enzymatic acylation of a bifunctional molecule in 2-methyl-2-butanol: kinetic modelling
TRANSCRIPT

EK
EFa
b
a
ARRA
KEAOLBK
1
ltaafcsacatwt
0d
Enzyme and Microbial Technology 46 (2010) 338–346
Contents lists available at ScienceDirect
Enzyme and Microbial Technology
journa l homepage: www.e lsev ier .com/ locate /emt
nzymatic acylation of a bifunctional molecule in 2-methyl-2-butanol:inetic modelling
ric Hussona, Veronica Garcia-Matillaa, Catherine Humeaub, Isabelle Chevalota,rantz Fourniera, Ivan Marca,∗
LRGP - UPR 3349 CNRS, Nancy Université, 2 avenue de la forêt de Haye, 54500 Vandœuvre-lès-Nancy, FranceLIBio, Nancy Université, 2 avenue de la forêt de Haye, 54500 Vandœuvre-lès-Nancy, France
r t i c l e i n f o
rticle history:eceived 19 June 2009eceived in revised form 23 October 2009ccepted 18 November 2009
eywords:nzymatic acylationmino alcoholrganic solventipasei bi ping-pong mechanisminetic modelling
a b s t r a c t
The kinetics of enzymatic acylation of a bifunctional molecule exhibiting both an amino and a hydroxylfunction, considered as a peptide-like model molecule, is studied. The acylation of 6-amino-1-hexanolby ethyl oleate in 2-methyl-2-butanol catalysed by Novozym 435® has been investigated and mod-elled. This acylation process generated two products issued respectively from the O-acylation of the6-amino-1-hexanol, and from the N-acylation of the O-acyl product, which leads to the N,O-diacyl prod-uct. Experimental results confirmed the sequential reaction scheme. An experimental approach wasadopted to identify the impact of several operating conditions on the reaction performances, such asinitial substrates molar ratio and stirring rate. The kinetics of enzyme deactivation was also studied andincluded in the kinetic model. The establishment of the kinetic model was based on three major hypothe-ses: (i) a sequential bi bi ping-pong enzymatic mechanism, (ii) an interaction between acyl-acceptor andthe acyl-enzyme formation steps, (iii) the absence of influence of the residual hydrolysis of ethyl oleate on
the kinetics. Under this hypothesis, the kinetic parameters of the model were quantified using a genetic-evolutionary algorithm as an innovative identification method. The simplified model made it possible todescribe the kinetics of the 6-amino-1-hexanol acylation for some ranges of 6-amino-1-hexanol/ethyloleate molar ratios (2/1, 1/1, 1/2, 1/4) with a single kinetic parameter set. The case of a significant excessof 6-amino-1-hexanol (4/1) or ethyl oleate (1/8) constituted the limit of the kinetic model. However,these extreme molar conditions are not suitable to develop bioconversion processes involving bioactiveilable
peptides that are not ava. Introduction
The acylation process of peptides allows to enhance theipophilic properties of these molecules, and to improve bothheir transport across biological membranes and their stability,s described in several works concerning the acylation of insulinnd carnosine [1–5]. In a few cases, the covalent attachment withatty acid may maintain or improve the therapeutic or biologi-al activities of peptides [6,7]. The use of chemical processes toynthesize acylated peptides requires drastic reaction conditionsnd numerous functions protection and/or activation steps. Lipase-atalysed acylation in organic solvent constitutes a promising
lternative approach [8–12]. As peptides are generally polyfunc-ional molecules, their selective acylation constitutes a challengehich depends on both the chemo-selectivity and the regioselec-ivity of the reaction. Thus, understanding the enzyme efficiency
∗ Corresponding author.E-mail address: [email protected] (I. Marc).
141-0229/$ – see front matter © 2010 Published by Elsevier Inc.oi:10.1016/j.enzmictec.2009.11.008
in large quantities.© 2010 Published by Elsevier Inc.
and selectivity toward N- or O-acylation is necessary before extend-ing such conventional enzymatic processes to peptides acylation.Several studies investigated the enzymatic acylation of amino alco-hols such as ethanolamine, diethanolamine or 6-amino-1-hexanol[13–17]. Concerning the lipase-catalysed acylation of 6-amino-1-hexanol in organic solvent, the reaction led to the synthesis of twoproducts: the O-acyl product and the N,O-diacyl product. To explainthese observations, the authors suggested that the reaction could besequential. The two products might result from a first O-acylation ofthe 6-amino-1-hexanol and then from the N-acylation of the O-acylproduct, leading to the N,O-diacylated product [17]. Until now, toour knowledge, modelling of the enzymatic acylation of a bifunc-tional molecule has not yet been studied. The establishment of akinetic model would help to verify such hypothesis and to qualifythe most suitable operating conditions. In the literature, only mod-
els concerning the lipase-catalysed acylation of primary alcoholsin organic solvents have already been developed. Most are basedon the well-known bi bi ping-pong mechanism, characterised bythe formation of an acyl-enzyme intermediate, and were used todescribe O-acylation reactions catalysed by the lipase B of Candida
crobia
aelotfftet
rwithusepiietTsid
2
2
Si7D2f
2
veamal5oDdtvlcbs
et
2
SluFitm
E. Husson et al. / Enzyme and Mi
ntarctica (CAL B) in an organic solvent [18,19]. The kinetic mod-lling of O-acylation processes catalysed by other enzymes like theipases of Mucor miehei and Pseudomonas cepacia was also basedn this mechanism [20–22]. The bi bi-ordered mechanism, charac-erised by the formation of a tertiary complex, has been used in veryew studies concerning trans-esterification reactions in solvent-ree systems catalysed by the CAL B [23,24]. As the stability ofhe enzyme is in most cases a key issue in enzymatic processes,nzyme deactivation was taken into account, allowing to improvehe agreement between simulated and experimental data [25–27].
The aim of the present work is to investigate the trans-acylationeaction of a bifunctional model molecule, the 6-amino-1-hexanol,ith ethyl oleate in 2-methyl-2-butanol at 55 ◦C catalysed by the
mmobilized lipase B of C. antarctica. 6-amino-1-hexanol exhibitswo potential acylable sites: one primary amino group and oneydroxyl group. The presence of these two functional groups leds to consider this substrate as a peptide-like model. In the firsttep, an experimental approach is performed to study the influ-nce of substrates molar ratio and stirring rate conditions on theerformances of the reaction. The deactivation of the lipase dur-
ng the incubation time in 2-methyl-2-butanol at 55 ◦C was alsonvestigated. In the second step, a kinetic model based on thisxperimental approach was developed and the kinetic parame-ers were identified using a genetic-evolutionary algorithm [28,29].he most suitable model was proposed and validated for severalubstrates molar ratios by comparing model predictions to exper-mental results. In the final section, the limits of this model areiscussed.
. Materials and methods
.1. Chemicals and enzyme
6-Amino-1-hexanol (97%) and ethyl oleate (99%) were purchased fromigma–Aldrich (Steinheim, Germany). Novozym 435® (a lipase B from C. antarcticammobilized on an acrylic resin, E.C. 3.1.1.3) with propyl laurate synthesis activity of000 PLU g−1 and protein grade of [1–10%] came from Novo Nordisk A/S (Bagsraerd,enmark). In this study 7000 PLU g−1 corresponds to 1 UA (arbitrary unit). 2-Methyl--butanol, methanol, trifluoroacetic acid (TFA) with 99% of purity were acquiredrom Carlo Erba (Rodano, Spain).
.2. Acylation procedure
The enzymatic trans-acylation reactions of 6-amino-1-hexanol in organic sol-ent were carried out in a batch stirred reactor (Wheaton®) of 50 mL volumequipped with a condensation system to prevent solvent evaporation. In a typicalcylation reaction, 6-amino-1-hexanol and ethyl oleate was added to 10 mL of 2-ethyl-2-butanol previously dehydrated on 4 Å molecular sieves. The initial water
ctivity of the media was quantified with a thermoconstanter Novasina® (Switzer-and) and was inferior to 0.1. Reactions were performed at different stirring rates at5 ◦C during 96 h. After the total dissolution of the substrates over 12 h, the acylationf 6-amino-1-hexanol was started by the addition of 10 g L−1 of lipase preparation.uring the enzymatic reaction, samples of 50 �L were withdrawn, filtered theniluted 20× with methanol/TFA (100/0.1, v/v). The substrates and the products ofhe reaction were separated and quantified by HPLC (injection volume of 2 �L). Toalidate the repeatability of the experiments, each reaction was performed in trip-icate and results were expressed as mean values with standard deviations. Theonditions used for the purification and the identification of the acylated productsy mass spectrometry and nuclear magnetic resonance, were described in a previoustudy [17].
To confirm that no chemical reaction occurred between 6-amino-1-hexanol andthyl oleate, control experiments without enzyme were also carried out and showedhe absence of product whatever the applied experimental conditions.
.3. Analyses by HPLC
The time course of each reaction was monitored using HPLC (LC 10 AD – VP,himadzu, France) equipped with a UV detector at 214 nm and a light-scattering
ow temperature evaporative detector (Shimadzu, France) in this order. The col-mn was a newly developed C18 amide 125 × 2.1 mm – 5 �m (Altima® , Altech,rance) maintained at 25 ◦C. The mobile phase (0.2 mL min−1 flow rate) consistedn methanol/water/TFA (80/20/0.1, v/v/v). A constant elution gradient was appliedo reach methanol/TFA (100/0.1, v/v) after 5 min. This methanol concentration wasaintained for 18 min then gradually lowered to reach the initial methanol/water
l Technology 46 (2010) 338–346 339
ratio at the end of the run (i.e. 34 min). Calibrations were performed using standardsubstrates and purified products. The substrate conversion rate at the thermody-namic equilibrium was obtained applying the following equation:
r(%) =(
1 − [Substrate]equilibrium
[Substrate]initial
)× 100. (1)
Initial reaction rates were calculated by derivation of a second order polynomialapproximation of the concentration profile, built on the five first experimental data(Software: Matlab® , MathWorks, USA).
2.4. Study of the enzymatic stability in the course of the reaction
The biocatalyst was incubated in 2-methyl-2-butanol in Wheaton® reactor at55 ◦C for pre-defined durations (0, 2, 6, 10, 24 and 48 h). After this time, the substrates(6-amino-1-hexanol 0.12 M and ethyl oleate 0.24 M) previously solubilised wereadded to the reactor. The reaction was performed over 1 h. The initial consumptionrate of ethyl oleate was assessed for each incubation time.
To study the protective effect of 6-amino-1-hexanol on the enzymatic stability inthe 2-methyl-2-butanol, the biocatalyst was incubated in this solvent in Wheaton®
reactor at 55 ◦C for 10 h, in presence of 0.12 M of 6-amino-1-hexanol. After this time,ethyl oleate 0.24 M was added to the reactor and the reaction was performed over1 h. The initial rate of ethyl oleate consumption was then quantified and compared tothe initial rate obtained in the case of 10 h of incubation without 6-amino-1-hexanol(control).
The residual enzymatic activity (Ar) was expressed as the ratio of the initial rateof ethyl oleate disappearance after a period t of incubation (At) over the initial rateof ethyl oleate disappearance without incubation (A0):
Ar = At
A0(2)
2.5. Tool for model development
Following experimental investigations, several models based on classical mech-anisms were studied. Seven differential equations describing mass balances weresolved using the numerical integration tools present in the Matlab® software. Iden-tification of the kinetic parameters was performed on each of the 10 experimentaldata sets. The optimisation tool was an innovative genetic-evolutionary algorithmdeveloped in our laboratory. Its main features have been described in the literature[28,29]. The most suitable kinetic model is described in Section 3.
3. Results–discussion
The present work deals with the N,O-enzymatic trans-acylationof 6-amino-1-hexanol, a peptide-like model molecule. A previousstudy has shown that the Novozym 435®-catalysed acylation of 6-amino-1-hexanol (B) by an acyl donor (AR) in 2-methyl-2-butanolat 55 ◦C made it possible to obtain two acylated products. Thefirst one was O-acyl aminohexanol (AB) and seemed to be rapidlyconverted into N,O-diacyl aminohexanol (ABA) thus suggesting asequential reaction. The N,O-diacyl product was the major productat thermodynamic equilibrium [17]. Fig. 1 represents the assumedreaction scheme of this enzymatic acylation.
3.1. Preliminary tests
The first step consisted in evaluating the biocatalyst concen-tration which prevents enzyme concentration limitation in thecourse of the reaction. A convenient Novozym 435® concentra-tion of 10 g L−1 was defined to prevent such limitation (datanot shown). Under non-limiting biocatalyst concentration con-ditions, enzymatic trans-acylations of 6-amino-1-hexanol wereperformed with a quantitative monitoring of each substrate andproduct. The time profiles of the main reactants and productsconcentrations show that the kinetics of the reaction can bedivided into two parts (Fig. 2). The first part corresponds to afast reaction which takes place during the first hour. During thisfirst period, the two substrates were rapidly converted with ini-
tial apparent rates of consumption of 6-amino-1-hexanol andethyl oleate of 0.40 ± 0.02 M h−1, and 0.55 ± 0.02 M h−1 respectively(corresponding to 0.67 ± 0.03 �mol min−1 per mg of biocatalyst,and 0.92 ± 0.03 �mol min−1 mg−1 respectively). The synthesis ofboth O-oleyl and N,O-dioleyl products was observed. In this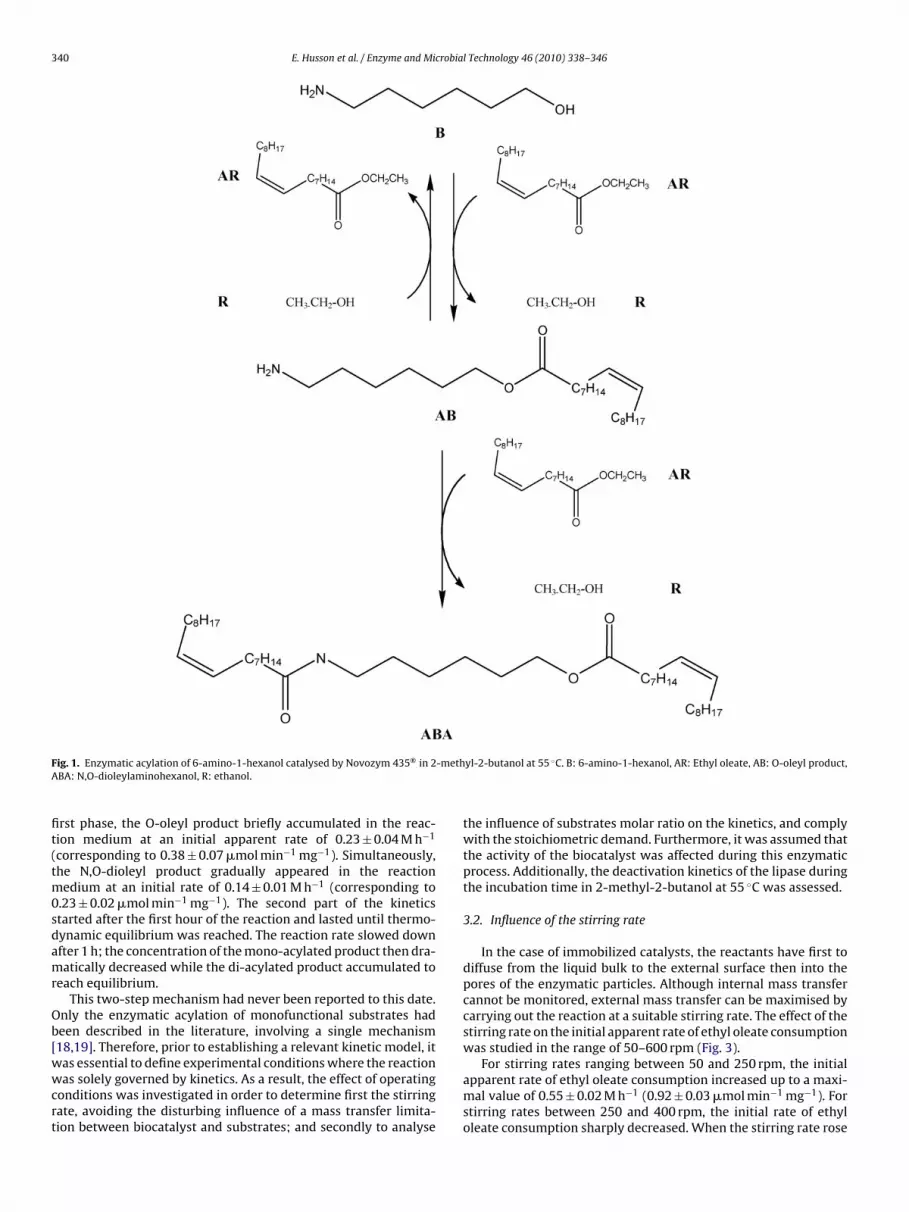
340 E. Husson et al. / Enzyme and Microbial Technology 46 (2010) 338–346
F -methA
fit(tm0sdamr
Ob[wwcrt
ig. 1. Enzymatic acylation of 6-amino-1-hexanol catalysed by Novozym 435® in 2BA: N,O-dioleylaminohexanol, R: ethanol.
rst phase, the O-oleyl product briefly accumulated in the reac-ion medium at an initial apparent rate of 0.23 ± 0.04 M h−1
corresponding to 0.38 ± 0.07 �mol min−1 mg−1). Simultaneously,he N,O-dioleyl product gradually appeared in the reaction
edium at an initial rate of 0.14 ± 0.01 M h−1 (corresponding to.23 ± 0.02 �mol min−1 mg−1). The second part of the kineticstarted after the first hour of the reaction and lasted until thermo-ynamic equilibrium was reached. The reaction rate slowed downfter 1 h; the concentration of the mono-acylated product then dra-atically decreased while the di-acylated product accumulated to
each equilibrium.This two-step mechanism had never been reported to this date.
nly the enzymatic acylation of monofunctional substrates hadeen described in the literature, involving a single mechanism18,19]. Therefore, prior to establishing a relevant kinetic model, it
as essential to define experimental conditions where the reactionas solely governed by kinetics. As a result, the effect of operatingonditions was investigated in order to determine first the stirringate, avoiding the disturbing influence of a mass transfer limita-ion between biocatalyst and substrates; and secondly to analyse
yl-2-butanol at 55 ◦C. B: 6-amino-1-hexanol, AR: Ethyl oleate, AB: O-oleyl product,
the influence of substrates molar ratio on the kinetics, and complywith the stoichiometric demand. Furthermore, it was assumed thatthe activity of the biocatalyst was affected during this enzymaticprocess. Additionally, the deactivation kinetics of the lipase duringthe incubation time in 2-methyl-2-butanol at 55 ◦C was assessed.
3.2. Influence of the stirring rate
In the case of immobilized catalysts, the reactants have first todiffuse from the liquid bulk to the external surface then into thepores of the enzymatic particles. Although internal mass transfercannot be monitored, external mass transfer can be maximised bycarrying out the reaction at a suitable stirring rate. The effect of thestirring rate on the initial apparent rate of ethyl oleate consumptionwas studied in the range of 50–600 rpm (Fig. 3).
For stirring rates ranging between 50 and 250 rpm, the initialapparent rate of ethyl oleate consumption increased up to a maxi-mal value of 0.55 ± 0.02 M h−1 (0.92 ± 0.03 �mol min−1 mg−1). Forstirring rates between 250 and 400 rpm, the initial rate of ethyloleate consumption sharply decreased. When the stirring rate rose

E. Husson et al. / Enzyme and Microbial Technology 46 (2010) 338–346 341
Fig. 2. Concentration profiles relative to the enzymatic acylation of the 6-amino-1-hexanol (0.12 M) with ethyl oleate (0.24 M) catalysed by Novozym 435® in 2-methyl-2-butanol at 55 ◦C and 250 rpm in a Wheaton® reactor. (�) Ethyl oleate, (�)6-amino-1-hexanol, (�) N,O-dioleyl aminohexanol and (©) O-oleyl aminohexanol.
Fig. 3. Influence of the stirring rate on the performances of the enzymatic acylationof the 6-amino-1-hexanol (0.12 M) with ethyl oleate (0.24 M) catalysed by Novozym435® (10 g L−1) in 2-methyl-2-butanol at 55 ◦C.
Fig. 4. Influence of the 6-amino-1-hexanol/ethyl oleate molar ratio on the substrateconversion during the enzymatic acylation reaction catalysed by Novozym 435®
(10 g L−1) in 2-methyl-2-butanol at 55 ◦C.
higher than 400 rpm, the initial apparent rate seemed to level offat a low value. For low stirring rates, the reaction medium was notcorrectly mixed so the efficient collisions between substrates andbiocatalyst particles could be prevented by mass transfer limitation.Increasing the stirring rate lowered this limitation, thus directlyaffecting the reaction rate. When the stirring rate was higher than250 rpm, the performance of the reaction appeared to be affectednegatively. It was observed that under these stirring conditions anddue to the centrifugal force, the biocatalyst particles were dissem-inated and stuck along the reactor wall.
According to these results, the best trade-off between transferlimitation and effective particles distribution in the reaction mediawas obtained at a stirring rate of 250 rpm. The kinetic model wasestablished using experimental data collected at a 250 rpm stirringrate.
3.3. Influence of the 6-amino-1-hexanol/ethyl oleate molar ratio
Prior to building the kinetic model, the influence of 6-amino-1-hexanol/ethyl oleate molar ratio on the reaction kinetics wasexperimentally studied for 10 distinct molar ratios. Fig. 4 rep-resents the variations of the substrate conversion yield duringacylation for five molar ratios: one equimolar ratio (0.12 M/0.12 M),two molar ratios with an excess of acyl donor (0.12 M/0.24 Mand 0.12 M/0.48 M) and two molar ratios with an excess of 6-amino-1-hexanol (0.24 M/0.12 M and 0.48 M/0.12 M). When ethyloleate was in excess, the 6-amino-1-hexanol conversion yieldwas close to 100% and the reaction appeared to be faster thanwhen the 6-amino-1-hexanol was in excess. This result can beexplained through the stoichiometry of the reaction. Actually, onemole of ethyl oleate is required to produce one mole of O-oleylaminohexanol; one additional mole of this acyl donor is necessaryto synthesize one mole of the N,O-diacylated. The total conver-sion of substrates (99 ± 1%) was obtained for a 1/2 molar ratiovalue, indicating that such stoichiometric conditions were the mostappropriate, while minimizing substrates quantities.
The experimental profiles showed that only O- and N,O-acylation occurred in the reaction medium so that the experimentaldata obtained with distinct substrates molar ratio seems to describethe same reaction set and can be used to identify and then to vali-date the kinetic model.
3.4. Stability of Novozym 435® in 2-methyl-2-butanol
Enzyme activity is the key parameter controlling the wholereaction process. It is necessary to ensure that it is fully charac-
342 E. Husson et al. / Enzyme and Microbia
FtN
tsmd4Ips62pitwadcwetct2eapauciit
d
E
Eadr
E
Ett
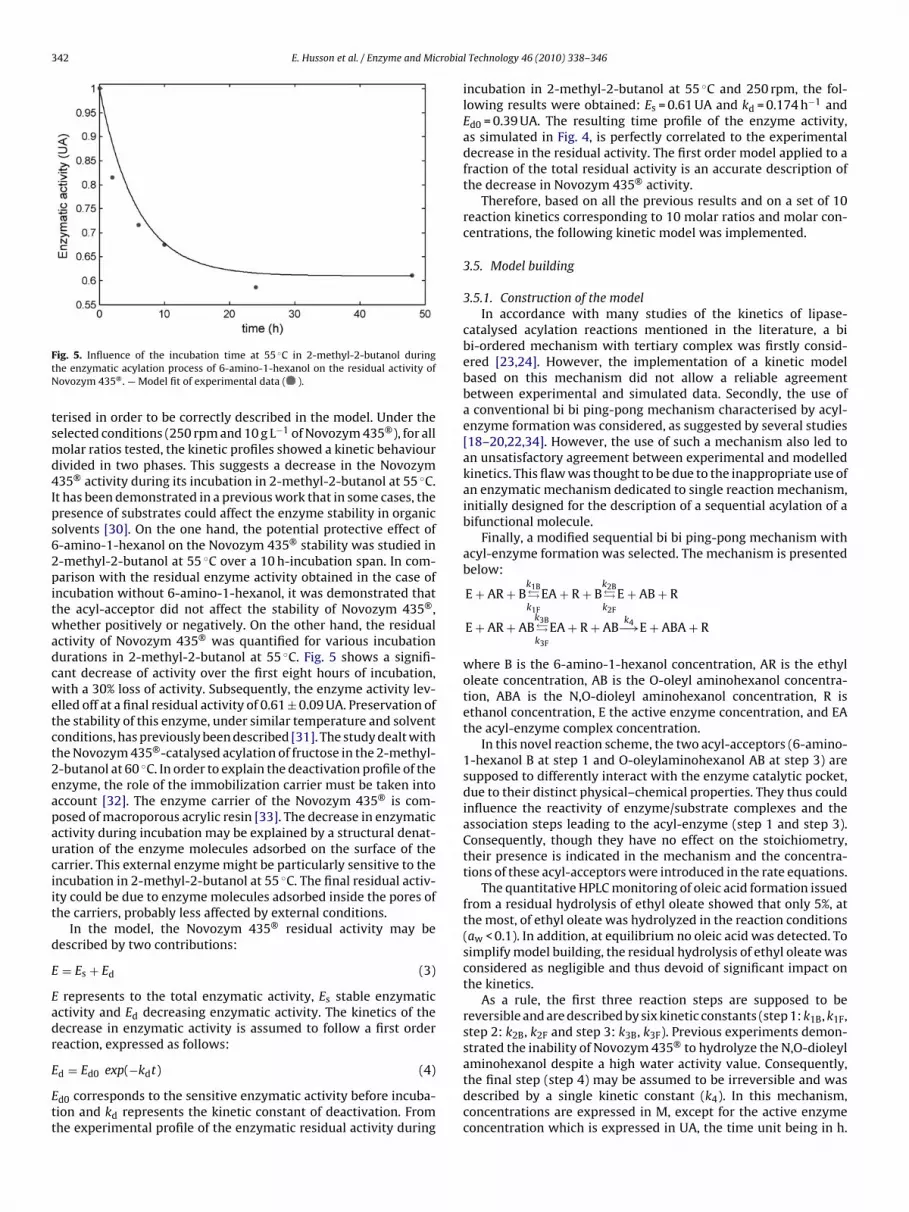
ig. 5. Influence of the incubation time at 55 ◦C in 2-methyl-2-butanol duringhe enzymatic acylation process of 6-amino-1-hexanol on the residual activity ofovozym 435® . — Model fit of experimental data ( ).
erised in order to be correctly described in the model. Under theelected conditions (250 rpm and 10 g L−1 of Novozym 435®), for allolar ratios tested, the kinetic profiles showed a kinetic behaviour
ivided in two phases. This suggests a decrease in the Novozym35® activity during its incubation in 2-methyl-2-butanol at 55 ◦C.t has been demonstrated in a previous work that in some cases, theresence of substrates could affect the enzyme stability in organicolvents [30]. On the one hand, the potential protective effect of-amino-1-hexanol on the Novozym 435® stability was studied in-methyl-2-butanol at 55 ◦C over a 10 h-incubation span. In com-arison with the residual enzyme activity obtained in the case of
ncubation without 6-amino-1-hexanol, it was demonstrated thathe acyl-acceptor did not affect the stability of Novozym 435®,hether positively or negatively. On the other hand, the residual
ctivity of Novozym 435® was quantified for various incubationurations in 2-methyl-2-butanol at 55 ◦C. Fig. 5 shows a signifi-ant decrease of activity over the first eight hours of incubation,ith a 30% loss of activity. Subsequently, the enzyme activity lev-
lled off at a final residual activity of 0.61 ± 0.09 UA. Preservation ofhe stability of this enzyme, under similar temperature and solventonditions, has previously been described [31]. The study dealt withhe Novozym 435®-catalysed acylation of fructose in the 2-methyl--butanol at 60 ◦C. In order to explain the deactivation profile of thenzyme, the role of the immobilization carrier must be taken intoccount [32]. The enzyme carrier of the Novozym 435® is com-osed of macroporous acrylic resin [33]. The decrease in enzymaticctivity during incubation may be explained by a structural denat-ration of the enzyme molecules adsorbed on the surface of thearrier. This external enzyme might be particularly sensitive to thencubation in 2-methyl-2-butanol at 55 ◦C. The final residual activ-ty could be due to enzyme molecules adsorbed inside the pores ofhe carriers, probably less affected by external conditions.
In the model, the Novozym 435® residual activity may beescribed by two contributions:
= Es + Ed (3)
represents to the total enzymatic activity, Es stable enzymaticctivity and Ed decreasing enzymatic activity. The kinetics of theecrease in enzymatic activity is assumed to follow a first ordereaction, expressed as follows:
d = Ed0 exp(−kdt) (4)
d0 corresponds to the sensitive enzymatic activity before incuba-ion and kd represents the kinetic constant of deactivation. Fromhe experimental profile of the enzymatic residual activity during
l Technology 46 (2010) 338–346
incubation in 2-methyl-2-butanol at 55 ◦C and 250 rpm, the fol-lowing results were obtained: Es = 0.61 UA and kd = 0.174 h−1 andEd0 = 0.39 UA. The resulting time profile of the enzyme activity,as simulated in Fig. 4, is perfectly correlated to the experimentaldecrease in the residual activity. The first order model applied to afraction of the total residual activity is an accurate description ofthe decrease in Novozym 435® activity.
Therefore, based on all the previous results and on a set of 10reaction kinetics corresponding to 10 molar ratios and molar con-centrations, the following kinetic model was implemented.
3.5. Model building
3.5.1. Construction of the modelIn accordance with many studies of the kinetics of lipase-
catalysed acylation reactions mentioned in the literature, a bibi-ordered mechanism with tertiary complex was firstly consid-ered [23,24]. However, the implementation of a kinetic modelbased on this mechanism did not allow a reliable agreementbetween experimental and simulated data. Secondly, the use ofa conventional bi bi ping-pong mechanism characterised by acyl-enzyme formation was considered, as suggested by several studies[18–20,22,34]. However, the use of such a mechanism also led toan unsatisfactory agreement between experimental and modelledkinetics. This flaw was thought to be due to the inappropriate use ofan enzymatic mechanism dedicated to single reaction mechanism,initially designed for the description of a sequential acylation of abifunctional molecule.
Finally, a modified sequential bi bi ping-pong mechanism withacyl-enzyme formation was selected. The mechanism is presentedbelow:
E + AR + Bk1B�k1F
EA + R + Bk2B�k2F
E + AB + R
E + AR + ABk3B�k3F
EA + R + ABk4−→E + ABA + R
where B is the 6-amino-1-hexanol concentration, AR is the ethyloleate concentration, AB is the O-oleyl aminohexanol concentra-tion, ABA is the N,O-dioleyl aminohexanol concentration, R isethanol concentration, E the active enzyme concentration, and EAthe acyl-enzyme complex concentration.
In this novel reaction scheme, the two acyl-acceptors (6-amino-1-hexanol B at step 1 and O-oleylaminohexanol AB at step 3) aresupposed to differently interact with the enzyme catalytic pocket,due to their distinct physical–chemical properties. They thus couldinfluence the reactivity of enzyme/substrate complexes and theassociation steps leading to the acyl-enzyme (step 1 and step 3).Consequently, though they have no effect on the stoichiometry,their presence is indicated in the mechanism and the concentra-tions of these acyl-acceptors were introduced in the rate equations.
The quantitative HPLC monitoring of oleic acid formation issuedfrom a residual hydrolysis of ethyl oleate showed that only 5%, atthe most, of ethyl oleate was hydrolyzed in the reaction conditions(aw < 0.1). In addition, at equilibrium no oleic acid was detected. Tosimplify model building, the residual hydrolysis of ethyl oleate wasconsidered as negligible and thus devoid of significant impact onthe kinetics.
As a rule, the first three reaction steps are supposed to bereversible and are described by six kinetic constants (step 1: k1B, k1F,step 2: k2B, k2F and step 3: k3B, k3F). Previous experiments demon-strated the inability of Novozym 435® to hydrolyze the N,O-dioleyl
aminohexanol despite a high water activity value. Consequently,the final step (step 4) may be assumed to be irreversible and wasdescribed by a single kinetic constant (k4). In this mechanism,concentrations are expressed in M, except for the active enzymeconcentration which is expressed in UA, the time unit being in h.
crobia
Fk
As
3
artsbstomTfite
fpl
J
TKt00
E. Husson et al. / Enzyme and Mi
or each reaction, on the assumption of a first order model, theinetic rates are written as follows:
r1F = k1F(E AR B) r2F = k2F(EA R B) r3F = k3F(E AB AR)r1B = k1B(EA R B) r2B = k2B(E AB R) r3B = k3B(EA R AB)r4 = k4(EA R AB)
(5)
mass balance for each compound yielded the differential equationet describing the acylation model:
dBdt
= −r2F + r2B
dARdt
= −r1F + r1B − r3F + r3B
dRdt
= +r1F − r1B + r3F − r3B
dABdt
= +r2F − r2B − r4
dABAdt
= +r4
dEAdt
= +r1F − r1B − r2F + r2B + r3F − r3B − r4
dEd
dt= −kdEd E = Es + Ed
(6)
.5.2. Identification of the kinetic constantsEq. (5) indicate that the kinetic constants (k1B, k1F) and (k3B, k3F)
re the parameters controlling the association and the dissociationates of the acyl-enzyme, for the O-acyl product synthesis and forhe N,O-dioleyl product synthesis respectively. k2B and k2F repre-ent the kinetic constants describing the equilibrium of the esterond synthesis/hydrolysis rate. k4 corresponds to the kinetic con-tant controlling the amide bond establishment rate which yieldshe N,O-dioleyl product. To identify the kinetic constants, databtained from 10 experiments achieved with different substrateolar ratios and concentration levels were first individually used.
he experimental conditions were classified into three categories:rst reactions with an excess of acyl donor (five experiments), thenhose with an excess of acyl-acceptor (two experiments) and finallyquimolar reactions (three experiments).
In order to identify the seven kinetic parameters, an objectiveunction describing the error between experimental concentrationrofiles and modelled concentration profiles was defined as fol-
ows:
= wB
N∑i=1
(Bexp(ti) − Bmod(ti))2 + wAR
N∑i=1
(ARexp(ti) − ARmod(ti))2
N∑
+ wABi=1
(ABexp(ti) − ABmod(ti))2
+ wABA
N∑i=1
(ABAexp(ti) − ABAmod(ti))2
(7)
able 1inetic constants for enzymatic acylation of 6-amino-1-hexanol performed in 2-methy
aining the kinetic parameters determined for nine acyl-acceptor/acyl donor molar rat.06 M/0.12 M; 0.06 M/0.24 M; 0.06 M/0.48 M; 0.24 M/0.12 M). Second row: reference par.48 M/0.12 M molar ratio.
Experiments Kinetic constants (M−1 UA−1 h−1)
k1F (×104) k1B (×101)
All experiments (9) [1.11; 8.56] [2.22; 5.17]0.12 M/0.24 M (6-amino-1-hexanol/ethyl oleate) 5.74 4.870.48 M/0.12 M (6-amino-1-hexanol/ethyl oleate) 1.20 5.59
l Technology 46 (2010) 338–346 343
where N is the number of experimental measurements for eachspecies, ti, i = 1, . . ., N, the corresponding time of measurement andwj, the relative weight for each species error. The most suitable val-ues for identification turned out to be wj = 0.25 ∀j∈{B, AR, AB, ABA}.A genetic-evolutionary algorithm was used for the identificationof the kinetic parameters, i.e. to minimize the model-experimenterror criteria (7). This method facilitated the identification process,as it made it possible to easily define boundaries of the search spacefor each kinetic parameter. It is suitable to solve such problems asit is capable of scanning an extended search space (the solution,i.e. the parameter vector, belongs to �7) in order to locate globaloptimum. It provided the optimal kinetic parameter vector leadingto the minimal experiment-model error. This optimisation methodalso supplied one set of kinetic parameter vectors which can beconsidered as equivalent regarding the same error criterion (7). Theanalysis of the resulting solution set delivered important informa-tion regarding error sensitivity, thus providing a kind of numericalconfidence interval. From the identification conducted on each ofthe 10 experimental data, 10 parameter sets were determined (datanot shown). Nine experiments led to parameters in the same orderof magnitude. One experiment (0.06 M 6-amino-1-hexanol/0.48 Methyl oleate) led to an outlier vector. This result is illustrated inTable 1 which presents the largest numerical interval containingthe nine parameter sets. The outlier parameter set is described inthe last row.
The optimal parameter vector, identified for a 6-amino-1-hexanol/ethyl oleate ratio (0.12 M/0.24 M), is defined in Table 1.For both synthesis steps, the acyl-enzyme association appears to bemuch more difficult to achieve than the acyl-enzyme dissociation.The order of magnitude is about 1000 times lower for the forwardkinetic constant (k1F and k3F) than for the backward one (k1B andk3B). As far as acyl-enzyme association and mono-acylation reactionare concerned, it seemed that the monoacylation was controlled bythe acylation reaction (k2F). As far as acyl-enzyme association andN-O-acylation reaction are concerned, the N-O-acylation reaction(k4) seemed to monitor the reaction.
The optimisation algorithm provides a set of 1000 parametervectors which can be considered as equivalent or acceptable withrespect to the model-experiment error (7). The distribution of thecorresponding kinetic parameter values is described in Fig. 6. It ispresented as a matrix scatter plot. For each kinetic parameter, theplot on the diagonal shows the qualitative distribution of its accept-able values. The rest of the figure matrix shows the two-by-twocorrelations. For example, the figure in the last row and 6th columndepicts the values of the k4 (last) and k3B (6th) parameters in theacceptable vectors. It thus describes the correlation between k4 andk3B as a numerical (but not statistical) confidence ellipsoid. Sucha numerical confidence interval showed that the identification ledon the one hand to uncorrelated (spherical) distribution around the
point (k4, k3B) = (0.048, 0.05) M−1 UA−1 h−1) and also to quite accu-rate parameter values (k4 ranged from 0.044 to 0.055 M−1 UA−1 h−1and k3B ranged from 0.035 to 0.065 M−1 UA−1 h−1). In the light ofthe narrow distributions observed for all parameters in Fig. 6, theparameter identification was considered as satisfactory and the
l-2-butanol and catalysed by Novozym 435® . First row: numerical intervals con-ios (0.12 M/0.12 M; 0.24 M/0.24 M; 0.48 M/0.48 M; 0.12 M/0.24 M; 0.12 M/0.48 M;ameter set used for simulations. Last row: parameter set identified for the extreme
k2F (×101) k2B (×104) k3F (×105) k3B (×102) k4 (×102)
[1.72; 9.76] [1.38; 9.77] [3.24; 29.3] [1.08; 9.71] [1.27; 12.3]4.26 4.61 4.57 1.54 7.170.85 2.86 7.31 1.46 27.8

344 E. Husson et al. / Enzyme and Microbial Technology 46 (2010) 338–346
F ino-1-hexanol performed with a molar ratio 0.12 M (6-amino-1-hexanol)/0.24 M (ethylo h−1.
ktoo
rt0
(
t(r0tittes
s(tirakvalk
3
we
ig. 6. Identification of kinetic constants for the enzymatic trans-acylation of 6-amleate) using a genetic-evolutionary algorithm. Values were expressed in UA−1 M−1
inetic constants were considered as reliable. Fig. 7(a) shows howhe resulting model accurately predicts the concentration profilef all species for a molar ratio 6-amino-1-hexanol (0.12 M)/ethylleate (0.24 M).
In order to check the relevance of this model, the maximalates of ethyl oleate consumption deduced from both experimen-al data and simulation, were compared for the reference reaction.12 M/0.24 M molar ratio.
In the model, the initial rate of ethyl oleate removal is given by8):
dARdt
= −r1F + r1B − r3F + r3B (8)
The maximal experimental rate of ethyl oleate consump-ion for a biocatalyst concentration of 10 g L−1 was 0.55 M h−1
0.92 �mol min−1 mg−1). The calculated reaction rates during theeaction showed that the rate values reached a maximum of.58 M h−1 (0.97 ± 0.03 �mol min−1 mg−1) which was very close tohe experimental value, with a deviation of 5%. Thus the reliabil-ty of the model was clearly established as well as its suitabilityo (i) describe the sequential kinetics of the enzymatic acyla-ion of 6-amino-1-hexanol in organic solvent, under the definedxperimental conditions and (ii) accurately predict the influence ofubstrates concentration on the reaction performances.
From the identification carried out for experiments (at sub-trates ratios 2/1, 1/1, 1/2, 1/4 obtained at different concentrations)Table 1), the kinetic parameters turned out to be essentially con-ained in the same join confidence interval. The parameter set,nitially identified from the reaction with a 0.12 M/0.24 M molaratio between the amino alcohol and ethyl oleate, was chosen asreference set and was used for the simulation of our enzymatic
inetics. The reference parameter set is described in Table 1. Thisector was assumed to reliably describe the kinetics with a goodgreement regarding all the various molar ratios and concentrationevels tested. It constitutes an important and original feature of thisinetic model, as it allows a very wide range of applications.
.5.3. Validation of the modelThe simulation of other reaction conditions was carried out
ith the reference parameter set (Table 1). Fig. 7(b) represents thexperimental and predicted concentration profiles for the reaction
Fig. 7. Concentration profiles of substrates and products relative to the enzy-matic acylation of the 6-amino-1-hexanol with ethyl oleate catalysed by Novozym435® in 2-methyl-2-butanol at 55 ◦C and 250 rpm. The symbols correspond to theexperimental data and the lines correspond to the modelled curves. 6-Amino-1-hexanol/ethyl oleate molar ratios were 0.12 M/0.24 M (a) and 0.06 M/0.12 M (b).
E. Husson et al. / Enzyme and Microbial Technology 46 (2010) 338–346 345
Fig. 8. Concentration profiles of substrates and products relative to the enzymaticacylation of the 6-amino-1-hexanol with ethyl oleate catalysed by Novozym 435®
itm
wIr0
is
mtodFf(betomtrh
Fig. 9. Concentration profiles of substrates and products relative to the enzymatic®
n 2-methyl-2-butanol at 55 ◦C and 250 rpm. Symbols correspond to the experimen-al data and lines correspond to modelled curves. 6-Amino-1-hexanol/ethyl oleate
olar ratio was 0.06 M/0.48 M.
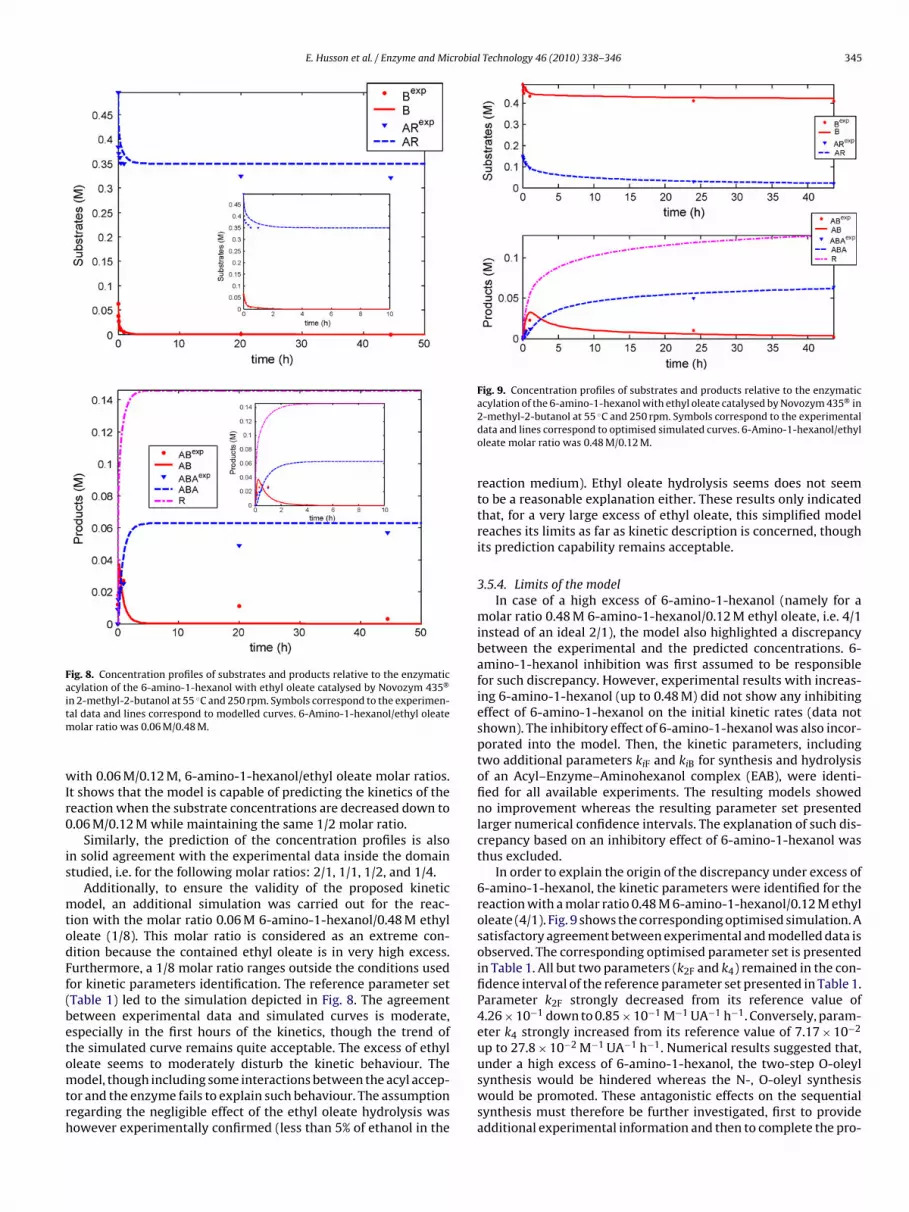
ith 0.06 M/0.12 M, 6-amino-1-hexanol/ethyl oleate molar ratios.t shows that the model is capable of predicting the kinetics of theeaction when the substrate concentrations are decreased down to.06 M/0.12 M while maintaining the same 1/2 molar ratio.
Similarly, the prediction of the concentration profiles is alson solid agreement with the experimental data inside the domaintudied, i.e. for the following molar ratios: 2/1, 1/1, 1/2, and 1/4.
Additionally, to ensure the validity of the proposed kineticodel, an additional simulation was carried out for the reac-
ion with the molar ratio 0.06 M 6-amino-1-hexanol/0.48 M ethylleate (1/8). This molar ratio is considered as an extreme con-ition because the contained ethyl oleate is in very high excess.urthermore, a 1/8 molar ratio ranges outside the conditions usedor kinetic parameters identification. The reference parameter setTable 1) led to the simulation depicted in Fig. 8. The agreementetween experimental data and simulated curves is moderate,specially in the first hours of the kinetics, though the trend ofhe simulated curve remains quite acceptable. The excess of ethylleate seems to moderately disturb the kinetic behaviour. The
odel, though including some interactions between the acyl accep-or and the enzyme fails to explain such behaviour. The assumptionegarding the negligible effect of the ethyl oleate hydrolysis wasowever experimentally confirmed (less than 5% of ethanol in the
acylation of the 6-amino-1-hexanol with ethyl oleate catalysed by Novozym 435 in2-methyl-2-butanol at 55 ◦C and 250 rpm. Symbols correspond to the experimentaldata and lines correspond to optimised simulated curves. 6-Amino-1-hexanol/ethyloleate molar ratio was 0.48 M/0.12 M.
reaction medium). Ethyl oleate hydrolysis seems does not seemto be a reasonable explanation either. These results only indicatedthat, for a very large excess of ethyl oleate, this simplified modelreaches its limits as far as kinetic description is concerned, thoughits prediction capability remains acceptable.
3.5.4. Limits of the modelIn case of a high excess of 6-amino-1-hexanol (namely for a
molar ratio 0.48 M 6-amino-1-hexanol/0.12 M ethyl oleate, i.e. 4/1instead of an ideal 2/1), the model also highlighted a discrepancybetween the experimental and the predicted concentrations. 6-amino-1-hexanol inhibition was first assumed to be responsiblefor such discrepancy. However, experimental results with increas-ing 6-amino-1-hexanol (up to 0.48 M) did not show any inhibitingeffect of 6-amino-1-hexanol on the initial kinetic rates (data notshown). The inhibitory effect of 6-amino-1-hexanol was also incor-porated into the model. Then, the kinetic parameters, includingtwo additional parameters kiF and kiB for synthesis and hydrolysisof an Acyl–Enzyme–Aminohexanol complex (EAB), were identi-fied for all available experiments. The resulting models showedno improvement whereas the resulting parameter set presentedlarger numerical confidence intervals. The explanation of such dis-crepancy based on an inhibitory effect of 6-amino-1-hexanol wasthus excluded.
In order to explain the origin of the discrepancy under excess of6-amino-1-hexanol, the kinetic parameters were identified for thereaction with a molar ratio 0.48 M 6-amino-1-hexanol/0.12 M ethyloleate (4/1). Fig. 9 shows the corresponding optimised simulation. Asatisfactory agreement between experimental and modelled data isobserved. The corresponding optimised parameter set is presentedin Table 1. All but two parameters (k2F and k4) remained in the con-fidence interval of the reference parameter set presented in Table 1.Parameter k2F strongly decreased from its reference value of4.26 × 10−1 down to 0.85 × 10−1 M−1 UA−1 h−1. Conversely, param-eter k4 strongly increased from its reference value of 7.17 × 10−2
up to 27.8 × 10−2 M−1 UA−1 h−1. Numerical results suggested that,under a high excess of 6-amino-1-hexanol, the two-step O-oleyl
synthesis would be hindered whereas the N-, O-oleyl synthesiswould be promoted. These antagonistic effects on the sequentialsynthesis must therefore be further investigated, first to provideadditional experimental information and then to complete the pro-
3 crobia
pi
dan
4
melomlwkald1hbhathTca
qt
tmt
etosmpp
A
R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[33] Arroyo M, Sanchez-Montero JM, Sinisterra JV. Thermal stabilization of immo-
46 E. Husson et al. / Enzyme and Mi
osed model. These results open an interesting track for furthernvestigations.
However, such extreme molar conditions are not suitable toevelop bioconversion processes involving bioactive peptides thatre not available in large quantities. This limitation does thereforeot reduce the quality of the proposed model.
. Conclusion
In the present study, an accurate and efficient original kineticodel has been developed to describe the sequential N,O-
nzymatic acylation of a peptide-like molecule, in organic solvent,eading to the synthesis of two products. The kinetic model is basedn an experimental approach involving a bi bi ping-pong enzy-atic mechanism in two steps, and integrates enzyme activity
oss. The effect of the acyl-acceptor on the acyl-enzyme formationas added to the mechanism as a relevant impacting factor. The
inetic parameters were identified using a genetic-evolutionarylgorithm. A single parameter set was proposed and allowed simu-ations which stood in good agreement with experimental data foristinct initial molar ratios of substrates concentrations (2/1, 1/1,/2, 1/4). Such good agreement thus consolidates the validity of theypotheses used in the model: (i) sequential reaction, (ii) enzymatici bi ping-pong mechanism, (iii) absence of impact for residualydrolysis of ethyl oleate on kinetics, and (iv) significant impact ofcyl-acceptor concentration on the acyl-enzyme formation. Addi-ionally, the maximal initial rates of reaction predicted by the modelave been compared with those estimated from experimental data.he prediction of maximal initial rates was in accordance with theorresponding observation. This confirmed the coherence, accuracynd reliability of the proposed kinetic model.
An enzyme denaturation phenomenon was also highlighted anduantified. It would be interesting to precisely understand the fac-ors affecting the stability of the biocatalyst under such conditions.
Additionally, the effect of the acyl-acceptors concentration onhe kinetics of the acyl-enzyme formation had to be included in the
odel. Similarly, further investigations are needed to understandhe type of interaction involved in such complex systems.
Though several limits to the model are clearly identified forxtreme initial molar ratios, the latter allows satisfactory predic-ion of enzymatic acylation of 6-amino-1-hexanol for a wide rangef regular initial ratios. This investigation thus constituted the firsttep of a promising breakthrough in the development of a kineticodel of enzymatic acylation of polyfunctional molecules such as
eptides and should allow a better control and optimisation ofeptide acylation processes.
cknowledgement
Authors are grateful to Ms. M. Faure for the proofreading.
eferences
[1] Asada H, Douen T, Mizokoshi Y, Fujita T, Murakami M, Yamamoto A, et al.Stability of acyl derivatives of insulin in the small intestine: relative impor-tance of insulin association characteristics in aqueous solution. Pharm Res1994;11(8):1115–20.
[2] Pegova A, Abe H, Boldyrev A. Hydrolysis of carnosine and related compoundsby mammalian carnosinases. Comp Biochem Physiol 2000;127(4):443–6.
[3] Veuillez F, Deshusses J, Buri P. Synthesis and characterization of an acylated di-peptide (Myr-Trp-Leu) with modified transmucosal transport properties. Eur J
Pharm Biopharm 1999;48(1):21–6.[4] Zweytick D, Pabst G, Abuja PM, Jilek A, Blondelle SE, Andra J, et al. Influenceof N-acylation of a peptide derived from human lactoferricin on membraneselectivity. Biochim Biophys Acta 2006;1758(9):1426–35.
[5] Yelamos B, Roncal F, Albar JP, Rodriguez-Crespo I, Gavilanes F. Influence of acy-lation of a peptide corresponding to the amino-terminal region of endothelial
[
l Technology 46 (2010) 338–346
nitric oxide synthase on the interaction with model membranes. Biochemistry2006;45(4):1263–70.
[6] Jagadeeshwar Rao R, Tiwari AK, Kumar US, Reddy SV, Ali AZ, Rao JM. Novel 3-O-acyl mesquitol analogues as free-radical scavengers and enzyme inhibitors:synthesis, biological evaluation and structure-activity relationship. Bioorg MedChem Lett 2003;13(16):2777–80.
[7] Babizhayev MA. Biological activities of the natural imidazole-containing pep-tidomimetics n-acetylcarnosine, carcinine and l-carnosine in ophthalmic andskin care products. Life Sci 2006;78(20):2343–57.
[8] Klibanov A. Enzymes that work in organic solvents. Chemtech1986;16(6):354–9.
[9] Klibanov AM. Improving enzymes by using them in organic solvents. Nature2001;409(6817):241–6.
10] Zaks A. New enzymatic properties in organic media. In: Enzymatic reactionsin organic media. London: Blackie Academic & Professional (An imprint ofChapman and Hall); 1996. p. 70–93.
11] Zaks A, Klibanov AM. Enzyme-catalyzed processes in organic solvents. Proc NatlAcad Sci U S A 1985;82(10):3192–6.
12] Husson E, Humeau C, Paris C, Vanderesse R, Framboisier X, Marc I, et al. Enzy-matic acylation of polar dipeptides: influence of reaction media and molecularenvironment of functional groups. Proc Biochem 2009;44:428–34.
13] Fernandez-Perez M, Otero C. Selective enzymatic synthesis of amide surfac-tants from diethanolamine. Enzyme Microb Technol 2003;33(5):650–60.
14] Fernandez-Perez M, Otero C. Enzymatic synthesis of amide surfactants fromethanolamine. Enzyme Microb Technol 2001;28(6):527–36.
15] Furutani T, Furui M, Ooshima H, Kato J. Acylation of [beta]-amino alcohol by acylmigration following enzyme-catalyzed esterification. Enzyme Microb Technol1996;19(8):578–84.
16] Furutani T, Ooshima H, Kato J. Preparation of N-, O-diacyl ethanolaminefrom N-acyl ethanolamine using lipase preparations. Enzyme Microb Technol1997;20(3):214–20.
17] Husson E, Humeau C, Blanchard F, Framboisier X, Marc I, Chevalot I. Chemo-selectivity of the N,O-enzymatic acylation in organic media and in ionic liquids.J Mol Catal B: Enzym 2008;55:110–7.
18] Romero MD, Calvo L, Alba C, Daneshfar A. A kinetic study of isoamyl acetate syn-thesis by immobilized lipase-catalyzed acetylation in n-hexane. J Biotechnol2007;127(2):269–77.
19] Yadav GD, Devi KM. Immobilized lipase-catalysed esterification and transester-ification reactions in non-aqueous media for the synthesis of tetrahydrofurfurylbutyrate: comparison and kinetic modeling. Chem Eng Sci 2004;59(2):373–83.
20] Chulalaksananukul W, Condoret JS, Delorme P, Willemot RM. Kineticstudy of esterification by immobilized lipase in n-hexane. FEBS Lett1990;276(1–2):181–4.
21] Jose A, Arcos CGHJCO. Kinetics of the lipase-catalyzed synthesis of glucoseesters in acetone. Biotechnol Bioeng 2001:104–10.
22] Dabkowska K, Szewczyk KW. Kinetics of enantioselective enzymatic acylationof mandelic acid. J Biotechnol 2007;131(2, Suppl. 1):S79–80.
23] Yadav GD, Trivedi AH. Kinetic modeling of immobilized-lipase catalyzed trans-esterification of n-octanol with vinyl acetate in non-aqueous media. EnzymeMicrob Technol 2003;32(7):783–9.
24] Garcia MJ, Rebolledo F, Gotor V. Chemoenzymatic aminolysis and ammonolysisof [beta]-ketoesters. Tetrahedron Lett 1993;34(38):6141–2.
25] Torres CF, Tore AM, Fornari T, Senorans FJ, Reglero G. Ethanolysis of a wastematerial from olive oil distillation catalyzed by three different commerciallipases: a kinetic study. J Biochem Eng 2007;34(2):165–71.
26] Truppo MD, Pollard DJ, Moore JC, Devine PN. Production of (S)-[gamma]-fluoroleucine ethyl ester by enzyme mediated dynamic kinetic resolution:comparison of batch and fed batch stirred tank processes to a packed bedcolumn reactor. Chem Eng Sci 2008;63(1):122–30.
27] Segel IH. Enzyme kinetics: behavior and analysis of rapid equilibrium andsteady-state enzyme systems. New York: John Wiley & Sons; 1975.
28] Muniglia L, Nandor Kiss L, Fonteix C, Marc I. Multicriteria optimization of asingle-cell oil production. Eur J Operation Res 2004;153(2):360–9.
29] Perrin E, Mandrille A, Oumoun M, Fonteix C, Marc I. Optimisation globale parstratégie d’evolution: technique utilisant la génétique des individus diploıdes.Operation Res 1997;31(2):161–201.
30] Pirozzi D, Greco G. Activity and stability of lipases in the synthesis of butyllactate. Enzyme Microb Technol 2004;34(2):94–100.
31] Coulon D, Ismail A, Girardin M, Rovel B, Ghoul M. Effect of different biochem-ical parameters on the enzymatic synthesis of fructose oleate. J Biotechnol1996;51(2):115–21.
32] Fernandez-Lorente G, Cabrera Z, Godoy C, Fernandez-Lafuente R, PalomoJM, Guisan JM. Interfacially activated lipases against hydrophobic supports:effect of the support nature on the biocatalytic properties. Proc Biochem2008;43(10):1061–7.
bilized lipase B from Candida antarctica on different supports: effect of wateractivity on enzymatic activity in organic media. Enzyme Microb Technol1999;24(1–2):3–12.
34] Arcos JA, Hill CG, Otero Jr C. Kinetics of the lipase-catalyzed synthesis of glucoseesters in acetone. Biotechnol Bioeng 2001;73(2):104–10.