epiphytism, anatomy and regressive evolution in trichomanoid filmy ferns (hymenophyllaceae)
TRANSCRIPT

Epiphytism, anatomy and regressive evolution intrichomanoid filmy ferns (Hymenophyllaceae)
JEAN-YVES DUBUISSON1*, SOPHIE BARY1,2, ATSUSHI EBIHARA3,EUGÉNIE CARNERO-DIAZ4, ELODIE BOUCHERON-DUBUISSON4 andSABINE HENNEQUIN1
1Université Pierre et Marie Curie, UMR 7207 CNRS-MNHN-UPMC, Centre de Recherche sur laPaléobidoversité et les Paléoenvironnements, MNHN, Bâtiment de Géologie, CP48, 57 rue Cuvier,F-75005 Paris, France2Département Systématique et Evolution, MNHN, 57 rue Cuvier, F-75005 Paris, France3Department of Botany, National Museum of Nature and Science, 4-1-1 Amakubo, Tsukuba 305-0005,Japan4Université Pierre et Marie Curie, UR5 Physiologie Cellulaire et Moléculaire des Plantes, Casier 156,9 Quai St Bernard, F-75005 Paris, France
Received 17 December 2012; revised 28 June 2013; accepted for publication 13 August 2013
The few studies on the evolution of epiphytism in ferns have mostly focused on xerophytic and humus-collectingstrategies, neglecting hygrophytes that are abundant in rainforests, such as the trichomanoids (Hymenophyl-laceae). Using a phylogenetic approach, we studied the acquisition of epiphytism in this lineage, with the aim ofidentifying ecological anatomical adaptations and verifying the regressive epiphytic ‘bryophyte-like’ strategypreviously suggested for the group. Inferred evolution of anatomy and morphology, regression and ecology (moreparticularly colonial epiphytism) were analysed and compared using a maximum likelihood approach. Regressiveevolution of anatomy and morphology is revealed in the three clades of colonial epiphytes, probably linked to theselection of water acquisition by blades rather than by regressed roots. However, the ‘bryophyte-like’ strategy isrestricted to some taxa (especially Didymoglossum). Furthermore, a relationship is revealed between largemetaxylem and climbing habit. Diversification of colonial epiphytes (and some individual epiphytes) and hemi-epiphytism would have occurred in the upper Cretaceous and Tertiary, in accordance with the timing ofdiversification of modern ferns and the evolution of epiphytism in other fern families in the first angiosperm-dominated forests. This was here performed by selecting hygrophilous strategies that are unique in vascularplants. © 2013 The Linnean Society of London, Botanical Journal of the Linnean Society, 2013, 173, 573–593.
ADDITIONAL KEYWORDS: bryophytes – comparative methods − ecology − phylogeny – Trichomanes.
INTRODUCTION
Epiphytes, including ferns, lycophytes and mosses,are one of the main components of rainforest diversityat the pantropical level, especially in montane cloudforests, in which epiphyte diversity is highest (Gentry& Dodson, 1987). However, studies on epiphytism andrelated strategies in ferns are relatively scarce incomparison with angiosperms (Benzing, 1990;
Dubuisson, Hennequin & Schneider, 2009). With 29%of the total number of fern species occupying thishabitat, ferns are nevertheless the second group ofvascular plants in terms of epiphyte diversity, aftermonocots with 31% epiphytic species, mostly Orchi-daceae. In addition, most ferns and lycophytes aremore diverse than orchids in many palaeotropicalrainforests, forming 36–72% of the total epiphyticdiversity (e.g. in Australia, New Zealand and theMicronesian islands; Oliver, 1930; Hosokawa, 1943;Wallace, 1981). Obligate epiphytic ferns are subjected*Corresponding author. E-mail: [email protected]
bs_bs_banner
Botanical Journal of the Linnean Society, 2013, 173, 573–593. With 7 figures
© 2013 The Linnean Society of London, Botanical Journal of the Linnean Society, 2013, 173, 573–593 573

to the same environmental constraints as seed plantsand should also be naturally selectively adapted tothis habitat. The epiphytic habitat is quite constrain-ing; the lack of relation to the soil implies a high riskof desiccation and the obligatory capacity to use rain-water or air moisture quickly and directly and/or tolimit or compensate the water loss from evaporation.Like the majority of epiphytic seed plants, strict epi-phytic ferns, especially those belonging to Polypodi-ales (sensu Smith et al., 2006; Christenhusz, Zhang &Schneider, 2011), generally display xeromorphic adap-tive traits, enabling them to avoid and/or enduredrought (Schneider et al., 2004a; see reviews byDubuisson et al., 2009; Hietz, 2010). Other fernsdisplay growth habits allowing them to accumulatehumus and to entrap nutrients and water. These aremesophile humus collectors, illustrated by the well-known bird’s nest fern (Asplenium nidus L., Aspleni-aceae), the drynarioid ferns [species of Drynaria(Bory) J.Sm., Polypodiaceae; Janssen & Schneider,2005] and the staghorn ferns (species of PlatyceriumDesv., Polypodiaceae; Kreier & Schneider, 2006). Thehumus-collecting strategy is sometimes combinedwith a mutualistic interaction with ants, as in Micro-gramma bifrons (Hook.) Lellinger (Polypodiaceae) andrelatives (e.g. Lecanopteris Reinw. genus, Polypodi-aceae; Haufler et al., 2003). The final type of epiphyticfern, hygrophilous epiphytes or hygrophytes, lacksboth xerophytic traits and humus-collector strategies(Dubuisson et al., 2009). Consequently, these epi-phytes are restricted to humid environments.
As recently reviewed by Dubuisson et al. (2009,2011), most members of Hymenophyllaceae, or filmyferns, are obligatory hygrophytes. They constitute apertinent group in which to investigate adaptivestrategies related to hygrophilous epiphytism inferns. Despite the strong limitations or constraintsrelated to the epiphytic habit, members of Hymeno-phyllaceae have succeeded in diversifying undersuch constraints, with > 60% epiphytic species dis-tributed worldwide on most continents and numer-ous oceanic islands. Several species of filmy ferns aredesiccation tolerant and have the capacity to survivea period of dehydration and revive after rehydration,similar to some species of epiphytic Polypodiaceae.However, the dehydration is limited to a short period(a few hours) (Hietz & Briones, 1998, 2001; Nitta,2006) and is supposedly more problematic in an epi-phytic context because plants do not have the pos-sibility to absorb water directly from the soil tocompensate for dehydration. A regressive evolutionin morphology has been suggested previously for epi-phytic trichomanoids, one of the two major clades inthe family and the most morphologically and ecologi-cally diversified (Schneider, 2000; Dubuisson et al.,2003). Trichomanoids here refer to the distinct clade
that traditionally corresponds to the genus Tricho-manes L. s.l. and currently includes eight genera(Abrodictyum C.Presl, Callistopteris Copel., Cephalo-manes C.Presl, Crepidomanes C.Presl, Didymoglos-sum Desv., Polyphlebium Copel., Trichomanes andVandenboschia Copel.; Ebihara et al., 2006). Theregressive evolution in trichomanoids would illus-trate tendencies towards a ‘bryophyte-like’ strategy,allowing filmy ferns to reduce their requirements ina constraining habitat. Recent phylogenetic and com-parative analyses have revealed that regressive evo-lution in trichomanoids also concerns anatomy(Dubuisson et al., 2011). The anatomical regression,illustrated by the acquisition of subcollateral andcollateral types and related to reductions in stemand stele size, was observed in three groups embed-ded in a clade clustering hemi-epiphytic and epi-phytic taxa, each involving only colonial epiphytes(i.e. individuals having the ability to colonize a broadsurface of substrate). Furthermore, Dubuisson et al.(2011) have shown that not all epiphytes displaymorphological and anatomical regression, especiallywhen non-colonial (i.e. individuals that do not havethe ability to colonize a broad surface of substrate).Further phylogenetic comparative analyses are thusneeded to test the relationships between habit (herehygrophilous epiphytism) and morphological andanatomical regression.
The present study is analogous to and derived froma previous study (Dubuisson et al., 2011). The aims ofthis study are to infer the evolution of the ecology andrelated morphological and anatomical changes intrichomanoids, relying on an expanded molecularphylogenetic analysis representing morphological,ecological and geographical variability in the lineage,and on adequate statistical tests. By completing anexisting dataset with additional anatomical and eco-logical data, we statistically test potential relation-ships among selected characters and betweencharacters and ecology, with the aim of proposinghypotheses on adaptive strategies that have enabledfilmy ferns to colonize the epiphytic habitat of rain-forests and to verify the ‘bryophyte-like’ strategy sug-gested previously.
MATERIAL AND METHODSTAXONOMIC SAMPLING
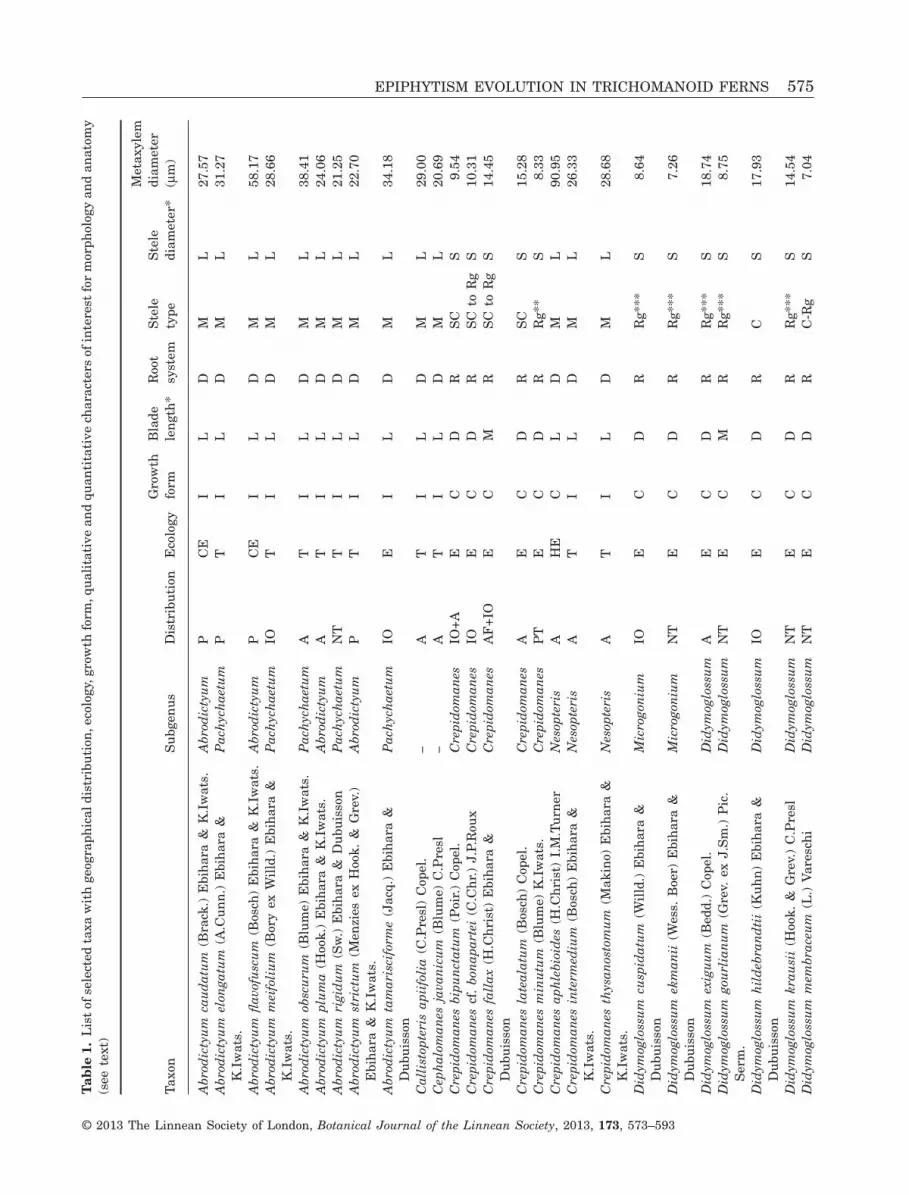
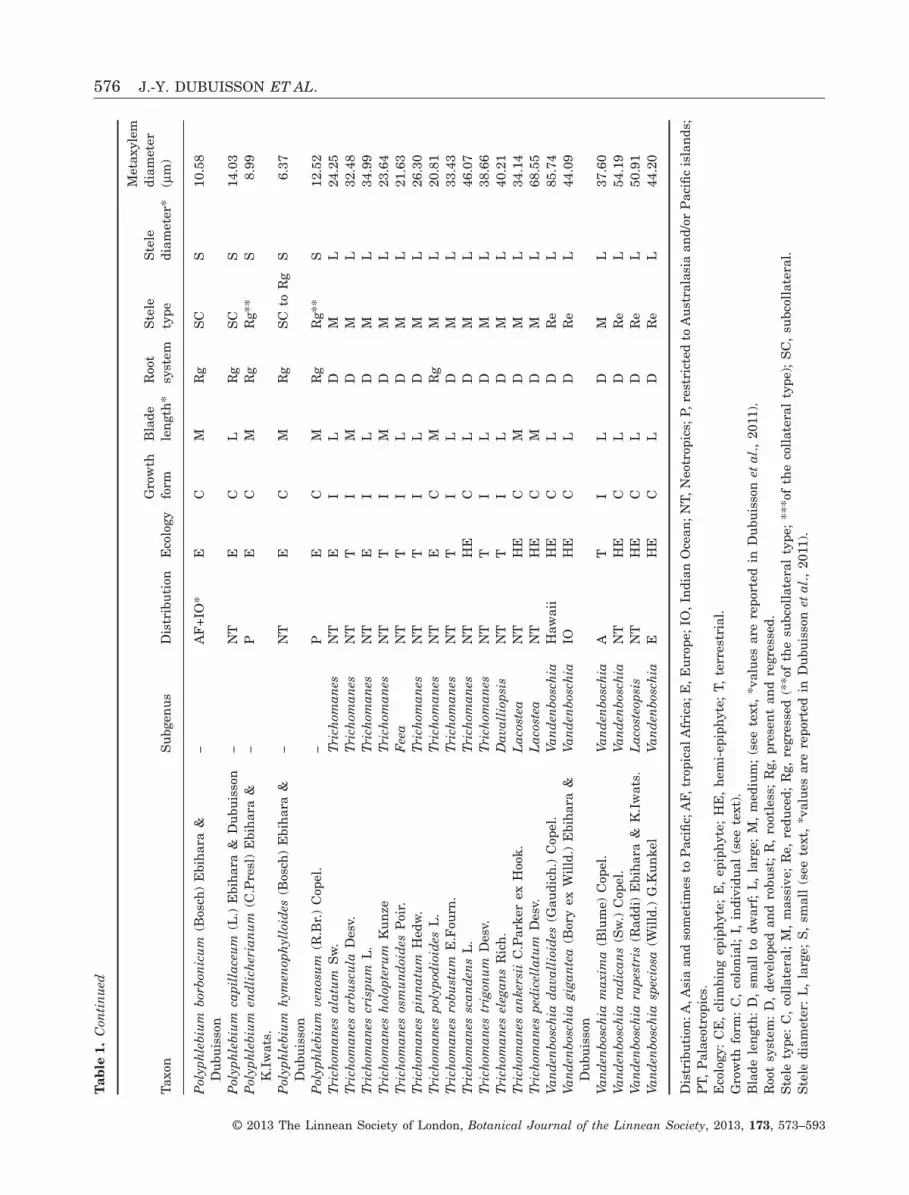
Here, we follow the strategy selected in the previousstudy (Dubuisson et al., 2011) with 50 trichomanoidspecies representative of the morphological, anatomi-cal and ecological variability of the lineage and of itsglobal geographical distribution. The selected speciesare listed in Table 1 with details of their ecology,growth forms and geographical distribution.
574 J.-Y. DUBUISSON ET AL.
© 2013 The Linnean Society of London, Botanical Journal of the Linnean Society, 2013, 173, 573–593
Tab
le1.
Lis
tof
sele
cted
taxa
wit
hge
ogra
phic
aldi
stri
buti
on,e
colo
gy,g
row
thfo
rm,q
ual
itat
ive
and
quan
tita
tive
char
acte
rsof
inte
rest
for
mor
phol
ogy
and
anat
omy
(see
text
)
Taxo
nS
ubg
enu
sD
istr
ibu
tion
Eco
logy
Gro
wth
form
Bla
dele
ngt
h*
Roo
tsy
stem
Ste
lety
peS
tele
diam
eter
*
Met
axyl
emdi
amet
er(μ
m)
Abr
odic
tyu
mca
ud
atu
m(B
rack
.)E
bih
ara
&K
.Iw
ats.
Abr
odic
tyu
mP
CE
IL
DM
L27
.57
Abr
odic
tyu
mel
onga
tum
(A.C
un
n.)
Ebi
har
a&
K.I
wat
s.P
ach
ych
aetu
mP
TI
LD
ML
31.2
7
Abr
odic
tyu
mfl
avof
usc
um
(Bos
ch)
Ebi
har
a&
K.I
wat
s.A
brod
icty
um
PC
EI
LD
ML
58.1
7A
brod
icty
um
mei
foli
um
(Bor
yex
Wil
ld.)
Ebi
har
a&
K.I
wat
s.P
ach
ych
aetu
mIO
TI
LD
ML
28.6
6
Abr
odic
tyu
mob
scu
rum
(Blu
me)
Ebi
har
a&
K.I
wat
s.P
ach
ych
aetu
mA
TI
LD
ML
38.4
1A
brod
icty
um
plu
ma
(Hoo
k.)
Ebi
har
a&
K.I
wat
s.A
brod
icty
um
AT
IL
DM
L24
.06
Abr
odic
tyu
mri
gid
um
(Sw
.)E
bih
ara
&D
ubu
isso
nP
ach
ych
aetu
mN
TT
IL
DM
L21
.25
Abr
odic
tyu
mst
rict
um
(Men
zies
exH
ook.
&G
rev.
)E
bih
ara
&K
.Iw
ats.
Abr
odic
tyu
mP
TI
LD
ML
22.7
0
Abr
odic
tyu
mta
mar
isci
form
e(J
acq.
)E
bih
ara
&D
ubu
isso
nP
ach
ych
aetu
mIO
EI
LD
ML
34.1
8
Cal
list
opte
ris
apii
foli
a(C
.Pre
sl)
Cop
el.
–A
TI
LD
ML
29.0
0C
eph
alom
anes
java
nic
um
(Blu
me)
C.P
resl
–A
TI
LD
ML
20.6
9C
repi
dom
anes
bipu
nct
atu
m(P
oir.
)C
opel
.C
repi
dom
anes
IO+A
EC
DR
SC
S9.
54C
repi
dom
anes
cf.
bon
apar
tei
(C.C
hr.
)J.
P.R
oux
Cre
pid
oman
esIO
EC
DR
SC
toR
gS
10.3
1C
repi
dom
anes
fall
ax(H
.Ch
rist
)E
bih
ara
&D
ubu
isso
nC
repi
dom
anes
AF
+IO
EC
MR
SC
toR
gS
14.4
5
Cre
pid
oman
esla
teal
atu
m(B
osch
)C
opel
.C
repi
dom
anes
AE
CD
RS
CS
15.2
8C
repi
dom
anes
min
utu
m(B
lum
e)K
.Iw
ats.
Cre
pid
oman
esP
TE
CD
RR
g**
S8.
33C
repi
dom
anes
aph
lebi
oid
es(H
.Ch
rist
)I.
M.T
urn
erN
esop
teri
sA
HE
CL
DM
L90
.95
Cre
pid
oman
esin
term
ediu
m(B
osch
)E
bih
ara
&K
.Iw
ats.
Nes
opte
ris
AT
IL
DM
L26
.33
Cre
pid
oman
esth
ysan
osto
mu
m(M
akin
o)E
bih
ara
&K
.Iw
ats.
Nes
opte
ris
AT
IL
DM
L28
.68
Did
ymog
loss
um
cusp
idat
um
(Wil
ld.)
Ebi
har
a&
Du
buis
son
Mic
rogo
niu
mIO
EC
DR
Rg*
**S
8.64
Did
ymog
loss
um
ekm
anii
(Wes
s.B
oer)
Ebi
har
a&
Du
buis
son
Mic
rogo
niu
mN
TE
CD
RR
g***
S7.
26
Did
ymog
loss
um
exig
uu
m(B
edd.
)C
opel
.D
idym
oglo
ssu
mA
EC
DR
Rg*
**S
18.7
4D
idym
oglo
ssu
mgo
url
ian
um
(Gre
v.ex
J.S
m.)
Pic
.S
erm
.D
idym
oglo
ssu
mN
TE
CM
RR
g***
S8.
75
Did
ymog
loss
um
hil
deb
ran
dti
i(K
uh
n)
Ebi
har
a&
Du
buis
son
Did
ymog
loss
um
IOE
CD
RC
S17
.93
Did
ymog
loss
um
krau
sii
(Hoo
k.&
Gre
v.)
C.P
resl
Did
ymog
loss
um
NT
EC
DR
Rg*
**S
14.5
4D
idym
oglo
ssu
mm
embr
aceu
m(L
.)V
ares
chi
Did
ymog
loss
um
NT
EC
DR
C-R
gS
7.04
EPIPHYTISM EVOLUTION IN TRICHOMANOID FERNS 575
© 2013 The Linnean Society of London, Botanical Journal of the Linnean Society, 2013, 173, 573–593
Tab
le1.
Con
tin
ued
Taxo
nS
ubg
enu
sD
istr
ibu
tion
Eco
logy
Gro
wth
form
Bla
dele
ngt
h*
Roo
tsy
stem
Ste
lety
peS
tele
diam
eter
*
Met
axyl
emdi
amet
er(μ
m)
Pol
yph
lebi
um
borb
onic
um
(Bos
ch)
Ebi
har
a&
Du
buis
son
–A
F+I
O*
EC
MR
gS
CS
10.5
8
Pol
yph
lebi
um
capi
llac
eum
(L.)
Ebi
har
a&
Du
buis
son
–N
TE
CL
Rg
SC
S14
.03
Pol
yph
lebi
um
end
lich
eria
nu
m(C
.Pre
sl)
Ebi
har
a&
K.I
wat
s.–
PE
CM
Rg
Rg*
*S
8.99
Pol
yph
lebi
um
hym
enop
hyl
loid
es(B
osch
)E
bih
ara
&D
ubu
isso
n–
NT
EC
MR
gS
Cto
Rg
S6.
37
Pol
yph
lebi
um
ven
osu
m(R
.Br.
)C
opel
.–
PE
CM
Rg
Rg*
*S
12.5
2Tr
ich
oman
esal
atu
mS
w.
Tric
hom
anes
NT
EI
LD
ML
24.2
5Tr
ich
oman
esar
busc
ula
Des
v.Tr
ich
oman
esN
TT
IM
DM
L32
.48
Tric
hom
anes
cris
pum
L.
Tric
hom
anes
NT
EI
LD
ML
34.9
9Tr
ich
oman
esh
olop
teru
mK
un
zeTr
ich
oman
esN
TT
IM
DM
L23
.64
Tric
hom
anes
osm
un
doi
des
Poi
r.F
eea
NT
TI
LD
ML
21.6
3Tr
ich
oman
espi
nn
atu
mH
edw
.Tr
ich
oman
esN
TT
IL
DM
L26
.30
Tric
hom
anes
poly
pod
ioid
esL
.Tr
ich
oman
esN
TE
CM
Rg
ML
20.8
1Tr
ich
oman
esro
bust
um
E.F
ourn
.Tr
ich
oman
esN
TT
IL
DM
L33
.43
Tric
hom
anes
scan
den
sL
.Tr
ich
oman
esN
TH
EC
LD
ML
46.0
7Tr
ich
oman
estr
igon
um
Des
v.Tr
ich
oman
esN
TT
IL
DM
L38
.66
Tric
hom
anes
eleg
ans
Ric
h.
Dav
alli
opsi
sN
TT
IL
DM
L40
.21
Tric
hom
anes
anke
rsii
C.P
arke
rex
Hoo
k.L
acos
tea
NT
HE
CM
DM
L34
.14
Tric
hom
anes
ped
icel
latu
mD
esv.
Lac
oste
aN
TH
EC
MD
ML
68.5
5Va
nd
enbo
sch
iad
aval
lioi
des
(Gau
dich
.)C
opel
.Va
nd
enbo
sch
iaH
awai
iH
EC
LD
Re
L85
.74
Van
den
bosc
hia
giga
nte
a(B
ory
exW
illd
.)E
bih
ara
&D
ubu
isso
nVa
nd
enbo
sch
iaIO
HE
CL
DR
eL
44.0
9
Van
den
bosc
hia
max
ima
(Blu
me)
Cop
el.
Van
den
bosc
hia
AT
IL
DM
L37
.60
Van
den
bosc
hia
rad
ican
s(S
w.)
Cop
el.
Van
den
bosc
hia
NT
HE
CL
DR
eL
54.1
9Va
nd
enbo
sch
iaru
pest
ris
(Rad
di)
Ebi
har
a&
K.I
wat
s.L
acos
teop
sis
NT
HE
CL
DR
eL
50.9
1Va
nd
enbo
sch
iasp
ecio
sa(W
illd
.)G
.Ku
nke
lVa
nd
enbo
sch
iaE
HE
CL
DR
eL
44.2
0
Dis
trib
uti
on:A
,Asi
aan
dso
met
imes
toP
acifi
c;A
F,tr
opic
alA
fric
a;E
,Eu
rope
;IO
,In
dian
Oce
an;N
T,N
eotr
opic
s;P,
rest
rict
edto
Au
stra
lasi
aan
d/or
Pac
ific
isla
nds
;P
T,P
alae
otro
pics
.E
colo
gy:
CE
,cl
imbi
ng
epip
hyt
e;E
,ep
iph
yte;
HE
,h
emi-
epip
hyt
e;T,
terr
estr
ial.
Gro
wth
form
:C
,co
lon
ial;
I,in
divi
dual
(see
text
).B
lade
len
gth
:D
,sm
all
todw
arf;
L,
larg
e;M
,m
ediu
m;
(see
text
,*v
alu
esar
ere
port
edin
Du
buis
son
etal
.,20
11).
Roo
tsy
stem
:D
,de
velo
ped
and
robu
st;
R,
root
less
;R
g,pr
esen
tan
dre
gres
sed.
Ste
lety
pe:
C,
coll
ater
al;
M,
mas
sive
;R
e,re
duce
d;R
g,re
gres
sed
(**o
fth
esu
bcol
late
ral
type
;**
*of
the
coll
ater
alty
pe);
SC
,su
bcol
late
ral.
Ste
ledi
amet
er:
L,
larg
e;S
,sm
all
(see
text
,*v
alu
esar
ere
port
edin
Du
buis
son
etal
.,20
11).
576 J.-Y. DUBUISSON ET AL.
© 2013 The Linnean Society of London, Botanical Journal of the Linnean Society, 2013, 173, 573–593

ANATOMICAL DATA AND NEW QUANTITATIVE
CHARACTERS
The anatomical diversity was fully investigated byDubuisson et al. (2011). In that paper, four stele types(massive, reduced, subcollateral, collateral) and twocortex types (homogeneous and heterogeneous) aredefined and described. Quantitative measures wereperformed on stem thickness and stele diameter, andallowed us to define two classes for stele diameterwith a limit at 150 μm. Many trichomanoids areclimbing taxa (in a hemi-epiphytic context, see nextsection) and Carlquist (1991) showed that, in angio-sperm lianas, vessels are significantly larger thanthose observed in non-climbing relatives, enablingfaster water conduction in long, climbing stems. Inthe present study, we added data on metaxylem diam-eter in order to test its potential relationship with aclimbing habit. Following the recommendationsdetailed in the previous study, for each speciesstudied, we performed measures on at least threespecimens collected from different localities and onten sections per specimen, collected at different posi-tions on the stem, whenever possible, but especiallyfor colonial long-creeping taxa. In the case of rare,short monocaulous species, measures were made onten sections from the single rhizome of one specimen.Then, at least ten measures (of the largest diameterfor not fully cylindrical cells) were performed on theten largest metaxylem cells, if available. For smallsteles with few tracheids and/or with less than tenmetaxylem cells, measures were made on all availablemetaxylem cells. Although the absence of xylem hasbeen reported for a few species of Hymenophyllaceae(Ogura, 1972), all specimens selected and studiedhere had at least one tracheid. In the case of a singletracheid, we measured this single cell. As explained inthe previous analysis, and as a result of heterogeneityin the data acquisition procedure, intraspecific vari-ability was not taken into account. We thus assumethat the data (means) used here are representative ofthe taxon and are useful for interspecific comparativestudies, but do not reflect their entire naturalvariability.
Correlations among selected anatomical characterswere graphically evaluated using a standard regres-sion procedure to evidence potential clustering. Thisapproach enabled us to categorize quantitative char-acters and to formulate hypothetical correlations,which were then phylogenetically tested.
PHYLOGENETIC APPROACHES AND INFERRED
EVOLUTION OF SELECTED HABITS AND CHARACTERS
The choice of the phylogenetic framework and therooting strategy is explained in detail in our previous
related study (Dubuisson et al., 2011). Followingreviews on ecology in trichomanoids (Dubuisson et al.,2003, 2009), we defined here four main ecologicaltypes: terrestrials; climbing (hemi)-epiphytes; indi-vidual epiphytes; and colonial epiphytes. Terrestrialscorrespond to taxa that complete their entire life cyclerooted on soil or, possibly, in trichomanoids, on crev-ices of rocks in which humus accumulates. In terres-trial taxa, epiphytism is sometimes observed, but isconsidered accidental and not obligatory. According toBenzing (1990), hemi-epiphytes correspond to taxathat occur in both habitats, firstly terrestrial andsecondarily epiphytic for secondary hemi-epiphytism,and conversely for primary hemi-epiphytism. Sometrichomanoids are secondary hemi-epiphytes sensuBenzing, beginning their growth terrestrially, thenclimbing on trunks and secondarily becoming epi-phytic. For example, Vandenboschia gigantea (Bory exWilld.) Ebihara & Dubuisson colonizes the forestunderstorey before climbing on trunks (Dubuissonet al., 2003). Other hemi-epiphytes, such as Vanden-boschia collariata (Bosch) Ebihara & K.Iwats., begintheir growth at the bases of trees, developing roots inthe soil and climbing on trunks with long-scandent,rootless stems. They are considered as primary hemi-epiphytes, because the juvenile stage is epiphytic atthe bases of trees before roots reach the soil (Nitta &Epps, 2009). The attribution of a precise hemi-epiphytic category to every species requires furtherfield investigations, and the separation is not alwaysclear cut. In addition, Dubuisson et al. (2003) alsodistinguished lianescence for Trichomanes subgenusLacostea Bosch, with species having a rooted terres-trial part and climbing rootless stems. Because strat-egies in Lacostea are comparable with those observedin V. collariata and, in order to avoid confusion with‘lianas’ (a term usually restricted to woody vines), wedecided to group here under ‘hemi-epiphytic habit’ s.l.all trichomanoids that begin their growth terrestri-ally or at the bases of trees and complete their growthas epiphytes after climbing on trunks. Uncertaintyremains as to the precise type of hemi-epiphytism forseveral climbing taxa. Hemi-epiphytism thereforeconcerns all Vandenboschia [except Vandenboschiamaxima (Blume) Copel.] and all Trichomanes subge-nus Lacostea and some Trichomanes subgenus Tricho-manes (e.g. Trichomanes scandens L.), and alwaysimplies a climbing habit on a trunk for mature parts.A particular case is formed by Abrodictyum caudatum(Brack.) Ebihara & K.Iwats. and Abrodictyum flavo-fuscum (Bosch) Ebihara & K.Iwats. These taxa areholo-epiphytic, living their entire lives as epiphytes(see hereafter), but always display a robust long-creeping stem (with rare branching) that climbs ver-tically along tree trunks. We categorized such speciesas climbing epiphytes. Our term epiphytes corre-
EPIPHYTISM EVOLUTION IN TRICHOMANOID FERNS 577
© 2013 The Linnean Society of London, Botanical Journal of the Linnean Society, 2013, 173, 573–593

sponds here to holo-epiphytes and concerns taxa thatobligatorily complete their entire life cycle on otherplants (phorophytes), especially trees or tree ferns.This habit also involves the saxicolous habit, in whichconstraints are similar in terms of accessibility towater and nutrients, and because numerous species(especially belonging to Crepidomanes and Didymo-glossum) occur in both habitats. In previous studies(Dubuisson et al., 2003, 2009, 2011), two types ofgrowth form were distinguished. The individualgrowth form concerns taxa with erect to short-creeping rhizomes that rarely branch, not allowingindividuals to colonize a large area. In contrast, thecolonial growth form concerns taxa with branchedlong-creeping rhizomes, enabling individuals to colo-nize their substrate, sometimes over a relatively largearea (e.g. V. gigantea, see Dubuisson et al., 2003; andFig. 6 for colonial epiphytes). In addition to these twomain growth forms, a few species display long-creeping rhizomes that do not or only rarely branch,and individuals do not colonize a large area (e.g.A. caudatum, A. flavofuscum and Trichomanes robus-tum E.Fourn.). The growth form of these speciesis thus considered here as individual. Most terrestrialspecies display an individual growth form. Allhemi-epiphytes are colonial with branched long-creeping scandent rhizomes. If not (i.e. all remainingepiphytes), the growth on phorophytes has, a priori,no preferential direction. Holo-epiphytism (s.l.) canthus be divided into individual (non-climbing) epi-phytism, colonial (non-climbing) epiphytism andclimbing epiphytism. In our coding, climbing holo-epiphytes (s.l.) are treated together with hemi-epiphytes. Phylogenetic relationships between growthform and anatomy are discussed in Dubuisson et al.(2011).
We also used characters previously defined for theblade length and anatomy (Dubuisson et al., 2011) asfollows. The blade length represents the plant sizeand is divided into three classes: large plants (withleaves longer than 12 cm), small to dwarf plants (withleaves shorter than 6 cm) and medium-sized plants(with leaves of 6–12 cm). The stele type was coded bycombining collateral with subcollateral types andmassive with reduced types. We defined two classesfor the stele diameter with a limit of 150 μm. Asalready highlighted in our previous study, anatomicalregression of the stem also seems to be accompaniedby a regression in the adventitious root system in atleast four groups (Trichomanes polypodioides L.,Polyphlebium, Didymoglossum and Crepidomanessubgenus Crepidomanes), which are all colonial epi-phytes. We also investigated the root system by defin-ing two classes: developed root system and regressedroot system (to rootless). Coding for all characters isreported in Table 1.
As for the previous study, the evolution of selectedcharacters was inferred from the consensus phyloge-netic tree (including branch lengths) using themaximum likelihood (ML) method as implementedin the Mesquite package ver. 2.73 (Maddison &Maddison, 2010). We tested two models implementedin Mesquite. The first (MK1 for ‘Markov K-state one-parameter’) assumes equivalent probabilities forforward (apomorphic) and backward (reversal)changes for a single character. The second(AsymmMK for ‘Asymmetrical Markov K-state two-parameter’) attributes distinct probabilities for rever-sal and apomorphic changes. This difference inchange rates appears to be more realistic, especiallyin the case of regressive evolution, when reductionand loss of characters are often irreversible (the irre-versibility constraint is particular to the AsymmMKmodel when the backward rate is constrained to below to negligible). However, the AsymmMK model canonly be selected for binary characters. For bothmodels, rates of changes are directly estimated basedon our data.
In accordance with previous hypotheses (Dubuissonet al., 2011), we particularly propose here to testcorrelations between morphological and anatomicalregression and epiphytism and between metaxylemdiameter and climbing habit of (hemi)-epiphytes.Morphological and anatomical regression is repre-sented by small to dwarf size (leaves shorter than6 cm), regressed root system, small stele (< 150 μm)and subcollateral and collateral stele types. We testedcorrelations between characters on the phylogenetictree using the method of Pagel & Meade (2006) ondiscrete data implemented in their BayesTraitspackage (available from http://www.evolution.rdg.ac.uk). For each pair of characters tested, theprogram calculates the likelihood of the distributionof character states expected on the trees under theassumption that both characters are dependent (orevolutionarily correlated), and the likelihood of theexpected distribution under the assumption that bothcharacters are independent (or not evolutionarily cor-related). We used the ML method implemented in theBayesTraits package. The analyses were performedon a random sampling of 100 trees among the treesproduced by the Bayesian analysis used to recon-struct the phylogenetic tree. The resulting likelihoodswere compared using the likelihood ratio test statistic(LR) (Felsenstein, 1981), where LR = 2[log-likelihood(dependent model) – log-likelihood (independentmodel)] is distributed as a χ2 with four degrees offreedom. A significant χ2 value proposes that theevolutionary correlation between the two charactersis statistically significant. The LR used here corre-sponds to the mean of the LR provided for each of the100 sampled trees.
578 J.-Y. DUBUISSON ET AL.
© 2013 The Linnean Society of London, Botanical Journal of the Linnean Society, 2013, 173, 573–593
RESULTSMETAXYLEM DIAMETER
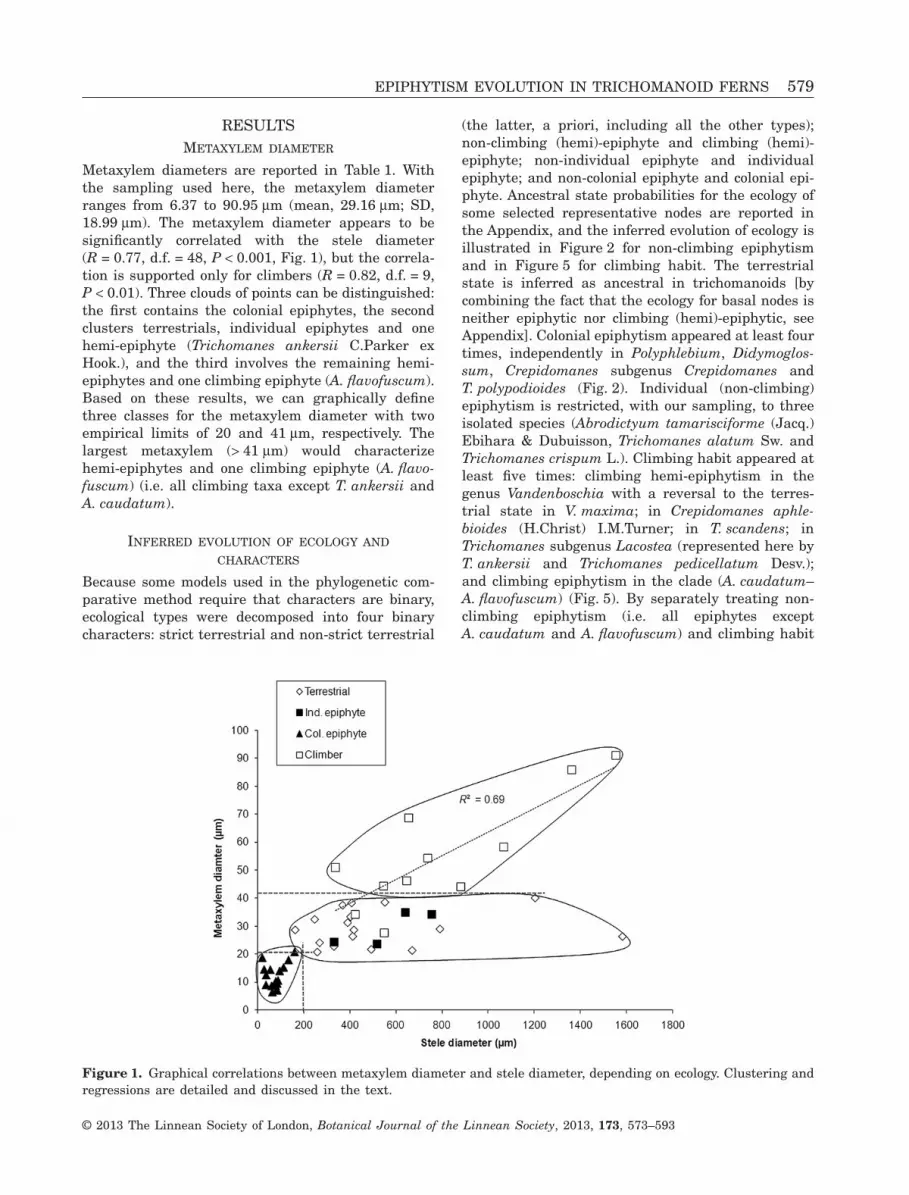
Metaxylem diameters are reported in Table 1. Withthe sampling used here, the metaxylem diameterranges from 6.37 to 90.95 μm (mean, 29.16 μm; SD,18.99 μm). The metaxylem diameter appears to besignificantly correlated with the stele diameter(R = 0.77, d.f. = 48, P < 0.001, Fig. 1), but the correla-tion is supported only for climbers (R = 0.82, d.f. = 9,P < 0.01). Three clouds of points can be distinguished:the first contains the colonial epiphytes, the secondclusters terrestrials, individual epiphytes and onehemi-epiphyte (Trichomanes ankersii C.Parker exHook.), and the third involves the remaining hemi-epiphytes and one climbing epiphyte (A. flavofuscum).Based on these results, we can graphically definethree classes for the metaxylem diameter with twoempirical limits of 20 and 41 μm, respectively. Thelargest metaxylem (> 41 μm) would characterizehemi-epiphytes and one climbing epiphyte (A. flavo-fuscum) (i.e. all climbing taxa except T. ankersii andA. caudatum).
INFERRED EVOLUTION OF ECOLOGY AND
CHARACTERS
Because some models used in the phylogenetic com-parative method require that characters are binary,ecological types were decomposed into four binarycharacters: strict terrestrial and non-strict terrestrial
(the latter, a priori, including all the other types);non-climbing (hemi)-epiphyte and climbing (hemi)-epiphyte; non-individual epiphyte and individualepiphyte; and non-colonial epiphyte and colonial epi-phyte. Ancestral state probabilities for the ecology ofsome selected representative nodes are reported inthe Appendix, and the inferred evolution of ecology isillustrated in Figure 2 for non-climbing epiphytismand in Figure 5 for climbing habit. The terrestrialstate is inferred as ancestral in trichomanoids [bycombining the fact that the ecology for basal nodes isneither epiphytic nor climbing (hemi)-epiphytic, seeAppendix]. Colonial epiphytism appeared at least fourtimes, independently in Polyphlebium, Didymoglos-sum, Crepidomanes subgenus Crepidomanes andT. polypodioides (Fig. 2). Individual (non-climbing)epiphytism is restricted, with our sampling, to threeisolated species (Abrodictyum tamarisciforme (Jacq.)Ebihara & Dubuisson, Trichomanes alatum Sw. andTrichomanes crispum L.). Climbing habit appeared atleast five times: climbing hemi-epiphytism in thegenus Vandenboschia with a reversal to the terres-trial state in V. maxima; in Crepidomanes aphle-bioides (H.Christ) I.M.Turner; in T. scandens; inTrichomanes subgenus Lacostea (represented here byT. ankersii and Trichomanes pedicellatum Desv.);and climbing epiphytism in the clade (A. caudatum–A. flavofuscum) (Fig. 5). By separately treating non-climbing epiphytism (i.e. all epiphytes exceptA. caudatum and A. flavofuscum) and climbing habit
Figure 1. Graphical correlations between metaxylem diameter and stele diameter, depending on ecology. Clustering andregressions are detailed and discussed in the text.
EPIPHYTISM EVOLUTION IN TRICHOMANOID FERNS 579
© 2013 The Linnean Society of London, Botanical Journal of the Linnean Society, 2013, 173, 573–593

as detailed above, we do not reveal any relationshipbetween these two habits, suggesting that climbinghemi-epiphytic or epiphytic taxa or clades would havebeen derived directly and independently from terres-trial ancestors. When we treat ecology as one multi-state character involving epiphytism and climbinghabit, colonial epiphytism is inferred to haveappeared in the common ancestor of the clade calledHE [as ‘(Hemi)-Epiphytic’ clade, because it involvesepiphytes and climbing (hemi)-epiphytes] with a prob-
ability of 0.71 (not detailed in Appendix, arrow inFig. 2). This result suggests that both hemi-epiphytism in Vandenboschia and in Cr. aphlebioideswould have been independently derived from colonialepiphytism with an additional reversal to terrestrialhabit in [Crepidomanes intermedium (Bosch) Ebihara& K. Iwats.–Crepidomanes thysanostomum (Makino)Ebihara & K.Iwats.] and V. maxima. Outside the HEclade, the inferred changes are identical to thoserevealed by the binary treatment.
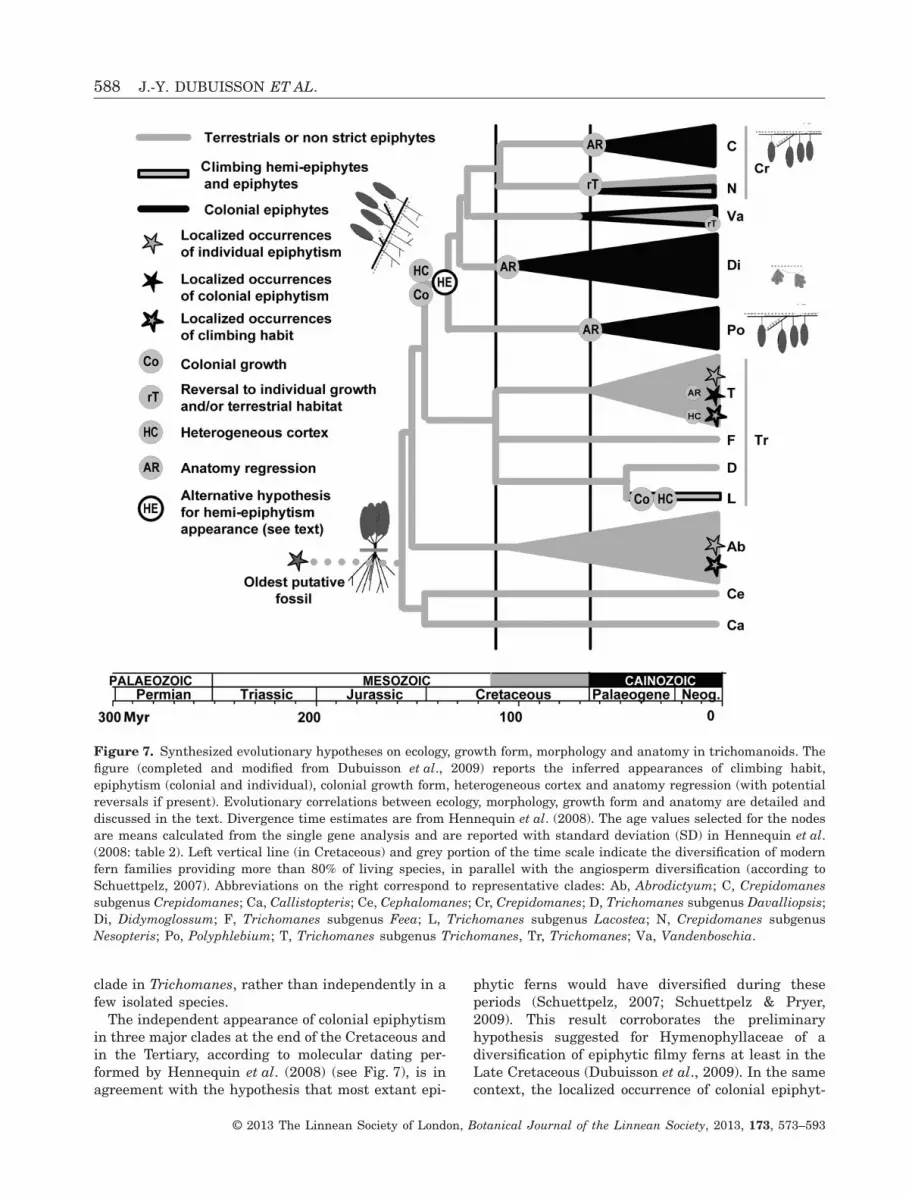
Figure 2. Evolution of holo-epiphytism in trichomanoids inferred using maximum likelihood. We excluded here climbingepiphytism that concerns only two taxa (Abrodictyum caudatum and A. flavofuscum) which are treated as climbers (seeFig. 5). The arrow indicates the alternative inferred appearance of colonial epiphytism when ecology is treated involvingall the states (see text). The partial coloration of some internal nodes is proportional to the change probabilities (seeAppendix). On nodes, abbreviations correspond to representative clades also reported in the Appendix: Ab, Abrodictyum;C, Crepidomanes subgenus Crepidomanes; Ca, Callistopteris; Ce, Cephalomanes; Cr, Crepidomanes; D, Trichomanessubgenus Davalliopsis; Di, Didymoglossum; HE, ‘(Hemi)-Epiphytic’ clade (see text); L, Trichomanes subgenus Lacostea;N, Crepidomanes subgenus Nesopteris; Po, Polyphlebium; Tr, Trichomanes; Va, Vandenboschia.
580 J.-Y. DUBUISSON ET AL.
© 2013 The Linnean Society of London, Botanical Journal of the Linnean Society, 2013, 173, 573–593

We found similar evolutionary scenarios for allbinary characters, regardless of the model selected(MK1 or AsymmMK, see Appendix). Consequently,only the evolution with the MK1 model is presentedin Figures 3−5. Inferred evolutions of stele type andstele diameter were detailed in the previous study(Dubuisson et al., 2011) and are reported in Figure 3.The evolution of blade length and root system isshown in Figure 4. The large leaves are inferred asancestral in trichomanoids and dwarfism (i.e. appear-ance of small to dwarf leaves) evolved at least twice:in Didymoglossum with a reversal to medium size inDidymoglossum gourlianum (Grev. ex J. Sm.) Pic.Serm.; and in Crepidomanes subgenus Crepidomaneswith a reversal to medium size in Crepidomanesfallax (H.Christ) Ebihara & Dubuisson (Fig. 4).Medium size appeared at least four times: inPolyphlebium [excluding Polyphlebium capillaceum(L.) Ebihara & Dubuisson]; in Trichomanes subgenusLacostea; in the clade (Trichomanes arbuscula Desv.–Trichomanes holopterum Kunze); and in T. polypodi-oides. The evolution of the root system is fullycongruent with the appearance of colonial epiphytism(Fig. 4). Metaxylem evolution (here the appearance oflarge metaxylem) is reported in Figure 5. Largemetaxylem appeared (probably from medium-sizedmetaxylem) five times: in Vandenboschia with areversal in V. maxima; in Cr. aphlebioides; in T. pedi-cellatum; in T. scandens; and in A. flavofuscum.
PHYLOGENETIC COMPARATIVE ANALYSES
When treating ecology as a multistate character(where the appearance of colonial epiphytism isinferred in the common ancestor of the HE clade, seeFig. 2), we demonstrated no significant phylogeneticcorrelation between epiphytism and any morphologi-cal or anatomical character. Conversely, when apply-ing a binary treatment for each ecological type,graphics (Fig. 3A, B) and analyses clearly indicate asignificant phylogenetic correlation between colonialepiphytism and small stele (LR = 22.542, P < 0.001),and between colonial epiphytism and collateral/subcollateral stele (LR = 18.608, P < 0.001). Further-more, the inferred evolution of small metaxylem (notshown here) is identical to the evolution of thecollateral/subcollateral stele; the correlation withcolonial epiphytism could therefore also be applied tosmall metaxylem. Colonial epiphytism, root systemregression (which fully evolves in parallel with colo-nial epiphytism), stele diameter, stele type and smallmetaxylem thus appear to be strongly evolutionarilylinked. Concerning dwarfism, we evidenced no phylo-genetic correlation between small to dwarf sizes andcolonial epiphytism (LR = 9.338, P > 0.05), but thegraphics strongly show that dwarfism is restricted to
two colonial epiphytic clades: Didymoglossum andCrepidomanes subgenus Crepidomanes. However, ifwe combine medium sizes with small to dwarf sizes,the correlation between size and colonial epiphytismbecomes significant (LR = 16.552, P < 0.01), suggest-ing that colonial epiphytism is accompanied by areduction in size in the three colonial epiphytic clades(by adding Polyphlebium) and in colonial epiphyticT. polypodioides, and thus leads to dwarfism in onlytwo of the three colonial epiphytic clades, as devel-oped above. We found no phylogenetic correlationbetween individual epiphytism and any anatomical ormorphological character. The climbing habit appearsto be phylogenetically correlated only with largemetaxylem (Fig. 5; LR = 33.494, P < 0.001).
DISCUSSIONEVOLUTION OF ECOLOGY IN TRICHOMANOIDS
Two distinct inferences of the appearance of colonialepiphytism were obtained depending on the treat-ment of ecology as a multistate character (colonialepiphytism ancestral in HE clade, but with weaksupport; probability 0.71) or as a binary character(three occurrences in the HE clade, see Fig. 2; changeprobabilities > 0.92). As already discussed in the pre-vious study (Dubuisson et al., 2011), the resultobtained with the multistate treatment is also statis-tically expected. Indeed, colonial epiphytism is themost frequently observed state in terminal taxa of theHE clade and therefore has a higher probability to beinferred for the common ancestor of the HE cladethan has hemi-epiphytism, which is less frequent interminal taxa and concentrated in the subclade cor-responding to Vandenboschia. Biologically, the obser-vation in a broad clade of a characteristic shared bythe majority of species does not necessarily imply thatthe other, less frequent traits observed in the sameclade evolved first. This is especially true if the mostcommon trait illustrates one or several recent eventsof rapid and high diversification subsequent to theoccurrence of the other characters, leading to an over-representation in extant taxa. We could suggest anequivalent bias in the study by Hennequin et al.(2008), which proposed a similar evolutionary patternfor epiphytism s.l. in trichomanoids. Therefore, weprefer to select here the evolutionary hypothesis pro-vided with the binary treatment, where change prob-abilities (here the appearance of colonial epiphytism)are also the highest. The choice of this scenario is alsoreinforced as being the unique hypothesis revealing asignificant correlation between a habit (colonial epi-phytism) and important changes in morphology(reduction in size, regression in root system) andanatomy (acquisition of subcollateral/collateral types,
EPIPHYTISM EVOLUTION IN TRICHOMANOID FERNS 581
© 2013 The Linnean Society of London, Botanical Journal of the Linnean Society, 2013, 173, 573–593
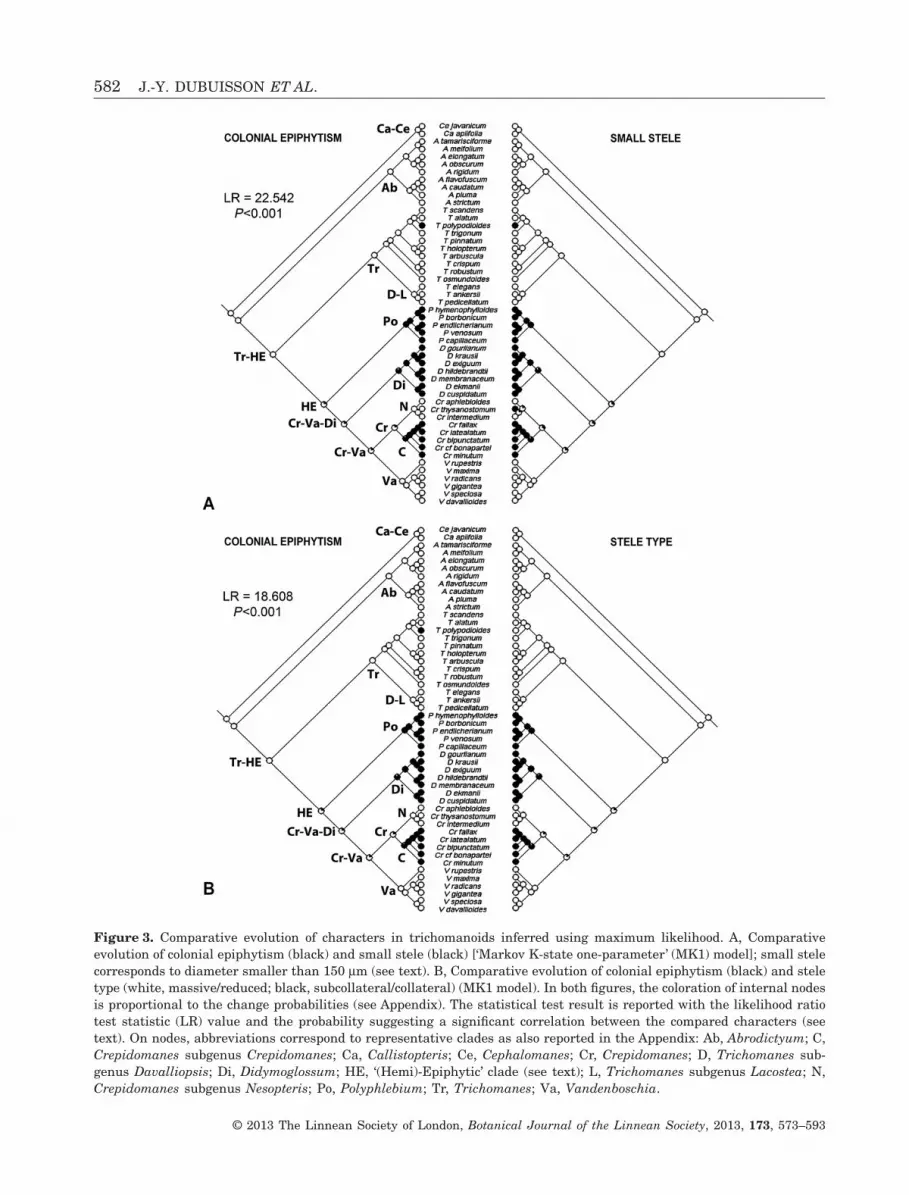
Figure 3. Comparative evolution of characters in trichomanoids inferred using maximum likelihood. A, Comparativeevolution of colonial epiphytism (black) and small stele (black) [‘Markov K-state one-parameter’ (MK1) model]; small stelecorresponds to diameter smaller than 150 μm (see text). B, Comparative evolution of colonial epiphytism (black) and steletype (white, massive/reduced; black, subcollateral/collateral) (MK1 model). In both figures, the coloration of internal nodesis proportional to the change probabilities (see Appendix). The statistical test result is reported with the likelihood ratiotest statistic (LR) value and the probability suggesting a significant correlation between the compared characters (seetext). On nodes, abbreviations correspond to representative clades as also reported in the Appendix: Ab, Abrodictyum; C,Crepidomanes subgenus Crepidomanes; Ca, Callistopteris; Ce, Cephalomanes; Cr, Crepidomanes; D, Trichomanes sub-genus Davalliopsis; Di, Didymoglossum; HE, ‘(Hemi)-Epiphytic’ clade (see text); L, Trichomanes subgenus Lacostea; N,Crepidomanes subgenus Nesopteris; Po, Polyphlebium; Tr, Trichomanes; Va, Vandenboschia.
582 J.-Y. DUBUISSON ET AL.
© 2013 The Linnean Society of London, Botanical Journal of the Linnean Society, 2013, 173, 573–593

reduction in stele diameter and appearance of smallmetaxylem). These changes could illustrate a regres-sive evolution linked to an ecological specialization, asdiscussed later.
INDIVIDUAL EPIPHYTISM
Even in rainforest, a significant decrease in relativeair hygrometry is often possible during a few hours aday, generating water loss in plants. Under relativedrought conditions, even for a short period, terrestrialand climbing hygrophilous species are supposed to be
able to compensate for water loss by absorbing watervia roots anchored in an ever-wet soil. Terrestrialhygrophilous species are often observed near water-falls, along streams or on shady crevices below rocksin forests, where the soil is never dry, and somespecies, such as the Neotropical Trichomanes pinna-tum Hedw. and the Asiatic Cephalomanes javanicum(Blume) C.Presl, often grow in temporarily floodedareas. A decrease in relative air hygrometry is moreproblematic for epiphytic species that have no directconnection to the soil and rely on selected alternativeadaptations to endure water loss. Nitta (2006)
Figure 4. Comparative evolution of blade length, root system and colonial epiphytism in trichomanoids inferred usingmaximum likelihood [‘Markov K-state one-parameter’ (MK1) model]. The coloration of internal nodes is proportional tothe change probabilities (see Appendix). The statistical test shows no significant correlation between small to dwarf size,regression in root system and colonial epiphytism (LR = 9.338, P > 0.05). A significant correlation is nevertheless revealedif small to dwarf size is combined with medium size (LR = 16.552, P < 0.01). On nodes, abbreviations correspond torepresentative clades as also reported in the Appendix: Ab, Abrodictyum; C, Crepidomanes subgenus Crepidomanes; Ca,Callistopteris; Ce, Cephalomanes; Cr, Crepidomanes; D, Trichomanes subgenus Davalliopsis; Di, Didymoglossum; HE,‘(Hemi)-Epiphytic’ clade (see text); L, Trichomanes subgenus Lacostea; N, Crepidomanes subgenus Nesopteris; Po,Polyphlebium; Tr, Trichomanes; Va, Vandenboschia.
EPIPHYTISM EVOLUTION IN TRICHOMANOID FERNS 583
© 2013 The Linnean Society of London, Botanical Journal of the Linnean Society, 2013, 173, 573–593

demonstrated on a restricted and local sample thatcolonial epiphytic members of Hymenophyllaceaewere more drought tolerant than terrestrial types. Inforests in which it rains daily, rapid rehydration isalso facilitated by the absorption of rainwater directlyby the lamina that lacks a cuticle, regardless ofthe ecology and growth form. Individual epiphytes(A. caudatum, A. flavofuscum, A. tamarisciforme,T. crispum and T. alatum) exhibit a morphology andanatomy similar to those of terrestrial relatives and,a priori, no particular adaptation to epiphytism.
These species are found in wet areas where the riskof drought is low. Epiphytic A. tamarisciforme occurson La Réunion Island above 1000 m where annualrainfall exceeds 6000 mm without a significant dryseason, whereas the terrestrial sister species Abrod-ictyum meifolium (Bory ex Willd.) Ebihara & K.Iwats.occurs above 300 m where annual rainfall can bebelow 3000–4000 mm (Grangaud, 2010; J.-Y. Dubuis-son, pers. observ.). Furthermore, except for the wide-spread T. crispum that appears to be opportunistic(Proctor, 1977; Lellinger, 1994; J.-Y. Dubuisson, pers.
Figure 5. Comparative evolution of climbing habit (black) and large metaxylem (black) in trichomanoids inferred usingmaximum likelihood [‘Markov K-state one-parameter’ (MK1) model]. Large metaxylem corresponds to diameter largerthan 41 μm (see text). The coloration of internal nodes is proportional to the change probabilities (see Appendix). Thestatistical test result is reported with the likelihood ratio test statistic (LR) value and the probability, suggesting asignificant correlation between the compared characters (see text). On nodes, abbreviations correspond to representativeclades as also reported in the Appendix: Ab, Abrodictyum; C, Crepidomanes subgenus Crepidomanes; Ca, Callistopteris;Ce, Cephalomanes; Cr, Crepidomanes; D, Trichomanes subgenus Davalliopsis; Di, Didymoglossum; HE, ‘(Hemi)-Epiphytic’clade (see text); L, Trichomanes subgenus Lacostea; N, Crepidomanes subgenus Nesopteris; Po, Polyphlebium; Tr,Trichomanes; Va, Vandenboschia.
584 J.-Y. DUBUISSON ET AL.
© 2013 The Linnean Society of London, Botanical Journal of the Linnean Society, 2013, 173, 573–593

observ.), all individual epiphytes are mostly observedon tree fern trunks (Cyatheaceae; Brownlie, 1969;Proctor, 1977; Ebihara, 2002; Grangaud, 2010; J.-Y.Dubuisson, pers. observ.) with their stems and rootsystem embedded in the adventitious root mantlecovering the tree fern trunk, but there does notappear to be a strict specific relationship. This sub-strate is likely to retain more water than smoothbark, where such species are not or rarely observed.Most individual epiphytes are rarely observed above10 m on trunks, especially if they grow on tree ferns(exceptionally > 20 m for Cyatheaceae in the IndianOcean; Janssen & Rakotondrainibe, 2008). Lower ontree fern trunks, the environment of epiphytes issimilar to conditions of the understorey with lowhygrometric fluctuations compared with higher up inthe canopy. This environment is thus favourable forepiphytic hygrophytes that are also named ‘understo-rey epiphytes’. In Madagascar and the MascareneIslands, A. tamarisciforme is found on various treefern species, but not exclusively (Grangaud, 2010;J.-Y. Dubuisson, pers. observ.). In the area, tree fernsradiated quite recently in the upper Tertiary (Plio-cene) according to Janssen et al. (2008). We thereforesuggest that individual epiphytism may thus haveappeared several times by parallelism from terres-trial clades, probably at least in the Tertiary (seeFig. 7), and that it does not illustrate any ecologicaldiversification, radiation or ‘success’, contrary to colo-nial epiphytism as discussed below. The presence ofthese species as epiphytes on tree ferns rather than ina terrestrial habitat, as displayed by their close rela-tives, could be conditioned by preferences of thegametophyte.
COLONIAL EPIPHYTISM
All colonial epiphytic species belonging to Didymo-glossum, Polyphlebium and Crepidomanes subgenusCrepidomanes share regressive morphology andanatomy (reduction in size, stem thickness, stele typeand diameter, metaxylem diameter and reduction inroot system to rootless type). These regressive ten-dencies appear to be statistically well related tocolonial epiphytism acquisition (see Figs 3 and 4). Asfor individual climbing and non-climbing epiphytes,they are understorey epiphytes and not or only rarelyobserved in canopy situations. Some species are gen-eralist and found on various substrates [such asDidymoglossum cuspidatum (Willd.) Ebihara &Dubuisson observed on tree trunks and branches,emerging roots, wet rocks or stumps; Grangaud, 2010;J.-Y. Dubuisson, pers. obs.], whereas others seem tobe more specialized [such as Didymoglossum godma-nii (Hook.) Ebihara & Dubuisson, mostly occurring onthe palm Welfia georgii H.Wendl.; Moran & Russel,
2004], even though they are phylogenetically related(D. godmanii was not sampled here but belongs to thesame subgenus, Microgonium C.Presl, as D. cuspida-tum). Colonial epiphytic species have the ability tocolonize a broad surface with their highly branchedand long-creeping filiform rhizomes (see Fig. 6), andto adhere to smooth bark and hard substrates (includ-ing rocks) by using a dense cauline indumentum inaddition to branched root-like shoots for rootless taxa(sensu Schneider, 2000), rather than their few roots ifpresent.
On vertical substrates, leaves are pendant orappressed to the substrate. In Polyphlebium, pendantleaves of various sizes are always dissected and some-times have capillary segments (e.g. P. capillaceum). Insubgenus Crepidomanes, pendant leaves are alwaysdissected (Fig. 6A), even in the smallest species, suchas Crepidomanes minutum (Blume) K.Iwats., whichoften exhibits a finely dissected flabellate lamina(Fig. 6B). In Didymoglossum, a few species [e.g.D. gourlianum and Didymoglossum krausii (Hook. &Grev.) C.Presl] are pinnatifid with pendant leaves;most other species display lobed to rounded laminathat are pendant or appressed to substrates, espe-cially the smallest ones (Fig. 6C). Didymoglossumhildebrandtii (Kuhn) Ebihara & Dubuisson from theWestern Indian Ocean Islands and Didymoglossumtahitense (Nadeaud) Ebihara & K.Iwats. from Poly-nesia are related species (Ebihara et al., 2007), andare characterized by rounded to peltate leaves tightlyappressed on the smooth barks of tree trunks(Fig. 6D, E), resulting in a habit that is unusual forferns and more frequently observed in thallose liver-worts and hornworts. Regardless of leaf architectureand size, all colonial epiphytes, with their large colo-nies of pendant or appressed thin blades lacking acuticle, have the capacity to capture rainwaterdirectly via the lamina rather than via their reducedroot system. They especially capture the rainwaterflowing along vertical substrates during and afterdaily rainfall. This could enable fast rehydration inthe case of short periods of drought. The preferentialuse of leaves for water absorption, probably by diffu-sion, could explain the reduction in the root systemand in vascular tissues in the stems, which wouldhave been counter-selected.
EPIPHYTISM AND DWARFISM
Dwarfism, here defined as the reduction in leaf size,for epiphytic filmy ferns is preponderant only inDidymoglossum and Crepidomanes subgenus Crepi-domanes, and the smallest species are observed inDidymoglossum (with Didymoglossum nummulariumBosch displaying leaves that do not exceed 0.3 cmin length). This contrasts with one of the largest
EPIPHYTISM EVOLUTION IN TRICHOMANOID FERNS 585
© 2013 The Linnean Society of London, Botanical Journal of the Linnean Society, 2013, 173, 573–593

Figure 6. Representative colonial epiphytic trichomanoids in situ. A, Crepidomanes bipunctatum (Poir.) Copel., colonizinga tree trunk in Mauritius rainforest, with pendant highly dissected leaves that can attain 5–6 cm in length (photographby J.-Y. Dubuisson). B, Crepidomanes minutum (Blume) K.Iwats., minute species growing on a tree trunk in Mauritiusrainforest; detail of two leaves of a colony that can colonize a broad surface; the largest leaf illustrated here bears twosori on a flabellate dissected lamina; leaves of this species rarely exceed 2.5 cm in length in Mascarenes (photograph byJ.-Y. Dubuisson). C, Didymoglossum barklianum (Hook. ex Baker) J.P.Roux, dwarf species colonizing a tree trunk inMauritian rainforest with simple leaves that do not exceed 0.8 cm in length, mostly pendant or appressed on the substrate(photograph by J.-Y. Dubuisson). D, Didymoglossum hildebrandtii (Kuhn) Ebihara & Dubuisson colonizing a stump in arainforest on Grande Comore (Comoros Islands) (photograph by G. Rouhan). E, Detail of D. hildebrandti, rounded andpeltate leaves, strongly appressed on tree bark, the central one bearing many marginal sori, leaves rarely exceeding 3 cmin length (photograph by G. Rouhan).
586 J.-Y. DUBUISSON ET AL.
© 2013 The Linnean Society of London, Botanical Journal of the Linnean Society, 2013, 173, 573–593

trichomanoid species, Callistopteris polyantha (Hook.)Copel., a terrestrial taxon that shows highly divided,erect leaves exceeding 50 cm in length (Dubuissonet al., 2003). The gross morphological resemblance tobryophytes, especially to thallose liverworts, is mostlyobserved in Didymoglossum (see Fig. 6D, E). Thisresemblance is, of course, superficial; dwarf filmyferns are still vascular plants (even with regressedconducting tissues). The lobed to rounded leaf thatreinforces this gross resemblance in Didymoglossumcould be the consequence of developmental con-straints linked to sorus position. Members of Hyme-nophyllaceae are characterized by sori producedmarginally in the continuity of a single vein. Inspecies with highly dissected leaves, the ultimatesegments contain a unique vein, resulting in a singlesorus per segment. Polyphlebium and Crepidomanesspecies display an anadromous venation combinedwith marginal sori always localized on lateral veins,rarely also on the terminal ones, whereas Didymo-glossum species have catadromous venation combinedwith marginal sori that always develop on terminalveins and sometimes on the lateral ones (Dubuisson,1997). This implies that Polyphlebium and Crepido-manes species always have dissected leaves withsingle-veined segments. Consequently, the leaf of thesmallest Crepidomanes species is reduced to a singleor few segments with a limited number of sori (asobserved in Cr. minutum, see Fig. 6B, Crepidomanesintramarginale (Hook. & Grev.) Copel., or Crepido-manes kurzii (Bedd.) Tagawa & K. Iwatsuki). InDidymoglossum, the obligatory terminal sorus posi-tion seems to be accompanied by the ability to ‘later-ally’ develop a lamina involving more than one veinand often additional false veins (Dubuisson, 1997),allowing the leaf to acquire a rounded to peltatearchitecture and to bear many sori on the largestleaves (see Fig. 6E).
The ontogeny of fern sporophytes is well documentedand it has been shown that, in species with highlydissected mature leaves, the first (or juvenile) leaves ofthe sporophyte are generally less dissected than thefollowing ones, thus indicating an increase in leafdissection with age, defined as a heteroblastic devel-opment (Bower, 1963; Allsopp, 1965). Didymoglossumspecies, with their lobed to rounded leaves, couldillustrate a developmental heterochronic process, andespecially paedomorphy (here neoteny; as defined byGould, 1988), with mature fertile individuals retainingthe morphology of a young sporophyte. Developmentalconstraints related to the sorus position and the het-erochronic process hypothesis could explain whydwarfism and the gross morphological resemblance tothallose liverworts are exacerbated in catadromousDidymoglossum. The gross resemblance to thalloseliverworts, as a consequence of regressive evolution in
morphology and anatomy leading to dwarfism, com-bined with probable preferential water absorption bythe lamina, seems to have been selected for Didymo-glossum. It is therefore not unexpected to observe thedwarf species sympatrically and often mixed withliverworts and mosses.
Because medium-sized to large colonial epiphytes(as observed in Polyphlebium) are, a priori, not lessefficient at growing in similar habitats and in captur-ing rainwater than minute taxa, the selection ofdwarfism in filmy ferns should be explained by addi-tional factors. The smallest species of Didymoglossumand Crepidomanes are found mostly in the shadiestplaces, at the base of trunks and under largebranches, or on very wet rocks, and always close tothe substrate and mostly mixed with bryophytes.These represent potentially various microhabitats inwhich high levels of hygrometry are maintainedthroughout the year. These conditions are requiredfor hygrophilous mosses and, especially, liverworts tolimit water loss. We suggest that dwarf epiphyticspecies of filmy ferns could be less drought tolerantthan the largest ones, hence their colonization of thebryophyte-dominated microhabitats. However, thishypothesis remains speculative in the absence ofstudies on the drought tolerance of dwarf trichoman-oids. Dwarfism related to epiphytism is not rare inother groups of plants, but it always involves xero-phyte strategies (e.g. succulent blades in small Lem-maphyllum C. Presl and Microgramma C. Preslspecies, Polypodiaceae; dwarfism in orchids and bro-meliads; see Benzing & Ott, 1981). The ‘bryophyte-like’ ecology of dwarf filmy fern species appears inaccordance with an epiphytic ‘bryophyte-like’ strategythat is unique in vascular plants, but which is not theunique epiphytism strategy in general selected forHymenophyllaceae.
DIVERSIFICATION OF EPIPHYTISM IN TRICHOMANOIDS
The few colonial epiphytes outside of the HE cladeinclude T. polypodioides (in subgenus Trichomanes),often observed on tree fern trunks and having thinbranched creeping stems with a small stele, pendantleaves of medium size and filiform roots (Proctor,1977; Lellinger, 1994; J.-Y. Dubuisson, pers. observ.).This species has a massive stele, a character alsopresent in its terrestrial relatives. Other colonial epi-phytes, such as Trichomanes anadromum Rosenst.and Trichomanes paucisorum R.C.Moran & B.Øllg.(Moran & Øllgaard, 1998), were not available for thisstudy, but they have filiform long-creeping stems andundoubtedly belong to Trichomanes subgenus Tricho-manes. If a close relationship was to be confirmedbetween T. polypodioides and these taxa, this wouldsuggest a colonial epiphytic specialization in a small
EPIPHYTISM EVOLUTION IN TRICHOMANOID FERNS 587
© 2013 The Linnean Society of London, Botanical Journal of the Linnean Society, 2013, 173, 573–593
clade in Trichomanes, rather than independently in afew isolated species.
The independent appearance of colonial epiphytismin three major clades at the end of the Cretaceous andin the Tertiary, according to molecular dating per-formed by Hennequin et al. (2008) (see Fig. 7), is inagreement with the hypothesis that most extant epi-
phytic ferns would have diversified during theseperiods (Schuettpelz, 2007; Schuettpelz & Pryer,2009). This result corroborates the preliminaryhypothesis suggested for Hymenophyllaceae of adiversification of epiphytic filmy ferns at least in theLate Cretaceous (Dubuisson et al., 2009). In the samecontext, the localized occurrence of colonial epiphyt-
Figure 7. Synthesized evolutionary hypotheses on ecology, growth form, morphology and anatomy in trichomanoids. Thefigure (completed and modified from Dubuisson et al., 2009) reports the inferred appearances of climbing habit,epiphytism (colonial and individual), colonial growth form, heterogeneous cortex and anatomy regression (with potentialreversals if present). Evolutionary correlations between ecology, morphology, growth form and anatomy are detailed anddiscussed in the text. Divergence time estimates are from Hennequin et al. (2008). The age values selected for the nodesare means calculated from the single gene analysis and are reported with standard deviation (SD) in Hennequin et al.(2008: table 2). Left vertical line (in Cretaceous) and grey portion of the time scale indicate the diversification of modernfern families providing more than 80% of living species, in parallel with the angiosperm diversification (according toSchuettpelz, 2007). Abbreviations on the right correspond to representative clades: Ab, Abrodictyum; C, Crepidomanessubgenus Crepidomanes; Ca, Callistopteris; Ce, Cephalomanes; Cr, Crepidomanes; D, Trichomanes subgenus Davalliopsis;Di, Didymoglossum; F, Trichomanes subgenus Feea; L, Trichomanes subgenus Lacostea; N, Crepidomanes subgenusNesopteris; Po, Polyphlebium; T, Trichomanes subgenus Trichomanes, Tr, Trichomanes; Va, Vandenboschia.
588 J.-Y. DUBUISSON ET AL.
© 2013 The Linnean Society of London, Botanical Journal of the Linnean Society, 2013, 173, 573–593

ism in Trichomanes subgenus Trichomanes (T. polypo-dioides, see Fig. 2), localized occurrences of individualepiphytism (A. tamarisciforme, T. alatum, T. crispum,see Fig. 2) and multiple acquisitions of a climbinghabit (see Fig. 5) could also be dated back to theTertiary (Hennequin et al., 2008) during the develop-ment of current tropical rainforests dominated byangiosperms. These Tertiary tropical rainforests prob-ably provided ecological facilities for the appearanceof such habits and for the diversification of colonialepiphytes (Behrensmeyer et al., 1992; Willis &McElwain, 2002; Schuettpelz & Pryer, 2009).
ANATOMY AND CLIMBING HABIT
Concerning the climbing habit, our analysis stronglysuggests that, by convergence with angiosperm lianas,as demonstrated by Carlquist (1991), the largest sizefor equivalent conducting tissues (here tracheids)would have been selected in climbing trichomanoidtaxa, with the consequence of a potentially higherefficiency of water conduction in the long robust stemsof these species. Extant hemi-epiphytic climbing filmyferns produce medium-sized to large fertile leaves afew metres above the ground (to maximize sporedispersion) under conditions in which water loss, as forstrict epiphytes, is not negligible. By keeping a rela-tionship to the ground via a developed root system,hemi-epiphytes have the possibility to absorb ground-water, the conduction of which to more distant leavesmay be facilitated by the large stele and metaxylem. Incontrast, colonial non-climbing holo-epiphytes havelong-creeping, lax and filiform rhizomes with reducedconducting tissues that are sufficient for a colonialstrategy on the epiphytic substrate. Dubuisson et al.(2011) showed that climbing taxa outside the HE cladeand all taxa belonging to the HE clade have a hetero-geneous cortex, which appears to be significantlyrelated to a colonial strategy and is conserved incolonial epiphytes. The present study infers thecommon ancestor of the HE clade as an individualterrestrial. However, as already proposed in the pre-vious study, we cannot reject the possibility that thisancestor would have been hemi-epiphytic, becauseinference is strongly conditioned by the current under-representation of hemi-epiphytic taxa in the HE clade.The extant distribution of hemi-epiphytism in the HEclade is limited to two subclades and a few isolatedtaxa in trichomanoids (Dubuisson et al., 2011), but thiscould hide the possibility that hemi-epiphytic taxawere dominant during the clade diversification, esti-mated to have occurred during the Cretaceous in theunderstorey of early angiosperm-dominated forests.With this alternative hypothesis, regressed colonialepiphytes would have derived from robust hemi-epiphytes in the HE clade.
IMPORTANCE OF THE GAMETOPHYTE GENERATION
In ferns, the sporophyte ecology is strongly linked tothe gametophyte habitat, which determines wherethe sporophyte will grow, but fern gametophyteecology has not yet been studied in sufficient detail. Intrichomanoids, one study has suggested that therecruitment and growth habit of the hemi-epiphyticV. collariata depends on the habitat of the gameto-phytes, which seems to be restricted to the bases oftrees (Nitta & Epps, 2009). The few studies performedon the subject have provided promising insights, suchas strong differences in ecological preferences andbiology of the gametophyte generation between ter-restrial, epiphytic and climbing species. Dassler &Farrar (2001) suggested that the success of epiphyticferns, especially in long-distance dispersal and colo-nization, could be related to the perennial and long-lived strategy of epiphytic gametophytes, comparedwith short-lived terrestrial ones (at least in Polypo-diales). Watkins, Mack & Mulkey (2007) and Watkinset al. (2007) further showed that epiphytic gameto-phytes are more drought tolerant than terrestrialones. The long-lived strategy and relative droughttolerance of epiphytic gametophytes could also berelated to competition with epiphytic bryophytes andto the increase in the outcrossing probability in ahabitat in which gametophyte establishment fromspores is limited (see also the review of Farrar et al.,2008). Members of Hymenophyllaceae are character-ized by perennial, long-lived and colonial gameto-phytes, and some populations, especially in temperateareas, are also known to subsist only as gametophytesreproducing asexually by gemmae (Farrar, 1990,1992; Rumsey et al., 1999). The gametophytes of filmyferns thus appear (pre-)adapted to epiphytic habitats.In this sense, we disagree with Farrar et al. (2008),who proposed that gametophytes of terrestrial Hyme-nophyllaceae [citing Abrodictyum rigidum (Sw.)Ebihara & Dubuisson and Trichomanes osmundoidesPoir.] would have retained the form of their epiphyticrelatives, assuming terrestrial species to be derived inthe family. We propose, instead, that gametophytes ofterrestrial species probably exhibit the ancestralstate, because terrestrial habitat is inferred as ances-tral. Epiphytic sporophytes, as in Didymoglossum,would have more or less retained the properties oftheir gametophytes, probably in a competitive contextwith liverworts and other mosses. A precise compara-tive study on the ecological preferences, biology anddrought tolerance of gametophytes of terrestrial,hemi-epiphytes and individual or colonial epiphytesin Hymenophyllaceae using phylogenetic analysis istherefore needed. The ecological preference of thegametophyte could also explain why some speciesare opportunistic and some specialized (e.g. how
EPIPHYTISM EVOLUTION IN TRICHOMANOID FERNS 589
© 2013 The Linnean Society of London, Botanical Journal of the Linnean Society, 2013, 173, 573–593

individual climbing or non-climbing epiphytes aremostly observed on tree ferns, whereas their relativesare terrestrial). Filmy ferns are a pertinent model forthe study of ecology evolution, not only of the fernsporophyte, but also of the gametophyte, one of thefuture directions of study in fern ecology proposed byWalker, Mehltreter & Sharpe (2010).
CONCLUSIONS AND PROSPECTIVE WORK
Evolutionary scenarios of ecology, growth forms(according to our previous study; Dubuisson et al.,2011), gross morphology and anatomy for the tricho-manoids are summarized in Figure 7, with a proposedtime scale following Hennequin et al. (2008). The firsttrichomanoids were inferred as terrestrial, probablyrobust plants displaying individual growth, standardwell-developed protosteles, developed root systemsand erect leaves (as observed for all extant terrestrialtrichomanoids). A colonial growth form appears tohave been selected in a clade during the lower Cre-taceous, probably in the understorey of the firstangiosperm-dominated forests. Although our analysesdid not statistically propose that these first colonialfilmy ferns would be climbing hemi-epiphytes, thehypothesis of a hemi-epiphytic appearance related tocoloniality in the HE clade is not fully rejected, assuggested by the heterogeneous cortex that mayhave been selected in relation to a climbing habit(Dubuisson et al., 2011). Colonial epiphytism couldhave appeared in the HE clade, maybe from hemi-epiphytism, at least in three clades that diversifiedduring the upper Cretaceous or the Tertiary. This firstperiod corresponds to the time estimated for thediversification of most modern fern lineages in the‘shadow’ of the angiosperms (Schneider et al., 2004b),and the second period corresponds to epiphytismacquisition in other fern families (Schuettpelz, 2007;Schuettpelz & Pryer, 2009). Morphological and ana-tomical regression is observed in these three colonialepiphytic clades and would be related to water acqui-sition by the pendant or appressed blades ratherthan by a root system. A ‘bryophyte-like’ strategy isobserved in one clade (Didymoglossum). Outside theHE clade, parallel acquisition of climbing habit andepiphytism (individual and colonial) could probablyhave occurred more or less recently in the Tertiary.
This study deliberately excluded the Hymenophyl-lum sister clade, most members of which are colonialepiphytic or epilithic species with pendant leaves(with few individual epiphytic exceptions), whichwould have diversified in parallel with trichomanoidcolonial epiphytic clades (Hennequin et al., 2008).This genus differs from the trichomanoids in anato-mical diversity [with three stele types: reduced;subcollateral (rarely collateral); and dorsiventral:
Hennequin et al., 2006] and in the absence of cladescomprising dwarf forms resembling thallose liver-worts. An analogous comparative study in this lineageis required to provide hypotheses on the strategiesthat allowed Hymenophyllum to succeed in diversify-ing in the epiphytic habitat at the pantropical levelwith similar species richness, and to reveal potentialparallel evolution or distinct strategies in the twolineages at the family level.
ACKNOWLEDGEMENTS
This work, and especially the field trips for specimenacquisition, was supported by UMR 7207 ‘Centre deRecherche sur la Paléobiodiversité et les Paléoenvi-ronnements‘, IFR 101 ‘Institut d’Ecologie, Biodiver-sité, Evolution, Environnement‘ and the PPF MNHN‘Etat et Structure Phylogénétique de la BiodiversitéActuelle et Fossile’. The molecular work for providingthe phylogenetic framework was partly supported byJSPS fellowship for A. Ebihara. We thank G. Rouhan,F. Rakotondrainibe and J.-N. Labat for the use of Pherbarium specimens, C. Chaussidon, G. Rouhan, F.Rakotondrainibe, C. Reeb, E. Grangaud, A. LeThomas and H. Schneider for discussions on filmyfern anatomy, biology and ecology, and T. Dufour, N.Salel and E. Watroba for precious help in anatomyinvestigations. We also thank two anonymous review-ers for their pertinent commentaries, suggestions andcorrections.
REFERENCES
Allsopp A. 1965. Heteroblastic development in cormophytes.In: Ruhland W, ed. Handbuch der Pflanzenphysiologie.Berlin: Springer-Verlag, 1172–1221.
Behrensmeyer AK, Damuth JD, DiMichele WA, Potts R,Sues H-D, Wing SL. 1992. Terrestrial ecosystems throughtime: evolutionary paleoecology of terrestrial plants andanimals. Chicago, IL: University of Chicago Press.
Benzing DH. 1990. Vascular epiphytes. Cambridge: Cam-bridge University Press.
Benzing DH, Ott DW. 1981. Vegetative reduction in epi-phytic Bromeliaceae and Orchidaceae: its origin and signifi-cance. Biotropica 13: 131–140.
Bower FO. 1963. The ferns (Filicales), Vol. 1. Analyticalexamination of the criteria of comparison (reprint edition).New Delhi: Today & Tomorrow’s Book Agency.
Brownlie G. 1969. Flore de la Nouvelle Calédonie etDépendances, volume 3. Ptéridophytes. Paris: MuséumNational d’Histoire Naturelle.
Carlquist S. 1991. Anatomy of vine and liana stems: a reviewand synthesis. In: Putz FE, Mooney HA, eds. The biology ofvines. Cambridge: Cambridge University Press, 53–71.
Christenhusz MJM, Zhang X-C, Schneider H. 2011. Alinear sequence of extant families and genera of lycophytesand ferns. Phytotaxa 19: 7–54.
590 J.-Y. DUBUISSON ET AL.
© 2013 The Linnean Society of London, Botanical Journal of the Linnean Society, 2013, 173, 573–593

Dassler CL, Farrar DR. 2001. Significance of gametophyteform in long-distance colonization by tropical epiphyticferns. Brittonia 53: 352–369.
Dubuisson J-Y. 1997. Systematic relationships within thegenus Trichomanes sensu lato (Hymenophyllaceae, Filicop-sida): cladistic analysis based on anatomical and morpho-logical data. Botanical Journal of the Linnean Society 123:265–296.
Dubuisson JY, Hennequin S, Bary S, Ebihara A,Boucheron-Dubuisson E. 2011. Anatomical diversity andregressive evolution in trichomanoid filmy ferns (Hymeno-phyllaceae): a phylogenetic approach. Comptes RendusBiologies 334: 880–895.
Dubuisson J-Y, Hennequin S, Rakotondrainibe F,Schneider H. 2003. Ecological diversity and adaptive ten-dencies in the tropical fern Trichomanes L. (Hymenophyl-laceae) with special reference to climbing and epiphytichabits. Botanical Journal of the Linnean Society 142: 41–63.
Dubuisson J-Y, Hennequin S, Schneider H. 2009. Epi-phytism in ferns: diversity and evolution. Comptes RendusBiologies 332: 120–128.
Ebihara A. 2002. Hymenophyllaceae Flora of WesternOceania and its origin. MSc Dissertation, University ofTokyo.
Ebihara A, Dubuisson J-Y, Iwatsuki K, Hennequin S, ItoM. 2006. A taxonomic revision of Hymenophyllaceae.Blumea 51: 221–280.
Ebihara A, Iwatsuki K, Ito M, Hennequin S, DubuissonJ-Y. 2007. A global molecular phylogeny of the fern genusTrichomanes (Hymenophyllaceae) with special reference tostem anatomy. Botanical Journal of the Linnean Society155: 1–27.
Farrar DR. 1990. Species and evolution in asexually repro-ducing independent fern gametophytes. Systematic Botany15: 98–111.
Farrar DR. 1992. Trichomanes intricatum: the independentTrichomanes gametophyte in the eastern United States.American Fern Journal 82: 68–74.
Farrar DR, Dassler CL, Watkins JJE, Skelton C. 2008.Gametophyte ecology. In: Ranker TA, Haufler CH, eds. Thebiology and evolution of ferns and lycophytes. Cambridge:Cambridge University Press, 224–258.
Felsenstein J. 1981. Evolutionary trees from DNAsequences: a maximum likelihood approach. Journal ofMolecular Evolution 17: 368–376.
Gentry AH, Dodson CH. 1987. Diversity and biogeographyof Neotropical vascular epiphytes. Annals of the MissouriBotanical Garden 74: 205–233.
Gould SJ. 1988. The uses of heterochrony. In: McKinney ML,ed. Heterochrony in evolution: a multidisciplinary approach.New York: Plenum Press, 1–13.
Grangaud E. 2010. Guide des fougères et plantes alliées desMascareignes. Paris: MNHN/Biotope.
Haufler CH, Grammer WA, Hennipman E, Ranker TA,Smith AR, Schneider H. 2003. Systematics of the ant-ferngenus Lecanopteris (Polypodiaceae): testing phylogenetichypotheses with DNA sequences. Systematic Botany 28:217–227.
Hennequin S, Ebihara A, Ito M, Iwatsuki K, DubuissonJ-Y. 2006. New insights into the phylogeny of Hymenophyl-lum (Hymenophyllaceae): revealing the polyphyly of Meco-dium. Systematic Botany 31: 271–284.
Hennequin S, Schuettpelz E, Pryer KM, Ebihara A,Dubuisson J-Y. 2008. Divergence times and the evolutionof epiphytism in filmy ferns (Hymenophyllaceae) revisited.International Journal of Plant Sciences 169: 1278–1287.
Hietz P. 2010. Fern adaptations to xeric environments. In:Mehltreter K, Walker LR, Sharpe JM, eds. Fern ecology.Cambridge: Cambridge University Press, 140–176.
Hietz P, Briones O. 1998. Correlation between water rela-tions and within-canopy distribution of epiphytic ferns in aMexican cloud forest. Oecologia 114: 305–316.
Hietz P, Briones O. 2001. Photosynthesis, chlorophyll fluo-rescence and within-canopy distribution of epiphytic fernsin a Mexican cloud forest. Plant Biology 3: 279–287.
Hosokawa T. 1943. Studies on the life forms of vascularepiphytes and the epiphyte flora of Ponape, Micronesia.Transactions of the Natural History Society of Taiwan 33:35–55.
Janssen T, Bystriakova N, Rakotondrainibe F, CoomesD, Labat J-N, Schneider H. 2008. Neoendemism in Mada-gascan tree ferns results from recent, coincident diversifi-cation bursts. Evolution 62: 1876–1889.
Janssen T, Rakotondrainibe F. 2008. A revision of theindusiate scaly tree ferns (Cyatheaceae, Cyathea subgen.Alsophila sect. Alsophila) in Madagascar, the Comoros andthe Seychelles. Adansonia 30: 221–376.
Janssen T, Schneider H. 2005. Exploring the evolution ofhumus collecting leaves in drynarioid ferns (Polypodiaceae,Polypodiidae) based on phylogenetic evidence. Plant System-atics and Evolution 252: 175–197.
Kreier HP, Schneider H. 2006. Phylogeny and biogeogra-phy of the staghorn fern genus Platycerium (Polypodiaceae,Polypodiidae). American Journal of Botany 93: 217–225.
Lellinger DB. 1994. Hymenophyllaceae. In: Gorts-van RijnARA, ed. Flora of the Guianas, series B: ferns and fern allies,Fasc.3. Königstein: Koeltz Scientific Books.
Maddison WP, Maddison DR. 2010. Mesquite: a modularsystem for evolutionary analysis, version 2.7.3. Available at:http://mesquiteproject.org
Moran RC, Øllgaard B. 1998. New species of ferns (Polypo-diopsida) from Ecuador. Nordic Journal of Botany 18: 431–439.
Moran RC, Russel RV. 2004. The occurrence of Trichomanesgodmanii (Hymenophyllaceae) on Welfia georgii (Arecaceae)at the La Selva Biological Station, Costa Rica. AmericanFern Journal 94: 70–76.
Nitta JH. 2006. Distribution, ecology, and systematics of thefilmy ferns (Hymenophyllaceae) of Moorea, French Polyne-sia. Water Resources Center Archives, Biology and Geomor-phology of Tropical Islands (ESPM 107/IB 158), Universityof California, Berkeley, CA. Available at: http://repositories.cdlib.org/wrca/moorea/nitta
Nitta JH, Epps MJ. 2009. Hemi-epiphytism in Vanden-boschia collariata (Hymenophyllaceae). Brittonia 61: 392–397.
EPIPHYTISM EVOLUTION IN TRICHOMANOID FERNS 591
© 2013 The Linnean Society of London, Botanical Journal of the Linnean Society, 2013, 173, 573–593

Ogura Y. 1972. Comparative anatomy of vegetative organs ofthe pteridophytes. Berlin: Gebrüder Borntraeger.
Oliver WRB. 1930. New Zealand epiphytes. Journal ofEcology 18: 1–50.
Pagel M, Meade A. 2006. Bayesian analysis of correlatedevolution of discrete characters by reversible-jump Markovchain Monte Carlo. The American Naturalist 167: 808–825.
Proctor GR. 1977. Hymenophyllaceae. In: Howard RA, ed.Flora of the Lesser Antilles, Leeward and Windward Islands,Vol. 2, Pteridophyta. Jamaica Plain: Arnold Arboretum,Harvard University, 64–97.
Rumsey FJ, Vogel JC, Russell SJ, Barrett JA, Gibby M.1999. Population genetics and conservation biology of theendangered fern Trichomanes speciosum (Hymenophyl-laceae) in Scotland. Biological Journal of the LinneanSociety 66: 333–344.
Schneider H. 2000. Morphology and anatomy of roots in thefilmy fern tribe Trichomaneae H. Schneider (Hymenophyl-laceae, Filicatae) and the evolution of rootless taxa. Botani-cal Journal of the Linnean Society 132: 29–46.
Schneider H, Schuettpelz E, Pryer KM, Cranfill R,Magallón S, Lupia R. 2004b. Ferns diversified in theshadow of angiosperms. Nature 428: 553–557.
Schneider H, Smith AR, Cranfill R, Hildebrand TJ,Haufler CH, Ranker TA. 2004a. Unrevealing the phylog-eny of polygrammoid ferns (Polypodiaceae and Grammiti-
daceae): exploring aspects of the diversification of epiphyticplants. Molecular Phylogenetics and Evolution 31: 1041–1063.
Schuettpelz E. 2007. The evolution and diversification ofepiphytic ferns. PhD Thesis, Duke University, Durham, NC.
Schuettpelz E, Pryer KM. 2009. Evidence for a Cenozoicradiation of ferns in an angiosperm-dominated canopy. Pro-ceedings of the National Academy of Sciences of the UnitedStates of America 106: 11 200–11 205.
Smith AR, Pryer KM, Schuettpelz E, Korall P,Schneider H, Wolf PG. 2006. A classification for extantferns. Taxon 55: 705–731.
Walker LR, Mehltreter K, Sharpe JM. 2010. Current andfuture directions in fern ecology. In: Mehltreter K, WalkerLR, Sharpe JM, eds. Fern ecology. Cambridge: CambridgeUniversity Press, 360–378.
Wallace BJ. 1981. The Australian vascular epiphytes: floraand ecology. PhD Thesis, University of New England, NSW.
Watkins JJE, Mack MC, Mulkey SS. 2007. Gametophyteecology and demography of epiphytic and terrestrial tropicalferns. American Journal of Botany 94: 701–708.
Watkins JJE, Mack MC, Sinclair TR, Mulkey SS. 2007.Ecological and evolutionary consequences of desiccation tol-erance in tropical fern gametophytes. New Phytologist 176:708–717.
Willis KJ, McElwain JC. 2002. The evolution of plants.Oxford: Oxford University Press.
592 J.-Y. DUBUISSON ET AL.
© 2013 The Linnean Society of London, Botanical Journal of the Linnean Society, 2013, 173, 573–593
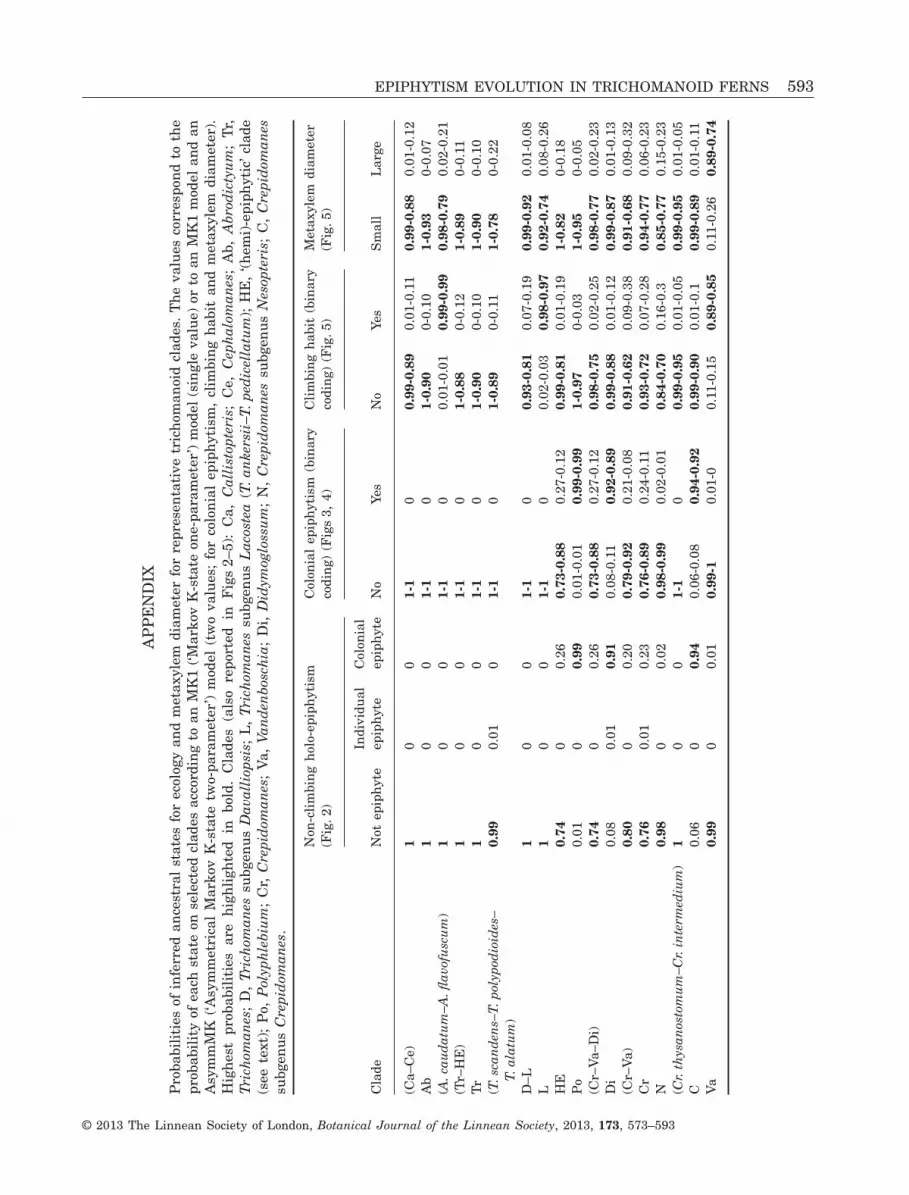
AP
PE
ND
IX
Pro
babi
liti
esof
infe
rred
ance
stra
lst
ates
for
ecol
ogy
and
met
axyl
emdi
amet
erfo
rre
pres
enta
tive
tric
hom
anoi
dcl
ades
.T
he
valu
esco
rres
pon
dto
the
prob
abil
ity
ofea
chst
ate
onse
lect
edcl
ades
acco
rdin
gto
anM
K1
(‘Mar
kov
K-s
tate
one-
para
met
er’)
mod
el(s
ingl
eva
lue)
orto
anM
K1
mod
elan
dan
Asy
mm
MK
(‘Asy
mm
etri
cal
Mar
kov
K-s
tate
two-
para
met
er’)
mod
el(t
wo
valu
es;
for
colo
nia
lep
iph
ytis
m,
clim
bin
gh
abit
and
met
axyl
emdi
amet
er).
Hig
hes
tpr
obab
ilit
ies
are
hig
hli
ghte
din
bold
.C
lade
s(a
lso
repo
rted
inF
igs
2–5)
:C
a,C
alli
stop
teri
s;C
e,C
eph
alom
anes
;A
b,A
brod
icty
um
;Tr
,Tr
ich
oman
es;
D,
Tric
hom
anes
subg
enu
sD
aval
liop
sis;
L,
Tric
hom
anes
subg
enu
sL
acos
tea
(T.a
nke
rsii
–T.p
edic
ella
tum
);H
E,
‘(hem
i)-e
piph
ytic
’cla
de(s
eete
xt);
Po,
Pol
yph
lebi
um
;C
r,C
repi
dom
anes
;V
a,Va
nd
enbo
sch
ia;
Di,
Did
ymog
loss
um
;N
,C
repi
dom
anes
subg
enu
sN
esop
teri
s;C
,C
repi
dom
anes
subg
enu
sC
repi
dom
anes
.
Cla
de
Non
-cli
mbi
ng
hol
o-ep
iph
ytis
m(F
ig.2
)C
olon
ial
epip
hyt
ism
(bin
ary
codi
ng)
(Fig
s3,
4)C
lim
bin
gh
abit
(bin
ary
codi
ng)
(Fig
.5)
Met
axyl
emdi
amet
er(F
ig.5
)
Not
epip
hyt
eIn
divi
dual
epip
hyt
eC
olon
ial
epip
hyt
eN
oYe
sN
oYe
sS
mal
lL
arge
(Ca–
Ce)
10
01-
10
0.99
-0.8
90.
01-0
.11
0.99
-0.8
80.
01-0
.12
Ab
10
01-
10
1-0.
900-
0.10
1-0.
930-
0.07
(A.c
aud
atu
m–A
.flav
ofu
scu
m)
10
01-
10
0.01
-0.0
10.
99-0
.99
0.98
-0.7
90.
02-0
.21
(Tr–
HE
)1
00
1-1
01-
0.88
0-0.
121-
0.89
0-0.
11Tr
10
01-
10
1-0.
900-
0.10
1-0.
900-
0.10
(T.s
can
den
s–T.
poly
pod
ioid
es–
T.al
atu
m)
0.99
0.01
01-
10
1-0.
890-
0.11
1-0.
780-
0.22
D–L
10
01-
10
0.93
-0.8
10.
07-0
.19
0.99
-0.9
20.
01-0
.08
L1
00
1-1
00.
02-0
.03
0.98
-0.9
70.
92-0
.74
0.08
-0.2
6H
E0.
740
0.26
0.73
-0.8
80.
27-0
.12
0.99
-0.8
10.
01-0
.19
1-0.
820-
0.18
Po
0.01
00.
990.
01-0
.01
0.99
-0.9
91-
0.97
0-0.
031-
0.95
0-0.
05(C
r–V
a–D
i)0.
740
0.26
0.73
-0.8
80.
27-0
.12
0.98
-0.7
50.
02-0
.25
0.98
-0.7
70.
02-0
.23
Di
0.08
0.01
0.91
0.08
-0.1
10.
92-0
.89
0.99
-0.8
80.
01-0
.12
0.99
-0.8
70.
01-0
.13
(Cr–
Va)
0.80
00.
200.
79-0
.92
0.21
-0.0
80.
91-0
.62
0.09
-0.3
80.
91-0
.68
0.09
-0.3
2C
r0.
760.
010.
230.
76-0
.89
0.24
-0.1
10.
93-0
.72
0.07
-0.2
80.
94-0
.77
0.06
-0.2
3N
0.98
00.
020.
98-0
.99
0.02
-0.0
10.
84-0
.70
0.16
-0.3
0.85
-0.7
70.
15-0
.23
(Cr.
thys
anos
tom
um
–Cr.
inte
rmed
ium
)1
00
1-1
00.
99-0
.95
0.01
-0.0
50.
99-0
.95
0.01
-0.0
5C
0.06
00.
940.
06-0
.08
0.94
-0.9
20.
99-0
.90
0.01
-0.1
0.99
-0.8
90.
01-0
.11
Va
0.99
00.
010.
99-1
0.01
-00.
11-0
.15
0.89
-0.8
50.
11-0
.26
0.89
-0.7
4
EPIPHYTISM EVOLUTION IN TRICHOMANOID FERNS 593
© 2013 The Linnean Society of London, Botanical Journal of the Linnean Society, 2013, 173, 573–593