fibronectin receptors of mononuclear phagocytes: binding characteristics and biochemical isolation
TRANSCRIPT

Experimental Cell Research 181 (1989) 420-431
Fibronectin Receptors of Mononuclear Phagocytes: Binding Characteristics and Biochemical Isolation
ANGELES GARCIA-PARDO,’ ORLANDO C. FERREIRA, JAY VALINSKY, and CELSO BIANCO
The Lindsley F. Kimball Research Institute, The New York Blood Center, 310 East 67th Street, New York, New York 10021
Fibronectin receptors on mononuclear phagocytes are involved in the localization of monocytes at intlammatory sites and in the subsequent expression of macrophage-like phenotypes. In this study, we have investigated the hypothesis that proteolytically derived fragments of tibronectin may interfere with binding of tibronectin to monocytes in the extracellular matrix. We report on the reactivity of U937 cells with an 80-kDa tryptic fragment of fibronectin which contains the cell-binding domain but lacks the gelatinicolla- gen-binding domain. U937 cells attached to surfaces coated with the 80-kDa fragment as well as with intact fibronectin. Preincubation of the cells with the 80-kDa fragment inhibited attachment to both surfaces while intact tibronectin had little or no inhibitory effect. The K, for inhibition of attachment (0.5 @4) was consistent with the Kd for binding of the 3H-labeled 80-kDa fragment (0.34 p&f) to U937 cells in suspension. There were 4-5x 10’ 80-kDa binding sites per cell. The relatively high affinity of the 80-kDa fragment for the monocyte surface permitted the isolation and characterization of fibronectin- binding proteins from U937 cells and peripheral blood monocytes by affinity chromatogra- phy. When octylglucoside lysates of lactoperoxidase iodinated cells were applied to 80- kDa-Sepharose columns, a polypeptide complex of 152/125 kDa was eluted with the synthetic peptide GRGDSPC, but not with GRGESP. This complex resolved into a single diffuse band of 144 kDa upon reduction. Binding of the protein complex to the affinity column required divalent cations. The complex bound to wheat germ agglutinin and could be specifically eluted by N-acetylglucosamine. Similar cell-surface proteins were isolated from peripheral blood ITIOnOCyteS. 0 1989 Academic Press, Inc.
Many of the functions of mononuclear phagocytes are regulated by specific interactions between plasma membrane receptors and components of the extra- cellular matrix, notably libronectin (Fn) [l-3]. The attachment of monocytes to surfaces containing Fn not only serves to localize the cells in the matrix (e.g., at sites of inflammation) [4] but also might lead to expression of macrophage- associated activities (e.g., expression of membrane receptors, enhanced phago- cytosis, appearance of oxidative metabolites, and secretion of growth factors) [ 1, 3, 5, 61.
As in the case of fibroblasts [7, 81 and platelets [9], the interaction of mono- cytes with Fn is mediated by specific cell-surface receptors. Monocyte receptors recognize the RGDS (Arg-Gly-AspSer) sequence of the cell-binding domain of Fn [lO-131, are trypsin sensitive [14], and require Mg2+ for binding 1151. All these properties are common to members of the integrin family of receptors [16, 171. Although the structural composition of monocyte Fn receptors was recently
’ To whom reprint requests should be addressed at Mononuclear Cell Biology, The Lindsley F. Kimball Research Institute, New York Blood Center, 310 E 67th Street, New York, NY 10021.
Copyright @ 1989 by Academic Press, Inc. 420 All rights of reproduction in any form reserved 0014.4827/89 SOi.

Fibronectin receptors of monocytes 421
reported [18], other properties of these cell-surface molecules, as well as the binding parameters of the monocyte/Fn interaction, have not been studied.
Interactions between mononuclear phagocytes and Fn occur preferentially in solid phases (e.g., extracellular matrix, blood clots, opsonized particles) rather than in solution. This is especially relevant in light of the high concentrations of Fn normally found in plasma [ 191 and the relatively low affinity and specificity of binding of intact, soluble Fn reported for monocytes [14, 201, fibroblasts [21], and hepatocytes [22].
In contrast, some soluble proteolytic fragments of Fn bind to cell receptors with relatively high affinity and several of these fragments have pronounced biological activities [23-271. It has been postulated [28] that these fragments, if present in sufficiently high concentrations in duo, could compete for binding to cellular Fn receptors. In the case of monocytes, this competition could interfere with the ability of the cells to migrate and subsequently localize at inflammatory sites.
In order to test this hypothesis, we have examined the interaction of an 80-kDa tryptic fragment of Fn, which contains the cell-binding domain, but lacks the N- terminal fibrin- and collagen-binding domains, with the human monocytoid cell line U937. We demonstrate the binding of this fragment to suspensions of U937 cells as well as the ability of the fragment to inhibit the attachment of cells to Fn- coated surfaces. The relatively high affinity of the 80-kDa fragment for cellular Fn receptors has also allowed receptor isolation and characterization following afftnity chromatography.
MATERIALS AND METHODS Purification of the 80-klla tryptic fragment offibronectin. Human plasma tibronectin, purified by
affinity chromatography on gelatin-Sepharose [29], was the generous gift of Drs. B. Horowitz and R. Schulman (New York Blood Center, NY). An IO-kDa fragment containing the cell-binding domain, but not the collagen-binding domain, was isolated from a tryptic digest of Fn by aftinity and ion- exchange chromatography. Fn (600 mg) was digested with TPCK-trypsin (Cooper Biomedical, Malvem, PA) (1: 200, w/w) for 90 min at 37°C in a 25 mM Tris-Cl buffer, pH 7.6, containing 50 n&f NaCl and 0.5 mit4 NarEDTA (digestion buffer). Phenylmethylsulfonyl fluoride (PMSF) was added to a final concentration of 1 m&4 and the digest was applied to a heparin-Sepharose column (Pharmacia, Inc., Piscataway, NJ), at 5 mg protein/ml of swollen gel. After extensive washing with the digestion buffer, a fraction containing an IO-kDa fragment, as a predominant species, was eluted by addition of 0.1 M NaCl to the buffer and applied to a gelatin-Sepharose column (Pharmacia) at 2 mg protein/ml gel. Polypeptides containing the gelatin-binding domain were retained by the column. The flow- through fraction from the column, which included the 80-kDa fragment, was dialyzed against starting buffer (10 m&f Tris-HCl, 50 mM NaCl, pH 6.0) and applied to a DEAE-Sephacel column (Pharmacia). The column was first washed with starting buffer and the 80-kDa fragment was then eluted with starting buffer containing 0.2 M NaCl. Purity of the fragment was assessed by SDS-PAGE.
To prepare affinity columns containing the 80-kDa fragment, the purified protein was concentrated on an Amicon YM-10 membrane filter, dialyzed against 0.1 M NaHCOJ0.5 M NaCl, pH 8.5, and coupled to cyanogen bromide-activated Sepharose (Pharmacia) at 3-4 mg/ml gel, according to the manufacturer’s instructions. Identical procedures were used for the preparation of Fn-Sepharose columns.
The 80-kDa Fn fragment was radiolabeled by reductive methylation using 25 mCi of NaB[‘H]H4 (100 mCi/mmol, Du Pont Inc., Wilmington, DE) per milligram of protein and formaldehyde following published procedures [30]. The labeled fragment was extensively dialyzed against PBS at 4°C and stored at -70°C until use. Specific activities of approximately 10 Ci/mol were routinely obtained.

422 Garcia-Pardo et al.
Binding of ‘H-labeled BO-kDa to U937 cells. U937 cells were obtained from the American Type Culture Collection and expanded in RPM1 1640 containing 10% heat-inactivated fetal bovine serum (FBS; Irvine Scientific, Santa Ana, CA) in l-liter roller bottles. Cultures, initiated at 16 cells/ml, were terminated on the fourth day of incubation. The cells were collected by centrifugation, washed three times with RPM1 containing 10 m&f Hepes and 1.5 % BSA (binding medium), and resuspended in this medium at 7~10~ cells/ml. The ‘H-labeled IO-kDa fragment alone, or mixed with an excess of unlabeled fragment, was added to the cells in a final volume of 400 ul and the suspension was incubated at an ambient temperature for 30 min on a rotary shaker. At the end of the incubation, the cells were centrifuged at 200g and washed three times in binding medium. The final pellet was resuspended in 50 ul of this medium, added to Hydro-Solv No. 2 liquid scintillation fluid (Anoroc Scientific, South Hackensack, NJ), and counted. Total and specific binding, as well as equilibrium constants, were calculated by Scatchard analysis using Enztitter programs (R. J. Leatherow, Elsevier Science, Amsterdam).
Cell attachment assays. Cell attachment assays were performed on flat-bottom, 96-well Linbro tissue culture plates (Flow Laboratories, McLean, VA). The wells were coated with 100 ul of appropriate dilutions of Fn or the 80-kDa fragment for 1 h at room temperature, washed three times with PBS, and then incubated with 100 ul of BSA (10 mg/ml) in PBS for 1 h at 37°C to block nonspecific binding sites. Prior to use, the plates were rinsed three times with PBS containing 0.9 mM CaCl* and 0.5 mM MgCl*.
For attachment assays, 100 ul of a suspension containing 5x 10’ cells/ml in RPMI, 1% BSA, and 10 mM Hepes was added to each well. The plates were incubated at 37°C for 30 min in an atmosphere of 5 % CO*. Nonadherent cells were removed by three gentle washes with Hanks’ BSS containing Ca2+ and Me, and attached cells were fixed by addition of 1.25 % glutaraldehyde at room temperature. For attachment-inhibition assays, U937 cells (1.25~ lO?ml in RPMI, 10 rnM Hepes, 1% BSA) were preincubated in a total volume of 0.4 ml with appropriate amounts of the IO-kDa fragment or Fn for 30 min at room temperature. The mixture was diluted to 1 ml, and a loo-u1 (5X104 cells) aliquot was transferred to wells coated with 1.25 ug F&m*. After a 30-min incubation, the wells were washed gently and attached cells were fixed with glutaraldehyde. Cells were stained with 0.1% toluidine blue and attachment was assessed, in both types of assays, on a Nikon Diaphot inverted microscope, with the aid of an image analyzer (Artek Counter, Model 982).
In some experiments, U937 cells (10’ in 1 ml) were pretreated with 200 ug of TPCK-trypsin at 37°C for 40 min. Soybean trypsin inhibitor (500 ug) was added to both trypsin-treated and control preparations and the cells were washed twice with RPM1 containing 10 mM Hepes and 1% BSA prior to assay.
Cell-surface labeling and preparation of lysates. U937 cells and moncytes were surface iodinated exactly as described [7, 311. Labeled cells were lysed for 10 min at 4°C in 1 ml of PBS containing 200 mM octylglucoside (Sigma Chemical Co., St. Louis, MO) 1 mM CaCh, 1 mM MgCl*, 0.1% aprotinin (Sigma), and 3 mM PMSF.
Affinity chromatography of cell lysates on BO-kDa fragment-Sepharose columns. Octylglucoside lysates from 10’ cells were applied to a 2-ml 80-kDa fragment-Sepharose column equilibrated with PBS containing 1 mM CaCl*, 1 rnM MgC&, 50 mM octylglucoside, and 1 m&f PMSF (column buffer). The column was eluted first with column buffer, then sequentially with column buffer containing 0.3 M NaCl, 1 mg/ml of the synthetic peptide GRGESP (Peninsula Laboratories, Belmont, CA), 1 mg/ml of the synthetic peptide GRGDSPC (Peninsula), and 8 M urea. Affinity chromatography of cell lysates on Fn-Sepharose columns was performed exactly as above.
Affinity chromatography on gelatin-Fn columns. Fn (8 mg) was bound to 2 ml of gelatin-Sepharose on a rotary mixer at room temperature for 1 h. The amount of Fn bound was determined by absorbance at 280 nm of the protein solution before and after incubation, and was found to be over 95% of the starting material. Lysates of U937 cells were applied to Fn-gelatin-Sepharose columns and eluted exactly as described for IO-kDa fragment-Sepharose columns.
AfjTnity chromatography on wheat germ agglutinin (WGA). The radioactive protein specifically eluted by GRGDSPC from 80-kDa fragment-Sepharose columns was incubated with 0.5 ml WGA-Se- pharose (Sigma) for 2-3 h at an ambient temperature on a rotary mixer. The beads were then washed twice by centrifugation with column buffer and treated for 15 min with 0.5 M N-acetylglucosamine (Sigma) in column buffer; the eluate was analyzed by SDS-PAGE and autoradiography.
Immunoprecipitation. Aliquots (0.5 ml) of the materials specifically eluted from the IO-kDa-Sephar- ose column with GRGDSPC were added to 0.9 ml of a buffer containing 20 mM Tris-HCl, pH 7.5, 0.15 M NaCl, 1 mM CaCl,, 2 mM PMSF, 2 mg/ml BSA, and 1% NP-40 [32] and preadsorbed with 60 ul of a 20% suspension (v/v) of formalin-fixed Staphylococcus uureus (Bethesda Research Lab., Gaithersburg, MD) for 90 min at 4°C. The mixture was centrifuged at 10,OOOg for 15 min, the pellets
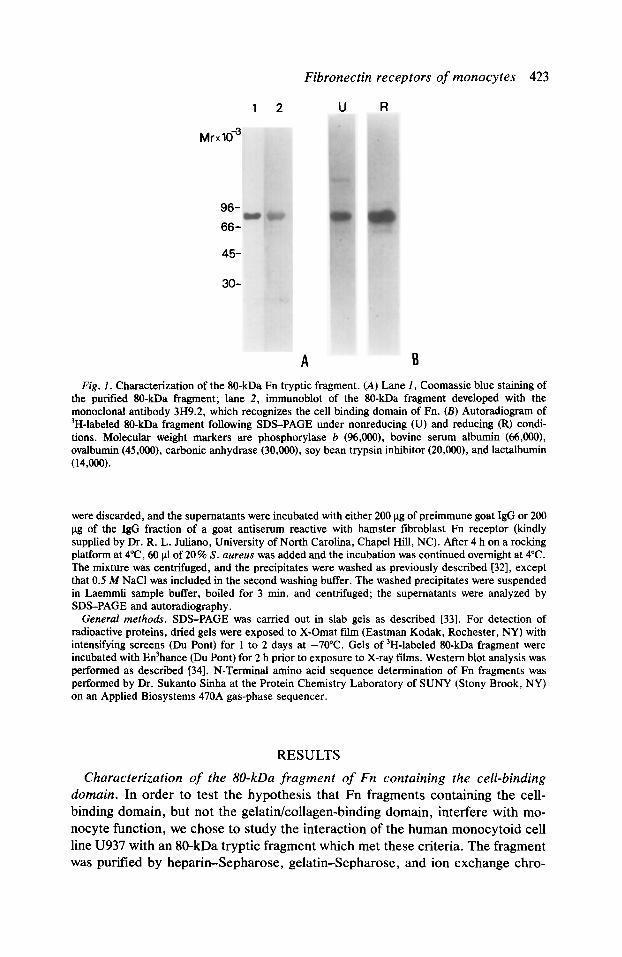
Fibronectin receptors of monocytes 423
Fig. 1. Characterization of the 80-kDa Fn tryptic fragment. (A) Lane I, Coomassie blue staining of the purified IO-kDa fragment; lane 2, immunoblot of the 80-kDa fragment developed with the monoclonal antibody 3H9.2, which recognizes the cell binding domain of Fn. (B) Autoradiogram of ‘H-labeled 80-kDa fragment following SDS-PAGE under nonreducing (U) and reducing (R) condi- tions. Molecular weight markers are phosphorylase b (96,000), bovine serum albumin (66,000), ovalbumin (45,000), carbonic anhydrase (30,000), soy bean trypsin inhibitor (20,000), and lactalbumin (14,Qw.
were discarded, and the supematants were incubated with either 200 ug of preimmune goat IgG or 200 ug of the IgG fraction of a goat antiserum reactive with hamster libroblast Fn receptor (kindly supplied by Dr. R. L. Juliano, University of North Carolina, Chapel Hill, NC). After 4 h on a rocking platform at 4”C, 60 ul of 20% 5. nureus was added and the incubation was continued overnight at 4°C. The mixture was centrifuged, and the precipitates were washed as previously described [32], except that 0.5 M NaCl was included in the second washing buffer. The washed precipitates were suspended in Laemmli sample buffer, boiled for 3 min, and centrifuged; the supematants were analyzed by SDS-PAGE and autoradiography.
General methods. SDS-PAGE was carried out in slab gels as described [33]. For detection of radioactive proteins, dried gels were exposed to X-Omat film (Eastman Kodak, Rochester, NY) with intensifying screens (Du Pont) for 1 to 2 days at -70°C. Gels of ‘H-labeled BO-kDa fragment were incubated with En’hance (Du Pont) for 2 h prior to exposure to X-ray films. Western blot analysis was performed as described [34]. N-Terminal amino acid sequence determination of Fn fragments was performed by Dr. Sukanto Sinha at the Protein Chemistry Laboratory of SUNY (Stony Brook, NY) on an Applied Biosystems 470A gas-phase sequencer.
RESULTS
Characterization of the 80-kDa fragment of Fn containing the cell-binding domain. In order to test the hypothesis that Fn fragments containing the cell- binding domain, but not the gelatin/collagen-binding domain, interfere with mo- nocyte function, we chose to study the interaction of the human monocytoid cell line U937 with an 80-kDa tryptic fragment which met these criteria. The fragment was purified by heparin-Sepharose, gelatin-Sepharose, and ion exchange chro-

424 Garcia-Pardo et al.
1000~- UI
% 0 600~- P 5 2
600--
4 z 400--
$ f 200--
1 0, - 0.1 1.0 10.0
Fibronectin or 60 kD Fragment (pg/cmz)
Fig. 2. Attachment of U937 cells to fibronectin and the 80-kDa tryptic fragment. Microtiter wells were coated with the indicated amounts of Fn (A) or the IO-kDa fragment (0). U937 cells (5X lO’/ml) were allowed to attach for 30 min at 37°C. Values represent the means of three separate experiments.
matography and was similar to other fragments which displayed binding affinities for fibroblast [21] and hepatocyte [22] Fn receptors.
The purity of the SO-kDa fragment was ascertained in part by SDS-PAGE (Fig. 1 A, lane I). The fragment also reacted in immunoblots with monoclonal antibod- ies which recognize the portion of Fn containing the cell-binding domain (Fig. IA, lane 2). One of these antibodies, 3H9.2, developed in our laboratory, not only recognizes a region encompassing the cell-binding domain but also inhibits the attachment of monocytes to gelatin-Fn-coated surfaces (unpublished results). A second antibody that reacted with the SO-kDa fragment (data not shown) was N294 (Mallinkrodt, Maryland Heights, MO). The N-terminal amino acid se- quence of the 80-kDa fragment was determined to be SD()VPSPR()LQF. The fragment begins, therefore, at position 874 of the Fn molecule [35], and, based on its apparent molecular weight, extends beyond the RGDS cell-binding sequence, which is located at positions 1493-1496 [35]. Purity of the 80-kDa fragment preparation was also assessed after tritiation by reductive alkylation. More than 95 % of the radioactivity was TCA precipitable and the labeled fragment ran as a single band on SDS-gels under reducing and nonreducing conditions (Fig. 1 B).
Attachment of U937 cells to Fn and &I-kDa fragment-coated surfaces. U937 cells attach to both Fn and to 80-kDa fragment-coated plates in a dose-dependent manner (Fig. 2). Saturation was achieved when 2.5 pg/cm* of Fn or 5 pg/cm’ of the fragment was deposited on the microtest wells. On a molar basis, however, Fn is approximately five times more effective than the fragment.
Table 1 summarizes some of the characteristics of the attachment of U937 cells to plastic wells coated with Fn or with the 80-kDa fragment. Preincubation of U937 cells with trypsin prevented cell attachment. Cells attached to either Fn- or 80-kDa-coated wells could be removed by incubation with 5 mM NaZEDTA. This indicates that divalent cations were required for the binding of U937 cells to Fn, and that the cell-surface molecules involved in this interaction were sensitive to
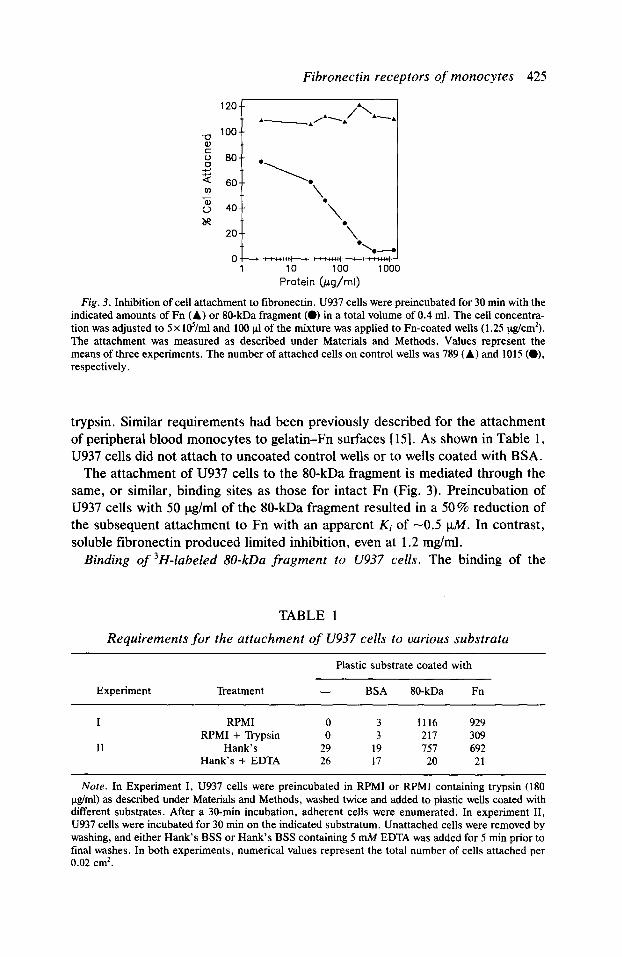
Fibronectin receptors of monocytes 425
120 :: , ,,..,. A-, ~ lOO-- (u
5 0
=: a m
II
::I '\.,
J 40.- 8? l \
20 -- l \., .-’ 01 : :::+++I : : ::+H : :::Hut
1 .
. 1 1 10 100 1000
Protein (w/ml)
Fig. 3. Inhibition of cell attachment to fibronectin. U937 cells were preincubated for 30 min with the indicated amounts of Fn (A) or IO-kDa fragment (0) in a total volume of 0.4 ml. The cell concentra- tion was adjusted to 5 x Id/ml and 100 pi of the mixture was applied to Fn-coated wells (1.25 &cm*). The attachment was measured as described under Materials and Methods. Values represent the means of three experiments. The number of attached cells on control wells was 789 (A) and 1015 (O), respectively.
trypsin. Similar requirements had been previously described for the attachment of peripheral blood monocytes to gelatin-Fn surfaces [ 151. As shown in Table 1, U937 cells did not attach to uncoated control wells or to wells coated with BSA.
The attachment of U937 cells to the 80-kDa fragment is mediated through the same, or similar, binding sites as those for intact Fn (Fig. 3). Preincubation of U937 cells with 50 Ctg/ml of the SO-kDa fragment resulted in a 50% reduction of the subsequent attachment to Fn with an apparent Ki of -0.5 u.M. In contrast, soluble fibronectin produced limited inhibition, even at 1.2 mg/ml.
Binding of 3H-labeled 80-kDa fragment to U937 cells. The binding of the
TABLE 1
Requirements for the attachment of U937 cells to various substrata
Plastic substrate coated with
Experiment Treatment - BSA 80-kDa Fn
I RPM1 0 3 1116 929 RPM1 + Trypsin 0 3 217 309
II Hank’s 29 19 1.57 692 Hank’s + EDTA 26 17 20 21
Nore. In Experiment I, U937 cells were preincubated in RPM1 or RPM1 containing trypsin (180 ug/ml) as described under Materials and Methods, washed twice and added to plastic wells coated with different substrates. After a 30-min incubation, adherent cells were enumerated. In experiment II, U937 cells were incubated for 30 min on the indicated substratum. Unattached cells were removed by washing, and either Hank’s BSS or Hank’s BSS containing 5 mM EDTA was added for 5 min prior to final washes. In both experiments, numerical values represent the total number of cells attached per 0.02 cm’.
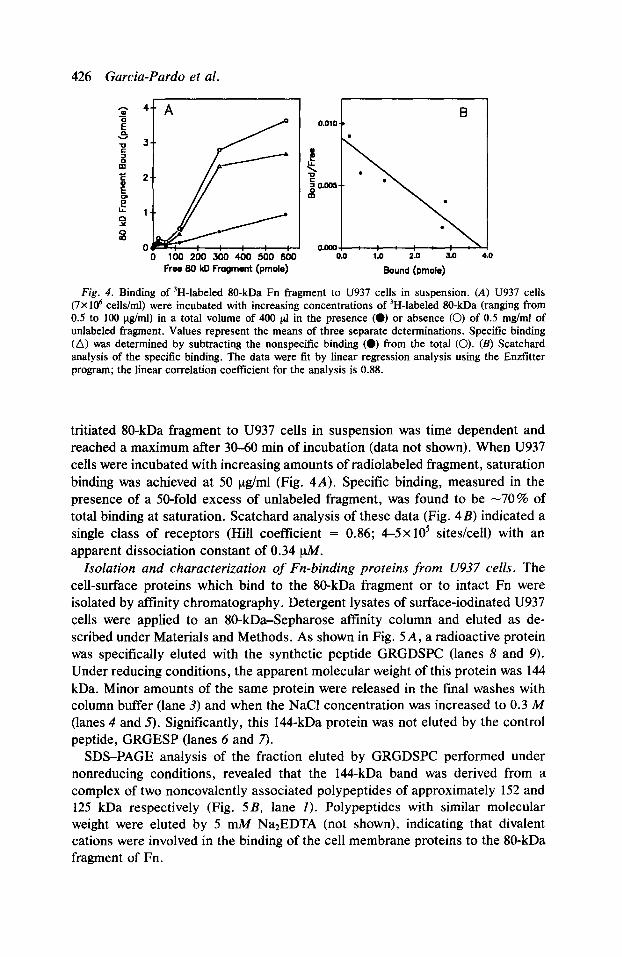
426 Garcia-Pardo et al.
0 100200300400500600 Free 80 kD Fmgment (pmole)
0.0 1.0 2.0 3.0 Bound (pmole)
D
Fig. 4. Binding of ‘H-labeled IO-kDa Fn fragment to U937 cells in suspension. (A) U937 cells (7X IO6 cells/ml) were incubated with increasing concentrations of ‘H-labeled %O-kDa (ranging from 0.5 to 100 &nl) in a total volume of 400 pl in the presence (0) or absence (0) of 0.5 mg/ml of unlabeled fragment. Values represent the means of three separate determinations. Specific binding (A) was determined by subtracting the nonspecific binding (0) from the total (0). (II) Scatchard analysis of the specific binding. The data were tit by linear regression analysis using the Enzfitter program; the linear correlation coefficient for the analysis is 0.88.
tritiated 80-kDa fragment to U937 cells in suspension was time dependent and reached a maximum after 30-60 min of incubation (data not shown). When U937 cells were incubated with increasing amounts of radiolabeled fragment, saturation binding was achieved at 50 ug/ml (Fig. 4A). Specific binding, measured in the presence of a 50-fold excess of unlabeled fragment, was found to be -70% of total binding at saturation. Scatchard analysis of these data (Fig. 4B) indicated a single class of receptors (Hill coefficient = 0.86; 4-5x 10’ sites/cell) with an apparent dissociation constant of 0.34 @f.
Isolation and characterization of Fn-binding proteins from U937 cells. The cell-surface proteins which bind to the 80-kDa fragment or to intact Fn were isolated by affinity chromatography. Detergent lysates of surface-iodinated U937 cells were applied to an 80-kDa-Sepharose affinity column and eluted as de- scribed under Materials and Methods. As shown in Fig. 5A, a radioactive protein was specifically eluted with the synthetic peptide GRGDSPC (lanes 8 and 9). Under reducing conditions, the apparent molecular weight of this protein was 144 kDa. Minor amounts of the same protein were released in the final washes with column buffer (lane 3) and when the NaCl concentration was increased to 0.3 M (lanes 4 and 5). Significantly, this 144-kDa protein was not eluted by the control peptide, GRGESP (lanes 6 and 7).
SDS-PAGE analysis of the fraction eluted by GRGDSPC performed under nonreducing conditions, revealed that the 144-kDa band was derived from a complex of two noncovalently associated polypeptides of approximately 152 and 125 kDa respectively (Fig. 5B, lane I). Polypeptides with similar molecular weight were eluted by 5 nuI4 Na*EDTA (not shown), indicating that divalent cations were involved in the binding of the cell membrane proteins to the 80-kDa fragment of Fn.

Fibronectin receptors of monocytes 427
1 23456769 10 11
-200
e
-116 -93
A
12 34
Fig. 5. Analysis of fibronectin-binding proteins from U937 cells. (A) Lysates from 10’ U937 cells were applied to 2 ml of 80-kDa fragment-Sepharose and 0.5 ml fractions were eluted as described under Materials and Methods. Aliquots (50~$) were lyophilized, redissolved in sample buffer, and analyzed on 7.5% SDS-polyacrylamide gels under reducing conditions. Lane I, lysate starting material; lanes 2-3, flow-through; lanes 4-11, material eluted with column buffer containing 0.3 M NaCl (lanes 4-5), GRGESP peptide (lanes 67), GRGDSPC peptide (lanes 8-9), and 8 M urea (lanes IO-II). Protein bands were detected by autoradiography. (B) Comparison of fibronectin-binding proteins from U937 cells and from peripheral blood monocytes. Fractions eluted from the IO-kDa atlinity column with the GRGDSPC peptide were analyzed on 7.5 % gels under nonreducing (lanes 1 and 2) and reducing conditions (lanes 3 and 4). Lanes 1 and 3, lysates of U937 cells; lanes 2 and 4, lysates of monocytes. Protein bands were detected by autoradiography. Molecular weight markers are myosin (200,000), @galactosidase (116,500), phosphorylase b (92,500), bovine serum albumin (66,200), and ovalbumin (45,000).
The same 152/125-kDa radioactive proteins were eluted from columns of Fn-Sepharose or gelatin-Fn-Sepharose either by 5 mM Na*EDTA or by the GRGDSPC peptide. In control experiments, specific binding of the 152/125-kDa complex to gelatin-Sepharose columns was not observed (data not shown).
Relationship to other Fn receptors. The Fn-binding proteins of surface iodinat- ed U937 cells could be immunoprecipitated with a goat antibody to the Fn receptor of hamster fibroblasts [36]. As shown in Fig. 6, both the 152- and the 125-kDa radioactive bands were precipitated by this antibody but not by a preimmune goat IgG (lanes 2 and 1, respectively).
The Fn-binding proteins of U937 cells were also compared to those of human peripheral blood monocytes [14, 151. Detergent lysates of surface iodinated monocytes were applied to the 80-kDa-Sepharose column and eluted as before. A polypeptide with an M, in the range of 136142 kDa was specifically eluted with GRGDSPC (Fig. 5B, lane 4) as revealed by SDS-PAGE performed under reduc- ing conditions; this protein resolved to a 148/122-kDa complex under nonreducing conditions (lane 2).
DISCUSSION
In this paper, we describe experiments on the interactions of U937 cells with Fn and with a proteolytically derived fragment of Fn, which may serve as a model

428 Garcia-Pardo et al.
Fig. 6. Immunoprecipitation of the GRGDSPC-eluted fraction from the 80-kDa-Sepharose affinity column. GRGDSPC-eluted fractions from 80-kDa fragment-Sepharose columns were treated with preimmune goat IgG (lane I) or goat IgG with specificity for hamster Fn receptor (lane 2). Samples were analyzed on 6% acrylamide gels under nonreducing conditions and proteins bands were detected by autoradiography as described under Materials and Methods. Molecular weight markers are the same as those in Fig. 5.
for the interactions of monocytes with matrices encountered at inflammatory sites. We have demonstrated that U937 cells bind to surfaces coated with Fn or an 80-kDa-derived fragment, and that the 80-kDa fragment inhibits attachment of these cells to intact Fn. On a molar basis, attachment of U937 cells to Fn (monomer molecular weight 220,000) was fivefold more effective than attachment of U937 cells to the 80-kDa fragment. This difference may reflect specific conformations attainable only by the intact molecule, or the presence of addition- al binding domains in the intact molecule.
Addition of 0.5 t&f of the 80-kDa fragment inhibited attachment of U937 cells to Fn-coated plates by 50 %. This apparent Ki for inhibition of attachment by the 80-kDa fragment is consistent with the Kd for binding of the 3H-labeled 80-kDa fragment (-0.34 p&f) determined by Scatchard analysis. Less than 10% inhibi- tion was observed with soluble, intact Fn at concentrations of 1.2 mg/ml. The inability of soluble, intact Fn to inhibit attachment of U937 cells supports previous observations suggesting that soluble Fn has low affinity for receptors on several types of cells [20-221. This is also supported by the finding that the Fn receptors of circulating monocytes are never saturated [15, 371.
Scatchard analysis also indicated the presence of approximately 4-5X lo5 80- kDa fragment binding sites per U937 cell. This value is similar to that obtained for the binding of a related Fn fragment to fibroblasts [38]. While the Hill coefficient calculated in our experiments (0.86) indicates the existence of a single class of binding sites in U937 cells, we note that the data presented in Fig. 4B may not be sufficient to exclude the possibility of additional classes of binding sites.

Fibronectin receptors of monocytes 429
In preliminary experiments, we have compared the attachment of U937 cells and peripheral blood monocytes to gelatin-fin-coated plates. Approximately 8 to lo-fold more Fn was required to achieve 50% attachment of U937 cells than was required for monocytes. The attachment of U937 cells to gelatin-Fn surfaces could be enhanced 4- to 6-fold, however, following treatment of the cells with phorbol myristate acetate (PMA) (0. C. Ferreira, unpublished observations). At this time, we have not determined the number of Fn receptors on peripheral blood monocytes or on PMA-stimulated U937 cells. While differences in receptor number could account for the observed enhancement in binding, several other explanations could be invoked: (1) unlike monocytes, the U937 cells Fn receptors may not be in a fully active state, perhaps because of receptor distribution in the membrane, or because interactions with other membrane proteins may be re- quired. Induction of differentiation might lead to membrane reorganization and to an associate increase in Fn-mediated adhesion; (2) monocytes and PMA-treated U937 cells synthesize Fn [39-40], which may contribute to the enhanced attach- ment of the cells.
The experiments described in this report suggest that the 80-kDa fragment and substrate-bound Fn react with the same receptor on U937 cells, and that similar protein complexes can be isolated from cell lysates by affinity chromatography on 80-kDa-Sepharose or gelatin-Sepharose-Fn columns. Identical results were obtained when Fn was directly coupled to cyanogen bromide-activated Sephar- ose, confirming that Fn immobilization leads to the expression of high-affinity binding sites for cell-surface receptors.
Chromatography on 80-kDa-Sepharose revealed that (a) the Fn-binding com- plex of U937 cells could be specifically eluted by the synthetic peptide GRGDSPC; (b) binding of the complex required divalent ions, a finding consist- ent with the ionic requirements for attachment of monocytes to gelatin-Fn-coated surfaces [15]; (c) the complex bound to WGA-Sepharose, indicating that the monocyte receptor, like other Fn receptors [7,41], is a glycoprotein; and, finally, (d) the complex was immunoprecipitated by a polyclonal antibody raised against the hamster tibroblast receptor.
When we employed Fn as an affinity ligand, the only protein specifically eluted with GRGDSPC or with EDTA was the 152/125-kDa complex. Recent reports have pointed to the existence of receptors which recognize sequences of Fn which are distinct from RGDS [32, 42, 431. Our experiments suggest that if these receptors exist in monocytoid cells, they are likely to be of lower affinity than the RGDS-related receptors, or binding of these receptors to Fn does not require divalent ions.
The Fn-binding proteins of U937 cells and peripheral blood monocytes are similar, in both apparent molecular weight and chain composition, to other members of the integrin family of receptors [16, 171. The receptor subunits of monocytes had a slightly faster electrophoretic mobility than those of U937 cells. This may reflect different patterns of glycosylation or heterogeneities in primary structure. Brown and co-workers have recently described what they believe to be two types of monocyte receptors. One class was isolated by affinity chromatogra-

430 Garcia-Pardo et al.
phy on columns of immobilized cell-binding fragments and the second on col- umns of a synthetic decapeptide derived from the cell-binding domain [181. The 152/125-kDa complex described here is similar to the first type of receptor reported by these investigators.
The ability of proteolytic fragments of Fn to bind to monocyte receptors, and perhaps to inhibit the interactions of monocytes with Fn bound to a solid phase, supports the contention that these fragments may play a role in uiuo. Excessive production of fragments of the type described at inflammatory sites may interfere with monocyte localization and subsequent differentiation. Further studies are required for validation of this hypothesis.
The authors thank Judith Miller and Donna Strauss for their expert technical assistance and Saida Perez for help in the preparation of the manuscript. This work was supported by Grants AI19751 and HL33860 from the National Institutes of Health (Bethesda, MD). O.C.F. is a recipient of a fellowship from the Conselho National de Desenvolvimento Cientifico e Tecnologico (CNPq), Brazil.
REFERENCES 1. Bianco, C. (1983) Ann. N.Y. Acad. Sci. 408, 602. 2. Brown, E. J. (1986) J. Leukocyte Biol. 39, 579. 3. Wright, S. D., Craigmyle, L. S., and Silverstein, S. C. (1983) J. Exp. Med. 158, 1338. 4. Clark, R. A. F., Dvorak, H. F., and Colvin, R. B. (1981) J. Zmmunol. 126, 787. 5. Bianco, C., and Edelson, P. J. (1977) in Immune Effector Mechanisms in Disease. (Siskind, G.,
and Weksler, M., Eds.), p. 1, Grune and Stratton, Inc., New York. 6. Pommier, C. G., Inada, S., Fries, L. F., Takahashi, T., Frank, M. M., and Brown, E. J. (1983) .Z.
Exp. Med. 157, 1844. 7. Pytela, R., Pierschbacher, M. D., and Ruoslahti, E. (1985) Cell 40, 191. 8. Akiyama, S. K., Yamada, S. S., and Yamada, K. M. (1986) .Z. Cell Biol. 102, 442. 9. Gardner, J. M., and Hynes, R. 0. (1985) Cell 42, 439.
10. Pierschbacher, M. D., and Ruoslahti, E. (1982) J. Biol. Chem. 257, 9593. 11. Yamada, K. M., and Kennedy, D. W. (1985) J. Cell. Biochem. 2.8, 99. 12. Ruoslahti, E., and Piershbacher, M. D. (1986) Cell 44, 517. 13. Wright, S. D., and Meyer, B. C. (1985) .Z. Exp. Med. 162, 762. 14. Hosein, B., and Bianco, C. (1985) J. Exp. Med. 162, 157. 15. Bevilacqua, M. P., Amrani, D., Mosesson, M. W., and Bianco, C. (1981) .Z. Exp. Med. 153, 42. 16. Hynes, R. 0. (1987) Cell 48, 549. 17. Ruoslahti, E., and Pierschbacher, M. D. (1987) Science 238, 491. 18. Brown, E. J., and Goodwin, J. L. (1988) J. Exp. Med. 167, 777. 19. Mosesson, M. W., and Umfleet, R. A. (1970) J. Biol. Chem. 245, 5728. 20. Molnar, J., Hoekstra, S., Ku, C. S. L., and Van Alten, P. (1987) J. Cell. Physiol. 131, 374. 21. Akiyama, S. K., and Yamada, K. H. (1985) J. Biol. Chem. 260, 4492. 22. Johansson, S. (1985) .Z. Biol. Chem. 260, 1557. 23. Postlethwaite, A. E., Keski-Oja, J., Balian, G., and Kang, A. H. (1981) J. Exp. Med. 153, 494. 24. De Petro, G., Barlati, S., Vartio, T., and Vaheri, A. (1981) Proc. Nat/. Acad. Sci. USA 78.4965. 25. Czop, J. K., and Austen, H. F. (1982) J. Zmmunol. 129, 2678. 26. Colvin, R. B., and Kradin, R. L. (1983) Suru. Synth. Pathol. Res. 2, 10. 27. Hormann, H., Richter, H., and Jelinic, V. (1987) Thrombosis Res. 46, 39. 28. Brown, R. A. (1983) Lancer 2, 1058. 29. Engvall, E., and Ruoslahti, E. (1977) Znt. J. Cancer 20, 1. 30. Tack, B. F., Dean, J., Eilat, D., Lorenz, P. E., and Schecter, A. N. (1980) J. Biol. Chem. 255,
8842. 31. Lebien, T. W., BouC, D. R., Bradley, J. G., and Kersey, J. H. (1982) .Z. Zmmunol. 129, 2287. 32. Bemardi, P., Patel, V. P., and Lodish, H. F. (1987) J. Cell Biol. 105, 489. 33. Laemmli, U. K. (1970) Nature (London) 227, 680. 34. Towbin, H., Staehlin, T., and Gordon, J. (1979) Proc. Nat/. Acad. Sci. USA 76, 4350.

Fibronectin receptors of monocytes 43 1
35. Komblihtt, A. R., Umezawa, K., Vibe-Pedersen, K., and Baralle, F. (1985) EMBO J. 4, 1755. 36. Brown, P. J., and Juliano, R. L. (1986) J. Cell Biol. 103, 1595. 37. Crinnell, F., Billingham, R. E., and Burgess, L. (1981) J. Znuesr. Dermatol. 76, 181. 38. Akiyama, S. K., Hasegawa, E., Hasegawa, T., and Yamada, K. M. (1985) J. Biol. Ckem. 260,
13256. 39. Alitalo, K., Hovi, T., and Vaheri, A. (1980) J. Exp. Med. 151, 602. 40. Minta, J. O., and Pambrun, L. (1985) Amer. J. Pntkol. 119, 111. 41. Johansson, S., Forsberg, E., and Lundgren, B. (1987) J. Biol. Ckem. 262, 7819. 42. Humphries, M. J., Komoriya, A., Akiyama, S. K., Olden, K., and Yamada, K. M. (1987) J. Biol.
Chew. 262, 6886. 43. McCarthy, J. B., Chelberg, M. K., Mickelson, D. J., and Furcht, L. T. (1988) Biochemistry 27,
1380.
Received August 23, 1988 Revised version received November 17, 1988
28-898334 Printed in Sweden