estrogen potentiates the behavioral and nucleus accumbens dopamine response to continuous...
TRANSCRIPT

Estrogen potentiates the behavioral and nucleusaccumbens dopamine response to continuous haloperidoltreatment in female rats
Dan Madularu,* Waqqas M. Shams* and Wayne G. BrakeDepartment of Psychology, Centre for Studies in Behavioural Neurobiology, Concordia University, Montreal, Quebec, Canada
Keywords: amphetamine, antipsychotic, estradiol, microdialysis, schizophrenia
Abstract
Estrogen has been shown to enhance the effects of antipsychotics in humans. To investigate the mechanisms of how this mayoccur, the current study examined estradiol’s effects on dopaminergic transmission and behavior in amphetamine-sensitized andnon-sensitized female rats. Sixty-four ovariectomized female Sprague–Dawley rats were used for this study. Half of the rats weresensitized to four once-daily injections of 1 mg/kg amphetamine and the other half served as controls. Rats received chronicadministration of either low-dose haloperidol (0.25 mg/kg/day) or saline vehicle via osmotic minipumps implanted subcutaneously.The groups were further subdivided with respect to estradiol treatment: low chronic estrogen (subcutaneous estradiol implant,0.36 mg/pellet: 90-day release, plus an additional oil vehicle injection every second day) and high pulsatile estrogen (subcutane-ous estradiol implant plus an additional 10 lg/kg estradiol injection every second day). Motor activity was assessed at day 2 andday 12 during haloperidol treatment, while nucleus accumbens dopamine availability was assessed via microdialysis 10 days intoantipsychotic treatment. Haloperidol treatment along with high, but not low, estradiol replacement was effective in reducingamphetamine-induced locomotor activity in sensitized rats. High estradiol treatment also augmented the effects of chronic halo-peridol in reducing dopaminergic release in sensitized rats. These data suggest that estradiol levels affect both the behavioraland the dopamine responses to chronic antipsychotic treatment.
Introduction
There are significant sex differences in patients with schizophreniawith respect to time of onset and symptom manifestation (Anger-meyer & Kuhn, 1988; Hafner et al., 1991; Riecher-Rossler et al.,1994; Hafner, 2003). Women have been shown to differ in symptomseverity depending on the phase of the menstrual cycle (Hallonquistet al., 1993). Studies on medicated pre-menopausal women withschizophrenia suggest an interaction between estrogen levels andtheir response to antipsychotic medications, which all have in com-mon that they are dopamine (DA) D2 receptor (D2R) antagonists.For example, previous research has shown that women receivingestrogen in addition to antipsychotic treatment respond better thanthose with antipsychotic treatment alone (Kulkarni et al., 1996,2001; Akhondzadeh et al., 2003). These findings implicate ovariansteroids in schizophrenia onset, symptomatology and treatmentresponsiveness.The aim of the current study is to further investigate the possible
interactions between antipsychotic treatment, estrogen and the dopa-minergic system in a rodent model, by using female, D-amphetamine
sulphate (AMPH)-sensitized rats. Behaviors elicited by AMPH sensi-tization are thought to reflect some of the positive and cognitivesymptoms of schizophrenia (Tenn et al., 2003; Featherstone et al.,2007). These changes are further thought to correspond to nucleusaccumbens (NAcc) DA transmission changes in both rodents andnon-human primates (Tenn et al., 2003; Castner et al., 2005; Peleg-Raibstein et al., 2008). In a previous study, locomotor activity wasrecorded in response to an acute injection of AMPH in male ratsreceiving chronic antipsychotic treatment over a period of 12 days(Samaha et al., 2007). Chronic continuous antipsychotic treatmentbecame progressively ineffective at blocking AMPH-induced loco-motion, with the higher doses resulting in a potentiated response toAMPH 5 days after treatment cessation. In the current study, weadministered the typical antipsychotic haloperidol (HAL), at thelower concentration of the chronic regimen used by Samaha et al.(2007) which is still shown to reflect effective doses in humans (Ka-pur et al., 2000; Samaha et al., 2007, 2008) to either AMPH-sensi-tized or non AMPH-sensitized female rats. These ovariectomized(OVX) rats received either chronic low alone, or chronic low plusphasic high 17b-estradiol (E2) replacement to simulate two differentestrogen levels during different phases of the estrous cycle in youngfemales (Quinlan et al., 2008). Following an AMPH challenge, loco-motor activity was recorded and NAcc DA and its metabolites weremeasured using in vivo microdialysis.
Correspondence: Wayne G. Brake, as above.E-mail: [email protected]
*Both authors contributed equally to this work.
Received 13 December 2012, revised 26 September 2013, accepted 27 September 2013
© 2013 Federation of European Neuroscience Societies and John Wiley & Sons Ltd
European Journal of Neuroscience, pp. 1–9, 2013 doi:10.1111/ejn.12401
European Journal of Neuroscience

It has been suggested that antipsychotic administration may leadto DA receptor supersensitivity, which could lead to a reboundeffect when drug administration is discontinued (Antelman et al.,1986; Samaha et al., 2008); such a rebound was observed in malerats following discontinuation of continuous HAL at a higher con-centration than used here (Samaha et al., 2007). To examine thisphenomenon in females, HAL administration was discontinued for1 week, after which locomotor activity in response to an additionalAMPH challenge was examined.
Materials and methods
Animals
Sixty-four female Sprague–Dawley rats (Charles River Laboratories,Montreal, QC, Canada) weighing 220–250 g were pair-housed andwere the original N of this study. Cages were located in a 21 °Croom with a 12-h reverse light–dark cycle (lights off at 09.00 h),with ad libitum access to food and water. Bedding consisted of a50 : 50 mixture of corncob and beta-chip. All testing and surgicalprocedures were performed during the dark phase of the diurnalcycle. All animal protocols were previously approved by ConcordiaUniversity’s animal research ethics committee and were in accor-dance with the guidelines put forth by the Canadian Council onAnimal Care as well as the European Communities Council Direc-tive of 24 November 1986.
Drugs
HAL (0.25 mg/kg/day; Sandoz Canada Inc., QC, Canada) wasadministered subcutaneously (SC) using Alzet osmotic minipumps(model: 2002, 14-day delivery, at a rate of 0.5 lL/h; Durect, Cuper-tino, CA, USA). This dose, when administered chronically, has beenpreviously shown to result in ~ 75% D2R occupancy in the striatumof rats (Samaha et al., 2007, 2008), similar to D2R striatal occupan-cies observed following effective antipsychotic doses in humans(Kapur et al., 2000).AMPH (1 mg/kg, 0.5 mg/kg or 0.25 mg/kg; Sigma–Aldrich) was
dissolved in 0.9% saline and administered intraperitoneally (IP).These doses were selected based on previous studies inducingbehavioural sensitization to AMPH as well as studies examining theefficacy of antipsychotics in response to an AMPH challenge (e.g.Samaha et al., 2007).All rats were implanted with subcutaneous E2 pellets to provide a
chronic low dose of E2 (0.36 mg/pellet, 90-day release; InnovativeResearch of America, Sarasota, FL, USA). Additionally, half of theanimals received a subcutaneous injection of E2 every second day(20 lg/kg dissolved in sesame seed oil) in a volume of 0.5 mL/kgbody weight, providing an intermittent phasic high dose. The low-E2 rats also received an injection of sesame oil vehicle every secondday as a control. These doses were chosen to mimic the levels ofE2 in estrous and proestrous young cycling rats (Overpeck et al.,1978; Quinlan et al., 2008).
Surgery
Rats were anesthetized using isoflurane (Inhalation Anaesthetic,Richmond Hill, ON, Canada), and two 8.3-mm stainless steel cannu-lae (21-gauge; Plastics-One, Ronaoke, VA, USA) were stereotaxi-cally implanted, bilaterally, toward both the left and right NAcc atthe following coordinates from bregma: anteroposterior (AP),1.8 mm, lateral–medial (LM), 3.0 mm and dorsoventral (DV),
5.4 mm, at a 10° angle. Cannulae were anchored into place withskull-screws using dental cement. Obturators (26-gauge; Plastics-One) were inserted into each cannula. Following surgery, animalswere single-housed for the remainder of the experiment and werehandled every day for ~ 5 min/day.All surgical procedures, i.e. OVX and E2 pellet and cannula
implantations, were performed at the same time in order to avoidmultiple sessions of general anesthesia. All rats were ovariectomizedvia bilateral lumbar incisions (1 cm). Post-ovariectomy, rats wereimplanted with E2 pellets in the nape region. They were adminis-tered the analgesic drug Anafen (0.1 mL/rat, SC; Merial CanadaInc., Morgan Baie d’Urfe, QC, Canada) and the antibiotic penicillinG (0.2 mL/rat, intramuscular; CDMV, St Hyscinthe, QC, Canada).Antibiotic ointment (By/Par Pharmaceuticals Inc., Brampton, ON,Canada) was also applied to the incision. Rats were allowed a weekto recover in their home cages following surgery.
AMPH sensitization and challenge
To avoid possible stress associated with the introduction of a novelenvironment, rats were habituated to the locomotor boxes for 2 h/day for 2 consecutive days. In addition, each rat received an IPinjection of saline 1 day before the induction phase of AMPH sensi-tization. Half of the rats were then administered a single dailyAMPH (1 mg/kg IP) injection (sensitized group; SEN) and halfwere administered saline (non-sensitized group; NON) for four con-secutive days while locomotor activity was recorded (Robinson,1984; Robinson & Becker, 1986).Spontaneous locomotor behaviour was monitored in activity
chambers (Truscan Activity Monitoring System; Coulbourn Instru-ments, Allentown, PA, USA). Each chamber (39 9 42 9 50 cm)had four transparent Plexiglas walls and a removable plastic trayat the bottom. Chambers were placed in sound-attenuating boxesin a dark room. Locomotor activity was monitored for a period of120 min, by recording infrared beam interruptions on two sensorrings placed around the chambers (on the outside of the Plexiglaswalls), creating a 16 9 16 beam matrix. The monitoring sessionwas divided into pre-injection (30 min) and post-injection(90 min) components, during which the TRUSCAN Softwarerecorded total time spent moving. All rats were tested throughoutthe experiment in the same respective activity chamber at thesame time of day.After a 1-week AMPH withdrawal period, rats were administered
an initial AMPH challenge (0.5 mg/kg IP) to determine whetherthey exhibited sensitization to the locomotor stimulating effects ofAMPH (see Fig. 1 for experimental timeline). The doses selectedfor the subsequent challenge injections were based on a pilot study,in which it was observed that AMPH doses > 0.25 mg/kg resultedin stereotypy (data not shown).As stated previously, rats were divided into two main groups,
SEN (n = 32) and NON (n = 32). Within each of these groups andfollowing the initial AMPH challenge, rats were assigned to one offour E2 groups: SEN with low E2 replacement (n = 16), SEN withhigh E2 replacement (n = 16), NON with low E2 replacement(n = 16) and NON with high E2 replacement (n = 16). Thesegroups were each then further divided into two conditions depend-ing upon whether they received chronic HAL or chronic saline(SAL). The final group designations were as follows: sensitized,with high E2 replacement and HAL (HE; HE/SEN; n = 8), sensi-tized with high E2 replacement and SAL (SE; SE/SEN; n = 8),sensitized with low E2 replacement and HAL (He; He/SEN; n = 8),low E2 replacement and SAL (Se; Se/SEN; n = 8), non-sensitized
© 2013 Federation of European Neuroscience Societies and John Wiley & Sons LtdEuropean Journal of Neuroscience, 1–9
2 D. Madularu et al.

with high E2 and HAL (HE/NON; n = 8), non-sensitized with highE2 and SAL (SE/NON; n = 8), non-sensitized with low E2 andHAL (He/NON; n = 8) and non-sensitized with low E2 and SAL(Se/NON; n = 8). Rats were administered four subsequent AMPH(0.25 mg/kg, IP) challenges on days 2, 10 and 12 of HAL or SALtreatment and 1 week after discontinuation of HAL treatment. Loco-motor activity was assessed on days 2 and 12 in order to compareshort- versus long-term HAL treatment.
In vivo microdialysis
Apparatus
In vivo microdialysis was performed in response to an AMPH chal-lenge, with dialysate samples being collected from both hemisphereson day 10 of HAL treatment. Microdialysis testing occurred insidefour separate operant chambers located inside foam-insulated isola-tion units that minimized noise and other environmental stimuli(Coulbourn Instruments, Whitehall, PA, USA). The operant cham-bers (28 9 18 9 19 cm) were equipped with a fan and a houselight. The ceiling of the isolation unit had a small opening thatallowed for unobstructed passage of the microdialysis probe tubinginto the operant chamber. The operant chambers had grid floors witha plastic tray underneath filled with beta chip.
Probes
Probes were assembled according to previously reported methods(Sorge et al., 2005). They consisted of 20-lm-diameter polyethyl-ene (PE) tubing (70–75 cm long; Plastics-One, Roanoke, VA,USA) with one end connected to the stainless steel shaft of adual-channel liquid swivel (HRS Scientific, Montreal, QC, Can-ada). The swivel was located on top of the isolation unit and wasconnected to a variable speed electric syringe infusion pump (Har-vard Apparatus, South Natick, MA, USA). Dialysate was collectedfrom the outlet of the probe into 0.5-mL Eppendorf tubes (Sigma–Aldrich). The other end of the PE tubing was attached to a probetip consisting of 26-gauge stainless steel tubing, 22 mm in length(Fisher Scientific, Nepean, ON, Canada) and a 2.5-mm-long semi-permeable membrane (280 lm OD, 220 lm ID, with a molecularweight cutoff of 13 000; Fisher Scientific). The outer end mem-brane was occluded with epoxy syringe glue (Henkel, Mississauga,ON, Canada) to create a closed system for the flow of dialysate.Small-diameter fused silica tubing (Polymicro Technologies, Pheo-nix, AZ, USA) extended into the probe 0.5 mm from the glued tipof the semi-permeable membrane. A stainless steel collar wasscrewed onto the cannula to secure the probe.
Dialysate collection
Ten days following minipump implantation, rats were anesthetized andmicrodialysis probes were lowered into each guide cannula 5 h beforedialysate sampling began. When lowered, the probe extended 3.0 mmbeyond the guide cannula directing the probe tip andmembrane towardsthe center of the NAcc. Artificial cerebrospinal fluid (aCSF; in mM:Na+, 145; K+, 2.7; Ca2+, 1.2; Mg2+, 1.0; Cl�, 150; ascorbate, 0.2; andNa2HPO4, 2; pH 7.4 � 0.1; Sigma) was perfused through the probeduring a period of 5 h to prevent occlusion and stabilize the baseline, ata rate of 1.0 lL/min. Following this period, six baseline dialysatesamples were collected. Each sample was collected for 10 min at a flowrate of 1.0 lL/min (resulting in 10 lL of dialysate per sample). Sam-ples were immediately placed on dry ice and stored at �80 °C. Afterbaseline, rats were administered AMPH (0.25 mg/kg IP) and another12 samples were collected every 10 min for a period of 2 h. Upon com-pletion of microdialysis, the probes were removed and replaced withobturators and the animals were returned to their home cages.
High performance liquid chromatography (HPLC)
Dialysate samples were thawed and immediately analyzed for DAand its metabolites, dihydroxyphenylacetic acid (DOPAC) and ho-movanillic acid (HVA), using HPLC with electrochemical detection.The samples were loaded through manual injection ports (Reo-dyn 7125; 20-lL loop) onto C-18 reverse-phase columns (5 lm,15 cm; Higgins Analytical, Mountain View, CA, USA). DA and itsmetabolites were measured on separate independent channels withdual-channel ESA coulometric detectors (Coulochem III, Waltham,MA, USA; with a 5011 model analytical cell) for reduction and/oroxidation currents. Mobile phase was circulated through at a flowrate of 1.1 mL/min by Waters 515 HPLC pumps (Waters, QC,Canada), and consisted of: 20% acetonitrile, 40 mL; sodium dodecylsulphate, 0.076 M; EDTA, 0.1 M; NaPO4, 0.058 M; and citric acid,0.27 M; pH 3.35. Known amounts of standard DA and its metabo-lites (concentrations: DA, 0.384 pg/lL; DOPAC, 90 pg/lL; HVA,90 pg/lL; Sigma–Aldrich) were used to calibrate the system usingestimates from peak heights by comparison with standard injections.Extracellular levels of DA (elution time ~6.5 min) and its metabo-lites (DOPAC elution time, ~2.25 min; HVA elution time,~ 3.7 min) were analyzed using the EZCHROM Chromatography Soft-ware Data system (Scientific Software, San Ramon, CA, USA).
Histology and plasma E2 levels
Following the final AMPH challenge, rats were decapitated andbrains were removed and flash-frozen for later histology, while
Fig. 1. Schematic of the experimental timeline. After OVX, cannula and estrogen replacement surgery, rats received four daily injections of either AMPH orSAL IP. One week after the last AMPH injection, all rats received an AMPH challenge. Following the challenge, rats were implanted subcutaneously witheither HAL- or SAL-containing delivery minipumps. At 2 and 12 days into HAL or SAL treatment, as well as 1 week after minipump removal, all rats wereinjected with AMPH and locomotor activity was measured. Dialysate collection occurred 10 days into HAL or SAL treatment.
© 2013 Federation of European Neuroscience Societies and John Wiley & Sons LtdEuropean Journal of Neuroscience, 1–9
Estradiol and continuous antipsychotic treatment 3

blood was collected from a subset of rats (n = 14) to determine cir-culating E2 levels. Blood was stored on ice and immediately centri-fuged. Plasma was then collected and stored at �20 °C untilassayed. E2 was measured using an enzyme-linked immunosorbent
assay (ELISA) kit (Life Technologies, Frederick, MD, USA). Theassay antibodies have 100% cross-reactivity with E2 and 0.2% and0.05% cross-reactivity with estrone and estriol, respectively. Therange of the assay is between 0 and 2000 pg/mL and the reportedinter-assay variation is 7–9%. Brains were sliced along the coronalplane at 40 lm using a cryostat. Sections were mounted onto glassslides and stained with Cresyl Violet to confirm probe placements.
Statistical analysis
Samaha et al. (2007) showed that during a 12-day chronic HALtreatment regimen, male rats respond to the locomotor activity-reducing effects of HAL in response to AMPH by day 2 but thiseffect disappears by day 12. To examine whether this effect is simi-lar in females and whether E2 levels might influence it, here day 2HAL treatment was compared to day 12 in both SEN and NONfemales with either high or low E2 replacement. Spontaneous activ-ity was expressed as total moving time during 5-minute bins follow-ing AMPH. Data were analyzed using eight-two-way mixed ANOVAs,comparing SE, Se, HE and He on days 2 and 12 into treatment forboth SEN and NON groups. Between-subjects factors were day (2,12) and time following AMPH injection served as within-subjectfactor. A separate three-way ANOVA was conducted on AMPH-
Fig. 2. Mean (+SEM) locomotor activity in response to an acute AMPHinjection (0.5 mg/kg, IP) in rats following amphetamine withdrawal. Thegreater activity of the amphetamine-treated rats suggests that behavioural sen-sitization has occurred. The SEN group showed significantly (*P < 0.0001)greater activity than the NON group in response to an AMPH challenge.
A B
C D
Fig. 3. Mean (�SEM) locomotor activity of amphetamine-sensitized (SEN) groups (A, B, C and D) on days 2 and 12 of treatment in response to amphetamine(AMPH) challenge (0.25 mg/kg, IP). HAL: haloperidol, SAL: saline, E2: estrogen. Asterisk indicates day 12 significantly lower than day 2 (P < 0.01) in ratsreceiving high E2 replacement in conjunction with HAL treatment.
© 2013 Federation of European Neuroscience Societies and John Wiley & Sons LtdEuropean Journal of Neuroscience, 1–9
4 D. Madularu et al.

induced locomotor activity following a week of HAL withdrawal,with E2 and HAL treatment as between-subjects factors and time aswithin-subject factor.For in vivo microdialysis, concentrations of DA and its metabo-
lites are expressed percentages of baseline. That is, the three sam-ples taken prior to drug injection were averaged as baseline andsubsequent samples were converted to percentages of this value.Four two-way mixed ANOVAs were performed on DA levels withsensitization (SEN vs. NON) as the between-subjects factor and timeas the within-subject factor.To determine whether sensitization had occurred, an independent-
samples t-test was used, comparing average time spent moving inresponse to an AMPH challenge for the SEN compared to the NONgroup. Plasma estradiol levels were compared between the high E2and low E2 groups using an independent-samples t-test.
Results
Behavioural assessment
SEN rats
Three rats died during microdialysis testing (day 10) and another ratdied during surgery; thus a final N of 60 was used for the locomotor
analyses. Expression of sensitization was measured by administeringhalf the dose (i.e. 0.5 mg/kg) of AMPH used for induction (i.e.1 mg/kg) following a 1-week withdrawal period. The locomotorresponse of the SEN group was significantly greater than that ofthe NON rats in response to a low-dose challenge AMPH injec-tion (t34 = 2.12, P < 0.0001), showing that sensitization to the loco-motor activating effects of AMPH had occurred in the SEN group(Fig. 2).Amphetamine-sensitized, HAL-treated rats with high E2 replace-
ment (HE group) showed a difference in AMPH-induced locomotoractivity (Fig. 3A), where HAL significantly reduced AMPH-inducedactivity on day 12 compared to day 2 of treatment (F1,6 = 17.98,P = 0.005). No other comparisons were statistically significant(Fig. 3B–D). These findings indicate that HAL had little or nobehavioural effect in female rats after 2 days of treatment but did soafter 12 days, notably only in females with high levels of E2.
NON rats
There was a significant difference in AMPH-induced locomotoractivity between days 2 and 12 in the SAL-treated group (Fig. 4C)receiving high E2 replacement (F1,6 = 13.39, P = 0.011). Therewere no differences in activity in the other NON groups, suggesting
A B
C D
Fig. 4. Mean (�SEM) locomotor activity of non-amphetamine sensitized (NON) groups (A, B, C and D) on days 2 and 12 of treatment in response to amphet-amine (AMPH) challenge (0.25 mg/kg, IP). HAL: haloperidol, SAL: saline, E2: estrogen. Asterisk indicates day 12 significantly higher than day 2 (P < 0.05)in NON rats with high E2 replacement, in the absence of HAL treatment.
© 2013 Federation of European Neuroscience Societies and John Wiley & Sons LtdEuropean Journal of Neuroscience, 1–9
Estradiol and continuous antipsychotic treatment 5
that high E2 replacement exacerbated the effects of AMPH after10 days of treatment (Fig. 4A, B and D).Taken together, the behavioural findings show that although in
AMPH-sensitized rats high E2 replacement enhanced the locomotoractivity-reducing effects of HAL 12 days into treatment, high E2replacement alone increased locomotor activity in non-sensitized ratsafter chronic administration of AMPH.
HAL withdrawal
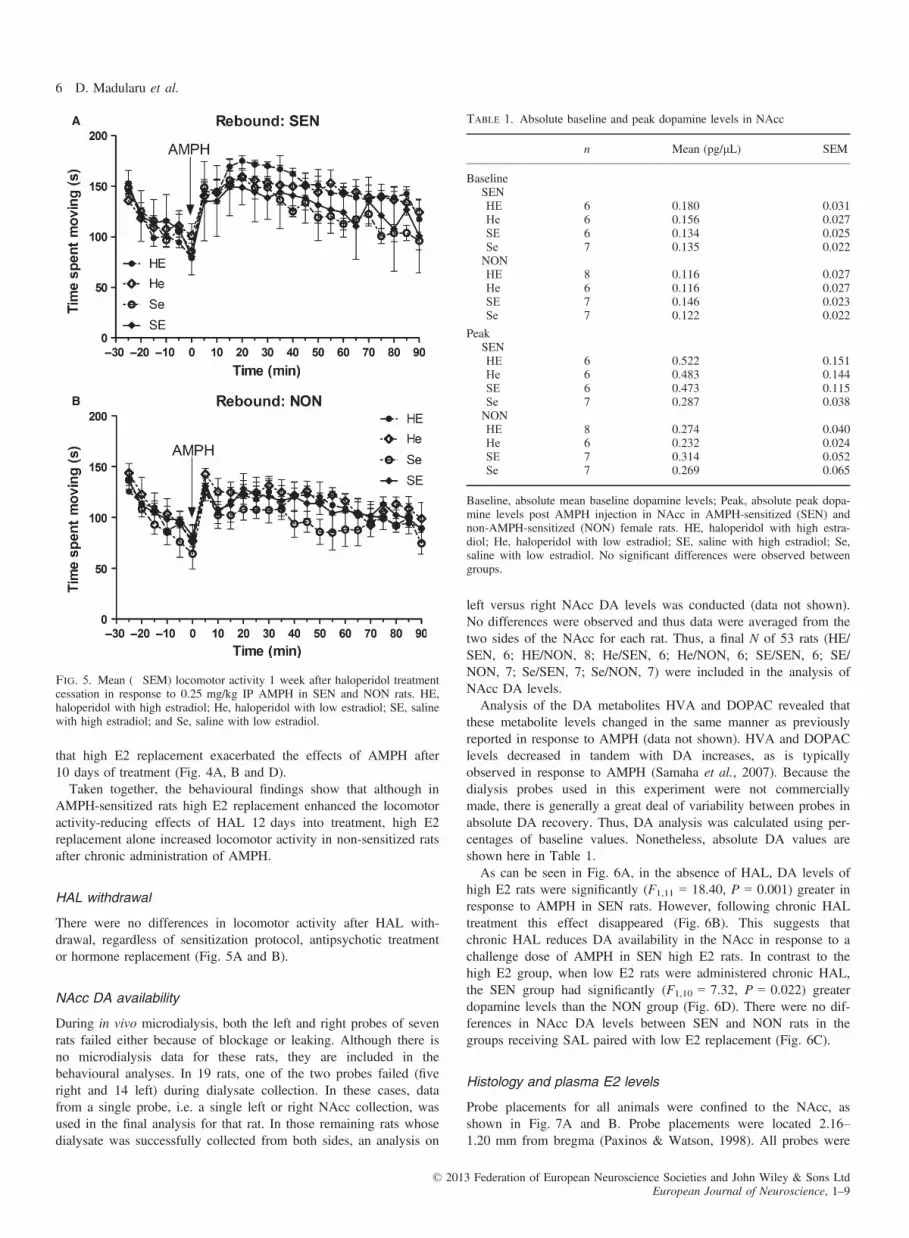
There were no differences in locomotor activity after HAL with-drawal, regardless of sensitization protocol, antipsychotic treatmentor hormone replacement (Fig. 5A and B).
NAcc DA availability
During in vivo microdialysis, both the left and right probes of sevenrats failed either because of blockage or leaking. Although there isno microdialysis data for these rats, they are included in thebehavioural analyses. In 19 rats, one of the two probes failed (fiveright and 14 left) during dialysate collection. In these cases, datafrom a single probe, i.e. a single left or right NAcc collection, wasused in the final analysis for that rat. In those remaining rats whosedialysate was successfully collected from both sides, an analysis on
left versus right NAcc DA levels was conducted (data not shown).No differences were observed and thus data were averaged from thetwo sides of the NAcc for each rat. Thus, a final N of 53 rats (HE/SEN, 6; HE/NON, 8; He/SEN, 6; He/NON, 6; SE/SEN, 6; SE/NON, 7; Se/SEN, 7; Se/NON, 7) were included in the analysis ofNAcc DA levels.Analysis of the DA metabolites HVA and DOPAC revealed that
these metabolite levels changed in the same manner as previouslyreported in response to AMPH (data not shown). HVA and DOPAClevels decreased in tandem with DA increases, as is typicallyobserved in response to AMPH (Samaha et al., 2007). Because thedialysis probes used in this experiment were not commerciallymade, there is generally a great deal of variability between probes inabsolute DA recovery. Thus, DA analysis was calculated using per-centages of baseline values. Nonetheless, absolute DA values areshown here in Table 1.As can be seen in Fig. 6A, in the absence of HAL, DA levels of
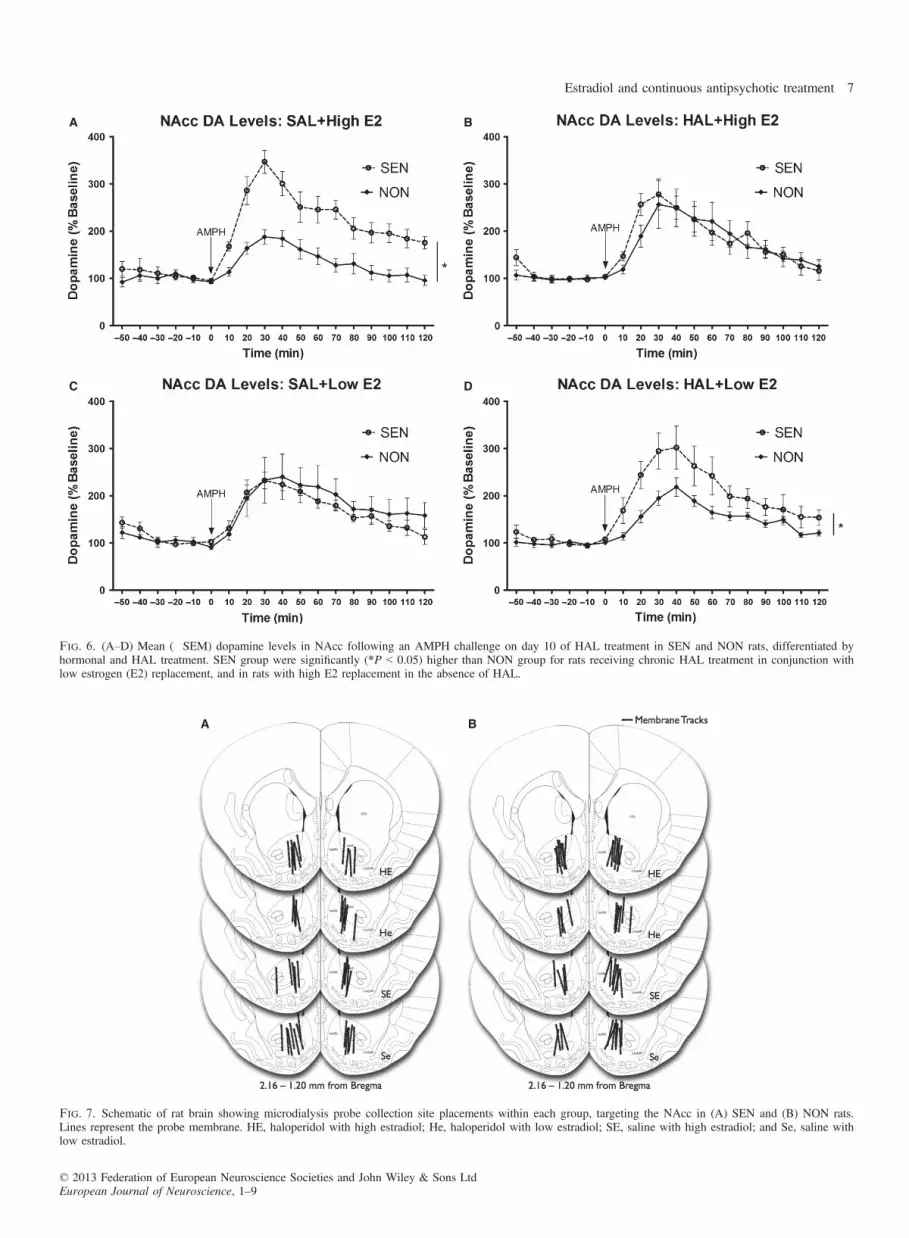
high E2 rats were significantly (F1,11 = 18.40, P = 0.001) greater inresponse to AMPH in SEN rats. However, following chronic HALtreatment this effect disappeared (Fig. 6B). This suggests thatchronic HAL reduces DA availability in the NAcc in response to achallenge dose of AMPH in SEN high E2 rats. In contrast to thehigh E2 group, when low E2 rats were administered chronic HAL,the SEN group had significantly (F1,10 = 7.32, P = 0.022) greaterdopamine levels than the NON group (Fig. 6D). There were no dif-ferences in NAcc DA levels between SEN and NON rats in thegroups receiving SAL paired with low E2 replacement (Fig. 6C).
Histology and plasma E2 levels
Probe placements for all animals were confined to the NAcc, asshown in Fig. 7A and B. Probe placements were located 2.16–1.20 mm from bregma (Paxinos & Watson, 1998). All probes were
A
B
Fig. 5. Mean (�SEM) locomotor activity 1 week after haloperidol treatmentcessation in response to 0.25 mg/kg IP AMPH in SEN and NON rats. HE,haloperidol with high estradiol; He, haloperidol with low estradiol; SE, salinewith high estradiol; and Se, saline with low estradiol.
Table 1. Absolute baseline and peak dopamine levels in NAcc
n Mean (pg/lL) SEM
BaselineSENHE 6 0.180 0.031He 6 0.156 0.027SE 6 0.134 0.025Se 7 0.135 0.022NONHE 8 0.116 0.027He 6 0.116 0.027SE 7 0.146 0.023Se 7 0.122 0.022
PeakSENHE 6 0.522 0.151He 6 0.483 0.144SE 6 0.473 0.115Se 7 0.287 0.038NONHE 8 0.274 0.040He 6 0.232 0.024SE 7 0.314 0.052Se 7 0.269 0.065
Baseline, absolute mean baseline dopamine levels; Peak, absolute peak dopa-mine levels post AMPH injection in NAcc in AMPH-sensitized (SEN) andnon-AMPH-sensitized (NON) female rats. HE, haloperidol with high estra-diol; He, haloperidol with low estradiol; SE, saline with high estradiol; Se,saline with low estradiol. No significant differences were observed betweengroups.
© 2013 Federation of European Neuroscience Societies and John Wiley & Sons LtdEuropean Journal of Neuroscience, 1–9
6 D. Madularu et al.
A B
C D
Fig. 6. (A–D) Mean (�SEM) dopamine levels in NAcc following an AMPH challenge on day 10 of HAL treatment in SEN and NON rats, differentiated byhormonal and HAL treatment. SEN group were significantly (*P < 0.05) higher than NON group for rats receiving chronic HAL treatment in conjunction withlow estrogen (E2) replacement, and in rats with high E2 replacement in the absence of HAL.
A B
Fig. 7. Schematic of rat brain showing microdialysis probe collection site placements within each group, targeting the NAcc in (A) SEN and (B) NON rats.Lines represent the probe membrane. HE, haloperidol with high estradiol; He, haloperidol with low estradiol; SE, saline with high estradiol; and Se, saline withlow estradiol.
© 2013 Federation of European Neuroscience Societies and John Wiley & Sons LtdEuropean Journal of Neuroscience, 1–9
Estradiol and continuous antipsychotic treatment 7

located both within the core and shell of the NAcc. ELISA results(Fig. 8) indicate an approximate two-fold increase in E2 levels(13.31 � 3.55 pg/mL) in high E2 rats compared to low E2 rats(6.59 � 0.85 pg/mL) 1 day following the last high E2 injection(t13 = 2.12, P = 0.026).
Discussion
Previous studies suggest that E2 may have antipsychotic-like proper-ties, possibly through its interaction with the dopaminergic system(Kulkarni et al., 2001). The aim of this study was to investigate thisinteraction in chronic low-dose HAL-treated, AMPH-sensitized andnon-sensitized female rats using behavioural and neurochemicalanalyses.A significant effect of HAL in sensitized rats was observed on
day 12 compared to day 2, when AMPH-induced locomotor hyper-activity was reduced in rats receiving high E2, but not low E2,replacement. The implications of these results are two-fold. Firstly,short-term (i.e. 2 days) antipsychotic treatment had no effect inreducing AMPH-induced locomotion at this dose in female rats, incontrast to previous findings in male rats (Samaha et al., 2007) andhumans (Stern et al., 1993). Secondly, long-term (i.e. 12 days) low-dose HAL treatment was effective only in female rats receiving highE2 replacement in sensitized rats. These results partly contradict pre-vious findings by Samaha et al. (2007), who observed that at day 12neither high nor low doses of HAL proved to be effective in reduc-ing AMPH-induced locomotion in male rats. Our findings suggestthat E2 has antipsychotic-like effects when paired with a long-termHAL regimen in AMPH-sensitized female rats. One of the possiblereasons behind the discrepancy between the current and previousfindings is probably the fact that the previous study (Samaha et al.,2007) used male rats and females have been shown to require lowerdoses of antipsychotic drugs (Melkersson et al., 2001).Haloperidol withdrawal had no effect on AMPH-induced locomo-
tion, regardless of whether the rats were sensitized or not (Fig. 5).The study by Samaha et al. (2007) yielded similar results, wheremale rats treated with a low dose of HAL (0.25 mg/kg) failed toshow a potentiated response to AMPH after a 5-day period of anti-psychotic withdrawal, while rats treated with a higher dose did showa potentiated response to AMPH (Samaha et al., 2007). It would beinteresting to see in future studies whether females show a with-drawal effect at a higher dose of HAL.Amphetamine sensitization enhanced the NAcc DA response to
acute AMPH when rats received high E2 replacement (Fig. 6A).
When high E2 replacement rats were administered chronic HAL,this effect went away (Fig. 6B). That is to say, HAL was effectivein reducing the higher NAcc DA levels observed in SEN rats whenthey were given high E2. By comparison, in rats administered lowE2 replacement, HAL did not reduce NAcc DA levels in SEN rats(Fig. 6D) to the same degree as seen in high E2 rats (Fig 6B). Inother words, NAcc DA levels were significantly higher in SEN ratscompared to NON rats when HAL was accompanied by low E2replacement. Finally, there were no differences in DA availabilitybetween SEN and NON low E2 rats in the absence of HAL treat-ment (Fig. 6C). Although it has been established that AMPH sensiti-zation and acute DA release in response to psychostimulants are atleast in part mediated by estrogen, it is unclear why high levels ofE2 replacement yield differential neurochemical as well as behavio-ural effects compared to low E2.The mechanisms by which E2 is effective in reducing AMPH-
induced locomotion when paired with prolonged HAL treatment areunknown. The effects of E2 on striatal DA are not limited only torelease, but also to DA receptor state. D2Rs are thought to be onlyfunctional when they are in their high affinity state (D2High) and areinactive when they are in their low affinity state (Seeman, 2006,2010, 2011, 2013). An in vitro study showed that an acute physio-logical dose of E2 administered to OVX rat striatal tissue producesa rapid conversion of DA D2Rs from their high to low affinity state(Levesque & Dipaolo, 1988). Similarly, the affinity state of DAD2Rs fluctuates across the estrous cycle with the most DA D2Rs inthe high affinity state during diestrus when estrogen is low and mostin the low affinity state during behavioural estrus and proestrus(Dipaolo et al., 1988). In addition, chronic replacement of E2 inOVX rats results in an increase in striatal DA D1 receptor (D1R)binding, suggesting that E2 affects both the affinity state of D2Rsand the binding of D1Rs (Levesque & Dipaolo, 1989).Previous research showed that although HAL treatment alone
increased D2High availability (Samaha et al., 2007; Seeman, 2009),when paired with AMPH, HAL reduces by 60% AMPH-elevatedD2High receptors (Seeman, 2009). One could speculate that differentlevels of circulating estrogen might influence the affinity state ofDA D2R such that increased levels of estrogen might result in ashift in DA D2R affinity from its high state into a low one. Thiscould potentially explain how E2 enhances the behavioural effectsof HAL. Future studies should investigate the potential effects ofestrogen replacement on the state of the DA D2R in the striatum ofsensitized rats. On the other hand, such a postsynaptic mechanismmay not explain how E2 affects the NAcc DA response to HAL.We have evidence that E2 affects D2R autoreceptors in the dorsalstriatum such that autoreceptor function is less sensitive in high E2rats (Hussain et al., 2013). This effect may be direct via estrogenreceptors; our recent findings show that both ERa (estrogen receptoralpha) and GPER-1 (g-protein-coupled estrogen receptor 1) areindeed located on DA terminals in the NAcc (Almey, A., Milner,T.A. & Brake, W.G., unpublished observations), although we havepreviously shown that this is not the case in the dorsal striatum(Almey et al., 2012). Thus, E2 may be acting at both pre- and post-synaptic sites in the NAcc to modulate the effects of HAL, andpossibly via different mechanisms.HAL became effective only in AMPH-sensitized rats receiving high
E2 replacement, and only with prolonged treatment. These data mirrorprevious research on humans, where estrogen, when added to antipsy-chotic treatment, significantly reduces schizophrenic symptoms(Kulkarni et al., 1996, 2001; Akhondzadeh et al., 2003). In addition,the neurochemical analysis points at a direct link between NAcc DAavailability and E2 levels, whereby locomotor activity in response to
Fig. 8. Mean (+SEM) E2 levels in rats with high and low hormone replace-ment 24 h following the final E2 injection. The high estrogen group showeda significant (*P < 0.05) increase in circulating estrogen. E, high estrogenreplacement; e, low estrogen replacement.
© 2013 Federation of European Neuroscience Societies and John Wiley & Sons LtdEuropean Journal of Neuroscience, 1–9
8 D. Madularu et al.

AMPH seems to be at least in part driven by this relationship.Although earlier studies have shown that estrogen replacement signifi-cantly increased postsynaptic striatal DA levels, as well as AMPH-induced stereotypy (Hruska et al., 1980), additional reports suggestthat similar treatment reduced DA levels in the NAcc (Chavez et al.,2010). To our knowledge, this is the first study to demonstrate, in sen-sitized female rats, differences in NAcc DA availability in response toAMPH that are mediated by levels of E2. It is also the first to showthat antipsychotic treatment efficacy does not decrease in female ratswhen administered chronically at a lower dose.
Conflict of Interest
The authors have no conflict of interest to declare.
Acknowledgements
This work was supported by a grant to W.G.B. from the Natural Sciencesand Engineering Research Council (NSERC) of Canada. The Center forStudies in Behavioral Neurobiology (CSBN) is a ‘group de recherche’funded by the Fonds de Recherche Qu�ebec–Sant�e. We would like to thankMarie-Pierre Cossette for her technical assistance with microdialysis.
Abbreviations
AMPH, D-amphetamine sulphate; D2R, dopamine D2 receptor; DA, dopa-mine; DOPAC, dihydroxyphenylacetic acid; E2, estradiol; HAL, haloperidol;HE, high E2 replacement with HAL; He, low E2 replacement with HAL;HPLC, high performance liquid chromatography; HVA, homovanillic acid;IP, intraperitoneally; NAcc, nucleus accumbens; NON, non-sensitized group;OVX, ovariectomized; SAL, saline; SE, high E2 replacement with SAL; Se,low E2 replacement with SAL; SEN, sensitized group.
References
Akhondzadeh, S., Nejatisafa, A.A., Amini, H., Mohammadi, M.R., Larijani,B., Kashani, L., Raisi, F. & Kamalipour, A. (2003) Adjunctive estrogentreatment in women with chronic schizophrenia: a double-blind, random-ized, and placebo-controlled trial. Prog. Neuropsychopharmacol. Biol.Psychiatry, 27, 1007–1012.
Almey, A., Filardo, E.J., Milner, T.A. & Brake, W.G. (2012) Estrogen recep-tors are found in glia and at extranuclear neuronal sites in the dorsal stria-tum of female rats: evidence for cholinergic but not dopaminergiccolocalization. Endocrinology, 153, 5373–5383.
Angermeyer, M.C. & Kuhn, L. (1988) Gender differences in age at onset ofschizophrenia. An overview. Eur. Arch. Psy. Clin. N., 237, 351–364.
Antelman, S.M., Kocan, D., Edwards, D.J., Knopf, S., Perel, J.M. & Stiller,R. (1986) Behavioral effects of a single neuroleptic treatment grow withthe passage of time. Brain Res., 385, 58–67.
Castner, S.A., Vosler, P.S. & Goldman-Rakic, P.S. (2005) Amphetaminesensitization impairs cognition and reduces dopamine turnover in primateprefrontal cortex. Biol. Psychiat., 57, 743–751.
Chavez, C., Hollaus, M., Scarr, E., Pavey, G., Gogos, A. & van den Buuse,M. (2010) The effect of estrogen on dopamine and serotonin receptor andtransporter levels in the brain: an autoradiography study. Brain Res.,1321, 51–59.
Dipaolo, T., Falardeau, P. & Morissette, M. (1988) Striatal D-2 dopamineagonist binding-sites fluctuate during the rat estrous-cycle. Life Sci., 43,665–672.
Featherstone, R.E., Kapur, S. & Fletcher, P.J. (2007) The amphetamine-induced sensitized state as a model of schizophrenia. Prog. Neuropsycho-pharmacol. Biol. Psychiatry, 31, 1556–1571.
Hafner, H. (2003) Gender differences in schizophrenia. Psychoneuroendo-crino., 28(Suppl 2), 17–54.
Hafner, H., Behrens, S., De Vry, J. & Gattaz, W.F. (1991) An animal modelfor the effects of estradiol on dopamine-mediated behavior: implicationsfor sex differences in schizophrenia. Psychiat. Res., 38, 125–134.
Hallonquist, J.D., Seeman, M.V., Lang, M. & Rector, N.A. (1993) Variationin symptom severity over the menstrual cycle of schizophrenics. Biol. Psy-chiat., 33, 207–209.
Hruska, R.E., Ludmer, L.M. & Silbergeld, E.K. (1980) Characterization ofthe striatal dopamine receptor supersensitivity produced by estrogen treat-ment of male rats. Neuropharmacology, 19, 923–926.
Hussain, D., Cossette, M. & Brake, W.G. (2013) Dorsal Striatal dopaminelevels and memory system bias in ovariectomized rats with chronic low orphasic high estradiol replacement. Submitted.
Kapur, S., Zipursky, R., Jones, C., Remington, G. & Houle, S. (2000)Relationship between dopamine D(2) occupancy, clinical response, andside effects: a double-blind PET study of first-episode schizophrenia. Am.J. Psychiat., 157, 514–520.
Kulkarni, J., de Castella, A., Smith, D., Taffe, J., Keks, N. & Copolov, D.(1996) A clinical trial of the effects of estrogen in acutely psychoticwomen. Schizophr. Res., 20, 247–252.
Kulkarni, J., Riedel, A., de Castella, A.R., Fitzgerald, P.B., Rolfe, T.J., Taffe,J. & Burger, H. (2001) Estrogen - a potential treatment for schizophrenia.Schizophr. Res., 48, 137–144.
Levesque, D. & Dipaolo, T. (1988) Rapid conversion of high into low stria-tal D2-dopamine receptor agonist binding states after an acute physiologi-cal dose of 17-beta-Estradiol. Neurosci. Lett., 88, 113–118.
Levesque, D. & Dipaolo, T. (1989) Chronic Estradiol treatment increasesovariectomized rat striatal D-1 dopamine-receptors. Life Sci., 45, 1813–1820.
Melkersson, K.I., Hulting, A.L. & Rane, A.J. (2001) Dose requirement andprolactin elevation of antipsychotics in male and female patients withschizophrenia or related psychoses. Brit. J. Clin. Pharmaco., 51, 317–324.
Overpeck, J.G., Colson, S.H., Hohmann, J.R., Applestine, M.S. & Reilly, J.F.(1978) Concentrations of circulating steroids in normal prepubertal and adultmale and female humans, chimpanzees, rhesus monkeys, rats, mice, and ham-sters: a literature survey. J. Toxicol. Env. Health Sci., 4, 785–803.
Paxinos, G. & Watson, C. (1998) The Rat Brain in Stereotaxic Coordinates:Fourth Edition. Academic Press, San Diego, CA.
Peleg-Raibstein, D., Knuesel, I. & Feldon, J. (2008) Amphetamine sensitiza-tion in rats as an animal model of schizophrenia. Behav. Brain Res., 191,190–201.
Quinlan, M.G., Hussain, D. & Brake, W.G. (2008) Use of cognitive strate-gies in rats: the role of estradiol and its interaction with dopamine. Horm.Behav., 53, 185–191.
Riecher-Rossler, A., Hafner, H., Stumbaum, M., Maurer, K. & Schmidt, R.(1994) Can estradiol modulate schizophrenic symptomatology? Schizo-phrenia Bull., 20, 203–214.
Robinson, T.E. (1984) Behavioral sensitization: characterization of enduringchanges in rotational behavior produced by intermittent injections ofamphetamine in male and female rats. Psychopharmacology, 84, 466–475.
Robinson, T.E. & Becker, J.B. (1986) Enduring changes in brain andbehavior produced by chronic amphetamine administration: a review andevaluation of animal models of amphetamine psychosis. Brain Res., 396,157–198.
Samaha, A.N., Seeman, P., Stewart, J., Rajabi, H. & Kapur, S. (2007) “Break-through” dopamine supersensitivity during ongoing antipsychotic treatmentleads to treatment failure over time. J. Neurosci., 27, 2979–2986.
Samaha, A.N., Reckless, G.E., Seeman, P., Diwan, M., Nobrega, J.N. &Kapur, S. (2008) Less is more: antipsychotic drug effects are greater withtransient rather than continuous delivery. Biol. Psychiat., 64, 145–152.
Seeman, P. (2006) Targeting the dopamine D2 receptor in schizophrenia.Expert Opin. Ther. Tar., 10, 515–531.
Seeman, P. (2009) Schizophrenia model of elevated D2(High) receptors:haloperidol reverses the amphetamine-induced elevation in dopamine D2(High). Schizophr. Res., 109, 191–192.
Seeman, P. (2010) Dopamine D2 receptors as treatment targets in schizophre-nia. Clin. Schizophr. Relat. Psychoses, 4, 56–73.
Seeman, P. (2011) All roads to schizophrenia lead to dopamine supersensitiv-ity and elevated dopamine D2(high) receptors. CNS Neurosci. Ther., 17,118–132.
Seeman, P. (2013) Are dopamine D2 receptors out of control in psychosis?Prog. Neuropsychopharmacol. Biol. Psychiatry, 46C, 146–152.
Sorge, R.E., Rajabi, H. & Stewart, J. (2005) Rats maintained chronically onbuprenorphine show reduced heroin and cocaine seeking in tests of extinc-tion and drug-induced reinstatement. Neuropsychopharmacol., 30, 1681–1692.
Stern, R.G., Kahn, R.S., Harvey, P.D., Amin, F., Apter, S.H. & Hirschowitz,J. (1993) Early response to haloperidol treatment in chronic schizophrenia.Schizophr. Res., 10, 165–171.
Tenn, C.C., Fletcher, P.J. & Kapur, S. (2003) Amphetamine-sensitized ani-mals show a sensorimotor gating and neurochemical abnormality similarto that of schizophrenia. Schizophr. Res., 64, 103–114.
© 2013 Federation of European Neuroscience Societies and John Wiley & Sons LtdEuropean Journal of Neuroscience, 1–9
Estradiol and continuous antipsychotic treatment 9