evaluation of macaca mulatta as a model for genotoxicity studies
TRANSCRIPT

E
VDa
b
c
d
a
ARRAA
KRMEHPCFLE
1
metcrtrttu
1d
Mutation Research 673 (2009) 21–28
Contents lists available at ScienceDirect
Mutation Research/Genetic Toxicology andEnvironmental Mutagenesis
journa l homepage: www.e lsev ier .com/ locate /gentoxCommuni ty address : www.e lsev ier .com/ locate /mutres
valuation of Macaca mulatta as a model for genotoxicity studies
asily N. Dobrovolskya,∗ , Joseph G. Shaddocka , Roberta A. Mittelstaedta , Mugimane G. Manjanathaa ,aishiro Miuraa,b, Makoto Uchikawac, Donald R. Mattisond, Suzanne M. Morrisa
US FDA, National Center for Toxicological Research, Division of Genetic and Reproductive Toxicology, 3900 NCTR Rd., HFT-120, Jefferson, AR 72079, USATeijin Pharma Ltd., Tokyo, JapanJapanese Red Cross, Tokyo Blood Center, Tokyo, JapanUS NIH, NICHD, Bethesda, MD 20852, USA
r t i c l e i n f o
rticle history:eceived 2 October 2008eceived in revised form 6 November 2008ccepted 10 November 2008vailable online 27 November 2008
eywords:hesus monkeyutation
NUPRTIG-Aloninglow cytometryymphocytes
a b s t r a c t
We have investigated the use of peripheral blood from the nonhuman primate (NHP) rhesus monkey(Macaca mulatta) as a model system for mutation detection. The rhesus monkey is metabolically closerto humans than most common laboratory animals, and therefore may be a relevant model for hazardidentification and human risk assessment. To validate the model, conditions were determined for invitro selection and expansion of 6-thioguanine-resistant (6-TGr) HPRT mutant and proaerolysin-resistant(ProAERr) PIG-A mutant lymphocytes from peripheral blood obtained by routine venipuncture. Also,flow cytometric methods were developed for the rapid detection of PIG-A mutant erythrocytes. The flowcytometric analysis of PIG-A mutant erythrocytes was based on enumerating cells deficient in surfacemarkers attached to the cellular membrane via glycosylphosphatidyl inositol (GPI) anchors. Mutant cellswere enumerated over an extended period of time in peripheral blood of male monkeys receiving dailydoses of the electrolyte replenisher PrangTM (a common carrier for oral delivery of drugs in NHPs), andin the blood of one male monkey treated with a single i.p. dose of 50 mg/kg of N-ethyl-N-nitrosourea at∼2 years of age and another similar injection at approximately 3.5 years of age. The spontaneous PIG-A
−6
rythrocytesand HPRT T-cell mutant frequency (MF) was low in animals receiving Prang (0–8 × 10 ), and treatmentwith ENU resulted in a clearly detectable increase in the frequency of ProAERr and 6-TGr lymphocytes(up to ∼28 × 10−6 and ∼30 × 10−6, respectively). Also, the ENU-treated animal had higher frequency ofGPI-deficient erythrocytes (46.5 × 10−6 in the treated animal vs. 7.8 ± 4.2 × 10−6 in control animals). Ourresults indicate that the rhesus monkey can be a valuable model for the identification of agents that mayimpact upon human health as mutagens and that the PIG-A gene can be a useful target for detection of
nd re
mutation in both white a. Introduction
Due to their cognitive and learning capabilities, nonhuman pri-ates (NHPs), for example, the rhesus monkey, are frequently
mployed for behavioral studies. However, rodent models, ratherhan NHP models, are most commonly utilized in mutation andarcinogenesis studies, mainly because of the shorter lifespan ofodents as well as the increased costs associated with the main-enance of NHPs. Nevertheless, the NHP model may be a moreelevant model for human mutagenesis and carcinogenesis due to
he closer evolutionary relationship between humans and the NHPshan between humans and rodents, and it may be advantageous tose NHPs in some situations.∗ Corresponding author. Tel.: +1 870 543 7549; fax: +1 870 543 7393.E-mail address: [email protected] (V.N. Dobrovolsky).
383-5718/$ – see front matter. Published by Elsevier B.V.oi:10.1016/j.mrgentox.2008.11.006
d blood cells.Published by Elsevier B.V.
An NHP-based model for mutation detection, for instance,may be useful for studying neurotropic agents/medications thatare suspected human mutagens, but are negative in other muta-tion detection models. The mutagenicity of such agents maybe studied in the NHP model at doses that are clinically rel-evant and trigger behavioral changes characteristic of humanpatients.
Unfortunately, little information is available on the frequencyand spectrum of either spontaneous or induced mutation in theNHP genome. One group used cynomolgus monkey as a model fordetection of mutation in limited trials [1–5]. We have used a closelyrelated species – the rhesus monkey (Macaca mulatta) – to deter-mine if the endogenous HPRT and PIG-A genes can be useful for
monitoring in vivo mutagenesis.The HPRT gene is well characterized; it has been used as thetarget for mutation detection in humans [6,7] and various lab-oratory animals [8,9]. The PIG-A gene is a novel target for thedetection of in vivo mutation. The X-linked PIG-A gene is involved

22 V.N. Dobrovolsky et al. / Mutation Research 673 (2009) 21–28
Fig. 1. Schematic diagrams of basic principles of PIG-A mutation detection. (A) Phenotypic differences between wild-type and PIG-A mutant cells. Upper half: in wild-typecells, GPI anchors are synthesized and retain coupled markers (e.g., CD48, CD55, CD59 and CD90) at the cell surface; normal precursors in bone marrow release normaldifferentiated cells into peripheral blood. Lower half: in PIG-A mutant cells, GPI anchors are not synthesized and specific markers are not expressed at the cell surface; themutant precursor cells release differentiated cells that lack GPI-anchored markers into peripheral blood. Note that other (e.g., transmembrane) surface markers are unaffectedby PIG-A mutation. PIG-A mutant cells (both RBCs and WBCs) can be detected rapidly by flow cytometry using appropriate labeling techniques. (B) Toxicity of proaerolysin( y binda d kill wo in a l
itiafbemtt[
ProAER) is potentiated through GPI anchors. Bacterial protoxin ProAER selectivelerolysin; aerolysin aggregates into multimers that form pores in the membrane anf ProAER. The frequency of PIG-A mutant ProAER-resistant cells can be determined
n the synthesis of glycosyl phosphatidyl inositol (GPI) moleculeshat anchor specific markers at the cell surface [10,11]. Mutationn the PIG-A gene renders the cell deficient for GPIs and GPI-nchored proteins [12]. Therefore, flow cytometry can be usedor the identification of PIG-A mutant cells that are characterizedy the absence of GPI-anchored markers otherwise ubiquitously
xpressed in wild-type cells (Fig. 1A) [13]. Alternatively, PIG-Autants can be clonally expanded in a medium containing the bac-erial toxin, proaerolysin (ProAER) (Fig. 1B) [14], in an assay similaro the clonal expansion of HPRT mutants with 6-thioguanine (6-TG)6].
s cell surface GPI anchors and gets proteolytically converted into ultimate toxinild-type cells. PIG-A mutant cells lack GPI anchors and are resistant to cytotoxicity
imiting dilution cloning assay.
2. Materials and methods
2.1. Animals and animal care
All animal procedures were approved by the NCTR Institutional Animal Care andUse Committee. Twelve free-ranging juvenile male rhesus monkeys of similar age(judged by their weight) were obtained from Alpha Genesis, Inc. (Yemassee, SC). Ten
were designated as untreated controls; they received twice daily 0.5 ml/kg doses ofthe electrolyte replenisher PrangTM (Bio-Serv; Frenchtown, NJ) by oral ingestion. Theanimal designated for ENU treatment was added to the experiment about 1.5 monthsafter the start of the Prang treatment of other animals. One additional animal wasdesignated as a reserve, and it did not receive any treatment. The control animalswere enrolled in daily behavior testing activities that are believed to be nongeno-
ation
to
2
icesJ
2
mVdcpVpppiaasAfB
2
iatpinu
2
ccPPBsPG
2
hDo(Tca
2
tH(b2a1gPw
V.N. Dobrovolsky et al. / Mut
oxic. Several additional animals of various ages were used at the preliminary stagef the experiment for optimizing conditions for culturing peripheral T-lymphocytes.
.2. Mutagen exposure
N-ethyl-N-nitrosourea (ENU; ISOPAC® cat. no.: N3385, ENU content approx-mately 65%; Sigma, St. Louis, MO) was reconstituted in warm water at aoncentration of 1% (weight/volume), 0.2 �m filter sterilized, and promptly deliv-red to one animal in a single i.p. injection of ∼50 mg/kg (e.g., 24.6 ml of the stockolution per 3.2 kg animal). Only animal #32 was treated with ENU two times: onune 30, 2006, and November 26, 2007.
.3. HPRT assay and PIG-A T-cell clonal assay
The lymphocyte HPRT assay was performed as described by Harbach [1,2] withinor variations. Blood (∼7 ml) was collected from each animal into heparinizedacutainerTM tubes by venipuncture and the lymphocyte fraction was isolated byensity gradient centrifugation in Histopaq 1077 tubes (Sigma). Growth medium forulturing of lymphocytes contained 20% of T-cell growth factor (TCGF) supplementroduced by human LAK cells (a generous gift from Dr. R.J. Albertini, University ofermont). The cells were primed for 48 h in the growth medium containing 1 �g/mlhytohaemagglutinin (PHA; Remel, Lenexa, KS), counted and dispersed into 96-welllates (96WPs) at the desired concentrations (4 cells/well for cloning efficiency (CE)lates, 2 plates per animal; 1 × 104 cells/well for selection plates, 6 plates per animal)
n the medium containing 0.125 �g/ml PHA. Each well with monkey lymphocyteslso contained 1 × 104 feeder cells (HPRT-deleted human TK6 lymphoblastoid cells,gift from Dr. Pat O’Neill, University of Vermont) lethally irradiated at 90 Gy. For
election of HPRT mutants, 6-TG (Sigma) was used at 2–4 �g/ml. For selection of PIG-mutants, the cells were plated into 96WPs at the same concentration as were used
or the HPRT assay, with the exception that ProAER (Protox Biochemicals, Victoria,C) was used as selecting agent at final concentration of 2 nM.
.4. Plate scoring
After 12–14 days of culture, positive wells on the 96 WPs were identifiedn a computerized, semi-automated procedure by adding the viability indicatorlamarBlue® (TREK Diagnostic Systems, Cleveland, OH) to the wells and scanninghe plates in a SpectraFluor® fluorescence reader (TECAN; Durham, NC) as describedreviously [15,16]. Positive wells on each plate were identified as having a 1.4-fold
ncrease in fluorescence over the background level on the same plate (in a typicalegative well). CEs for cultures with and without selection agents were calculatedsing the Poisson distribution. MF was calculated as [CEwith selection]/[CEwithout selection].
.5. HPRT cDNA sequencing
RNA was extracted directly from cells grown in individual wells of 96WPs; totalDNA was synthesized using an oligo-dT primer as described previously [16]. HPRTDNA was amplified in two rounds, first using primers P9 and P10, followed by nestedCR primers P18 and P15 [1,2]. The nested PCR product was sequenced using primers18, P15, and internal primers P2 and P3 on a 3130xl Genetic Analyzer (Appliediosystems, Foster City, CA) and the mutations were identified with SeqScape v2.5oftware (Applied Biosystems). Primer sequences: P9 CTGCTCCGCCACCGGCTTCC;10 GATAATTTTACTGGCGATGT; P18 CCTCCTCCTGAGCAGTCAGCC; P15 GGACTCCA-ATGTTTCC; P2 GCTCAAGGGGGGCTATAAAT; P3 GCCAGTGTCAATTATATCTT.
.6. Antibodies
FITC-labeled anti-human CD59 (clone p282(H19)) and PE-Cy5-labeled anti-uman CD45 (clone TU116) monoclonal antibodies were purchased from Becton,ickinson and Company (BD; Milpitas, CA). Anti-human glycophorin (GYP) mon-clonal antibody (clone CBC-2(2E6)) was described previously [17]. The antibodymouse IgG1 isotype) produced by the clone was purified and concentrated atokyo Blood Center using affinity chromatography on protein G carrier followingonventional techniques. The secondary detection molecule, PE-Cy5.5-labeled goatnti-mouse IgG1 polyclonal antibody, was purchased from Invitrogen (Carlsbad, CA).
.7. Flow cytometry
Flow cytometry of purified RBCs was performed on a FACSAria flow cytome-er (BD) at a rate of 10,000–20,000 events per second. The RBC fraction isolated byistopaq density gradient centrifugation was washed in phosphate buffered saline
PBS) and resuspended in a volume of PBS equal to the initial volume of collectedlood. 4 �l of RBC suspension were used in a labeling reaction in 200 �l of PBS,
0 �l of FITC-anti-human-CD59 antibody and 20 �l of PE-Cy5-anti-human-CD45ntibody. The labeling was performed at room temperature in the dark in a standard.5 ml microcentrifuge tube; after 1 h, the cells were sedimented by 5 min centrifu-ation at 500 × g and the pellet was resuspended in 1 ml of 1% paraformaldehyde inBS. The region occupied by CD59-negative PIG-A mutant cells on flow cytogramsas identified by running unlabeled cells.Research 673 (2009) 21–28 23
In addition, the positive identification of mutant RBCs was performed on severalsamples by incubation with anti-human GYP antibody, followed by the additionof the secondary PE-Cy5.5-labeled antibody, brief centrifugation and hybridizationwith FITC-anti human CD59 antibody. FITC fluorescence was measured on a PMTequipped with 530/30 nm filter, PE-Cy5 and PE-Cy5.5 fluorescence was measuredon a PMT equipped with 695/40 nm filter. Only the blue 488 nm laser was used forthe excitation of fluorophores.
3. Results
3.1. CE variability
Initial trials of limited dilution culturing of lymphocytes fromseveral older NHPs (3–9 years of age) indicated that the optimalconcentration of the TCGF supplement (supporting the highest CEof cells without selection agent in the medium) was 20%. Laterexperiments with younger animals indicated that the variabilityin lymphocyte CE among different animals was high, and that ahigh TCGF concentration does not necessarily increase CEs. Overthe extended time period of sampling (over 1.5 years), some ani-mals consistently exhibited low CEs, while others demonstratedconsistently high CEs in the culture medium with 20% TCGF. Someanimals showed variable CEs over time. CE ranged from 1% to26%.
3.2. HPRT MF
Relatively low HPRT MFs were consistently detected in thePrang-treated animals over the course of 18-months-long treat-ment (Fig. 2). Only the ENU-treated animal produced an elevatedHPRT MF after the first treatment and over all subsequent sam-pling time-points. Some control animals never produced a singleHPRT mutant clone over the duration of the study, while othersproduced only one mutant clone at several sampling times. For agiven number of detected 6-TGr clones, MF is inversely related tothe CE of the sample. This suggests that for an animal with low CE,a minimal variation in the number of 6-TGr clones may result in asubstantial variation in MF. In the cultures with zero 6-TGr clones(with subsequent zero HPRT MFs), a low limit for detection of MFcould be estimated with the assumption that at least 1 mutant clonewas found. With this adjustment, an “estimated non-zero” MF wasidentified for all samples. In several cases, the “estimated non-zero”HPRT MF was between 5 and 15 × 10−6. This value was higher thanthe MF calculated for the samples with actual 6-TGr clones, butlower than the peak MF determined for the ENU-treated animal.After the first ENU injection, a rapid increase in the HPRT lympho-cyte MF (up to 30 × 10−6 in about 4 months) was detected and wasfollowed by a decline in MF over the subsequent 12 months. Thesecond ENU treatment did not result in a significant change in theHPRT MF (at least at 2 months after the treatment).
3.3. ENU-induced HPRT mutation spectrum
The publicly available reference sequence for rhesus mon-key HPRT cDNA was partially computer predicted (e.g., GenBankaccession number DR773101). There was a substantial differencebetween the human HPRT cDNA sequence and the predictedsequence, specifically, at the 5′-end. We used human sequence-based primers P9, P10, P15, and P18 designed by Harbach for theamplification of HPRT cDNA of cynomolgus monkey [2]. Theseprimers reliably amplified DNA fragments of the expected size(especially after nested PCR) using cDNA template derived by
reverse transcription of total RNA from lymphocyte clones grown inindividual wells of CE 96WPs. The HPRT cDNA sequence of severalCE clones (presumably wild-type) was determined and found to beidentical to the HPRT sequence from cynomolgus monkey. Muta-tions were identified in all 6-TGr clones from which a cDNA product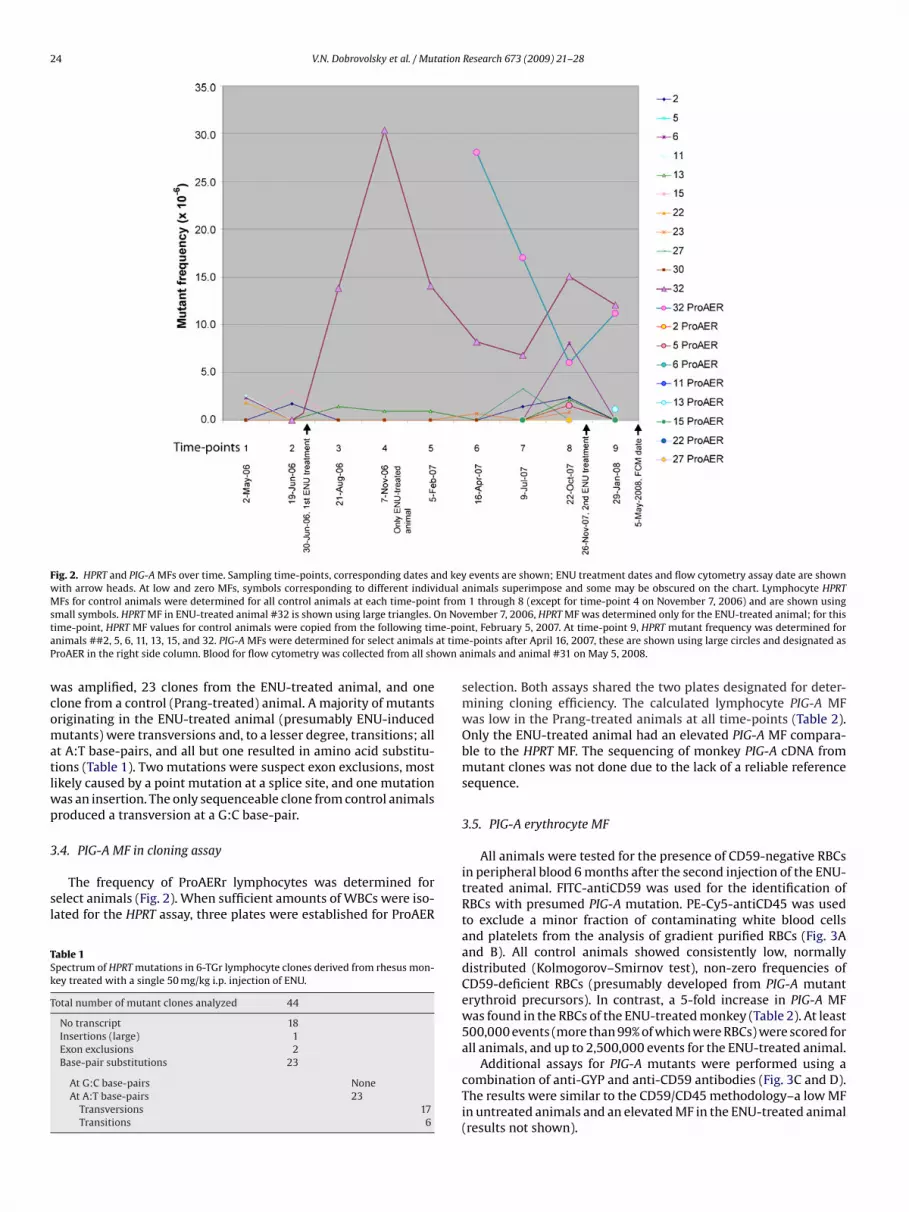
24 V.N. Dobrovolsky et al. / Mutation Research 673 (2009) 21–28
Fig. 2. HPRT and PIG-A MFs over time. Sampling time-points, corresponding dates and key events are shown; ENU treatment dates and flow cytometry assay date are shownwith arrow heads. At low and zero MFs, symbols corresponding to different individual animals superimpose and some may be obscured on the chart. Lymphocyte HPRTMFs for control animals were determined for all control animals at each time-point from 1 through 8 (except for time-point 4 on November 7, 2006) and are shown usings n Not e-poa at timP own a
wcomatlwp
3
sl
TSk
T
mall symbols. HPRT MF in ENU-treated animal #32 is shown using large triangles. Oime-point, HPRT MF values for control animals were copied from the following timnimals ##2, 5, 6, 11, 13, 15, and 32. PIG-A MFs were determined for select animalsroAER in the right side column. Blood for flow cytometry was collected from all sh
as amplified, 23 clones from the ENU-treated animal, and onelone from a control (Prang-treated) animal. A majority of mutantsriginating in the ENU-treated animal (presumably ENU-inducedutants) were transversions and, to a lesser degree, transitions; all
t A:T base-pairs, and all but one resulted in amino acid substitu-ions (Table 1). Two mutations were suspect exon exclusions, mostikely caused by a point mutation at a splice site, and one mutationas an insertion. The only sequenceable clone from control animalsroduced a transversion at a G:C base-pair.
.4. PIG-A MF in cloning assay
The frequency of ProAERr lymphocytes was determined forelect animals (Fig. 2). When sufficient amounts of WBCs were iso-ated for the HPRT assay, three plates were established for ProAER
able 1pectrum of HPRT mutations in 6-TGr lymphocyte clones derived from rhesus mon-ey treated with a single 50 mg/kg i.p. injection of ENU.
otal number of mutant clones analyzed 44
No transcript 18Insertions (large) 1Exon exclusions 2Base-pair substitutions 23
At G:C base-pairs NoneAt A:T base-pairs 23
Transversions 17Transitions 6
vember 7, 2006, HPRT MF was determined only for the ENU-treated animal; for thisint, February 5, 2007. At time-point 9, HPRT mutant frequency was determined fore-points after April 16, 2007, these are shown using large circles and designated asnimals and animal #31 on May 5, 2008.
selection. Both assays shared the two plates designated for deter-mining cloning efficiency. The calculated lymphocyte PIG-A MFwas low in the Prang-treated animals at all time-points (Table 2).Only the ENU-treated animal had an elevated PIG-A MF compara-ble to the HPRT MF. The sequencing of monkey PIG-A cDNA frommutant clones was not done due to the lack of a reliable referencesequence.
3.5. PIG-A erythrocyte MF
All animals were tested for the presence of CD59-negative RBCsin peripheral blood 6 months after the second injection of the ENU-treated animal. FITC-antiCD59 was used for the identification ofRBCs with presumed PIG-A mutation. PE-Cy5-antiCD45 was usedto exclude a minor fraction of contaminating white blood cellsand platelets from the analysis of gradient purified RBCs (Fig. 3Aand B). All control animals showed consistently low, normallydistributed (Kolmogorov–Smirnov test), non-zero frequencies ofCD59-deficient RBCs (presumably developed from PIG-A mutanterythroid precursors). In contrast, a 5-fold increase in PIG-A MFwas found in the RBCs of the ENU-treated monkey (Table 2). At least500,000 events (more than 99% of which were RBCs) were scored for
all animals, and up to 2,500,000 events for the ENU-treated animal.Additional assays for PIG-A mutants were performed using acombination of anti-GYP and anti-CD59 antibodies (Fig. 3C and D).The results were similar to the CD59/CD45 methodology–a low MFin untreated animals and an elevated MF in the ENU-treated animal(results not shown).

V.N. Dobrovolsky et al. / Mutation Research 673 (2009) 21–28 25
Fig. 3. Typical dot-plot cytograms and histograms of surface marker expression analysis in RBC fraction of peripheral blood from rhesus monkeys. (A and B) Staining withFITC-anti-CD59 and PE-Cy5-anti-CD45. 500,000 events were processed, out of which more than 99.5% fall into the singlets gate, almost all of which were CD45-negative(close to 99.8%); the gate for CD59-negative mutants was chosen to cover approximately 90% of unstained cells (flow cytometric data from unstained samples are not shown);when calculating mutant frequency the number of cells in the gate for mutants was divided by 500,000 and by 0.9. (A) Animal #2, three cells in the gate for 90% of mutants;calculated frequency of CD59-negative RBCs is 6.7 × 10−6. (B) ENU-treated animal #32, 23 cells in the gate for 90% of mutants; calculated frequency of CD59-negative RBCs is46.5 × 10−6 (average of several separate 500,000-cell assays). (C and D) Staining with FITC-anti-CD59, anti-GYP and PE-Cy5.5-anti-mouse-IgG1. 500,000 events were processed.T lot cyts r PE-CG (e.g., it orrec
4
tsc
he diagonally elongated shape of the population of double-stained cells on dot-pignificant fraction of cells after staining may be present in aggregates (with higheYP-positive cells (potential mutants) was consistently lower in untreated animals
o distinctly isolate and enumerate a single-cell fraction of RBCs not permitted the c
. Discussion
The identification and characterization of genotoxicity usinghe most relevant biological models is of utmost importance forcreening drugs, food additives, and environmental pollutants forarcinogenic potential. For this purpose many in vivo assays have
ograms and the two-peak distribution of labeled cells on histograms suggest thaty5.5 and FITC signals for the aggregates). Although the number of CD59-negativen animal #31, panel C) than in the ENU-treated animal #32 (panel D), the inabilityt calculation of the frequency of CD59-negative RBCs.
been developed, most notably the assay for detection of HPRT
mutant lymphocytes in human peripheral blood [6]. Later the assaywas adopted for detection of mutants among spleen cells in labrodents [8,9]. Only limited use of the HPRT assay has been made inNHPs despite the fact that rhesus monkeys are frequently used inbiomedical research. In the present study we have demonstrated
26 V.N. Dobrovolsky et al. / Mutation Research 673 (2009) 21–28
Table 2Comparison of frequencies of CD59-deficient RBCs and ProAER-resistant (ProAERr) T-lymphocytes in peripheral blood of individual animals as determined by flow cytometryand clonal assay, respectively. RBCs that lack CD59 at the surface and ProAERr lymphocytes are presumed to originate from PIG-A mutant hematopoietic stem cells.
Animal Treatment/status Frequency of CD59-deficientRBCs (×10−6)
Frequency of ProAERr T-cells determined in clonal assays onvarious dates (×10−6)
May 2008 April 2007 July 2007 October 2007 January 2008
2 Prang/control 6.7 – – – 05 Prang/control 13.3 – – 1.5 06 Prang/control 13.3 – – – 011 Prang/control 2.2 – – – 013 Prang/control 6.7 – – – 1.115 Prang/control 13.3 – 0 – 022 Prang/control 4.4 – – 0 –23 Prang/control 8.9 – – – –27 Prang/control 2.2 – – 0 –30 Prang/control 8.9 – – – –31 Extra 5.5 – – – –
Average and standard deviation for the above group 7.8 ± 4.2
32 ENU 46.5 27.8 17 6 12
“N y in Mc Secti
ttcbass[tassiigwutdtnfOsm
mTdolhibastb
dfdt
–” Means the assay has not been done for this animal on this date.ote: T-cell ProAER selection clonal assay in January 2008 and flow cytometry assalonal assays on animal #32 were performed after a single treatment with ENU (see
hat the HPRT lymphocyte MF in response to ENU treatment ofhe rhesus monkey is similar to that of the evolutionary closeynomolgous monkey. Although these are two different species,oth have identical HPRT cDNA protein coding sequences. The aver-ge lymphocyte CE was higher in cynomolgous monkeys, thoughubstantial variability between individual animals and within theame animal at different time-points was observed for both species4]. The kinetics of induction of HPRT mutant lymphocytes afterreatment with ENU were practically identical between rhesusnd cynomolgous species, an initial increase in the HPRT MF andubsequent decline, which suggests that mechanisms of negativeelection against HPRT mutants are similar in both species. Surpris-ngly, the second treatment with the same dose of ENU did not resultn a second spike in the HPRT MF as was described for cynomol-ous monkeys [4]. It remains to be determined if those differencesere due to individual variations in the small number of animalssed or due to species differences. The spectrum of HPRT muta-ion determined for monkeys was similar to ENU-induced mutationetermined for the cynomolgous monkey and other species (mostlyransversions and transitions at A:T base-pairs) [1,18–21]. This isot surprising for the direct-acting alkylating agent, ENU, which
orms not only promutagenic O6-ethylguanine, but also O2- and4-ethylthymine DNA adducts. The latter two adducts are more per-istent than O6-ethylguanine due to a lower rate of repair and areore likely to cause mutation [22].The value of the NHP HPRT mutation detection assay is under-
ined by technical issues in the procedure. An expansion of a single-cell into a visibly identifiable viable clone requires an extraor-inary combination of growth factors. Serum-free formulationsf growth medium described for successful expansion of human
ymphocytes [23] proved to be ineffective for monkey cells. Theuman T-cell growth factor is no longer available commercially; and
n-house production of the growth factor requires technical capa-ilities that may be beyond the scope of most laboratories [24]. Itlso should be noted that monkey lymphocyte cultures are moreusceptible to microbial contamination than human or rodent cul-ures. It is more difficult to maintain an aseptic environment duringlood collection from monkeys.
Because of the challenges described above, it is important toevelop an assay where mutant cells could be rapidly discriminatedrom wild-type. Thus, the second part of the study describes theevelopment of a novel assay for rapid detection of in vivo muta-ion in the monkey endogenous PIG-A gene using flow cytometry. In
ay 2008 on animal #32 were performed after two treatments with ENU. All otheron 2).
humans, mutation in the PIG-A gene of blood stem cells results in anacquired genetic disorder, paroxysmal nocturnal hemoglobinurea(PNH) [25,26]. Diagnosis of the disease relies on the detection of asignificant fraction (up to 99% [27]) of GPI-deficient hematopoieticcells of all lineages. Currently, molecular diagnostics and charac-terization of PNH is performed by the flow cytometric detectionof cells deficient in GPI-anchored markers using appropriate anti-bodies [28] or by the detection of GPI-deficient cells using FLAER,a fluorescently labeled genetically modified bacterial toxin ProAER[29]. Unmodified ProAER is used to suppress wild-type cells andpromote the selective clonal expansion of PIG-A mutants for sub-sequent sequence analysis of PIG-A gene mutation [14,30]. UsingProAER for selective expansion of PIG-A mutant nucleated cellsit was demonstrated that, in healthy individuals, cells with PNHphenotype are present at low frequencies [31,32]. We report a sim-ilar finding in rhesus monkey: a low frequency of ProAER-resistantlymphocytes in control animals and an increased frequency in theENU-treated animal, a frequency similar to the frequency of HPRTmutant 6-TGr lymphocytes.
In studies aimed at understanding the clinical progression of thedisease, it was noted that the location of the PIG-A gene on the X-chromosome, accompanied by the ability to discriminate mutantcells by phenotype, could lead to the use of this system as a highthroughput mutation assay [33]. Theoretically, the Pig-A mutationdetection assay can be developed for most mammalian species dueto the conservative nature of GPI synthesis and the unique positionof Pig-A gene on the X-chromosome.
Our strategy for the identification of PIG-A mutants in theperipheral blood of rhesus monkeys was partially inspired by thesuccessful implementation of the Pig-A mutation assay in rats in ourlaboratory [34,35] and by others [36]. PIG-A mutants were identi-fied by the absence of GPI-anchored CD59 marker. The CD59 markeris ubiquitously expressed on cells of blood and solid tissues, and inhumans, it is the erythrocyte marker suggested for the most reliablediagnosis of PNH [37].
During the development of the monkey FCM assay, a consider-able difference between the rat and the monkey RBC Pig-A assayswas noted. In rats, whole blood could be used for labeling and sub-
sequent flow cytometry. In monkeys, the fractionation of blood bydensity gradient centrifugation was required. Without density cen-trifugation, whole blood produced noisy data, and the frequency ofCD59-negative events was unusually high. One explanation for thisis that the anti-human CD45 and CD59 that were used to detect
ation
mcaCrtmnioftow(hc
awitssmrbtayto
lmkiiovt0mtsaitbcdimmmiouo
mpttdm
V.N. Dobrovolsky et al. / Mut
utants in monkeys may lack the specificity associated with humanells. In our rodent studies, a low expression of CD59 was found insubstantial fraction (5–10%) of CD45+ WBCs. In the rat assay, theD45+ cells were eliminated (using highly specific anti-rat CD45),esulting in a purer population for analysis of nonfluorescent cellshat are presumed to be Pig-A mutant RBCs [34]. Essentially, this
ay be considered as a limitation of the assay. To confirm that theseonfluorescent cells are truly RBCs, either an additional positive
dentification marker needs to be used or the cells should be sortedut and visually identified under microscope (which has been doneor cells from high-dose ENU-treated rats, unpublished results). Inhe case of rhesus monkeys, the exact cell-type specific reactivityf anti-human CD45 has not been comprehensively detailed; so,hen using whole blood, the probability of scoring nucleated cells
that are naturally low in expression and cross-reactivity with anti-uman antibodies to both CD45 and CD59) as PIG-A mutant RBCsannot be excluded.
Using a positive label for RBCs (anti-GYP antibody), we wereble to confirm that the frequency of CD59-negative erythrocytesas increased in the ENU-treated monkey. But MF quantitation
n control and treated animals was uncertain due to aggrega-ion of erythrocytes in the presence of anti-GYP. Earlier studiesuggested that the reaction of antibody with glycophorin causestructural changes in the surface of RBCs [38], which may pro-ote aggregation. A concern with the use of anti-GYP is that the
are CD59-negative mutants will aggregate with CD59+ RBCs ande classified as non-mutants. In excessively aggregated sampleshe PIG-A MF may be artificially underestimated as a result. Thenti-CD59 and anti-GYP combination was not useful for the anal-sis of whole blood samples, probably due to the same reasonshat anti-CD59 and anti-CD45 were not efficient for the analysisf unprocessed blood.
In addition, the anti-GYP antibody used in this study was notabeled with a fluorescent dye; fluorescent polyclonal goat-anti
ouse IgG was used as a secondary antibody for labeling mon-ey RBCs, which required additional centrifugation/washing stepsn labeling protocol. These extra steps could adversely affect stain-ng the rare population of PIG-A mutant RBCs. The expected fractionf mutant RBCs in Prang- or ENU-treated animals was low by con-entional flow cytometry standards; even in animals treated withhe most powerful mutagens, the mutant frequency rarely exceeds.1% (1 × 10−3, or 1000 mutants per 1,000,000 cells). Any additionalanipulation with the cells, such as mechanical stress (e.g., cen-
rifugation) or pretreatment (e.g., to allow penetration of specifictains inside the cells), should be avoided in performing the PIG-Assay. The GPI anchor that holds markers used in the assay (CD59n the case of rats and monkeys) is weak; essentially, bulky pro-eins are retained at the outer surface of the cellular membraney two hydrophobic lipid tails without transmembrane domains orovalent links. Compromising the integrity of the cell membraneue to mechanical stress (e.g., induced by centrifugation or react-
ng antibodies with surface markers, not necessarily GPI-anchoredarkers, e.g. glycophorin [38]) may cause the loss of GPI-anchoredarkers. Also, changes in the medium can affect GPI-anchoredarkers. As an example, preservation of blood in cold methanol (as
n storing RBCs for the micronucleus assay) increases the frequencyf CD59-deficient cells dramatically, so that the marker becomesseless for the detection of Pig-A mutation in rats (unpublishedbservation).
In conclusion, we have developed assays for detecting in vivoutation in the endogenous HPRT and PIG-A genes in the lym-
hocytes and erythrocytes of rhesus monkeys. In control animals,he PIG-A MF in RBCs was higher than the PIG-A MF in T-cells andhan the HPRT MF in T-cells. This suggests that the flow cytometricetection of mutation may be more sensitive than the detection ofutation using cloning at limiting dilution, primarily because no
[
Research 673 (2009) 21–28 27
long-term culture of cells is needed for performing the flow cyto-metric assay. The values for induced MFs in the HPRT and PIG-Agenes in T-lymphocytes of the ENU-treated animal were similar,while the RBC PIG-A mutant frequency was slightly higher and wasdetectable long past the treatment, when mutation in lymphocyteswas no longer observable. This difference in the kinetics of mutationinduction and manifestation may suggest that bone marrow cellsthat subsequently produced the mutant cell types may undergovariable degrees of negative selection over time, and that the PIG-A mutant RBC precursors may experience the least selection. Thepersistence of ENU-induced PIG-A mutant RBCs in the peripheralblood of rhesus monkey suggests that RBCs could be a useful targetfor identification of genotoxic hazards using fast and cost-efficientflow cytometry methodology.
Conflict of interest
No conflicts of interest to declare.
Acknowledgements
We would like to thank the following people for their importantcontributions to the study. We thank Dr. R. Albertini for provid-ing human T-cell growth factor supplement, Dr. Patrick O’Neill forproviding HPRT-deficient human lymphoblastoid TK6 cell line. Spe-cial thanks to employees of animal care facility for multiple bloodsamplings of experimental animals. Also we thank Drs. CharlotteHotchkiss and Jeff Carraway for administering ENU and monitoringhealth of monkey colony at NCTR. This work was partially supportedby the NICHD through the U.S. Congressional Best Pharmaceuticalsfor Children Act of 2002. The views presented in this article do notnecessary reflect those of the U.S. Food and Drug Administration.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.mrgentox.2008.11.006.
References
[1] P.R. Harbach, A.L. Filipunas, Y. Wang, C.S. Aaron, DNA sequence analysis of spon-taneous and N-ethyl-N-nitrosourea-induced hprt mutations arising in vivo incynomolgus monkey T-lymphocytes, Environ. Mol. Mutagen. 20 (1992) 96–105.
[2] P.R. Harbach, S.S. Mattano, D.M. Zimmer, Y. Wang, C.S. Aaron, DNA sequenceanalysis of spontaneous hprt mutations arising in vivo in cynomolgus monkeyT-lymphocytes, Environ. Mol. Mutagen. 26 (1995) 218–225.
[3] P.R. Harbach, S.S. Mattano, D.M. Zimmer, A.L. Filipunas, Y. Wang, C.S. Aaron, DNAsequence analysis of hprt mutants persisting in peripheral blood of cynomolgusmonkeys more than two years after ENU treatment, Environ. Mol. Mutagen. 33(1999) 42–48.
[4] D.M. Zimmer, C.S. Aaron, In vivo mutagenesis in the cynomolgus monkey:time course of HPRT mutant frequency at long time points following ethyl-nitrosourea exposure, Environ. Mol. Mutagen. 29 (1997) 117–123.
[5] D.M. Zimmer, P.R. Harbach, S.S. Mattano, R.L. Yu, W.B. Mattes, C.S. Aaron, Lackof response to multiple genotoxic agents at the hprt locus in peripheral bloodT-lymphocytes of cynomolgus monkeys, Environ. Mol. Mutagen. 33 (1999)123–131.
[6] R.J. Albertini, K.L. Castle, W.R. Borcherding, T-cell cloning to detect the mutant6-thioguanine-resistant lymphocytes present in human peripheral blood, Proc.Natl. Acad. Sci. U.S.A. 79 (1982) 6617–6621.
[7] J.P. O’Neill, L.M. Sullivan, R.J. Albertini, In vitro induction, expression and selec-tion of thioguanine-resistant mutants with human T-lymphocytes, Mutat. Res.240 (1990) 135–142.
[8] I.M. Jones, K. Burkhart-Schultz, A.V. Carrano, A method to quantify spontaneousand in vivo induced thioguanine-resistant mouse lymphocytes, Mutat. Res. 147(1985) 97–105.
[9] A. Aidoo, S.M. Morris, D.A. Casciano, Development and utilization of the rat
lymphocyte hprt mutation assay, Mutat. Res. 387 (1997) 69–88.10] K.Y. Hwa, Glycosyl phosphatidylinositol-linked glycoconjugates: structure,biosynthesis and function, Adv. Exp. Med. Biol. 491 (2001) 207–214.
[11] A. Tiede, I. Bastisch, J. Schubert, P. Orlean, R.E. Schmidt, Biosynthesis of gly-cosylphosphatidylinositols in mammals and unicellular microbes, Biol. Chem.380 (1999) 503–523.

2 ation
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
8 V.N. Dobrovolsky et al. / Mut
12] N. Inoue, Y. Murakami, T. Kinoshita, Molecular genetics of paroxysmal nocturnalhemoglobinuria, Int. J. Hematol. 77 (2003) 107–112.
13] S.J. Richards, A.C. Rawstron, P. Hillmen, Application of flow cytometry tothe diagnosis of paroxysmal nocturnal hemoglobinuria, Cytometry 42 (2000)223–233.
14] R.A. Brodsky, G.L. Mukhina, K.L. Nelson, T.S. Lawrence, R.J. Jones, J.T. Buck-ley, Resistance of paroxysmal nocturnal hemoglobinuria cells to the glycosyl-phosphatidylinositol-binding toxin aerolysin, Blood 93 (1999) 1749–1756.
15] V.N. Dobrovolsky, J.G. Shaddock, R.H. Heflich, 7, 12-dimethylbenz[a]anthracene-induced mutation in the Tk gene of Tk+/− mice: automatedscoring of lymphocyte clones using a fluorescent viability indicator, Environ.Mol. Mutagen. 36 (2000) 283–291.
16] V.N. Dobrovolsky, J.G. Shaddock, R.H. Heflich, Analysis of in vivo mutation in theHprt and Tk genes of mouse lymphocytes, in: P. Keohavong, S.G. Grant (Eds.),Molecular Toxicolgy Protocols, Humana Press Inc., 2004.
17] A. Blancher, W.W. Socha, M.E. Reid, Reactivity of anti-glycophorin monoclonalantibodies (Mabs) in tests with red cells of non-human primates, Transfus. Clin.Biol. 4 (1997) 81–85.
18] J.G. Jansen, G.R. Mohn, H. Vrieling, C.M. van Teijlingen, P.H. Lohman, A.A. vanZeeland, Molecular analysis of hprt gene mutations in skin fibroblasts of ratsexposed in vivo to N-methyl-N-nitrosourea or N-ethyl-N-nitrosourea, CancerRes. 54 (1994) 2478–2485.
19] V.N. Dobrovolsky, T. Chen, R.H. Heflich, Molecular analysis of in vivo mutationsinduced by N-ethyl-N-nitrosourea in the autosomal Tk and the X-linked Hprtgenes of mouse lymphocytes, Environ. Mol. Mutagen. 34 (1999) 30–38.
20] R.A. Mittelstaedt, B.A. Smith, R.H. Heflich, Analysis of in vivo mutation inducedby N-ethyl-N-nitrosourea in the hprt gene of rat lymphocytes, Environ. Mol.Mutagen. 26 (1995) 261–269.
21] T.R. Skopek, V.E. Walker, J.E. Cochrane, T.R. Craft, N.F. Cariello, Mutationalspectrum at the Hprt locus in splenic T cells of B6C3F1 mice exposed to N-ethyl-N-nitrosourea, Proc. Natl. Acad. Sci. U.S.A. 89 (1992) 7866–7870.
22] S.M. Bronstein, T.R. Skopek, J.A. Swenberg, Efficient repair of O6-ethylguanine,but not O4-ethylthymine or O2-ethylthymine, is dependent upon O6-alkylguanine-DNA alkyltransferase and nucleotide excision repair activities inhuman cells, Cancer Res. 52 (1992) 2008–2011.
23] S.M. Hou, F.J. Van Dam, F. de Zwart, C. Warnock, M. Mognato, J. Turner, N. Pod-lutskaja, A. Podlutsky, R. Becker, Y. Barnett, C.R. Barnett, L. Celotti, M. Davies, E.Huttner, B. Lambert, A.D. Tates, Validation of the human T-lymphocyte cloningassay—ring test report from the EU concerted action on HPRT mutation (EUC-
AHM), Mutat. Res. 431 (1999) 211–221.24] H. Inouye, J.A. Hank, B.J. Alter, F.H. Bach, TCGF production for cloning and growthof functional human T lymphocytes, Scand. J. Immunol. 12 (1980) 149–154.
25] M. Bessler, P.J. Mason, P. Hillmen, T. Miyata, N. Yamada, J. Takeda, L. Luzzatto, T.Kinoshita, Paroxysmal nocturnal haemoglobinuria (PNH) is caused by somaticmutations in the PIG-A gene, EMBO J. 13 (1994) 110–117.
[
Research 673 (2009) 21–28
26] J. Nishimura, Y. Murakami, T. Kinoshita, Paroxysmal nocturnal hemoglobinuria:an acquired genetic disease, Am. J. Hematol. 62 (1999) 175–182.
27] J. Piedras, X. Lopez-Karpovitch, Flow cytometric analysis of glycosyl-phosphatidyl-inositol-anchored proteins to assess paroxysmal nocturnalhemoglobinuria clone size, Cytometry 42 (2000) 234–238.
28] S.E. Hall, W.F. Rosse, The use of monoclonal antibodies and flow cytometryin the diagnosis of paroxysmal nocturnal hemoglobinuria, Blood 87 (1996)5332–5340.
29] R.A. Brodsky, G.L. Mukhina, S. Li, K.L. Nelson, P.L. Chiurazzi, J.T. Buckley, M.J.Borowitz, Improved detection and characterization of paroxysmal nocturnalhemoglobinuria using fluorescent aerolysin, Am. J. Clin. Pathol. 114 (2000)459–466.
30] K.L. Nelson, S.M. Raja, J.T. Buckley, The glycosylphosphatidylinositol-anchoredsurface glycoprotein Thy-1 is a receptor for the channel-forming toxin aerolysin,J. Biol. Chem. 272 (1997) 12170–12174.
31] R. Hu, G.L. Mukhina, S. Piantadosi, J.P. Barber, R.J. Jones, R.A. Brodsky, PIG-Amutations in normal hematopoiesis, Blood 105 (2005) 3848–3854.
32] R.E. Ware, C.V. Pickens, C.M. DeCastro, T.A. Howard, Circulating PIG-A mutantT lymphocytes in healthy adults and patients with bone marrow failure syn-dromes, Exp. Hematol. 29 (2001) 1403–1409.
33] D.J. Araten, K. Nafa, K. Pakdeesuwan, L. Luzzatto, Clonal populations ofhematopoietic cells with paroxysmal nocturnal hemoglobinuria genotype andphenotype are present in normal individuals, Proc. Natl. Acad. Sci. U.S.A. 96(1999) 5209–5214.
34] D. Miura, V.N. Dobrovolsky, Y. Kasahara, Y. Katsuura, R.H. Heflich, Developmentof an in vivo gene mutation assay using the endogenous Pig-A gene: I. Flowcytometric detection of CD59-negative peripheral red blood cells and CD48-negative spleen T-cells from the rat, Environ. Mol. Mutagen. (2008).
35] D. Miura, V.N. Dobrovolsky, R.A. Mittelstaedt, Y. Kasahara, Y. Katsuura, R.H.Heflich, Development of an in vivo gene mutation assay using the endogenousPig-A gene: II. Selection of Pig-A mutant rat spleen T-cells with proaerolysinand sequencing Pig-A cDNA from the mutants, Environ. Mol. Mutagen.(2008).
36] S.M. Bryce, J.C. Bemis, S.D. Dertinger, In vivo mutation assay based onthe endogenous Pig-a locus, Environ. Mol. Mutagen. 49 (2008) 256–264.
37] P.M. Hernandez-Campo, J. Almeida, M.L. Sanchez, M. Malvezzi, A. Orfao, Normalpatterns of expression of glycosylphosphatidylinositol-anchored proteins ondifferent subsets of peripheral blood cells: a frame of reference for the diagnosis
of paroxysmal nocturnal hemoglobinuria, Cytometry B Clin. Cytom. 70B (2006)71–81.38] D.W. Knowles, J.A. Chasis, E.A. Evans, N. Mohandas, Cooperative action betweenband 3 and glycophorin A in human erythrocytes: immobilization of band3 induced by antibodies to glycophorin A, Biophys. J. 66 (1994) 1726–1732.