr R. J. Mullen, University of Utah, Salt Lake City, UT, U.S.A.), mouse monoclonal anti-p75 NTR (1:20, clone IgG- 192, a gift from Dr E. M. Johnson, Washington University, St Louis, WA, U.S.A.), mouse monoclonal anti-ChAT (1:1000, Chemicon), and rabbit polyclonal anti-TrkA (1:5000, a gift from Dr L. F. Reichardt, University of California, San Francisco, CA, U.S.A.).
Double-immunostainings (ChAT+p75 NTR) were first per- formed for ChAT and then for p75 NTR using the chro- mogens Vector SG for ChAT (blue precipitate) and Vector Red for p75 NvR (red precipitate). Since both primary anti- bodies are mouse monoclonal antibodies and the same horse anti-mouse secondary antibody was used in the two consecutive immunostainings, we stained some sections from animals with 48 h of re-perfusion after MCAO accord- ing to the complete protocol but omitted the primary antibody for either p75 NvR or CHAT.
Morphometric analysis
The number of DARPP-32 and NeuN-positive neurons in the striatum was quantified using stereological procedures, which allow for unbiased estimates of the number of cells within a defined brain structure by random sampling the fields of study. 39 Counting of immunopositive cells was performed at the following representative rostrocaudal levels of the striatum: 1. AP 1.7 mm; 2. AP 0.7 mm; 3. AP -0 .3mm; 4. AP - 1 . 4 m m anterior or posterior from bregma according to the atlas of Paxinos and Watson. 67 For stereological analysis, the image of the brain section was displayed on the screen of a computer using an Olympus BH-2 microscope and a CCD-IRIS color video camera, both controlled by CAST-GRID ® software (Olympus, Denmark). The border of the striatum ipsilateral to the MCAO was first marked at low magnification (x 4). Sampling and cell counting were then performed at x 40 magnification. Cell counts from the intact, contralateral side were used for comparison.
All cholinergic neurons in the striatum immunoreactive for CHAT, TrkA or p75 NTR were counted manually in the microscope at x 10 magnification at the four levels defined above.
Statistical analysis Evaluation of differences in the number of immunoposi-
tive cells between the striatum ±psi- and contralateral to MCAO was performed using one-way ANOVA followed by
Bonferroni-Dunn post hoc test. Physiological parameters were compared using Student's unpaired t-test. Significance was set at P<0.05.
RESULTS
Quantification of DARPP-32- and choline acetyl- transferase-expressing cells
Immunocytochemical detection of DARPP-32 provides an excellent tool for the demonstrat ion of nearly all medium-sized GABAergic projection neurons in the striatum. 65,66 The distribution of DARPP-32 immunoreactivity in the intact striatum, contralateral to the MCAO, agreed well with that reported previously. 65,66 Both at 48 h and one week following the ischemic insult, there was a major loss of DARPP-32 immunoreactivi ty in the lateral striatum ipsilateral to the M C A O in all coronal sections throughout the rostrocaudal extent of this structure. The medial part was clearly spared (Figs 1, 2A) except most caudally, where the lesion was affecting the entire striatum (Fig. 1). Microscopic examination of the lesioned areas showed only scattered surviving DARPP-32-posi t ive cells, in many cases close to blood vessels (Fig. 2E).
The number of DARPP-32-expressing striatal neurons was quantified using a stereological method on both the lesioned and non-lesioned side at four representative coronal levels (Fig. 1). At 48 h after the ischemic insult, there was a marked reduction (range 56 88%, mean 73% as compared to contra- lateral side; Fig. 3A) of DARPP-32-stained neurons at all levels in the striatum ipsilateral to the MCAO. In the animals killed five days later, at one week after MCAO, the loss of DARPP-32-posi t ive cells (range 49-83%; mean 68%, Fig. 3B) was very similar to that observed at 48 h (Fig. 3B). This indicates that death of DARPP-32-posi t ive projection neurons following 30 min of M C A O is complete already at the earlier time point. However, in order to explore the possi- bility that the lack of DARPP-32 staining after ischemia was due to down-regulation of DARPP-32 protein levels and not to cell death, we also per- formed Haematoxylin Eosin staining and counted

1116 z. Kokaia et al.
AP 1 .7ram
A P 0 . 7 m m
AP-0.3 m m
AP -1.4 m m
Fig. 1. Focal ischemia causes loss of striatal projection neurons. Photographs of anti-DARPP-32-stained sections taken at four representative coronal levels through the rostrocaudal extent of the striatum. Sections were obtained from a rat subjected to 30 rain of MCAO and re-perfused after one week. Asterisks denote lesioned striatum. Right column shows cartoons of the corresponding anterio-posterior (AP) levels with coordinates according to the rat brain atlas by Paxinos and Watson. 6v Filled areas on the cartoons
depict regions where cell counting was performed. Scale bar=3 mm.
striatal neurons in alternate sections stained with an antibody against the neuron specific marker NeuN. The pattern of cell loss observed in these sections closely matched that observed using DARPP-32 immunocytochemistry (data not shown). Outside the striatum, e.g., in the cerebral cortex, no obvious neuronal loss was observed.
Cholinergic cells were identified immuno- cytochemically using an antibody against the acetylcholine-synthesizing enzyme CHAT. In agree- ment with previous reports, 15 large numbers of ChAT-positive cells were found in the medial septum, vertical and horizontal limbs of the diagonal band of
Broca and in the nucleus basalis. In the intact hemi- sphere, scattered ChAT-positive cells were detected in the striatum, with higher numbers in the ventro- lateral and dorsal parts (Fig. 4C-F). Ipsilateral to the MCAO, the distribution and density of CHAT- immunoreactive cells both in the medial and lateral parts of the striatum were similar to what was observed on the contralateral side. This was in sharp contrast to the major loss of DARPP-32- (Fig. 4A, B) and NeuN-positive neurons in the lesioned area. Quantification of the number of ChAT-positive cells was performed at the same four levels of the striatum used for counting of DARPP-32-positive neurons.

p75 NvR expression in focal ischemia I117
DARPP-32 p75 NTR
Z
I Z © cd3
Fig. 2. Focal ischemia induces expression of p75 NTR in lesioned striatum. Adjacent coronal sections through the striatum of a rat subjected to 30 min of MCAO and 48 h of re-perfusion. The sections were stained with antibodies against DARPP-32 (A, C, E) and p75 NvR (B, D, F), respectively. Dashed squares in A and B indicate those parts of the intact, non-lesioned striatum which are shown at higher magnification in C and D, respectively. Similarly, open squares in A and B depict lesioned areas of the striatum enlarged in E and F, respectively. Arrows in C-F point to areas which are enlarged and presented as framed inserts on the same images. Note the high number of DARPP-32-positive cells in the intact part of striatum (C), and just few, scattered such cells in the lesioned part (E). Also note that p75NTR-positive cells are only observed in the lesioned part of the striatum (compare D and F). Scale bar= 1.8 mm for
A and B; 150/am for C F; 390 ~tm for inserts on C F.

1118 Z. Kokaia et al.
150 rA
1 0 0
"~ so
:~.
& 0
150 e~ o
100
Z
5o
o
Forty eight hours of reperfusion
T l
One week of reperfusion
AP1.7mm AP 0.7 mm AP -0.3 m m AP-L4mm
Fig. 3. Quantification of loss of striatal projection neurons after focal ischemia. Number of DARPP-32-positive neurons in the striatum ipsilateral and contralateral to the MCAO after 48 h (A) and one week (B) of reperfusion. Quantification was performed at the four representative coronal levels indicated in Fig. 1. Means± S.E.M. *P<0.05; one-way ANOVA followed by Bonferroni-Dunn post hoc
test.
No significant differences in the number of CHAT- positive neurons were observed between the intact and lesioned striatum at any level, irrespective of whether the animals were killed at 48 h or one week after the MCAO (Fig. 5A, B). In summary, there was a selective sparing of ChAT-positive, cholinergic interneurons in the striatum after 30 min of focal ischemia.
Quantification of TrkA- and p75 neurotrophin receptor-expressing cells
As described previously in immunocytochemical studies, 74"76 TrkA-positive neurons were found in the nucleus accumbens, medial septum, vertical and horizontal limbs of the diagonal band of Broca, nucleus basalis and in the thalamic paraventricular and reuniens nuclei. In the non-lesioned striatum, numerous TrkA-positive neurons were observed, with a similar distribution to that of CHAT- immunoreactive neurons. No change of either the number (Fig. 6A, B) or distribution (Fig. 4G-J) of TrkA-immunopositive cells was observed in the striatum ipsilateral to MCAO at 48 h or one week after the insult. Both in the lesioned and non-lesioned striatum, the numbers and distributional patterns of
TrkA- and ChAT-immunopositive cells were very similar (Fig. 4G-J, C F), suggesting that these cells represent the same population of cholinergic interneurons. 74,76
Large numbers of strongly p75NVR-immuno - reactive neurons were detected in the medial septal nucleus, vertical and horizontal limbs of the diagonal band of Broca and in the nucleus basalis (Fig. 2 B ) . 36 '46"83 Occasionally, very lightly stained p75 NTR- positive cells were also observed in the intact stria- tum, contralateral to the MCAO, primarily in the most ventrolateral part. At 48 h after the insult, there was a marked increase (range 6-35-fold, mean 14- fold) of the number of p75YXR-immunoreactive neurons at all four examined striatal levels ipsilateral to the MCAO (Fig. 6C). The p75YVa-positive cells were primarily located in the lateral part of the striatum, in the area of cell loss as demonstrated with DARPP-32 immunostaining (Fig. 2). In the animals subjected to one week of reperfusion, distinct striatal cells expressing high levels of p75 NTR immuno- reactivity were no longer observed (Fig. 4K-N). However, at this time-point, p75 NTR staining was profuse in blood vessels within the lesioned area of the striatum (Fig. 4M, N).
In order to identify the type of striatal neuron expressing p75 NTR after MCAO, we performed double-staining immunocytochemistry using anti- bodies against ChAT (giving blue staining) and p75 NTR (red staining) (Fig. 7). In the striatum ipsi- lateral to the MCAO, the majority of labelled cells (about 95%) in the lesioned area were double-stained with red and blue colour (Fig. 7B), indicating that almost all cholinergic neurons had started to express p75 NTR. In the area of the lesion, we could not detect any cells showing only red colour which would represent non-cholinergic neurons expressing p75 NTR. Only few cells in the striatum ipsilateral to MCAO, but all stained cells in the contralateral striatum, showed blue colour only (Fig. 7A), indi- cating that they were cholinergic neurons not ex- pressing p75 NTR. Omission of the p75 NTR antibody resulted in only blue staining of forebrain cholinergic neurons, and exclusion of the ChAT antibody gave rise to only red staining in p75NXR-immunopositive neurons, including the ones in the striatum ipsilateral to MCAO (Fig. 7C).
D I S C U S S I O N
The major findings of the present study can be summarized as follows: first, 30 min of MCAO lead to massive death of DARPP-32-positive, GABAergic projection neurons and selective sparing of CHAT- positive, cholinergic interneurons in the striatum. Second, this ischemic insult induces transient expres- sion of p75 NxR in striatal cholinergic interneurons.
In agreement with previous studies, 2"52'6° MCAO for 30 min caused damage mainly located to the lateral striatum, and very little neuronal death in the

p75 NTR expression in focal ischemia 1119
DARPP-32
ChAT
TrkA
p75 NTR
Fig. 4. Cholinergic striatal interneurons are selectively spared after focal ischemia, which induces expression of p75 NxR in neurons. Photographs of the striatum from adjacent coronal sections immuno- stained for DARPP-32 (A, B), ChAT (C F), TrkA (G-J) and p75 NvR (K-N), respectively. Sections were taken from a rat subjected to 30 min of MCAO and one week of reperfusion. B, E, F, I, J, M and N, and A, C, D, G, H, K and L are from the striatum ipsilateral and contralateral to the MCAO, respectively. C, F, G, J, K and N show enlargements of areas in D, E, H, I, L and M. Note the disappearance of DARPP-32-positive staining after MCAO (A and B), and the similar pattern and number of CHAT- and TrkA-positive cells (D and E, and H and I, respectively) on the side ipsilateral to the MCAO and on the contralateral, intact side. Also note the absence of p75NXR-positive neurons but the strong immuno- reactivity in blood vessels at one week of reperfusion (M, N). Scale bar=300 gm for A, B, D, E, H, I, L
and M; 50 ~m for C, F, G, J, K and N.
cerebral cortex. The extent of str iatal cell death, as demons t r a t ed using the D A R P P - 3 2 ant ibody, was maximal already after 48 h of reperfusion, and no fur ther change occurred up to one week. Similar da ta were ob ta ined using immunos ta in ing with the specific
neurona l marke r NeuN. Because focal ischemia in- duces inhibi t ion of prote in synthesis,~'z'2°63 it might be argued that the observed disappearance of D A R P P - 3 2 immunoreac t iv i ty was due to low levels of the DARPP-32 protein, caused by suppressed

1120 Z. Kokaia et al.
Forty eight hours of repeffusion 150 1 5 0
1 0 0 1 0 0
50 "~ 50 z
0 eL • ~ ~ 0
t~ One week of reperfusion 150 b~ 150 <
100
Z
Z
50
100
50
0 0
AP1.7 m m A P 0 . T m m A P - 0 3 m m AP-1.4 m m "~
Fig. 5. Quantification of striatal cholinergic interneurons ~ 60 after focal ischemia. Number of ChAT-positive neurons in the striatum ipsilateral to the MCAO and in the intact, contralateral striatum after 48 h (A) and one week (B) of ~ 40 reperfusion. Quantification of ChAT-positive, cholinergic "~t..
D, neurons was performed at four representative coronal levels "d 20 (see Fig. 1). Means :t: S.E.M. No significant differences ~1 between the sides were found (P>0.05; one-way ANOVA "~
followed by Bonferroni Dunn post hoc test). ~ 0
synthesis, and did not reflect neuronal death. How- ever, protein synthesis in the lateral striatum is fully recovered at one week after 30 min of MCAO, 2 whereas we observed a similar loss of DARPP-32 and NeuN immunostaining at 48 h and one week. In addition, Haematoxylin Eosin staining revealed the same pattern of striatal damage after 48 h and one week of reperfusion as that shown with DARPP-32 and NeuN immunocytochemistry. Taken together, these data strongly support the conclusion that 30min of M C A O lead to pronounced death of striatal projection neurons, and that this process is complete at least at 48 h after the insult.
In contrast to the high vulnerability of striatal projection neurons, cholinergic interneurons were found to be resistant to 30 min of focal ischemia. Previous studies have also indicated a differential sensitivity of these neuronal populations to brain insults. Cholinergic interneurons were less vulnerable to early postnatal hypoxia-ischemia in rats as com- pared to other striatal neurons. 14,43 Bilateral carotid artery occlusion for 5 min in gerbils induced massive loss of projection neurons in the dorsolateral stria- tum with sparing of cholinergic interneurons. 19 Simi- lar to the present data, Goto eta[. 37 found a marked
Forty eight hours of reperfusion
One week of reperfusion
Forty eight hours of reperfusion
AP 1.7 m m AP 0.7 m m AP .03 m m AP -1.4 m m
Fig. 6. Quantification of TrkA- and p75NXR-expressing striatal neurons after focal ischemia. Number of TrkA- (A, B) and p75NVR-positive (C) neurons in the striatum ipsilat- eral and contralateral to the MCAO after 48 h (A, C) and one week (B) of reperfusion. Quantification was performed at four representative coronal levels (see Fig. 1). Means+S.E.M. *P<0.05; one-way ANOVA followed by
Bonferroni-Dunn post hoc test.
reduction of medium-sized spiny neurons following 30 rain of MCAO in rats, whereas cholinergic in- terneurons seemed to be unaffected. Striatal cholin- ergic interneurons in the rat have also been found to be more resistant to excitotoxic lesions as compared to projection neurons, 9"11.25,30,31 and in patients with Hunt ington 's disease, cell death in the striatum pri- marily affects projection neurons while cholinergic interneurons are spared. 61 The finding that the thera- peutic window for protection against excitotoxic damage by N-methyl-D-aspartate (NMDA) receptor antagonists is much longer for striatal cholinergic interneurons also indicates a higher resistance of these neurons. 32'59
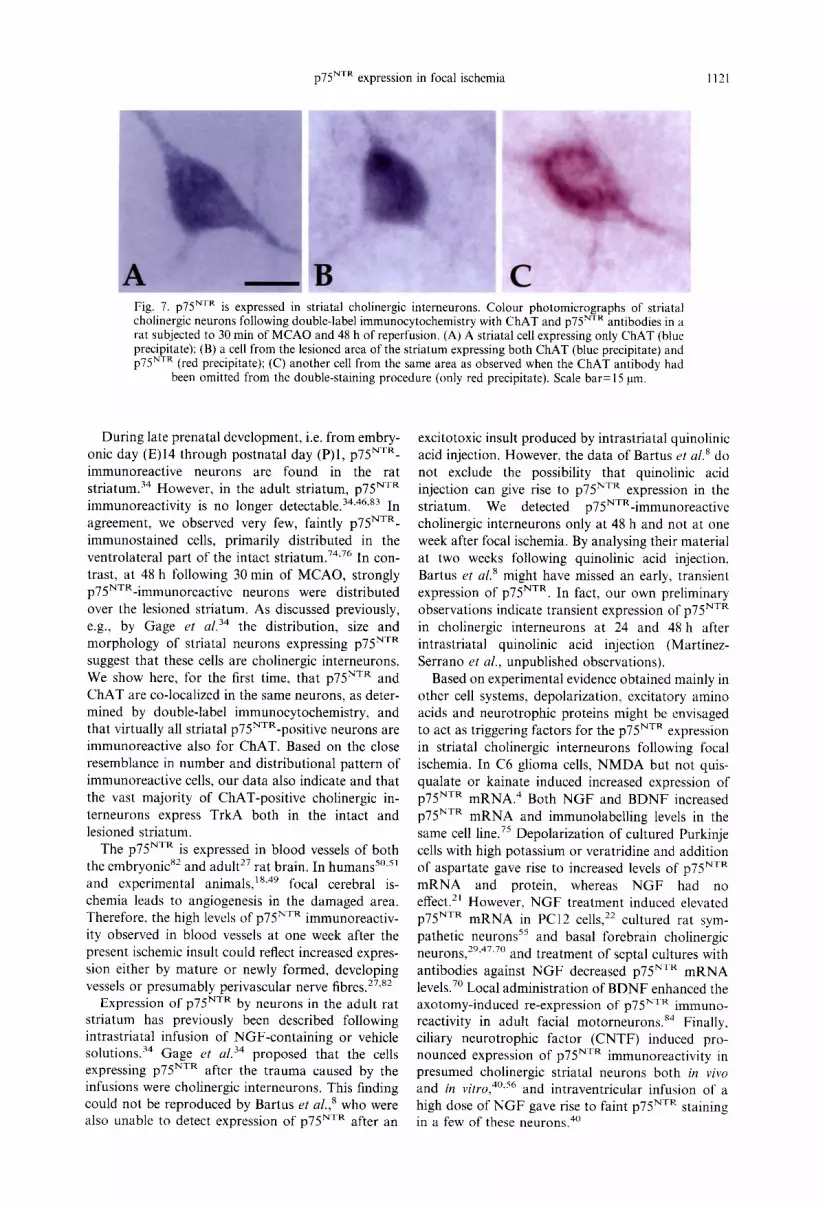
p75 NTR expression in focal ischemia 1121
Fig. 7. p75 NTR is expressed in striatal cholinergic interneurons. Colour photomicrographs of striatal cholinergic neurons following double-label immunocytochemistry with ChAT and p75 NTR antibodies in a rat subjected to 30 min of MCAO and 48 h of reperfusion. (A) A striatal cell expressing only ChAT (blue precipitate); (B) a cell from the lesioned area of the striatum expressing both ChAT (blue precipitate) and p75 NTR (red precipitate); (C) another cell from the same area as observed when the ChAT antibody had
been omitted from the double-staining procedure (only red precipitate). Scale bar= 15 gin.
During late prenatal development, i.e. from embry- onic day (E)14 through postnatal day (P)I, p75 NxR- immunoreactive neurons are found in the rat striatum. 34 However, in the adult striatum, p75 NvR immunoreactivity is no longer detectable. 34'46's3 In agreement, we observed very few, faintly p75 NvR- immunostained cells, primarily distributed in the ventrolateral part of the intact striatum. 74"76 In con- trast, at 48 h following 30 min of MCAO, strongly p75NTR-immunoreactive neurons were distributed over the lesioned striatum. As discussed previously, e.g., by Gage et al. 34 the distribution, size and morphology of striatal neurons expressing p75 NvR suggest that these cells are cholinergic interneurons. We show here, for the first time, that p75 N'rR and ChAT are co-localized in the same neurons, as deter- mined by double-label immunocytochemistry, and that virtually all striatal p75NVR-positive neurons are immunoreactive also for CHAT. Based on the close resemblance in number and distributional pattern of immunoreactive cells, our data also indicate and that the vast majority of ChAT-positive cholinergic in- terneurons express TrkA both in the intact and lesioned striatum.
The p75 NTR is expressed in blood vessels of both the embryonic s2 and adult 27 rat brain. In humans s°'5~ and experimental animals, 18'49 focal cerebral is- chemia leads to angiogenesis in the damaged area. Therefore, the high levels of p75 NTR immunoreactiv- ity observed in blood vessels at one week after the present ischemic insult could reflect increased expres- sion either by mature or newly formed, developing vessels or presumably perivascular nerve fibres. 27'82
Expression of p75 NTR by neurons in the adult rat striatum has previously been described following intrastriatal infusion of NGF-containing or vehicle solutions. 34 Gage et al. 34 proposed that the cells expressing p75 NTR after the trauma caused by the infusions were cholinergic interneurons. This finding could not be reproduced by Bartus et al., 8 who were also unable to detect expression of p75 NTR after an
excitotoxic insult produced by intrastriatal quinolinic acid injection. However, the data of Bartus et al. 8 do not exclude the possibility that quinolinic acid injection can give rise to p75 NTR expression in the striatum. We detected p75NXR-immunoreactive cholinergic interneurons only at 48 h and not at one week after focal ischemia. By analysing their material at two weeks following quinolinic acid injection, Bartus el al. 8 might have missed an early, transient expression of p75 NTR. In fact, our own preliminary observations indicate transient expression of p75 NTR in cholinergic interneurons at 24 and 48 h after intrastriatal quinolinic acid injection (Martinez- Serrano et al., unpublished observations).
Based on experimental evidence obtained mainly in other cell systems, depolarization, excitatory amino acids and neurotrophic proteins might be envisaged to act as triggering factors for the p75 NTR expression in striatal cholinergic interneurons following focal ischemia. In C6 glioma cells, NMDA but not quis- qualate or kainate induced increased expression of p75 NTR mRNA. 4 Both NGF and BDNF increased p75 NTR mRNA and immunolabelling levels in the same cell line. 75 Depolarization of cultured Purkinje cells with high potassium or veratridine and addition of aspartate gave rise to increased levels of p75 NTR mRNA and protein, whereas NGF had no effect. 2~ However, NGF treatment induced elevated p75 NrR mRNA in PC12 cells, 22 cultured rat sym- pathetic neurons 55 and basal forebrain cholinergic neurons, 29'47'7° and treatment of septal cultures with antibodies against NGF decreased p75 NTR mRNA levels. 7° Local administration of BDNF enhanced the axotomy-induced re-expression of p75 NTR immuno- reactivity in adult facial motorneurons. 84 Finally, ciliary neurotrophic factor (CNTF) induced pro- nounced expression of p75 NTR immunoreactivity in presumed cholinergic striatal neurons both in vivo
and in vitro, 4°'56 and intraventricular infusion of a high dose of NGF gave rise to faint p75 NTR staining in a few of these neurons. 4°

1122 Z. Kokaia et al.
The functional consequences of the transient ex- pression of p75 NTR in striatal cholinergic inter- neurons following MCAO remain to be elucidated. In particular, it seems highly warranted to explore whether the expression of p75 NTR plays any role for the high resistance to ischemic damage exhibited by these neurons as compared to the projection neurons. Both neuron types express the high-affinity receptors for BDNF and NT-3, i.e. TrkB and TrkC, 3'12'62'85
respectively, whereas the high-affinity N G F receptor, TrkA, is only expressed by cholinergic interneurons. Some experimental evidence indicate that p75 NTR can facilitate the dimerization of Trk receptors, thereby increasing the functional responses to the neurotrophins. 7"24'41'8° It is well established that administration of neurotrophins can promote the survival of striatal neurons both in vitro and in FIFO. 33"57"64"78"81 Hypothetically, the protective action of endogeneous neurotrophins, which are all able to interact with their specific high-affinity receptors on striatal cholinergic interneurons, might be amplified by the expression of p75 NTR, leading to the selective survival of these neurons following ischemia.
Another possibility is that the p75 NTR expression in cholinergic interneurons constitutes the first step of a death process triggered by the ischemic insult. If this is the case, the p75NXR-mediated degeneration of these neurons is efficiently prevented by some other mechanism after 30 rain of MCAO. As discussed by Kaplan and Miller, 44 several studies have indicated that apoptotic death induced by p75 yxR is sup- pressed in the presence of high levels of TrkA activity. For example, in cultures of sympathetic neurons, which express both p75 NTR and TrkA, BDNF induced apoptosis but this did not occur when TrkA was activated to higher levels. 44 Van der Zee et al. 77 described that p75NXR-positive, TrkA- negative cholinergic neurons in the medial septum die through apoptosis during postnatal development whereas cholinergic neurons expressing both p75 NTR
and TrkA survive. The apoptotic death did not occur in transgenic mice lacking p75 NTR or in control mice injected with a p75NVR-inhibiting peptide. Also, in accordance with the concept of TrkA inhibiting the apoptosis-mediating action of p75 NTR are recent data by Sauer et al. 73 They found atrophy and
hypoplasia of medial septal cholinergic neurons in mice heterozygous for a deletion of the N G F gene, probably due to low levels of NGF. Disruption of the p75 NTR gene completely prevented the atrophy and restored normal neuronal numbers in the medial septum. It is tempting to speculate that death of cholinergic interneurons following 30 rain of MCAO does not occur, despite expression of p75 NTR, due to concomitant activation of TrkA receptors on the same cells. In fact, the co-activation of p75 NTR and TrkA could promote the survival of these neurons. 44 Interestingly, Lee et al. 52 reported that striatal N G F levels decrease after focal ischemia. This implies that with further reduction of N G F levels caused, e.g., by a more severe insult, the activation of TrkA might be insufficient to suppress apoptosis induced by p75 NTR.
CONCLUSION
In conclusion, our findings demonstrate selective sparing of striatal cholinergic interneurons and tran- sient expression of p75 NTR in these neurons following focal cerebral ischemia. Further experiments are needed to clarify whether the expression of p75 NxR contributes to the high resistance of the striatal cholinergic neurons to ischemic damage, or if it initiates a death pathway, which is inhibited under the present experimental conditions. It also seems highly warranted to explore the possibility that high p75 NTR expression is a more general response to brain insults affecting the striatum. In any case, the model used here should be highly useful to clarify the role of the neurotrophins and their high- and low- affinity receptors in the adult brain, in particular their involvement in neuronal death and survival.
Acknowledgements--We thank Cristina Ciornei, Alicja Flasch and Ulla Jarl for excellent technical assistance. This work was supported by the Swedish Medical Research Council, the Arbetsmarknadens F6rs~kringsaktiebolag, the Swedish Stroke Foundation, the Wiberg Foundation, the Kock Foundation, the Zo6ga Foundation, the Medical Faculty of the University of Lund, The Royal Physio- graphic Society and the Elsa and Thorsten Segerfalk Foundation. Z. K. was partly supported by funds from I. Beritashvili Institute of Physiology, Georgian Academy of Sciences.
REFERENCES
1. Abe K., Araki T. and Kogure K. (1988) Recovery from edema and of protein synthesis differs between the cortex and caudate following transient focal cerebral ischemia in rats. J. Neurochem. 51, 1470 1476.
2. Abe K., Kawagoe J., Araki T., Aoki M. and Kogure K. (1992) Differential expression of heat shock protein 70 gene between the cortex and caudate after transient focal cerebral ischaemia in rats. Neurol. Res. 14, 381-385.
3. Altar C. A., Siuciak J. A., Wright P., Ip N. Y., Lindsay R. M. and Wiegand S. J. (1994) In situ hybridization of trkB and trkC receptor mRNA in rat forebrain and association with high-affinity binding of [12SI]BDNF, [~25I]NT-4/5 and [125I]NT-3. Eur. J. Neurosci. 6, 1389 1405.
4. Amano T., Yamakuui T., Okabe N., Kuwahara R., Ozawa F. and Hishinuma F. (1992) Regulation of nerve growth factor and nerve growth factor receptor production by NMDA in C6 glioma cells. Molec. Brain Res. 14, 35~,2.
5. Andsberg G., Kokaia Z., Bj6rklund A., Lindvall O. and Martinez-Serrano A. (1998) Amelioration of ischemia- induced neuronal death in the rat striatum by NGF-secreating neural stem cells. Eur. J. Neurosci. (submitted).
6. Barbacid M. (1995) Neurotrophic factors and their receptors. Curr. Opin. Cell Biol. 7, 148 155.

p75 Nva expression in focal ischemia 1123
7. Barker P. A. and Shooter E. M. (1994) Disruption of NGF binding to the low affinity neurotrophin receptor p75LNTR reduces NGF binding to TrkA on PC12 cells. Neuron 13, 203-215.
8. Bartus R. T., Dean R. L., Abelleira S., Charles V. and Kordower J. H. (1996) Dissociation of p75 receptors and nerve growth factor neurotrophic effects: lack of p75 immunoreactivity in striatum following physical trauma, excitotoxicity and NGF administration. Rest. Neurol. Neurosci. 10, 49-59.
9. Beal M. F., Kowall N. W., Swartz K. J., Ferrante R. J. and Martin J. B. (1989) Differential sparing of somatostatin-neuropeptide Y and cholinergic neurons following striatal excitotoxin lesions. Synapse 3, 3847.
10. Bederson J. B., Pitts L. H., Tsuji M., Nishimura M. C., Davis R. L. and Bartkowski H. (1986) Rat middle cerebral artery occlusion: evaluation of the model and development of neurologic examination. Stroke 17, 472476.
11. Boegman R. J., Smith Y. and Parent A. (1987) Quinolinic acid does not spare striatal neuropeptide Y-immunoreactive neurons. Brah~ Res. 415, 178-182.
12. Boissiere F., Hunot S., Faucheux B., Mouatt-Prigent A., Agid Y. and Hirsch E. C. (1994) High affinity neurotrophin receptors in cholinergic neurons in the human brain. C. R. Acad. Sei., Paris" 317, 997 1003.
13. Bonfoco E., Krainc D., Ankarcrona M., Nicotera P. and Lipton S. A. (1995) Apoptosis and necrosis: two distinct events induced, respectively, by mild and intense insults with N-methyl-D-aspartate or nitric oxide/superoxide in cortical cell cultures. Proc. natn. Acad. Sci. U.S.A. 92, 7162 7166.
14. Burke R. E. and Karanas A. L. (1990) Quantitative morphological analysis of striatal cholinergic neurons in perinatal asphyxia. Ann. Neurol. 27, 81 88.
15. Butcher L. L., Oh J. D., Woolf N. J., Edwards R. H. and Roghani A. (1992) Organization of central cholinergic neurons revealed by combined in situ hybridization histochemistry and choline-O-acetyltransferase immunocyto- chemistry. Neuroehem. Int. 21,429M45.
16. Chao M. V. and Hempstead B. L. (1995) p75 and Trk: a two-receptor system. Trends Neurosci. 18, 316 321. 17. Charriaut-Marlangue C., Margaill I., Represa A., Popovici T., Plotkine M. and Ben-Ari Y. (1996) Apoptosis and
necrosis after reversible focal ischemia: an in situ DNA fragmentation analysis. J. cerebr. Blood Flow Metab. 16, 186 194.
18. Chen H. H., Chien C.-H. and Liu H. M. (1994) Correlation between angiogenesis and basic fibroblast growth factor expression in experimental brain infarct. Stroke 25, 1651 1657.
19. Chesselet M. F., Gonzales C., Lin C. S., Polsky K. and Jin B. K. (1990) Ischemic damage in the striatum of adult gerbils: relative sparing of somatostatinergic and cholinergic interneurons contrasts with loss of efferent neurons. Expl Neurol. 110, 209 218.
20. Christensen T., Bruhn T., Frank L. and Diemer N. H. (1996) Differential effect of NMDA and AMPA receptor blockade on protein synthesis in the rat infarct borderzone. Acta neurol, scand. 93, 160-167.
21. Cohen-Cory S., Elliott R. C., Dreyfus C. F. and Black I. B. (1993) Depolarizing influences increase low-affinity NGF receptor gene expression in cultured Purkinje neurons. Expl Neurol. 119, 165 175.
22. Cosgaya J. M., Garcia-Villalba P., Perona R. and Aranda A. (1996) Comparison of the effects of retinoic acid and nerve growth factor on PC12 cell proliferation, differentiation, and gene expression. J. Neurochem. 66, 89 98.
23. Curtis R., Adryan K. M., Stark J. L., Park J. S., Compton D. L., Weskamp G., Huber L. J., Chao M. V., Jaenisch R.. Lee K. F., Lindsay R. M. and DiStefano P. S. (1995) Differential role of the low affinity neurotrophin receptor (p75) in retrograde axonal transport of the neurotrophins. Neuron 14, 1201-121 l.
24. Davies A. M., Lee K.-F. and Jaenisch R. (1993) p75-deficient trigeminal sensory neurons have an altered response to NGF but not to other neurotrophins. Neuron 11,565-574.
25. Davies S. W. and Roberts P. J. (1988) Sparing of cholinergic neurons following quinolinic acid lesions of the rat striatum. Neuroscience 26, 387 393.
26. De Ryck M., Van Reempts J., Borgers M., Wauquier A. and Janssen P. A. (1989) Photochemical stroke model: flunarizine prevents sensorimotor deficits after neocortical infarcts in rats. Stroke 20, 1383 1390.
27. del Valle M. E., Alvarez-M6ndez J. C., Calzada B. and Vega J. A. (1992) Nerve growth factor receptor immunoreactivity in non-nervous structures of the adult rat brain. Eur. J. Histochem. 36, 435444.
28. Du C., Hu R., Csernansky C. A., Liu X. Z., Hsu C. Y. and Choi D. W. (1996) Additive neuroprotective effects of dextrorphan and cycloheximide in rats subjected to transient focal cerebral ischemia. Brain Res. 718, 233 236.
29. Figueiredo B. C., Skup M., Bedard A. M., Tetzlaff W. and Cuello A. C. (1995) Differential expression of p l40trk, p75NGFR and growth-associated phosphoprotein-43 genes in nucleus basalis magnocellularis, thalamus and adjacent cortex following neocortical infarction and nerve growth factor treatment. Neuroseience 68, 2945.
30. Figueredo-Cardenas G., Anderson K. D., Chen Q., Veenman C. L. and Reiner A. (1994) Relative survival of striatal projection neurons and interneurons after intrastriatal injection of quinolinic acid in rats. Expl Neurol. 129, 37 56.
31. Forloni G. L., Angeretti N., Rizzi M. and Vezzani A. (1992) Chronic infusion of quinolinic acid in rat strialum: effects on discrete neuronal populations. J. neurol. Sci. 108, 129-136.
32. Foster A. C., Gill R. and Woodruff G. N. (1988) Neuroprotective effects of MK-801 in vivo: selectivity and evidence for delayed degeneration mediated by NMDA receptor activation. J. Neurosci. 8, 47454754.
33. Frim D. M., Uhler T. A., Short M. P., Ezzedine Z. D., Klagsbrun M., Breakefield X. O. and Isacson O. (1993) Effects of biologically delivered NGF, BDNF and bFGF on striatal excitotoxic lesions. NeuroReport 4, 367 370.
34. Gage F. H., Batchelor P., Chen K. S., Chin D., Higgins G. A., Koh S., Deputy S., Rosenberg M. B., Fischer W. and Bj6rklund A. (1989) NGF receptor reexpression and NGF-mediated cholinergic neuronal hypertrophy in the damaged adult neostriatum. Neuron 2, 1177-I 184.
35. Gerfen C. R. (1992) The neostriatal mosaic: multiple levels of compartmental organization. Trends Neurosci. 15, 133 139.
36. Gibbs R. B., McCabe J. T., Buck C. R., Chao M. V. and Pfaff D. W. (1989) Expression of NGF receptor in the rat forebrain detected with in situ hybridization and immunohistochemistry. Molec. Brain Res. 6, 275 287.
37. Goto S., Nagahiro S., Korematsu K. and Ushio Y. (1993) Striatonigral involvement following transient focal cerebral ischemia in the rats: an immunohistochemical study on a reversible ischemia model. Acta neuropath. 85, 515 520.
38. Graybiel A. M. (1990) Neurotransmitters and neuromodulators in the basal ganglia. Trends" Neurosci. 13, 244 254. 39. Gundersen H. J., Bendtsen T. F., Korbo L., Marcussen N., Moller A., Nielsen K., Nyengaard J. R., Pakkenberg B.,
Sorensen F. B., Vesterby A. and West M. J. (1988) Some new, simple and efficient stereological methods and their use in pathological research and diagnosis. Acta pathol, microbiol, scand. 96, 379-394.

1124 Z. Kokaia et al.
40. Hagg T., Quon D., Higaki J. and Varon S. (1992) Ciliary neurotrophic factor prevents neuronal degeneration and promotes low affinity NGF receptor expression in the adult rat CNS. Neuron 8, 145-158.
41. Hantzopoulos P. A., Suri C., Glass D. J., Goldfarb M. P., Yancopoulos G. D. (1994) The low affinity NGF receptor, p75, can collaborate with each of the Trks to potentiate functional responses to the neurotrophins. Neuron 13, 187- 201.
42. Heimer L., Zahm D. S. and Alheid G. F. (1995) Basal ganglia. In The Rat Nervous System (ed Paxinos G.), pp. 579-628. Academic, San Diego.
43. Johnston M. V. and Hudson C. (1987) Effects of postnatal hypoxia-ischemia on cholinergic neurons in the developing rat forebrain: choline acetyltransferase immunocytochemistry. Brain Res. 431, 41 50.
44. Kaplan D. R. and Miller F. D. (1997) Signal transduction by the neurotrophin receptors. Curr. Opin. Cell Biol. 9, 213-221.
45. Kawaguchi Y., Wilson C. J., Augood S. J. and Emson P. C. (1995) Striatal interneurones: chemical, physiological and morphological characterization. Trends Neurosci. 18, 527-535.
46. Kiss J., McGovern J. and Patel A. J. (1988) Immunohistochemical localization of cells containing nerve growth factor receptors in the different regions of the adult rat forebrain. Neuroscience 27, 731 748.
47. Kojima M., Takahashi N., Ikeuchi T. and Hatanaka H. (1992) Nerve growth factor (NGF)-mediated up-regulation of low-affinity NGF receptor gene expression in cultured basal forebrain cholinergic neurons from postnatal 3-day-old rats. Molec. Brain Res. 16, 267 273.
48. Kokaia Z., Zhao Q., Kokaia M., Elm6r E., Metsis M., Smith M.-L., Siesj6 B. K. and Lindvall O. (1995) Regulation of brain-derived neurotrophic factor gene expression after transient middle cerebral artery occlusion with and without brain damage. Expl Neurol. 136, 73 88.
49. Kovfics Z., Ikezaki K., Samoto K., Inamura T. and Fukui M. (1996) VEGF and fit. Expression time kinetics in rat brain infarct. Stroke 27, 1865 1872.
50. Krupinski J., Kaluza J., Kumar P., Kumar S. and Wang J. M. (1994) Role of angiogenesis in patients with cerebral ischemic stroke. Stroke 25, 1794~1798.
51. Krupinski J., Kumar P., Kumar S. and Kaluza J. (1996) Increased expression of TGF-bl in brain tissue after ischemic stroke in humans. Stroke 27, 852-857.
52. Lee T.-H., Kato H., Kogure K. and Itoyama Y. (1996) Temporal profile of nerve growth factor-like immunoreactivity after transient focal cerebral ischemia in rats. Brain Res. 713, 199 210.
53. Li Y., Chopp M., Jiang N. and Zaloga C. (1995) In situ detection of DNA fragmentation after focal cerebral ischemia in mice. Molec. Brain Res. 28, 164-168.
54. Linnik M. D., Zobrist R. H. and Hatfield M. D. (1993) Evidence supporting a role for programmed cell death in focal cerebral ischemia in rats. Stroke 24, 2002 2008.
55. Ma Y., Campenot R. B. and Miller F. D. (1992) Concentration-dependent regulation of neuronal gene expression by nerve growth factor. J. Cell Biol. 117, 135-141.
56. Magal E., Burnham P. and Varon S. (1991) Effect of CNTF on low-affinity NGF receptor expression by cultured neurons from different rat brain regions. J. Neurosci. Res. 30, 560-566.
57. Martinez-Serrano A. and Bj6rklund A. (1996) Protection of the neostriatum against excitotoxic damage by neurotrophin-producing, genetically modified neural stem cells. J. Neurosci. 16, 46044616.
58. Martinez-Serrano A., Hantzopoulos P. A. and Bj6rklund A. (1996) Ex vivo gene transfer of brain-derived neurotrophic factor to the intact rat forebrain: neurotrophic effects on cholinergic neurons. Eur. J. Neurosci. 8, 727 735.
59. Massieu L., Thedinga K. H., McVey M. and Fagg G. E. (1993) A comparative analysis of the neuroprotective properties of competitive and uncompetitive N-methyl-D-aspartate receptor antagonists in vivo: implications for the process of excitotoxic degeneration and its therapy. Neuroscience 55, 883 892.
60. Memezawa H., Smith M.-L. and Siesj6 B. K. (1992) Penumbral tissues salvaged by reperfusion following middle cerebral artery occlusion in rats. Stroke 23, 552 559.
61. Menkes J. H. (1988) Huntington disease: finding the gene and after. Pediatr. Neurol. 4, 73-78. 62. Merlio J.-P., Ernfors P., Jaber M. and Persson H. (1992) Molecular cloning of rat trkC and disribution of
cells expressing messenger RNAs for members of the trk family in the rat central nervous system. Neuroscience 51, 513 532.
63. Mies G., Ishimaru S., Xie Y., Seo K. and Hossmann K. A. (1991) Ischemic thresholds of cerebral protein synthesis and energy state following middle cerebral artery occlusion in rat. J. cerebr. Blood Flow Metab. 11,753-761.
64. Nakao N., Kokaia Z., Odin P. and Lindvall O. (1995) Protective effects of BDNF and NT-3 but not PDGF against hypoglycemic injury to cultured striatal neurons. Expl Neurol. 131, 1 10.
65. Ouimet C. C. and Greengard P. (1990) Distribution of DARPP-32 in the basal ganglia: an electron microscopic study. J. Neurocytol. 19, 39 52.
66. Ouimet C. C., Miller P. E., Hemmings H. C. Jr, Walaas S. I. and Greengard P. (1984) DARPP-32, a dopamine- and adenosine 3':5'-monophosphate-regulated phosphoprotein enriched in dopamine-innervated brain regions, lIl. Immunocytochemical localization. J. Neurosci. 4, 111 124.
67. Paxinos G. and Watson C. (1997) The Rat Brain in Stereotaxic Coordinates. 3rd edn, Academic, San Diego. 68. Phelps P. E., Houser C. R. and Vaughn J. E. (1985) Immunocytochemical localization of choline acetyltransferase
within the rat neostriatum: a correlated light and electron microscopic study of cholinergic neurons and synapses. J. comp. Neurol. 238, 286 307.
69. Reiner A. and Anderson K. D. (1990) The patterns of neurotransmitter and neuropeptide co-occurrence among striatal projection neurons: conclusions based on recent findings. Brain Res. Rev. 15, 251 265.
70. Roback J. D., Diede S. J., Downen M., Lee H. J., Kwon J., Large T. H., Otten U. and Wainer B. H. (1992) Expression of neurotrophins and the low-affinity NGF receptor in septal and hippocampal reaggregate cultures: local physiologic effects of NGF synthesized in the septal region. Devl Brain Res. 70, 123-133.
71. Rodriguez-Tebar A., Dechant G. and Barde Y.-A. (1990) Binding of brain-derived neurotrophic factor to the nerve growth factor receptor. Neuron 4, 487492.

p75 N'vR expression in focal ischemia 1125
72. Rodriguez-Tebar A., Dechant G., Gotz R. and Barde Y.-A. (1992) Binding of neurotrophin-3 to its neuronal receptors and interactions with nerve growth factor and brain-derived neurotrophic factor. Eur. molec. Biol. Org J. 11, 917- 922.
73. Sauer H., Nishimura M. C. and Philips H. S. (1996) Deletion of the p75NTFR gene attenuates septal cholinergic cell loss in mice heterozygous for a deletion of the NGF gene. Soc. Neurosci. Abstr. 22, 513.
74. Sobreviela T., Clary D. O., Reichardt L. F., Brandabur M. M., Kordower J. H. and Mufson E. J. (1994) TrkA-immunoreactive profiles in the central nervous system: colocalization with neurons containing p75 nerve growth factor receptor, choline acetyltransferase, and serotonin. J. comp. Neurol. 350, 587 611.
75. Spoerri P. E., Romanello S., Petrelli L., Negro A., Dal-Toso R., Leon A. and Skaper S. D. (1992) Nerve growth factor (NGF) receptors in a central nervous system gliai cell line: upregulation by NGF and brain-derived neurotrophic l~actor. J. Neurosci. Res. 33, 82-90.
76. Steininger T. L., Wainer B. H., Klein R., Barbacid M. and Palfrey H. C. (1993) High-affinity nerve growth factor receptor (Trk) immunoreactivity is localized in cholinergic neurons of the basal forebrain and striatum in the adult rat brain. Brain Res. 612, 330-335.
77. Van der Zee C. E. E. M., Ross G. M., Riopelle R. J. and Hagg T. (1996) Survival of cholinergic forebrain neurons in developing p75NGFR-deficient mice. Science 274, 1729 1732.
78. Ventimiglia R., Mather P. E., Jones B. E. and Lindsay R. M. (1995) The neurotrophins BDNF, NT-3 and NT-4/5 promote survival and morphological and biochemical differentiation of striatal neurons in vitro. Eur. J. Neurosci. 7, 213 222.
79. yon Bartheld C. S., Williams R., Lefcort F., Clary D. O., Reichardt L. F. and Bothwell M. (1996) Retrograde transport of neurotrophins from the eye to the brain in chick embryos: roles of the p75NTR and trkB receptors. J. Neurosci. 16, 2995-3008.
80. Weskamp G. and Reichardt L. F. (1991) Evidence that biological activity of NGF is mediated through a novel subclass of high affinity receptors. Neuron 6, 649-663.
81. Widmer H. R. and Hefti F. (1994) Neurotrophin-4/5 promotes survival and differentiation of rat striatal neurons developing in culture. Eur. J. Neurosci. 6, 1669 1679.
82. Yan Q. and Johnson E. M. Jr (1988) An immunohistochemical study of the nerve growth factor receptor in developing rats. J. Neurosci. 8, 3481 3498.
83. Yan Q. and Johnson E. M. Jr (1989) Immunohistochemical localization and biochemical characterization of nerve growth factor receptor in adult rat brain. J. comp. Neurol. 290, 585-598.
84. Yan Q., Matheson C., Lopez O. T. and Miller J. A. (1994) The biological responses of axotomized adult motoneurons to brain-derived neurotrophic factor. J. Neurosci. 14, 5281-5291.
85. Yan Q., Radeke M. J., Matheson C. R., Talvenheimo J., Welcher A. A. and Feinstein S. C. (1997) lmmuno- cytochemical localization of TrkB In the central nervous system of the adult rat. J. comp. Neurol. 378, 135-157.
86. Zhao Q., Smith M.-L. and Siesj6 B. K. (1994) The w-conopeptide SNX-Il l , an N-type calcium channel blocker, dramatically ameliorates brain damage due to transient focal ischaemia. Acta physiol, scand. 150, 459-461.
(Accepted 29 October 1997)