distribution of p75 and trk-neurotrophin receptor proteins in adult human sympathetic ganglia
TRANSCRIPT

Anat Embryol (1996) 193:577-583 �9 Springer-Verlag 1996 n
O R I G I N A L A R T I C L E
O. Garcia-Suarez �9 F.J. Naves �9 M.E. Del Valle I. Esteban �9 E. Bronzetti �9 E. Vazquez �9 J.A. Vega
Distribution of p75 and trk-neurotrophin receptor proteins in adult human sympathetic ganglia
Accepted: 17 November 1995
Abstract We investigated the expression of immunore- activity (IR) for low- (p75) and high-affinity (trk pro- teins) neurotrophin-receptor proteins in adult human paravertebral-sympathetic ganglion neurons. Mouse monoclonal antibodies against the pan-neurotrophin-re- ceptor p75, and rabbit polyclonal antibodies against spe- cific epitopes of the intracytoplasmic domain on trk neu- rotrophin-receptor proteins were used in fresh unfixed and formaldehyde-fixed paraffin-embedded sympathetic ganglia. All adult human paravertebral-sympathetic neu- rons displayed trkA neurotrophin-receptor-like protein IR, 10% express trkC neurotrophin-receptor-like protein IR, 37-44% show p75 IR, and no IR was obtained for trkB neurotrophin-receptor-like protein. The intensity of immunostaining was independent of the neuron size. La- belling of non-neuronal tissues, especially blood-vessel walls, was observed for p75, trkA and trkC neurotro- phin-receptor proteins. These results indicate that over- lapping exists in the expression of p75 and trk neurotro- phin-receptor proteins in adult human paravertebral-sym- pathetic neurons, and suggest that neurotrophins might act on these neurons.
Key words trk neurotrophin-receptor proteins - p75 protein �9 Paravertebral-sympathetic ganglia �9 Human
O. Garcia-Suarez �9 FJ. Naves. M.E. Del Valle �9 I. Esteban J.A. Vega ([~) Departamento de Morfologfa y Biologfa Celular, Facultad de Medicina, Universidad de Oviedo, E-33006 Oviedo, Spain
E. Bronzetti Dipartimento di Scienze Cardiovascolari e Respiratorie, Universit5 "La Sapienza", I-0 l 61 Roma, Italy
E. Vazquez Departamento de Bioquimica, Biologfa Molecular y Fisiolog/a, Facultad de Medicina, Universidad de Valladolid, C.S.I.C., E-47005 Valladolid, Spain
Introduction
The neurons of the paravertebral sympathetic ganglia, and a subpopulation of the prevertebral ones, are under the control of neurotrophins (see Bothwell 1995; Chao and Hempstead 1995) for the development (Levi-Mon- talcini 1987) and expression and maintenance of their phenotypic characteristics (Longo et al. 1993; Klein 1994; Snider 1994). The key role of neurotrophins in de- veloping sympathetic ganglia has been recently con- firmed by selective gene knockout studies (Lee et al. 1992; Klein et al. 1993; Crowley et al. 1994; Ernfors et al. 1994; Farifias et al. 1994; Jones et al. 1994; Klein 1994; Smeyne et al. 1994; Snider 1994). Nevertheless, only limited infoxrnation is so far available about the dis- tribution of p75 or trk proteins, considered to be the low- and high-affinity neurotrophin receptors (see Bothwell 1995) in adult human sympathetic ganglia (Sobue et al. 1989; Yamamoto et al. 1993; Vega et al. 1994b), since most of the studies analyzed the occurrence of nenrotro- phin-receptors' mRNAs (Schecterson and Bothwell 1992; Yamamoto et al. 1993; Ehrhard and Otten 1994; Dixon and McKinnon 1994). A recent study reporting the expression of trk-neurotrophin receptors in different parts of the human fetal and adult peripheral nervous system did not consider the sympathetic ganglia (Shelton et al. 1995).
In this study we describe the distribution of the immu- noreactivity (IR) for p75 and trk-neurotrophin-receptor proteins in adult human paravertebral-sympathetic gan- glia. This study might serve as a baseline for future re- searches on neuropathies involving sympathetic ganglia, such as sympathetic disfunction in aging (Schmidt et al. 1993; Andrews and Cowen 1994), injury (Schmidt et al. 1988), hypertension (Kondo et al. 1990; Messina and Bell 1991), and diabetes (Hellweg and Hartung 1990; Schmidt et al. 1993; Apfel et aI. 1994).

578
Materials and methods
Treatment of the tissue
Lumbar-paravertebral-sympathetic ganglia (n=22) were obtained during routine vascular surgery from 17 subjects of both sexes (13 males and 4 females) and of different ages (ranging from 26 to 52 years). The subjects were free of neurological or metabolic disor- ders, and received identical symptomatic treatment. Four addition- al ganglia were obtained from two subjects (males of 26 and 32 years who died in traffic incidents) during removal of organs for transplantation. In each case the ganglia were removed within 45 min after of the circulation had stopped. Obtaining the tissues was in compliance with Spanish Law (Law 30/79 and Royal De- cree 426/80) for brain death and organ transplantation.
The tissues were collected in an ice-cold physiological solution, dissected, and fixed for 24 h with 10% formaldehyde in 0.1 M phos- phate buffer saline (pH 7.4), dehydrated and embedded in paraffin (n=18), or embedded in a cryoprotectant medium and stored at -70~ Serial sections 10 ~tm thick were mounted on gelatin-coated microscope slides and processed for indirect peroxidase anti-peroxi- dase immunohistochemistry. The normal histology of the analyzed tissue samples was ascertained with hematoxylin-eosin staining.
Immunohistochemistry
Sections were washed in HC1-TRIS buffer (0.05 M, pH 7.5) con- taining 0.1% Triton X- 100, 0.1% bovine serum albumin and 0.1% sheep serum. Then, after blocking endogenous peroxidase activity (1% H202 in cold 100% methanol) and non specific binding (fetal- calf serum), sections were incubated with rabbit-polyclonal (pAbs) and mouse-monoclonal (mAbs) primary antibodies (see below). The incubation was carried out overnight at 4~ in a hu- mid chamber. Thereafter, sections were washed and incubated with peroxidase-labelled sheep anti-rabbit IgG immunoglobulin, or with peroxidase-labelled sheep anti-mouse IgG immunoglobu- lin, both diluted 1:100 (Ammersham). Finally, after washing, sec- tions were incubated for 5-10 rain in 3.3'-diaminobenzidine (0.5 mg/ml in TBS 0.2 M, pH 7.5) containing 0.01% H202. Con- trol sections were processed as described below, using pre-ab- sorbed rabbit sera instead of the primary antibodies.
To label neurotrophin receptor proteins pAbs directly against specific epitopes of the intracellular domain of trkA (diluted 1:100), trkB (diluted 1:50 and 1:100) and trkC proteins (diluted 1:200) (Santa Cruz Biotechnology, USA) were used. Moreover, two mAbs against p75 (clone 8211, Boeringer Mannheim; clone ME20.4, Amersham), both diluted 5 gg/ml, were also employed. Primary pAbs were raised against a peptide corresponding to resi- dues 763-777 (trkA protein), 794-808 (trkB protein) and 798-812 (trkC protein) mapping within the COO-domain of the deduced human trkA aminoacid sequence, and the predicted mouse trkB and trkC proteins (manufacturer's instructions). The specificity of the immunoreactivity was controlled by incubating representative sections with specifically pre-absorbed rabbit sera (5 gg of peptide in 1 ml of the primary antibody solution) (Santa Cruz Biotechnol- ogy, catalog references sc- l l8 P, sc-12 P and sc- l l7 P for trkA, trkB and trkC proteins, respectively). Moreover, to avoid cross-re- activity of trk proteins to each other, aliquots of each trk protein's antibodies were absorbed with the control peptides for the other two trk proteins (5 ~tg+5 gg in 1 ml of the primary antibody solu- tion). Additional controls were carried out by omitting the primary antibodies in the incubation. In the sections incubated with the preabsorbed antisera no specific immunoreactivity was observed. Moreover, in the cross-reactivity controls the distribution and in- tensity of the immunostaining did not change with respect to that of the non-preabsorbed primary antibody.
Quantitative study
The neuron size and the intensity of immunostaining developed in the neuron-cell bodies of sympathetic ganglia for each investigat-
ed antibody were evaluated by automatic quantitative image analy- sis (MIP System, Servicio de Anfilisis de Imfigenes de la Universi- dad de Oviedo, Spain) as reported in detail in a previous report (Vega et al. 1994b). Measurements were made on two sections, 70 ~tm apart, per case and antibody from the fixed material, and on one section from un-fixed material. Five randomly selected fields per section (2.5 mm 2) were evaluated, and the number of neurons evaluated was 30-50 per section. In the neuronal profiles with ap- parent nuclei the cell body size was measured by evaluating the mean diameter (in ~tm); neurons were divided into three groups re- ferred to as small (<25 gm), intermediate (26-50 gm) and large (>51 gm). The intensity of immunostaining was also estimated and expressed in arbitrary units of grey level, ranging from 1 (black) to 256 (white). According to the intensity of immunostain- ing, neuronal profiles were divided into four groups (64 units of grey level each) referred in the text as strong (1-64), high (65-128), intermediate (129-192) and low (193-256); nerve cell bodies showing an intensity of immunostaining _>240 were consid- ered as unreactive. Results are expressed as the percentage of la- belled neuron profiles in each pre-established neuron-size sub- class, and as mean+S.D, values of the intensity of immunostain- ing. It must be clear, however, that the aim of this part of the study was not to establish the distribution of neuron size in adult human sympathetic ganglia (see Schmidt et al. 1993; Vega et al. 1994a,b) but to analyze whether any relation exists between neuron size and intensity of immunostaining for neurotrophin-receptor proteins. Furthermore, data of optic microdensitometry are approximate for each antigen, and cannot be compared among them.
Results
In the ana lyzed paraver tebra l - sympathe t ic gangl ia no sig- nif icant age-dependent changes were encountered in the neuron density. The only not iceable structural change was an increase in the number o f lypofusc in neuronal bodies (data not shown). On the other hand, no differences were found in ei ther the dis tr ibut ion or the intensi ty o f immu- nosta ining be tween f ixed and unfixed mater ia l observed for any of the invest igated antigens (data not shown).
t r kA neuro t roph in- recep tor - l ike prote ins
Al l adult human paraver tebra l -sympathet ic neurons dis- p layed t rkA neurotrophin-receptor- l ike protein IR (Figs. 1A, 2A,B), showing a diffuse cy toplasmic pattern of stain- ing. The intensi ty of immunos ta in ing was intermediate and independent o f the neuron size (Table 1). In addi t ion to neurons, IR was also present in non-neuronal tissues, main ly in the connective septa and b lood-vesse l walls, and occas ional ly in the support ing satell i te cells (Fig. 2A,B).
trkB neuro t roph in- recep tor - l ike prote ins
In no case posi t ive immunos ta in ing was ob ta ined for trkB neuro t rophin receptor - l ike protein, ne i ther in neu- rons nor in non-neurona l t issues (Fig. 1B).
trkC neuro t rophin- recep tor - l ike prote ins
Only a smal l percen tage o f pos tgang l ion ic sympathe t i c neurons (10%) d i sp l ayed trkC neuro t rophin receptor - l ike

579
Fig. 1 Serial sections of a lumbar paravertebral sympathetic gan- glia showing the distribution of immunoreactivity for trkA (A), trkB (B), trkC (C, arrowheads) and p75 (D, E) neurotrophin-re-
ceptor proteins; in control sections (trkA pre-absorbed with the corresponding peptide, F) no immunoreactivity was observed. Bar 10 gm

580
Fig. 2 trkA neurotrophin-receptor-like protein IR was observed in all neurons and some in non-neuronal tissues, mainly blood ves- sels (A, B arrowheads); conversely trkC neurotrophin-receptor- like proteins IR were exclusively found in some neurons (C, D
large arrows) and the initial segment of their processes (C, D small arrows); a subset of neurons (E, F large arlvws) and non- neuronal tissues also display p75 IR. Bar 20/.tin
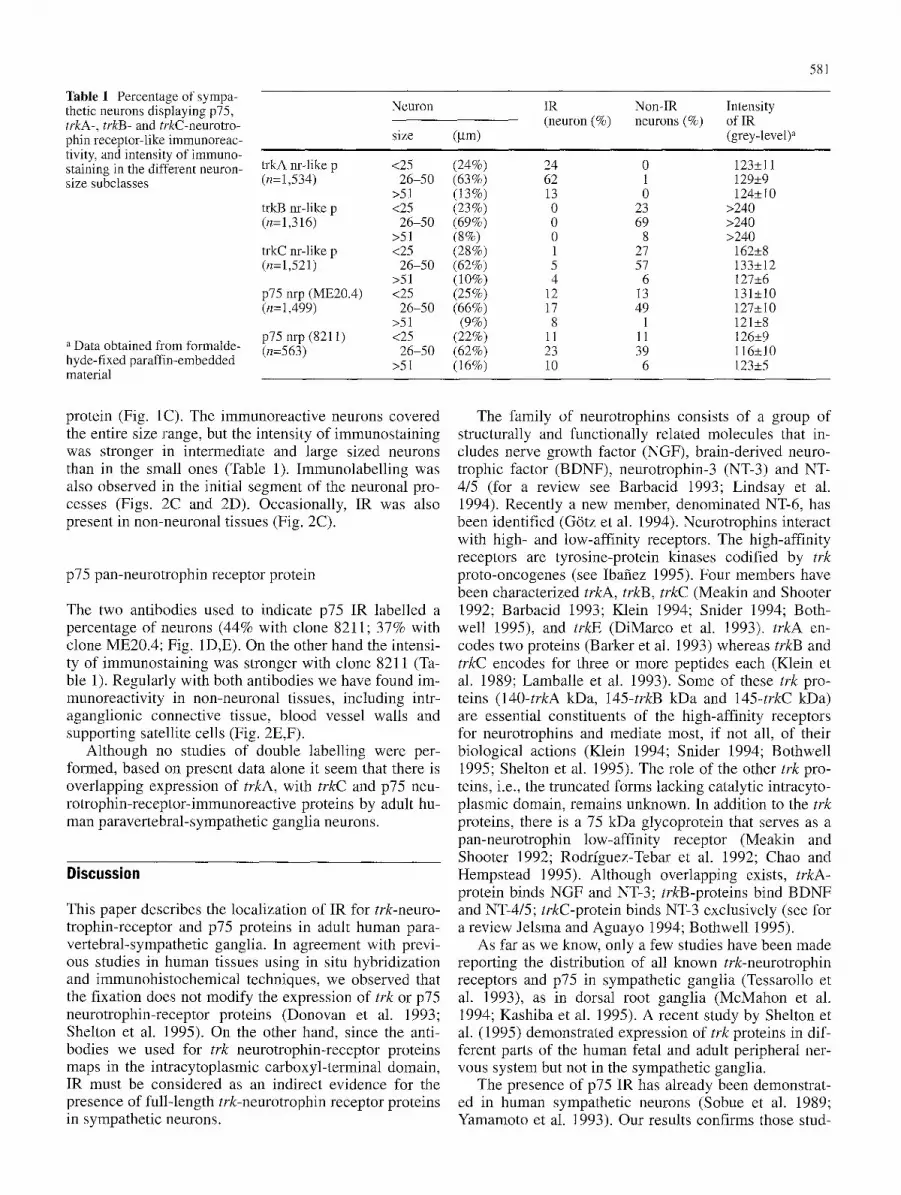
Table 1 Percentage of sympa- thetic neurons displaying p75, trkA-, trkB- and trkC-neurotro- phin receptor-like immunoreac- tivity, and intensity of immuno- staining in the different neuron- size subclasses
a Data obtained from formalde- hyde-fixed paraffin-embedded material
Neuron
size (gm)
trkA nr-like p <25 (24%) (n=1,534) 26-50 (63%)
>51 (13%) trkB nr-like p <25 (23%) (n=1,316) 26-50 (69%)
>51 (8%) trkC nr-like p <25 (28%) (n=1,521) 26-50 (62%)
>51 (10%) p75 nrp (ME20.4) <25 (25%) (n=1,499) 26-50 (66%)
>51 (9%) p75 nrp (8211) <25 (22%) (n=563) 26-50 (62%)
>51 (16%)
IR Non-IR Intensity (neuron (%) neurons (%) of IR
(grey-level) a
24 0 123• 62 1 129• 13 0 124• 0 23 >240 0 69 >240 0 8 >240 1 27 162• 5 57 133• 4 6 127•
12 13 131• 17 49 127• 8 1 121•
11 11 126• 23 39 116• 10 6 123•
581
protein (Fig. 1C). The immunoreactive neurons covered the entire size range, but the intensity of immunostaining was stronger in intermediate and large sized neurons than in the small ones (Table 1). Immunolabelling was also observed in the initial segment of the neuronal pro- cesses (Figs. 2C and 2D). Occasionally, 1R was also present in non-neuronal tissues (Fig. 2C).
p75 pan-neurotrophin receptor protein
The two antibodies used to indicate p75 IR labelled a percentage of neurons (44% with clone 8211; 37% with clone ME20.4; Fig. 1D,E). On the other hand the intensi- ty of immunostaining was stronger with clone 8211 (Ta- ble 1). Regularly with both antibodies we have found im- munoreactivity in non-neuronal tissues, including intr- aganglionic connective tissue, blood vessel walls and supporting satellite cells (Fig. 2E,F).
Although no studies of double labelling were per- formed, based on present data alone it seem that there is overlapping expression of trkA, with trkC and p75 neu- rotrophin-receptor-immunoreactive proteins by adult hu- man paravertebral-sympathetic ganglia neurons.
Discussion
This paper describes the localization of IR for trk-neuro- trophin-receptor and p75 proteins in adult human para- vertebral-sympathetic ganglia. In agreement with previ- ous studies in human tissues using in situ hybridization and immunohistochemical techniques, we observed that the fixation does not modify the expression of trk or p75 neurotrophin-receptor proteins (Donovan et al. 1993; Shelton et al. 1995). On the other hand, since the anti- bodies we used for trk neurotrophin-receptor proteins maps in the intracytoplasmic carboxyl-terminal domain, IR must be considered as an indirect evidence for the presence of full-length trk-neurotrophin receptor proteins in sympathetic neurons.
The family of neurotrophins consists of a group of structurally and functionally related molecules that in- cludes nerve growth factor (NGF), brain-derived neuro- trophic factor (BDNF), neurotrophin-3 (NT-3) and NT- 4/5 (for a review see Barbacid 1993; Lindsay et al. 1994). Recently a new member, denominated NT-6, has been identified (G6tz et al. 1994). Neurotrophins interact with high- and low-affinity receptors. The high-affinity receptors are tyrosine-protein kinases codified by trk proto-oncogenes (see Ibafiez 1995). Four members have been characterized trkA, trkB, trkC (Meakin and Shooter 1992; Barbacid 1993; Klein 1994; Snider 1994; Both- well 1995), and trkE (DiMarco et al. 1993). trkA en- codes two proteins (Barker et al. 1993) whereas trkB and trkC encodes for three or more peptides each (Klein et al. 1989; Lamballe et al. 1993). Some of these trk pro- teins (140-trkA kDa, 145-trkB kDa and 145--trkC kDa) are essential constituents of the high-affinity receptors for neurotrophins and mediate most, if not all, of their biological actions (Klein 1994; Snider 1994; Bothwell 1995; Shelton et al. 1995). The role of the other trk pro- teins, i.e., the truncated forms lacking catalytic intracyto- plasmic domain, remains unknown. In addition to the trk proteins, there is a 75 kDa glycoprotein that serves as a pan-neurotrophin low-affinity receptor (Meakin and Shooter 1992; Rodrfguez-Tebar et al. 1992; Chao and Hempstead 1995). Although overlapping exists, trkA- protein binds NGF and NT-3; lrkB-proteins bind BDNF and NT-4/5; trkC-protein binds NT-3 exclusively (see for a review Jelsma and Aguayo 1994; Bothwell 1995).
As far as we know, only a few studies have been made reporting the distribution of all known trk-neurotrophin receptors and p75 in sympathetic ganglia (Tessarollo et al. 1993), as in dorsal root ganglia (McMahon et al. 1994; Kashiba et al. 1995). A recent study by Shelton et al. (1995) demonstrated expression of trk proteins in dif- ferent parts of the human fetal and adult peripheral ner- vous system but not in the sympathetic ganglia.
The presence of p75 IR has already been demonstrat- ed in human sympathetic neurons (Sobue et al. 1989; Yamamoto et al. 1993). Our results confirms those stud-

582
ies indicating that only a subpopulation (37-43%) of adult human paravertebral-sympathetic neurons displays p75 IR. On the other hand, we also corroborate the oc- currence of p75 IR in the ganglionic non-neuronal tis- sues, especially in the blood vessels (Yamamoto et al. 1993; Vega et al. 1994b). Evidence regarding the role of p75 in the paravertebral-sympathetic neurons is still con- flicting, and it is therefore unknown (see Chao and Hempstead 1995). In fact, mice carrying a mutation of the gene encoding p75 did not change the size of sympa- thetic ganglia, their neurochemical characteristics, or the density of sympathetic innervation (Lee et al. 1992).
Regarding trkA-neurotrophin-receptor-like proteins, our results suggest that all postganglionic neurons of paravertebral-sympathetic ganglia express IR for those proteins. These findings are consistent with the expres- sion of mRNA trkA in a subpopulation of human (Yama- moto et al. 1993) and rat sympathetic ganglia (Schecter- son and Bothwell 1992; Dixon and McKinnon 1994), probably corresponding to both trkAI and trkAII neuro- trophin-receptor-like proteins (Barker et al. 1993). In ad- dition to neurons, and in disagreement with Yamamoto et al. (1993) we also observed IR in non-neuronal tissues, especially again in blood vessel walls, which confirms previous studies from our laboratory (Vega et al. 1994b) and others (Donovan et al. 1993). From a functional point of view these results lend support to the view that paravertebral-sympathetic neurons continue to depend on trkA neurotrophin-receptor-like protein ligands (mainly NGF) for survival and maintenance of functions into adulthood (see Korsching 1993). Moreover, our results are consistent with observations in gene knockouts in mice for trkA or NGF which show a massive reduction in the number of sympathetic neurons (Crowley et al. 1994; Smeyne et al. 1994).
We have not found trkB neurotrophin-receptor-like proteins IR in human sympathetic ganglia. Most of the studies carried out in other mammalian species lend sup- port to our findings. In fact, the mRNAs encoding trkB proteins are barely expressed in the rat paravertebral ganglia (Klein et al. 1989; Dixon and McKinnon 1994), and in mouse embryos they were undetected at later fetal developmental stages (Escandon et al. 1994). On the contrary, Ehrhard and Ottern (1994) showed trkB mRNA in postnatal rats, and Schecterson and Bothwell (1992) found trkB mRNA in 20% of the rat sympathetic gan- glia. In support of our findings, sympathetic ganglion neurons are not responsive to BDNF, although many sympathetic neurons may respond poorly into adulthood (Maisonpierre et al. 1990; see Korsching 1993). On the other hand the absence of trkB-neurotrophin-receptor- like protein is consistent with the small reduction of sympathetic neurons in trkB and BDNF knockout mice (Klein et al. 1993; Jones et al. 1994).
trkC mRNAs are expressed in the sympathetic ganglia of mouse embryos (Lamballe et al. 1993, 1994; Tessa- rollo et al. 1993) and postnatal rats (Dixon and McKin- non 1994), but data have not been reported in humans. Present results indicate that a subpopulation (10%) of
adult human paravertebral-sympathetic neurons displays trkC neurotrophin-receptor-like protein IR, suggesting a possible role of NT-3 on these neurons. However, the role of NT-3/trkC neurotrophin receptor protein in sym- pathetic ganglia still remains unclear. Thus, while trkC- knockout mice (Klein et al. 1994; Davies et al. 1995) showed slight neuronal loss (see Davies et al. 1995) in the NT-3 knockout-mice sympathetic neurons were re- duced to one-half of normal (Ernfors et al. 1994; Farifias et al. 1994). On the other hand, and in agreement with the present results, trkC-neurotrophin-receptor-like pro- teins IR are also expressed by blood vessels of human fe- tal and adult tissues (Shelton et al. 1995).
In summary, adult human postganglionic sympathetic neurons placed in the paravertebral ganglia express im- munoreactivity for low- and high-affinity neurotrophin receptor proteins, although in a different percentage for each one. This study might serve as a baseline for future researches on neuropathies involving sympathetic gan- glia (Anand et al 1991; Hellweg and Hartung 1990; Mes- sina and Bell 1991; Andrews and Cowen 1994; Apfel et al. 1994) since probably neurotrophins will be used in the future as therapeutic molecules (see Lindsay et al. 1994; Ibafiez 1995)
Acknowledgements This study was supported by a grant from the University of Oviedo (DF/95-218M) to FJN. The authors thank Prof. J. Represa for critical reading of the manuscript,
References
Anand R Rudge P, Mathias CJ, Springall DR, Ghatei MA, Naher- Noe M, Sharief M, Misra VR Polak JM, Bloom SR, Thomas PK (1991) New autonomic and sensory neuropathy with loss of adrenergic sympathetic function and sensory neuropeptides. Lancet 337:1253-1254
Andrews TJ, Cowen T (1994) Nerve growth factor enhances the dendritic arborization of sympathetic ganglion cells undergo- ing atrophy in aged rats. J Neurocytol 23:234-241
Apfel SC, Arezzo JC, Browlee M, Federoff H, Kessler JA (1994) Nerve growth factor administration protects against experi- mental diabetic sensory neuropathy. Brain Res 634:7-12
Barbacid M (1993) Nerve growth factor: a tale of two receptors. Oncogene 8:2033-2042
Barker PA, Lomen-Hoerth C, Gensch EM, Meakin SO, Glass DJ, Shooter EM (1993) Tissue-specific alternative splicing gener- ates two isoforms of the trkA receptor. J Biol Chem 268: 15150-15157
BothwelI M (1995) Functional interactions of neurotrophins and neurotrophin receptors. Annu Rev Neurosci 18:223-253
Chao MV, Hempstead B L (1995) p75 and Trk: a two-receptor system. Trends Neurosci 18:321-326
Crowley C, Spencer SD, Nishimura MC, Chen KS, Pitts-Meek S, Armanini MR Ling LH, McMahon SB, Shelton DL, Levinson AD, Phillips HS (1994) Mice lacking nerve growth factor dis- play peripheral loss of sensory and sympathetic neurons yet de- velop basal forebrain cholinergic neurons. Cell 76:100 1-1011
Davies AM, Minichiello L, Klein R (1995) Developmental changes in NT3 signalling via TrkA and TrkB in embryonic neurons. EMBO J 14:4482-4489
Di Marco E, Cutuli N, Guerra L, Cancedda R, De Luca M (1993) Molecular cloning of trkE, a novel trk-related putative tyrosine kinase receptor isolated from normal human keratinocytes and widely expressed by normal human tissues. J Biol Chem 268: 24290-24295

Dixon JE, McKinnon D (1994) Expression of the trk gene family of neurotrophins receptors in prevertebral sympathetic ganglia. Brain Res Dev Brain Res 77:177-182
Donovan MJ, Hampstead BL, Horvath C, Chao MV, Schofield D (1993) Immunohistochemical localization of Trk receptor pro- tein in pediatric small round blue cell tumors. Am J Pathol 143:1560-1567
Ehrhard PB, Otten U (1994) Postnatal ontogeny of the neurotro- phin receptors trk and trkB mRNA in rat sensory and sympa- thetic ganglia. Neurosci Lett 166:207-210
Ernfors R Lee FK, Kucera J, Jaenisch R (1994) Lack of neurotro- phin-3 leads to deficiencies in the peripheral nervous system and loss of limb proprioceptive afferents. Cell 77:503-512
Escandon E, Soppter D, Rosenthal A, Mendoza-Ramirez J-L, Sz6nyi E, Burton LE, Henderson CE, Parada LF, Nikolics K (1994) Regulation of neurotrophin receptor expression during embryonic and postnatal development. J Neurosci 14: 2054- 2068
Farifias I, Jones KR, Backus C, Wang X-Y, Reichardt LF (1994) Severe sensory and sympathetic deficits in mice lacking neu- rotrophin-3. Nature 369:658-661
Gatz R, K6ster R, Winkler C, Raulf F, Lottspeich F, Schartl M, Thoenen H (1994) Neurotrophin-6 is a new member of the nerve growth factor family. Nature 372:266-269
Helweg R, Hartung HD (1990) Endogenous levels of nerve growth factor (NGF) are altered in experimental diabetes mellitus: a possible role for NGF in pathogenesis of diabetic neuropathy. J Neurosci Res 26:258-267
Ibafiez CF (1995) Neurotrophic factors: from structure-function studies to designing effective therapeutics. Trends Biol Tech- nol 13:217-227
Jelsma TN, Aguayo AJ (1994) Trophic factors. Curr Opin Neuro- biol 4:717-725
Jones KR, Farifias I, Backus C, Reichardt LF (1994) Targeted dis- ruption of the BDNF gene perturbs brain and sensory neuron development but not motor neuron development. Cell 76: 989-999
Kashiba H, Noguchi K, Ueda Y, Senba E (1995) Coexpression of trk family members and low-affinity neurotrophins receptors in rat dorsal root ganglion neurons. Mol Brain Res 30: 158- 164
Klein R (1994) Role of neurotrophins in mouse neuronal develop- ment. FASEB J 8:738-744
Klein R, Parada LF, Coullier F, Barbacid M (1989) trkB, a novel tyrosine protein kinase receptor expressed during mouse neu- ral development. EMBO J 8:3701-3709
Klein R, Smeyne RJ, Wurst W, Long LK, Auerbach BA, Joyner AL, Barbacid M (1993) Targeted disruption of the trkB neuro- trophin receptor gene results in nervous system lesions and neonatal death. Cell 75:113-122
Klein R, Silos-Santiago I, Smeyne RJ, Lira SA, Brambilla R, Bry- ant S, Zhang L, Snider WD, Barbacid M (1994) Disruption of the neurotrophin-3 receptor gene trkC eliminates Ia muscle af- ferents and results in abnormal movements. Nature 368: 249-251
Kondo M, Terada M, Shimizu D, Fujiwara T, Tabei R (1990) Mor- phometrie study of the superior cervical and stellate ganglia of spontaneously hypertensive rats during the prehypertensive stage. Virchows Arch B Celt Pathol 58:371-376
Korsching S (1993) The neurotrophic factor concept: a reexamina- tion. J Neurosci 13:2739-2748
Lamballe F, P Tapley, M Barbacid (1993) trkC encodes multiple neurotrophin-3 receptors with distinct biological properties and substrate specificities. EMBO J 12:3083-3094
Lamballe E Smeyne RJ, Barbacid M (1994) Developmental ex- pression of trkC, the neurotrophin-3 receptor, in the mammali- an nervous system. J Neurosci 14:14-28
Lee K-F, Li E, Huber LJ, Landis SC, Sharpe AH, Chao MV, Jaen- isch AH (1992) Targeted mutation of the gene encoding the low affinity NGF receptor p75 leads to deficits in the peripher- al sensory nervous system. Cell 69:737-749
583
Levi-Montalcini R (1987) The nerve growth factor 35 years later. Science 237:1154-1162
Lindsay RM, Wiegand S J, Altar CA, Stefano PS (1994) Neurotro- phic factors: from molecule to man. Trends Neurosci 17: 182-190
Longo FM, Holtzman DM, Grimes ML, Mobley WC (1993) Nerve growth factor: actions in the peripheral and central ner- vous system. In: Loughlin SE, Fallon JH (eds) Neurotrophic factors. Academic Press, New York London, pp 209-256
Maisonpierre PC, Belluscio L, Squinto S, Ip NY, Furth ME, Lind- say RM, Yancopoulos GD (1990) Neurotrophin-3: a neurotro- phic factor related to NGF and BDNF. Science 247: 1446- 1451
McMahon SB, Armanini MR Ling LH, Phillips HS (1994) Ex- pression and coexpression of Trk receptors in a subpopulation of adult primary sensory neurons projecting to identified pe- ripheral targets. Neuron 12:1161-1171
Meakin SO, Shooter EM (1992) The nerve growth factor family of receptors. Trends Neurosci 15:323-331
Messina A, Bell C (1991) Are genetically hypertensive rats defi- cient in nerve growth factor. Neuroreport 2:45.48
Rodrfguez-Tebar A, Dechant G, G6tz R, Barde Y-A (1992) Bind- ing of neurotrophin-3 to its neuronal receptors and interactions with nerve growth factor and brain-derived neurotrophic fac- tor. EMBO J 11:917-922
Schecterson LC, Bothwell M (1992) Novel roles for neurotrophins are suggested by BDNF and NT-3 mRNA expression in devel- oping neurons. Neuron 9:449.463
Schmidt RE, McAtee S J, Plurad DA, Parvin CA, Cogswell BE, Roth KA (1988) Differential susceptibility of prevertebral and paravertebral sympathetic ganglia to experimental injury. Brain Res 460:214-226
Schmidt RE, Chae HY, Parvin CA, Roth KA (1990) Neuroaxonal dystrophy in aging human sympathetic ganglia. Am J Pathol 136:1327-1338
Schmidt RE, Plurad SB, Parvin CA, Roth KA (1993) Effect of di- abetes and aging on human sympathetic autonomic ganglia. Am J Pathol 145:143-153
Shelton DL, Sutherland J, Gripp J, Camerato T, Armanini MR Phillips HS, Carroll K, Spencer SD, Levinson AD (1995) Hu- man trks: molecular cloning, tissue distribution, and expres- sion of extracellular domain inmunoadhesins. J Neurosci 15: 477-491
Smeyne RJ, Klein R, Schnapps A, Long LK, Bryant S, Lewin A, Lira SA, Barbacid M (1994) Severe sensory and sympathetic neuropathies in mices carrying a disrupted trkfNGF receptor gene. Nature 368:246-249
Snider WD (1994) Functions of the neurotrophins during nervous system development: what the knockouts are teaching us. Cell 77:627-638
Sobue G, Yasuda T, Mitsuma T, Pleasure D (1989) Nere growth factor receptor immunoreactivity in the neuronal perikarya of human sensory and sympathetic ganglia. Neurology 39: 937- 941
Tessarollo L, Tsoulfas R Martin-Zanca D, Gilbert DJ, Jenkins NA, Copeland NG, Parada LF (1993) trkC, a receptor for neurotro- phin-3, is widely expressed in the developing nervous system and in non-neuronal tissues. Development 118:463-475
Vega JA, Humara JM, Naves FJ, Esteban I, Del Valle ME (1994a) Immunoreactivity for phosphorylated 200-kDa neurofilament subunit is heterogeneously expressed in human sympathetic and primary sensory neurons. Anat Embryol 190:453-459
Vega JA, Vfizquez E, Naves FJ, Del Valle ME, Calzada B, Represa JJ (1994b) Immunohistochemical localization of the high-af- finity NGF receptor (gp 140-trkA) in the adult human dorsal root and sympathetic ganglia and in the nerves and sensory corpuscles supplying digital skin. Anat Rec 240:579-588
Yamamoto M, Sobue G, Mutoh T, Li M, Doyu M, Mitsuma T, Kim- ata K (1993) Gene expression of high- (p140 t~k) and low-affinity nerve growth factor receptor (LNGFR) in the adult and aged hu- man peripheral nervous system. Neurosci Lett 158:39.43