extracellular ca2+ sensing contributes to excess ca2+ accumulation and vacuolar fragmentation in a...
TRANSCRIPT

IntroductionChanges in the cytosolic Ca2+ concentration are thought toparticipate in a variety of physiological processes in yeast,including cell-cycle control (Hartley et al., 1996; Iida et al.,1990a); escape from G0/G1 arrest following carbon sourcelimitation (Eilam and Othman, 1990; Eilam et al., 1990;Kaibuchi et al., 1986); glucose and galactose sensing (Tokes-Fuzesi et al., 2002); the mating response (Nakajima-Shimadaet al., 1991; Withee et al., 1997); protein processing in thesecretory pathway (Durr et al., 1998); and adaptation toenvironmental stress (Batiza et al., 1996; Denis and Cyert,2002; Mori et al., 1998). The basic features of Ca2+ signalingin Saccharomyces cerevisiaeappear to be remarkably similarto the mechanisms used in mammalian cells. Ca2+ acts asa signaling ion in yeast through the activation of thecalmodulin/calcineurin pathway, which leads to alterations inthe transcription of a large number of genes (Matheos et al.,1997; Stathopoulos and Cyert, 1997; Yoshimoto et al., 2002).
A relatively small number of Ca2+ transporters appear tomaintain cellular Ca2+ homeostasis in yeast. Among these arethe vacuolar Ca2+-ATPase Pmc1p (Cunningham and Fink,1994b); the vacuolar Ca2+/H+ exchanger Vcx1p/Hum1p(Cunningham and Fink, 1996; Pozos et al., 1996); theendoplasmic reticulum (ER) Ca2+-ATPase Cod1p/Spf1p(Bonilla et al., 2002; Cronin et al., 2000; Cronin et al., 2002;Suzuki and Shimma, 1999); and the Golgi Ca2+-ATPase Pmr1p
(Antebi and Fink, 1992; Rudolph et al., 1989; Sorin et al.,1997). Remarkably, the action of this small group of Ca2+
transporters maintains the resting cytosolic Ca2+ level between50 and 200 nM when environmental Ca2+ concentrations rangefrom <1 µM to >100 mM (Batiza et al., 1996; Iida et al., 1990b;Miseta et al., 1999b). When the level of environmental Ca2+
is elevated, the calmodulin/calcineurin signaling pathwayactivates the expression of many genes (including thoseencoding Pmr1p and Pmc1p) by the transcription factorTcn1p/Crz1p. By contrast, the level of Vcx1p is slightlyreduced by high environmental Ca2+, and its activity is furtherrepressed by calcineurin activation through a post-translationalmechanism (Cunningham and Fink, 1996; Matheos et al.,1997; Stathopoulos-Gerontides et al., 1999; Yoshimoto et al.,2002).
The vacuole is the principle Ca2+ storage organelle in yeast,and normally contains >95% of the total cellular Ca2+ (Dunnet al., 1994; Eilam et al., 1985). However, it has been shownthat the loss of the Golgi-localized Ca2+ transporter Pmr1pcauses an increased sensitivity to high environmental Ca2+
when vacuolar Ca2+ transport is compromised, indicating thatthe Golgi apparatus also plays an important role in Ca2+
sequestration (Miseta et al., 1999b; Tanida et al., 1995).Furthermore, Pmr1p has also been reported to be involved inmaintaining the resting Ca2+ concentration within the ER(Strayle et al., 1999), whereas both Pmr1p and Pmc1p
1637
Previous studies have suggested that yeast strains lackingthe Ca2+-ATPase Pmr1p are unable to maintain anadequate level of Ca2+ within the Golgi apparatus. It isthought that this compartmental store depletion induces asignal that causes an increased rate of Ca2+ uptake andaccumulation in a manner similar to the capacitative Ca2+
entry (CCE) response in non-excitable mammalian cells. Toexplore this model further, we examined cellular Ca2+
uptake and accumulation in a pmr1∆ strain grown in thepresence of a reduced level of divalent cations. We foundthat the level of Ca2+ uptake and accumulation in a pmr1∆strain increased as the concentration of divalent cationsin the growth medium decreased. These results areinconsistent with a model in which cellular Ca2+ uptake and
accumulation are determined solely by the depletion ofCa2+ in an intracellular compartment. Instead, our resultssuggest that a second regulatory mechanism couplescellular Ca2+ uptake to the availability of Ca2+ in theextracellular environment. Furthermore, we found thatvarious conditions that increase the level of cytosolic Ca2+
correlate with vacuolar fragmentation in wild-type (WT),pmr1∆ and pmr1∆/pmc1∆ yeast strains. This suggests thatvacuolar fragmentation might function as a normalphysiological response to Ca2+ stress that increases thevacuolar surface/volume ratio, thereby maximizing thesequestration of this important signaling molecule.
Key words: Yeast, Pmr1p, Ca2+ homeostasis, Vacuole
Summary
Extracellular Ca 2+ sensing contributes to excess Ca 2+
accumulation and vacuolar fragmentation in a pmr1 ∆mutant of S. cerevisiaeRichard Kellermayer 1, David P. Aiello 1, Attila Miseta 2 and David M. Bedwell 1,*1Department of Microbiology, University of Alabama at Birmingham, Birmingham, AL 35294, USA2Department of Clinical Chemistry, University Medical School, 7624 Peçs, Hungary*Author for correspondence (e-mail: [email protected])
Accepted 16 January 2003Journal of Cell Science 116, 1637-1646 © 2003 The Company of Biologists Ltddoi:10.1242/jcs.00372
Research Article

1638
influence Ca2+-dependent functions within the secretorypathway such as protein degradation in the ER and proteinsorting in the Golgi apparatus (Durr et al., 1998). The abilityof these transporters to influence Ca2+-dependent processes inmultiple organelles might be due to their movement throughthese compartments during the transit to their final cellulardestinations. However, it has been reported that the distributionof Pmc1p in Golgi fractions increases in a pmr1∆ strain,suggesting that its abundance in compartments of the secretorypathway might be influenced by the lumenal Ca2+
concentration (Marchi et al., 1999). These findings illustrateboth the complexity and the sophisticated regulatorymechanisms that control cellular Ca2+ homeostasis in yeast.
In mammalian cells, free Ca2+ located in the ER serves asa mobilizable pool that can be released into the cytosol inresponse to an appropriate stimulus. The resulting increase incytosolic Ca2+ can then activate signaling pathways that alterthe expression of many genes in a coordinated manner (Putney,1992). In many non-excitable cells, the release of ER Ca2+ canalso induce a store depletion signal that results in the influx ofCa2+ ions across the plasma membrane in a process termedcapacitative Ca2+ entry (CCE). Recent studies have providedevidence that yeast cells might also utilize a mechanism thatcouples intracellular store depletion to Ca2+ uptake across theplasma membrane. A pmr1∆ mutant has been shown to exhibita higher rate of Ca2+ uptake than the WT strain (Antebi andFink, 1992; Halachmi and Eilam, 1996; Rudolph et al., 1989;Sorin et al., 1997). This led to the model that the depletion ofGolgi Ca2+ stores can stimulate Ca2+ uptake into yeast cells ina manner analogous to CCE in mammalian cells (Csutora etal., 1999; Durr et al., 1998; Locke et al., 2000). In contrast tothe pmr1∆ mutant, a pmc1∆ strain exhibits a reduced level oftotal cellular Ca2+ (Cunningham and Fink, 1994b). SincePmc1p is the only known vacuolar Ca2+-ATPase in yeast andthe vacuole normally contains the bulk of total cellular Ca2+,this suggests that Ca2+ uptake across the plasma membrane iscoupled to the ability of the cell to remove it efficiently fromthe cytosol.
In the current study, we examined how a reduced level ofdivalent cations in the growth medium influences cellular Ca2+
homeostasis in the pmr1∆ strain. We found a large increase inCa2+ uptake and accumulation under these conditions, which ledto activation of the calcineurin signaling pathway. Consistentwith this high level of Ca2+ uptake, we found that the PMC1gene was required for growth of the pmr1∆ mutant under thesegrowth conditions. Our observation that cellular Ca2+ uptakeincreases as the concentration of environmental Ca2+ decreasessuggests that an extracellular Ca2+ sensor is capable of couplingCa2+ uptake to extracellular Ca2+ levels. Finally, we foundthat conditions of cellular Ca2+ stress result in a vacuolarfragmentation phenotype in both WT and mutant yeast strains.
This might serve as an adaptive mechanism to maintain cellularCa2+ homeostasis under these stress conditions.
Materials and MethodsStrains and plasmids usedThe yeast strains used in this study are described in Table 1. ThePmc1p-GFP fusion plasmid was constructed as follows. A 4.9 kbHindIII fragment containing the PMC1 gene was isolated frompKC44 (gift from Kyle Cunningham) and ligated into pBlueKS+.Site-directed mutagenesis was performed according to theQuickChange Site Directed Mutagenesis (Stratagene) protocol togenerate a BssHII site at the 3′ end of PMC1 (5 codons upstreamof the stop codon) using the oligonucleotides DB571 (5′-CTGATAGTCC TTGGCGCGCC AACTTTTATT AATAGACGC-3′)and DB572 (5′-GCGTCTATTA ATAAAAGTTG GCGCGCCAAGGACTATCAG-3′). A 635 bp BglII/SnaBI fragment was isolated fromthe resulting plasmid and cloned back into pBlueKS+/PMC1for DNAsequencing. A BssHII DNA fragment carrying the GFP gene wasgenerated using primers DB573 (5′-CGATAAGGAT TGGCGCG-CCA AATTCATGAG-3′) and DB574 (5′-CGATAAGGAT TGGCG-CGCCA AATTCATGAG-3′) and the template pRSETB-GFP (giftfrom Kelly Thatchell), and ligated into the BssHII site ofpBlueKS+/PMC1. The correct GFP coding sequence was confirmedby automated DNA sequencing and a HindIII fragment carrying thePMCI-GFP construct was ligated into the low-copy yeast shuttlevector pSEYC58 (Emr et al., 1986). This final plasmid was thentransformed into the pmc1∆ (YDB0224) and the pmr1∆/pmc1∆(YDB0276) strains. The HindIII fragment carrying the completePMC1-GFP open reading frame was also inserted into the high-copyshuttle vector pSEY8 (Emr et al., 1986), digested with ApaI, and thedigested linear DNA was transformed into the pmc1∆ (YDB0224) andthe pmr1∆/pmc1∆ (YDB0276) strains to integrate it into the genomeof each strain. With both approaches, the construct reversed theEGTA-sensitive phenotype of the pmr1∆/pmc1∆ strain and the highCa2+-sensitive phenotype of the pmc1∆ strain.
Culture mediaBacterial strains were grown on standard media (Miller, 1992). Yeaststrains were maintained on YP medium containing 2% D-glucose(YPD) or synthetic minimal medium containing 2% D-glucose (SMD)and other supplements as required (Burke et al., 2000). Culture mediawere routinely buffered with 40 mM Mes-Tris, pH 5.5.
Total cellular Ca2+ measurementsTotal cellular Ca2+ was measured using flame photometry as describedpreviously (Miseta et al., 1999a). Cells were grown in 30-40 ml ofYPD or the same medium supplemented with either 1 mM EGTA or50 mM CaCl2. Cultures were harvested at a cell density of ~1OD600/ml, harvested by centrifugation and washed with 20 ml ofYPD. Cells were then transferred to previously weighed microfugetubes and harvested by centrifugation in a micro-centrifuge (16,000g) for 5 minutes. After aspirating the supernatant, a second spin was
Journal of Cell Science 116 (8)
Table 1. Yeast strains used in this studyStrain Genotype Source
SEY6210 MATα, ura3-52, leu2-3,112, his3-∆200, trp1-∆901, lys2-801, suc2-∆9 S. Emr, UCSD, San Diego, CAYDB224 MATα, ura3-52, leu2-3,112, his3-∆200, trp1-∆901, lys2-801, suc2-∆9, pmc1∆::TRP1 Miseta et al., 1999aYDB225 MATα, ura3-52, leu2-3,112, his3-∆200, trp1-∆901, lys2-801, suc2-∆9, vcx1∆::URA3 Miseta et al., 1999aYDB279 MATa, ura3-52, leu2-3,112, his3-∆200, trp1-∆901, lys2-801, suc2-∆9, pmr1∆::LEU2 Miseta et al., 1999aYDB254 MATα, ura3-52, leu2-3,112, his3-∆200, trp1-∆901, lys2-801, suc2-∆9, pmc1∆::TRP1, vcx1∆::URA3 Miseta et al., 1999aYDB276 MATa, trp1-∆901, ura3-52, his3-∆200, leu2-3,112, pmc1∆::TRP1, pmr1∆::LEU2 This studyYDB289 MATa, trp1-∆901, ura3-52, his3-∆200, leu2-3,112, lys2-80, vcx1∆::URA3, pmr1∆::LEU2 This study

1639Ca2+ homeostasis in yeast
conducted for 3 minutes. Samples were weighed, then dried in aSpeedvac. The dried samples were weighed again and resuspended inHCl for flame photometric measurements.
Ca2+ uptakeCa2+ uptake measurements were similar to a method describedpreviously (Halachmi and Eilam, 1996). Cells were grown in YPD to0.7-1.0 OD600/ml, harvested, washed twice with ddH2O, then re-suspended in buffer containing 25 mM Mes-Tris, pH 6.0supplemented with 20 mM glucose to 1 OD600/ml and incubated at30°C for 10 minutes. Uptake was initiated by the addition of 1 µCi/ml45Ca2+. At the indicated time-points, 1 ml aliquots were collected byfiltration through 0.45 µm membrane filters (Gelman Sciences) usinga vacuum manifold. Membranes were immediately washed with two5 ml aliquots of ice-cold wash buffer (20 mM MgCl2, 0.2 mM LaCl3),dried, and the cell-associated radioactivity was measured by liquidscintillation counting.
Northern analysisRNA extraction and northern blot analysis were carried out asdescribed previously (Bonetti et al., 1995). Strains were grownovernight in YPD medium or YPD supplemented with either 1 mMEGTA or 50 mM CaCl2 to ~1 OD600/ml. A 0.44 kb probe for thePMC1 mRNA was generated by PCR using primers DB490 (5′-ATGTCTAGACAAGACGAAAA-3 ′) and DB491 (5′-ATACTGTG-GAGGTTGCATCC-3′). As control, an ACT1 probe was generatedusing the primers DB154 (5′-GCGCGGAATTCAACGTTCCAG-CCTTCTACG-3′) and DB155 (5′-GGATGGAACAAAGCTTCTGG-3′). Probes were labeled with [α-32P]dATP using the random hexamermethod (Sambrook et al., 1989). The specific band representing thePMC1 mRNA was confirmed by its absence in RNA extractedfrom the pmc1∆ strain YDB0224. Gels were quantitated byPhosphorImager analysis (Molecular Dynamics). The relative mRNAlevels were normalized using the ACT1 mRNA as internal controlafter background correction.
Aequorin assayAequorin assays were carried out as described earlier (Miseta et al.,1999a). The two-micron-based plasmid pDB617 expressing afunctional apoaequorin gene (pAEQ) was transformed into yeast.Cells containing pAEQ were grown in SMD medium and harvestedin the early logarithmic growth phase (0.5-1.0 A600units/ml). 10 A600units of cells were harvested and re-suspended in 0.2 ml of aequorintest medium, which consists of SMD medium (which normallycontains 1 mM Ca2+) supplemented with 2 mM EGTA and 40 mMMES-Tris, pH 6.5. To convert the apoaequorin to aequorin, 20 µl of590 µM coelenterazine (dissolved in methanol) was added, and thecells were incubated for 20 minutes at room temperature. The cellswere then briefly centrifuged, and the supernatant containing excesscoelenterazine was removed. The cell pellets were washed again in0.5 ml of aequorin test medium, re-suspended to a cell density of 1OD600/0.1 ml and incubated at room temperature for 20 minutesbefore initiating the experiment. Bafilomycin A1 (5 µM) was addedfrom a 100 µM stock solution (dissolved in DMSO) 10 minutes priorto the measurements (Abe and Horikoshi, 1995). After detecting thebaseline light emission, 100 mM Ca2+ was administered into thechamber to generate a Ca2+ shock. A Berthold Lumat 9050luminometer was used to collect aequorin light emission (L) data at200 millisecond intervals. The data were downloaded directly to acomputer and transferred to Microsoft Excel 5.0 for analysis. Aftermeasuring maximal light emission from crude cell extracts upon Ca2+
addition (Lmax), L/Lmax values were plotted on a standard curve toestimate the free cytosolic Ca2+ concentrations, as previouslydescribed (Miseta et al., 1999a).
Light microscopyCells expressing the Pmc1p-GFP fusion protein and/or stained withFM 4-64 were collected, re-suspended in fresh YPD medium, orYPD medium supplemented with Ca2+ or EGTA, to 10 OD600units/ml and 10 µl of the culture was mounted on slides coated withconcanavalin A, covered with a coverslip and viewed immediately.FM 4-64 staining was carried out as published (Vida and Emr, 1995)with minor modifications. Briefly, yeast cells were grown in theindicated media to mid-log phase, 2 OD600 units of cells werecollected and re-suspended in 100 µl of YPD (Ca2+ was omitted atthis step because it decreased total fluorescence). A 1 µl aliquot ofan FM 4-64 stock (4 mM in DMSO) was added to the cells, and theywere stained for 10-15 minutes at 30°C. Cells were collected andre-suspended in 200 µl fresh media supplemented with Ca2+ orEGTA and incubated at 30°C for 40-50 minutes. Light microscopywas conducted using a Leitz Orthoplan microscope withepifluorescence optics and Hoffman Modulation Contrast optics.The images were acquired with a Photometrics CH250 liquid-cooledCCD high-resolution monochromatic camera (Roper Scientific;Tucson, AZ) and analyzed by IPLab Spectrum software fromScanalytics (Fairfax, VA).
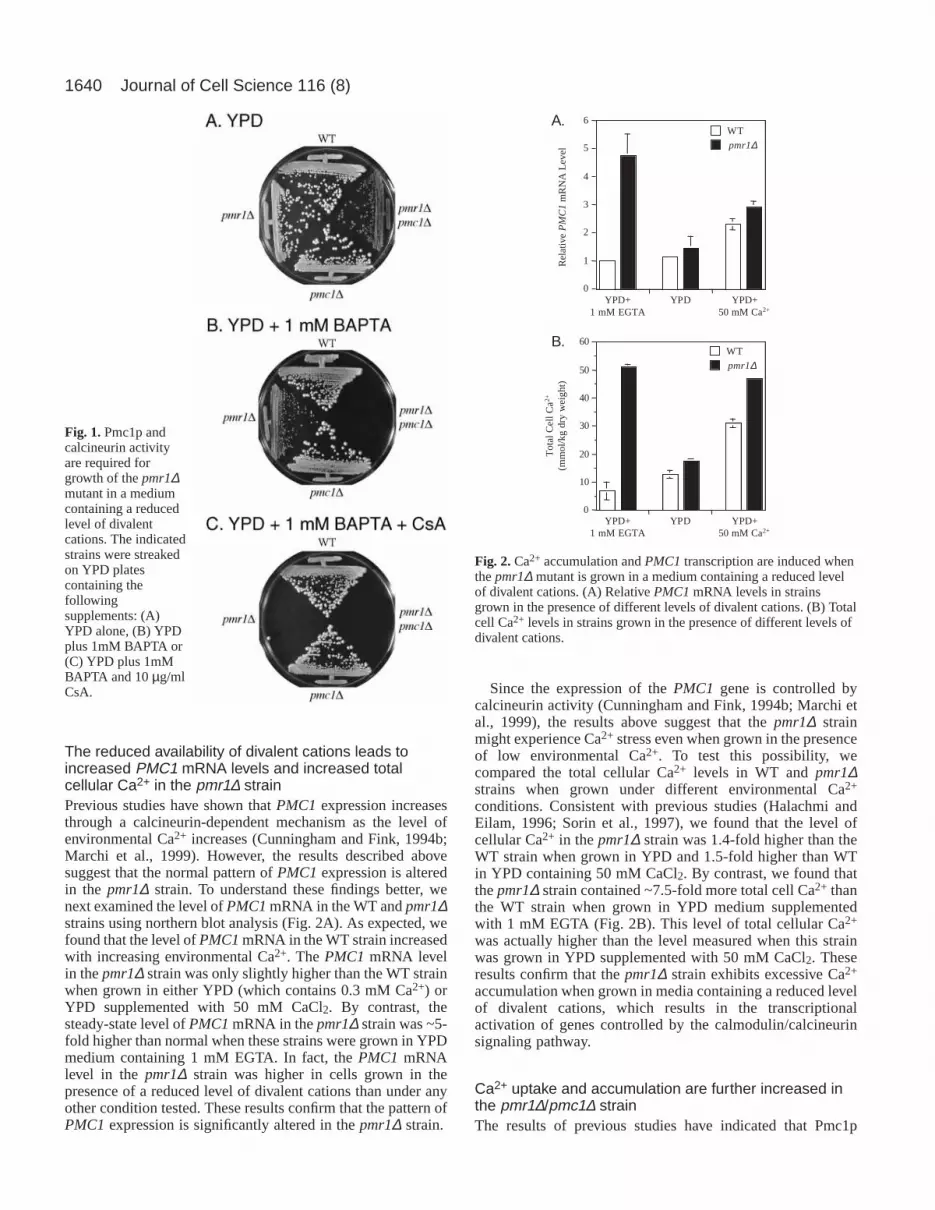
ResultsPMC1 expression is required for growth of the pmr1∆mutant in an environment containing a reduced level ofCa2+ and other divalent cationsWe first examined whether the loss of Pmc1p function alteredthe ability of thepmr1∆ strain to grow in the presence ofchelating agents that reduced the level of Ca2+ and otherdivalent cations. We found that the pmr1∆ strain could growon YPD plates containing the Ca2+ chelator 1,2 bis(2-aminophenoxy) ethane-N,N,N,N-tetraacetic acid (BAPTA; 1mM), whereas the pmr1∆/pmc1∆ strain could not (Fig.1A,B). Similar results were obtained with YPD platescontaining 2 mM EGTA (data not shown). These results wereconsistent with the previous conclusion that Pmc1p plays arole in Ca2+ uptake into secretory compartments (Bonilla etal., 2002; Durr et al., 1998; Locke et al., 2000). However,it was surprising that Pmc1p influenced cell growth whenthe availability of Ca2+ and other divalent cations wasreduced, since previous studies have shown that PMC1expression and activity is normally induced by calcineurinwhen cellular Ca2+ stress increases (Cunningham and Fink,1994b; Marchi et al., 1999). To determine whethercalcineurin activation is required for growth of the pmr1∆strain under these conditions, we challenged these strainswith cyclosporin A (CsA) on YPD plates containing 1 mMBAPTA (Fig. 1C). We found that neither the pmr1∆ nor thepmr1∆/pmc1∆ strains were able to grow in the presence ofboth CsA and BAPTA, demonstrating that calcineurin activityis necessary for growth of the pmr1∆ mutant when theenvironmental concentration of divalent cations is decreased.CsA inhibition was also observed when these strains weregrown on YPD plates containing 2 mM EGTA, but not onYPD plates containing CsA that lacked one of these chelatingagents (data not shown). When taken together, these resultsindicate that one or more genes regulated by the calcineurinsignaling pathway, including Pmc1p, plays an importantrole in Ca2+ homeostasis in the pmr1∆ strain when theconcentration of divalent cations in the environmental isreduced.
1640
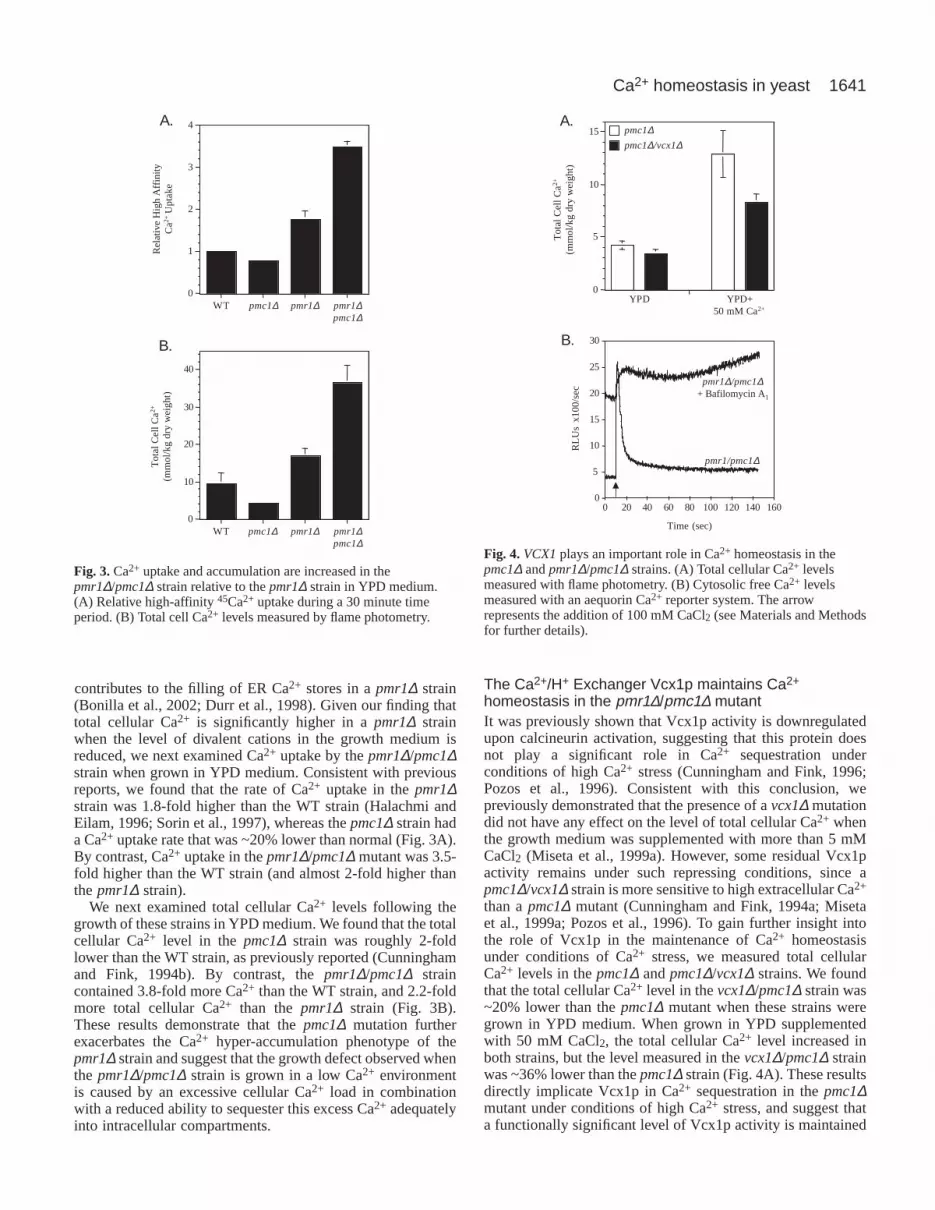
The reduced availability of divalent cations leads toincreased PMC1 mRNA levels and increased totalcellular Ca2+ in the pmr1∆ strainPrevious studies have shown that PMC1 expression increasesthrough a calcineurin-dependent mechanism as the level ofenvironmental Ca2+ increases (Cunningham and Fink, 1994b;Marchi et al., 1999). However, the results described abovesuggest that the normal pattern of PMC1expression is alteredin the pmr1∆ strain. To understand these findings better, wenext examined the level of PMC1mRNA in the WT and pmr1∆strains using northern blot analysis (Fig. 2A). As expected, wefound that the level of PMC1mRNA in the WT strain increasedwith increasing environmental Ca2+. The PMC1 mRNA levelin the pmr1∆ strain was only slightly higher than the WT strainwhen grown in either YPD (which contains 0.3 mM Ca2+) orYPD supplemented with 50 mM CaCl2. By contrast, thesteady-state level of PMC1mRNA in the pmr1∆ strain was ~5-fold higher than normal when these strains were grown in YPDmedium containing 1 mM EGTA. In fact, the PMC1 mRNAlevel in the pmr1∆ strain was higher in cells grown in thepresence of a reduced level of divalent cations than under anyother condition tested. These results confirm that the pattern ofPMC1expression is significantly altered in the pmr1∆ strain.
Since the expression of the PMC1 gene is controlled bycalcineurin activity (Cunningham and Fink, 1994b; Marchi etal., 1999), the results above suggest that the pmr1∆ strainmight experience Ca2+ stress even when grown in the presenceof low environmental Ca2+. To test this possibility, wecompared the total cellular Ca2+ levels in WT and pmr1∆strains when grown under different environmental Ca2+
conditions. Consistent with previous studies (Halachmi andEilam, 1996; Sorin et al., 1997), we found that the level ofcellular Ca2+ in the pmr1∆ strain was 1.4-fold higher than theWT strain when grown in YPD and 1.5-fold higher than WTin YPD containing 50 mM CaCl2. By contrast, we found thatthe pmr1∆ strain contained ~7.5-fold more total cell Ca2+ thanthe WT strain when grown in YPD medium supplementedwith 1 mM EGTA (Fig. 2B). This level of total cellular Ca2+
was actually higher than the level measured when this strainwas grown in YPD supplemented with 50 mM CaCl2. Theseresults confirm that the pmr1∆ strain exhibits excessive Ca2+
accumulation when grown in media containing a reduced levelof divalent cations, which results in the transcriptionalactivation of genes controlled by the calmodulin/calcineurinsignaling pathway.
Ca2+ uptake and accumulation are further increased inthe pmr1∆/pmc1∆ strainThe results of previous studies have indicated that Pmc1p
Journal of Cell Science 116 (8)
Fig. 1.Pmc1p andcalcineurin activityare required forgrowth of the pmr1∆mutant in a mediumcontaining a reducedlevel of divalentcations. The indicatedstrains were streakedon YPD platescontaining thefollowingsupplements: (A)YPD alone, (B) YPDplus 1mM BAPTA or(C) YPD plus 1mMBAPTA and 10 µg/mlCsA.
0
1
2
3
4
5
6
Rel
ativ
e PM
C1
mR
NA
Lev
el
WT
pmr1∆
YPD+1 mM EGTA
YPD YPD+50 mM Ca2+
0
10
20
30
40
50
60
Tot
al C
ell C
a2+
(mm
ol/k
g d
ry w
eig
ht)
WT
pmr1∆
YPD+1 mM EGTA
YPD YPD+50 mM Ca2+
A.
B.
Fig. 2.Ca2+ accumulation and PMC1transcription are induced whenthe pmr1∆ mutant is grown in a medium containing a reduced levelof divalent cations. (A) Relative PMC1mRNA levels in strainsgrown in the presence of different levels of divalent cations. (B) Totalcell Ca2+ levels in strains grown in the presence of different levels ofdivalent cations.
1641Ca2+ homeostasis in yeast
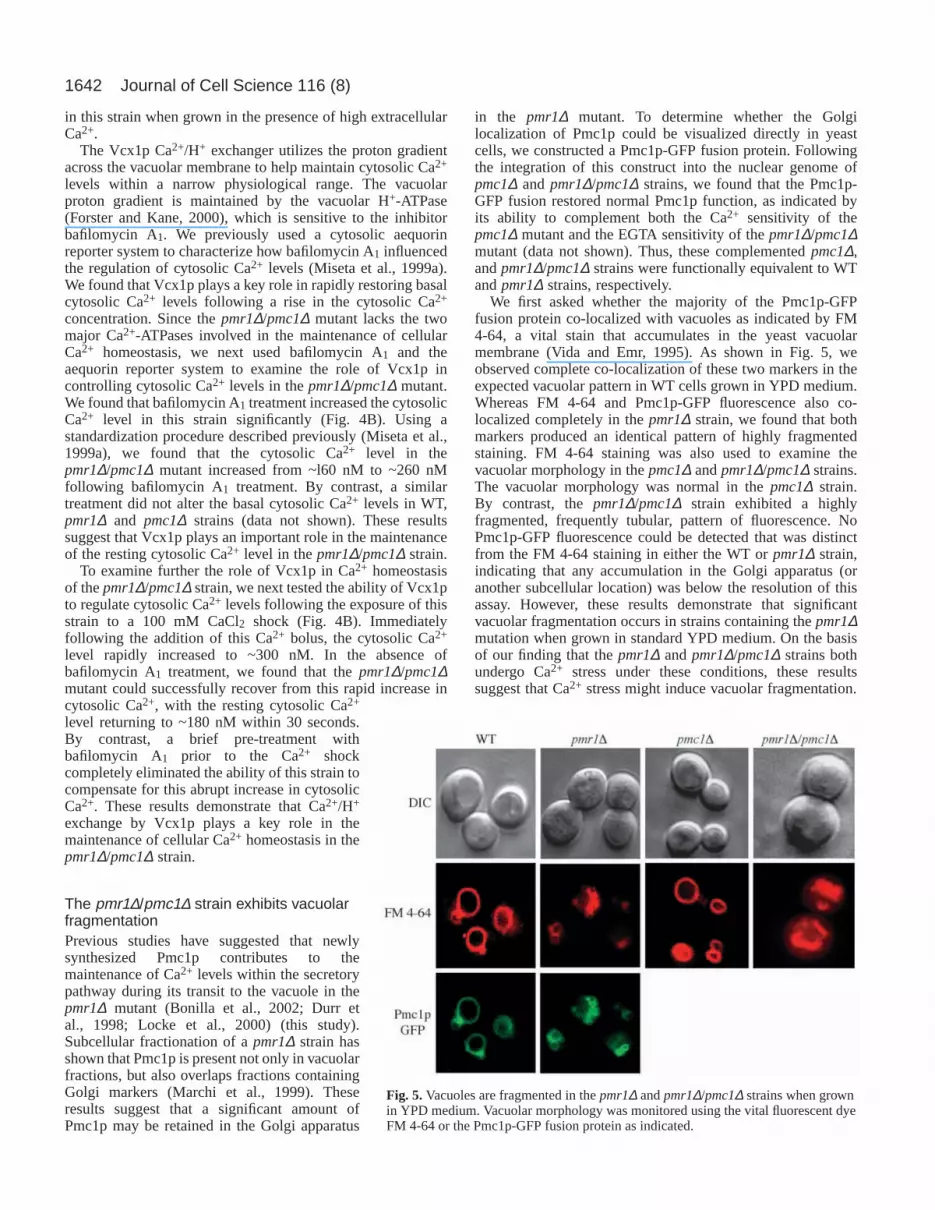
contributes to the filling of ER Ca2+ stores in a pmr1∆ strain(Bonilla et al., 2002; Durr et al., 1998). Given our finding thattotal cellular Ca2+ is significantly higher in a pmr1∆ strainwhen the level of divalent cations in the growth medium isreduced, we next examined Ca2+ uptake by thepmr1∆/pmc1∆strain when grown in YPD medium. Consistent with previousreports, we found that the rate of Ca2+ uptake in the pmr1∆strain was 1.8-fold higher than the WT strain (Halachmi andEilam, 1996; Sorin et al., 1997), whereas the pmc1∆ strain hada Ca2+ uptake rate that was ~20% lower than normal (Fig. 3A).By contrast, Ca2+ uptake in the pmr1∆/pmc1∆ mutant was 3.5-fold higher than the WT strain (and almost 2-fold higher thanthe pmr1∆ strain).
We next examined total cellular Ca2+ levels following thegrowth of these strains in YPD medium. We found that the totalcellular Ca2+ level in the pmc1∆ strain was roughly 2-foldlower than the WT strain, as previously reported (Cunninghamand Fink, 1994b). By contrast, the pmr1∆/pmc1∆ straincontained 3.8-fold more Ca2+ than the WT strain, and 2.2-foldmore total cellular Ca2+ than the pmr1∆ strain (Fig. 3B).These results demonstrate that the pmc1∆ mutation furtherexacerbates the Ca2+ hyper-accumulation phenotype of thepmr1∆ strain and suggest that the growth defect observed whenthe pmr1∆/pmc1∆ strain is grown in a low Ca2+ environmentis caused by an excessive cellular Ca2+ load in combinationwith a reduced ability to sequester this excess Ca2+ adequatelyinto intracellular compartments.
The Ca2+/H+ Exchanger Vcx1p maintains Ca2+
homeostasis in the pmr1∆/pmc1∆ mutantIt was previously shown that Vcx1p activity is downregulatedupon calcineurin activation, suggesting that this protein doesnot play a significant role in Ca2+ sequestration underconditions of high Ca2+ stress (Cunningham and Fink, 1996;Pozos et al., 1996). Consistent with this conclusion, wepreviously demonstrated that the presence of a vcx1∆ mutationdid not have any effect on the level of total cellular Ca2+ whenthe growth medium was supplemented with more than 5 mMCaCl2 (Miseta et al., 1999a). However, some residual Vcx1pactivity remains under such repressing conditions, since apmc1∆/vcx1∆ strain is more sensitive to high extracellular Ca2+
than a pmc1∆ mutant (Cunningham and Fink, 1994a; Misetaet al., 1999a; Pozos et al., 1996). To gain further insight intothe role of Vcx1p in the maintenance of Ca2+ homeostasisunder conditions of Ca2+ stress, we measured total cellularCa2+ levels in the pmc1∆ and pmc1∆/vcx1∆ strains. We foundthat the total cellular Ca2+ level in the vcx1∆/pmc1∆ strain was~20% lower than the pmc1∆ mutant when these strains weregrown in YPD medium. When grown in YPD supplementedwith 50 mM CaCl2, the total cellular Ca2+ level increased inboth strains, but the level measured in the vcx1∆/pmc1∆ strainwas ~36% lower than the pmc1∆ strain (Fig. 4A). These resultsdirectly implicate Vcx1p in Ca2+ sequestration in the pmc1∆mutant under conditions of high Ca2+ stress, and suggest thata functionally significant level of Vcx1p activity is maintained
0
1
2
3
4
Rel
ativ
e H
igh
Affi
nity
Ca2+
Upt
ake
WT pmc1∆ pmr1∆ pmr1∆pmc1∆
A.
Tot
al C
ell C
a2+
(mm
ol/k
g d
ry w
eig
ht)
0
10
20
30
40
WT pmc1∆ pmr1∆ pmr1∆pmc1∆
B.
Fig. 3.Ca2+ uptake and accumulation are increased in thepmr1∆/pmc1∆ strain relative to the pmr1∆ strain in YPD medium.(A) Relative high-affinity 45Ca2+ uptake during a 30 minute timeperiod. (B) Total cell Ca2+ levels measured by flame photometry.
0
5
10
15 pmc1∆pmc1∆/vcx1∆
YPD YPD+50 mM Ca2+
Tot
al C
ell C
a2+
(mm
ol/k
g d
ry w
eig
ht)
A.
B.
0
5
10
15
20
25
30
0 20 40 60 80 100 120 140 160
Time (sec)
RL
Us
x10
0/se
c
pmr1/pmc1∆
pmr1∆/pmc1∆+ Bafilomycin A1
Fig. 4.VCX1plays an important role in Ca2+ homeostasis in thepmc1∆ and pmr1∆/pmc1∆ strains. (A) Total cellular Ca2+ levelsmeasured with flame photometry. (B) Cytosolic free Ca2+ levelsmeasured with an aequorin Ca2+ reporter system. The arrowrepresents the addition of 100 mM CaCl2 (see Materials and Methodsfor further details).
1642
in this strain when grown in the presence of high extracellularCa2+.
The Vcx1p Ca2+/H+ exchanger utilizes the proton gradientacross the vacuolar membrane to help maintain cytosolic Ca2+
levels within a narrow physiological range. The vacuolarproton gradient is maintained by the vacuolar H+-ATPase(Forster and Kane, 2000), which is sensitive to the inhibitorbafilomycin A1. We previously used a cytosolic aequorinreporter system to characterize how bafilomycin A1 influencedthe regulation of cytosolic Ca2+ levels (Miseta et al., 1999a).We found that Vcx1p plays a key role in rapidly restoring basalcytosolic Ca2+ levels following a rise in the cytosolic Ca2+
concentration. Since the pmr1∆/pmc1∆ mutant lacks the twomajor Ca2+-ATPases involved in the maintenance of cellularCa2+ homeostasis, we next used bafilomycin A1 and theaequorin reporter system to examine the role of Vcx1p incontrolling cytosolic Ca2+ levels in the pmr1∆/pmc1∆ mutant.We found that bafilomycin A1 treatment increased the cytosolicCa2+ level in this strain significantly (Fig. 4B). Using astandardization procedure described previously (Miseta et al.,1999a), we found that the cytosolic Ca2+ level in thepmr1∆/pmc1∆ mutant increased from ~l60 nM to ~260 nMfollowing bafilomycin A1 treatment. By contrast, a similartreatment did not alter the basal cytosolic Ca2+ levels in WT,pmr1∆ and pmc1∆ strains (data not shown). These resultssuggest that Vcx1p plays an important role in the maintenanceof the resting cytosolic Ca2+ level in the pmr1∆/pmc1∆ strain.
To examine further the role of Vcx1p in Ca2+ homeostasisof the pmr1∆/pmc1∆ strain, we next tested the ability of Vcx1pto regulate cytosolic Ca2+ levels following the exposure of thisstrain to a 100 mM CaCl2 shock (Fig. 4B). Immediatelyfollowing the addition of this Ca2+ bolus, the cytosolic Ca2+
level rapidly increased to ~300 nM. In the absence ofbafilomycin A1 treatment, we found that thepmr1∆/pmc1∆mutant could successfully recover from this rapid increase incytosolic Ca2+, with the resting cytosolic Ca2+
level returning to ~180 nM within 30 seconds.By contrast, a brief pre-treatment withbafilomycin A1 prior to the Ca2+ shockcompletely eliminated the ability of this strain tocompensate for this abrupt increase in cytosolicCa2+. These results demonstrate that Ca2+/H+
exchange by Vcx1p plays a key role in themaintenance of cellular Ca2+ homeostasis in thepmr1∆/pmc1∆ strain.
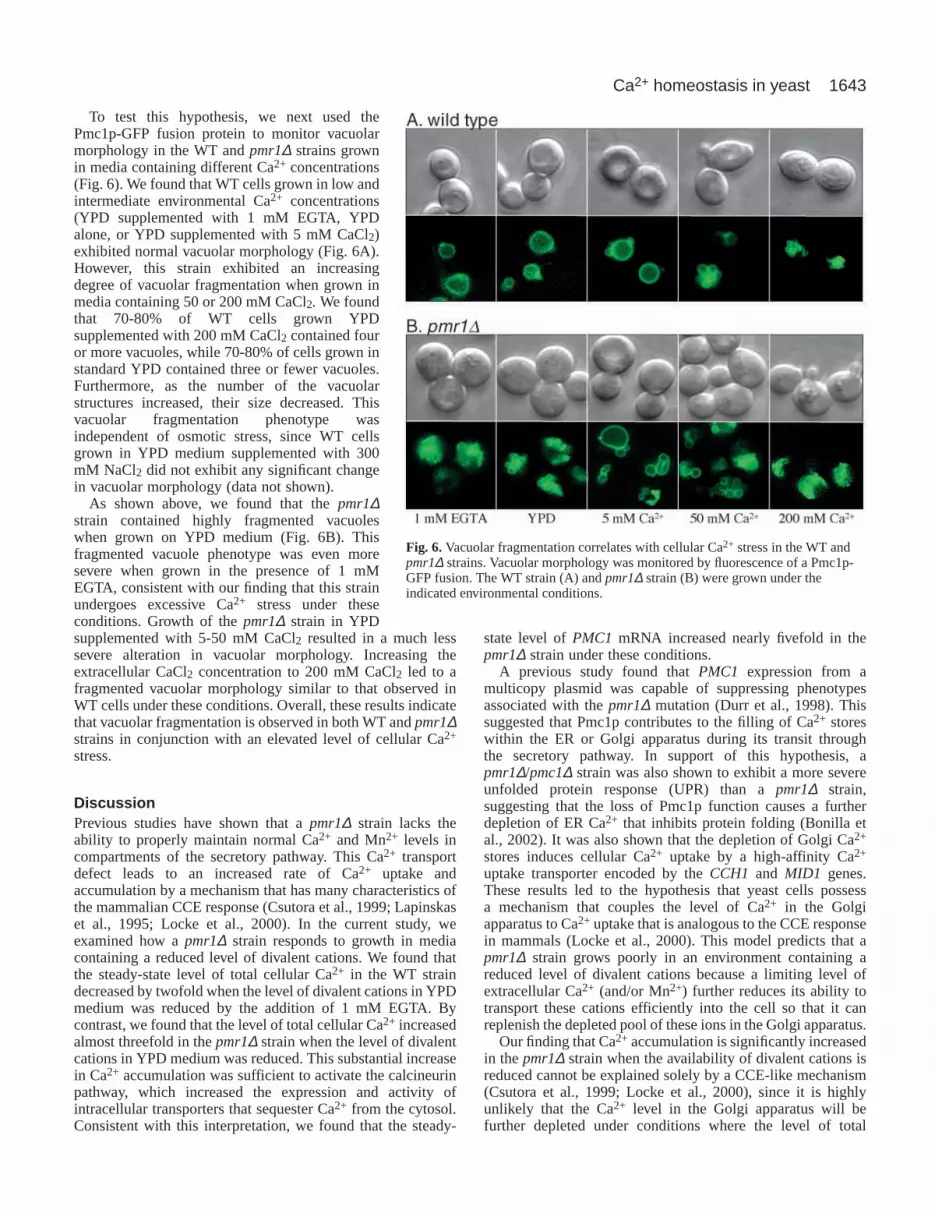
The pmr1∆/pmc1∆ strain exhibits vacuolarfragmentationPrevious studies have suggested that newlysynthesized Pmc1p contributes to themaintenance of Ca2+ levels within the secretorypathway during its transit to the vacuole in thepmr1∆ mutant (Bonilla et al., 2002; Durr etal., 1998; Locke et al., 2000) (this study).Subcellular fractionation of a pmr1∆ strain hasshown that Pmc1p is present not only in vacuolarfractions, but also overlaps fractions containingGolgi markers (Marchi et al., 1999). Theseresults suggest that a significant amount ofPmc1p may be retained in the Golgi apparatus
in the pmr1∆ mutant. To determine whether the Golgilocalization of Pmc1p could be visualized directly in yeastcells, we constructed a Pmc1p-GFP fusion protein. Followingthe integration of this construct into the nuclear genome ofpmc1∆ and pmr1∆/pmc1∆ strains, we found that the Pmc1p-GFP fusion restored normal Pmc1p function, as indicated byits ability to complement both the Ca2+ sensitivity of thepmc1∆ mutant and the EGTA sensitivity of the pmr1∆/pmc1∆mutant (data not shown). Thus, these complemented pmc1∆,and pmr1∆/pmc1∆ strains were functionally equivalent to WTand pmr1∆ strains, respectively.
We first asked whether the majority of the Pmc1p-GFPfusion protein co-localized with vacuoles as indicated by FM4-64, a vital stain that accumulates in the yeast vacuolarmembrane (Vida and Emr, 1995). As shown in Fig. 5, weobserved complete co-localization of these two markers in theexpected vacuolar pattern in WT cells grown in YPD medium.Whereas FM 4-64 and Pmc1p-GFP fluorescence also co-localized completely in the pmr1∆ strain, we found that bothmarkers produced an identical pattern of highly fragmentedstaining. FM 4-64 staining was also used to examine thevacuolar morphology in the pmc1∆ and pmr1∆/pmc1∆ strains.The vacuolar morphology was normal in the pmc1∆ strain.By contrast, the pmr1∆/pmc1∆ strain exhibited a highlyfragmented, frequently tubular, pattern of fluorescence. NoPmc1p-GFP fluorescence could be detected that was distinctfrom the FM 4-64 staining in either the WT or pmr1∆ strain,indicating that any accumulation in the Golgi apparatus (oranother subcellular location) was below the resolution of thisassay. However, these results demonstrate that significantvacuolar fragmentation occurs in strains containing the pmr1∆mutation when grown in standard YPD medium. On the basisof our finding that the pmr1∆ and pmr1∆/pmc1∆ strains bothundergo Ca2+ stress under these conditions, these resultssuggest that Ca2+ stress might induce vacuolar fragmentation.
Journal of Cell Science 116 (8)
Fig. 5.Vacuoles are fragmented in the pmr1∆ and pmr1∆/pmc1∆ strains when grownin YPD medium. Vacuolar morphology was monitored using the vital fluorescent dyeFM 4-64 or the Pmc1p-GFP fusion protein as indicated.
1643Ca2+ homeostasis in yeast
To test this hypothesis, we next used thePmc1p-GFP fusion protein to monitor vacuolarmorphology in the WT and pmr1∆ strains grownin media containing different Ca2+ concentrations(Fig. 6). We found that WT cells grown in low andintermediate environmental Ca2+ concentrations(YPD supplemented with 1 mM EGTA, YPDalone, or YPD supplemented with 5 mM CaCl2)exhibited normal vacuolar morphology (Fig. 6A).However, this strain exhibited an increasingdegree of vacuolar fragmentation when grown inmedia containing 50 or 200 mM CaCl2. We foundthat 70-80% of WT cells grown YPDsupplemented with 200 mM CaCl2 contained fouror more vacuoles, while 70-80% of cells grown instandard YPD contained three or fewer vacuoles.Furthermore, as the number of the vacuolarstructures increased, their size decreased. Thisvacuolar fragmentation phenotype wasindependent of osmotic stress, since WT cellsgrown in YPD medium supplemented with 300mM NaCl2 did not exhibit any significant changein vacuolar morphology (data not shown).
As shown above, we found that the pmr1∆strain contained highly fragmented vacuoleswhen grown on YPD medium (Fig. 6B). Thisfragmented vacuole phenotype was even moresevere when grown in the presence of 1 mMEGTA, consistent with our finding that this strainundergoes excessive Ca2+ stress under theseconditions. Growth of the pmr1∆ strain in YPDsupplemented with 5-50 mM CaCl2 resulted in a much lesssevere alteration in vacuolar morphology. Increasing theextracellular CaCl2 concentration to 200 mM CaCl2 led to afragmented vacuolar morphology similar to that observed inWT cells under these conditions. Overall, these results indicatethat vacuolar fragmentation is observed in both WT and pmr1∆strains in conjunction with an elevated level of cellular Ca2+
stress.
DiscussionPrevious studies have shown that a pmr1∆ strain lacks theability to properly maintain normal Ca2+ and Mn2+ levels incompartments of the secretory pathway. This Ca2+ transportdefect leads to an increased rate of Ca2+ uptake andaccumulation by a mechanism that has many characteristics ofthe mammalian CCE response (Csutora et al., 1999; Lapinskaset al., 1995; Locke et al., 2000). In the current study, weexamined how a pmr1∆ strain responds to growth in mediacontaining a reduced level of divalent cations. We found thatthe steady-state level of total cellular Ca2+ in the WT straindecreased by twofold when the level of divalent cations in YPDmedium was reduced by the addition of 1 mM EGTA. Bycontrast, we found that the level of total cellular Ca2+ increasedalmost threefold in the pmr1∆ strain when the level of divalentcations in YPD medium was reduced. This substantial increasein Ca2+ accumulation was sufficient to activate the calcineurinpathway, which increased the expression and activity ofintracellular transporters that sequester Ca2+ from the cytosol.Consistent with this interpretation, we found that the steady-
state level of PMC1 mRNA increased nearly fivefold in thepmr1∆ strain under these conditions.
A previous study found that PMC1 expression from amulticopy plasmid was capable of suppressing phenotypesassociated with the pmr1∆ mutation (Durr et al., 1998). Thissuggested that Pmc1p contributes to the filling of Ca2+ storeswithin the ER or Golgi apparatus during its transit throughthe secretory pathway. In support of this hypothesis, apmr1∆/pmc1∆ strain was also shown to exhibit a more severeunfolded protein response (UPR) than a pmr1∆ strain,suggesting that the loss of Pmc1p function causes a furtherdepletion of ER Ca2+ that inhibits protein folding (Bonilla etal., 2002). It was also shown that the depletion of Golgi Ca2+
stores induces cellular Ca2+ uptake by a high-affinity Ca2+
uptake transporter encoded by the CCH1 and MID1 genes.These results led to the hypothesis that yeast cells possessa mechanism that couples the level of Ca2+ in the Golgiapparatus to Ca2+ uptake that is analogous to the CCE responsein mammals (Locke et al., 2000). This model predicts that apmr1∆ strain grows poorly in an environment containing areduced level of divalent cations because a limiting level ofextracellular Ca2+ (and/or Mn2+) further reduces its ability totransport these cations efficiently into the cell so that it canreplenish the depleted pool of these ions in the Golgi apparatus.
Our finding that Ca2+ accumulation is significantly increasedin the pmr1∆ strain when the availability of divalent cations isreduced cannot be explained solely by a CCE-like mechanism(Csutora et al., 1999; Locke et al., 2000), since it is highlyunlikely that the Ca2+ level in the Golgi apparatus will befurther depleted under conditions where the level of total
Fig. 6.Vacuolar fragmentation correlates with cellular Ca2+ stress in the WT andpmr1∆ strains. Vacuolar morphology was monitored by fluorescence of a Pmc1p-GFP fusion. The WT strain (A) and pmr1∆ strain (B) were grown under theindicated environmental conditions.

1644
cellular Ca2+ is threefold higher. We also found that the pmr1∆strain exhibited less vacuolar fragmentation in the presenceof 5 mM or 50 mM CaCl2 than either higher or lowerconcentrations (see Fig. 6), suggesting that a moderate increasein extracellular Ca2+ can reduce the rate of cellular uptake (andthe resulting level of Ca2+ stress). When taken together, theseresults suggest that a mechanism exists that can couple the rateof cellular Ca2+ uptake to the extracellular Ca2+ concentration.The results of a previous study also support the existence of anextracellular Ca2+-sensing mechanism. We have shown that theloss of the major isoform of phosphoglucomutase (encoded bythe PGM2 gene) causes a large increase in Ca2+ uptake andaccumulation when a pgm2∆ strain is grown in YP mediumcontaining galactose as carbon source (Fu et al., 2000).Furthermore, we found that a pgm2∆/pmr1∆ strain is unable togrow in YP galactose medium, presumably because thecombination of these mutations leads to excessive Ca2+ uptakeand accumulation that results in an inhibition of cell growth.However, we found that growth of the pgm2∆/pmr1∆ straincould be restored on YP galactose medium when 100 mMCaCl2 was added to the growth medium (Fu et al., 2000). Inlight of our current results, we propose that this increase in theconcentration of CaCl2 in the growth medium can restoregrowth of the pgm2∆/pmr1∆ strain by reducing Ca2+ uptakeand accumulation through the action of an extracellular Ca2+-sensing mechanism.
While several mechanisms could be used to monitor thelevel of extracellular Ca2+, the most straightforward methodwould utilize a Ca2+ sensor on the cell surface (Fig. 7). Thiscell-surface Ca2+ sensor could be functionally equivalent to theextracellular Ca2+-sensing receptor of mammalian cells, whichcan respond to extremely small changes in the free Ca2+
concentration in the blood (Brown et al., 1995; Hebert et al.,1997). The results of the current study provide strong evidencethat an extracellular Ca2+-sensing mechanism can also play animportant role in coupling the level of environmental Ca2+ tocellular Ca2+ uptake, homeostasis and signaling in yeast. Toour knowledge, such a mechanism has not been proposed
previously for yeast cells. This might be due to the fact thatthis mechanism works in conjunction with other redundantprocesses that together tightly control cellular Ca2+
homeostasis. Such overlapping mechanisms could explain whyyeast mutants that maintain abnormally high levels of Ca2+
uptake and accumulation (such as the pmr1∆ and pgm2∆strains) were necessary to obtain evidence of this novel controlmechanism. These mutant strains have shed new light onthe mechanisms that couple cellular Ca2+ uptake andaccumulation. For example, the inability of the pmr1∆ strainto grow in the presence of a reduced level of divalent cationsallowed us to show that excessive Ca2+ accumulation isresponsible for this growth defect. This problem is furtherexacerbated in the pmr1∆/pmc1∆ strain in two ways. First, theloss of Pmc1p further aggravates the reduced ability to properlyfill the ER store depletion caused by the pmr1∆ mutation.Second, the pmc1∆ mutation diminishes the ability of the cellto sequester excess Ca2+ in the vacuole. As a result, thecytosolic Ca2+ load becomes more severe in the pmr1∆/pmc1∆strain, whereas its ability to sequester excess Ca2+ adequatelyinto the vacuole is decreased. Together, these consequencesresult in an inability to grow in a Ca2+-depleted environment.
In order to understand better the functional interplaybetween Ca2+ transporters in yeast, we also examined the roleof Vcx1p in maintaining cytosolic Ca2+ homeostasis in thepmr1∆/pmc1∆ strain. We found that Vcx1p plays an importantrole in Ca2+ homeostasis over a much broader range ofextracellular Ca2+ concentrations than previously appreciated(Pozos et al., 1996). Genetic studies have suggested that Vcx1pmight play a less important role in Ca2+ homeostasis than theCa2+-ATPases Pmc1p and Pmr1p. Consistent with thisconclusion, we have observed that a pmr1∆/vcx1∆ strain is nomore sensitive to chelating agents than a pmr1∆ strain. Thisresult indicated that Pmc1p alone is sufficient to sequester thehigh cellular Ca2+ that accumulates under these conditions, andsuggested that Vcx1p plays only a secondary role in Ca2+
homeostasis (R. Kellermayer and D. Bedwell, unpublished).However, in the current study, we show that a pmr1∆/pmc1∆mutant can successfully cope with a considerable level of Ca2+
stress, whereas cellular Ca2+ homeostasis is completelydisrupted when its vacuolar proton gradient is disrupted bybafilomycin A1. Since it has been shown that vacuolar vesiclesderived from a strain lacking Vcx1p do not possess anyCa2+/H+ exchange activity (Pozos et al., 1996), these findingsstrongly indicate that Vcx1p plays the predominant role inmaintaining Ca2+ homeostasis in the pmr1∆/pmc1∆ mutant. Inaddition, the previous observation that the combination of thepmr1∆, pmc1∆ and vcx1∆ mutations together causes syntheticlethality reinforces the importance of Vcx1p in Ca2+ regulationin the absence of these two Ca2+-ATPases (Cunningham andFink, 1996; Miseta et al., 1999a). These results suggest that thePmr1p, Pmc1p and Vcx1p Ca2+ transporters are all capable ofindependently maintaining cellular Ca2+ homeostasis in manyyeast strains. Apparently, subtle differences in the extent ofVcx1p inhibition by calcineurin are responsible for conflictingreports regarding the viability of pmr1∆/pmc1∆ strains indifferent genetic backgrounds (Cunningham and Fink, 1996;Locke et al., 2000) (this report).
We also used a Pmc1p-GFP fusion protein to show thatvacuolar fragmentation coincides with Ca2+ stress. Our findingthat vacuolar fragmentation occurs under diverse conditions
Journal of Cell Science 116 (8)
Vacuole
Ca2+ Ca2+
ER/Golgi
Ca2+
CCESignal
StoreDepletion
Ca2+
Ca2+
Ca2+
Ca2+
Ca2+
ExtracellularSensing
Ca2+
Ca2+
Ca2+
1
2
Fig. 7.Model showing how cellular Ca2+ uptake in yeast iscoordinately regulated by two distinct mechanisms. The firstmechanism is a CCE-like response that couples cellular Ca2+ uptaketo Ca2+ store depletion in the ER/Golgi apparatus. The secondmechanism couples cellular Ca2+ uptake to the level of Ca2+ in theextracellular environment.

1645Ca2+ homeostasis in yeast
that lead to cellular Ca2+ stress in both WT and mutant strainsstrongly suggests a direct relationship between these twoevents. The data presented does not allow us to determinewhether it is high Ca2+ in the cytosol or vacuole that leadsto this fragmentation phenomenon. However, in otherexperiments we found that a pmc1∆/vcx1∆ strain exhibitsfragmented vacuoles in media containing 50 mM CaCl2 in amanner similar to the WT strain (R. Kellermayer and D.Bedwell, unpublished). Since this mutant accumulates muchless Ca2+ in the vacuole than the WT strain (Cunningham andFink, 1996; Miseta et al., 1999a; Pozos et al., 1996), theseresults strongly suggest that an elevated cytosolic Ca2+ causesvacuolar fragmentation in S. cerevisiae.
A recent study examined a large collection of knockoutstrains from the SaccharomycesGenome Deletion Project fordefects in homotypic vacuolar fusion (Seeley et al., 2002).Surprisingly, 714 out of 4828 deletion strains examined(~15%) exhibited alterations in vacuole morphology. Afterexcluding a large number of genes thought to influencevacuole morphology by indirect means, 137 genes (~3%) werechosen as candidate VAM genes that were thought to play adirect role in vacuolar morphology. Among these were avariety of genes that encoded proteins previously relatedto homotypic vacuolar fusion, including fusion catalysts,enzymes of lipid metabolism, SNARES, GTPases and theireffectors, protein kinases, phosphatases, and cytoskeletalproteins. Another group of genes encoded proteins involvedin cation transport (including the PMR1 gene). Becausevacuolar fragmentation was associated with the deletion ofthese genes, it was reasoned that the loss of these geneproducts caused defects in vacuolar fusion, resulting in thevacuolar fragmentation phenotype. However, on the basis ofthe strong correlation between cellular Ca2+ stress andvacuolar fragmentation we observed in both WT and mutantyeast strains, we propose that an elevation of cytosolic Ca2+
might lead to vacuolar fragmentation as a regulatory responseto aid in Ca2+ sequestration in WT yeast, rather than simplybeing the result of a defect in vacuolar fusion associated withhigh cytosolic Ca2+. On a per unit volume basis, multiplesmall vacuoles provide a greater surface area than fewer,larger vacuoles. This increased surface/volume ratio could aidin accommodating the function of the increased number ofPmc1p transporters in the vacuolar membrane that are inducedby calcineurin activation, thus allowing vacuolar Ca2+
sequestration to proceed more efficiently.In both the current study and a prior study of genes that
influence homotypic vacuolar fusion (Seeley et al., 2002),it was shown that a pmr1∆ strain exhibits a vacuolarfragmentation phenotype. By contrast, another study found thata pmr1∆ mutation reversed the vacuolar fragmentationphenotype associated with oxidative stress in a sod1∆ strain(Corson et al., 1999). These results led to the conclusion thatoxidative stress associated with the sod1∆ mutation alteredcellular iron homeostasis, leading to oxidative damage thatsomehow led to vacuolar fragmentation. It was proposed thatthe pmr1∆ mutation suppressed this vacuolar fragmentationphenotype by raising the concentration of Mn2+ in the cytosol,which acted to scavenge free radicals and reduce oxidativestress. Thus, pmr1∆ mutations have been associated with boththe induction and reversal of vacuolar fragmentation. Thesemarkedly different effects suggest the existence of a complex
regulatory mechanism that allows the cytosolic levels ofdifferent divalent cations to influence vacuolar morphology indistinct ways. Further studies are needed to determine how thiscomplex physiological adaptation is carried out.
The authors thank Lianwu Fu for fruitful discussions, AlbertTousson for providing assistance in digital imaging, and KyleCunningham, Kelly Tatchell and Aaron Straight for generouslyproviding strains and other reagents. This work was supported byAmerican Heart Association (Southeast Affiliate) grant 0255121B (toD.M.B.) and Hungarian National Foundation grant OTKA-T038144(to A.M.).
ReferencesAbe, F. and Horikoshi, K. (1995). Hydrostatic pressure promotes the
acidification of vacuoles in Saccharomyces cerevisiae. FEMS Microbiol.Lett. 130, 307-312.
Antebi, A. and Fink, G. R. (1992). The yeast Ca2+-ATPase homologue,PMR1, is required for normal Golgi function and localizes in a novel Golgi-like distribution. Mol. Biol. Cell3, 633-654.
Batiza, A. F., Schulz, T. and Masson, P. H. (1996). Yeast respond tohypotonic shock with a calcium pulse. J. Biol. Chem.271, 23357-23362.
Bonetti, B., Fu, L., Moon, J. and Bedwell, D. M. (1995). The efficiency oftranslation termination is determined by a synergistic interplay betweenupstream and downstream sequences in Saccharomyces cerevisiae. J. Mol.Biol. 251, 334-345.
Bonilla, M., Nastase, K. K. and Cunningham, K. W. (2002). Essential roleof calcineurin in response to endoplasmic reticulum stress. EMBO J.21,2343-2353.
Brown, E. M., Vassilev, P. M. and Hebert, S. C. (1995). Calcium ions asextracellular messengers. Cell 83, 679-682.
Burke, D., Dawson, D. C. and Stearns, T. (2000). Methods in Yeast Genetics:a Cold Spring Harbor Laboratory Course Manual. Cold Spring Harbor, NY:Cold Spring Harbor Laboratory Press.
Corson, L. B., Folmer, J., Strain, J. J., Culotta, V. C. and Cleveland, D. W.(1999). Oxidative stress and iron are implicated in fragmenting vacuoles ofSaccharomyces cerevisiaelacking Cu, Zn-superoxide dismutase. J. Biol.Chem.274, 27590-27596.
Cronin, S. R., Khoury, A., Ferry, D. K. and Hampton, R. Y. (2000).Regulation of HMG-CoA reductase degradation requires the P-type ATPaseCod1p/Spf1p. J. Cell Biol.148, 915-924.
Cronin, S. R., Rao, R. and Hampton, R. Y. (2002). Cod1p/Spf1p is a P-typeATPase involved in ER function and Ca2+ homeostasis. J. Cell Biol.157,1017-1028.
Csutora, P., Su, Z., Kim, H. Y., Bugrim, A., Cunningham, K. W., Nuccitelli,R., Keizer, J. E., Hanley, M. R., Blalock, J. E. and Marchase, R. B.(1999). Calcium influx factor is synthesized by yeast and mammalian cellsdepleted of organellar calcium stores. Proc. Natl. Acad. Sci. USA96, 121-126.
Cunningham, K. W. and Fink, G. R. (1994a). Ca2+ transport inSaccharomyces cerevisiae. J. Exp. Biol.196, 157-166.
Cunningham, K. W. and Fink, G. R. (1994b). Calcineurin-dependent growthcontrol in Saccharomyces cerevisiaemutants lacking PMC1, a homolog ofplasma membrane Ca2+ ATPases. J. Cell Biol.124, 351-363.
Cunningham, K. W. and Fink, G. R. (1996). Calcineurin inhibits VCX1-dependent H+/Ca2+ exchange and induces Ca2+ ATPases in Saccharomycescerevisiae. Mol. Cell. Biol.16, 2226-2237.
Denis, V. and Cyert, M. S. (2002). Internal Ca2+ release in yeast is triggeredby hypertonic shock and mediated by a TRP channel homologue. J. CellBiol. 156, 29-34.
Dunn, T., Gable, K. and Beeler, T. (1994). Regulation of cellular Ca2+ byyeast vacuoles. J. Biol. Chem.269, 7273-7278.
Durr, G., Strayle, J., Plemper, R., Elbs, S., Klee, S. K., Catty, P., Wolf, D.H. and Rudolph, H. K. (1998). The medial-Golgi ion pump Pmr1 suppliesthe yeast secretory pathway with Ca2+ and Mn2+ required for glycosylation,sorting, and endoplasmic reticulum-associated protein degradation. Mol.Biol. Cell 9, 1149-1162.
Eilam, Y. and Othman, M. (1990). Activation of Ca2+ influx by metabolicsubstrates in Saccharomyces cerevisiae: role of membrane potential andcellular ATP levels. J. Gen. Microbiol.136, 861-866.
Eilam, Y., Lavi, H. and Grossowicz, N. (1985). Mechanism of stimulation of

1646
Ca2+ uptake by miconazole and ethidium bromide in yeasts: role of vacuolesin Ca2+ detoxification. Microbios44, 51-66.
Eilam, Y., Othman, M. and Halachmi, D. (1990). Transient increase in Ca2+
influx in Saccharomyces cerevisiaein response to glucose: effects ofintracellular acidification and cAMP levels. J. Gen. Microbiol.136, 2537-2543.
Emr, S. D., Vassarotti, A., Garrett, J., Geller, B. L., Takeda, M. andDouglas, M. G. (1986). The amino terminus of the yeast F1-ATPase beta-subunit precursor functions as a mitochondrial import signal. J. Cell Biol.102, 523-533.
Forster, C. and Kane, P. M. (2000). Cytosolic Ca2+ homeostasis is aconstitutive function of the V-ATPase in Saccharomyces cerevisiae. J. Biol.Chem.275, 38245-38253.
Fu, L., Miseta, A., Hunton, D., Marchase, R. B. and Bedwell, D. M. (2000).Loss of the major isoform of phosphoglucomutase results in altered calciumhomeostasis in Saccharomyces cerevisiae. J. Biol. Chem.275, 5431-5440.
Halachmi, D. and Eilam, Y. (1996). Elevated cytosolic free Ca2+
concentrations and massive Ca2+ accumulation within vacuoles, in yeastmutant lacking PMR1, a homolog of Ca2+-ATPase. FEBS Lett.392, 194-200.
Hartley, A. D., Bogaerts, S. and Garrett, S. (1996). cAMP inhibits budgrowth in a yeast strain compromised for Ca2+ influx into the Golgi. Mol.Gen. Genet. 251, 556-564.
Hebert, S. C., Brown, E. M. and Harris, H. W. (1997). Role of the Ca2+-sensing receptor in divalent mineral ion homeostasis. J. Exp. Biol. 200, 295-302.
Iida, H., Sakaguchi, S., Yagawa, Y. and Anraku, Y. (1990a). Cell cyclecontrol by Ca2+ in Saccharomyces cerevisiae. J. Biol. Chem.265, 21216-21222.
Iida, H., Yagawa, Y. and Anraku, Y. (1990b). Essential role for induced Ca2+
influx followed by [Ca2+]i rise in maintaining viability of yeast cells late inthe mating pheromone response pathway. A study of [Ca2+]i in singleSaccharomyces cerevisiaecells with imaging of fura-2. J. Biol. Chem.265,13391-13399.
Kaibuchi, K., Miyajima, A., Arai, K. and Matsumoto, K. (1986). Possibleinvolvement of RAS-encoded proteins in glucose-inducedinositolphospholipid turnover in Saccharomyces cerevisiae. Proc. Natl.Acad. Sci. USA83, 8172-8176.
Lapinskas, P. J., Cunningham, K. W., Liu, X. F., Fink, G. R. and Culotta,V. C. (1995). Mutations in PMR1suppress oxidative damage in yeast cellslacking superoxide dismutase. Mol. Cell. Biol.15, 1382-1388.
Locke, E. G., Bonilla, M., Liang, L., Takita, Y. and Cunningham, K. W.(2000). A homolog of voltage-gated Ca2+ channels stimulated by depletionof secretory Ca2+ in yeast. Mol. Cell. Biol.20, 6686-6694.
Marchi, V., Sorin, A., Wei, Y. and Rao, R. (1999). Induction of vacuolarCa2+-ATPase and H+/Ca2+ exchange activity in yeast mutants lacking Pmr1,the Golgi Ca2+-ATPase. FEBS Lett.454, 181-186.
Matheos, D. P., Kingsbury, T. J., Ahsan, U. S. and Cunningham, K. W.(1997). Tcn1p/Crz1p, a calcineurin-dependent transcription factor thatdifferentially regulates gene expression in Saccharomyces cerevisiae. GenesDev.11, 3445-3458.
Miller, J. H. (1992). A Short Course in Bacterial Genetics. Cold SpringHarbor, NY: Cold Spring Harbor Laboratory Press.
Miseta, A., Fu, L., Kellermayer, R., Buckley, J. and Bedwell, D. M. (1999a).The Golgi apparatus plays a significant role in the maintenance of Ca2+
homeostasis in the vps33∆ vacuolar biogenesis mutant of Saccharomycescerevisiae. J. Biol. Chem.274, 5939-5947.
Miseta, A., Kellermayer, R., Aiello, D. P., Fu, L. and Bedwell, D. M.(1999b). The vacuolar Ca2+/H+ exchanger Vcx1p/Hum1p tightly controlscytosolic Ca2+ levels in S. cerevisiae. FEBS Lett.451, 132-136.
Mori, I. C., Iida, H., Tsuji, F. I., Isobe, M., Uozumi, N. and Muto, S. (1998).Salicylic acid induces a cytosolic Ca2+ elevation in yeast. Biosci. Biotechnol.Biochem.62, 986-989.
Nakajima-Shimada, J., Iida, H., Tsuji, F. I. and Anraku, Y. (1991).Monitoring of intracellular calcium in Saccharomyces cerevisiaewith anapoaequorin cDNA expression system. Proc. Natl. Acad. Sci. USA88, 6878-6882.
Pozos, T. C., Sekler, I. and Cyert, M. S. (1996). The product of HUM1, anovel yeast gene, is required for vacuolar Ca2+/H+ exchange and is relatedto mammalian Na+/Ca2+ exchangers. Mol. Cell. Biol.16, 3730-3741.
Putney, J. W., Jr (1992). Inositol phosphates and calcium entry. Adv. SecondMessenger Phosphoprotein Res.26, 143-160.
Rudolph, H. K., Antebi, A., Fink, G. R., Buckley, C. M., Dorman, T. E.,LeVitre, J., Davidow, L. S., Mao, J. I. and Moir, D. T. (1989). The yeastsecretory pathway is perturbed by mutations in PMR1, a member of a Ca2+
ATPase family. Cell 58, 133-145.Sambrook, J., Fritsch, E. F. and Maniatis, T. (1989). Molecular Cloning: a
Laboratory Manual. Cold Spring Harbor, N.Y.: Cold Spring HarborLaboratory Press.
Seeley, E. S., Kato, M., Margolis, N., Wickner, W. and Eitzen, G. (2002).Genomic analysis of homotypic vacuole fusion. Mol. Biol. Cell13, 782-794.
Sorin, A., Rosas, G. and Rao, R. (1997). PMR1, a Ca2+-ATPase in yeastGolgi, has properties distinct from sarco/endoplasmic reticulum and plasmamembrane calcium pumps. J. Biol. Chem.272, 9895-9901.
Stathopoulos, A. M. and Cyert, M. S. (1997). Calcineurin acts through theCRZ1/TCN1-encoded transcription factor to regulate gene expression inyeast. Genes Dev.11, 3432-3444.
Stathopoulos-Gerontides, A., Guo, J. J. and Cyert, M. S. (1999). Yeastcalcineurin regulates nuclear localization of the Crz1p transcription factorthrough dephosphorylation. Genes Dev.13, 798-803.
Strayle, J., Pozzan, T. and Rudolph, H. K. (1999). Steady-state freeCa2+ in the yeast endoplasmic reticulum reaches only 10 µM and ismainly controlled by the secretory pathway pump pmr1. EMBO J. 18,4733-4743.
Suzuki, C. and Shimma, Y. I. (1999). P-type ATPase spf1mutants show anovel resistance mechanism for the killer toxin SMKT. Mol. Microbiol. 32,813-823.
Tanida, I., Hasegawa, A., Iida, H., Ohya, Y. and Anraku, Y. (1995).Cooperation of calcineurin and vacuolar H+-ATPase in intracellular Ca2+
homeostasis of yeast cells. J. Biol. Chem.270, 10113-10119.Tokes-Fuzesi, M., Bedwell, D. M., Repa, I., Sipos, K., Sumegi, B., Rab, A.
and Miseta, A. (2002). Hexose phosphorylation and the putative calciumchannel component Mid1p are required for the hexose-induced transientelevation of cytosolic calcium response in Saccharomyces cerevisiae. Mol.Microbiol. 44, 1299-1308.
Vida, T. A. and Emr, S. D. (1995). A new vital stain for visualizing vacuolarmembrane dynamics and endocytosis in yeast. J. Cell Biol.128, 779-792.
Withee, J. L., Mulholland, J., Jeng, R. and Cyert, M. S. (1997). An essentialrole of the yeast pheromone-induced Ca2+ signal is to activate calcineurin.Mol. Biol. Cell8, 263-277.
Yoshimoto, H., Saltsman, K., Gasch, A. P., Li, H. X., Ogawa, N., Botstein,D., Brown, P. O. and Cyert, M. S. (2002). Genome-wide analysis of geneexpression regulated by the calcineurin/Crz1p signaling pathway inSaccharomyces cerevisiae. J. Biol. Chem.10, 31079-31088.
Journal of Cell Science 116 (8)