estrategias termorregulatorias y fisiología digestiva del lagarto liolaemus darwinii, del desierto...
TRANSCRIPT

1
“Estrategias termorregulatorias
y fisiología digestiva del lagarto
Liolaemus darwinii
en el desierto del Monte”
Licenciatura en Ecología
Universidad Champagnat
Andrea Anabella Barauna,
Director: Dr. Eduardo Alfredo Sanabria
Co-directora: Dra. María Eugenia Cabrillana
Mendoza, 2013

2
Índice
Pág.
Agradecimientos……………………………………………………………4
Resumen…………………………………………………………………….6
Introducción………………………………………………………………...8
Fundamentación……………………………………………………...9
Objetivo……………………………………………………………………13
Área de estudio…………………………………………………………….14
Clima………………………………………………………………..14
Fauna………………………………………………………………..15
Flora………………………………………………………………...16
Descripción de la especie………………………………………………….17
Materiales y Métodos……………………………………………………..19
Captura……………………………………………………………...19
Trabajo de Laboratorio……………………………………………...19
Medición de la Temperatura corporal selecta……………………20
Obtención de tejidos y plasma………………………………………21
Actividad enzimática en intestino delgado y páncreas…………….23
Concentración de proteínas totales y albúmina………………….24
Determinación de proteínas totales…………………………24
Determinación de albúmina………………………………….25
Curva de calibración………………………………………………..25

3
Análisis de Datos……………………………………………………………27
Resultados…………………………………………………………………28
Temperatura corporal selecta……………………………………….28
Actividad enzimática en intestino delgado y páncreas…………..30
Concentración de proteínas totales y albúmina…………………..30
Discusión……………………………………………………………………32
Temperatura corporal selecta………………………………………32
Actividad enzimática en intestino delgado y páncreas……………35
Concentración de proteínas totales y albúmina………………….36
Conclusiones generales……………………………………………………37
Bibliografía…….……………………………………………………………39

4
Agradecimientos
Este trabajo no hubiera sido posible sin la ayuda y el apoyo de muchas personas que
me acompañaron en cada etapa y a lo largo de toda la carrera.
Quiero agradecer a mis padres, Norma y Alejandro, por darme la posibilidad de
estudiar, por alentar siempre mis sueños y acompañarme en cada decisión.
A mi novio, Gustavo, por ser mi apoyo emocional, por acompañarme en todo desde
que empecé la carrera, bancar mis ausencias por viajes al campo, cursos, congresos, sin
jamás limitarme a crecer y siempre incentivar mi formación.
A mis hermanos Gabriel y Antonella, y mis cuñados, Mariela y Agustín, por estar
siempre conmigo, sacarme de apuros las veces que lo necesité y ser siempre
incondicionales.
A mi director, Eduardo, y su esposa Lorena, por enseñarme tanto, por ser tan
pacientes, tan comprensivos y divertidos.
A mi codirectora, María Eugenia, siempre predispuesta a enseñarme, a explicarme
con mucha paciencia todo, del modo más alegre y llevadero.
Tuve la suerte de tener un director y una codirectora que siempre me hicieron sentir
muy cómoda trabajando con ellos, que sabían muchísimo y me enseñaban transmitiendo su
pasión por lo que hacen.
A mi querida amiga Rocío Aguilar, por darme una mano enorme cuando más lo
necesitaba, poniéndome en contacto con mi director. Por todo lo que me enseñó tanto en

5
campo como en oficina y por corregir este trabajo. Me alegra que podamos tener proyectos
en común para seguir trabajando juntas.
A mi otra querida amiga, Daniela Rodríguez, por siempre darme fuerza, por confiar
en mí y hacer que yo también confíe en que soy capaz de hacer lo que me proponga con
esfuerzo. Por enseñarme tantas cosas que no se aprenden en ninguna universidad y por
permitir que siempre aprenda a su lado.
A Paola Sassi, por su calidez y predisposición para ayudarme, por corregir este
trabajo y por sus excelentes aportes profesionales.
A Yamil Rodríguez, Emanuel Ontivero, Exequiel Gonzalez y Ariel Cataldo, por ir
al campo y capturar prácticamente todos los lagartos utilizados en este estudio.
A mis compañeros de la facultad, con quienes compartí innumerables viajes,
salidas, momentos, fue grandioso conocerlos a todos y hoy son mis amigos además de
colegas.
A mis amigos de siempre, Flor, Barby, Maty y Matias, por estar siempre a mi lado,
por acompañarme a lo largo de toda esta experiencia y contar siempre con su amistad.

6
Resumen
El comportamiento de selección de temperaturas corporales elevadas frente a la
ingesta, es frecuentemente observado en organismos del orden Squamata. No se ha
estudiado este patrón en el género Liolaemus, por ello se puso a prueba en la especie
Liolaemus darwinii (Squamata: Liolaemidae), un lagarto insectívoro, habitante de los
Médanos Grandes, Caucete, Provincia de San Juan. Se determinó la temperatura corporal
selecta (Tsel) bajo dos condiciones experimentales: “ayuno”/“no ayuno” en un gradiente
linear térmico en los meses de octubre y marzo. Además, se midió la actividad de la enzima
digestiva tripsina en intestino delgado y páncreas de lagartos aclimatados a diferentes
temperaturas. En el primer estudio, los lagartos capturados en octubre presentaron una Tsel
significativamente menor (t(gl=722)=18,518; p<0.00002) en el tratamiento “no ayuno”
(30.9±0.2ºC) que en el “ayuno” (32.3±0.2ºC). En cambio, para marzo, la Tsel fue
significativamente mayor (t(gl=498)=5.73; p<0.0001) en el tratamiento “no ayuno”
(32.5±0.2ºC), que en el “ayuno” (30.5±0.2ºC). En el segundo estudio, las mediciones
realizadas en intestino delgado no mostraron diferencias significativas a las diferentes
temperaturas de aclimatación (Kruskal-Wallis test: H (2, N=11)=2,09; p=0,35). En contraste,
las mediciones realizadas en tejido pancreático mostraron una mayor actividad enzimática a
40°C, siendo esta estadísticamente diferente (Kruskal-Wallis test: H (2, N=9)=6,44; p=0,039)
con la actividad obtenida a los 15°C. Los resultados obtenidos en el primer estudio,
muestran que las diferencias estacionales encontradas entre octubre y marzo, podrían
deberse a los ciclos circanuales de los reptiles, como almacenamiento de grasas y
aletargamiento. Respecto al segundo estudio, los resultados reflejan que un aumento de la
temperatura corporal no tiene efecto en el intestino delgado, pero si favorece la secreción

7
de enzima tripsina en el páncreas de los lagartos y estimula las contracciones peristálticas
del esófago.

8
Introducción
El presente trabajo de investigación tiene como propósito acercarnos al
conocimiento de las estrategias termorregulatorias y su relación con los procesos
fisiológicos digestivos en el lagarto Liolaemus darwinii (Reptilia: Squamata: Liolaemidae).
Desde que el control comportamental de la temperatura corporal fue descripto
primariamente por Cowles y Bogert (1944), la termorregulación comportamental ha sido
considerada un factor de importancia en la fisiología digestiva de los reptiles.
Existen evidencias de que las especies pertenecientes al orden Squamata seleccionan
temperaturas corporales más altas luego de alimentarse (Brown y Roberts, 2008). La
hipótesis de la presencia de este patrón, está ampliamente basada en el supuesto de que
temperaturas corporales más elevadas, luego de alimentarse, representan una respuesta
adaptativa comportamental que mejora la digestión y/o conserva energía en periodos de
baja disponibilidad alimentaria (van Marken Lichtenbelt, 1992).
No existen estudios precedentes sobre el comportamiento termofílico digestivo y su
correlación con la actividad de enzimas pancreáticas e intestinales para el género
Liolaemus, por lo que se puso a prueba en el lagarto Liolaemus darwinii (Bell, 1843), un
lacértido ovíparo, insectívoro y psamófilo, habitante de los Médanos Grandes,
departamento de Caucete, provincia de San Juan.
En este proyecto se plantea identificar una relación entre el comportamiento
termofílico y la digestión en dependencia de la temperatura ambiental. Se comparó la
temperatura corporal selecta bajo condiciones de presencia y ausencia de alimento en un
gradiente linear térmico. Además, se contrastó la actividad de la enzima tripsina, una

9
proteasa secretada en el páncreas de forma inactiva y activada en la porción anterior del
intestino delgado, entre organismos aclimatados a diferentes temperaturas.
Fundamentación La temperatura ambiental varía a lo largo del espacio y el tiempo, por lo que la
mayoría de los organismos combinan mecanismos comportamentales, fisiológicos y
morfológicos para regular su temperatura dentro de límites tolerables y de este modo
amortiguar la heterogeneidad ambiental (Angilletta, 2009).
En base a la fuente de origen del calor corporal, podemos distinguir entre
organismos endotermos y ectotermos (IUPS Thermal Commission, 2001). Los primeros
alcanzan su temperatura corporal fundamentalmente mediante una gran producción de calor
metabólico (Bennet et al., 2000), mientras que en los segundos, la fuente de energía
calórica necesaria para los procesos fisiológicos, es externa (Rome et al., 1992).
Con respecto a la capacidad metabólica de mantener una temperatura corporal
constante o no, se puede diferenciar entre organismos homeotermos y poiquilotermos,
respectivamente (IUPS Thermal Commission, 2001). Los límites entre homeotermos y
poiquilotermos son considerablemente borrosos, teniendo en cuenta que muchos
homeotermos exhiben una variación estacional de su temperatura corporal (Prinzinger et
al., 1991).
A su vez, podemos hacer una distinción entre organismos termorreguladores y
termoconformes. Siendo un perfecto termorregulador el organismo que mantiene una
temperatura corporal invariable a lo largo del tiempo aunque las temperaturas del ambiente
puedan variar. Por otro lado, un termoconforme poseerá un amplio rango de tolerancia
térmica, permitiéndole mantener su temperatura corporal cercana a la del medio ambiente

10
(Angilletta, 2009). Entre estas dos estrategias extremas existen intermedios, donde los
organismos tienen un compromiso entre el tiempo destinado a la termorregulación y el
empleado en el resto de las funciones biológicas (Huey y Slatkin, 1976).
Los termorreguladores, para mantener su temperatura corporal dentro un
determinado rango, requieren cambios conductuales, como movimientos entre sitios que
ofrecen microclimas adecuados para su calentamiento y enfriamiento (Withers y Campbell,
1985; Huey et al., 1989; Schultz, 1998; Díaz y Cabezas-Díaz, 2004). Este valor estable de
una variable, en un organismo saludable, por medio de su propia regulación, se denomina
“Set point” (Hertz et al., 1993).
Más aún, un termorregulador mantiene una media o varianza particular de su
temperatura corporal utilizando un mecanismo neural para percibir y responder a su
ambiente (Bicego et al., 2007).
Esta regulación comportamental de la temperatura posee un rol preponderante en los
organismos pertenecientes al orden Squamata, ya que las funciones fisiológicas y
conductuales dependen de la temperatura corporal adquirida, afectando a los organismos en
su distribución espacial a diferentes escalas (Hutchinson, 1957; Haramura, 2007), actividad
estacional y diaria (Huey y Stevenson, 1979), desempeño locomotor (salto, natación,
locomoción) (Marvin, 2003; Aguilar y Cruz, 2010), tasa de crecimiento (Lillywhite et al.,
1973), tasa de digestión y comportamiento termofílico (Witters y Sievert, 2001); entre
otros.
Los animales pertenecientes al orden Squamata pueden además, disminuir su tasa
metabólica mediante un descenso de su temperatura corporal o aumentar la energía
destinada al crecimiento y la reproducción, consumiendo y digiriendo más alimento cuando
las presas son abundantes (Keith et al., 1984; Du et al., 2000; Wang et al., 2003; Kooijman,

11
2010). Este incremento en la tasa digestiva permite comidas frecuentes, lo cual es
acompañado por un incremento de la temperatura corporal (Du et al., 2000; Wang et al.,
2003).
Consecuentemente, la selección de temperaturas más elevadas dentro de su rango de
temperaturas selectas, les confiere un beneficio ya que la búsqueda, captura, digestión y
asimilación de los alimentos dependen fuertemente de la temperatura corporal (Greenwald
y Kanter, 1979; Waldschmidt et al., 1986; Angilletta, 2001). Si el alimento no estuviera
disponible ya sea porque se encuentra en regiones subóptimas o con una alta tasa de
depredación; el costo-beneficio de alimentarse haría que estos organismos seleccionen
temperaturas óptimas más bajas (Brett 1971; Elliott, 1982; Jonassen et al., 1999).
Este comportamiento efectivamente se observa en algunas especies de lagartos
como: Anolis carolinensis (Brown y Griffin, 2005), Crotaphytus collaris (Sievert, 1989);
en un gecko del género Eublepharis (Autumn y De Nardo, 1995) como así también en un
integrante de la familia Pygopodidae, Lialis burtonis (Bradshaw et al., 1980).
No obstante, Brown y Griffin (2005), han argumentado que la magnitud de este
efecto probablemente es muy pequeña para representar un ahorro energético significativo
en ectotermos como los reptiles. De hecho, no se ha comprobado este patrón en el lagarto
Sceloporus jarrovi (Schuler et al., 2001).
Todas estas respuestas derivan de las sensibilidades térmicas de los procesos
anabólicos y catabólicos, las cuales varían de acuerdo a la cantidad de comida que puede
ser procesada, digerida y absorbida por el estómago (Schuler et al., 2011). En tales
procesos, la actividad enzimática está estrechamente vinculada con la temperatura corporal
y se ve aumentada con el ascenso de la misma (Müller-Esterl, 2008). Los organismos
provenientes de regiones cálidas evolucionaron con niveles enzimáticos estables y ciertas

12
estructuras bioquímicas alcanzan su estabilidad funcional a altas temperaturas (Hochochka
y Somero, 2002).
La digestión es el proceso mediante el cual se realiza la conversión de sustancias
complejas en otras más simples para favorecer su absorción. Los alimentos sufren
transformaciones físicas y químicas a lo largo del tracto digestivo. Particularmente, una vez
arribado al intestino delgado, el alimento es degradado por secreciones biliares y enzimas
pancreáticas (Blanco, 2006). Las enzimas digestivas exócrinas del páncreas en reptiles,
incluyen compuestos amilolíticos (amilasa), proteolíticos (quimotripsina, tripsina,
carboxipeptidasa y elastasa), lipolíticos y quitinasa, en reptiles insectívoros. Las enzimas
producidas por el páncreas son altamente dependientes de la temperatura corporal y del pH,
el cual debe ser mayor a 6.0. (Yarto Jaramillo, 2011).

13
Objetivo
Objetivo General Determinar la existencia de un patrón de comportamiento termofílico en relación a
la digestión, en el lagarto L. darwinii.
Objetivos particulares Comparar la temperatura corporal selecta por L. darwinii en un gradiente térmico
linear, a lo largo de su horario de actividad, bajo diferentes condiciones
experimentales (ayuno vs. no ayuno).
Medir la concentración de la enzima tripsina, a tres temperaturas ecológicamente
relevantes, establecidas a partir de las mediciones de las temperaturas corporales
selectas.

14
Área de estudio
El área denominada Médanos Grandes se
encuentra ubicada a 42 km al sureste de la
ciudad de San Juan, en el departamento de
Caucete. Comprende una amplia región de
aproximadamente 2000 Km2.
El sitio de colecta seleccionado fue la
Estación Guayamas, ubicada al norte del
Sistema de Médanos Grandes (31.7445 Lat. S,
68.1045 Long. O; 670 m.s.n.m.).
Fig. 1: Caucete, provincia de San Juan
(tomado de www.wikimedia.org,
Autor: Enrique Guardia, 2011).
Clima Conforme a la Ecorregión del Monte, las precipitaciones se encuentran concentradas
en verano, con una variación anual entre 80 y 250 mm en promedio (Cabrera, 1976). En el
área de estudio en particular, la precipitación tiene un promedio anual de 89 mm. La
temperatura máxima media anual es de 25.7 °C y la mínima media anual es de 10.4°C
(Cabrera, 1976).

15
Fauna La fauna de insectos es bien conocida en la zona norte del Monte, donde existe una
alta proporción de géneros y especies endémicas pertenecientes a diferentes familias (Roig-
Juñent et al., 2001).
Entre los reptiles más representativos se encuentran la iguana colorada (Tupinambis
rufescens); la falsa yarará (Pseudotomodon trigonatus), la yarará ñata (Bothrops
ammodytoides), la falsa coral (Lystrophis semicinctus); lagartos como Liolaemus darwinii,
L. gracilis, los matuastos Liosaurus paronae y L. belli; los teidos Cnemidophorus
longicauda y Teius oculatus. También está presente la tortuga terrestre Chelonoidis
chilensis. Respecto a los anfibios, podemos mencionar a la ranita del monte (Pleurodema
nebulosa) y el sapo común, Rhinella arenarum, (Chebez, 1988; Bertonatti y González,
1992; Chebez, 1994; García Fernández et al., 1997).
Las aves incluyen gauchos (Agriornis sp.), dormilonas (Muscisaxicola sp.), la
martineta común (Eudromia elegans), la monterita canela (Poospiza ornata), el inambú
pálido (Nothura darwinii) y el loro barranquero, Cyanoliseus patagonus, (García Fernández
et al., 1997).
Los mamíferos están representados por especies de tamaño grande como el guanaco
(Lama guanicoe) y el puma (Felix concolor); por especies de tamaño mediano como la
vizcacha (Lagostomus maximus), el zorro colorado (Lycalopex culpaeus) y el zorro gris (L.
griseus); y por especies de tamaño pequeño como los cuises (Microcavia australis, Galea
musteloides), los tuco-tucos (Ctenomys mendocinus), el zorrino chico (Conepatus
castaneus) y el huroncito, Lyncodon patagonicus, (García Fernández et al., 1997; Díaz y
Ojeda, 2001).

16
Flora El tipo de vegetación predominante es el matorral o la estepa arbustiva xerófila,
sammófila o halófila. Desde el punto de vista florístico la provincia se caracteriza por la
presencia, casi constante, de especies del género Larrea (L. divaricata y L. cuneifolia) y
Prosopis (P. flexuosa, P. alpataco) arbustivos. Otros géneros de Zigofiláceas como
Bulnesia y Plectrocarpa sólo se hallan en la parte norte de la Provincia.
Además, se encuentran cactáceas como Tephrocactus articulatus y Opuntia
sulphurea (Cabrera, 1976).

17
Descripción de la especie
Dentro del género Liolaemus hay un grupo de especies que se caracteriza por la
presencia de un parche de escamas agrandadas en la cara posterior del fémur, este conjunto
de especies es llamado “grupo de L. boulengeri”. Integrando este grupo de especies, se
reconoce entre otros, un clado monofilético denominado grupo L. darwinii, al cual
pertenece el lagarto bajo estudio con el mismo nombre, Liolaemus darwinii (Fig. 2 y 3).
Este grupo, se caracteriza entre otras particularidades, por presentar las coronas de los
dientes posteriores con bordes rectos y un marcado dicromatismo sexual (Etheridge, 1993).
La especie L. darwinii, de pequeño a mediano tamaño corporal (LHC: 65 mm), es
común y típica de ambientes de Monte. Su distribución es muy amplia, desde La Rioja
hasta Chubut. (Cruz et al., 2012). En cuanto a su estado de conservación, se encuentra
categorizada como No Amenazada (NA) (Abdala et al., 2012).
Los machos pueden tener una coloración vivaz que incluye manchas pectorales,
ventrales y en muslos intensamente negras. Las hembras, (como en muchas especies del
género Liolaemus) son más pálidas (Cruz et al., 2012).

18
Fig. 2: Macho adulto de Liolaemus darwinii, de los Médanos Grandes de Caucete,
provincia de San Juan (Foto: Eduardo A. Sanabria, 2013).
Fig. 3: Hembra y macho adultos de Liolaemus darwinii, de los Médanos Grandes de
Caucete, provincia de San Juan (Foto: Andrea A. Barauna, 2013).

19
Materiales y Métodos
Captura Los ejemplares (machos y hembras adultos) fueron capturados durante las primeras
horas de su horario de actividad en los meses de octubre 2012 (n=29) y marzo 2013 (n=20),
realizando caminatas al azar por el área de estudio. Se utilizó un lazo hecho de hilo dental
sujeto a una vara de 2 metros de largo aproximadamente.
Inmediatamente después de la captura se les midió la temperatura corporal en
campo con un termómetro digital TES 1312 (TES Electrical Electronic Corp., Taipei,
Taiwan, ± 0.1ºC). También se registró temperatura del aire a 1 cm de la superficie, del
sustrato al sol y a la sombra, y la velocidad del viento con un anemómetro digital (Kaise
AR 816, ±0.2 ºC – 0.1 m/s). Estos datos se tomaron con la finalidad de conocer las
condiciones ambientales a las que se expone el animal, en los momentos de captura del año,
y así poder hacer una mejor recreación en laboratorio.
La época del año selecta para la captura, está relacionada con el comienzo y el fin
del periodo anual de actividad de los lagartos, teniendo en cuanta su ciclo circanual (Hill et
al., 2004).
Trabajo de Laboratorio Los lagartos capturados en ambos meses, se separaron en dos grupos y fueron
sometidos simultáneamente a tratamientos opuestos, ayuno vs. no ayuno (Tabla 1), a lo
largo de cuatro días.

20
OCTUBRE 2012 MARZO 2013
AYUNO 15 10
NO AYUNO 14 10
Tabla 1: Cantidad de individuos (n) por mes
y por tratamiento
Durante la fase de tratamiento se los colocó en recintos de plástico individuales para
asegurar que cada animal comiera el alimento que se le suministró. Cada recinto contaba
con una provisión de arena, para imitar su sustrato natural. Se los mantuvo a temperatura
ambiente (24°C ± 2°C), respetando su fotoperíodo natural 12:12 (horas luz/oscuridad).
Diariamente, se rociaba a los individuos con agua, para evitar una posible deshidratación.
A cada individuo del grupo “no ayuno”, alrededor del mediodía, se le proporcionó
por día una larva o una pupa de Tenebrio sp. (1,40% de su masa corporal). Previamente, se
registró el peso de cada larva o pupa, para llevar un control de la ingesta de los animales.
Medición de la temperatura corporal selecta Concluida la etapa de aclimatación a los tratamientos, se dispuso a los individuos en
un terrario con carriles de 1,5 metros de largo. El terrario posee paredes de vidrio. Por
debajo, corren placas térmicas, provistas de sensores que permiten mantener un gradiente
linear térmico entre 25-55°C. Este rango es ampliamente conservado en el género
Liolaemus (Medina et al., 2009). Las placas están aisladas del vidrio por una plancha de
telgopor, la cual a su vez está cubierta por arena, sirviendo ésta como sustrato para los
animales testeados.
El terrario se ubicó en un lugar cerrado, separado del ruido y de cualquier otra
perturbación que pudiera alterar el comportamiento de los lagartos. La temperatura

21
ambiente se mantuvo constante (20°C ± 2°C) con la ayuda de un aire acondicionado. La
iluminación también fue constante durante el experimento, provista por luz blanca.
Los lagartos fueron previamente aclimatados al carril, de 07:00 a 08:00 hs., y luego
se comenzaron a registrar sus temperaturas corporales selectas cada media hora durante
todo su periodo diario de actividad (08:00 a 20:00 hs). Las temperaturas corporales selectas
se registraron con un termómetro digital TES 1312 (TES Electrical Electronic Corp.,
Taipei, Taiwan, ± 0.1°C), que cuenta con una termocupla tipo sonda (TPK-01) de
aproximadamente 0,8 mm de espesor. Esta se introducía en el orificio cloacal de los
animales (~5 mm).
Se eligió este método porque la mejor evidencia de las preferencias térmicas de un
organismo, provienen de la medición de su temperatura corporal selecta en un gradiente
termal, donde lo único que varía espacialmente es la temperatura (Angilletta, 2009).
Controlando las variables ambientales, fácilmente se puede cuantificar la frecuencia
de las temperaturas selectas. La temperatura corporal selecta provee una valiosa
información sobre las estrategias termorregulatorias en ambientes donde no existen
restricciones ecológicas (Angilletta, 2009).
Obtención de tejidos y plasma Se mantuvo a quince individuos en incubadora a 25°C, con fotoperíodo de 12:12
(horas luz/oscuridad), durante 5 días, con el fin de producir su vaciamiento gástrico.
Posteriormente, se los dividió en 3 grupos de 5 ejemplares cada uno, fueron colocados en
recipientes individuales y alimentados ad libitum con larvas de Tenebrio sp., mientras se
incubaba a cada grupo durante 3 días a 15°, 33° y 40°C.

22
Se eligieron estas tres temperaturas de incubación en base a la frecuencia de
temperaturas seleccionadas en el gradiente linear térmico.
Cumplido el tiempo de incubación, los ejemplares se sacrificaron mediante una
sobredosis de anestesia (2.5 ml de 2% Xylocaine y 2% Lidocaine HCl, Astra Zeneca Labs,
Buenos Aires, Argentina) la cual fue inyectada vía intramuscular a la altura del abdomen.
Posteriormente se disecaron y se les extrajo sangre y órganos (páncreas e intestino
delgado).
La sangre se extrajo directamente del corazón y se colectó con capilares
heparinizados (para evitar su coagulación). Luego, se centrifugó a 2000 rpm durante 15
minutos. Finalizado el centrifugado, se separó el plasma y se lo congeló a -20°C, para
posteriormente medir la concentración de proteínas totales y albúmina.
Esta medición tiene la finalidad de conocer los niveles séricos de proteínas en el
momento de la extracción de los órganos. Es preciso saber si la relación entre la albúmina y
las globulinas es normal, pues la sumatoria de estas concentraciones equivale a la
concentración de las proteínas totales (Blanco, 2006).
Los órganos extraídos para el análisis fueron: la porción anterior del intestino
delgado (duodeno) y el páncreas. El primero de ellos se cortó longitudinalmente, para su
ulterior lavado de la cavidad del lúmen con cloruro de sodio (NaCl) al 1% (solución
fisiológica), durante 1-2 minutos. Luego se lo secó en papel secante y se lo almacenó a
-20ºC. El páncreas se extrajo y almacenó a -20°C.
Actividad enzimática en intestino delgado y páncreas Ambos órganos se homogenizaron para la determinación de la actividad enzimática.
Para la homogeneización del intestino delgado y páncreas, se utilizó un homogenizador

23
(Whirlotone N° Cat.: 59880-10, Matheson Scientific, USA), equipado con una sonda
(S25N-18G).
El intestino delgado se descongeló, se pesó y se suspendió en una solución de tritón
X100 (Sigma Aldrich, N° Cat.: T9284, Industria Argentina) al 10%, en una proporción de 6
ml por cada gramo de tejido. Luego de homogeneizar, se lo centrifugó a 14.000 rpm
durante 60 minutos a 4°C. Se colectó el sobrenadante para el análisis y se almacenó a
-80°C.
El páncreas se descongeló, pesó y se suspendió en una solución 154 mM de NaCl
(Dalton, N° Cat.: 7647, Industria Argentina), en una proporción 1 a 4 (volumen de tejido:
volumen de solución). Luego de homogeneizar se centrifugó a 11.400 rpm durante 20
minutos a 4°C. Posterior al centrifugado, se separó el sobrenadante y también fue
almacenado a -80°C.
La medición de la actividad enzimática se realizó en los sobrenadantes obtenidos en
el paso previo. El protocolo utilizado fue el del Kit Thermo Scientific Pierce Protein
Biology products, USA. Este Kit de Ensayo Colorimétrico de Proteasa (Parte Nº23263)
utiliza la caseína completamente succinilada como un sustrato para el ensayo. La hidrólisis
de este sustrato en presencia de la proteasa, resulta en la liberación de fragmentos de
péptidos con grupos amino terminales libres.
Los péptidos reaccionan con ácido sulfónico trinitrobenceno (TNBSA), dando como
resultado productos de color amarillo (TNB-péptido), los cuales se detectaron a una
longitud de onda de 500 nm en un lector de placa Thermo Scientific Multiskan FC (Thermo
Fisher Scientific, Shangai, China).
El procedimiento se realizó según indicaciones del fabricante (Rao S.K. et al., 1997;
Tian M. et al., 2004).

24
El valor obtenido de actividad enzimática en cada órgano fue relativizado al peso
del mismo previo a la homogenización, expresado en: UI/ml/gr (Unidades
Internacionales/mililitro/gramo).
Concentración de proteínas totales y albúmina
Determinación de proteínas totales Se utilizó un método colorimétrico, en el que los enlaces peptídicos de las proteínas
reaccionan con el ión cúprico, en medio alcalino, para dar un complejo color violeta con
máximo de absorción a 540 nm, cuya intensidad es proporcional a la concentración de
proteínas totales en la muestra.
Procedimiento En un primer tubo marcado como Blanco (B), se colocaron 50 ul de agua destilada y
3,5 ml de Reactivo A (complejo EDTA/Cu 13 mmol/l en hidróxido de sodio 875 mmol/l y
alquil aril poliéter, AAP). En un segundo tubo marcado como suero patrón (P) se colocaron
50 ul de de la misma (Suero Patrón: solución de albúmina y globulinas de origen bovino,
con título conocido de proteínas) y 3,5 ml de Reactivo A. Luego, se procesó la cantidad de
tubos necesarios para la cantidad de condiciones experimentales previstas. Se colocaron 50
ul de la muestra (plasma de los lagartos) y 3,5 ml de Reactivo A en cada tubo. El volumen
final de la reacción fue de 3,55 ml.
Se mezcló cada preparado con una varilla y se incubó a 37°C durante 10 minutos. Se leyó
en un espectrofotómetro (Sunostik SBA 733, China) a 540 nm y se llevó a cero con el Blanco de
Reactivo.

25
Determinación de albúmina La albúmina reacciona específicamente (sin separación previa) con la forma
aniónica de la 3,3',5,5'-tetrabromo cresolsulfon ftaleína (BCF), en presencia de un exceso
de colorante, en medio tamponado a pH 3,8. El aumento de absorbancia a 625 nm respecto
del Blanco de reactivo, es proporcional a la cantidad de albúmina presente en la muestra.
Procedimiento Para este caso, se destinó un tubo para el procesamiento de el blanco de reactivo, en
donde se colocaron 3,5 ml de Reactivo B (solución de 3,3',5,5'-tetrabromo cresolsulfon
ftaleína, en polioxietilén lauril éter); otro tubo para el procesamiento del suero patrón de
concentración conocida, en donde se colocaron 10 ul de la misma y 3,5 ml de Reactivo B; y
finalmente, un tubo para cada condición experimental a medir (desconocido), en donde se
colocaron 10 ul de la muestra y 3,5 ml de Reactivo B.
Nuevamente, se mezcló cada preparado con una varilla y se mantuvieron a 25°C
durante 10 minutos. Se leyó en el espectrofotómetro a 625 nm y se llevó a cero con el
Blanco de Reactivo.
En este caso, también se preparó una curva de calibración con cantidades crecientes
de suero patrón (10 y 20 ul), con un volumen de reactivo de 3,5 ml en todos los casos.
Curva de calibración Para constatar que el colorímetro tenga una respuesta lineal en las longitudes de
onda fijadas para las reacciones, se prepararon curvas de calibración para las proteínas
totales (Fig. 4a) y la albúmina (Fig. 4b) con cantidades crecientes de suero patrón (50 y
100 ul) y un volumen de reactivo de 3,5 ml en todos los casos.
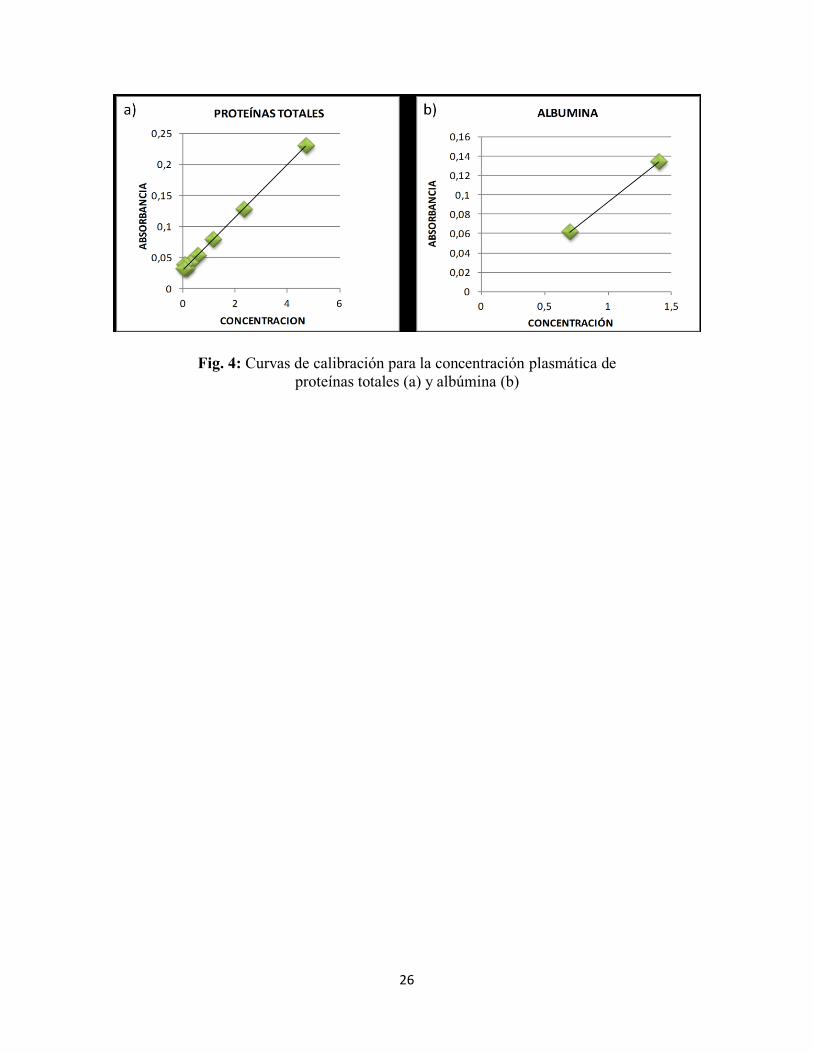
26
Fig. 4: Curvas de calibración para la concentración plasmática de
proteínas totales (a) y albúmina (b)

27
Análisis de datos
Para el análisis de los datos obtenidos del comportamiento termofílico de los
lagartos, se realizaron comparaciones de la temperatura corporal selecta entre los grupos
propuestos en las dos experiencias (ayuno vs. no ayuno). En el caso de los análisis
enzimáticos, se comparó la actividad enzimática a las diferentes temperaturas presentadas
para el estudio (15, 33, 40ºC).
En ambos casos se aseguró la independencia estadística de los datos. La normalidad
se comprobó mediante la prueba de Kolmogorov-Smirnov y se realizó un análisis de la
varianza con el peso corporal como covariable (ANCOVA). La homocedasticidad se
demostró mediante la prueba de Levenne. Para todos los análisis el nivel de significancia
fue de α=0,05. De todos los datos muestrales se extrajeron los estadísticos descriptivos de
media y desviación estándar. Todos los análisis se efectuaron con el paquete estadístico
STATISTICA 7.0.
28
Resultados
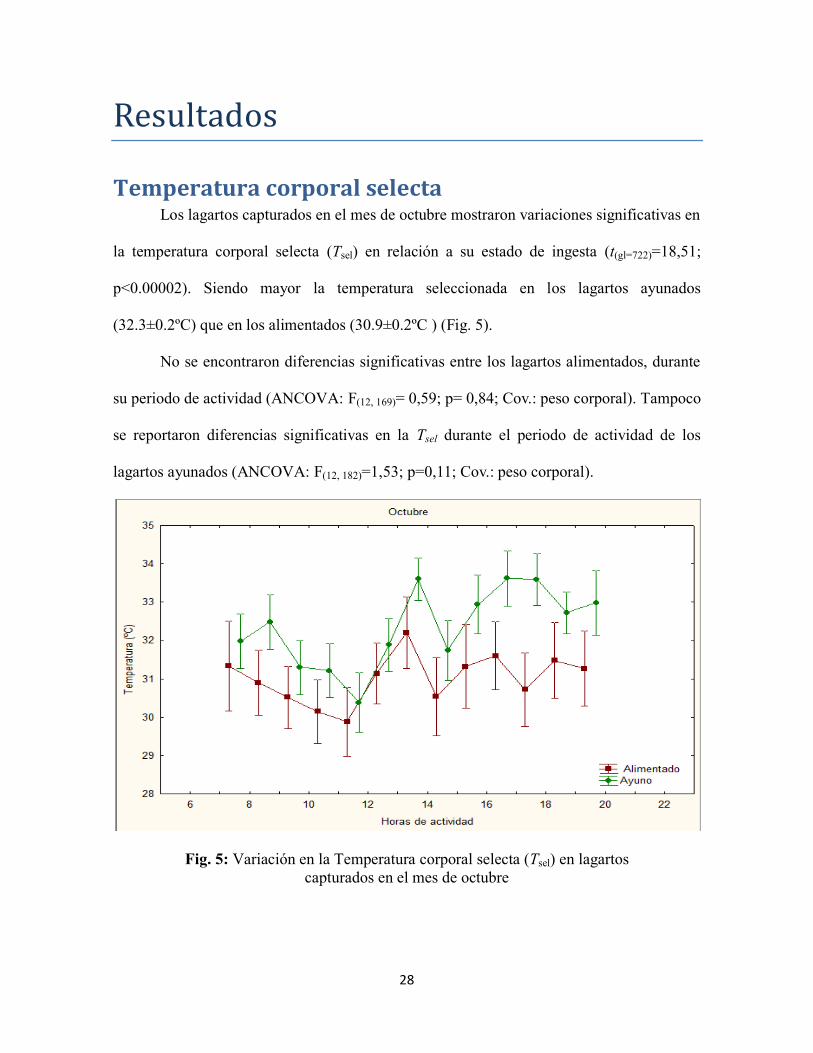
Temperatura corporal selecta Los lagartos capturados en el mes de octubre mostraron variaciones significativas en
la temperatura corporal selecta (Tsel) en relación a su estado de ingesta (t(gl=722)=18,51;
p<0.00002). Siendo mayor la temperatura seleccionada en los lagartos ayunados
(32.3±0.2ºC) que en los alimentados (30.9±0.2ºC ) (Fig. 5).
No se encontraron diferencias significativas entre los lagartos alimentados, durante
su periodo de actividad (ANCOVA: F(12, 169)= 0,59; p= 0,84; Cov.: peso corporal). Tampoco
se reportaron diferencias significativas en la Tsel durante el periodo de actividad de los
lagartos ayunados (ANCOVA: F(12, 182)=1,53; p=0,11; Cov.: peso corporal).
Fig. 5: Variación en la Temperatura corporal selecta (Tsel) en lagartos
capturados en el mes de octubre
29
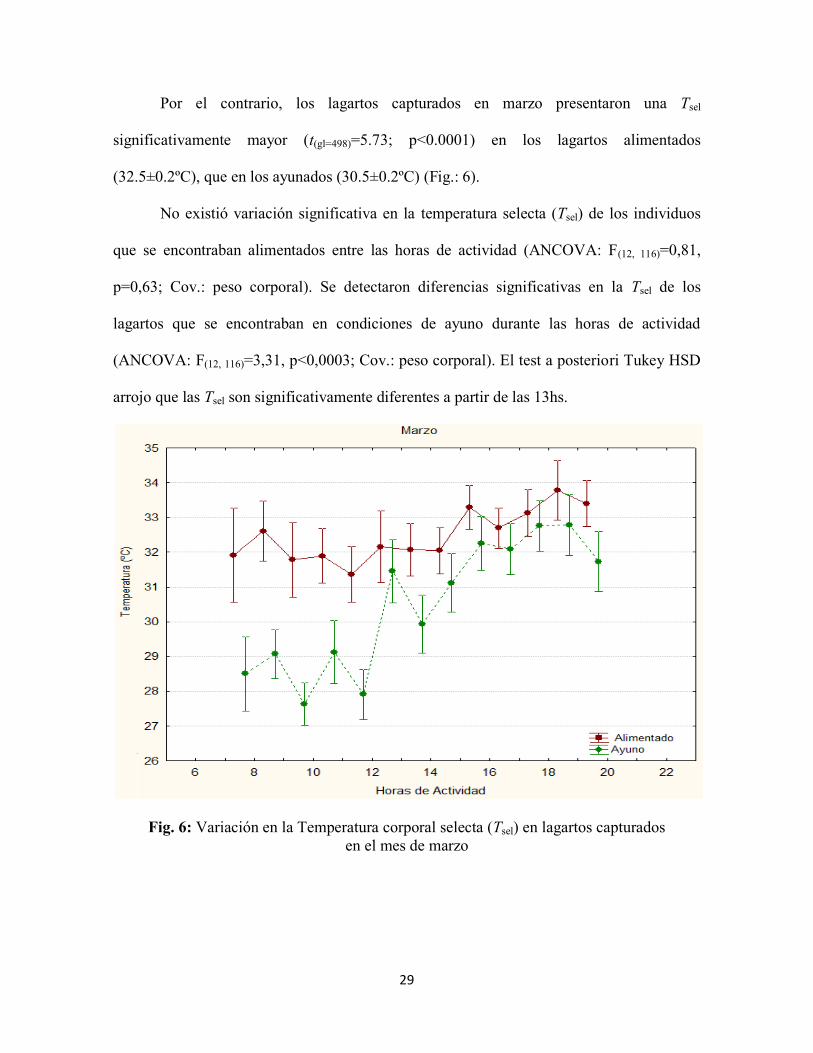
Por el contrario, los lagartos capturados en marzo presentaron una Tsel
significativamente mayor (t(gl=498)=5.73; p<0.0001) en los lagartos alimentados
(32.5±0.2ºC), que en los ayunados (30.5±0.2ºC) (Fig.: 6).
No existió variación significativa en la temperatura selecta (Tsel) de los individuos
que se encontraban alimentados entre las horas de actividad (ANCOVA: F(12, 116)=0,81,
p=0,63; Cov.: peso corporal). Se detectaron diferencias significativas en la Tsel de los
lagartos que se encontraban en condiciones de ayuno durante las horas de actividad
(ANCOVA: F(12, 116)=3,31, p<0,0003; Cov.: peso corporal). El test a posteriori Tukey HSD
arrojo que las Tsel son significativamente diferentes a partir de las 13hs.
Fig. 6: Variación en la Temperatura corporal selecta (Tsel) en lagartos capturados
en el mes de marzo

30
Actividad enzimática en intestino delgado y páncreas En mediciones realizadas en intestino delgado no se observaron diferencias
significativas a las diferentes temperaturas de aclimatación (Kruskal-Wallis test: H (2,
N=11)=2,09; p=0,35; Fig. 7a). En contraste, las mediciones realizadas en tejido pancreático
mostraron una mayor actividad enzimática a 40°C, siendo esta estadísticamente diferente
con la actividad obtenida a los 15°C, (Kruskal-Wallis test: H (2, N=9)=6,44; p=0,039; Fig.
7b).
Fig. 7: Variación de la actividad enzimática de tripsina en intestino delgado (a) y
páncreas (b) en función a la temperatura de incubación
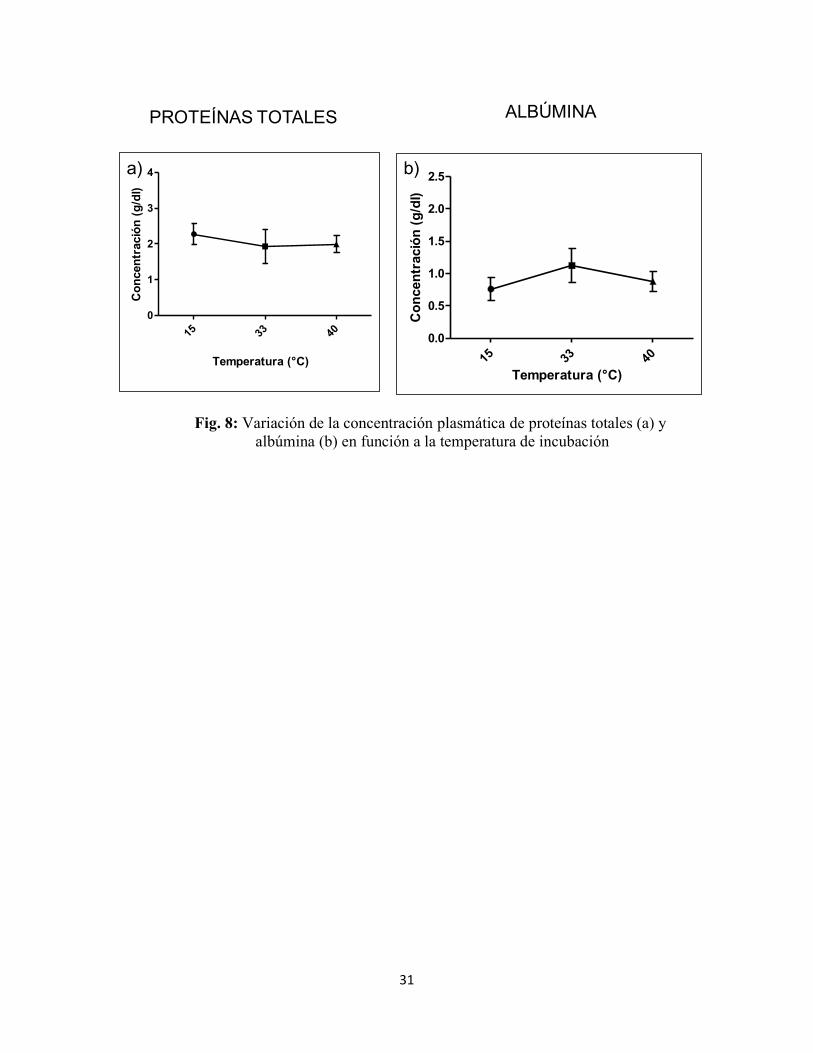
Concentración de proteínas totales y albúmina No se lograron detectar diferencias significativas para los niveles de proteínas
totales en relación a las temperaturas de incubación seleccionadas (Kruskal-Wallis test: H
(2, N=15)=1,52; p=0,46. Fig. 8a). De la misma manera, no se encontró diferencia significativa
en los niveles de albúmina en relación a las temperaturas de incubación seleccionadas
(Kruskal-Wallis test: H (2, N=15)=1,35; p=0,5. Fig. 8b).
15 33 40
0
10
20
30
40
50
Temperatura (°C)
Acti
vid
ad
de T
rip
sin
a (
UI/m
l/g
r)
15 33 40
0
1000
2000
*
Temperatura (°C)
Acti
vid
ad
de T
rip
sin
a (
UI/m
l/g
r)
INTESTINO DELGADO PÁNCREAS
a) b)
31
Fig. 8: Variación de la concentración plasmática de proteínas totales (a) y
albúmina (b) en función a la temperatura de incubación
15 33 40
0
1
2
3
4
Temperatura (°C)
Co
ncen
tració
n (
g/d
l)
PROTEÍNAS TOTALES
15 33 40
0.0
0.5
1.0
1.5
2.0
2.5
Temperatura (°C)
Co
ncen
tració
n (
g/d
l)
ALBÚMINA
a) b)

32
Discusión
Temperatura corporal selecta Muchos estudios sugieren que existe un comportamiento termofílico en ectotérmos
luego de alimentarse (Witters y Sievert, 2001), mientras que otros sostienen que no existe
dicho efecto (Schuler et al., 2011).
Los lagartos capturados en el mes de octubre, exhibieron una temperatura corporal
selecta significativamente más elevada en condiciones de ayuno que de alimentación, en
promedio. Estos resultados contradicen los modelos de termorregulación óptima, basados
en costos y beneficios energéticos, los cuales predicen que los animales disminuirán su
temperatura corporal selecta cuando la disponibilidad de alimento sea restringida (Huey,
1982).
En contraste, los resultados obtenidos para este caso, implican que los lagartos
expuestos a un déficit alimentario estarían afinando su comportamiento termorregulatorio
hacia beneficios no energéticos como el desarrollo, la inmunidad y la locomoción.
Probablemente el crecimiento se vea comprometido, como consecuencia del costo que
insume mantener temperaturas corporales elevadas durante condiciones de ayuno (Schuler
et al., 2011).
Los beneficios no energéticos están bien documentados en ectotérmos (Angilletta,
2009). La necesitad de capturar presas y evitar depredadores es un fuerte incentivo para
mantener elevadas temperaturas que permitan la locomoción (Schuler et al., 2011).
Esta situación no puede ser mantenida en el tiempo, y cuando estos lagartos se
enfríen por debajo de su óptimo térmico, su habilidad para localizar y dominar presas
disminuirá, tal como describen Greenwald (1974), Díaz (1994), y Van Damme y colegas

33
(1991). Además, estos animales fríos no podrían escapar efectivamente de un depredador
(Christian y Tracy, 1981; Cooper, 2000), y deberían detectarlos tempranamente o adoptar
defensas alternativas (Rand, 1964; Hertz et al., 1982; Goode y Duvall, 1989; Passek y
Gilligham, 1997 y Shine et al., 2000).
Por el contrario, los lagartos capturados en el mes de marzo, seleccionaron en
promedio, temperaturas significativamente más elevadas en condiciones de alimentación
que de ayuno. Los animales ayunados tienen mayor necesidad de forrajear que los nutridos
(Schuler et al., 2011). Esto representaría una dificultad para los primeros, ya que una
depresión térmica podría perjudicar su habilidad para defender recursos alimenticios de sus
conspecíficos (Garland et al., 1990; Robson y Miles, 2000).
Las diferencias estacionales encontradas, podrían deberse al ritmo biológico de los
reptiles. Estos ritmos son variaciones fisiológicas y conductuales rítmicas a lo largo de un
día (ciclo circadiano) o de un año (ciclo circanual). Los ritmos son endógenos y persisten
incluso cuando se coloca a los animales en un ambiente de laboratorio, privados de
información ambiental sobre la época del año (Hill et al., 2004). Entre los ciclos
circanuales de interés para este estudio, se encuentran la acumulación de grasa y el
aletargamiento (Hill et al., 2004).
Los lagartos colectados en octubre están saliendo del periodo de letargo, y los
reptiles ayunados seleccionarían temperaturas corporales selectas elevadas para poder
llevar a cabo una captura de presas potenciales más eficiente. Los alimentados
seleccionarían temperaturas más bajas para retener más tiempo la comida en el tracto
digestivo y asegurar una buena digestión.
Los lagartos de marzo seleccionaron temperaturas corporales selectas más bajas
durante horas de la mañana. Probablemente esta variación diaria de la temperatura corporal

34
selecta podría deberse a que los lagartos en esta época del año forrajean durante horas de la
tarde (de 13 a 20 hs.) donde las temperaturas son mayores y quizás la disponibilidad de
alimento también. Por otro lado, la selección de temperaturas más bajas durante la mañana
les permitiría resguardar el alimento ya al disminuir su tasa metabólica desciende también
el gasto energético.
Los lagartos capturados en marzo, están por entrar en un periodo de aletargamiento.
La selección de temperaturas elevadas en lagartos alimentados, podría ser explicada por la
necesidad por fijar una mayor cantidad de reservas en forma de grasa, antes de que llegue el
periodo de baja disponibilidad de recursos alimenticios. Los lagartos ayunados se
ahorrarían de perder las bajas reservas de grasa presentes en su cuerpo, seleccionando
temperaturas corporales más bajas.
En este estudio, se les presentó a los lagartos un conflicto entre termorregulación y
alimentación, donde se vieron expuestos a riesgos de supervivencia y/o fecundidad. Hacer
un correcto análisis de los costos y beneficios de la termorregulación requiere un gran
esfuerzo por el hecho de que los organismos utilizan diversos mecanismos para
termorregular y cada uno tiene su costo particular (Lustick, 1983; Chappell y Whitman,
1990).
Actividad enzimática en intestino delgado y páncreas No se hallaron diferencias significativas entre las concentraciones de enzima
tripsina presentes en las porciones de intestino delgado analizadas. En apariencia, la
aclimatación a diferentes temperaturas no tuvo ningún efecto sobre la actividad enzimática
intestinal de los lagartos. Estudios similares realizados en especies pertenecientes a los

35
géneros Dipsosaurus, Uma, Anolis y Gerrhonotus, encontraron que si existía un máximo de
actividad en una enzima fosfatasa alcalina a altas temperaturas (Licht, 1964).
Volviendo a este trabajo, en el tejido pancreático se observó una mayor actividad
enzimática a los 40°C, mostrando una diferencia significativa con la actividad obtenida a
los 15°C.
Esta variación solo observada en páncreas, tiene lógica funcional, ya que este
órgano es el encargado de sintetizar y almacenar esta (y muchas otras) enzimas digestivas
(Hill et al., 2004). Estos resultados, podrían indicar que a bajas temperaturas, la actividad
de tripsina es escasa o nula, debido a que estaría por debajo del óptimo térmico requerido
por la enzima (Müller-Esterl, 2008).
Aparentemente, el aumento de la temperatura conlleva un aumento en la secreción
de tripsina en el páncreas de los lagartos. Se ha observado este incremento de la actividad
enzimática relacionado con la temperatura en reptiles, ya en estudios muy tempranos, como
los realizados por Riddle (1909) en una tortuga del género Emydoidea.
Probablemente la selección de temperaturas mayores durante el ayuno, observada en
los lagartos durante el mes de octubre, los prepararía para la digestión de presas
potencialmente capturadas. El ascenso térmico corporal, registrado en los lagartos
alimentados en marzo, favorecería la digestión de las presas ingeridas durante el forrajeo
(Stevens y Hume, 2004).
También se podría inferir que, a bajas temperaturas se ve desfavorecido el
movimiento peristáltico muscular. Este consiste en movimientos rítmicos, ondulatorios y
automáticos, realizados por el sistema digestivo para hacer avanzar la comida (Hill et al.,
2004). El peristaltismo empieza en el esófago y empuja el bolo alimenticio hacia el
estómago, donde las contracciones musculares y los jugos gástricos ayudan a desmenuzar

36
la comida (Hill et al., 2004). En estudios llevados a cabo en lagartos del género Varanus y
Ctenosaura (Mackay, 1968), se corroboró que las temperaturas corporales elevadas
incrementaban la amplitud y frecuencia de las contracciones peristálticas.
El ascenso de actividad enzimática observado en páncreas, conforme a la
temperatura tiene un límite, pues las altas temperaturas extremas implican un estrés físico
que superan todo efecto positivo en las tasas catalíticas (Angilletta, 2009).
Son necesarios más estudios para entender la variación en el efecto termodinámico.
Hasta ahora, la mayor parte de la evidencia encontrada, sugiere que los organismos se
benefician de la adaptación a altas temperaturas, combinado con una precisa
termorregulación (Angilletta, 2009).
Concentración de proteínas totales y albúmina La albúmina aporta la fuente de aminoácidos esenciales requeridos por el páncreas
para sintetizar enzimas. Como son esenciales, solo llegan al cuerpo de manera externa. Una
deficiencia en la concentración de albúmina plasmática se vería reflejada en una
interrupción de la síntesis enzimática (Blanco, 2006).
No se encontraron diferencias significativas entre los niveles séricos de proteínas
totales y albúmina en los lagartos aclimatados a distintas temperaturas. Esto indicaría que
los individuos de los tres grupos estarían en las mismas condiciones nutricionales (Blanco,
2006). De este modo, se suprimió una posible variable de error en los resultados de
actividad enzimática obtenidos.

37
Conclusiones Generales
Este trabajo contribuye a la comprensión de aspectos termorregulatorios
(comportamentales y fisiológicos) de una especie típica de la Ecorregión del Monte, como
lo es Liolaemus darwinii. Conocer el rol que juega la temperatura en una función
fisiológica como la digestión, permite predecir cómo se verían alteradas otras funciones
como el crecimiento, la reproducción, la fecundidad, entre otros.
La especie L. darwinii es un buen modelo representativo del género Liolaemus, por
su similitud con un gran número de lagartos emparentados, pertenecientes a este género tan
diverso y por su alta plasticidad fenotípica. Los resultados obtenidos con este modelo,
permitirían hacer inferencias sobre los resultados esperados en trabajos futuros.
La Ecorregión del Monte posee escasas precipitaciones anuales y una marcada
estacionalidad. Si nos paramos frente a un escenario de inminente cambio climático, las
especies de flora y fauna presente en regiones desérticas, son más vulnerables ya que están
sujetas a una gran amplitud térmica. Un pequeño cambio en la temperatura media anual en
la región, podría provocar el desplazamiento de las especies más íntimamente relacionadas
y dependientes de la temperatura ambiente.
Para el caso de la medición de la temperatura corporal selecta, en el mes de octubre
obtuvimos resultados inversos a los esperados, pero en el mes de marzo, los lagartos
mostraron un comportamiento coincidente con la mayor parte de la bibliografía analizada
en este trabajo.
Sería interesante evaluar qué ocurre en los meses intermedios, e incluso, en las
épocas del año que se encuentran inactivos.

38
Respecto a los análisis de actividad enzimática, los resultados obtenidos fueron
lógicos y compatibles con lo esperado. Aclimatar a los animales a temperaturas más altas
(cercanas a la crítica), nos llevaría a visualizar el punto de inflexión por sobre el cual la
actividad enzimática ya no es favorecida por el ascenso térmico, y podría tener
consecuencias letales.

39
Bibliografía
-Abdala C.S., Acosta J.L., Acsta J.C., Alvarez B.B., Arias F., Ávila L.J., Blanco G.M.,
Bonino M., Boretto J.M., Brancatelli G., Breitman M.F., Cabrera M.R., Cairo S., Corvalan
V., Hernando A., Ibarguengoytia N.R., Kacoliris F., Laspur A., Montero M., Morando M.,
Pelegrin N., Fulvio-Perez C.H., Quinteros S., Semhan R.V., Tedesco M.E., Vega L., Zalba
S.M., 2012. Categorización del estado de conservación de las lagartijas y anfisbenas de la
República Argentina: 215-302. En: A. Giraudo Ed., Categorización del estado de
conservación de la herpetofauna de la República Argentina. Cuadernos de herpetología Vol
26, Suplemento 1. Revista de la Asociación herpetológica argentina. Jujuy.
-Aguilar R. y Cruz F. B., 2010. Refuge Use in a Patagonian Nocturnal Lizard, Homonota
darwini: The Role of Temperature. Journal of Herpetoltogy, Vol. 44, No. 2, pp. 236–241,
2010
-Angilletta M.J. Jr., 2009. Thermal adaptation: A theorical and Empirical Synthesis. Oxford
University Press Inc., New York. Cap. 4, Pág. 88-125.
-Angilletta M.J. Jr., 2001. Thermal and physiological constraints on energy assimilation in
a wide spread lizard (Sceloporus undulatus). Ecology 82, 3044–3056.
-Autumn, K. y De Nardo, D.F., 1995. Behavioral thermoregulation increases growth rate in
a nocturnal lizard. Journal of Herpetology 29, 157–162.
-Bell T., 1843. The zoology of the voyage of H.M.S. Beagle, under the command of
Captain Fitzroy, R.N. during the years 1832 to 1836. Edited and superintended by Charles

40
Darwin, naturalist to the expedition. Part 5. Reptiles. London, Smith, Elder and Co., (1842-
1843), vi + 51 pp.
-Bennet A.F., Hicks J.W. & Cullum A.J., 2000. An experimental test of the
thermoregulatory hypothesis for the evolution of endothermy. Evolution, 54(5), pp. 1768-
1773.
-Bertonatti C. y González F. 1992. “Lista de Vertebrados Argentinos Amenazados de
Extinción”, Boletín Técnico N°8, Buenos Aires, Fundación Vida Silvestre Argentina, Pág.
33.
-Bicego K.C., Barros R.C. y Branco L.G., 2007. Physiology of temperature regulation:
comparative aspects. Comparative Biochemistry and Physiology A 147:616–639.
-Blanco A., 2006. Química Biológica - 8va Ed. - Buenos Aires: El Ateneo. Cap. 11, pág.:
195-212.
-Bradshaw S.D., Gans C. y Saint Girons H., 1980. Behavioral thermoregulation in a
pygopodid lizard, Lialis burtonis. Copeia 1980, 738–743.
-Brett J.R., 1971. Energetic responses of salmon to temperature: a study of some thermal
relations in the physiology and fresh water ecology of Sockeye salmon (Oncorhynchus
nerka). American Zoologist 11, 99–113.
-Brown R.P. y Griffin S., 2005. Lower selected body temperatures after food deprivation in
the lizard Anolis carolinesis. Journal of Thermal Biology 30, 79–83.
-Brown R.P. y Roberts N., 2008. Feeding state and selected body temperatures in the slow-
worm (Anguis fragilis). Herpetological Journal, 18: 59-62.

41
-Cabrera A.L., 1976. Regiones fitogeográficas de la República Argentina. In: ACME (Ed),
Enciclopedia Argentina de Agricultura y Jardinería, Buenos Aires, Argentina.
-Chappell M.A. y Whitman D.W. , 1990. Grasshopper thermoregulation, Pages 43–172 in
R. F. Chapman, and A. Joern, eds. Biology of Grasshoppers. NewYork,Wiley and Sons.
-Chebez J. C., 1988. “El deterioro de la Fauna”, El deterioro del Ambiente en la Argentina
(suelo, agua, vegetación, fauna), FECIC, 497 pp.
-Chebez J. C., 1994. Los que se van. Especies argentinas en peligro, Buenos Aires,
Editorial Albatros, 604 pp.
-Christian K.A. y Tracy, C.R., 1981. The effect of the thermal environment on the ability of
hatchling Galapagos land iguanas to avoid predation during dispersal. Oecologia 49, 218–
223.
-Cruz F.B., Abdala C.S. y Scrocchi G.J., 2012. Libro: Reptiles de La Rioja. ©CRILAR, 1°
Edición. Pág. 36.
-Cooper W.E., 2000. Effect of temperature on escape behavior by an ectothermic
vertebrate, the keeled earless lizard (Holbrookia propinqua). Behaviour137, 1299–1315.
-Cowles R.B. y Bogert C.M., 1944. A preliminary study of the thermal requirements of
desert reptiles. Bull. Am. Mus. Nat. Hist. 83, 265-296.
-Díaz G.B. y Ojeda R.A., 2001. Libro Rojo: Mamíferos amenazados de la Argentina, 2000,
p. 106. Environments, Nº 47, Pág. 77-94.

42
-Diaz J.A., 1994. Effects of body temperature on the predatory behavior of the lizard
Psammodromus algirus hunting winged and wingless prey. Herpetological Journal 4,145–
150.
-Díaz J.A. y Cabezas-Díaz S., 2004. Seasonal variation in the contribution of different
behavioural mechanisms to lizard thermoregulation. Funcional Ecology 18, 867-875.
-Du W.G., Yan S.J. y Ji X., 2000. Selected body temperature, thermal tolerance and
thermal dependence of food assimilation and locomotor performance in adult blue-tailed
skins, Eumeces elegans. J. Therm. Biol. 25, 197-202.
-Elliott J.M., 1982. The effects of temperature and ration size on the growth and energetics
of salmonids in captivity. Comparative Biochemistry and Physiology B 73, 81–91.
-Etheridge R., 1993. Lizards of the Liolaemus darwinii complex (Squamata:Iguania:
Tropiduridae) in northern Argentina. Museo regionale di Scienze naturali, Torino, 11: 137-
199.
-García Fernández J.J., Ojeda R.A., Fraga R.M., Díaz G.B. y Baigún R.J., 1997. Libro
Rojo: Mamíferos y Aves amenazados de la Argentina, Pág. 221.
-Garland Jr. T., Hankins E. y Huey R.B., 1990. Locomotor capacity and social dominance
in male lizards. Functional Ecology 4, 243–250.
-Goode M.J. y Duvall D., 1989. Body temperature and defensive behavior of free-ranging
prairie rattlesnakes, Crotalus viridisviridis. Animal Behaviour 38, 360–362.
-Greenwald O.E., 1974. Thermal dependence of striking and prey capture by gopher
snakes. Copeia 1974, 141–148.

43
-Greenwald O.E. y Kanter M.E., 1979. Effects of temperature and behavioral
thermoregulation on digestive efficiency and rate in corn snakes (Elaphe-Guttata-Guttata).
Physiological Zoology 52, 398–408.
-Haramura T., 2007. Microhabitat selection by tadpoles of Buergeria japonica inhabiting
the coastal area. J. Ethol, 25:3–7.
-Hertz P.E., Huey R.B. y Nevo E., 1982. Fight versus flight: body temperature influences
defensive responses of lizards. Animal Behaviour 30, 676–679.
-Hertz P.E., Huey R.B. y Stevenson R.D., 1993. Evaluating temperature regulation by field-
active ectotherms: the fallacy of the inappropriate question. Amer. Nat. 142, 796–818.
-Hill R.W., Wyse G.A. y Anderson M., 2004. Libro: Fisiología Animal. Ed. Médica
Panamericana S.A. Cap. 10, Pág.: 301-326
-Hochochka P.W. y Somero G.N., 2002. Biochemical Adaptation. Oxford, Oxford
University Press.
-Huey R.B., 1982. Temperature, physiology and the ecology of reptiles. In: Gans C., Pough
F.H. (Eds.), Biology of the reptilian. Academic Press, New York, Pág.: 25-91.
-Huey R.B., Peterson C.R., Arnold S.J. y Porter W.P., 1989. Hot rocks and no-so-hot rocks:
retreat-site selection by garter snakes and its thermal consequences. Ecology 70, 931-944.
-Huey R.B. y Slatkin M., 1976. Cost and benefits of lizards thermoregulation. Q. Rev. Biol.
51, 363–384.

44
-Huey R.B. y Stevenson R.D., 1979. Integrating Thermal Physiology and Ecology of
Ectotherms: A discusión of Approaches. Amer. Zool. 19: 357-366.
-Hutchinson G.E., 1957. Concluding remarks. Cold spring harbor symposi on quantitative
biology, 22, 415-427.
-IUPS Thermal Commission, 2001. 3rd
Ed. Glossary of terms for thermal physiology, The
Commission for Thermal Physiology of the International Union of Physiological Sciences.
The Japanese Journal of Physiology Vol. 51, No. 2
-Jonassen T.M., Imsland A.K. y Stefansson S.O., 1999. The interaction of temperature and
fish size on growth of juvenile halibut. Journal of Fish Biology 54, 556–572.
-Keith A.C., Tracy C.R. y Porter W.P., 1984. Diet, Digestion and Food Preferences of
Galapagos Land Iguanas. Herpetológica, Vol. 40, No. 2, pp. 205-212.
-Kooijman, S.A., 2010. Dynamc energy Budget theory for metabolic organisms. 3rd
Ed.
Amsterdam. Pp.508.
-Licht P., 1964. The temperature dependence of myosinadenosinetriphosphate and alkaline
phosphatase in lizards. Comp. Biochem. Physiol.12, 331–340
-Lillywhite B.H., Licht P. y Chelgren P., 1973. The Role of Behavioral Thermoregulation
in the Growth energetics of the toad, Bufo boreas. Ecology, 54 (2): 375–383.
-Lustick S.I., 1983. Cost–benefit of thermoregulation in birds: influences of posture,
microhabitat selection, and color, Pages 265–294 inW. P. Aspey, and S. I. Lustick, eds.
Behavioral Energetics. Columbus, Ohio State University Press.

45
-Mackay R.S., 1968. Observations on peristaltic activity versus temperature and circadian
rhythms in undisturbedVaranus flavescens andCtenosaura pectinata. Copeia, 252–259.
-Marvin G., 2003. Acuatic and Terrestrial Locomotor performance in a Seamiaquatic
Plethodontid Salamander (Pseudotriton ruber): Influence of acuatic temperature, Thermal
acclimatation and body size. Copeia (4): 704-713.
-Medina M., Gutierrez J., Scolaro A. e Ibargüengoytia N., 2009. Thermal responses to
environmental constraints in two populations of the oviparous lizard Liolaemus bibrioni in
Patagonia, Argentina. Journal of Thermal Biology 34, 32-40.
-Müller-Esterl W., 2008. Libro: Bioquímica: Fundamentos para Medicina y Ciencias de la
Vida. Versión española por Josep Joan Centelles Serra, Barcelona. Ed. Reverté. Pag. 165-
166.
-Passek K.M. y Gillingham J.C., 1997. Thermal influence on defensive behaviours of the
eastern garter snake, Thamnophis sirtalis. Animal Behaviour 54, 629–633.
-Prinzinger R., Pressmar A. y Schleucher E., 1991. Body temperature in birds. Comparative
Biochemistry and Physiology A 99:499–506.
-Rand A.S., 1964. Inverse relationship between temperature and shyness in the lizard
Anolis lineatopus. Ecology 45, 863–864.
-Randall D., Burggren W. y French K., 2001. Eckert Animal Physiology: Mecanisms and
Adaptations, fifth ed. W. H. Freeman and Co., New York.
-Rao S.K., Mathrubutham M., Karteron A., Sorensen K. y Cohen J.R., 1997. A versatile
microassay for elastase using succinylated elastin. Anal Biochem 150:222-7.

46
-Riddle O., 1909. The rate of digestion in cold blooded vertebrates. The influence of season
and temperature. Amer. J. Physiol.24, 447–458.
-Robson M.A. y Miles D.B., 2000. Locomotor performance and dominance in male tree
lizards, Urosaurus ornatus. Functional Ecology 14,338–344.
-Rome C., Stevens E. y Jhon-Alder H., 1992. The influence of temperature and thermal
acclimation on physiological function. Feder y Burggren (Eds.): Environmental Physiology
of the amphibians. The University of Chicago Press. United States.
-Roig-Juñent S., Flores G., Claver S. y Marvaldi A., 2001. “Monte desert (Argentina):
insect biodiversity and natural areas”, Journal of Arid
-Schuler M.S., Sears M.W. y Angilletta M.J.., 2011. Food consumption does not affect the
preferred body temperature of Yarrow’s spiny lizard (Sceloporus jarrovi). Journal of
Thermal Biology 36, 112-115.
-Schultz T.D., 1998. The utilization of patchy thermal microhabitats by the ectothermic
insect predator, Cicindelasex guttata. Ecological Entomology 23, 444–450.
-Shine R., Olsson M.M., Lemaster M.P., Moore I.T. y Mason R.T., 2000. Effects of sex,
body size, temperature, and location on the antipredator tactics of free-ranging gartersnakes
(Thamnophis sirtalis, Colubridae). Behavioral Ecology 11, 239–245.
-Sievert L., 1989. Postprandial temperature selection in Crotaphytus collaris. Copeia 1989,
983–989.
-Stevens C.E. y Hume I.D., 2004. Libro: Comparative physiology of the Vertebrate
Digestive System. Cambridge University Press. Cap. 2. Pág.: 11-23.

47
-Tian M., Huitema E., da Cunha L., Torto-Alalibo T. y Kamoun S., 2004. A Kazal-like
extracellular serine protease inhibitor from Phytophthora infestans targets the tomato
pathogenesis-related protease P69B. J Biol Chem 297(25):26370-7.
-Twining S.S., 1994. Fluorescein Isothiocuanate-Labeled Casein Assay for Proteolytic
Enzymes. Anal. Biochem., 143, 30-34.
-Van Damme R., Bauwens D. y Verheyen R.F., 1991. The thermal dependence of feeding
behavior, food consumption and gut passage time in the lizard Lacerta vivipara Jacquin.
FunctionalEcology 5,507–517.
-Van Marken Lichtenbelt W.D., 1992. Digestion in an ectothermic herbivore, the green
iguana (Iguana iguana): effect of food composition and body temperature. Physiological
Zoology 65, 649–673.
-Waldschmidt S.R., Jones S.M. y Porter W.P., 1986. The effect of body temperature and
feeding regime on activity, passage time, and digestive coefficient in the lizard Uta
stansburiana. Physiological Zoology 59, 376–383.
-Wang T., Zaar M., Arvedser S., Vedel-Smith C. y Overgaard J., 2003.Effects of
tempetature on the metabolic response to feeding in Python molurus. Comp. Biocherm.
Physiol. A 133, 519-527.
-Witters L.R. y Sievert L., 2001. Feeding causes thermophily in the woodhouse´s toad
(Bufo woodhousii). Journal of Thermal Biology 26, 205-208.
-Withers P.C. y Campbell J.D., 1985. Effects of environmental cost on thermoregulation in
the desert iguana. Physiological Zoology 58, 329–339.

48
-Yarto Jaramillo E., 2011. Alojamiento y problemas relacionados en reptiles: quemaduras,
problemas digestivos y respiratorios. 4to. Congreso LAVECCS-Santiago, Chile.