exploiting the brachypodium tool box in cereal and grass research
TRANSCRIPT

Tansley review
Exploiting the Brachypodium Tool Boxin cereal and grass research
Author for correspondence:Luis A. J. Mur
Tel: +44 (0)1970 622981Email: [email protected]
Received: 4 March 2011
Accepted: 25 March 2011
Luis A. J. Mur1, Joel Allainguillaume1, Pilar Catalan2, Robert Hasterok3,
Glyn Jenkins1, Karolina Lesniewska3, Ianto Thomas1 and John Vogel4
1Institute of Biological, Environmental and Rural Sciences, Aberystwyth, Wales SY23 3DA, UK;
2Department of Agriculture, University of Zaragoza, High Polytechnic School of Huesca, Ctra.
Cuarte km 1, ES–22071 Huesca, Spain; 3Department of Plant Anatomy and Cytology, Faculty of
Biology and Environmental Protection, University of Silesia, PL–40-032 Katowice, Poland; 4USDA
ARS Western Regional Research Center, Albany, CA 94710 USA
New Phytologist (2011) 191: 334–347doi: 10.1111/j.1469-8137.2011.03748.x
Key words: Brachypodium, comparativegenomics, functional genomics, genomesequencing, model grass.
Summary
It is now a decade since Brachypodium distachyon (Brachypodium) was suggested
as a model species for temperate grasses and cereals. Since then transformation
protocols, large expressed sequence tag (EST) databases, tools for forward and
reverse genetic screens, highly refined cytogenetic probes, germplasm collections
and, recently, a complete genome sequence have been generated. In this review,
we will describe the current status of the Brachypodium Tool Box and how it is
beginning to be applied to study a range of biological traits. Further, as genomic
analysis of larger cereals and forage grasses genomes are becoming easier, we will
re-evaluate Brachypodium as a model species. We suggest that there remains an
urgent need to employ reverse genetic and functional genomic approaches to
identify the functionality of key genetic elements, which could be employed subse-
quently in plant breeding programmes; and a requirement for a Pooideae
reference genome to aid assembling large pooid genomes. Brachypodium is an
ideal system for functional genomic studies, because of its easy growth require-
ments, small physical stature, and rapid life cycle, coupled with the resources
offered by the Brachypodium Tool Box.
Contents
Summary 334
I. Introduction 335
II. A challenge for the Brachypodium Tool Box: thelegacy of cereal domestication
335
III. Opening the Brachypodium Tool Box: what isBrachypodium?
336
IV. The Brachypodium Tool Box: where are we now? 337
V. Targets for the Brachypodium Tool Box: key traits 342
VI. Whence for the Brachypodium Tool box?Primus inter pares?
344
Acknowledgements 345
References 345
NewPhytologistReview
334 New Phytologist (2011) 191: 334–347
www.newphytologist.com� 2011 The Authors
New Phytologist � 2011 New Phytologist Trust

I. Introduction
The publication of the entire genome sequence forBrachypodium distachyon (which we will subsequentlyreferred to Brachypodium, except when we describe intra-generic variation) accession Bd21 early in 2010 (IBI, 2010)is a fitting end to a decade that started with the seriousproposal of Brachypodium as a model grass for temperatecereals and forage grasses (Draper et al., 2001). Equallyimportantly, the same decade has seen the development ofnext generation sequencing (NGS) technologies offering thecapability of generating unprecedented amounts of genomicsequence information (Metzker, 2010). When coupled withimproved bioinformatic approaches allowing genomes to bemore readily assembled (Cantacessi et al., 2010), the conse-quence is that large genome cereal species are now tractablefor studies that would previously have been the preserve ofmodel species. Even wheat is now within the sights ofgenome sequencers and thoughts are now focusing on howto exploit these sequence data in crop breeding programmes(Edwards & Batley, 2010). In the wake of these major tech-nological developments, we will reconsider in this reviewthe value of a Brachypodium model to cereal biology and tocrop breeding programmes. In doing so, it is not the inten-tion of the authors to describe the details of theBrachypodium genomic sequence (IBI, 2010).
II. A challenge for the Brachypodium Tool Box:the legacy of cereal domestication
Temperate cereals were domesticated by humanity fromwild grassland species in a multi-episodic process between10 000 and 3000 yr ago (Brown et al., 2009). However,domestication resulted in population bottlenecks – a loss ofgenetic diversity compared with wild ancestors – amongcultivated cereal varieties, with perhaps only 10–20% of thewild variation being used in modern wheat varieties(Langridge et al., 2006). This lack of variation thwartsattempts to improve elite germplasm, such as increasing itsyield (e.g. possibly by 71% between 1997 and 2050;Rosegrant & Cline, 2003) during a period of potential envi-ronmental change and population increase.
In order to meet this challenge, a number of approachesare available to cereal scientists. Agricultural practices, par-ticularly in the developing world, could be improved toincrease the areas under cultivation and maximize produc-tion but this should be also coupled with significantimprovements in the cereal gene pools (Rosegrant & Cline,2003). The latter could come about via screens of relativelyrestricted collections of elite germplasm or through exploi-tation of cereal landraces – semidomesticated variants ofcrop lines that have been locally maintained through tradi-tional farming and often are excellent sources of stresstolerance (Newton et al., 2010). More innovative processes
may involve the rehybridization of cereal ancestors to formnew ‘synthetic’ progeny. However, ‘linkage drag’ – the asso-ciation of a desired genetic trait with nonoptimal traits – isa problem, as to avoid ‘linkage drag’ a genetic modification(GM) strategy can be adopted where the focus is squarelyupon the transfer of well-defined genetic element(s).However, at least within a European context, this strategyhas met with considerable opposition (Schenkelaars, 2001;Hartl & Herrmann, 2009).
Against such a background, what are the roles for a modelspecies such as Brachypodium, which has no agriculturalvalue and is not easily introgressed into other cereals (Khan& Stace, 1999)? It is our contention that Brachypodium-based research will play a vital role on two fronts. First, theBrachypodium genome will act as a reference for the assem-bly of large temperate cereal genomes. Classically, plantgenomes are currently sequenced either by a global shotgunsequencing approach or by sequencing of large-insertoverlapping genomic clones (Venter et al., 1996). Theprevalence of repeated sequences in the genomes of plants,especially the cereals, makes the assembly of a completedgenome very difficult. This problem is aggravated by theplasticity of repeated sequences within the intragenicregions, which is greater than that observed in primates(Dubcovsky & Dvorak, 2007). This results in intragenicregions with large numbers of very similar nucleotidesequences, which has effectively prevented the use of theshotgun genome sequencing approach for diploid Triticeaespecies (Luo et al., 2010). Newer approaches such as high-information-content-fingerprinting (HICF) of bacterialartificial chromosome (BAC) clones (Luo et al., 2003) areundoubtedly aiding the assembly of large, complexgenomes. However, comparative genomic strategies using asmaller low-repetitive DNA reference genome, such asBrachypodium, still has value. An incidental, albeit notinsignificant, additional benefit will be that Brachypodiumwill contribute towards the supply of markers for mappingdifficult regions of large genome cereals. The latter role, theprovision of markers to fine map traits in (for example) het-erochromatic areas of crop plants, has already beendemonstrated (Turner et al., 2005; Griffiths et al., 2006;Spielmeyer et al., 2008). Second, its role as a functionalgenomic model to rapidly determine gene function is likelyto be much more valuable. In this context, the innate bio-logical traits of Brachypodium make it a useful model forall the grasses. Platform infrastructure (The BrachypodiumTool Box: Fig. 1b–c) is being developed with which func-tion can be assigned to hitherto intractable genes. Thus,within the context of breeding programmes, Brachypodiumcould be seen as a tool by which gene function, could berapidly assigned by alignment with other genomes, and bywhich tightly-linked markers to useful alleles could bedeveloped. Further, in those areas of the world where GMapproaches are widely accepted by the general public,
NewPhytologist Tansley review Review 335
� 2011 The Authors
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011) 191: 334–347
www.newphytologist.com

Brachypodium genes, or their cereal orthologues, could beused to generate transgenic crops.
III. Opening the Brachypodium Tool Box: whatis Brachypodium?
Brachypodium distachyon, commonly called purple falsebrome, is native to the countries around the Mediterraneanbasin, the Middle East, south-west Asia and north-east Africa(Schippmann, 1991). It has been widely naturalized beyondits natural range with a notable recent colonization in North(Mexico, USA) and South (Argentina, Ecuador, Peru, Uruguay)America, Australia and South Africa (Schippmann, 1991;Catalan 2003) and even the UK (England and Scotland)(Stace, 2010). Although in the wake of the completedB. distachyon genome sequence (IBI, 2010), this species hasreceived much of the attention, the Brachypodium genusencompasses between 15 and 18 species distributed world-wide (Catalan et al., 1997; Catalan & Olmstead, 2000). Thevarious Brachypodium species have different geographicalranges. B. pinnatum and B. sylvaticum are wide-ranging andfound throughout Eurasia, and B. retusum across theMediterranean, while B. rupestre is predominantly found inwestern Europe, and B. phoenicoides in the westernMediterranean. Other Brachypodieae are more geographi-cally restricted, B. arbuscula being endemic to the CanaryIslands, B. boissieri to southern Spain and B. kawakamii toTaiwan while B. mexicanum is found in Central and SouthAmerica, B. flexum in eastern and western tropical Africa and
South Africa, and B. bolusii in South Africa (Schippmann,1991; Catalan & Olmstead, 2000). The Brachypodieae havedifferent chromosome base numbers (x = 5, 7, 8 and 9).Brachypodium distachyon has a chromosome base number ofx = 5 and has been described as having different ploidies(2n = 10, 20 and 30; Robertson, 1981). As a rule, the2n = 10 cytotypes tend to be much smaller and many acces-sions require vernalization to induce flowering. By contrast,B. distachyon 2n = 20, 30 tend to be taller, have a lesserrequirement for vernalization and almost invariably (at leastfrom our observations) exhibit prominent anthesis and largerseeds (Schwartz et al., 2010).
Much early work initiated by Professor C. A. Stace(Leicester University, UK), focused on establishing thephylogenetic relationships between the Brachypodieae(Fig. 1d). Perhaps unsurprisingly, the long-rhizomatousperennial species (B. arbuscula, B. retusum, B. rupestre,B. phoenicoides, B. pinnatum and B. sylvaticum) are the mostclosely related, being distinct from both the geographicallyisolated short-rhizomatous B. mexicanum and the widespreadannual B. distachyon (Catalan et al. 1995; Catalan &Olmstead, 2000). Furthermore, the Brachypodieae have beenshown to be sister to the recently evolved core pooid’ grasses,which include the temperate cereals and forages of tribesTriticeae–Bromeae and Aveneae–Poeae, in all the molecularphylogenetic studies conducted so far. Based on sequence vari-ation (mean synonymous substitution rates, Ks) betweenorthologous genes from B. distachyon, sorghum (Sorghumbicolor), rice (Oryza sativa) and wheat (Triticum aestivum)genomes, it was estimated that B. distachyon diverged from acommon wheat ancestor 32–39 million yr ago (Mya), fromthe common rice ancestor 40–53 Mya and from that com-mon ancestor to sorghum 45–60 Mya (IBI, 2010).
Until recently, it was assumed that B. distachyon had anautopolyploid series with diploid (2n = 10), tetraploid(2n = 20) and hexaploid (2n = 30) chromosomal races(Robertson, 1981). However, cytogenetic studies usingfluorescence in situ hybridization (FISH) with total nuclearDNA (genomic in situ hybridization; GISH) and ribosomalprobes indicated that the 30 chromosome accessions ofB. distachyon were in fact allotetraploids (Hasterok et al.,2004). Subsequently, FISH analysis with BAC-based probesfrom a genomic library of B. distachyon (2n = 10), coupledwith genome size analysis, has further confirmed that diploidB. distachyon (2n = 10; 0.631 pg ⁄ 2C) genotypes may havehybridized with another diploid (2n = 20; 0.564 pg ⁄ 2C) (ofwhich only a single accession – ABR114 – currently exists) toform the allotetraploid (2n = 30; 1.265 pg ⁄ 2C) (Hasteroket al., 2006; E. Wolny pers. comm.). Given these observa-tions and clear morpho-anatomical differences between thecytotypes, it is entirely appropriate to consider each B.distachyon chromosome class as different species. Some of theauthors of this review will be formally presenting these newBrachypodium species in the near future. Forthwith, we will
• Small stature• Rapid seed-to-seed
Biologicalinfrastructure
cycle• Undemanding growth
conditions• Growing availability of
germplasm• Important traits
(b)(a)
Genomicinfrastructure
• Genome sequence• EST libraries
•• Genomic markers for forward genetic screens• Syntenic relationships
with major cereals• T-DNA tagged
populations• EMS/fast neutron-mutated populations
• Cytogenetic markers• Oligo-microarrays
• Bioinformatic resources
(c)Brachypodium sylvaticum
Brachypodium pinnatum
Brachypodium phoenicoidesBrachypodium rupestre
(d)
Triticeae
Brachypodium retusum
Brachypodium arbuscula
Brachypodium mexicanumBrachypodium distachyon
PoaceaeOryza
Fig. 1 Components of the Brachypodium Tool Box for genomic andpost genomic analyses. (a) Brachypodium distachyon growing in thewild (Castillo de Mur, Lleida, Catalunya, Spain:N42.09763 ⁄ E0.87750); bar, 1 cm. The Brachypodium toolboxcontains considerable (b) biological and (c) genomic resources. (d)Phylogenetic relationships within the Brachypodium genus and withthe Triticeae, Poaceae and Ehrhartoideae (adapted from Catalan &Olmstead, 2000).
336 Review Tansley reviewNewPhytologist
� 2011 The Authors
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011) 191: 334–347
www.newphytologist.com

refer to B. distachyon as the diploid 2n = 10 form. TheWolny & Hasterok (2009) study has been extended to exam-ine other species within the genus and has provided someinsights into the organization, phylogeny and evolution ofBrachypodium genomes at the chromosomal level. Genomicin situ hybridization studies revealed inter alia that B. retusum(2n = 38) is likely to be an allopolyploid comprising thegenome of B. distachyon or a close relative. Similarly, twoother species with 2n = 28, B. pinnatum and B. phoenicoides,are in fact allotetraploids, each containing the B. distachyongenome and either B. pinnatum (2n = 18) or B. sylvaticum(2n = 18), respectively. This complex evolution of theBrachypodium genus should be seen as an opportunity tostudy the factors governing genome polyploidization usingpowerful functional genomic tools that have been added tothe Brachypodium Tool Box. Many important cereal speciesare also allopolyploids and elements such the Ph1 locus,which diploidizes allohexaploid wheat, are of interest. Giventhis, the genus Brachypodium represents an ideal system withwhich to study the value of synthetic hybrids which exploitgenomic interactions to deliver beneficial traits not necessar-ily present in the parental diploid or diploid-like lines.
IV. The Brachypodium Tool Box: where are wenow?
Given the relatively short period since it was first suggestedas a model species, the genomic resources comprising theBrachypodium Tool Box have been very rapidly established.Most recently, a very high quality draft genome sequencewas published (IBI, 2010). The final genome assembly wasremarkably complete, with 99.6% of all the sequencesincorporated into the final assembly and only 0.4% of thesequence predicted to be missing based on paired end infor-mation. Similarly, the initial annotation was of high qualityand was facilitated by a large number of ESTs (Vogel et al.,2006). Thus, the genome sequence is now in place to serveas a foundation for a myriad of applications. For example, acombination expression and tiling Affymetrix microarrayhas been created for Brachypodium (T. Mockler, pers.comm.). Other genomic resources include BAC libraries(Hasterok et al., 2006; Huo et al., 2008), BAC endsequences and a physical map based on contigs derived froma BAC library using the SNaPshot high-information-content-fingerprinting (HICF) method (Huo et al., 2009).However, other challenges, which are considered later inthis text, must be overcome for the Brachypodium ToolBox to achieve its full potential.
1. Comparative genomics: Brachypodium as a Pooidreference genome?
The value of Brachypodium was first seen in its ability toact as a bridge species (Jenkins et al., 2005) to aid the clon-
ing of genes in temperate cereals with extremely largegenomes. A bridge species is necessary because largergenomes have many DNA repeats that effectively isolategenes or ‘gene islands’ in map-based cloning approaches(Devos, 2010). By contrast, small genome grasses havegene-rich euchromatic regions with one gene per 5–15 kb,on average. Brachypodium and rice have similar propor-tions of retrotransposon sequences (21.4% and 26%,respectively), which is much less than sorghum (54%) orbread wheat (over 80%) (IBI, 2010). Even before the gen-ome sequence was available, the chromosomal pairing locusPh1 was accurately mapped to a cluster of kinase geneswithin a heterochromatic segment of wheat chromosome5B using markers derived from a much smaller orthologousregion in B. sylvaticum (Griffiths et al., 2006). Interestingly,although the rice genome sequence was available, itssequence was too divergent from wheat in this region toprovide markers and it could not be used to fine map thePh1 locus. In other examples, markers from Brachypodiumwere used to identify the Lr34 ⁄ Yr18 wheat rust resistancegene (Spielmeyer et al., 2007) and the barley Ppd-H1 photo-period response gene (Turner et al., 2005).
The availability of the Bd21 genomic sequence, EST col-lections and resequencing data from other accessions (soonto be available) means that more genomic sequence dataand markers are now coming on line to facilitate genomiccomparisons. Simple sequence repeat (SSR) microsatellitemarkers are widely used as anchor markers in genetic map-ping, and in marker-assisted breeding. A large number ofSSR markers have been derived from B. distachyon(Azhaguvel et al., 2009; Vogel et al., 2009; Garvin et al.,2010) offering the possibility of rapidly identifying geneticloci associated with a trait and translating this to other cere-als. Garvin et al. (2010) populated the Brachypodiumgenome with a total of 139 SSR markers derived fromESTs, BAC end sequences (BES) and conserved ortholo-gous sequences (COS) from a range of grass species. Inmacrosyntenic comparisons, 13 out of 20 Brachypodiumlinkage groups have equivalents in the rice genomesequence. Azhaguvel et al. (2009) developed and used 160EST- and 21 derived genomic microsatellite markers toevaluate genetic diversity among the Brachypodium acces-sions and to relate the observed genetic races to the feedingpreferences of the wheat greenbug (Schizaphis graminumRondani) and the Russian wheat aphid (RWA) (Diuraphisnoxia). Further, phylogenetic analysis suggested thatBrachypodium is closer to Aegilops tauschii (the D genomedonor of common wheat) than to rice. When EST contigswere compared, Brachypodium exhibited orthology withwheat EST contigs from all 21 of the wheat chromosomes(Kumar et al., 2009).
Some astonishing examples of orthology betweenBrachypodium and the temperate cereals at a single genelevel have been reported. In Triticeae genomes, a tandem
NewPhytologist Tansley review Review 337
� 2011 The Authors
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011) 191: 334–347
www.newphytologist.com

duplication occurred in a globulin gene leading to the selec-tion of a high molecular weight glutenin gene – an essentialtrait in bread wheat. This globulin duplication was presentin Brachypodium but not in any tropical grass genome (Guet al., 2010). The Eps-A (m) 1 locus, which controlsmorphological changes, such as spiking in Brachypodiumand in one wheat genome progenitor (A. tauschii), is verysimilar to orthologues at this locus, including, crucially,Mot1 and FtsH4, which are tightly linked to the earliness perse phenotype (Faricelli et al., 2010). Nevertheless, somestudies have suggested other genomic relationships. Yuet al., (2009) associated the Hessian fly resistance gene H26(mapped to the wheat 3DL distal region 3DL3-0.81-1)with a Brachypodium supercontig. Encouragingly, 14 ofthe 15 ESTs were collinear between the distal region ofwheat 3DL and Brachypodium supercontig 13. However,of 46 STS (sequence tagged site) primer markers derivedfrom this supercontig, only one could be mapped to wheatchromosome 3D. The apparent lack of conservation wassuch that the authors advised caution when using theBrachypodium genomic sequence for molecular mappingand gene cloning in wheat.
Reconciliation of these seemingly contradictory data cameabout with the publication of the Brachypodium genomesequence. On a global level, Brachypodium and rice exhibita high degree of conservation of gene order such that entirerice chromosomes or chromosome arms can be mapped totheir Brachypodium counterparts. By contrast, the align-ment of diploid wheat ancestor (A. tauschii) and barleygenetic maps to the Brachypodium genome is more frag-mentary, although there are still large segments of conservedalignment. This indicates that there were a large number ofgenomic rearrangements in the lineage containing wheat andbarley after the divergence from the Brachypodium lineage.Thus, instead of simply using Brachypodium as a roadmapfor the wheat and barley genomes through simple pairwisecomparisons, it will be more effective to use multiple com-parisons of gene order in sequenced grass genomes.However, because Brachypodium shares a higher degree ofnucleotide sequence conservation with the temperate grains,markers created using Brachypodium sequence have a muchhigher conversion rate.
When colinearity of genes within seven large genefamilies was examined four exhibited a high degree of con-servation in gene order between cereal genomes, but thiswas not the case for the nucleotide binding site (NBS)-leucine-rich repeat (LRR) and F-box gene families (IBI,2010). This could indicate rapid diversification owing tostrong natural selection driven by pathogen pressure in thecase of NBS-LRR and by the regulation of both develop-mental and stress responsive traits in the case of F-boxes(Meyers et al., 2003; Xu et al., 2009). Thus, the degree ofsynteny between Brachypodium and wheat in any particularregion will vary owing to both the historical rearrangements
after the divergence of the lineage and evolutionary history ofthe gene, which may be accelerated in specific gene families.
Interestingly, when only gene sequence is compared, theoverwhelming majority of gene families are highly con-served between rice, sorghum, Brachypodium, wheat andbarley; only 265 out of 16 215 gene families were specificto the Pooideae (Brachypodium, wheat and barley). Thus,there are considerable genomic similarities between theEhrhartoideae (rice), Panicoideae (Sorghum) and Pooideae(B. distachyon, T. aestivum and Hordeum vulgare), undoubt-edly reflecting their relatively recent evolutionary divergence.Thus, in addition to serving as a structural model for thelarge genomes of the temperate grains, the overall similarityat the gene level indicates that, for the majority of traits,Brachypodium can serve as a functional genomic model forall the grasses. As a corollary to this, depending on the trait,Brachypodium might not be the best source of comparativegenomic information for the Triticeae and a comparativeapproach using all available genomes is advised. This may bemore important for traits that have come under selectionpressure during rapid evolutionary diversification, and aretherefore specific to the Triticeae, such as traits conferred bythe NBS-LRR and F-box’s genes (IBI, 2010).
2. Transformation and reverse genetic tools withinthe Brachypodium Tool Box
Efficient transformation is a prerequisite for a modern modelorganism and we are fortunate that Brachypodium has beenvery amenable in this regard. Biolistic bombardment-basedtransformation of a polyploid line was first reported in 2001(Draper et al., 2001). A more efficient method that workedon a diploid line was later reported in 2005 (Christiansenet al., 2005). However, Agrobacterium is the preferred trans-formation method, where it is important to have simple, lowcopy integration events and methods with efficiencies up to30% have been published (Vain et al., 2008; Vogel & Hill,2008). Today, the average efficiency in a production settingwhere the emphasis is on minimizing the labour per trans-genic line rather than maximizing the efficiency of eachtransformation is c. 45% (up-to-date methods are available athttp://brachypodium.pw.usda.gov/). With these improve-ments Brachypodium is arguably one of, if not the, mosteasily transformed grasses.
The efficiency of Brachypodium transformation makesfeasible the creation of collections of sequence indexed T-DNA mutants. An excellent example of the power of aT-DNA population to reveal gene function is the SALKT-DNA tagged Arabidopsis resource (Ecker, 2002). Sucha resource is a crucial component of the BrachypodiumTool Box. Two groups have established projects to createBrachypodium T-DNA mutants. Researchers at the JohnInnes Centre have reported the generation of a collectionof 4500 T-DNA lines. Analysis of 741 accessions showed
338 Review Tansley reviewNewPhytologist
� 2011 The Authors
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011) 191: 334–347
www.newphytologist.com
that 660 T-DNA loci could be assigned to a unique loca-tion in the Brachypodium genome sequence (Thole et al.,2010). Of these, c. 60% could be associated with ESTs.The T-DNA lines generated by the BrachyTAG pro-gramme are available as a community resource and havebeen distributed internationally since 2008 via the website (http://www.brachytag.org/). Similarly, researchers atthe USDA-ARS Western Regional Research Center aregenerating thousands of tagged lines. At the time of writ-ing > 10 000 have been created, and over 6000 flankingsequence tags (FSTs), have been assigned to a uniquelocation in the genome. Rather than concentrating onlyon T-DNA tagging, this effort is also using gene-trappingvectors to identify promoters with useful expression pat-terns, and activation tagging vectors that containtranscriptional enhancers to ‘activation tag’ nearby genes.Details and ordering instructions can be found at http://brachypodium.pw.usda.gov/TDNA.
In addition to insertional mutants, a method to mutage-nize seeds with ethyl methanesulphonate (EMS) has beenoptimized (http://brachypodium.pw.usda.gov), and irradia-tion with fast neutrons has also been used to create mutantcollections (D. Laudencia-Chingcuanco and M. Byrne,pers. comm.). These EMS mutants are very useful forforward genetic screens because of the large number ofmutants per plant. In addition, EMS mutants can also beused to identify mutations in particular genes for reversegenetic studies by a TILLING (Targeted Induced LocalLesions in Genomes) approach (McCallum et al., 2000)which employs a mismatch specific endonuclease to identifymutated PCR amplicons. The authors are aware of thecreation of one TILLING population (http://www-ijpb.versailles.inra.fr/en/institut/actualite.htm) which currentlyconsists of c. 6000 individuals.
Although these emerging mutagenic approaches areimpressive, it will take some time before sufficient genomecoverage is achieved for most Brachypodium genes to havea corresponding mutant. Given this, the development of atargeted gene disruption strategy based on virus-inducedgene silencing (VIGS) represents a significant advance as itrepresents an immediately applicable strategy throughwhich the expression of targeted genes can be disrupted.Barley stripe mosaic virus (BSMV) is a single-stranded tripar-tite RNA virus where infection of the host can occurfollowing simple rubbing of leaves with naked (nonenvel-oped or capsidated) genome. Derivatives of this virus,including fragments of a targeted gene, have been used tosuppress gene expression in barley and wheat (Holzberget al., 2002; Scofield et al., 2005). Recently, two studieshave shown the efficacy of BSMV-induced VIGS inBrachypodium. Demircan & Akkaya (2010) silenced a phy-toene desaturase gene and Pacak et al. (2010) suppressedthe expression of genes IPS1, PHR1 and PHO2 knownto participate in phosphate (Pi) uptake and reallocation.
Following these successes, we expect VIGS to be widelyemployed in Brachypodium research.
3. Cytogenetic tools for genome analyses
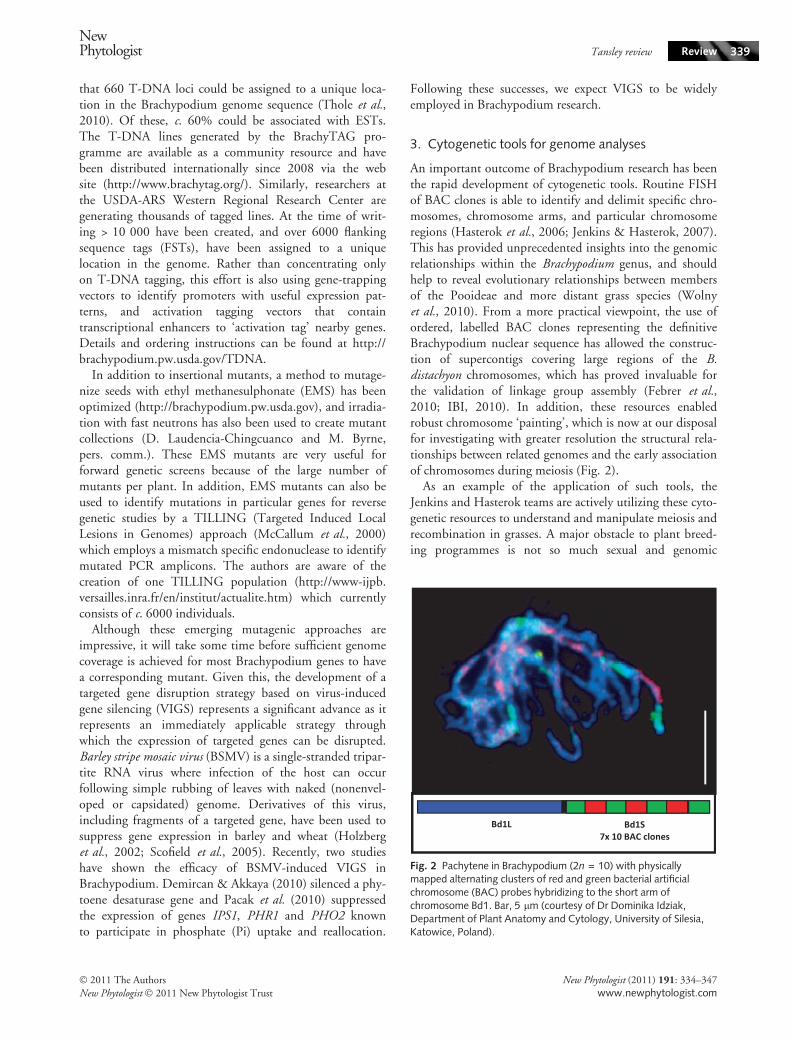
An important outcome of Brachypodium research has beenthe rapid development of cytogenetic tools. Routine FISHof BAC clones is able to identify and delimit specific chro-mosomes, chromosome arms, and particular chromosomeregions (Hasterok et al., 2006; Jenkins & Hasterok, 2007).This has provided unprecedented insights into the genomicrelationships within the Brachypodium genus, and shouldhelp to reveal evolutionary relationships between membersof the Pooideae and more distant grass species (Wolnyet al., 2010). From a more practical viewpoint, the use ofordered, labelled BAC clones representing the definitiveBrachypodium nuclear sequence has allowed the construc-tion of supercontigs covering large regions of the B.distachyon chromosomes, which has proved invaluable forthe validation of linkage group assembly (Febrer et al.,2010; IBI, 2010). In addition, these resources enabledrobust chromosome ‘painting’, which is now at our disposalfor investigating with greater resolution the structural rela-tionships between related genomes and the early associationof chromosomes during meiosis (Fig. 2).
As an example of the application of such tools, theJenkins and Hasterok teams are actively utilizing these cyto-genetic resources to understand and manipulate meiosis andrecombination in grasses. A major obstacle to plant breed-ing programmes is not so much sexual and genomic
Fig. 2 Pachytene in Brachypodium (2n = 10) with physicallymapped alternating clusters of red and green bacterial artificialchromosome (BAC) probes hybridizing to the short arm ofchromosome Bd1. Bar, 5 lm (courtesy of Dr Dominika Idziak,Department of Plant Anatomy and Cytology, University of Silesia,Katowice, Poland).
NewPhytologist Tansley review Review 339
� 2011 The Authors
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011) 191: 334–347
www.newphytologist.com

incompatibility, as the retention over many backcross gen-erations of undesirable wild germplasm. Purging potentialnew cultivars of this material is costly and time consuming,and largely governs the speed at which new cultivars arereleased. These negative effects of linkage drag are inverselyproportional to the extent of genetic recombination in suc-cessive backcross generations.
Recombination is assayed traditionally by direct cytologi-cal scoring of chiasmata at metaphase I of meiosis, whichhas shown that the chromosomes of temperate, large-genome cereals and grasses, such as wheat, barley, rye andryegrass, each have regularly two or more chiasmata.However, cytological inspection shows that the vast major-ity of these are confined to distal regions, meaning thatinterstitial and proximal chromosome segments seldomengage in recombination events. This uneven distributionof crossovers can also be inferred from measurements ofrecombination frequencies between genetic markers inmapping populations, which are then correlated to physicalchromosome positions using deletion and introgressionmapping or recombination nodules. Such studies in, forexample, wheat (Erayman et al., 2004; Saintenac et al.,2009) have revealed that there is a common gradient ofincreasing recombination from proximal to distal regions ofchromosomes. Because it is now known that the interstitialand proximal regions of cereal and grass chromosomes har-bour a sizeable proportion of the genes of the genome(albeit at a lower density), the inevitable conclusion is that alarge part of the genome of these species is not regularlyinvolved in recombination events, effectively consigningmany genes to recombination backwaters. This limits thepotential of genetic variation in these crops, it prolongslinkage drag in introgression programmes, and it confoundsthe scope of map-based cloning approaches. Indeed, it hasbeen acknowledged that ‘> 30% of wheat genes are inrecombination-poor regions and thus are inaccessible tomap-based cloning’ (Erayman et al., 2004). Clearly, itwould be desirable to crack open these recombination cold-spots in order to release new genetic variation which couldbe exploited in advanced breeding programmes. However,even if sites of recombination were shifted, map-based clon-ing strategies would still be constrained by the inordinatelylarge genome sizes of these species.
Preliminary studies of meiosis in a diploid and an allote-traploid accession of Brachypodium (Jenkins et al., 2005)have shown that chiasmata are not distally localized as inthe other members of the Poaceae mentioned above. Thus,if there is variation in recombination among Brachypodiumgermplasm there is a real prospect of establishing the geneticbasis of chiasma location and frequency. Variation in geno-mic recombination has been noted in Arabidopsis(Sanchez-Moran et al., 2002), rye (Rees, 1961), Lolium andFestuca (Jones & Rees, 1966; Rees & Dale, 1974), andbarley (Gale & Rees, 1970).
4. Looking into the Brachypodium Tool Box:bioinformatic tools
In the multinational effort required to developBrachypodium as a model, parallel development ofappropriate bioinformatic tools allows the collection, cura-tion and interrogation of genomic and post-genomic data.Readers unfamiliar with Brachypodium are recommendedto use the bioinformatic tools available through thehttp://www.brachypodium.org website. From this website,it is possible to access the Brachypodium genome browserBrachybase (http://www.brachybase.org), where the 8XAssembly genome can be viewed, downloaded it in its entirety,and related to EST databases and Affymetrix (BdArray) arrayprobes. A BLAST tool (http://www.brachybase.org/blast/)allows nucleotide and amino acid sequence comparisons.For excellent cross-species genomic comparisons and as arepository of much information on Brachypodium, theGramene website (http://www.gramene.org/Brachypodium_distachyon/) is also a good place to start.
Other databases (http://mips.helmholtz-muenchen.de/plant/brachypodium/) allow whole genome, proteinsequence ⁄ structures and motifs to be investigated and pro-moter regions to be accessed. http://www.modelcrop.org/has many of the functions of these other sites but also allowsthe Brachypodium physical map to be displayed and easilycompared with the rice and sorghum genomes. With http://www.phytozome.net it is possible to search for orthologousor homologous genes among all sequenced plant genomesbased on name or sequence. Matches may be extracted andalignments compared by progressive alignment algorithms(dynamic programming) while relationships can be dis-played using phylogenetic approaches or multivariateprincipal component analysis.
One result of all of these bioinformatic data and tools hasbeen the development of an Affymetrix array based on theBd21 genome sequence and EST databases (S. E. Fox et al.,unpublished data). The derived oligonucleotides were usedto generate the array representing unique single copysequences, with mean probe spacing of 42 bases with 95%of probe pairs < 126 nucleotides apart enabling studies ofgene-specific expression. S. E. Fox et al. (unpublished) areusing this array resource to characterize Bd21 expressionduring development, diurnal and circadian cycling as wellas abiotic or biotic stress to represent a ‘Bd21 expressionatlas’. It is expected that these data will be made availablevia a web portal analogous to the Arabidopsis genomic site(e.g. TAIR Microarray Experiments).
5. Diversity within the Brachypodium Tool Box:germplasm collections
If Brachypodium is to act as functional genomic model forthe Pooideae and other grasses, germplasm collections must
340 Review Tansley reviewNewPhytologist
� 2011 The Authors
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011) 191: 334–347
www.newphytologist.com

contain accessions with economically relevant traits andencompass sufficient variation for mapping projects to suc-ceed. Thus, an essential part of the Brachypodium Tool Kitmust be well-characterized germplasm collections (Filizet al., 2009). Sequence data from comprehensive germ-plasm collections will yield markers linked to specific lociand help identify gene function, for example, improvedstress tolerance or yields, enabling the isolation of corre-sponding alleles in grasses and cereals. Brachypodium genescould also be used directly to generate GM cereal deriva-tives. Beyond cereal ⁄ forage grass improvement strategies, awell-curated Brachypodium germplasm collection will alsoallow macro and micro level evolutionary trends to be mod-elled and related to past or present selection pressures. Itshould also be possible to predict losses in genetic diversityby genetics and ecological niche modelling, due to environ-mental change or human activities.
Until recently, a salient feature of Brachypodium researchhas been the relative paucity of available germplasm. Muchearly work focused on only seven inbred lines (Bd1-1, Bd2-3, Bd3-1, Bd18-1, Bd21, Bd21-3 and Bd29) developedfrom USDA collections (http://www.ars-grin.gov/npgs) andanother small collection of ABR accession (ABR1 throughABR7) originating mainly from Spain (Stace and Catalancollection, Leicester, UK). Now, however, there is a largecollection of Turkish germplasm available: 195 diploid linescollected from 53 locations in Turkey (Filiz et al., 2009;Vogel et al., 2009). Within this large collection consider-able variation in flowering time, seed size, and plantarchitecture was noted. Sixty-two wild accessions (firstgeneration) were analysed with 43 SSR markers and the vastmajority of individuals were homozygous, despite the pres-ence of multiple alleles in the local population. Underlaboratory conditions, intimately grown lines failed to out-cross. This reflects near-cleistogamy in the diploid lineswhich, while facilitating genotype conservation, poses aconsiderable barrier to full exploitation of theBrachypodium Tool Box as this makes crosses and deriva-tion of mapping families quite difficult. However,following on from an early demonstration of successfulcrossing of two Brachypodium accessions (Routledge et al.,2004), recombinant inbred lines have been derived (Garvinet al., 2008) and simple, very efficient step-by-step crossingprotocols are now available (see http://www.ars.usda.gov/SP2UserFiles/person/1931/BrachypodiumCrossing.pdf andhttp://brachypodium.pw.usda.gov). These relatively simpleprotocols represent an essential component of theBrachypodium Tool Box and we expect the populations ofrecombinant inbred lines developed by Garvin et al. (2008)will soon be added to by those from other groups.
We are now greatly expanding the germplasm collectionwith numerous collections from Northern Spain (Fig. 3a).In this collection, we have concentrated on developinginbred lines from various environments. Thus, we have
sampled individual seeds from 46 localities, from high alti-tudes in the foothills of the Pyrenees, to lowland areasaround the flood plain of the Ebro river basin, coastal areasaround Catalunya and a Balearic Island populationcollected from Ibiza (Table 1). Between 10 and 30 individ-uals were sampled from each site, with each sampleseparated from the others by at least 1 m to avoid samplingclose relatives. This collection is being developed specificallyto search for intrapopulation and interpopulation variationand to relate this genetic diversity to potential adaptation toenvironmental selection pressures. We also are seeking todetermine the relationships between sympatric 2n = 10 and2n = 30 Brachypodium populations, and we have at leastone site where the two ploidy-level species are sympatric(Table 1, Bierge, Huesca, Spain).
Preliminary assessment of genotypic variation in a total of38 wild individual samples, representative of most of theSpanish Brachypodium germplasm localities (Table 1,Fig. 3a), involved comparison with five individual samplesrepresenting different Turkish Brachypodium lines (Vogelet al., 2009) and Bd21. The number of alleles surveyed perlocus ranged from 2 to 14, with an average of 6.85 across the44 samples studied. Six out of 14 screened loci showed totalor predominant levels of homozygosity across the 44 germ-plasm lines assessed (with observed heterozygosity valuesranging from 0 to 0.159). Relationships among individualsamples were evaluated through Nei et al.’s (1983) Da dis-tance-based neighbour-joining (NJ) reconstruction. Theunrooted NJ phenogram (Fig. 3b) showed: a large Spanishclade formed by samples from medium to relatively high alti-tude Pyrenean and Prepyrenean localities (Fig. 3a‘highland’) and their close low altitude Ebro river valleylocalities (Fig. 3a ‘lowland’); a Turkish–Iraqi clade formedby most of the samples surveyed from different altitudinallocalities of Turkey, including the sequenced line Bd21 anda line developed from the same location, Bd21-3; and a‘Miscellaneous’ clade formed by the coastal Balearic isles(Ibiza: Fig. 3a ‘island’) samples plus one sample from lowaltitude Tekirdag (Tek10, Turkey) and four samples fromboth highland and lowland Iberian localities.
Although only a small number of Turkish–Iraqi germplasmrepresentatives were tested, a greater genetic diversity wasobserved in wild Brachypodium individuals from the westernMediterranean region compared with those from the eastern.Variation in DNA sequences within other temperateMediterranean grasses (e.g. Hordeum marinum, Jakob et al.2007), has similarly suggested greater diversity in the linesfrom the western part of the region, which may have containeda larger number of glacial refugia during the last ice age fromwhich a larger number of ‘bottlenecked’ populations wouldhave been derived. More exhaustive comparative phylogeo-graphical studies of Brachypodium populations across itsnative Mediterranean distribution area are currently underwayby Catalan’s group to test this hypothesis.
NewPhytologist Tansley review Review 341
� 2011 The Authors
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011) 191: 334–347
www.newphytologist.com

V. Targets for the Brachypodium Tool Box: keytraits
When Brachypodium was first suggested as a model, somekey traits were highlighted which were suggested to be ofrelevance to temperate cereals and grasses (Draper et al.,2001). Subsequently, a large number of groups have begunto define the molecular basis of these traits and, as a conse-quence, assess to what extent these are of relevance tocereals, grasses and biofuel crops.
1. Pathogen attack
Given the interests of the research groups which first sug-gested Brachypodium, many early papers focused onresponses to plant–pathogen interactions (Draper et al.,
2001). Following on from preliminary pathogen screenswhich included examining cereal-adapted fungal rust andmildew pathogens (Draper et al., 2001), the first well-charac-terized Brachypodium pathogen was Rice Blast (Magnaporthegrisea) (Routledge et al., 2004). Subsequent metabolomicanalyses have suggested that phospholipid processing andjasmonate accumulation play important roles in the inter-action of Brachypodium with M. grisea (Allwood et al., 2006).However, it is questionable how far Brachypodium should bepursued as a model for Rice Blast disease, given the availabil-ity of a genome sequence for rice and its excellent functionalgenomic infrastructure (Leung & An, 2004). Instead, there isa clear need for greater efforts to develop Brachypodiuminteractions for pathogens of temperate cereals. Perhaps mostpressingly well-characterized interactions involving rust fun-gal pathogens are required (Ayliffe et al., 2008). Following
(a)
(b)
33 29
2926
Turkey - Iraq
Miscellaneous
Spain
0.05
Fig. 3 The development of an ecotypicBrachypodium germplasm collection. (a) Sitesof Brachypodium sampling in north-eastSpain: ‘Lowland’, light blue circles;‘Highland’, orange circles; ‘Coastal’, greencircles; ‘Island’, dark blue circles. Insets areexample images of a ‘lowland’ (Alfranca,Zaragoza, Spain), ‘highland’ (Puerto delPerdon, Navarra, Spain), ‘coastal’ (PortLligat, Cadaques, Girona, Spain) and ‘Island’(Port des Torrents, Ibiza, Spain) region. (b)Preliminary assessment of genotypicvariation of this new germplasm collectionhas been undertaken using 14 bacterialartificial chromosomes end sequences (BES)-derived tetranucleotide- trinucleotide- anddinucleotide-based repeats simple sequencerepeat (SSR) markers described at the USDABrachypodium web site (http://brachypodium.pw.usda.gov/SSR/) (lociDB069I22, DB078E03, DB088E23,DB080E01, DH044F02, DB078M18,DH038F15, DB060E14, DB092H16,DB082M05, DB009L08, DB071F17,DH048N18, DH038H13, DB088J19).Descriptions of the germplasm together witheither confirmed or tentative estimations ofploidy are given in Table 1. Neighbour-joining phenogram from 96 shared SSRalleles based on Nei & Tajima (1983) Dadistance between 44 Spanish and Turkish–Iraqi Brachypodium wild-type germplasmlines. Bootstrap values on branches are basedon 10 000 replications. Symbols: blue circle,Spanish highlands; red circle, Spanishlowlands; dark green square, Turkish–Iraqi(the Bd21 whole sequenced-genome sampleis encircled); bright green square, Tekirdag10 (Turkey); purple triangle, Balearic coastal.
342 Review Tansley reviewNewPhytologist
� 2011 The Authors
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011) 191: 334–347
www.newphytologist.com
from our early work on wheat and barley adapted yellow(Puccinia striiformis) and brown (Puccinia recondita) rusts, wehave focused on crown rust (Puccinia coronata; J. V. R.Smith et al., unpublished). A model interaction betweenBrachypodium and the more important stem rust (Pucciniagraminis) has also been established (David Garvin, USDA, StPaul, MN, USA, pers. comm.). However, (to the authors’knowledge) no model interactions between Brachypodiumand other major cereal pathogens, including Blumeria (pow-dery mildew), Septoria (the cause of leaf blotch), Fusariumgraminearum (the cause of ear blight) and Rhizoctonia solanior Gaeumannomyces sp. (the cause of take all) have beenestablished. We note that a model interaction between aphidfeeding Schizaphis graminum and Diuraphis noxia andBrachypodium has been established (Azhaguvel et al., 2009)but others involving biting rather than piercing insects needto be developed.
Of relevance to plant pathology are the comparativestudies that have been made with defined resistance geneswithin the Brachypodium genome. Drader & Kleinhofs(2010) used genetic markers for barley chromosomes toconstruct a synthetic map with Brachypodium but with aparticular focus on rust resistance genes. The stem rustresistance genes Rpg1 and Rpg4 have very similar ortho-logues but with Rpg5 key motifs, consisting of the -lLRR
and a serine ⁄ threonine protein kinase domain that is sepa-rately encoded on two Brachypodium contigs. In a widergenomic comparison Li et al. (2010) examined NBS-LRRgenes in maize, sorghum, Brachypodium and rice. Theyobserved considerable variation between species, with only3.93% of NBS-LRR conforming to a conserved family.This compared with 96.1% conservation in selectedhousekeeping genes (Li et al., 2010). Although these com-parisons were with nontemperate grasses, the lack ofconservation was striking and most likely reflects rapidgene diversification as a result of pathogen-mediated selec-tion pressures.
2. Cell wall and bioenergy crop characteristics
Much of the funding for sequencing the Brachypodiumgenome originated from the US Department of Energywho foresaw its value as a model for energy grasses (DOE,2006). Biofuel crops can contribute to the energy economyby serving as either direct combustion inputs into powerstations or representing substrates for biofermentationprocesses leading the production of bioethanol or biodiesel(Chang, 2007; Gomez et al., 2008a,b). In particular,moving beyond simply fermenting easily extracted starchand sugar, the substantial resources captured within the lig-
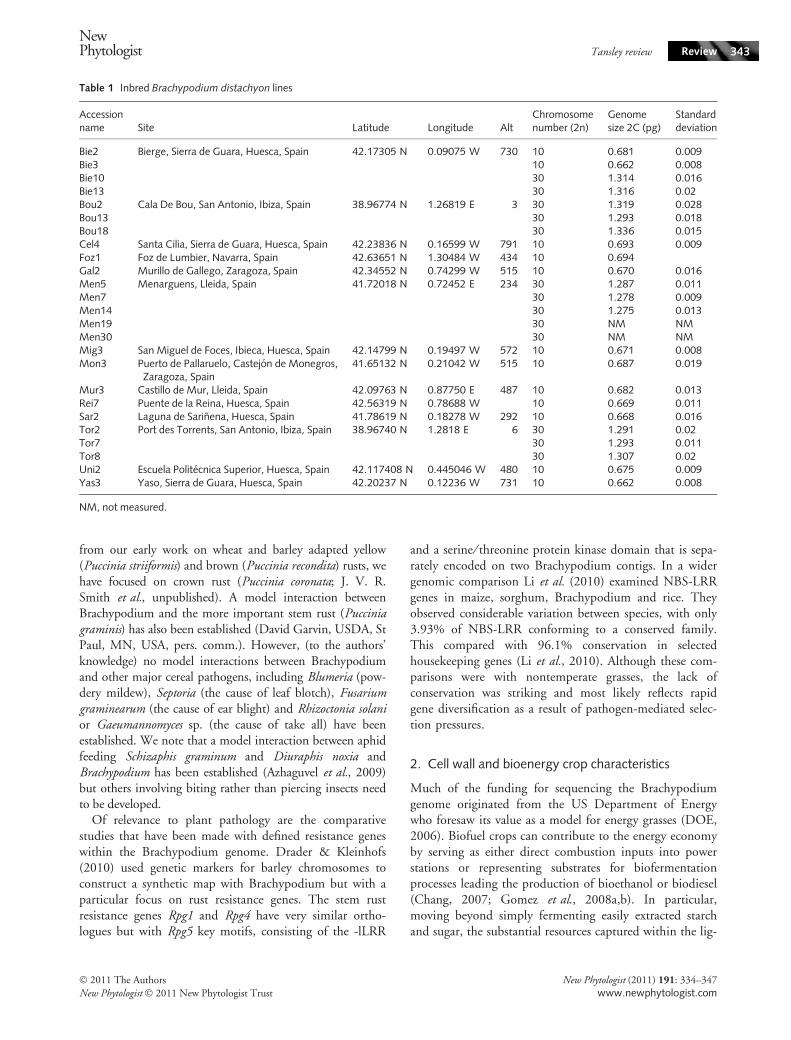
Table 1 Inbred Brachypodium distachyon lines
Accessionname Site Latitude Longitude Alt
Chromosomenumber (2n)
Genomesize 2C (pg)
Standarddeviation
Bie2 Bierge, Sierra de Guara, Huesca, Spain 42.17305 N 0.09075 W 730 10 0.681 0.009Bie3 10 0.662 0.008Bie10 30 1.314 0.016Bie13 30 1.316 0.02Bou2 Cala De Bou, San Antonio, Ibiza, Spain 38.96774 N 1.26819 E 3 30 1.319 0.028Bou13 30 1.293 0.018Bou18 30 1.336 0.015Cel4 Santa Cilia, Sierra de Guara, Huesca, Spain 42.23836 N 0.16599 W 791 10 0.693 0.009Foz1 Foz de Lumbier, Navarra, Spain 42.63651 N 1.30484 W 434 10 0.694Gal2 Murillo de Gallego, Zaragoza, Spain 42.34552 N 0.74299 W 515 10 0.670 0.016Men5 Menarguens, Lleida, Spain 41.72018 N 0.72452 E 234 30 1.287 0.011Men7 30 1.278 0.009Men14 30 1.275 0.013Men19 30 NM NMMen30 30 NM NMMig3 San Miguel de Foces, Ibieca, Huesca, Spain 42.14799 N 0.19497 W 572 10 0.671 0.008Mon3 Puerto de Pallaruelo, Castejon de Monegros,
Zaragoza, Spain41.65132 N 0.21042 W 515 10 0.687 0.019
Mur3 Castillo de Mur, Lleida, Spain 42.09763 N 0.87750 E 487 10 0.682 0.013Rei7 Puente de la Reina, Huesca, Spain 42.56319 N 0.78688 W 10 0.669 0.011Sar2 Laguna de Sarinena, Huesca, Spain 41.78619 N 0.18278 W 292 10 0.668 0.016Tor2 Port des Torrents, San Antonio, Ibiza, Spain 38.96740 N 1.2818 E 6 30 1.291 0.02Tor7 30 1.293 0.011Tor8 30 1.307 0.02Uni2 Escuela Politecnica Superior, Huesca, Spain 42.117408 N 0.445046 W 480 10 0.675 0.009Yas3 Yaso, Sierra de Guara, Huesca, Spain 42.20237 N 0.12236 W 731 10 0.662 0.008
NM, not measured.
NewPhytologist Tansley review Review 343
� 2011 The Authors
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011) 191: 334–347
www.newphytologist.com

nocellulosic cell wall is proving to be an attractive target(Gomez et al., 2008a).
Emerging biofuel crops such as Miscanthus and switch-grass (Panicum virgatum) therefore represent good sources ofbiomass with considerable potential. However, the conver-sion of lignocellulose to biofuel is a scientifically andeconomically demanding process whereby cell walls must beefficiently hydrolysed and the released sugars fermented. It isimperative to develop our understanding of the constructionof the cell wall and its degradation and saccharification(Wyman, 2007). Alfalfa mutants with altered lignin levelshave already proved useful in such studies (Chen & Dixon,2007) but, with its genomic infrastructure, a focus onBrachypodium represents a cost effective strategy to rapidlygain greater insights. Indeed, a characterization of saccharifi-cation of Brachypodium stem segments under differentpretreatment and hydrolysis conditions has already beenpublished (Gomez et al., 2008b). Crucially, the compositionof the Brachypodium cell wall was more similar to that ofbiomass grasses and cereals than that of Arabidopsis, con-firming it to be a more suitable model species (Gomez et al.,2008b; Opanowicz et al., 2008).
Christensen et al. (2010) compared the cell walls ofBrachypodium, H. vulgare and T. aestivum, focusing partic-ularly on hemicelluloses. This study showed that thepresence of (1,3;1,4)-beta-D-glucans correlated with cellwall elongation but this could not be related to thetranscriptional regulation of genes involved in (1,3-1,4)-beta-D-glucan synthesis (cellulose synthase-like F family(CSLF)). Similarly, in all three species arabinoxylansincreased during growth but in Brachypodium subsequentcrosslinking via ferulic acid dimers and ester-linked p-coumaric acid were more prominent. In a key study, VanHulle et al. (2010) demonstrated that by modulating genesinvolved in the Brachypodium cell wall biosynthesis theextractability of sugars could be improved. This importantstudy validates Brachypodium as a biofuel crop model. Thisearly work will surely be followed up by many.
Senescence is also an important trait in a biofuel crop, interms either of its delay to prolong plant greenness and thusincrease sugar levels for fermentative processes, or of improv-ing nutrient mobilization to overwintering organs inperennial crops. Brachypodium has been used along withswitchgrass to study fatty acid mobilization and the initiationand development of senescence (Yang & Ohlrogge, 2009).
3. Abiotic stress tolerance
Somewhat surprisingly, given its preference for growth onmarginal ground or at higher altitudes, the potential ofBrachypodium as a model to understand abiotic stress toler-ance has yet to be addressed. There has been some limitedwork in this area using B. pinnatum (Hurst & John, 1999)or B. rupestre (Liancourt et al., 2005). Although focusing on
switchgrass, Matts et al. (2010) used high-throughputsequencing and cross-genomic data mining to identify 27conserved miRNAs with possibly an additional 129 inBrachypodium. Some of the miRNAs proved to be modu-lated by cold, the vast majority of which were not conserved,suggesting some unique aspects to the response to cold inBrachypodium. As stresses such as drought and flooding, areimportant limiting factors to cereal crop yields (Cassman,1999), we fully expect other studies into abiotic stressresponses and tolerance to be published in the near future.
4. Grain characteristics
One surprising feature of Brachypodium is its large grainsize, relative to its stature. As has been frequently pointedout (Draper et al., 2001; Garvin et al., 2008; Opanowiczet al., 2008), in its length if not its width, Brachypodiumseeds are similar to those of rice and wheat. Furthermore, itslarge seed head is also very prominent in natural environ-ments (Fig. 1a). Thus, even though B. distachyon is anundomesticated grass species, workers have examined itsseed storage accumulation and localization (Gu et al., 2010;Larre et al., 2010). Proteomic analyses have revealed thepredominance of globulins which, based on their solubility,appeared to be similar to the rice seed storage protein glut-elin (Laudencia-Chingcuanco & Vensel, 2008).It appearsthat some Brachypodium species may have been among thefirst grains to be processed by Palaeolithic hunter gatherersc. 30 000 yr ago (Revedin et al., 2010).
Brachypodium has also been exploited in studies examin-ing flowering. One key study demonstrated that theflowering transition gene GIGANTEA functions in a similarway in Brachypodium as in Arabidopsis (Hong et al., 2010).In another bioinformatic screening of existing B. distachyonEST databases, Unver & Budak (2009) detected 26miRNA which were predicted to target as many as 246mRNAs. Most of the putative target mRNAs encoded tran-scription factors, which were predicted to regulate plantdevelopment and flowering time (Unver & Budak, 2009).The potential practical application of such studies wasshown by Olsen et al. (2006) who expressed Lolium andArabidopsis Terminal Flower 1 genes in Brachypodiumaccessions to delay heading.
VI. Whence for the Brachypodium Tool box?Primus inter pares?
It is very likely that, in the very near future, whole genomicsequence data will be available for all major cereal and foragecrops. Some such as Lolium and barley (H. vulgare) havemany attributes that make them good candidates for a modelspecies. Lolium is physically small and easy to grow whilebarley is easy to grow and has functional genomic resourcessuch as a TILLING population (Talame et al., 2008).
344 Review Tansley reviewNewPhytologist
� 2011 The Authors
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011) 191: 334–347
www.newphytologist.com

Further, Lolium has enormous value as a forage grass andbarley as a cereal, neither of which is the case forBrachypodium. However, such considerations betray amonolithic view where one model species dominatesresearch efforts, possibly influenced by the dominance thatArabidopsis has exerted on plant science. With the genomicdifficulties of using actual crop species rapidly disappearing,we suggest a pluralistic approach where research employsmodel, semimodel and crop species as required. Such a strat-egy would also augment comparative genomic approacheswhereby functions assigned in Brachypodium can be used toannotate as well as assemble grass and cereal genomes. Theintermediate evolutionary position of Brachypodiumbetween the core Pooideae (including Triticeae andAveneae–Poeae) and the Ehrhartoideae (including Oryzeae)makes it particularly valuable for such approaches.
Clearly, one species cannot adequately serve as a model ofall biological traits (e.g. photosynthesis in C4 grasses) but,as outlined above, Brachypodium comes close. It can serveas a host for some important cereal pathogens, act as a bio-fuel crop model as a way of gaining insight into cell wallstructures and biosynthesis, and can be used to study graindevelopment. Even more importantly, its ease of growthand small stature make it a low (logistical) cost but highgain (in terms of insights) species to work on. Whenthese inherent traits are combined with the establishedinfrastructure (e.g. extremely efficient transformation, excel-lent genome sequence, growing T-DNA collections,microarrays, expanding sequence resources, etc.) it becomesclear that Brachypodium will remain an important modelsystem. This stated, the Brachypodium Tool Box is notcomplete and the plant research community will benefitfrom continued investment, particularly in additionalreverse genetic tools.
The burgeoning number of germplasm collections alsorepresents both an opportunity and a challenge. Harvestinggenotypic variation will be essential to the functional exploi-tation of the Brachypodium genomic sequence(s); thegermplasm needs to be available to all. Precedents set byTAIR (http://www.arabidopsis.org) and NASC (http://arabidopsis.info/) demonstrate the need to have at least twogeographically disparate sites through which germplasm canbe maintained and distributed. This must be associatedwith a suitable web accessible interface, and http://www.brachypodium.org is clearly developing into such afacility. However, the infrastructure and resources (humanas well as computational) should not be underestimated;they are a challenge, especially as funds for such projects arebecoming scarcer (Abbott, 2009).
For those of us working with Brachypodium, furtherefforts are needed to develop the Brachypodium Tool Boxso that it could fulfil its potential. That potential could beas a model that is ‘first amongst equals’ (i.e. a preferredmodel species but complementary to others).
Acknowledgements
We thank Mrs Dorota Siwinska (University of Silesia,Katowice) for her technical assistance with flow cytometry.This work was partially supported by the Polish Ministry ofScience and Higher Education (grant N N303 570738).Pilar Catalan was funded by a Spanish Research GrantProject CGL2009-12955-C02-01. The establishment ofthe Mur-Catalan Spanish germplasm collection was sup-ported by the Genetics Society (UK).
References
Abbott A. 2009. Plant genetics database at risk as funds run dry. Nature462: 258–259.
Allwood JW, Ellis DI, Heald JK, Goodacre R, Mur LAJ. 2006.
Metabolomic approaches reveal that phosphatidic and phosphatidyl
glycerol phospholipids are major discriminatory non-polar metabolites
in responses by Brachypodium distachyon to challenge by Magnaporthegrisea. Plant Journal 46: 351–368.
Ayliffe M, Singh R, Lagudah E. 2008. Durable resistance to wheat stem
rust needed. Current Opinion in Plant Biology 11: 187–192.
Azhaguvel P, Li W, Rudd JC, Gill BS, Michels GJ Jr, Weng Y. 2009.
Aphid feeding response and microsatellite-based genetic diversity among
diploid Brachypodium distachyon (L.) Beauv accessions. Plant GeneticResources Characterization and Utilization 7: 72–79.
Brown TA, Jones MK, Powell W, Allaby RG. 2009. The complex origins
of domesticated crops in the Fertile Crescent. Trends in Ecology &Evolution 24: 103–109.
Cantacessi C, Jex AR, Hall RS, Young ND, Campbell BE, Joachim A,
Nolan MJ, Abubucker S, Sternberg PW, Ranganathan S et al. 2010. A
practical, bioinformatic workflow system for large data sets generated by
next-generation sequencing. Nucleic Acids Research 38: e171.
Cassman KG. 1999. Ecological intensification of cereal production
systems: yield potential, soil quality, and precision agriculture.
Proceedings of the National Academy of Sciences, USA 96: 5952–5959.
Catalan P, Kellogg EA, Olmstead RG. 1997. Phylogeny of Poaceae
subfamily Pooideae based on chloroplast ndhF gene sequences.
Molecular Phylogenetics and Evolution 8: 150–166.
Catalan P, Olmstead RG. 2000. Phylogenetic reconstruction of the genus
Brachypodium P-Beauv. (Poaceae) from combined sequences of
chloroplast ndhF gene and nuclear ITS. Plant Systematics and Evolution220: 1–19.
Chang MCY. 2007. Harnessing energy from plant biomass. CurrentOpinion in Chemical Biology 11: 677–684.
Chen F, Dixon RA. 2007. Lignin modification improves fermentable
sugar yields for biofuel production. Nature Biotechnology 25: 759–761.
Christensen U, Alonso-Simon A, Scheller HV, Willats WGT, Harholt J.
2010. Characterization of the primary cell walls of seedlings of
Brachypodium distachyon – a potential model plant for temperate grasses.
Phytochemistry 71: 62–69.
Christiansen P, Andersen CH, Didion T, Folling M, Nielsen KK. 2005.
A rapid and efficient transformation protocol for the grass Brachypodiumdistachyon. Plant Cell Reports 23: 751–758.
Demircan T, Akkaya MS. 2010. Virus induced gene silencing in
Brachypodium distachyon, a model organism for cereals. Plant Cell Tissueand Organ Culture 100: 91–96.
Devos KM. 2010. Grass genome organization and evolution. CurrentOpinion in Plant Biology 13: 139–145.
DOE, ED. 2006. Breaking the biological barriers to cellulosic ethanol: a jointresearch agenda. US Department of Energy, Office of Science and
Office of Energy Efficiency. URL http://genomicscience.energy.gov/
NewPhytologist Tansley review Review 345
� 2011 The Authors
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011) 191: 334–347
www.newphytologist.com

biofuels/2005workshop/2005low_lignocellulosic.pdf [accessed 29 April
2011].
Drader T, Kleinhofs A. 2010. A synteny map and disease resistance gene
comparison between barley and the model monocot Brachypodiumdistachyon. Genome 53: 406–417.
Draper J, Mur LAJ, Jenkins G, Ghosh-Biswas GC, Bablak P, Hasterok R,
Routledge APM. 2001. Brachypodium distachyon. A new model system
for functional genomics in grasses. Plant Physiology 127: 1539–1555.
Dubcovsky J, Dvorak J. 2007. Genome plasticity a key factor in the success
of polyploid wheat under domestication. Science 316: 1862–1866.
Ecker JR. 2002. A sequence-indexed library of insertion mutations in the
Arabidopsis genome. Plant Physiology (Rockville) 129: 405–406.
Edwards D, Batley J. 2010. Plant genome sequencing: applications for
crop improvement. Plant Biotechnology Journal 8: 2–9.
Erayman M, Sandhu D, Sidhu D, Dilbirligi M, Baenziger PS, Gill KS.
2004. Demarcating the gene-rich regions of the wheat genome. NucleicAcids Research 32: 3546–3565.
Faricelli ME, Valarik M, Dubcovsky J. 2010. Control of flowering time and
spike development in cereals: the earliness per se Eps-1 region in wheat,
rice, and Brachypodium. Functional & Integrative Genomics 10: 293–306.
Febrer M, Goicoechea JL, Wright J, McKenzie N, Song X, Lin J, Collura
K, Wissotski M, Yu Y, Ammiraju JS et al. 2010. An integrated physical,
genetic and cytogenetic map of Brachypodium distachyon, a model
system for grass research. PLoS One 18: e13461.
Filiz E, Ozdemir BS, Budak F, Vogel JP, Tuna M, Budak H. 2009.
Molecular, morphological, and cytological analysis of diverse
Brachypodium distachyon inbred lines. Genome 52: 876–890.
Gale MD, Rees H. 1970. Genes controlling chiasma frequency in
Hordeum. Heredity 25: 3 93–410.
Garvin DF, Gu YQ, Hasterok R, Hazen SP, Jenkins G, Mockler TC,
Mur LAJ, Vogel JP. 2008. Development of genetic and genomic
research resources for Brachypodium distachyon, a new model system for
grass crop research. Crop Science 48: S69–S84.
Garvin DF, McKenzie N, Vogel JP, Mockler TC, Blankenheim ZJ,
Wright J, Cheema JJS, Dicks J, Huo NX, Hayden DM et al. 2010. An
SSR-based genetic linkage map of the model grass Brachypodiumdistachyon. Genome 53: 1–13.
Gomez LD, Bristow JK, Statham ER, McQueen-Mason SJ. 2008a.
Analysis of saccharification in Brachypodium distachyon stems under mild
conditions of hydrolysis. Biotechnology for Biofuels 1: 15.
Gomez LD, Steele-King CG, McQueen-Mason SJ. 2008b. Sustainable
liquid biofuels from biomass: the writing’s on the walls. New Phytologist178: 473–485.
Griffiths S, Sharp R, Foote TN, Bertin I, Wanous M, Reader S, Colas I,
Moore G. 2006. Molecular characterization of Ph1 as a major
chromosome pairing locus in polyploid wheat. Nature 439: 749–752.
Gu YQ, Wanjugi H, Coleman-Derr D, Kong X, Anderson OD. 2010.
Conserved globulin gene across eight grass genomes identify
fundamental units of the loci encoding seed storage proteins. Functional& Integrative Genomics 10: 111–122.
Hartl J, Herrmann R. 2009. Do they always say no? German consumers
and second-generation GM foods. Agricultural Economics 40: 551–560.
Hasterok R, Draper J, Jenkins G. 2004. Laying the cytotaxonomic
foundations of a new model grass, Brachypodium distachyon (L.) Beauv.
Chromosome Research 12: 397–403.
Hasterok R, Marasek A, Donnison IS, Armstead I, Thomas A, King IP,
Wolny E, Idziak D, Draper J, Jenkins G. 2006. Alignment of the
genomes of Brachypodium distachyon and temperate cereals and grasses
using bacterial artificial chromosome landing with fluorescence in situhybridization. Genetics 173: 349–362.
Holzberg S, Brosio P, Gross C, Pogue GP. 2002. Barley stripe mosaic virus-
induced gene silencing in a monocot plant. Plant Journal 30: 315–327.
Hong SY, Lee S, Seo PJ, Yang MS, Park CM. 2010. Identification and
molecular characterization of a Brachypodium distachyon GIGANTEA
gene: functional conservation in monocot and dicot plants. PlantMolecular Biology 72: 485–497.
Huo NX, Lazo GR, Vogel JP, You FM, Ma YQ, Hayde DM, Coleman-
Derr D, Hill TA, Dvorak J, Anderson OD et al. 2008. The nuclear
genome of Brachypodium distachyon: analysis of BAC end sequences.
Functional & Integrative Genomics 8: 135–147.
Huo NX, Vogel JP, Lazo GR, You FM, Ma YQ, McMahon S, Dvorak J,
Anderson OD, Luo MC, Gu YQ. 2009. Structural characterization of
Brachypodium genome and its syntenic relationship with rice and
wheat. Plant Molecular Biology 70: 47–61.
Hurst A, John E. 1999. The biotic and abiotic changes associated with
Brachypodium pinnatum dominance in chalk grassland in south-east
England. Biological Conservation 88: 75–84.
International Brachypodium Initiative (IBI). 2010. Genome sequencing
and analysis of the model grass Brachypodium distachyon. Nature 463:
763–768.
Jakob SS, Ihlow A, Blattner FR. 2007. Combined ecological niche
modelling and molecular phylogeography revealed the evolutionary
history of Hordeum marinum (Poaceae) – niche differentiation, loss of
genetic diversity, and speciation in Mediterranean Quaternary refugia.
Molecular Ecology 16: 1713–1727.
Jenkins G, Hasterok R. 2007. BAC ‘landing’ on chromosomes of
Brachypodium distachyon for comparative genome alignment. NatureProtocols 2: 88–98.
Jenkins G, Mikhailova E, Phillips D, Hasterok R, Draper J, Jones N.
2005. Models and meiosis in the ‘omics era. In: Zwierzykowski Z,
Kosmala A, eds. Recent advances in genetics and breeding of grasses.Poznan, Poland: Institute of Plant Genetics PAS, 97–104.
Jones N, Rees H. 1966. Chiasma frequencies and potential genetic
variability of Lolium populations. Nature 211: 432–433.
Khan MA, Stace CA. 1999. Breeding relationships in the genus
Brachypodium (Poaceae: Pooideae). Nordic Journal of Botany 19:
257–269.
Kumar S, Mohan A, Balyan HS, Gupta PK. 2009. Orthology between
genomes of Brachypodium, wheat and rice. BMC Research Notes 2: 93.
Langridge P, Paltridge N, Fincher G. 2006. Functional genomics of
abiotic stress tolerance in cereals. Briefings in Functional Genomics &Proteomics 4: 343–354.
Larre C, Penninck S, Bouchet B, Lollier V, Tranquet O, Denery-Papini
S, Guillon F, Rogniaux H. 2010. Brachypodium distachyon grain:
identification and subcellular localization of storage proteins. Journal ofExperimental Botany 61: 1771–1783.
Laudencia-Chingcuanco DL, Vensel WH. 2008. Globulins are the main
seed storage proteins in Brachypodium distachyon. Theoretical andApplied Genetics 117: 555–563.
Leung H, An GL. 2004. Rice functional genomics: large-scale gene
discovery and applications to crop improvement. Advances in Agronomy82: 55–111.
Li J, Ding J, Zhang W, Zhang YL, Tang P, Chen JQ, Tian DC, Yang
SH. 2010. Unique evolutionary pattern of numbers of gramineous
NBS-LRR genes. Molecular Genetics and Genomics 283: 427–438.
Liancourt P, Corcket E, Michalet R. 2005. Stress tolerance abilities and
competitive responses in a watering and fertilization field experiment.
Journal of Vegetation Science 16: 713–722.
Luo MC, Ma Y, You FM, Anderson OD, Kopecky D, Simkova H, Safar
J, Dolezel J, Gill B, McGuire PE, Dvorak J. 2010. Feasibility of
physical map construction from fingerprinted bacterial artificial
chromosome libraries of polyploid plant species. BMC Genomics 11:
122–130.
Luo MC, Thomas C, You FM, Hsiao J, Ouyang S, Buell CR, Malandro
M, McGuire PE, Anderson OD, Dvorak J. 2003. High-throughput
fingerprinting of bacterial artificial chromosomes using the snapshot
labeling kit and sizing of restriction fragments by capillary
electrophoresis. Genomics 82: 378–389.
346 Review Tansley reviewNewPhytologist
� 2011 The Authors
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011) 191: 334–347
www.newphytologist.com

Matts J, Jagadeeswaran G, Roe BA, Sunkar R. 2010. Identification of
microRNAs and their targets in switchgrass, a model biofuel plant
species. Journal of Plant Physiology 167: 896–904.
McCallum CM, Comai L, Greene EA, Henikoff S. 2000. Targeted
screening for induced mutations. Nature Biotechnology 18: 455–457.
Metzker ML. 2010. Sequencing technologies – the next generation. NatureReviews Genetics 11: 31–46.
Meyers BC, Kozik A, Griego A, Kuang HH, Michelmore RW. 2003.
Genome-wide analysis of NBS-LRR-encoding genes in Arabidopsis.
Plant Cell 15: 809–834.
Nei M, Tajima F. 1983. Maximum likelihood estimation of the number
of nucleotide substitutions for restriction sites data. Genetics 105:
207–216.
Nei M, Tajima F, Tateno Y. 1983. Accuracy of estimated
phylogenetic trees from molecular data. Journal of Molecular Evolution19: 153–170.
Newton AC, Akar T, Baresel JP, Bebeli PJ, Bettencourt E,
Bladenopoulos KV, Czembor JH, Fasoula DA, Katsiotis A, Koutis K
et al. 2010. Cereal landraces for sustainable agriculture. A review.
Agronomy for Sustainable Development 30: 237–269.
Olsen P, Lenk I, Jensen CS, Petersen K, Andersen CH, Didion T,
Nielsen KK. 2006. Analysis of two heterologous flowering genes in
Brachypodium distachyon demonstrates its potential as a grass model
plant. Plant Science 170: 1020–1025.
Opanowicz M, Vain P, Draper J, Parker D, Doonan JH. 2008.
Brachypodium distachyon: making hay with a wild grass. Trends in PlantScience 13: 172–177.
Pacak A, Geisler K, Jørgensen B, Barciszewska-Pacak M, Nilsson L,
Nielsen TH, Johansen E, Grønlund M, Jakobsen I, Albrechtsen M.
2010. Investigations of barley stripe mosaic virus as a gene silencing
vector in barley roots and in Brachypodium distachyon and oat. PlantMethods 6: 26–33.
Rees H. 1961. Genotypic control of chromosome form and behaviour.
Botanical Review 27: 288–318.
Rees H, Dale PJ. 1974. Chiasmata and variability in Lolium and Festucapopulations. Chromosoma 47: 335–351.
Revedin A, Aranguren B, Becattini R, Longo L, Marconi E, Lippi MM,
Skakun N, Sinitsyn A, Spiridonova E, Svoboda J. 2010. Thirty
thousand-year-old evidence of plant food processing. Proceedings of theNational Academy of Sciences of the United States of America, 107:
18815–18819.
Robertson IH. 1981. Chromosome-numbers in Brachypodium Beauv
(Gramineae). Genetica 56: 55–60.
Rosegrant MW, Cline SA. 2003. Global food security: challenges and
policies. Science 302: 1917–1919.
Routledge APM, Shelley G, Smith JV, Talbot NJ, Draper J, Mur LAJ.
2004. Magnaporthe grisea interactions with the model grass
Brachypodium distachyon closely resemble those with rice (Oryza sativa).
Molecular Plant Pathology 5: 253–265.
Saintenac C, Falque M, Martin OC, Paux E, Feuillet C, Sourdille P.
2009. Detailed recombination studies along chromosome 3B provide
new insights on crossover distribution in wheat (Triticum aestivum L.).
Genetics 181: 393–403.
Sanchez-Moran E, Armstrong SJ, Santos JL, Franklin FCH, Jones GH.
2002. Variation in chiasma frequency among eight accessions of
Arabidopsis thaliana. Genetics 162: 1415–1422.
Schenkelaars P. 2001. Uncertainty and reluctance: Europe and GM foods.
Biotechnology and Development Monitor 47: 16–19.
Schippmann U. 1991. Revision of the European species of the genus
Brachypodium Palisot De Beauvois Poaceae. Boissiera 45: 1–250.
Schwartz CJ, Doyle MR, Manzaneda AJ, Rey PJ, Mitchell-Olds T,
Amasino RM. 2010. Natural variation of flowering time and
vernalization responsiveness in Brachypodium distachyon. BioenergyResearch 3: 38–46.
Scofield SR, Huang L, Brandt AS, Gill BS. 2005. Development of a virus-
induced gene-silencing system for hexaploid wheat and its use in
functional analysis of the Lr21-mediated leaf rust resistance pathway.
Plant Physiology 138: 2165–2173.
Spielmeyer W, Singh RP, McFadden H, Wellings CR, Huerta-Espino J,
Kong X, Appels R, Lagudah ES. 2008. Fine scale genetic and physical
mapping using interstitial deletion mutants of Lr34 Yr18: a disease
resistance locus effective against multiple pathogens in wheat. Theoreticaland Applied Genetics 116: 481–490.
Stace CA. 2010. New flora of the British Isles, 3rd edn. Cambridge, UK:
Cambridge University Press.
Talame V, Bovina R, Sanguineti MC, Tuberosa R, Lundqvist U, Salvi S.
2008. TILLMore, a resource for the discovery of chemically induced
mutants in barley. Plant Biotechnology Journal 6: 477–485.
Thole V, Worland B, Wright J, Bevan MW, Vain P. 2010. Distribution
and characterization of more than 1000 T-DNA tags in the genome of
Brachypodium distachyon community standard line Bd21. PlantBiotechnology Journal 8: 734–747.
Turner A, Beales J, Faure S, Dunford RP, Laurie DA. 2005. The pseudo-
response regulator Ppd-H1 provides adaptation to photoperiod in
barley. Science 310: 1031–1034.
Unver T, Budak H. 2009. Conserved microRNAs and their targets in
model grass species Brachypodium distachyon. Planta 230: 659–669.
Vain P, Worland B, Thole V, McKenzie N, Alves SC, Opanowicz M,
Fish LJ, Bevan MW, Snape JW. 2008. Agrobacterium-mediated
transformation of the temperate grass Brachypodium distachyon(genotype Bd21) for T-DNA insertional mutagenesis. PlantBiotechnology Journal 6: 941.
Van Hulle S, Roldan-Ruiz I, Van Bockstaele E, Muylle H. 2010.
Functional analysis of genes involved in cell wall biosynthesis of the
model species Brachypodium distachyon to improve saccharification.
Sustainable Use of Genetic Diversity in Forage and Turf Breeding 5:
479–482.
Venter JC, Smith HO, Hood L. 1996. A new strategy for genomesequencing. Nature 381: 364–366.
Vogel J, Hill T. 2008. High-efficiency Agrobacterium-mediated
transformation of Brachypodium distachyon inbred line Bd21-3. PlantCell Reports 27: 471–478.
Vogel JP, Gu YQ, Twigg P, Lazo GR, Laudencia-Chingcuanco D, Hayden
DM, Donze TJ, Vivian LA, Stamova B, Coleman-Derr D. 2006. EST
sequencing and phylogenetic analysis of the model grass Brachypodiumdistachyon. Theoretical and Applied Genetics 113: 186–195.
Vogel JP, Tuna M, Budak H, Huo NX, Gu YQ, Steinwand MA. 2009.
Development of SSR markers and analysis of diversity in Turkish
populations of Brachypodium distachyon. BMC Plant Biology 9: 88–99.
Wyman CE. 2007. What is (and is not) vital to advancing
cellulosicethanol. Trends in Biotechnology 25: 153–157.
Wolny E, Hasterok R. 2009. Comparative cytogenetic analysis of the
genomes of the model grass Brachypodium distachyon and its close
relatives. Annals of Botany 104: 873–881.
Wolny E, Lesniewska K, Hasterok R, Langdon T. 2011. Compact
genomes and complex evolution in the genus Brachypodium.
Chromosoma 120: 199–212.
Xu GX, Ma H, Nei M, Kong HZ. 2009. Evolution of F-box genes in
plants: different modes of sequence divergence and their relationships
with functional diversification. Proceedings of the National Academy ofSciences, USA 106: 835–840.
Yang ZL, Ohlrogge JB. 2009. Turnover of fatty acids during natural
senescence of Arabidopsis, Brachypodium, and switchgrass and in
Arabidopsis beta-oxidation mutants. Plant Physiology 150: 1981–1989.
Yu GT, Cai XW, Harris MO, Gu YQ, Luo MC, Xu SS. 2009. Saturation
and comparative mapping of the genomic region harboring Hessian fly
resistance gene H26 in wheat. Theoretical and Applied Genetics 118:
1589–1599.
NewPhytologist Tansley review Review 347
� 2011 The Authors
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011) 191: 334–347
www.newphytologist.com