permanent aggregations of a pelagic predator at shallow seamounts
TRANSCRIPT

1 3
Mar Biol (2014) 161:1349–1360DOI 10.1007/s00227-014-2423-9
OrIgInal PaPer
Permanent aggregations of a pelagic predator at shallow seamounts
Jorge Fontes · Mara Schmiing · Pedro Afonso
received: 2 October 2013 / accepted: 8 March 2014 / Published online: 11 april 2014 © Springer-Verlag Berlin Heidelberg 2014
these sites in order to preserve these mature fish aggrega-tions with high reproductive potential.
Introduction
Jacks and trevallies, a group of large fishes from the Caran-gidae family, are a major element of any tropical or sub-tropical reef fish assemblage, where they play a major ecological role as pelagic predators (Friedlander and DeMartini 2002; Heithaus et al. 2008). Jacks and treval-lies are an important target and accessory catch of tropical artisanal fisheries (Meyer et al. 2001), a prized trophy for sports anglers and spearfishers, and an iconic species for recreational divers worldwide.
The benefits of marine reserves and other local scale conservation efforts targeting this group are normally per-ceived as difficult to achieve. This perception stems from the fact that carangidae are typically considered highly mobile and migratory species (Hutson et al. 2007). Ideally, reserves should protect the fishes’ home range or, at least, their core activity area in order to substantially reduce the chances of it being caught elsewhere (russ 2002). Conse-quently, the spatial protection offered by marine reserves will be very limited for carangids unless the reserves match the scale of their movements. This should seldom be the case, as the large majority of marine reserves established to protect reef habitats are small in size (Halpern 2003). Yet, surprisingly little research has been conducted on the spatial ecology of carangids. a few studies used visual census or tag-recapture methods (Tanaka 1984; McClel-lan and Cummings 1996; gillanders et al. 2001; Demp-ster and Kingsford 2003; Hutson et al. 2007; afonso et al. 2008b), but only a handful have used telemetry techniques to investigate the movements and patterns of habitat use in
Abstract The effectiveness of marine reserves for highly mobile reef fishes such as jacks and trevallies is normally assumed to be small, even though we generally lack the understanding of their long-term movement patterns. In this work, we combined the analysis of multi-year landings and underwater visual census with acoustic telemetry to investigate the long-term movement patterns (up to 4 years) of almaco jack, Seriola rivoliana, a large reef top preda-tor, in one protected and one unprotected offshore shallow seamount in the azores, central north atlantic. although the analysis of visual census and landing data suggests a pronounced seasonal pattern of occurrence, we found that tagged fish were almost continuously detected at the sea-mounts for up to four consecutive years. Some individuals showed a few long periods of undetection of 1–2 months in the Formigas reserve, mostly in the fall and winter. Mobil-ity within this seamount increased during spring and sum-mer, and geostrophic current speed was negatively cor-related with detection probability. Overall, the acoustic telemetry results showed that adult almaco jacks were resi-dent year-round, refuting the traditional view that these fish are essentially migratory and a seasonal occurrence in the azores. given the intrinsic vulnerability of resident fish to fishing, our results highlight the importance of protecting
Communicated by J. Houghton.
Electronic supplementary material The online version of this article (doi:10.1007/s00227-014-2423-9) contains supplementary material, which is available to authorized users.
J. Fontes (*) · M. Schmiing · P. afonso Department of Oceanography and Fisheries, Institute of Marine research, University of the azores, rua Prof. Doutor Frederico Machado, 9901-862 Horta, Portugale-mail: [email protected]

1350 Mar Biol (2014) 161:1349–1360
1 3
their natural habitats (Wetherbee et al. 2004; Meyer et al. 2007; afonso et al. 2009; Brown et al. 2010; Capello et al. 2012). results from these studies show that the scale of movements of carangid fishes can be substantially differ-ent both between and within species. as revealed by tag-recapture studies, some of them are capable of large dis-persal of up to thousands of km along continental shelves (McClellan and Cummings 1996; gillanders et al. 2001), and many species are also known to aggregate around off-shore floating structures (fish aggregating devices—FaDs) during their juvenile stages (Dempster and Kingsford 2003; Brown et al. 2010). Contrastingly, telemetry studies also showed that some carangids are resident within a reef area or an island (e.g. Holland et al. 1996; Meyer et al. 2007; afonso et al. 2009). regardless of scale, there seems to be a common ontogenetic or seasonal nature of their migra-tions, most probably associated with changes in biological requirements such as feeding and reproduction (Tanaka 1984; McClellan and Cummings 1996; Dempster 2005).
Current opinion that jacks and trevallies share a highly migratory and mobile behaviour that precludes an effec-tive protection through the establishment of marine pro-tected areas is mainly based on common perception and traditional ecological knowledge (TeK) of fishermen rather than relying upon scientific evidences. This emphasizes the challenge but also the need for assessing the movement patterns and home range sizes of highly mobile marine fish towards a more complete understanding of their ecol-ogy and responses to protection (Brown et al. 2010). Man-agement of exploited fishes exhibiting seasonal spawning migrations can be challenging, especially when they leave core activity areas inside small marine reserves and travel long distances to reach spawning sites (Bolden 2000). These fishes may also become even more vulnerable to fishing if they aggregate in large groups to spawn, as seems to be the case in some jacks and trevallies (e.g. Sala et al. 2003; Meyer et al. 2007; afonso et al. 2008b, 2009). If such spawning sites can be identified, then even small marine protected areas (MPas) can be used to protect fish aggregations at these locations and times of year (Sadovy and Domeier 2005a; De Mitcheson et al. 2008).
The almaco jack, Seriola rivoliana (Cuvier, 1833), inhabits the reefs of all subtropical regions (Smith-Vaniz 1986). Despite its wide distribution, the ecology of almaco jack has received little attention, and work so far has been limited to aspects of its biology (Manooch and Haimovici 1983; Barreiros et al. 2003) or presence in fish assemblages at natural habitats or oil rigs (Stanley and Wilson 1997, 2003). In the azores archipelago, central north atlantic, S. rivoliana is a frequent sighting, during summer, on reefs, island slopes, and offshore shallow seamounts down to at least 100 m, (afonso 2002). During this period, the almaco jack feeds mainly on pelagic juvenile fishes (Barreiros et al.
2003). as in other areas, this jack is an important resource for local artisanal pole-and-line fishery, with an annual average 50 tons landed from 2000 to 2011 (Portuguese Pro-gram for the Collection of Fisheries data/azores), as well as for anglers and spearfishers (Diogo and Pereira 2013; Diogo and Pereira 2014). according to azorean fishermen TeK, the almaco jack is a highly migratory predator that visits the region during the summer but emigrates there-after. Hence, most fishermen consider that local marine reserves will have limited or no impact on this species.
In this study, empirically assess the movement patterns of the almaco jack in two offshore shallow seamounts by a long-term monitoring of tagged fish, in order (1) to quan-tify their level of residency, while comparing telemetry data with those based on visual census and commercial catches, and (2) to estimate the potential of seamount marine reserves for protecting this species.
Methods
landings
We extracted monthly landings of almaco jack from the azorean regional fish auction authority database which, by law, controls all commercial landings in the region. We aggregated reported landings from the traditional pole-and-line with live-bait fishery from 1978 to 2011 for all nine islands, to calculate monthly total landings, which we used as a proxy for the monthly average catch by the commer-cial fleet in the azores.
Underwater visual census
We derived the seasonal pattern of occurrence of almaco jack from a fishery-independent SCUBa visual census sur-vey (UVC) from the 15-year UVC database at the Univer-sity of the azores. We only used the fish counts from July 1997 to november 2000 because after this period, the sur-veys were restricted to summer and fall. Fish were counted at rocky reefs by standard belt transect from surface to 40 m depth, and a list of all species sighted at each dive was kept (for details see Schmiing et al. 2013). We selected sampling sites, for their habitat and geographical representativeness, which included rocky reefs at the Formigas and D. João de Castro (DJC) shallow seamount reserve and at the islands of Corvo, Faial, Pico, and Sta. Maria (Fig. 1). no fish counts were available for Princesa alice seamount. The surveys covered all months of the year, but some months could not be sampled every year, while the surveys at seamounts were restricted to June, august, and September. The number of census per month was variable. To account for this unbal-anced design, (1) we averaged the monthly frequency of
1351Mar Biol (2014) 161:1349–1360
1 3
occurrence of almaco jack (from species lists per dive) from 1997 to 2000, and (2) we standardized the observations by dividing the number of fish lists containing almaco jack by the total number of dives per month.
acoustic telemetry
Study sites
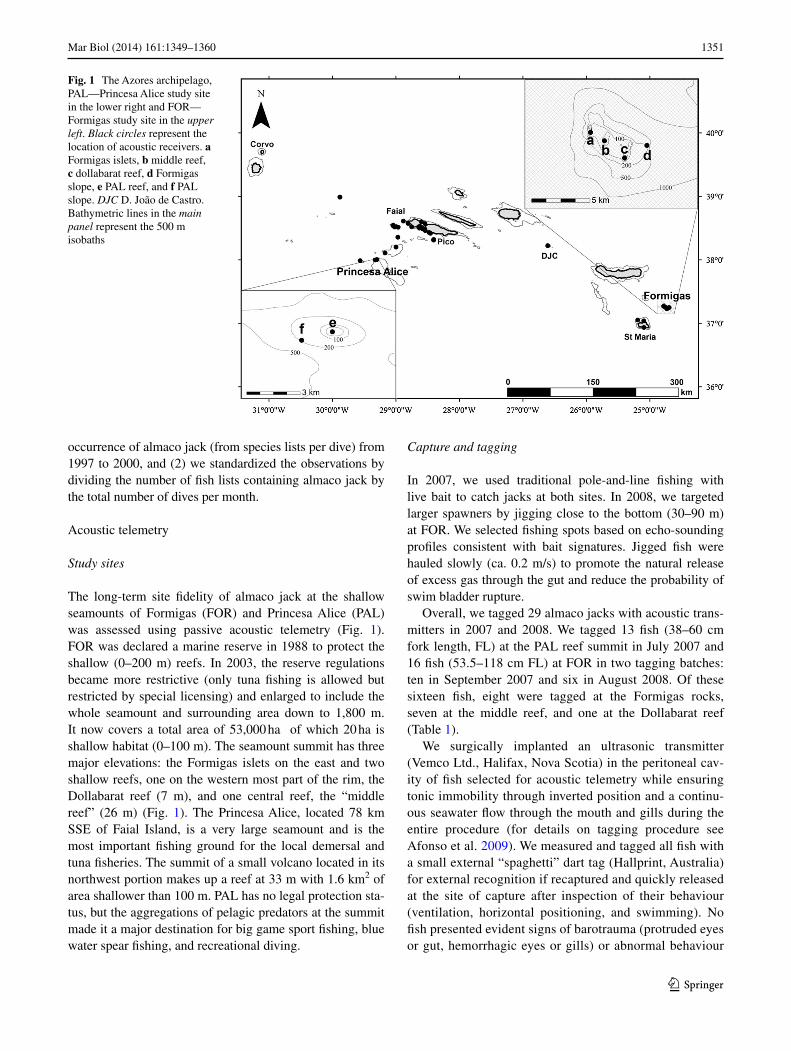
The long-term site fidelity of almaco jack at the shallow seamounts of Formigas (FOr) and Princesa alice (Pal) was assessed using passive acoustic telemetry (Fig. 1). FOr was declared a marine reserve in 1988 to protect the shallow (0–200 m) reefs. In 2003, the reserve regulations became more restrictive (only tuna fishing is allowed but restricted by special licensing) and enlarged to include the whole seamount and surrounding area down to 1,800 m. It now covers a total area of 53,000 ha of which 20 ha is shallow habitat (0–100 m). The seamount summit has three major elevations: the Formigas islets on the east and two shallow reefs, one on the western most part of the rim, the Dollabarat reef (7 m), and one central reef, the “middle reef” (26 m) (Fig. 1). The Princesa alice, located 78 km SSe of Faial Island, is a very large seamount and is the most important fishing ground for the local demersal and tuna fisheries. The summit of a small volcano located in its northwest portion makes up a reef at 33 m with 1.6 km2 of area shallower than 100 m. Pal has no legal protection sta-tus, but the aggregations of pelagic predators at the summit made it a major destination for big game sport fishing, blue water spear fishing, and recreational diving.
Capture and tagging
In 2007, we used traditional pole-and-line fishing with live bait to catch jacks at both sites. In 2008, we targeted larger spawners by jigging close to the bottom (30–90 m) at FOr. We selected fishing spots based on echo-sounding profiles consistent with bait signatures. Jigged fish were hauled slowly (ca. 0.2 m/s) to promote the natural release of excess gas through the gut and reduce the probability of swim bladder rupture.
Overall, we tagged 29 almaco jacks with acoustic trans-mitters in 2007 and 2008. We tagged 13 fish (38–60 cm fork length, Fl) at the Pal reef summit in July 2007 and 16 fish (53.5–118 cm Fl) at FOr in two tagging batches: ten in September 2007 and six in august 2008. Of these sixteen fish, eight were tagged at the Formigas rocks, seven at the middle reef, and one at the Dollabarat reef (Table 1).
We surgically implanted an ultrasonic transmitter (Vemco ltd., Halifax, nova Scotia) in the peritoneal cav-ity of fish selected for acoustic telemetry while ensuring tonic immobility through inverted position and a continu-ous seawater flow through the mouth and gills during the entire procedure (for details on tagging procedure see afonso et al. 2009). We measured and tagged all fish with a small external “spaghetti” dart tag (Hallprint, australia) for external recognition if recaptured and quickly released at the site of capture after inspection of their behaviour (ventilation, horizontal positioning, and swimming). no fish presented evident signs of barotrauma (protruded eyes or gut, hemorrhagic eyes or gills) or abnormal behaviour
Fig. 1 The azores archipelago, Pal—Princesa alice study site in the lower right and FOr—Formigas study site in the upper left. Black circles represent the location of acoustic receivers. a Formigas islets, b middle reef, c dollabarat reef, d Formigas slope, e Pal reef, and f Pal slope. DJC D. João de Castro. Bathymetric lines in the main panel represent the 500 m isobaths
1352 Mar Biol (2014) 161:1349–1360
1 3
(Jarvis and lowe 2008). The whole handling procedure lasted <4 min.
acoustic transmitters randomly emitted a coded signal every 60 to 180 s. Transmitters did not have a predefined switch-off date, but the manufacturer provides an expected battery life. FOr jacks were implanted with V16-4H r64 K (158-dB) coded transmitters of 1,470 days expected battery life. Pal jacks were implanted with V8-2l (330 and 580 days, 148-dB) and V13-1l (1,025 days, 147-dB) transmitters (Table 2).
Acoustic monitoring
In august 2007, we deployed three acoustic receivers (Vr2W, Vemco single frequency) at FOr, one at each of the three shallow reefs (about 25 m depth). We kept the receivers active for 1,410 days, i.e. until July 2011, 2 months before the estimated expiration date of the first batch transmitters, except the middle reef station which failed after 1,039 days. One additional receiver was deployed on the seamount slope (around 200 m depth) in august 2008 and definitely retrieved in September 2010. In July 2007, we deployed one receiver at Pal, at 36 m. This receiver was lost, along with the 324 days of post-release data, and was replaced in July 2008. a second receiver was deployed at deeper stands (around 200 m) in September
2008, 4 km away from the seamount summit. The shallow water receiver remained active for 832 days, and the deep receiver for 820 days. The receivers continuously moni-tored the presence of any coded transmitters in their vicin-ity, logging the exact time/date, and code of a given tag when in range. We moored the receivers about 2.5 m above the seafloor and retrieved them by SCUBa diving (shallow stations) or acoustic releases (deep stations, ar50 Sub Sea Sonics, San Diego, USa). We retrieved receivers once a year to download stored information. although we did not conduct listening range tests at the study sites, we assumed similar listening ranges as observed on other azorean coastal reefs (afonso et al. 2008a, b, afonso unpublished data) and seamounts (for details see afonso et al. 2012) of comparable characteristics. These tests showed maxi-mum listening ranges for the different transmitter models to be around 200 m (V8-2l), 500 m (V13-1l), and 800 m (V16-4H).
Data analysis
We screened the long-term monitoring data for spurious detections, which may occur whenever signals from differ-ent coded transmitters are emitted simultaneously and col-lide, resulting in “false” detections (Heupel et al. 2006). To evaluate levels of residency (site attachment) of tagged fish,
Table 1 Seriola rivoliana: summary data for 16 almaco jacks tagged with acoustic transmitters at the Formigas seamount reserve
Ir, residency index (ratio of days detected-monitoring period); IrD, residency index-detections (ratio of days detected-days between first and last detection); IForm, residency index at Formigas reef receiver; IMiddle, residency index at midlle reef receiver; IDoll, residency index at Dollabarat reef receiver; IDeep, residency index at deep station. Fl, fork length in cm; T. model, transmitter model; last det., last detection recorded in any active station in the Formigas reserve
Tag ID Fl (cm) T. model Tagging site Tagging date last det. Ir IrD I Form I Meio IDoll I Deep
2007
6666 64 V16 Dollabarat 5-Sep-07 15-Jul-11 0.89 0.89 0.23 0.04 0.66 0.03
6669 53.8 V16 Middle 7-Sep-07 12-Jul-11 0.71 0.71 0.37 0.08 0.34 0.03
6670 53.5 V16 Middle 7-Sep-07 21-Jul-09 0.38 0.79 0.16 0.08 0.22 0.03
6671 56.5 V16 Middle 7-Sep-07 11-Jul-11 0.70 0.70 0.36 0.15 0.34 0.02
6673 60.5 V16 Middle 7-Sep-07 15-Jul-11 0.78 0.78 0.35 0.10 0.43 0.02
6674 59.5 V16 Middle 7-Sep-07 15-Jul-11 0.76 0.76 0.54 0.21 0.22 0.01
6675 55.5 V16 Middle 7-Sep-07 15-Jul-11 0.79 0.79 0.34 0.04 0.45 0.04
6681 58 V16 Formigas 8-Sep-07 14-Jul-11 0.75 0.76 0.54 0.02 0.21 0.00
6682 59 V16 Formigas 8-Sep-07 15-Jul-11 0.61 0.62 0.45 0.02 0.16 0.02
6683 58 V16 Formigas 8-Sep-07 15-Jul-11 0.93 0.93 0.62 0.00 0.30 0.00
2008
6688 92 V16 Middle 28-aug-08 15-Jul-11 0.47 0.46 0.33 0.15 0.14 0.00
6690 69.5 V16 Formigas 28-aug-08 15-Jul-11 0.88 0.88 0.56 0.04 0.33 0.03
6691 95 V16 Formigas 28-aug-08 11-apr-10 0.52 0.92 0.23 0.01 0.28 0.00
6694 106 V16 Formigas 29-aug-08 15-Jul-11 0.64 0.64 0.38 0.07 0.26 0.01
6698 118 V16 Formigas 30-aug-08 15-Jul-11 0.73 0.73 0.47 0.01 0.26 0.00
6699 56 V16 Formigas 30-aug-08 14-Jul-11 0.82 0.82 0.43 0.03 0.39 0.02
1353Mar Biol (2014) 161:1349–1360
1 3
we calculated detection frequency indices after afonso et al. (2012) where the total number of days a fish was detected by receiver x (Ix) or the whole array of receivers (In) is divided by the maximum possible number of days that the fish could be detected. This denominator assumes the same value as the duration of the experiment or the transmitter′s predicted battery duration if it is shorter than the duration of the experiment. We computed an additional residence index (IrD), based on the ratio between the total number of days a fish was detected and the number of days elapsed between first and last detection (afonso et al. 2008a).
In order to preserve the length of the 4-year time series when calculating (In) at FOr, we only considered detec-tions from the receivers deployed at the three shallow reefs because only these were continuously active for 4 years. We corrected Ix by using only effective duty periods for each receiver. Ir indices were not calculated for fish #77, 80, 81, and 82 because the transmitter′s expected battery life was shorter than 1 year, and all data from that period from Pal were lost with the receiver. according to the manufacturer, transmitter behaviour may become unreli-able after the expiration date and could bias the detection patterns.
We used linear distance between stations where a given fish was detected as a proxy for fish mobility within the FOr receiver array, and minimum monthly traveled distance within the array was estimated as the sum of the linear distances between the stations sequentially visited over 1 month. For this analysis, we only used detections from September 2008 to September 2010, when all four stations were active, in order to have the best possible spatial cover of the seamount.
To further investigate residency and the seasonal patterns of detection, we modelled the probability of detection at the For-migas reserve as a function of explanatory covariates with a generalized linear mixed model, glMM (package lme4 Bates et al. 2011). We binned detections over 15-day periods, and a fish was considered present within a given bin if, at least, two valid detections were recorded within <24 from each other (for more details see afonso et al. 2012). In this analysis, the data set from the first batch of fish (i.e. those released in 2007) was truncated, taking the total period of monitoring of the second batch (1,051 days) as cut-off point. We applied a thorough ini-tial data exploration (i.e. visual inspection of data distribution and possible outliers, pairwise correlation of environmental covariates, variance inflation factor (VIF) analysis, and inter-action between variables) following Zuur et al. (2010).
available explanatory covariates included fish length, the time post-release (i.e. the number of days at liberty after being tagged), water temperature, season, surface chlorophyll-a concentration, and geostrophic current veloc-ity. The last two covariates were extracted from aVISO (http://www.aviso.oceanobs.com/en/home.html) using arc-gIS, with the Marine geospatial ecology Tools (MgeT), from a 20-by-20 km square centered at the Formigas reserve. Temperature was measured every 30 min with a temperature logger (Stowaway® tidbit) moored at 25 m on the Formigas rocks. all covariates, except season that was a categorical covariate, were averaged over 15 days to match the detection bins used. We excluded correlated covari-ates from the “full model” which thus included only time post-release, fish length, season, and geostrophic current velocity as fixed effect parameters. The tag ID was used
Table 2 Seriola rivoliana: summary data for 9 almaco jacks tagged with acoustic transmitters and monitored with acoustic receivers at the Princesa alice seamount
residency at Pal seamount starting July 2008;Ir, residency index (ratio of days detected-monitoring period);IrD, residency index-detections (ratio of days detected-days between first and last detection); Battery (days), manufacturer expected battery duration in days; recaptured, reported recapture; trans. expired, manufacturer transmitter expiration date before July 2008
Tag ID Fl (cm) T. model T. life (days) Tagging date last det. Ir IrD
77 54.5 V8 330 07/jul/08 21/out/08 Recaptured 0.39
80 60 V8 330 07/jul/08 15/out/10 Trans. expired 0.95
81 60 V8 330 07/jul/08 Trans. expired
82 58.5 V8 330 07/jul/08 25/mai/10 Trans. expired 0.86
84 39.5 V8 580 07/jul/08
85 59 V8 580 07/jul/08
86 57 V8 580 07/jul/08 14/ago/08 Recaptured 1.00
90 59.5 V13 1,025 07/jul/08 03/mar/09 0.29 1.00
91 38 V13 1,025 07/jul/08
92 50.5 V13 1,025 07/jul/08 28/mai/09 0.39 1.00
93 53 V13 1,025 07/jul/08 25/jul/10 0.87 0.96
94 58.5 V13 1,025 07/jul/08 15/jun/09 0.41 1.00
95 54 V13 1,025 07/jul/08 15/out/10 0.95 0.96
1354 Mar Biol (2014) 161:1349–1360
1 3
as random effect to account for repetitive measurements of each tag with time. We used stepwise backward model selection based on the minimization of the akaike Informa-tion Criterion (akaike 1973) to identify the best model.
We applied post hoc tests (repetitive glMMs that always used another season as baseline for the estimation) to analyse pairwise differences between seasons. Bonferroni correction was used to correct the significance levels for repetitive test-ing. Statistical models and data exploration were done with the software r (r-Development-Core-Team 2012).
Results
Commercial catches and UVC
The analysis of almaco jack landings in the azores, from 1978 to 2011, showed a pronounced seasonal pattern, with most of the landings occurring between June and Septem-ber (88 %) with a peak in July and august (Fig. 2).We found a similar seasonal pattern for the monthly occurrence of almaco jack in the species lists extracted from the UVC data set, with most relative counts occurring between June and September with a peak in august (Fig. 3).
Residency at seamounts
none of the tagged fish were detected by any acoustic receiver other than the ones at the tagging sites, i.e. FOr and Pal, respectively. at FOr, all tagged fish were almost continu-ously detected over the entire study period, except fish 6670 and 6691, which were detected only during the first 2 years (Fig. 4). The average overall IR index was 0.71, ranging from 0.38 to 0.93, while the IRD index ranged from 0.46 to 0.93 (Table 1). Despite the relatively high residency indices, we observed a coincident period of absence for most fish, roughly centered on January 2008. Fish 6688 went regularly unde-tected for about 2 months in autumn–winter periods (Fig. 4).
all data recorded by the shallow receiver during the first year of monitoring at Pal were lost, due to mooring failure. Of the 13 almaco jacks, we tagged at Pal, four were never detected, while the remaining nine individuals were immediately detected when we replaced the receiver in the shallow station. Two fish (# 77 and 86) were recap-tured almost 2 years after release. all individuals were con-tinuously recorded on a daily basis, until they ceased to be detected altogether or at the end of the study. Some trans-mitters largely exceeded the manufacturer expiration date (Fig. 5). average residency indices were 0.58 and 0.90 for Ir and IrD, respectively (Table 2).
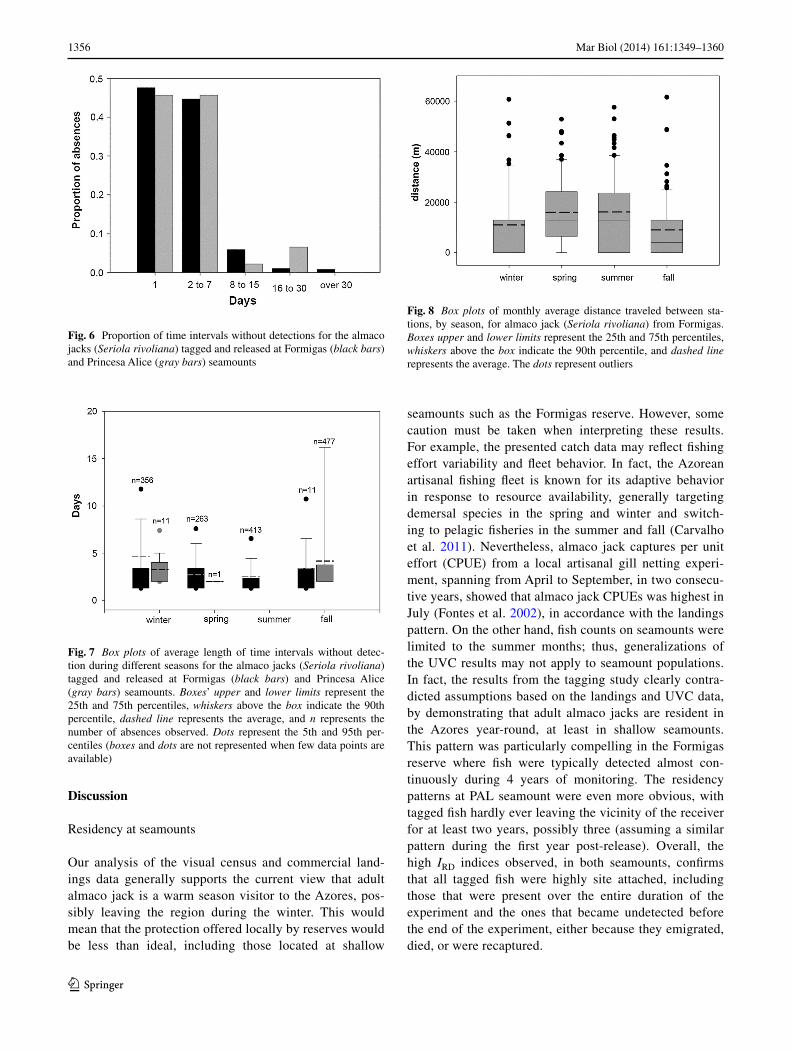
More than 90 % of all absence periods were <1 week long (92 % at FOr and 91 % at Pal), and from these, about half were 1 day long absences (Fig. 6). at Pal,
absences longer than 1 week were recorded only for fish 77, with two isolated detections post transmitter expiration date, prior to recapture, which we attributed to post expira-tion transmitter failure and treated as outliers in all statisti-cal analysis.
In both seamounts, the average absence period was longer in fall and winter than in spring and summer (nested anOVa, with season nested within year, F3,4 = 4.58; p < 0.0001). We observed no absences in the summer at Pal (Fig. 7) assum-ing the two isolated detections are outliers (see above). Con-versely, we observed an increase in mobility within the Formi-gas receiver array during spring and summer with an average distance traveled between stations significantly higher than in the fall and winter periods (nested anOVa, with season nested within year, F3,4 = 2.33; p = 0.03) (Fig. 8).
Fig. 2 Monthly cumulated catches of almaco jack (Seriola rivoliana) landed in the azores from 1978 to 2011
Fig. 3 Monthly frequency of occurrence of almaco jack (Seriola rivoliana) in underwater visual census dives conducted in the archi-pelago from 1997 to 2000. Numbers above columns represent the number of transects performed in island locations(bold) and offshore seamounts (regular)

1355Mar Biol (2014) 161:1349–1360
1 3
Detection probability
Season, geostrophic current speed, and time post-release had a significant effect on the detection probability (Table 3). Fewer detections were recorded during fall and winter in comparison with spring (Table 4), and detection
probability was inversely related to current speed and time post-release. However, as soon as one of the fishes that went unheard before the end of the study was removed from the analysis (#6670 and #6691), time post-release no longer had a significant effect on detection probability.
Fig. 4 Detection plot of almaco jack (Seriola rivoliana) tagged at the Formigas seamount reserve, illustrating periods of presence and absence on any given day during the duration of the study. Black squares indicate daily detections in all three receivers combined, and the doted vertical line indicates the day when the middle reef receiver was deactivated
Fig. 5 Detection plot of almaco jack (Seriola rivoliana) tagged at the Princesa alice (Pal) seamount, illustrating periods of presence and absence on any given day during the duration of the study. Black squares indicate daily detections in all three receivers combined. all data to the left of the dotted line (1st year post-release) were lost along with the acoustic receiver
1356 Mar Biol (2014) 161:1349–1360
1 3
Discussion
residency at seamounts
Our analysis of the visual census and commercial land-ings data generally supports the current view that adult almaco jack is a warm season visitor to the azores, pos-sibly leaving the region during the winter. This would mean that the protection offered locally by reserves would be less than ideal, including those located at shallow
seamounts such as the Formigas reserve. However, some caution must be taken when interpreting these results. For example, the presented catch data may reflect fishing effort variability and fleet behavior. In fact, the azorean artisanal fishing fleet is known for its adaptive behavior in response to resource availability, generally targeting demersal species in the spring and winter and switch-ing to pelagic fisheries in the summer and fall (Carvalho et al. 2011). nevertheless, almaco jack captures per unit effort (CPUe) from a local artisanal gill netting experi-ment, spanning from april to September, in two consecu-tive years, showed that almaco jack CPUes was highest in July (Fontes et al. 2002), in accordance with the landings pattern. On the other hand, fish counts on seamounts were limited to the summer months; thus, generalizations of the UVC results may not apply to seamount populations. In fact, the results from the tagging study clearly contra-dicted assumptions based on the landings and UVC data, by demonstrating that adult almaco jacks are resident in the azores year-round, at least in shallow seamounts. This pattern was particularly compelling in the Formigas reserve where fish were typically detected almost con-tinuously during 4 years of monitoring. The residency patterns at Pal seamount were even more obvious, with tagged fish hardly ever leaving the vicinity of the receiver for at least two years, possibly three (assuming a similar pattern during the first year post-release). Overall, the high IrD indices observed, in both seamounts, confirms that all tagged fish were highly site attached, including those that were present over the entire duration of the experiment and the ones that became undetected before the end of the experiment, either because they emigrated, died, or were recaptured.
Fig. 6 Proportion of time intervals without detections for the almaco jacks (Seriola rivoliana) tagged and released at Formigas (black bars) and Princesa alice (gray bars) seamounts
Fig. 7 Box plots of average length of time intervals without detec-tion during different seasons for the almaco jacks (Seriola rivoliana) tagged and released at Formigas (black bars) and Princesa alice (gray bars) seamounts. Boxes’ upper and lower limits represent the 25th and 75th percentiles, whiskers above the box indicate the 90th percentile, dashed line represents the average, and n represents the number of absences observed. Dots represent the 5th and 95th per-centiles (boxes and dots are not represented when few data points are available)
Fig. 8 Box plots of monthly average distance traveled between sta-tions, by season, for almaco jack (Seriola rivoliana) from Formigas. Boxes upper and lower limits represent the 25th and 75th percentiles, whiskers above the box indicate the 90th percentile, and dashed line represents the average. The dots represent outliers

1357Mar Biol (2014) 161:1349–1360
1 3
Despite the strong site-attachment behaviour, two fish (13 %) from FOr and four fish (31 %) from Pal stopped being detected before the end of the study or were never detected altogether. although we cannot exclude the pos-sibility that these fish emigrated from the seamounts or remained out of the detection range, we believe that they were caught by fishermen. Tuna fishing is very frequent at both seamounts, and two tagged fish were reported recap-tured by tuna boats at the Pal seamount. These recaptures were reported by skippers based close to our research insti-tute in Faial Island, and we suspect that some fish may have been recaptured by boats which do not typically land in Faial. In addition, poaching at FOr is frequently reported by dive operators and supported by our own observations over the past 15 years. It is unlikely that fish potentially caught by poachers would be reported. Moreover, none of the tagged almaco jacks were detected by the receiver array located at the major offshore reefs off Sta Maria and Faial islands or any other receiver within the regional network (Fig. 1).
Despite the long-term residency patterns, we recorded absences of 1–2 months, mainly during the first winter at Formigas seamount. noticeably, nearly all fish tagged in 2007 went undetected at around the same time. Such a pat-tern could have resulted from the deterioration of acous-tic conditions during inclement winter conditions and increased environmental noise, or a behavioural response, such as vertical seasonal migrations, or both. We do not
expect that these potential behaviour changes were moti-vated by biological cycles since the pattern was not cycli-cal and was not observed in the following winters. Tagged fish could have undertaken longer off seamount migrations and returned. However, none of these fish were detected in our receiver array around Sta. Maria Island, about 40 km away, and represents the closest available reef habitat, or in the any other receivers across the archipelago. even though inter-island movements have not typically been reported for other carangid or pelagic reef teleosts (Holland et al. 1996; Meyer and Holland 2005; Meyer et al. 2007; afonso et al. 2009), this possibility cannot be excluded.
Short periods without detections were frequent at FOr but rare at Pal seamount. This difference is most likely a result of the different receiver′s detection range and avail-ability of potential habitat at both sites. One receiver at the summit of Pal potentially covered most of the reef area shallower than 100 m, while the three shallow receivers at FOr covered <20 % of this habitat. In addition, the Formi-gas seamount summit shows greater morphologically com-plexity and concomitant physical barriers to the propaga-tion of the acoustic signals from transmitters. For example, fish on the east side of the Formigas islets would not be detected even if they were within detection range because the Formigas rocks would act as a barrier between the sta-tion, on the west side, and the fish. afonso et al. (2009) found that white trevally Pseudocaranx dentex from off-shore reefs occupied the down-current areas in relation to the reef summit when currents were strong and moved to the shallowest part of the reef when current was slowest. a similar behaviour might also occur in the almaco jack, and it would explain some temporal gaps of detection at FOr and the inverse relationship between geostrophic current strength and detection probability.
Our results also suggest that the almaco jacks increased their mobility within the Formigas seamount during spring and summer. This period includes the spawning season of almaco jack in the azores, which takes place between June and September (Fontes personal observation), and our tagged fish were all at about or larger than size at first matu-rity (afonso, unpublished data). Such increased mobility might be related to spawning activity, as reported for other carangidae. This behaviour has also been documented for white trevally where mature fish were less attached to their core areas and moved more widely within the receiver range of detection during the spawning season (afonso et al. 2009). In a similar study, Meyer et al. (2007) also found that giant trevally increased the frequency of visits between the core activity areas and spawning aggregations sites during spring and summer. It is possible that increased mobility is related to travels between feeding and spawn-ing sites within the seamount, or that it allows the fish to increase mating opportunities.
Table 3 Summary of the generalized linear mixed model
Spring is used as baseline for the estimation of the categorical vari-able season and, as such, does not appear in the model summary
estimate Se z value Pr(>|z|)
Intercept 7.2121363 0.8628939 8.358 <2e−16
Time Post rel. −0.0016886 0.0004425 −3.816 0.000136
Season (summer) −0.5916930 0.4563624 −1.297 0.194789
Season (winter) −1.3192956 0.4151139 −3.178 0.001482
Season (fall) −1.3222545 0.4303411 −3.073 0.00212
Geo_Curr −0.2151736 0.0568076 −3.788 0.000152
Table 4 Significant differences in tag detection between different seasons
a Bonferroni correction for multiple testing was applied. The 5 % significance level (p < 0.5) is corrected to p = 0.00833 (p = 0.01 is corrected to p = 0.00167). Significant values are printed bold
Spring Summer Fall winter
Spring
Summer 0.194789
Fall 0.002122 0.051876
Winter 0.001482 0.043077 0.992723

1358 Mar Biol (2014) 161:1349–1360
1 3
If almaco jacks are in fact more vagile in the summer, it could explain why they are more frequently recorded by divers in the summer surveys as the probability of encoun-ters is expected to increase as the fish should cover larger areas within the habitat boundaries. Similarly, this increased mobility could also affect their vulnerability to fishing gears.
It is also possible that the almaco jack moves during fall and winter towards deeper water where UVC surveys usually are not carried out. In fact, over 80 % of all valid detections at the deep station, from august 2008 to Sep-tember 2010, were recorded in the fall, partially supporting this hypothesis, even though no detections were recorded during the winter. If this is the case, it could also explain the marked seasonal pattern observed in landings since the pole-and-line fishing is limited to the first few meters of the water column where fishermen can see the jacks attracted to the live bait. In addition, the preferred bait fish (juve-nile, Scomber japonicas, Trachurus picturatus, and Pagel-lus sp.) is scarce during the colder months (Fontes personal observation), when captures are lowest. The hypothetical vertical migration towards deeper habitat in the fall and winter could also explain why fewer detections and longer absences were observed during this period (supplemen-tary material). Some caution must be taken when general-izing these results to other areas of the archipelago and to immature fish. For example, larger mature white trevally use offshore reefs as preferential spawning habitat, while immature fish prefer inshore habitats all year-round, as probably do a number of other pelagic predators (afonso et al. 2008b). Future studies are needed to clarify the long-term spatial behaviour of immature almaco jacks as well as that of coastal populations of adults.
Implications for management
Our findings have clear management implications given the growing body of evidence stressing the vulnerability and risks of exploiting fish aggregations, particularly spawning aggregations (Sadovy and Domeier 2005b; Sadovy et al. 2008). Moreover, in recent years, fisheries scientists and ecologists have also recognized that seamount-aggregating fishes have higher intrinsic vulnerability than other groups of fishes (Morato et al. 2006; Morato and Pitcher 2008). even though very little is known about the spatial ecology of sea-mount fishes (but see afonso et al. 2012), there has been a growing effort to protect and manage a number of both shal-low and deep water seamounts in the north atlantic, par-ticularly in the azores region (Cardigos 2006; Morato et al. 2010). Our results show that marine reserve at the Formigas shallow seamount can play an important role in protecting and managing the almaco jack. It is likely that other reserves proposed for some north atlantic seamounts where aggre-gations of jacks have been documented (e.g. the gorringe
seamount, abecassis et al. 2009) can effectively protect resident populations of mature almaco jack and, most impor-tantly, preserve significant spawning biomass given that the largest individuals are typically found on the seamounts.
This study suggests that resident almaco jacks are likely more vulnerable at Pal than at FOr because the fish location is highly predictable and essentially restricted to the seamount summit. Thus, even a small protected area around the summit of Pal could effectively protect a core subpopulation of this species in the long term. From an economic perspective, aggregations of large predators might be more valuable as an attraction for Scuba divers rather than as a resource for local fisheries (Sadovy et al. 2008). ecotourism in the azores, particularly the diving at shallow seamount, targeting sharks, mantas, and other large predators, has increased exponentially over the past cou-ple of years, while income and yields from fisheries show signs of decline. Yet, some of the most popular seamounts for dive tourism, such as the Pal, still have no protection status, while enforcement is deficient at DJC and Formigas reserves. More importantly, this study demonstrates the significance of small reserves to protect putative spawn-ing aggregation sites for the management and conservation of reef fishes that are traditionally seen as highly mobile, such as the almaco jack. Protecting these sites might have a beneficial effect on local populations because of the dispro-portionate contribution of the large size individuals to the reproductive potential of the population.
Acknowledgments We thank T. Morato, F. Cardigos, n. Serpa, M. Santos, J. Sousa, g. graça, M. Machete, J. Botelho, and the crews of r/V “arquipélago” and r/V “Águas Vivas” for their help with field work; F. Tempera, r. Medeiros, and g. graça for providing data and support in the production of bathymetric maps; and J. San-tos for telemetry database management. a special thank is due to r. S. Santos for stimulating discussions and for his continued support to this research. FCT/MCTeS-MeC provided individual support to J.F., M.S., and P.a. (SFrH/BD/12788/2003, SFrH/BPD/66532/2009, SFrH/BD/66117/2009, SFrH/BPD/27057/2006, and Ciência 2008/POPH/Qren/FeDer/COMPeTe, FrCT, M3.1.5/F/168/2012). IMar-DOP/Uaz is UI&D #531 and associated laboratory larSyS funded by FCT/MeC and azorean DrCTC (Oe, FeDer/COM-PeTe, Pro-Convergência). This paper is a contribution to the projects MarMaCII (InTerreg/05/MaC/4.2/a4), SeaMOV (PTDC/Mar/108232/2008), and MoniZeC (DrCT/0281M2.1.2/I/018/2011), and received additional funding through the FCT/MCTeS and DrCT “reequipamento” programs. research in this study was conducted in agreement with azorean and Portuguese laws and legal requirements for the handling of vertebrate animals.
References
abecassis D, Cardigos F, alamada F, gonçalves JMS (2009) new records on the ichthyofauna of the gorringe seamount (north-eastern atlantic). Mar Biol res 5(605):611
afonso P (2002) Spatial patterns in the littoral fish community of the azores. MS thesis, Coimbra

1359Mar Biol (2014) 161:1349–1360
1 3
afonso P, Fontes J, Holland Kn, Santos rS (2008a) Social status determines behaviour and habitat usage in a temperate parrot-fish: implications for marine reserve design. Mar ecol Prog Ser 359:215–227
afonso P, Fontes J, Morato T, Holland Kn, Santos rS (2008b) repro-duction and spawning habitat of white trevally, Pseudocaranx den-tex, in the azores, central north atlantic. Sci Mar (Barc) 72:373–381
afonso P, Fontes J, Holland Kn, Santos rS (2009) Multi-scale pat-terns of habitat use in a highly mobile reef fish, the white trevally Pseudocaranx dentex, and their implications for marine reserve design. Mar ecol Prog Ser 381:273–286. doi:10.3354/meps07946
afonso P, graça g, Berke g, Fontes J (2012) First observations on seamount habitat use of blackspot seabream (Pagellus bogara-veo) using acoustic telemetry. J exp Mar Biol ecol 1:436–437
akaike H (1973) Information theory and an extension of the maxi-mum likelihood principle. In: Petrov Bn, Csaki F (eds) 2nd Inter-national symposium information theory, Budapest, pp 267–281
Barreiros JP, Morato T, Santos rS, de Borba ae (2003) Interannual changes in the diet of the almaco jack, Seriola rivoliana (Perci-formes: Carangidae) from the azores. Cybium 27:37–40
Bates D, Maechler M, Bolker B (2011) lme4: linear mixed-effects models using S4 classes. r package version 0.999375-42. http://Cran.r-project.org/package=lme4
Bolden SK (2000) long-distance movement of a nassau grouper (Epinephelus striatus) to a spawning aggregation in the central Bahamas. Fish Bull 98:642–645
Brown H, Benfield MC, Keenan SF, Powers SP (2010) Movement patterns and home ranges of a pelagic carangid fish, Caranx crysos, around a petroleum platform complex. Mar ecol Prog Ser 403:205–218. doi:10.3354/meps08465
Capello M, Soria M, Cotel P, Potin g, Dagorn l, Freon P (2012) The heterogeneous spatial and temporal patterns of behavior of small pelagic fish in an array of Fish aggregating Devices (FaDs). J exp Mar Biol ecol 430:56–62. doi:10.1016/j.jembe.2012.06.022
Cardigos F (2006) Contribuição para a Implementação de Um plano de gestão no Sítio de Interesse Comunitário Banco D. João de Castro. In: Dias e, Cancela Ml, Fonsec l, Beja P, Dentinho T (eds) gestão de recursos marinhos. Principia, Cascais, p 385
Carvalho n, edwards-Jones g, Isidro e (2011) Defining scale in fish-eries: small versus large-scale fishing operations in the azores. Fish res 109:360–369. doi:10.1016/j.fishres.2011.03.006
De Mitcheson YS, Cornish a, Domeier M, Colin Pl, rus-sell M, lindeman KC (2008) a global baseline for spawn-ing aggregations of reef fishes. Conserv Biol 22:1233–1244. doi:10.1111/j.1523-1739.2008.01020.x
Dempster T (2005) Temporal variability of pelagic fish assemblages around fish aggregation devices: biological and physical influences. J Fish Biol 66:1237–1260. doi:10.1111/j.1095-8649.2005.00674.x
Dempster T, Kingsford MJ (2003) Homing of pelagic fish to fish aggregation devices (FaDs): the role of sensory cues. Mar ecol Prog Ser 258:213–222. doi:10.3354/meps258213
Diogo HMC, Pereira Jg (2013) Impact evaluation of spear fish-ing on fish communities in an urban area of São Miguel Island (azores archipelago). Fish Manage ecol 20:1–11. doi:10.1111/fme.12036
Diogo HMC, Pereira Jg (2014) assessing the potential biological implications of recreational inshore fisheries on subtidal fish communities of azores (northeast atlantic) using catch and effort data. J Fish Biol 8:952–970
Fontes J, Machete M, Santos rS (2002) Projecto BarCa—Bases para a Caracterização de duas Pescarias artesanais Costeiras dos açores: redes de emalhar e peixe-porco: relatório Final. Univer-sidade dos açores, Horta
Friedlander aM, DeMartini ee (2002) Contrasts in density, size, and biomass of reef fishes between the northwestern and the main
Hawaiian Islands: the effects of fishing down apex predators. Mar ecol Prog Ser 230:253–264
gillanders BM, Ferrell DJ, andrew nl (2001) estimates of move-ment and life-history parameters of yellowtail kingfish (Seriola lalandi): how useful are data from a cooperative tagging pro-gramme? Mar Freshw res 52:179–192. doi:10.1071/mf99153
Halpern BS (2003) The impact of marine reserves: do reserves work and does reserve size matter? ecol appl 13:S117–S137
Heithaus Mr, Frid a, Wirsing aJ, WormMichael B (2008) Predicting ecological consequences of marine top predator declines. Trends ecol evol 23:537
Heupel Mr, Simpfendorfer Ca, Collins aB, Tyminski JP (2006) residency and movement patterns of bonnethead sharks, Sphyrna tiburo, in a large Florida estuary. environ Biol Fishes 76:47–67. doi:10.1007/s10641-006-9007-6
Holland Kn, lowe Cg, Wetherbee BM (1996) Movements and dis-persal patterns of blue trevally (Caranx melampygus) in a fisher-ies conservation zone. Fish res 25:279–292
Hutson KS, Smith BP, godfrey rT, Whittington ID, Chambers CB, ernst I, gillanders BM (2007) a tagging study on yellowtail kingfish (Seriola lalandi) and Samson fish (S-hippos) in south australian waters. Trans r Soc S aust 131:128–134
Jarvis eT, lowe Cg (2008) The effects of barotrauma on the catch-and release survival of southern California nearshore and shelf rockfish (Scorpaenidae, Sebastes spp.). Can J Fish aquat Sci 65:1286–1296. doi:10.1139/F08-071
Manooch CS, Haimovici M (1983) Foods of greater amberjack, Seri-ola dumerili, and almaco jack, Seriola rivoliana (Pisces: Caran-gidae), from the South atlantic Bight. J elisha Mitchell Sci Soc 99:1–9
McClellan DB, Cummings nJ (1996) Preliminary analysis of tag and recapture data of the greater amberjack, Seriola dumerili, in the southeastern United States. In: Proceedings of the 49th gulf and Caribbean Fisheries Institute, pp 25–45
Meyer Cg, Holland Kn (2005) Movement patterns, home range size and habitat utilization of the bluespine unicornfish, Naso uni-cornis (acanthuridae) in a Hawaiian marine reserve. environ Biol Fishes 73:201–210
Meyer Cg, Holland Kn, Wetherbee BM, lowe Cg (2001) Diet, resource partitioning and gear vulnerability of Hawaiian jacks captured in fishing tournaments. Fish res 53(2):105–113
Meyer Cg, Holland Kn, Papastamatiou YP (2007) Seasonal and diel movements of giant trevally Caranx ignobilis at remote Hawaiian atolls: implications for the design of marine protected areas. Mar ecol Prog Ser 333:13–25. doi:10.3354/meps333013
Morato T, Pitcher TJ (2008) reconciling fisheries with conservation on seamounts. reconcil Fish Conserv, Vols I and Ii 49: 1623–1634
Morato T, Cheung WWl, Pitcher TJ (2006) Vulnerability of seamount fish to fishing: fuzzy analysis of life-history attributes. J Fish Biol 68:209–221. doi:10.1111/j.1095-8649.2005.00894.x
Morato T, Pitcher TJ, Clark Mr, Menezes g, Tempera F, Porteiro F, giacomello e, Santos rS (2010) Can we protect seamounts for research? a call for conservation. Oceanography 23:190–199
r-Development-Core-Team (2012) r: a language and environment for statistical computing. r-project.org, Vienna
russ gr (2002) Yet another review of marine reserves as reef fish-ery management tools. In: Sale PF (ed) Coral reef fishes: dynam-ics and diversity in a complex ecosystem. accademic press, San Diego, pp 421–443
Sadovy Y, Domeier M (2005a) are aggregation-fisheries sustain-able? reef fish fisheries as a case study. Coral reefs 24:254–262. doi:10.1007/s00338-005-0474-6
Sadovy Y, Domeier Ml (2005b) Perplexing problems of sexual pat-terns in the fish genus Paralabrax (Serranidae, Serraninae). J Zool 267:121–133. doi:10.1017/S0952836905007466

1360 Mar Biol (2014) 161:1349–1360
1 3
Sadovy Y, De Mitcheson YS, Cornish a, Domeier M, Colin Pl, rus-sell M, lindeman KC (2008) a global baseline for spawning aggregations of reef fishes. Conserv Biol 22:1233–1244
Sala e, aburto-Oropeza O, Paredes g, Thompson g (2003) Spawning aggregations and reproductive behavior of reef fishes in the gulf of California. Bull Mar Sci 72:103–121
Schmiing M, afonso P, Tempera F, Santos r (2013) Predictive habitat modelling of reef fishes with contrasting trophic ecologies. Mar ecol Prog Ser 478:201–216
Smith-Vaniz W (1986) Carangidae. In: Whitehead PJP, Bauchot M-l, Hureau J-C, nielsen J, Tortonese e (eds) Fishes of the northeast atlantic and the Mediterranean (FnaM). UneSCO, Paris, pp 815–844
Stanley Dr, Wilson Ca (1997) Seasonal and spatial variation in the abundance and size distribution of fishes associated with a petro-leum platform in the northern gulf of Mexico. Can J Fish aquat Sci 54:1166–1176. doi:10.1139/cjfas-54-5-1166
Stanley Dr, Wilson Ca (2003) Seasonal and spatial variation in the biomass and size frequency distribution of fish associated with oil and gas platforms in the northern gulf of Mexico. Fisheries, reefs, and Offshore Development, pp 123–153
Tanaka S (1984) Migration model and dynamics parameters of large-sized yellowtails in the Pacific along the Japanese coast inferred from tag recaptures after the year of release. Bull Jpn Soc Sci Fish 50:1341–1347
Wetherbee BM, Holland Kn, Meyer Cg, lowe Cg (2004) Use of a marine reserve in Kaneohe Bay, Hawaii by the giant trevally, Caranx ignobilis. Fish res 67:253–263. doi:10.1016/j.fishres.2003. 11.004
Zuur aF, Ieno en, elphick CS (2010) a protocol for data explora-tion to avoid common statistical problems. Methods ecol evol 1:3–14. doi:10.1111/j.2041-210X.2009.00001.x