fishing in the dark-local knowledge, night spearfishing and spawning aggregations in the western...
TRANSCRIPT

Biological Conservation xxx (2011) xxx–xxx
Contents lists available at SciVerse ScienceDirect
Biological Conservation
journal homepage: www.elsevier .com/ locate /biocon
Fishing in the dark-local knowledge, night spearfishing and spawningaggregations in the Western Solomon Islands
R.J. Hamilton a,⇑, M. Giningele b, S. Aswani c, J.L. Ecochard d
a The Nature Conservancy, Indo Pacific Resource Center, 51 Edmondstone, South Brisbane, QLD 4101, Australiab Dunde Community, Munda, Western Province, Solomon Islandsc Department of Anthropology and Interdepartmental Graduate Program in Marine Science, University of California, Santa Barbara, CA 93106-3210, USAd Technology and Information Systems, The Nature Conservancy, Arlington, VA, USA
a r t i c l e i n f o a b s t r a c t
Article history:Received 30 May 2011Received in revised form 21 November 2011Accepted 26 November 2011Available online xxxx
Keywords:Fish spawning aggregationLocal ecological knowledgeGroupersCommunity-based monitoringMarine protected areaCoral Triangle
0006-3207/$ - see front matter � 2011 Elsevier Ltd. Adoi:10.1016/j.biocon.2011.11.020
⇑ Corresponding author. Tel.: +61 7 3214 6913; faxE-mail address: [email protected] (R.J. Hamilton)
Please cite this article in press as: Hamilton, R.JSolomon Islands. Biol. Conserv. (2011), doi:10.1
Within the marine conservation community there is considerable interest in combining local knowledgeand science to achieve management objectives. Yet there remain few studies which have examined themerits and caveats of local knowledge, or shown how combining both knowledge systems has resulted inbetter management outcomes. This study outlines collaborative efforts to conserve fish spawning aggre-gations (FSAs) in Roviana Lagoon, Western Solomon Islands. Baseline information on FSAs was obtainedthrough local knowledge and spearfishing creel surveys. This information provided the starting point forestablishing a 2-year community-based underwater monitoring program at the largest known FSA inRoviana Lagoon, where the brown-marbled grouper (Epinephelus fuscoguttatus), camouflage grouper(Epinephelus polyphekadion) and squaretail coralgrouper (Plectropomus areolatus) co-aggregate. This par-ticipatory research shows that local knowledge on FSAs is utilised to maximise returns from fishing, withspearfishermen targeting aggregations at night during the lunar periods when abundances peak. Becauseof its shallow distribution P. areolatus is the most vulnerable of the three groupers to nighttime spearf-ishing, with two fishermen capable of removing 15–30% of the total spawning biomass in two nights.Underwater monitoring demonstrates that while fishermen provided accurate information on manyaspects of FSAs, their knowledge on spawning seasons was inaccurate for the FSA reported on here. Peakaggregations occurred from December to April each year, which differs from the traditionally recognisedgrouper season of October to January. A combination of local knowledge and science was used to developappropriate management measures for this FSA, with the aggregation declared a community-based mar-ine protected area (MPA) in 2006.
� 2011 Elsevier Ltd. All rights reserved.
1. Introduction
Within the marine conservation community there is consider-able interest in understanding how local knowledge of fisherscan be utilised to advance both management and conservationagendas (e.g. Drew, 2005; Johannes and Neis, 2007). Fishers canprovide important information on such things as inter-annual, sea-sonal, lunar, diel, tide- and habitat-related differences in speciesdistributions and abundance, as well as providing a historical per-spective on the state of fisheries (Johannes et al., 2000). To datemarine scientists and conservation practitioners have incorporatedfishers’ local knowledge into research programs, fisheries assess-ments, species evaluations and conservation planning processes(e.g. Sadovy and Cheung, 2003; Aswani and Hamilton, 2004; Dulvyand Polunin, 2004; Saénz-Arroyo et al., 2005; Silvano et al., 2006;
ll rights reserved.
: +61 7 3214 6999..
., et al. Fishing in the dark-local016/j.biocon.2011.11.020
Aswani et al., 2007; Almany et al., 2010; Game et al., 2011; Tayloret al., 2011). One of the most widely applied uses of fishers localknowledge is in the research and conservation of fish spawningaggregations (FSAs) (Hamilton et al., 2012). In many locations, fish-ers have known of FSAs for generations, or have experienced sea-sonal gluts in landings subsequently identified as FSAs (Johannes,1978; Colin et al., 2003). In recognition of this, and because ofthe practical difficulties of discovering FSAs that typically form athighly localised areas for brief periods of time, scientists that studyFSAs have often drawn on local knowledge in the initial stages oftheir research (e.g. Johannes et al., 1999; Robinson et al., 2008;Sadovy de Mitcheson et al., 2008).
Three large-bodied aggregating reef fishes that have receivedconsiderable attention in the past decade are the brown-marbledgrouper (Epinephelus fuscoguttatus), camouflage grouper (Epinephe-lus polyphekadion) and squaretail coralgrouper (Plectropomusareolatus). In the Indo-Pacific these three groupers frequentlyco-aggregate to spawn at predictable sites and times around the
knowledge, night spearfishing and spawning aggregations in the Western

2 R.J. Hamilton et al. / Biological Conservation xxx (2011) xxx–xxx
new or full moon (e.g. Johannes et al., 1999; Rhodes and Sadovy,2002; Pet et al., 2005; Hamilton et al., 2011). These groupers makeup important components of many small-scale commercial fisher-ies in the Pacific (e.g. Rhodes and Tupper, 2007) and they are threeof the most economically valuable species in the Southeast Asia-based live reef food fish trade (LRFFT) (Sadovy et al., 2003). As a re-sult of their predictable aggregating behaviour, all are highly sus-ceptible to overexploitation, with E. fuscoguttatus and E.polyphekadion listed as near threatened (Cornish, 2004; Russellet al., 2006, respectively) and P. areolatus listed as vulnerable (Thi-erry et al., 2008) in the 2008 IUCN Red List.
Globally there are numerous examples of where targeted fish-ing of FSAs has resulted in aggregation decline or loss (e.g. Sadovyand Domeier, 2005; Hamilton and Matawai, 2006; Rhodes et al.,2011), which has negative implications for local fisheries and thecommunities that depend on them (Sadovy and Domeier, 2005).Fisheries managers and conservation practitioners are increasinglyrecognising that FSAs need protection, with marine protected areas(MPAs) and closed seasons being the most commonly imple-mented conservation measures (e.g. Beets and Friedlander, 1999;Sadovy de Mitcheson et al., 2008; Rhodes et al., 2011). In some in-stances management measures for FSAs have been designed solelyon local knowledge (e.g. Hamilton et al., 2012). In other cases, long-term underwater monitoring has helped to establish a more com-plete picture of FSAs than local knowledge alone, while building lo-cal capacity and support for management (e.g. Hamilton et al.,2011).
The multi-species FSA reported on in this paper is the largestknown grouper aggregation in Roviana Lagoon, Western SolomonIslands. A Roviana spearfisherman discovered this FSA in 1995,and between 1995 and 2006 it was exploited by nighttime spear-fishermen. By 2004 Roviana spearfishermen perceived catches atthis FSA to be in steep decline, and when a non-government organi-sation (NGO) raised community awareness on the importance ofconserving FSAs, the customary owners of this FSA agreed to estab-lish a science-based, community-led underwater monitoring pro-gram at the aggregation site (Hamilton and Kama, 2004). Thepurpose of this monitoring was to obtain scientific information onaggregation seasons and status, with a view that this informationcould be utilised to develop appropriate community-based man-agement strategies for the FSA. Feedback on the scientific findingsresulted in the FSA being declared a no-take community-basedMPA in June 2006. While compliance with this MPA has generallybeen good, a limited amount of poaching by nighttime spearfisher-men has occurred since 2006. The aims of this paper are to (1)establish the temporal and spatial trends of E. fuscoguttatus, E.polyphekadion and P. areolatus at this site, (2) determine if small-scale commercial nighttime spear fisheries pose a significant threatto spawning aggregations of E. fuscoguttatus, E. polyphekadion and P.areolatus, and (3) evaluate whether or not local knowledge needs tobe independently validated before being used as the basis formanagement.
2. Methods
2.1. Environmental setting
The current study was conducted in Roviana Lagoon, WesternSolomon Islands (Fig. 1) in the Solomon Sea, which forms the mosteastern part of the Coral Triangle (Veron et al., 2009). Roviana La-goon is a body of shallow water approximately 50 km long en-closed between the New Georgia mainland and a series ofuplifted coral reef islands lying 2–3 km offshore. In Roviana fishingpractices are strongly influenced by local knowledge of the lagoonenvironment. Knowledge of the tides, lunar stages and seasons in-
Please cite this article in press as: Hamilton, R.J., et al. Fishing in the dark-localSolomon Islands. Biol. Conserv. (2011), doi:10.1016/j.biocon.2011.11.020
form fishers when, where and for what they should fish (e.g. Ham-ilton and Walter, 1999; Aswani and Hamilton, 2004; Aswani andVaccaro, 2008). October to the end of January is widely known asthe ‘pazara’ season, when groupers such as E. fuscoguttatus, E.polyphekadion, P. areolatus and the white-streaked grouper (Epi-nephelus ongus) are said to aggregate at a minimum of 14 FSAs inRoviana Lagoon, in the 10 days leading up to and including thenew moon (Hamilton and Kama, 2004). In Roviana Lagoon spawn-ing aggregations are targeted by subsistence and small-scale com-mercial fisheries, and in several cases it appears that FSAs havebeen fished almost to the point of extirpation (Hamilton and Kama,2004).
2.2. Site description
The studied FSA is located on a seaward facing reef promontorywithin the western region of Roviana Lagoon (Fig. 1). In order toprotect this FSA from further exploitation by outside entities, theexact location is not shown. The reef flat of the promontory is3 m deep, with the reef slope dropping steeply to below 200 m.Aggregations of all three species overlap over a linear reef distanceof ca. 300 m and between depths of 3–50 m (Fig. 2). P. areolatus aremost abundant between depths of 3–15 m, whereas E. fuscogutta-tus and E. polyphekadion are most abundant in the middle sectionof the FSA between depths of 5–50 m (Fig. 2).
This FSA is predominantly fished by nighttime spearfishermenalthough daytime spearfishing occasionally occurs here. Hookand line fishing is not practiced. Nighttime spearfishermen freedive with the aid of fins, mask, snorkel, a rubber powered spearand an underwater flashlight with a maximum 4-h battery life.Spearfishermen prefer to fish this site at night, as this method pro-duces higher catch rates than daytime spearfishing (authors, per-sonal observations), a pattern that is common in many parts ofthe Pacific (Gillett and Moy, 2006). Local spearfishermen reportedthat in the first 2 years that they exploited this site (1995–1996)the maximum catch of two spearfishermen exceeded 200 P. areol-atus a night (Hamilton and Kama, 2004). Fishermen noted declinesin maximum catches from 1997 onwards, and in 2001 the maxi-mum catch of two spearfishermen was 71 P. areolatus in a night(see Table 3). Commercial spearfishermen sell their catches at fish-eries centres in the Munda Township (Fig. 1). In 2004 approxi-mately ten spearfishermen were known to periodically fish thisFSA.
2.3. Creel survey
Between the 23rd of January 2001 and the 20th of April 2001one of the authors (MG) led 41 nighttime spearfishing trips overall of the outer reefs shown in Fig. 1. Twenty-seven percent(n = 11) of these fishing trips occurred at the FSA site and 73%(n = 30) occurred on outer reefs 1–12 km from the FSA. After eachfishing trip MG recorded the date, fisher name(s), location, timespent travelling and fishing, species caught and their frequencyand weight. In some instances species were clumped at the genuslevel. Fish names were recorded in Roviana and translated to scien-tific names by the senior author, who participated in several ofthese fishing trips. On two occasions in 2005 and one occasion in2010 one of the authors (MG) documented additional informationon three nighttime spearfishing trips at the FSA that he did not par-ticipate in. He did this through informal discussions with thespearfishermen who led these trips, and by recording the totalweights of gutted P. areolatus that were purchased from these fish-ermen at local fisheries centres in Munda. Since 2005 and 2010data represent gutted catches, these total weights were raised bya factor of 1.15 to represent the ungutted condition (Grandcourt,2005). Catch per unit effort (CPUE) for the 2005 and 2010 trips
knowledge, night spearfishing and spawning aggregations in the Western

Fig. 1. The lagoon and reef habitat at the western end of Roviana Lagoon. The location of the provincial centre Munda is shown.
R.J. Hamilton et al. / Biological Conservation xxx (2011) xxx–xxx 3
were estimated by assigning 4 h of effort to each spearfishermen,based on the authors knowledge of the maximum battery life ofthe underwater flashlights used in Roviana Lagoon.
2.4. Community-based underwater monitoring program
In March 2004 four Roviana spearfishermen who exploited FSAs(free diving) and were also experienced SCUBA divers receivedtraining in monitoring spawning aggregations (Rhodes, 2004). Fol-lowing this training community approval was sought and given toestablish a 2 year community-based monitoring program at theFSA reported on here (Hamilton and Kama, 2004). A monitoringteam and protocols were established, and over the next 2 yearsthe senior author provided scientific advice, mentoring, logisticaland field support to Roviana community monitors.
To determine the seasonality with which aggregations form a250 m by 10 m wide (2500 m2), 20 m deep belt transect was placedat the FSA in April 2004, and a second 200 m by 10 m wide(2000 m2), 10 m deep belt transect placed at the FSA in March2005 (Fig. 2). The upper and lower boundaries of transects weremarked by inserting 1 m long re-bar stakes into the reef at 10 m(horizontal) intervals, and white PVC pipe sleeves were placed overeach re-bar stake to increase visibility. The two transects covered12% of the total FSA area, exceeding the 10% minimum area thatis recommended in order to ensure sufficient precision (Pet et al.,2006). A HOBO� data logger (Water Temp Pro H2O-001, Onset,Southern MA, USA) was placed at the start of the deep transect(20 m) in October 2004 and recorded water temperatures hourlyuntil it was recovered in June 2006.
Please cite this article in press as: Hamilton, R.J., et al. Fishing in the dark-localSolomon Islands. Biol. Conserv. (2011), doi:10.1016/j.biocon.2011.11.020
Transects were monitored by underwater visual census (UVC)in the early afternoon on 100 days between the 19th April 2004and the 12th June 2006 (Table 1). Initially monitoring occurredonce a month on the new moon. The decision to monitor on thenew moon was based on local knowledge and UVC surveys con-ducted at this site in March 2004, which showed peak FSAs abun-dances persisted several days after the new moon had passed. Itwas expected that sampling on the new moon would provide a‘snapshot’ of the peak aggregation density. We became aware offull moon P. areolatus aggregations in October 2004 and began reg-ular full moon monitoring in March 2005.
To verify that new moon monitoring conformed to periods ofpeak density and abundance, intensive (daily) UVC surveys wereconducted just before and just after the new moon in Februaryand March 2005 (6 and 5 days respectively) and February 2006(7 days). To better understand daily aggregating patterns intensivesurveys were also conducted over all lunar phases between Marchand April 2006. One of the authors (MG) led all 100 UVC surveys,accompanied by another trained monitor. During monitoring, di-vers swam side-by-side along the midpoint of a transect, maintain-ing a position several metres above the aggregated fish. Monitorsrecorded the total number of each of the three species sightedwithin transect boundaries.
2.5. Calculation of FSA area
In order to estimate maximum FSA abundances the total aggre-gation area was estimated in March 2005 and 2006. In both yearswe placed permanent markers at the aggregation boundarieswhere densities declined rapidly and neared non-reproductive val-
knowledge, night spearfishing and spawning aggregations in the Western
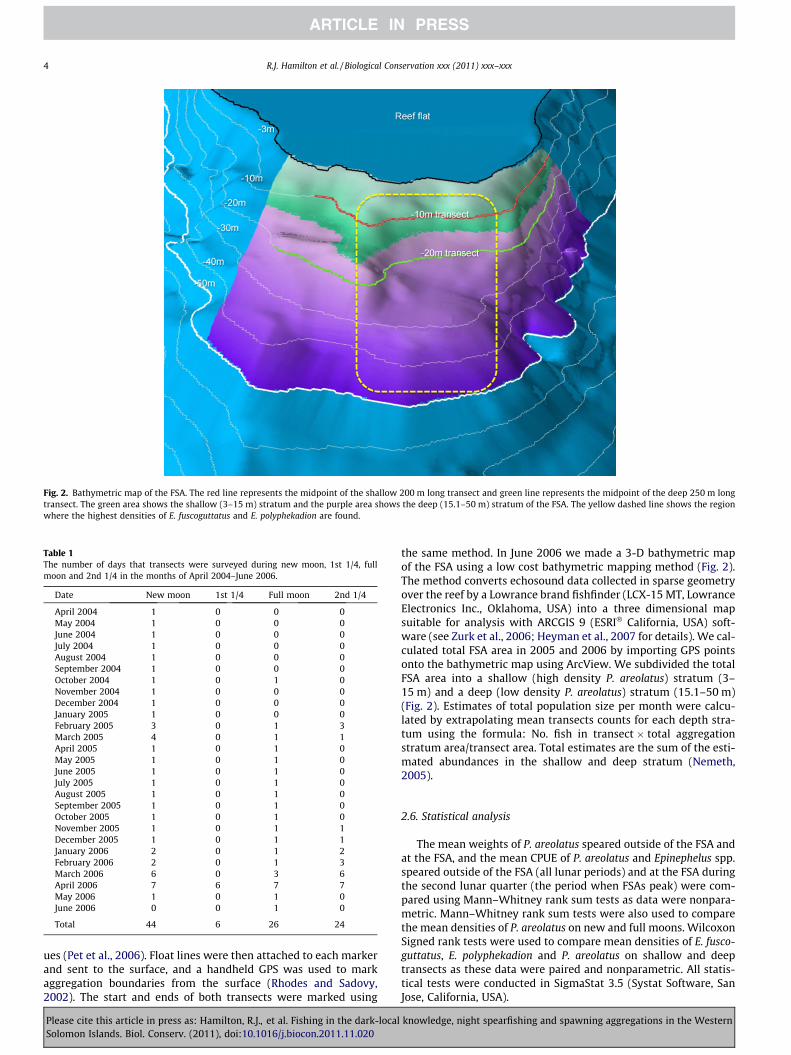
Fig. 2. Bathymetric map of the FSA. The red line represents the midpoint of the shallow 200 m long transect and green line represents the midpoint of the deep 250 m longtransect. The green area shows the shallow (3–15 m) stratum and the purple area shows the deep (15.1–50 m) stratum of the FSA. The yellow dashed line shows the regionwhere the highest densities of E. fuscoguttatus and E. polyphekadion are found.
Table 1The number of days that transects were surveyed during new moon, 1st 1/4, fullmoon and 2nd 1/4 in the months of April 2004–June 2006.
Date New moon 1st 1/4 Full moon 2nd 1/4
April 2004 1 0 0 0May 2004 1 0 0 0June 2004 1 0 0 0July 2004 1 0 0 0August 2004 1 0 0 0September 2004 1 0 0 0October 2004 1 0 1 0November 2004 1 0 0 0December 2004 1 0 0 0January 2005 1 0 0 0February 2005 3 0 1 3March 2005 4 0 1 1April 2005 1 0 1 0May 2005 1 0 1 0June 2005 1 0 1 0July 2005 1 0 1 0August 2005 1 0 1 0September 2005 1 0 1 0October 2005 1 0 1 0November 2005 1 0 1 1December 2005 1 0 1 1January 2006 2 0 1 2February 2006 2 0 1 3March 2006 6 0 3 6April 2006 7 6 7 7May 2006 1 0 1 0June 2006 0 0 1 0
Total 44 6 26 24
4 R.J. Hamilton et al. / Biological Conservation xxx (2011) xxx–xxx
ues (Pet et al., 2006). Float lines were then attached to each markerand sent to the surface, and a handheld GPS was used to markaggregation boundaries from the surface (Rhodes and Sadovy,2002). The start and ends of both transects were marked using
Please cite this article in press as: Hamilton, R.J., et al. Fishing in the dark-localSolomon Islands. Biol. Conserv. (2011), doi:10.1016/j.biocon.2011.11.020
the same method. In June 2006 we made a 3-D bathymetric mapof the FSA using a low cost bathymetric mapping method (Fig. 2).The method converts echosound data collected in sparse geometryover the reef by a Lowrance brand fishfinder (LCX-15 MT, LowranceElectronics Inc., Oklahoma, USA) into a three dimensional mapsuitable for analysis with ARCGIS 9 (ESRI� California, USA) soft-ware (see Zurk et al., 2006; Heyman et al., 2007 for details). We cal-culated total FSA area in 2005 and 2006 by importing GPS pointsonto the bathymetric map using ArcView. We subdivided the totalFSA area into a shallow (high density P. areolatus) stratum (3–15 m) and a deep (low density P. areolatus) stratum (15.1–50 m)(Fig. 2). Estimates of total population size per month were calcu-lated by extrapolating mean transects counts for each depth stra-tum using the formula: No. fish in transect � total aggregationstratum area/transect area. Total estimates are the sum of the esti-mated abundances in the shallow and deep stratum (Nemeth,2005).
2.6. Statistical analysis
The mean weights of P. areolatus speared outside of the FSA andat the FSA, and the mean CPUE of P. areolatus and Epinephelus spp.speared outside of the FSA (all lunar periods) and at the FSA duringthe second lunar quarter (the period when FSAs peak) were com-pared using Mann–Whitney rank sum tests as data were nonpara-metric. Mann–Whitney rank sum tests were also used to comparethe mean densities of P. areolatus on new and full moons. WilcoxonSigned rank tests were used to compare mean densities of E. fusco-guttatus, E. polyphekadion and P. areolatus on shallow and deeptransects as these data were paired and nonparametric. All statis-tical tests were conducted in SigmaStat 3.5 (Systat Software, SanJose, California, USA).
knowledge, night spearfishing and spawning aggregations in the Western
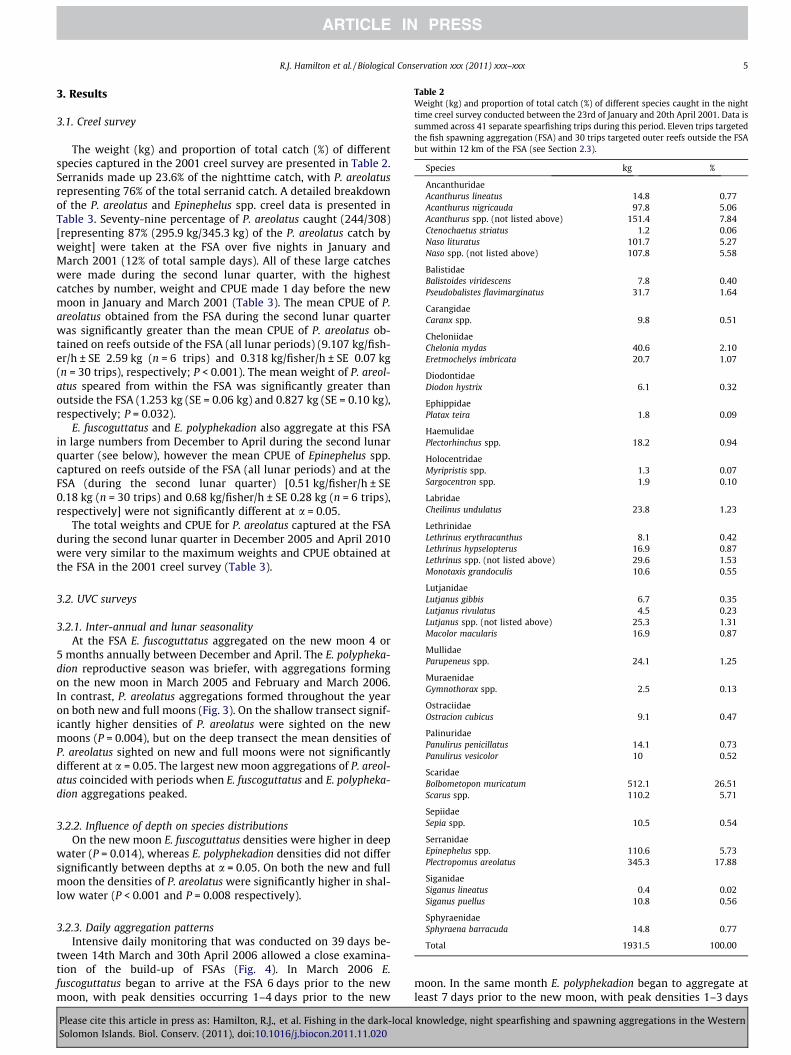
Table 2Weight (kg) and proportion of total catch (%) of different species caught in the nighttime creel survey conducted between the 23rd of January and 20th April 2001. Data issummed across 41 separate spearfishing trips during this period. Eleven trips targetedthe fish spawning aggregation (FSA) and 30 trips targeted outer reefs outside the FSAbut within 12 km of the FSA (see Section 2.3).
Species kg %
AncanthuridaeAcanthurus lineatus 14.8 0.77Acanthurus nigricauda 97.8 5.06Acanthurus spp. (not listed above) 151.4 7.84Ctenochaetus striatus 1.2 0.06Naso lituratus 101.7 5.27Naso spp. (not listed above) 107.8 5.58
BalistidaeBalistoides viridescens 7.8 0.40Pseudobalistes flavimarginatus 31.7 1.64
CarangidaeCaranx spp. 9.8 0.51
CheloniidaeChelonia mydas 40.6 2.10Eretmochelys imbricata 20.7 1.07
DiodontidaeDiodon hystrix 6.1 0.32
EphippidaePlatax teira 1.8 0.09
HaemulidaePlectorhinchus spp. 18.2 0.94
HolocentridaeMyripristis spp. 1.3 0.07Sargocentron spp. 1.9 0.10
LabridaeCheilinus undulatus 23.8 1.23
LethrinidaeLethrinus erythracanthus 8.1 0.42Lethrinus hypselopterus 16.9 0.87Lethrinus spp. (not listed above) 29.6 1.53Monotaxis grandoculis 10.6 0.55
LutjanidaeLutjanus gibbis 6.7 0.35Lutjanus rivulatus 4.5 0.23Lutjanus spp. (not listed above) 25.3 1.31Macolor macularis 16.9 0.87
MullidaeParupeneus spp. 24.1 1.25
MuraenidaeGymnothorax spp. 2.5 0.13
OstraciidaeOstracion cubicus 9.1 0.47
PalinuridaePanulirus penicillatus 14.1 0.73Panulirus vesicolor 10 0.52
ScaridaeBolbometopon muricatum 512.1 26.51Scarus spp. 110.2 5.71
SepiidaeSepia spp. 10.5 0.54
SerranidaeEpinephelus spp. 110.6 5.73Plectropomus areolatus 345.3 17.88
SiganidaeSiganus lineatus 0.4 0.02Siganus puellus 10.8 0.56
SphyraenidaeSphyraena barracuda 14.8 0.77
Total 1931.5 100.00
R.J. Hamilton et al. / Biological Conservation xxx (2011) xxx–xxx 5
3. Results
3.1. Creel survey
The weight (kg) and proportion of total catch (%) of differentspecies captured in the 2001 creel survey are presented in Table 2.Serranids made up 23.6% of the nighttime catch, with P. areolatusrepresenting 76% of the total serranid catch. A detailed breakdownof the P. areolatus and Epinephelus spp. creel data is presented inTable 3. Seventy-nine percentage of P. areolatus caught (244/308)[representing 87% (295.9 kg/345.3 kg) of the P. areolatus catch byweight] were taken at the FSA over five nights in January andMarch 2001 (12% of total sample days). All of these large catcheswere made during the second lunar quarter, with the highestcatches by number, weight and CPUE made 1 day before the newmoon in January and March 2001 (Table 3). The mean CPUE of P.areolatus obtained from the FSA during the second lunar quarterwas significantly greater than the mean CPUE of P. areolatus ob-tained on reefs outside of the FSA (all lunar periods) (9.107 kg/fish-er/h ± SE 2.59 kg (n = 6 trips) and 0.318 kg/fisher/h ± SE 0.07 kg(n = 30 trips), respectively; P < 0.001). The mean weight of P. areol-atus speared from within the FSA was significantly greater thanoutside the FSA (1.253 kg (SE = 0.06 kg) and 0.827 kg (SE = 0.10 kg),respectively; P = 0.032).
E. fuscoguttatus and E. polyphekadion also aggregate at this FSAin large numbers from December to April during the second lunarquarter (see below), however the mean CPUE of Epinephelus spp.captured on reefs outside of the FSA (all lunar periods) and at theFSA (during the second lunar quarter) [0.51 kg/fisher/h ± SE0.18 kg (n = 30 trips) and 0.68 kg/fisher/h ± SE 0.28 kg (n = 6 trips),respectively] were not significantly different at a = 0.05.
The total weights and CPUE for P. areolatus captured at the FSAduring the second lunar quarter in December 2005 and April 2010were very similar to the maximum weights and CPUE obtained atthe FSA in the 2001 creel survey (Table 3).
3.2. UVC surveys
3.2.1. Inter-annual and lunar seasonalityAt the FSA E. fuscoguttatus aggregated on the new moon 4 or
5 months annually between December and April. The E. polypheka-dion reproductive season was briefer, with aggregations formingon the new moon in March 2005 and February and March 2006.In contrast, P. areolatus aggregations formed throughout the yearon both new and full moons (Fig. 3). On the shallow transect signif-icantly higher densities of P. areolatus were sighted on the newmoons (P = 0.004), but on the deep transect the mean densities ofP. areolatus sighted on new and full moons were not significantlydifferent at a = 0.05. The largest new moon aggregations of P. areol-atus coincided with periods when E. fuscoguttatus and E. polypheka-dion aggregations peaked.
3.2.2. Influence of depth on species distributionsOn the new moon E. fuscoguttatus densities were higher in deep
water (P = 0.014), whereas E. polyphekadion densities did not differsignificantly between depths at a = 0.05. On both the new and fullmoon the densities of P. areolatus were significantly higher in shal-low water (P < 0.001 and P = 0.008 respectively).
3.2.3. Daily aggregation patternsIntensive daily monitoring that was conducted on 39 days be-
tween 14th March and 30th April 2006 allowed a close examina-tion of the build-up of FSAs (Fig. 4). In March 2006 E.fuscoguttatus began to arrive at the FSA 6 days prior to the newmoon, with peak densities occurring 1–4 days prior to the new
Please cite this article in press as: Hamilton, R.J., et al. Fishing in the dark-localSolomon Islands. Biol. Conserv. (2011), doi:10.1016/j.biocon.2011.11.020
moon. In the same month E. polyphekadion began to aggregate atleast 7 days prior to the new moon, with peak densities 1–3 days
knowledge, night spearfishing and spawning aggregations in the Western
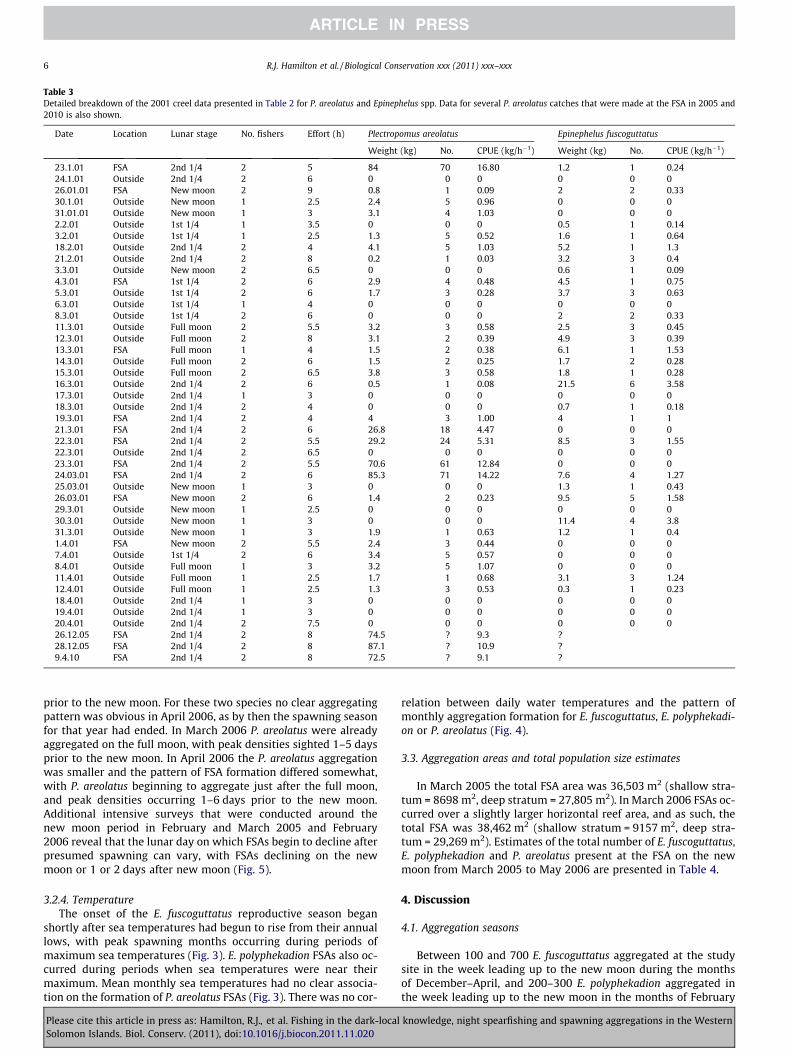
Table 3Detailed breakdown of the 2001 creel data presented in Table 2 for P. areolatus and Epinephelus spp. Data for several P. areolatus catches that were made at the FSA in 2005 and2010 is also shown.
Date Location Lunar stage No. fishers Effort (h) Plectropomus areolatus Epinephelus fuscoguttatus
Weight (kg) No. CPUE (kg/h�1) Weight (kg) No. CPUE (kg/h�1)
23.1.01 FSA 2nd 1/4 2 5 84 70 16.80 1.2 1 0.2424.1.01 Outside 2nd 1/4 2 6 0 0 0 0 0 026.01.01 FSA New moon 2 9 0.8 1 0.09 2 2 0.3330.1.01 Outside New moon 1 2.5 2.4 5 0.96 0 0 031.01.01 Outside New moon 1 3 3.1 4 1.03 0 0 02.2.01 Outside 1st 1/4 1 3.5 0 0 0 0.5 1 0.143.2.01 Outside 1st 1/4 1 2.5 1.3 5 0.52 1.6 1 0.6418.2.01 Outside 2nd 1/4 2 4 4.1 5 1.03 5.2 1 1.321.2.01 Outside 2nd 1/4 2 8 0.2 1 0.03 3.2 3 0.43.3.01 Outside New moon 2 6.5 0 0 0 0.6 1 0.094.3.01 FSA 1st 1/4 2 6 2.9 4 0.48 4.5 1 0.755.3.01 Outside 1st 1/4 2 6 1.7 3 0.28 3.7 3 0.636.3.01 Outside 1st 1/4 1 4 0 0 0 0 0 08.3.01 Outside 1st 1/4 2 6 0 0 0 2 2 0.3311.3.01 Outside Full moon 2 5.5 3.2 3 0.58 2.5 3 0.4512.3.01 Outside Full moon 2 8 3.1 2 0.39 4.9 3 0.3913.3.01 FSA Full moon 1 4 1.5 2 0.38 6.1 1 1.5314.3.01 Outside Full moon 2 6 1.5 2 0.25 1.7 2 0.2815.3.01 Outside Full moon 2 6.5 3.8 3 0.58 1.8 1 0.2816.3.01 Outside 2nd 1/4 2 6 0.5 1 0.08 21.5 6 3.5817.3.01 Outside 2nd 1/4 1 3 0 0 0 0 0 018.3.01 Outside 2nd 1/4 2 4 0 0 0 0.7 1 0.1819.3.01 FSA 2nd 1/4 2 4 4 3 1.00 4 1 121.3.01 FSA 2nd 1/4 2 6 26.8 18 4.47 0 0 022.3.01 FSA 2nd 1/4 2 5.5 29.2 24 5.31 8.5 3 1.5522.3.01 Outside 2nd 1/4 2 6.5 0 0 0 0 0 023.3.01 FSA 2nd 1/4 2 5.5 70.6 61 12.84 0 0 024.03.01 FSA 2nd 1/4 2 6 85.3 71 14.22 7.6 4 1.2725.03.01 Outside New moon 1 3 0 0 0 1.3 1 0.4326.03.01 FSA New moon 2 6 1.4 2 0.23 9.5 5 1.5829.3.01 Outside New moon 1 2.5 0 0 0 0 0 030.3.01 Outside New moon 1 3 0 0 0 11.4 4 3.831.3.01 Outside New moon 1 3 1.9 1 0.63 1.2 1 0.41.4.01 FSA New moon 2 5.5 2.4 3 0.44 0 0 07.4.01 Outside 1st 1/4 2 6 3.4 5 0.57 0 0 08.4.01 Outside Full moon 1 3 3.2 5 1.07 0 0 011.4.01 Outside Full moon 1 2.5 1.7 1 0.68 3.1 3 1.2412.4.01 Outside Full moon 1 2.5 1.3 3 0.53 0.3 1 0.2318.4.01 Outside 2nd 1/4 1 3 0 0 0 0 0 019.4.01 Outside 2nd 1/4 1 3 0 0 0 0 0 020.4.01 Outside 2nd 1/4 2 7.5 0 0 0 0 0 026.12.05 FSA 2nd 1/4 2 8 74.5 ? 9.3 ?28.12.05 FSA 2nd 1/4 2 8 87.1 ? 10.9 ?9.4.10 FSA 2nd 1/4 2 8 72.5 ? 9.1 ?
6 R.J. Hamilton et al. / Biological Conservation xxx (2011) xxx–xxx
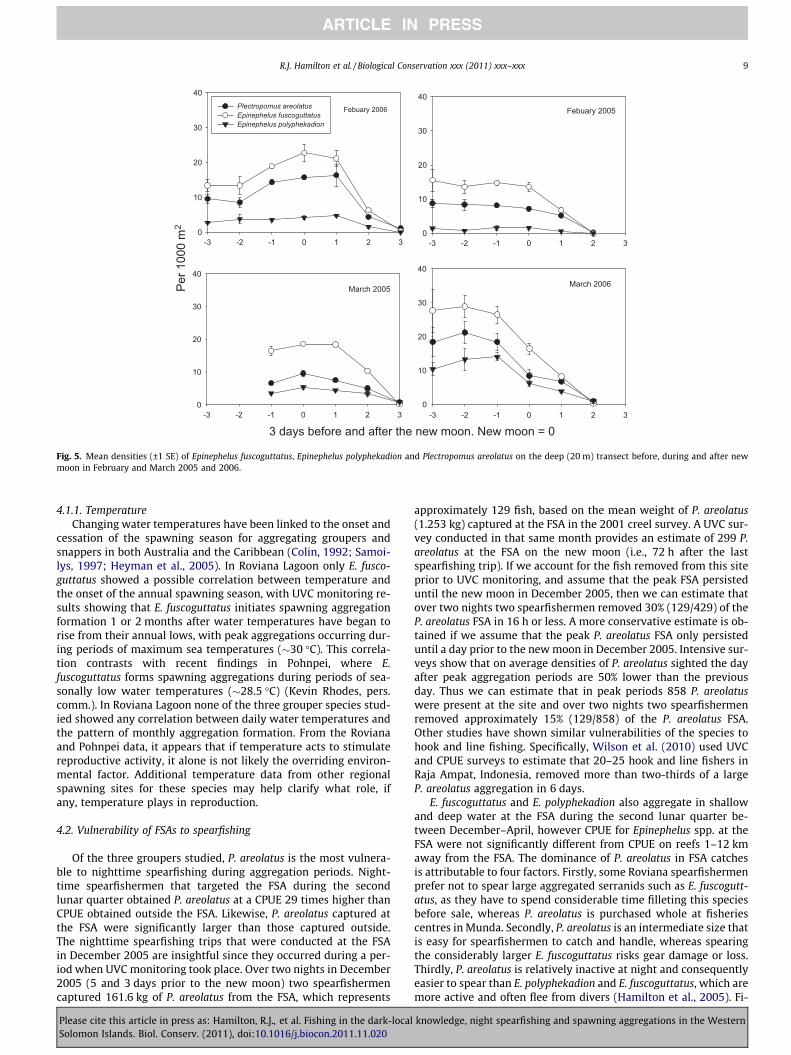
prior to the new moon. For these two species no clear aggregatingpattern was obvious in April 2006, as by then the spawning seasonfor that year had ended. In March 2006 P. areolatus were alreadyaggregated on the full moon, with peak densities sighted 1–5 daysprior to the new moon. In April 2006 the P. areolatus aggregationwas smaller and the pattern of FSA formation differed somewhat,with P. areolatus beginning to aggregate just after the full moon,and peak densities occurring 1–6 days prior to the new moon.Additional intensive surveys that were conducted around thenew moon period in February and March 2005 and February2006 reveal that the lunar day on which FSAs begin to decline afterpresumed spawning can vary, with FSAs declining on the newmoon or 1 or 2 days after new moon (Fig. 5).
3.2.4. TemperatureThe onset of the E. fuscoguttatus reproductive season began
shortly after sea temperatures had begun to rise from their annuallows, with peak spawning months occurring during periods ofmaximum sea temperatures (Fig. 3). E. polyphekadion FSAs also oc-curred during periods when sea temperatures were near theirmaximum. Mean monthly sea temperatures had no clear associa-tion on the formation of P. areolatus FSAs (Fig. 3). There was no cor-
Please cite this article in press as: Hamilton, R.J., et al. Fishing in the dark-localSolomon Islands. Biol. Conserv. (2011), doi:10.1016/j.biocon.2011.11.020
relation between daily water temperatures and the pattern ofmonthly aggregation formation for E. fuscoguttatus, E. polyphekadi-on or P. areolatus (Fig. 4).
3.3. Aggregation areas and total population size estimates
In March 2005 the total FSA area was 36,503 m2 (shallow stra-tum = 8698 m2, deep stratum = 27,805 m2). In March 2006 FSAs oc-curred over a slightly larger horizontal reef area, and as such, thetotal FSA was 38,462 m2 (shallow stratum = 9157 m2, deep stra-tum = 29,269 m2). Estimates of the total number of E. fuscoguttatus,E. polyphekadion and P. areolatus present at the FSA on the newmoon from March 2005 to May 2006 are presented in Table 4.
4. Discussion
4.1. Aggregation seasons
Between 100 and 700 E. fuscoguttatus aggregated at the studysite in the week leading up to the new moon during the monthsof December–April, and 200–300 E. polyphekadion aggregated inthe week leading up to the new moon in the months of February
knowledge, night spearfishing and spawning aggregations in the Western
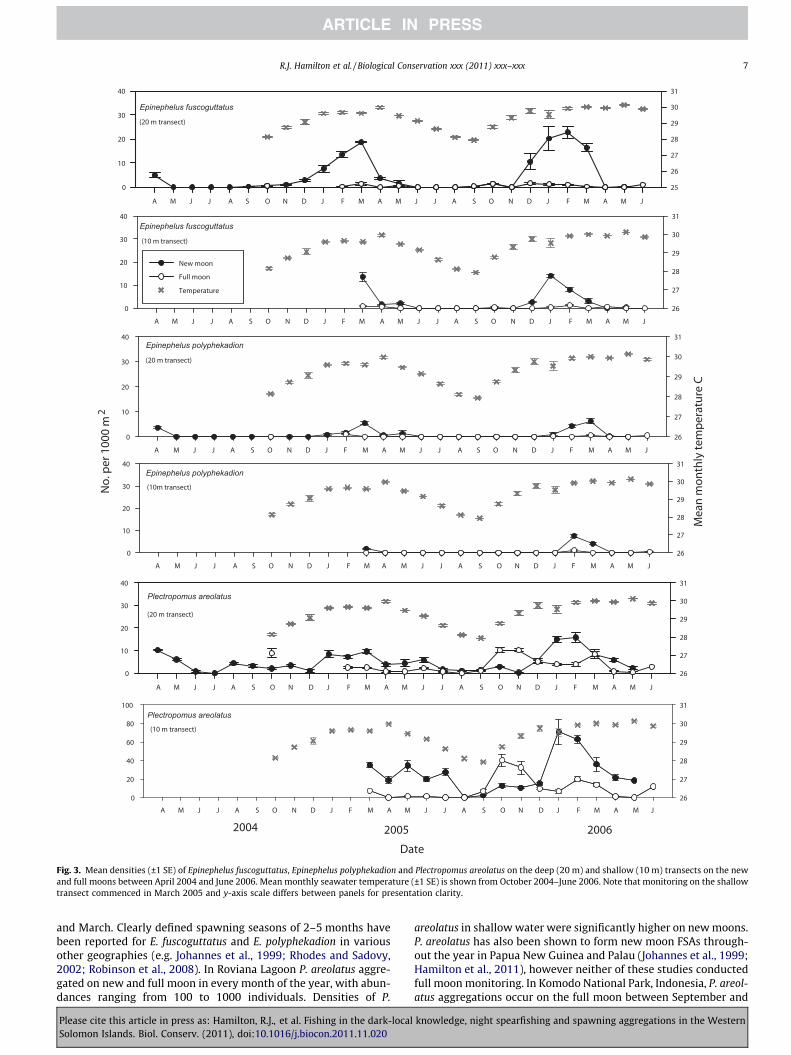
Fig. 3. Mean densities (±1 SE) of Epinephelus fuscoguttatus, Epinephelus polyphekadion and Plectropomus areolatus on the deep (20 m) and shallow (10 m) transects on the newand full moons between April 2004 and June 2006. Mean monthly seawater temperature (±1 SE) is shown from October 2004–June 2006. Note that monitoring on the shallowtransect commenced in March 2005 and y-axis scale differs between panels for presentation clarity.
R.J. Hamilton et al. / Biological Conservation xxx (2011) xxx–xxx 7
and March. Clearly defined spawning seasons of 2–5 months havebeen reported for E. fuscoguttatus and E. polyphekadion in variousother geographies (e.g. Johannes et al., 1999; Rhodes and Sadovy,2002; Robinson et al., 2008). In Roviana Lagoon P. areolatus aggre-gated on new and full moon in every month of the year, with abun-dances ranging from 100 to 1000 individuals. Densities of P.
Please cite this article in press as: Hamilton, R.J., et al. Fishing in the dark-localSolomon Islands. Biol. Conserv. (2011), doi:10.1016/j.biocon.2011.11.020
areolatus in shallow water were significantly higher on new moons.P. areolatus has also been shown to form new moon FSAs through-out the year in Papua New Guinea and Palau (Johannes et al., 1999;Hamilton et al., 2011), however neither of these studies conductedfull moon monitoring. In Komodo National Park, Indonesia, P. areol-atus aggregations occur on the full moon between September and
knowledge, night spearfishing and spawning aggregations in the Western
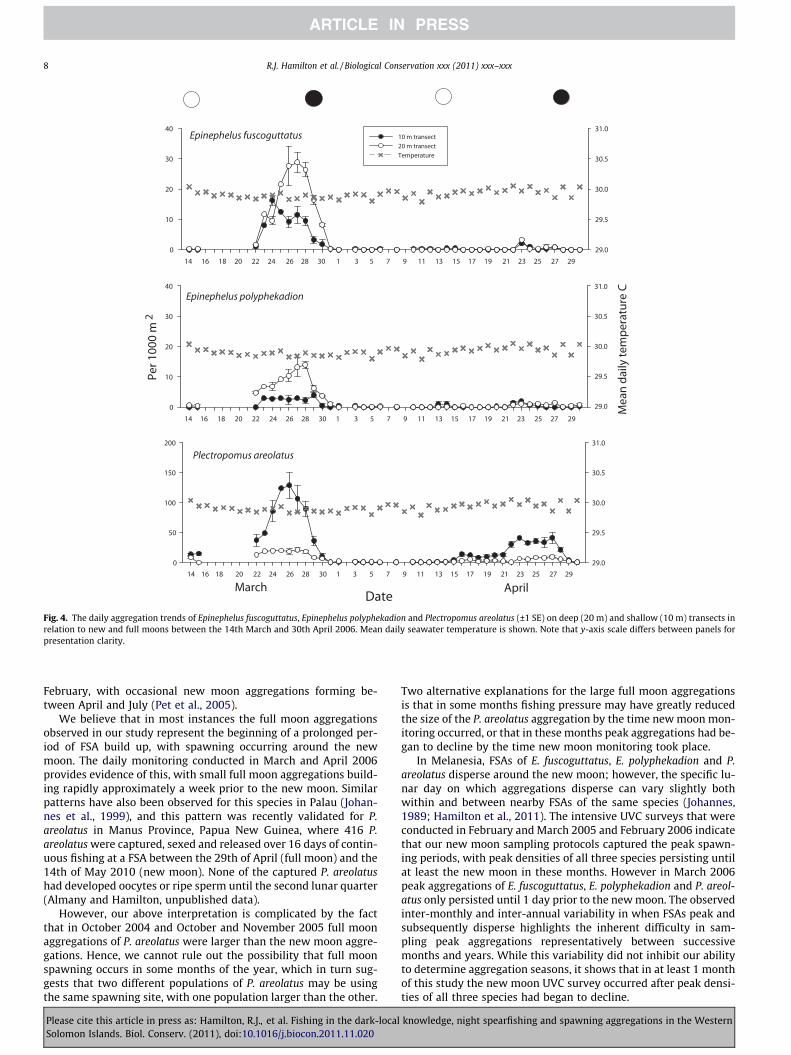
Fig. 4. The daily aggregation trends of Epinephelus fuscoguttatus, Epinephelus polyphekadion and Plectropomus areolatus (±1 SE) on deep (20 m) and shallow (10 m) transects inrelation to new and full moons between the 14th March and 30th April 2006. Mean daily seawater temperature is shown. Note that y-axis scale differs between panels forpresentation clarity.
8 R.J. Hamilton et al. / Biological Conservation xxx (2011) xxx–xxx
February, with occasional new moon aggregations forming be-tween April and July (Pet et al., 2005).
We believe that in most instances the full moon aggregationsobserved in our study represent the beginning of a prolonged per-iod of FSA build up, with spawning occurring around the newmoon. The daily monitoring conducted in March and April 2006provides evidence of this, with small full moon aggregations build-ing rapidly approximately a week prior to the new moon. Similarpatterns have also been observed for this species in Palau (Johan-nes et al., 1999), and this pattern was recently validated for P.areolatus in Manus Province, Papua New Guinea, where 416 P.areolatus were captured, sexed and released over 16 days of contin-uous fishing at a FSA between the 29th of April (full moon) and the14th of May 2010 (new moon). None of the captured P. areolatushad developed oocytes or ripe sperm until the second lunar quarter(Almany and Hamilton, unpublished data).
However, our above interpretation is complicated by the factthat in October 2004 and October and November 2005 full moonaggregations of P. areolatus were larger than the new moon aggre-gations. Hence, we cannot rule out the possibility that full moonspawning occurs in some months of the year, which in turn sug-gests that two different populations of P. areolatus may be usingthe same spawning site, with one population larger than the other.
Please cite this article in press as: Hamilton, R.J., et al. Fishing in the dark-localSolomon Islands. Biol. Conserv. (2011), doi:10.1016/j.biocon.2011.11.020
Two alternative explanations for the large full moon aggregationsis that in some months fishing pressure may have greatly reducedthe size of the P. areolatus aggregation by the time new moon mon-itoring occurred, or that in these months peak aggregations had be-gan to decline by the time new moon monitoring took place.
In Melanesia, FSAs of E. fuscoguttatus, E. polyphekadion and P.areolatus disperse around the new moon; however, the specific lu-nar day on which aggregations disperse can vary slightly bothwithin and between nearby FSAs of the same species (Johannes,1989; Hamilton et al., 2011). The intensive UVC surveys that wereconducted in February and March 2005 and February 2006 indicatethat our new moon sampling protocols captured the peak spawn-ing periods, with peak densities of all three species persisting untilat least the new moon in these months. However in March 2006peak aggregations of E. fuscoguttatus, E. polyphekadion and P. areol-atus only persisted until 1 day prior to the new moon. The observedinter-monthly and inter-annual variability in when FSAs peak andsubsequently disperse highlights the inherent difficulty in sam-pling peak aggregations representatively between successivemonths and years. While this variability did not inhibit our abilityto determine aggregation seasons, it shows that in at least 1 monthof this study the new moon UVC survey occurred after peak densi-ties of all three species had began to decline.
knowledge, night spearfishing and spawning aggregations in the Western
March 2005
-3 -2 -1 0 1 2 30
10
20
30
40March 2006
-3 -2 -1 0 1 2 30
10
20
30
40
Febuary 2005
-3 -2 -1 0 1 2 3
Per 1
000
m2
0
10
20
30
40
3 days before and after the new moon. New moon = 0
Febuary 2006
-3 -2 -1 0 1 2 30
10
20
30
40Plectropomus areolatusEpinephelus fuscoguttatusEpinephelus polyphekadion
Fig. 5. Mean densities (±1 SE) of Epinephelus fuscoguttatus, Epinephelus polyphekadion and Plectropomus areolatus on the deep (20 m) transect before, during and after newmoon in February and March 2005 and 2006.
R.J. Hamilton et al. / Biological Conservation xxx (2011) xxx–xxx 9
4.1.1. TemperatureChanging water temperatures have been linked to the onset and
cessation of the spawning season for aggregating groupers andsnappers in both Australia and the Caribbean (Colin, 1992; Samoi-lys, 1997; Heyman et al., 2005). In Roviana Lagoon only E. fusco-guttatus showed a possible correlation between temperature andthe onset of the annual spawning season, with UVC monitoring re-sults showing that E. fuscoguttatus initiates spawning aggregationformation 1 or 2 months after water temperatures have began torise from their annual lows, with peak aggregations occurring dur-ing periods of maximum sea temperatures (�30 �C). This correla-tion contrasts with recent findings in Pohnpei, where E.fuscoguttatus forms spawning aggregations during periods of sea-sonally low water temperatures (�28.5 �C) (Kevin Rhodes, pers.comm.). In Roviana Lagoon none of the three grouper species stud-ied showed any correlation between daily water temperatures andthe pattern of monthly aggregation formation. From the Rovianaand Pohnpei data, it appears that if temperature acts to stimulatereproductive activity, it alone is not likely the overriding environ-mental factor. Additional temperature data from other regionalspawning sites for these species may help clarify what role, ifany, temperature plays in reproduction.
4.2. Vulnerability of FSAs to spearfishing
Of the three groupers studied, P. areolatus is the most vulnera-ble to nighttime spearfishing during aggregation periods. Night-time spearfishermen that targeted the FSA during the secondlunar quarter obtained P. areolatus at a CPUE 29 times higher thanCPUE obtained outside the FSA. Likewise, P. areolatus captured atthe FSA were significantly larger than those captured outside.The nighttime spearfishing trips that were conducted at the FSAin December 2005 are insightful since they occurred during a per-iod when UVC monitoring took place. Over two nights in December2005 (5 and 3 days prior to the new moon) two spearfishermencaptured 161.6 kg of P. areolatus from the FSA, which represents
Please cite this article in press as: Hamilton, R.J., et al. Fishing in the dark-localSolomon Islands. Biol. Conserv. (2011), doi:10.1016/j.biocon.2011.11.020
approximately 129 fish, based on the mean weight of P. areolatus(1.253 kg) captured at the FSA in the 2001 creel survey. A UVC sur-vey conducted in that same month provides an estimate of 299 P.areolatus at the FSA on the new moon (i.e., 72 h after the lastspearfishing trip). If we account for the fish removed from this siteprior to UVC monitoring, and assume that the peak FSA persisteduntil the new moon in December 2005, then we can estimate thatover two nights two spearfishermen removed 30% (129/429) of theP. areolatus FSA in 16 h or less. A more conservative estimate is ob-tained if we assume that the peak P. areolatus FSA only persisteduntil a day prior to the new moon in December 2005. Intensive sur-veys show that on average densities of P. areolatus sighted the dayafter peak aggregation periods are 50% lower than the previousday. Thus we can estimate that in peak periods 858 P. areolatuswere present at the site and over two nights two spearfishermenremoved approximately 15% (129/858) of the P. areolatus FSA.Other studies have shown similar vulnerabilities of the species tohook and line fishing. Specifically, Wilson et al. (2010) used UVCand CPUE surveys to estimate that 20–25 hook and line fishers inRaja Ampat, Indonesia, removed more than two-thirds of a largeP. areolatus aggregation in 6 days.
E. fuscoguttatus and E. polyphekadion also aggregate in shallowand deep water at the FSA during the second lunar quarter be-tween December–April, however CPUE for Epinephelus spp. at theFSA were not significantly different from CPUE on reefs 1–12 kmaway from the FSA. The dominance of P. areolatus in FSA catchesis attributable to four factors. Firstly, some Roviana spearfishermenprefer not to spear large aggregated serranids such as E. fuscogutt-atus, as they have to spend considerable time filleting this speciesbefore sale, whereas P. areolatus is purchased whole at fisheriescentres in Munda. Secondly, P. areolatus is an intermediate size thatis easy for spearfishermen to catch and handle, whereas spearingthe considerably larger E. fuscoguttatus risks gear damage or loss.Thirdly, P. areolatus is relatively inactive at night and consequentlyeasier to spear than E. polyphekadion and E. fuscoguttatus, which aremore active and often flee from divers (Hamilton et al., 2005). Fi-
knowledge, night spearfishing and spawning aggregations in the Western
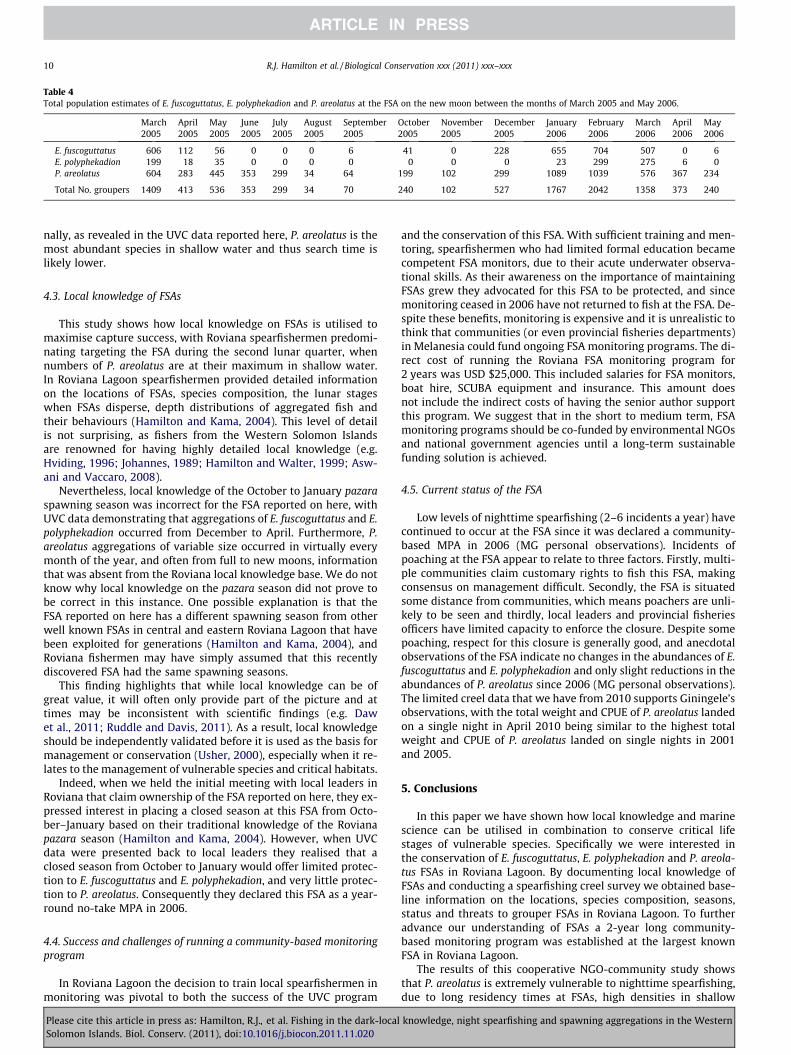
Table 4Total population estimates of E. fuscoguttatus, E. polyphekadion and P. areolatus at the FSA on the new moon between the months of March 2005 and May 2006.
March2005
April2005
May2005
June2005
July2005
August2005
September2005
October2005
November2005
December2005
January2006
February2006
March2006
April2006
May2006
E. fuscoguttatus 606 112 56 0 0 0 6 41 0 228 655 704 507 0 6E. polyphekadion 199 18 35 0 0 0 0 0 0 0 23 299 275 6 0P. areolatus 604 283 445 353 299 34 64 199 102 299 1089 1039 576 367 234
Total No. groupers 1409 413 536 353 299 34 70 240 102 527 1767 2042 1358 373 240
10 R.J. Hamilton et al. / Biological Conservation xxx (2011) xxx–xxx
nally, as revealed in the UVC data reported here, P. areolatus is themost abundant species in shallow water and thus search time islikely lower.
4.3. Local knowledge of FSAs
This study shows how local knowledge on FSAs is utilised tomaximise capture success, with Roviana spearfishermen predomi-nating targeting the FSA during the second lunar quarter, whennumbers of P. areolatus are at their maximum in shallow water.In Roviana Lagoon spearfishermen provided detailed informationon the locations of FSAs, species composition, the lunar stageswhen FSAs disperse, depth distributions of aggregated fish andtheir behaviours (Hamilton and Kama, 2004). This level of detailis not surprising, as fishers from the Western Solomon Islandsare renowned for having highly detailed local knowledge (e.g.Hviding, 1996; Johannes, 1989; Hamilton and Walter, 1999; Asw-ani and Vaccaro, 2008).
Nevertheless, local knowledge of the October to January pazaraspawning season was incorrect for the FSA reported on here, withUVC data demonstrating that aggregations of E. fuscoguttatus and E.polyphekadion occurred from December to April. Furthermore, P.areolatus aggregations of variable size occurred in virtually everymonth of the year, and often from full to new moons, informationthat was absent from the Roviana local knowledge base. We do notknow why local knowledge on the pazara season did not prove tobe correct in this instance. One possible explanation is that theFSA reported on here has a different spawning season from otherwell known FSAs in central and eastern Roviana Lagoon that havebeen exploited for generations (Hamilton and Kama, 2004), andRoviana fishermen may have simply assumed that this recentlydiscovered FSA had the same spawning seasons.
This finding highlights that while local knowledge can be ofgreat value, it will often only provide part of the picture and attimes may be inconsistent with scientific findings (e.g. Dawet al., 2011; Ruddle and Davis, 2011). As a result, local knowledgeshould be independently validated before it is used as the basis formanagement or conservation (Usher, 2000), especially when it re-lates to the management of vulnerable species and critical habitats.
Indeed, when we held the initial meeting with local leaders inRoviana that claim ownership of the FSA reported on here, they ex-pressed interest in placing a closed season at this FSA from Octo-ber–January based on their traditional knowledge of the Rovianapazara season (Hamilton and Kama, 2004). However, when UVCdata were presented back to local leaders they realised that aclosed season from October to January would offer limited protec-tion to E. fuscoguttatus and E. polyphekadion, and very little protec-tion to P. areolatus. Consequently they declared this FSA as a year-round no-take MPA in 2006.
4.4. Success and challenges of running a community-based monitoringprogram
In Roviana Lagoon the decision to train local spearfishermen inmonitoring was pivotal to both the success of the UVC program
Please cite this article in press as: Hamilton, R.J., et al. Fishing in the dark-localSolomon Islands. Biol. Conserv. (2011), doi:10.1016/j.biocon.2011.11.020
and the conservation of this FSA. With sufficient training and men-toring, spearfishermen who had limited formal education becamecompetent FSA monitors, due to their acute underwater observa-tional skills. As their awareness on the importance of maintainingFSAs grew they advocated for this FSA to be protected, and sincemonitoring ceased in 2006 have not returned to fish at the FSA. De-spite these benefits, monitoring is expensive and it is unrealistic tothink that communities (or even provincial fisheries departments)in Melanesia could fund ongoing FSA monitoring programs. The di-rect cost of running the Roviana FSA monitoring program for2 years was USD $25,000. This included salaries for FSA monitors,boat hire, SCUBA equipment and insurance. This amount doesnot include the indirect costs of having the senior author supportthis program. We suggest that in the short to medium term, FSAmonitoring programs should be co-funded by environmental NGOsand national government agencies until a long-term sustainablefunding solution is achieved.
4.5. Current status of the FSA
Low levels of nighttime spearfishing (2–6 incidents a year) havecontinued to occur at the FSA since it was declared a community-based MPA in 2006 (MG personal observations). Incidents ofpoaching at the FSA appear to relate to three factors. Firstly, multi-ple communities claim customary rights to fish this FSA, makingconsensus on management difficult. Secondly, the FSA is situatedsome distance from communities, which means poachers are unli-kely to be seen and thirdly, local leaders and provincial fisheriesofficers have limited capacity to enforce the closure. Despite somepoaching, respect for this closure is generally good, and anecdotalobservations of the FSA indicate no changes in the abundances of E.fuscoguttatus and E. polyphekadion and only slight reductions in theabundances of P. areolatus since 2006 (MG personal observations).The limited creel data that we have from 2010 supports Giningele’sobservations, with the total weight and CPUE of P. areolatus landedon a single night in April 2010 being similar to the highest totalweight and CPUE of P. areolatus landed on single nights in 2001and 2005.
5. Conclusions
In this paper we have shown how local knowledge and marinescience can be utilised in combination to conserve critical lifestages of vulnerable species. Specifically we were interested inthe conservation of E. fuscoguttatus, E. polyphekadion and P. areola-tus FSAs in Roviana Lagoon. By documenting local knowledge ofFSAs and conducting a spearfishing creel survey we obtained base-line information on the locations, species composition, seasons,status and threats to grouper FSAs in Roviana Lagoon. To furtheradvance our understanding of FSAs a 2-year long community-based monitoring program was established at the largest knownFSA in Roviana Lagoon.
The results of this cooperative NGO-community study showsthat P. areolatus is extremely vulnerable to nighttime spearfishing,due to long residency times at FSAs, high densities in shallow
knowledge, night spearfishing and spawning aggregations in the Western

R.J. Hamilton et al. / Biological Conservation xxx (2011) xxx–xxx 11
water and the ease with which it can be approached at night.Clearly FSAs of P. areolatus need to be protected from nighttimespearfishing wherever it is practiced. Conversely, the depth distri-butions and diver avoidance behaviour of deeper-dwelling E. fusco-guttatus and E. polyphekadion at FSAs offers them a greater degreeof protection from free diving spearfishers. However both E.polyphekadion, and to a lesser extent E. fuscoguttatus, are highlysusceptible to hook and line fisheries (Rhodes and Sadovy, 2002;Rhodes et al., 2011), and are therefore also deserving of protection.
In Roviana Lagoon the monitoring program served as an effec-tive vehicle for building community support for managing theFSA, and it also provided an independent validation of local knowl-edge on grouper spawning seasons. Results of the monitoringstudy showed that while local knowledge can be highly detailed,like all knowledge systems, it is not infallible (Johannes et al.,2000), with science-based underwater monitoring building a morecomplete picture of FSA seasons than local knowledge alone.Clearly, local knowledge is most useful in management and conser-vation if it is carefully collected, evaluated and validated, and thecultural sensitivities and confidentiality of local knowledge is re-spected (Ruddle et al., 1992; Usher, 2000; Daw, 2008; Hamiltonet al., 2012).
In this study it was local knowledge and the scientific results ofthe community-based monitoring program that led to the develop-ment of appropriate management measures for this FSA, with theaggregation declared a year-round community-based MPA in2006. Finally, while we cannot prove that the conservation effortsreported on here have assisted in the FSA remaining healthy, onething is clear; this site is one of the largest known multi-speciesFSAs in Melanesia, and its ongoing preservation is critical for thelong-term health of grouper fisheries in Roviana Lagoon.
Acknowledgments
First and foremost we would like to thank W. Kama for assistingwith the local knowledge survey and FSA monitoring. We alsothank S. Baso, G. Gadepeta and S. Kari for assisting with FSA mon-itoring. We are grateful to the Munda area communities who al-lowed us to work on their reefs. We thank N. Peterson forproducing Fig. 1, and G. Almany and K. Ruddle for improving anearlier version of this manuscript. Funding for this project was pro-vided by Oak Foundation, The David and Lucile Packard Foundationand The John D. and Catherine T. MacArthur Foundation.
References
Almany, G., Hamilton, R., Williamson, D., Evans, R., Jones, G., Matawai, M., Potuku, T.,Rhodes, K., Russ, G., Sawynok, B., 2010. Research partnerships with localcommunities: two case studies from Papua New Guinea and Australia. CoralReefs 29, 567–576.
Aswani, S., Hamilton, R.J., 2004. Integrating indigenous ecological knowledge andcustomary sea tenure with marine and social science for conservation ofBumphead Parrotfish (Bolbometopon muricatum) in the Roviana Lagoon,Solomon Islands. Environmental Conservation 31, 69–83.
Aswani, S., Albert, S., Sabetian, A., Furusawa, T., 2007. Customary management asprecautionary and adaptive principles for protecting coral reefs in Oceania.Coral Reefs 26, 1009–1021.
Aswani, S., Vaccaro, I., 2008. Lagoon ecology and social strategies: habitat diversityand ethnobiology. Human Ecology 36, 325–341.
Beets, J., Friedlander, A., 1999. Evaluation of a conservation strategy: a spawningaggregation closure for red hind, Epinephelus guttatus, in the US Virgin Islands.Environmental Biology of Fishes 55, 91–98.
Colin, P.L., 1992. Reproduction of the Nassau grouper, Epinephelus striatus (Pisces:Serranidae) and its relationship to environmental conditions. EnvironmentalBiology of Fishes 34, 357–377.
Colin, P.L., Sadovy, Y.J., Domeier, M.L., 2003. Manual for the Study and Conservationof Reef Fish Spawning Aggregations. Society for the Conservation of Reef FishAggregations.
Cornish, A., 2004. Epinephelus fuscoguttatus. In: IUCN 2010. IUCN Red List ofThreatened Species. Version 2010.2.
Daw, T.M., 2008. How Fishers Count: Engaging Fishers’ Knowledge in FisheriesScience and Management. Ph.D. Thesis, School of Marine Science and
Please cite this article in press as: Hamilton, R.J., et al. Fishing in the dark-localSolomon Islands. Biol. Conserv. (2011), doi:10.1016/j.biocon.2011.11.020
Technology, and School of Geography, Politics and Sociology, NewcastleUniversity, Newcastle upon Tyne, UK. <http://www.tinyurl.com/tdaw-thesis>(www document).
Daw, T.M., Robinson, J., Graham, N.A.J., 2011. Perceptions of trends in Seychellesartisanal trap fisheries: comparing catch monitoring, underwater visual censusand fishers’ knowledge. Environmental Conservation 38, 75–88.
Drew, J.A., 2005. Use of traditional ecological knowledge in marine conservation.Conservation Biology 19, 1286–1293.
Dulvy, N.K., Polunin, N.V.C., 2004. Using informal knowledge to infer human-induced rarity of a conspicuous reef fish. Animal Conservation 7, 365–374.
Game, E., Lipsett-Moore, G., Hamilton, R., Peterson, N., Atu, W., Kereseka, J., Watts,M., Possingham, H., 2011. Informed opportunism in practice; conservationplanning in the Solomon Islands. Conservation Letters 4, 38–46.
Gillett, R., Moy, W., 2006. Spearfishing in the Pacific Islands. Current Status andManagement Issues. FAO/FishCode Review No. 19. FAO, Rome.
Grandcourt, E., 2005. Demographic Characteristics of Selected EpinephelineGroupers (Family: Serranidae; Subfamily: Epinephelinae) from Aldabra Atoll,Seychelles. Atoll Research Bulletin No. 539.
Hamilton, R.J., Kama, W., 2004. Spawning Aggregations of Coral Reef Fish in RovianaLagoon, Western Province, Solomon Islands: Local Knowledge Field SurveyReport. (Unrestricted Access Version). Report Prepared for the Pacific IslandCountries Coastal Marine Program, The Nature Conservancy. TNC Pacific IslandCountries Report No. 5/04.
Hamilton, R.J., Matawai, M., 2006. Live reef food fish trade causes rapid declines inabundance of squaretail coralgrouper (Plectropomus areolatus) at a spawningaggregation site in Manus, Papua New Guinea. SPC Live Reef Fish InformationBulletin 16, 13–18.
Hamilton, R.J., Matawai, M., Potuku, T., Kama, W., Lahui, P., Warku, J., Smith, A.J.,2005. Applying local knowledge and science to the management of grouperaggregation sites in Melanesia. SPC Live Reef Fish Information Bulletin 14,7–19.
Hamilton, R.J., Potuku, T., Montambault, J., 2011. Community-based conservationresults in the recovery of reef fish spawning aggregations in the Coral Triangle.Biological Conservation 144, 1850–1858.
Hamilton, R.J., Sadovy de Mitcheson, Y., Aguilar-Perera, A., 2012. The role oflocal ecological knowledge in the conservation and management of reeffish spawning aggregations. In: Sadovy de Mitcheson, Y., Colin, P.L. (Eds.),Reef Fish Spawning Aggregations: Biology, Fisheries and Management. Fishand Fisheries Series 35. Springer Science + Business Media B.V., pp. 331–369.
Hamilton, R.J., Walter, R., 1999. Indigenous ecological knowledge and its role infisheries research design. A case study from Roviana Lagoon, Western Province,Solomon Islands. SPC Traditional Marine Resource Management and KnowledgeBulletin 11, 13–25.
Heyman, W.D., Ecochard, J.L.B., Biasi, F.B., 2007. Low-cost bathymetric mapping fortropical marine conservation—a focus on reef fish spawning aggregation sites.Marine Geodesy 30, 37–50.
Heyman, W.D., Kjerfve, B., Graham, R.T., Rhodes, K.L., Garbutt, L., 2005. Spawningaggregations of Lutjanus cyanopterus (Cuvier) on the Belize Barrier Reef over a 6year period. Journal of Fish Biology 67, 83–101.
Hviding, E., 1996. Guardians of Marovo Lagoon, Practice, Place, and Politics inMaritime Melanesia. University of Hawaii Press, Honolulu, Hawaii.
Johannes, R.E., 1978. Reproductive strategies of coastal marine fishes in the tropics.Environmental Biology of Fishes 3, 65–84.
Johannes, R.E., 1989. Spawning aggregations of the grouper Plectropomus areolatus(Ruppell) in the Solomon Islands. In: Choat, J.H., Barnes, D.J., Borowitzka, M.A.,Coll, J.C., Davies, P.J., Flood, P., Hatcher, B.G., Hopley, D., Hutchings, P.A., et al.(Eds.), Proceedings of the Sixth International Coral Reef Symposium, Townsville,Australia, pp. 751–755.
Johannes, R.E., Freeman, M.R., Hamilton, R.J., 2000. Ignore fishers’ knowledge andmiss the boat. Fish and Fisheries 1, 257–271.
Johannes, R.E., Neis, B., 2007. The value of anecdote. In: Haggan, N., Neis, B., Baird,I.G. (Eds.), Fishers’ Knowledge in Fisheries Science and Management. UNESCOPublishing, Paris.
Johannes, R.E., Squire, L., Graham, T., Sadovy, Y., Renguul, H., 1999. SpawningAggregations of Groupers (Serranidae) in Palau. Marine Research SeriesPublication No. 1. The Nature Conservancy, Arlington.
Nemeth, R.S., 2005. Population characteristics of a recovering US Virgin Islands redhind spawning aggregation following protection. Marine Ecology ProgressSeries 286, 81–97.
Pet, J.S., Mous, P.J., Muljadi, A.H., Sadovy, Y.J., Squire, L., 2005. Aggregations ofPlectropomus areolatus and Epinephelus fuscoguttatus (groupers, Serranidae) inthe Komodo National Park, Indonesia: monitoring and implications formanagement. Environmental Biology of Fishes 74, 209–218.
Pet, J.S., Mous, P.J., Rhodes, K., Green, A., 2006. Introduction to Monitoring ofSpawning Aggregations of Three Grouper Species from the Indo-Pacific. AManual for Field Practitioners. Version 2.0 (January 2006). The NatureConservancy Coral Triangle Center, Sanur, Bali, Indonesia, 98 p.
Robinson, J., Aumeeruddy, R., Jorgensen, T.L., Ohman, M.C., 2008. Dynamics ofcamouflage (Epinephelus polyphekadion) and brown marbled grouper(Epinephelus fuscoguttatus) spawning aggregations at a remote reef site,Seychelles. Bulletin of Marine Science 83, 415–431.
Rhodes, K.L., 2004. Solomon Islands Spawning Aggregation Monitoring TrainingWorkshop Report, Gizo, Western Province, Solomon Islands, 13–21 March 2004.Report Prepared for the Pacific Island Countries Coastal Marine Program, TheNature Conservancy. TNC Pacific Island Countries Report No. 2/04.
knowledge, night spearfishing and spawning aggregations in the Western

12 R.J. Hamilton et al. / Biological Conservation xxx (2011) xxx–xxx
Rhodes, K.L., Sadovy, Y., 2002. Temporal and spatial trends in spawningaggregations of camouflage grouper, Epinephelus polyphekadion (Bleeker 1849)in Pohnpei, Micronesia. Environmental Biology of Fishes 63, 27–39.
Rhodes, K.L., Taylor, B.M., McIIwain, J.L., 2011. Detailed demographic analysis of anEpinephelus polyphekadion spawning aggregation and fishery. Marine EcologyProgress Series 421, 183–198.
Rhodes, K.L., Tupper, M.H., 2007. A preliminary market-based analysis of thePohnpei, Micronesia, grouper (Serranidae: Epinepheline) fishery revealsunsustainable fishing practices. Coral Reefs 26, 335–344.
Ruddle, K., Davis, A., 2011. What is ‘‘Ecological’’ in local ecological knowledge?Lessons from Canada and Vietnam. Society and Natural Resources 24, 887–901.
Ruddle, K., Hviding, E., Johannes, R.E., 1992. Marine resources management in thecontext of customary tenure. Marine Resource Economics 7, 249–273.
Russell, B., Situ, A., Cornish, A., 2006. Epinephelus polyphekadion. In: IUCN 2010.IUCN Red List of Threatened Species. Version 2010.2.
Sadovy, Y., Cheung, W.L., 2003. Near extinction of a highly fecund fish: the one thatnearly got away. Fish and Fisheries 4, 86–89.
Sadovy de Mitcheson, Y.S., Cornish, A., Domeier, M., Colin, P.L., Russell, M.,Lindeman, K.C., 2008. A global baseline for spawning aggregations of reeffishes. Conservation Biology 22, 1233–1244.
Sadovy, Y., Domeier, M., 2005. Are aggregation-fisheries sustainable? Reef fishfisheries as a case study. Coral Reefs 24, 254–262.
Sadovy, Y.J., Donaldson, T.J., Graham, T.R., McGilvray, F., Muldoon, G.J., Phillips, M.J.,Rimme, M.A., Smith, A., Yeeting, B., 2003. While Stocks Last: The Live Reef FoodFish Trade. Asian Development Bank, Manila, 147 p.
Please cite this article in press as: Hamilton, R.J., et al. Fishing in the dark-localSolomon Islands. Biol. Conserv. (2011), doi:10.1016/j.biocon.2011.11.020
Saénz-Arroyo, A., Roberts, C.M., Torre, J., Carinõ-Olvera, M., 2005. Using fishers’anecdotes, naturalists’ observations and grey literature to reassess marinespecies at risk: the case of the Gulf grouper in the Gulf of California, Mexico. Fishand Fisheries 6, 121–133.
Samoilys, M.A., 1997. Periodicity of spawning aggregations of coral troutPlectropomus leopardus (Pisces: Serranidae) on the northern Great BarrierReef. Marine Ecology Progress Series 160, 149–159.
Silvano, R.A., MacCord, M.P., Lima, R.V., Begossi, A., 2006. When does this fishspawn? Fishermen’s local knowledge of migration and reproduction of Braziliancoastal fishes. Environmental Biology of Fishes 76, 371–386.
Taylor, R.B., Morrison, M.A., Shears, N.T., 2011. Establishing baselines for recovery ina marine reserve (Poor Knights Islands, New Zealand) using local ecologicalknowledge. Biological Conservation 144, 3038–3046.
Thierry, C., Sadovy, Y., Yeeting, B., 2008. Plectropomus areolatus. In: IUCN 2010. IUCNRed List of Threatened Species. Version 2010.2.
Usher, P.J., 2000. Traditional ecological knowledge in environmental assessmentand management. Arctic 53, 183–193.
Veron, J.E.N., Devantier, L.M., Turak, E., Green, A.L., Stuart Kininmonth, S., Stafford-Smith, M., Peterson, N., 2009. Delineating the Coral Triangle, Galaxea. Journal ofCoral Reef Studies 11, 91–100.
Wilson, J., Rhodes, K.L., Rotinsulu, C., 2010. Aggregation fishing and localmanagement within a marine protected area in Indonesia. SPC Live Reef FishInformation Bulletin 19, 7–13.
Zurk, L.M., Lotz, J., Ellis, T., McNames, J., Ecochard, J.L., 2006. Sonar mapping for coralreef conservation. Acoustical Society of America 120, 3060.
knowledge, night spearfishing and spawning aggregations in the Western