entomopathogenic nematodes - a review - aws
TRANSCRIPT

AGRICULTURAL RESEARCH COMMUNICATION CENTRE
www.arccjournals.com / indianjournals.comAgri. Reviews, 34 (3) : 163-175, 2013DOI- 10.5958/j.0976-0741.34.3.001
ENTOMOPATHOGENIC NEMATODES - A REVIEW
Sumit Vashisth* , Y.S. Chandel and P. K. Sharma
Department of Entomology,Himachal Pradesh Krishi Vishvavidyalaya, Palampur, 176 062, India
Received: 20-02-2012 Accepted: 19-10-2012
ABSTRACTThe entomopathogenic nematodes possessing balanced biological control attributes belong to
genera Steinernema and Heterorhabditis and are having mutualistic association with bacteria of thegenus Xenorhabdus for Steinernematidae and Photorhabdus for H eterorhabditidae.Entomopathogenic nematodes being highly lethal to many important insect-pests, are safe to non-target organisms and working with their symbiotic bacteria kill the insects within 24-28 hours ascompared to days and weeks required for insect killing in other biological control agents. Theirinfective juveniles (IJs) have been reported to tolerate short-term exposure to many chemical andbiological insecticides, fungicides, herbicides, fertilizers and growth regulators, hence providing anopportunity of tank-mixing and application together. Entomopathogenic nematodes are also reportedto be compatible with a number of agrochemicals and their use can offer a cost-effective alternativeto pest control. Only twelve species out of nearly eighty identified Steinernematids and Heterorhabditisnematode species have been commercialized. The EPNs have the great potential to be used inintegrated pest management systems and work done have been reviewed in this article to facilitatethe students and researchers to have an overview of the work done and proceed further to undertakethe advanced research in different aspects related to entomopathogenic nematodes.
Keywords: Entomopathogenic nematodes (EPN), Heterorhabditis, Photorhabdus,Steinernema, Xenorhabdus.
*Corresponding author’s e-mail: [email protected]
Nematodes are simple roundworms,colorless, unsegmented, and lacking appendages,may be free-living, predaceous, or parasitic. Manyof the parasitic species cause important diseases ofhumans plants and animals. The term ‘Entomophilicnematodes’ includes all relationships of insects andnematodes ranging from phoresis to parasitism andpathogenesis. ‘Entomogenous nematodes’ are thosethat have a facultative or obligate parasiticassociations with insects. Entomogenous nematodeshave several deleterious effects on their hostsincluding sterility, reduced fecundity, longevity andflight activity, delayed development, or otherbehavioral, physiological and morphologicalaberrations and in some cases, rapid mortality.Parasitic associations with insects have beendescribed from 23 nematode families. Seven of thesefamilies contain species that have potential forbiological control of insects (Koppenhofer and Kaya,2001). A very few cause insect death but these
species are difficult (e.g., tetradomatids) or expensive(e.g. mermithids) to mass produce, have narrow hostspecificity against pests of minor economicimportance, possess modest virulence (e.g.,sphaeruliids) or are otherwise poorly suited toexploit for pest control purposes. The only insect-parasi tic nematodes possessing an optimalbalance of biological control attributes areentomopathogenic or insecticidal nematodes in thegenera Steinernema and Heterorhabditis because oftheir ability to kill hosts quickly (1-4 d depending onnematode and host species). Mutualistic associationof these nematodes with bacteria of the genusXenorhabdus for Steinernematidae andPhotorhabdus for Heterorhabditidae have also beenreported by Boemare et al. (1993). However,although both the families belong to the same order,but are not closely related and have distinctlydifferent reproductive strategies (Blaxter et al. 1998).

164 AGRICULTURAL REVIEWS
Advantages:Entomopathogenic nematodes areextraordinarily lethal to many important insect pests,yet are safe for non target organisms (Georgis et al.,1991). This high degree of safety means that unlikechemicals, or even Bacillus thuringiensis, nematodeapplications do not require masks or other safetyequipment; and re-entry time, residues, groundwatercontamination, chemical trespass, and pollinatorsare not issues. Most biologicals require days or weeksto kill, yet nematodes, working with their symbioticbacteria, can kill insects within 24-48 hours. Dozensof different insect pests are susceptible to infection,yet no adverse effects have been shown againstbeneficial insects or other nontargets in field studies(Georgis et al., 1991; Akhurst and Smith, 2002).Nematodes are amenable to mass production anddo not require specialized application equipment asthey are compatible with standard agrochemicalequipments, including various sprayers (e.g.,backpack, pressurized, mist, electrostatic, fan, andaerial) and irrigation systems.
Life cycle: The entomopathogenic nematodes underSteinernematids and heterorhabditids have similarli fe histories. The free-l iving, non-feedingdevelopmentally arrested infective juveniles of thesenematode species have attributes of both insectparasitoids or predators, they have chemoreceptorsand are motile, like pathogens, highly virulent, killingtheir hosts quickly, and can be cultured easily andhave a high reproductive potential. When a host hasbeen located, the nematodes penetrate into the insectbody cavity, usually via natural body openings(mouth, anus, spiracles) or areas of thin cuticle.Wang and Gaugler (1999) compared the penetrationbehavior of S. glaseri and H. bacteriophora intoPopillia japonica larvae and found that S. glaseripenetrated primarily through the gut. H.bacteriophora was not efficient at penetrating thegut, presumably because of the thick peritrophicmembrane, but penetration through theintersegmental membranes of the cuticle. Once inthe body cavity, a symbiotic bacterium (Xenorhabdusfor steinernematids, Photorhabdus forheterorhabditids), that are motile, Gram-negative,faculat ively anaerobic rods in the fami lyEnterobacteriaceae released from the nematode gut,which multiplies rapidly and causes rapid insectdeath due to septicemia. Presently, eight species arerecognized but most isolates from recently described
entomopathogenic nematodes species have yet tobe determined (Hazir et al. 2003). The nematodesfeed upon the bacteria and liquefying host, andmature into adults. Steinernematid infective juvenilesmay become males or females, where asheterorhabditids develop into self-ferti lizinghermaphrodites although subsequent generationswithin a host produce males and females as well(Fig. 1). The life cycle is completed in a few days,and hundreds and thousands of new infectivejuveniles emerge in search of fresh hosts.
Mode of action:Entomopathogenic nematodes area nematode-bacterium complex. In this association,the nematode is dependent upon the bacterium forquickly killing its insect hosts, creating a suitableenvironment for its development by producingantibiotics that suppress competing secondarymicroorganisms and transforming the host tissuesinto a food source. The bacterium requires nematodefor protection from the external environment,penetration into the hosts haemocoel and inhibitionof the host’s antibacterial proteins.
Edaphic factors: Soil texture has an effect oninfective juvenile survival with survival being lowestin clay soils. The poor survival rate in clay soils isprobably due to lower oxygen levels in the smallersoil pores. Oxygen may also become a limiting factorin water-saturated soils and soils with contents oforganic matter. Soil pH does not have a strong effecton infective juvenile survival. Thus, infective juvenilesurvival at pH values between 4 and 8 does not vary,but survival declines at pH 10. Similarly, soil salinityhas only l imited negative effects onentomopathogenic nematode survival even atsalinity above the tolerance levels of most crop plants(Thurston et al., 1994). Sea water has no negativeeffects on survival of several Heterorhabditis species/strains (Griffin et al., 1994) and they have beenfrequently isolated from soils near the sea shore.However, high salinity (seawater) reduced the abilityto Heterorhabdistis to infect hosts but did improvetheir tolerance to high temperatures (Finnegan et al.,1999).
Production and storage technologyEntomopathogenic nematodes are mass
produced as biopesticides using in vivo or in vitromethods (Shapiro-Ilan and Gaugler 2002).

165Vol. 34, No. 3, 2013
In vivo production: Production methods forculturing entomopathogenic nematodes in insecthosts have been reported by various authors (Dutkyet al., 1964, Poinar 1979 and Kaya and Stock,1997). These references essentially describe systemsbased on the White trap (White, 1927), which takesadvantage of infective juveniles (IJ) naturalmigration away from the host cadaver uponemergence. Yield is affected by choice of nematodeand host species. Among nematode species, yield isgenerally proportional to host size (Blinova andIvanova, 1987 and Flanders et al., 1996) but yieldper milligram insect (within host species) andsusceptibility to infection is usually inverselyproportional to host size or age (Dutky et al., 1964and Blinova and Ivanova, 1987). In vivo productionrequires the least capital outlay and technicalexpertise (Friedman, 1990; Gaugler and Han, 2002).The most common insect host used for in vivolaboratory and commercial entomopathogenicnematode production is the last instar of the greaterwax moth, Galleria mellonella, because of its highsusceptibility to most nematodes, ease of rearing,wide availability and ability to produce high yields(Woodring and Kaya, 1988).
In vitro production: Production of entomo-pathogenic nematodes through in vitro requires athorough knowledge of biology and behavior of thenematode species produced. The only stage that canbe commercially used is the dauer juvenile (DJ), amorphologically distinct juvenile, formed as aresponse of depleting food sources and adverseenvironmental conditions. Mass production ofentomopathogenic nematodes has evolved from thefirst large scale in vitro soild media production byGlaser et al. (1940) to the three dimensional solidmedia in vitro process (Bedding, 1981 and 1984)and to the in vitro liquid fermentation productionmethod (Friedman, 1990). Currently, commercialnematodes are produced monoxenically using thesolid-media process developed by Bedding (1981and 1984) or the liquid-fermentation method. Thesolid media process has successfully producedpathogenic steinernematids and heterorhabditids,but the high labour cost is the main constraint(Bedding, 1990). The liquid-fermentation process,is highly efficient for several steinernematid speciesbut not for heterorhabditids (Gaugler and Georgis,1991).
Formulation of nematodes into a stableproduct has played a signi ficant role incommercialization of these biological-control agents.Active nematodes must be immobilized to preventdepletion of their lipid and glycogen reserves. Avariety of formulations have been developed tofacilitate nematode storage and application includingactivated charcoal, alginate and polyacrylamide gels,baits, clay, paste, peat, polyurethane sponge,vermiculite, and water-dispersible granules.Depending on the formulation and nematodespecies, successful storage under refrigeration rangesfrom one to seven months. Low temperature (2-50C)generally reduce metabolic activity and can thereforeenhance their shelf-life, some warm adapted speciessuch as H. indica and S. riobrave do not store wellattemperature below 100C (Strauch et al., 2000 andGrewal 2002)
Relative effectiveness and applicationparameters: Entomopathogenic nematodes arecertainly more specific and have no threat to theenvironment than chemical insecticides (Ehlers andPeters, 1995). In some instances, entomopathogenicnematodes have proven to be safe and effectivealternatives to chemical pesticides, but in numerousother cases they have failed to compete successfully(Georgis et al., 2006). Technological advances innematode production, formulation, quality control,application timing and delivery, and particularly inselecting optimal target habitats and target pests,have narrowed the efficacy gap between chemicaland nematode agents. S. feltiae is an efficacious andeconomical replacement for chemical insecticidesin the floriculture industry in the Netherlands,England and Germany (Jagdale et al., 2004)Entomopathogenic nematodes are remarkablyversatile in being useful against many soil and crypticinsect pests in diverse cropping systems, yet areclearly underutilized. Like other biological controlagents, nematodes are constrained by being livingorganisms that require specific conditions to beeffective. Desiccation and temperature extremes arethe most important abiotic factors affecting survivalof entomopathogenic nematodes (Glazer, 2002) andin soilinfective juveniles are subject to attack by avariety of microbial and invertebrate antagonists(Kaya, 2002). Thus, desiccation or ultraviolet lightrapidly inactivates nematodes in comparison tochemical insecticides. Similarly, nematodes are
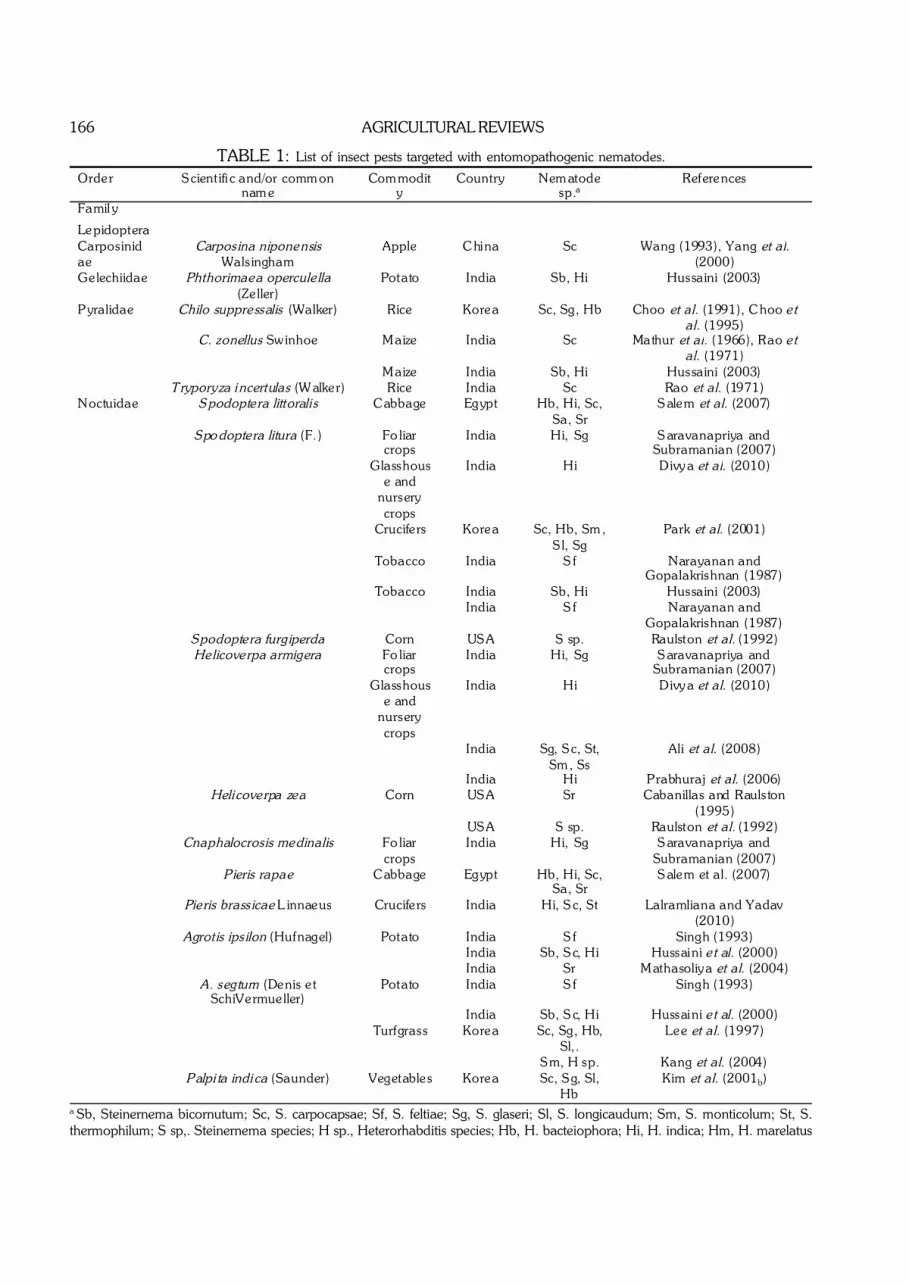
166 AGRICULTURAL REVIEWS
TABLE 1: List of insect pests targeted with entomopathogenic nematodes.
Order Famil y
Scientifi c and/or common name
Commodity
Country Nematode sp.a
References
Lepidoptera Carposinidae
Carposina niponensis Walsingham
Apple China Sc Wang (1993), Yang et al. (2000)
Gelechiidae Phthorimaea operculella (Zeller)
Potato India Sb, Hi Hussaini (2003)
Pyralidae Chilo suppressalis (Walker) Rice Korea Sc, Sg, Hb Choo et al. (1991), Choo et al. (1995)
C. zonellus Swinhoe Maize India Sc Mathur et al. (1966), Rao et al. (1971)
Maize India Sb, Hi Hussaini (2003) Tryporyza i ncertulas (Walker) Rice India Sc Rao et al. (1971) Noctuidae Spodoptera littoralis Cabbage Egypt Hb, H i, Sc,
Sa, Sr Salem et al. (2007)
Spo doptera litura (F.) Fo liar crops
India H i, Sg Saravanapriya and Subramanian (2007)
Glasshouse and
nursery crops
India H i Divya et al. (2010)
Crucifers Korea Sc, Hb, Sm, Sl, Sg
Park et al. (2001)
Tobacco India Sf Narayanan and Gopalakrishnan (1987)
Tobacco India Sb, Hi Hussaini (2003) India Sf Narayanan and
Gopalakrishnan (1987) Spodoptera furgiperda Corn USA S sp. Raulston et al. (1992) Helicoverpa armigera Fo liar
crops India H i, Sg Saravanapriya and
Subramanian (2007) Glasshous
e and nursery crops
India H i Divya et al. (2010)
India Sg, Sc, St, Sm, Ss
Ali et al. (2008)
India H i Prabhuraj et al. (2006) Helicoverpa zea Corn USA Sr Cabanillas and Raulston
(1995) USA S sp. Raulston et al. (1992) Cnaphalocrosis medinalis Fo liar
crops India H i, Sg Saravanapriya and
Subramanian (2007) Pieris rapae Cabbage Egypt Hb, H i, Sc,
Sa, Sr Salem et al. (2007)
Pieris brassicae L innaeus Crucifers India H i, Sc, St Lalramliana and Yadav (2010)
Agrotis ipsilon (Hufnagel) Potato India Sf Singh (1993) India Sb, Sc, H i Hussaini et al. (2000) India Sr Mathasoliya et al. (2004) A. segtum (Denis et
SchiVermueller) Potato India Sf Singh (1993)
India Sb, Sc, H i Hussaini et al. (2000) Turfgrass Korea Sc, Sg, Hb,
Sl,. Lee et al. (1997)
Sm, H sp. Kang et al. (2004) Palpi ta indi ca (Saunder) Vegetables Korea Sc, Sg, Sl,
Hb Kim et al. (2001b)
a Sb, Steinernema bicornutum; Sc, S. carpocapsae; Sf, S. feltiae; Sg, S. glaseri; Sl, S. longicaudum; Sm, S. monticolum; St, S.thermophilum; S sp,. Steinernema species; H sp., Heterorhabditis species; Hb, H. bacteiophora; Hi, H. indica; Hm, H. marelatus

167Vol. 34, No. 3, 2013
effective within a narrower temperature range(generally between 20 °C and 30 °C) than chemicals,and are more impacted by suboptimal soil type,thatch depth, and irrigation frequency (Georgis andGaugler, 1991; Shapiro-Ilan et al., 2006).Maintenance of high viability and virulence duringproduction, formulation and storage forms thebackbone of an effective quality control strategy.Effect of ageing of infective juveniles onpathogenicity was evaluated and compared byHussaini et al., 2005 with freshly hatched infectivejuveniles of the genera Steinernema andHeterorhabditis. S. tami and S. carpocapsaesurvived up to 40 days without mortality and loss ofinfectivity and S. abbasi with 2% mortality.Heterorhabditis spp. survived up to 20 days withoutmortality and declined to 60% and 20% after 40and 60 days of storage, respectively.
Therefore, based on the nematodes’ biology,applications should be made in a manner thatavoids direct sunlight, e.g., early morning or eveningapplications are often preferable. Soil in the treatedarea should be kept moist for at least two weeksafter applications (Klein, 1993). Application toaboveground target areas is difficult due to thenematode’s sensitivity to desiccation and UVradiation; however, improved formulations haveenhanced the efficacy of nematodes against aboveground target pests (e.g., Shapiro-Ilan et al., 2010).The most commonly used application method forentomopathogenic nematodes is spraying directlyon to the soil (or other) surface. Nematodes can beapplied with most commercially available sprayequipment including hand or ground sprayers, mistblowers etc. (Georgis et al., 1995). The infectivejuveniles can withstand pressure up to 1068 kPa andpass through all common nozzle type sprayers withopenings of about 100 µm (screen in nozzles shouldbe removed) in diameter. Nematodes can also bedelivered via irrigation systems including microjet,drip, sprinkler, etc. (Georgis et al., 1995 andCabanillas and Raulston, 1996). In all cases, thenematodes must be applied at a rate that is sufficientto kill the target pest; generally, 25 infective juvenilesper cm2 of treated area is required (rate can beincreased or decreased according to target pest andconditions) (Shapiro-Ilan et al., 2002). Selectivityof nematode species should be according to targetpest.
Appearance: Nematodes are formulated andapplied as infective juveni les as they aremorphologically, physiologically and behaviorallyadapted to act as the active ingradient of a biologicalpesticide. Infective juveniles range from 0.4 to 1.5mm in length. The location of the infective juvenilewithin the soil profile is one of the most importantfactor (Lewis, 2002). Formulation in water-dispersable granules is very successful with theambush forager S. carpocapsae, while the cruiseforaging S. feltiae and S. riobrave rapidly migrateout of the granules (Grewal, 2002). Disturbednematodes move actively, however sedentaryambusher species (e.g. S. carpocapsae, S.scapterisci) in water soon revert to a characteristic“J”-shaped resting position. Ambushing nematodespecies are usually associated with highly mobilesurface dwelling hosts. Entomopathogenicnematodes disperse horizontally and vertically afterapplication. S. carpocapsae infective juveniles moveupwards in soil column (Georgis and Poinar, 1983;Schroeder and Beavers, 1987) where as S. glaseriand H. bacteriophora move downwards, but theyalso disperse throughout the soil column. Lowtemperature or low oxygen levels will inhibitmovement of even active cruiser species (e.g., S.glaseri , H. bacteriophora). In short, lack ofmovement is not always a sign of mortality;nematodes may have to be stimulated (e.g., probes,acetic acid, gentle heat) to move before assessingviability. The infective juveniles do not feed, butrelies on stored energy reserves. Lipids (especiallytriglycerides) constitute up to 40% of the body weight(Selvan te al., 1993, Fitters et al., 1999) and aremost important energy reserve, though protein andthe carbohydrates, glycogen and trehalose, also yieldenergy (Qiu and Bedding, 2000). Good qualitynematodes tend to possess high lipid levels thatprovide a dense appearance, whereas nearlytransparent nematodes are often active but possesslow powers of infection.
Insects killed by most steinernematidnematodes become brown or tan, whereas insectskilled by heterorhabditids become red and the tissuesassume a gummy consistency. A dim luminescencegiven off by insects freshly killed by heterorhabditidsis a foolproof diagnostic for the genusHeterorhabdit is (the symbiotic bacteriaPhotorhabdus provide the luminescence). Black

168 AGRICULTURAL REVIEWS
Nematode species Product formulation Country
Steinernema carpocapsae ORTHO Biosafe USA Bio Vector USA
Exhibit USA Sanoplant
Boden Nutz;li nge Helix
X-GNAT Vector TL
USA USA USA
Switzerland Germany Canada
USA USA
S. feltiae Magent Nemasys Stealth
Entonem
USA UK UK
USA S. riobrave Vector MG
Bio Vector USA USA
S. scapteri sci USA Heterorhabditis bacteriophora Otinem USA H. megidis Nemasys UK S. carpocapsae Green commandos
Soil commandos India
TABLE 2: Commercial products available in international market.
cadavers with associated putrefaction indicate thatthe host was not killed by entomopathogenic species.Nematodes found within such cadavers tend to befree-living soil saprophages.
H abitat:Steinernematid and heterorhabditidnematodes are exclusively soil organisms. They areubiquitous, having been isolated from everyinhabited continent from a wide range of ecologicallydiverse soil habitats including cultivated fields,forests, grasslands, deserts, and even ocean beaches.When surveyed, entomopathogenic nematodes arerecovered from 2% to 45% of sites sampled(Hominick, 2002; Lalramliana and Yadav, 2010;Khatri-Chhetri et al. 2010).
Host range: Entomopathogenic nematodes havebeen tested against a large number of insect pestspecies with results varying from no effects toexcellent control. Because the symbiotic bacteriumkills insects so quickly, there is no intimate host-parasite relationship as is characteristic for otherinsect-parasitic nematodes. Consequently,entomopathogenic nematodes attacks a far widerspectrum of insects in the laboratory where the hostcontact is assured, environmental conditions areoptimal and no ecological or behavioral barriers toinfection exists(Gaugler 1981 and 1988) . Field hostrange is considerably more restricted, with somespecies being quite narrow in host specificity.Matching biology and ecology of both the nematode
and target pest are of great importance if nematodeapplications are to result in significant pestreductions. Nonetheless, when considered as a groupof nearly 80 species, entomopathogenic nematodesare useful against a large number of insect pests(Grewal et al., 2005). Although in field infections,impact on target populations has been frequentlyjudged modest or negligible, even whenconcentrations equivalent to billion of nematodesper hectare are applied (Finney and Walker, 1979).It would be premature to conclude that nematodesare safe for non target invertebrates in all habitats(Akhurst, 1990). Nevertheless, considerable evidenceindicates that nematodes possess a restricted hostrange in natural systems and, therefore, are not nearlyas threatening as their experimental host range mightsuggest (Kaya and Gaugler, 1993). Smitley et al.,(1992) were the fi rst to test the impact ofentomopathogenic nematodes on plant parasiticnematodes in a field trial and presently,entomopathogenic nematodes have been marketedfor control of certain plant parasitic nematodes,though efficacy has been variable depending onspecies (Lewis and Grewal, 2005).
Conservation:Reports and studies of naturaloccurrence and ecology of entomopathogenicnematodes are relatively uncommon. Informationfrom the manipulative experiments, however, canhelp to assess the effects of environmental factors,

169Vol. 34, No. 3, 2013
FIG. 1: Generalized life cycle of steinernematids andheterorhabditids. IJ = infective juvenile.
point out the gaps in our knowledge and help us tounderstand how this important group of naturalenemies of insects can be conserved (Lewis et al.,1998). The reasons for success or lack of success incontrolling insect pests, particularly in the soilenvironment, often remain unknown, underscoringthe need to obtain basic information on the biology,behavior, ecology and genetics of these nematodes.Research in behavioral ecology has clearlydemonstrated that these entomopathogenicnematodes are not generalist pathogens. Althoughmost of them have a broad host range in thelaboratory, in the field they are adapted to hosts in aparticular environment depending on their foragingstrategy. Their foraging strategy restricts much of theiractivity to certain soil stratum eliminating manyinsects from infection. Native populations are highlyprevalent, but, other than scattered reports ofepizootics, their impact on host populations isgenerally not well documented (Stuart et al., 2006).Epizootics of entomopathogenic nematode diseasesprobably occur regularly in soil, but they are difficultto detect and often go unrecorded (Kaya, 1987). Twoepizootics, both from Australia involving undescribedHeterorhabditis species, have been documented(Akhurst et al., 1992; Sexton and Williams 1981).In the first case, a significant reduction of thewhitefringed beetle, Graphagnathus leucoloma,larvae and adults was observed in lucerne field(Sexton and Williams 1981). In the second, twoundescribed species of Heterorhabditis were found
infecting four scarabaeid species in three adjoiningsugarcane fields (Akhurst et al., 1992). Underlaboratory conditions, the scarabaeid larvae are notsusceptible to the nematodes, but in the field infectedlarvae were readily recovered. This is largelyattributable to the mysterious nature of soil insects.Consequently, research and guidelines for conservingnative entomopathogenic nematodes are in need ofadvancement.
Compatibility: EPN infective juveniles (IJs) cantolerate short-term exposures (2-24 h) to manychemical and biological insecticides, fungicides,herbicides, fertilizers and growth regulators, and canthus be tank-mixed and applied together. This offersa cost-effective alternative to pest control, andfacilitates the use of nematodes in integrated pestmanagement systems. However, the actualconcentration of the chemical to which thenematodes will be exposed will vary depending uponthe application volume and system used (Alumai andGrewal, 2004). Compatibility has been tested withwell over 100 different chemical pesticides.Entomopathogenic nematodes are compatible (e.g.,may be tank-mixed) with most chemical herbicidesand fungicides as well as many insecticides (suchas bacterial or fungal products) (Koppenhöfer andGrewal, 2005). In fact, in some cases, combinationsof chemical agents with nematodes results insynergistic levels of insect mortality. Some chemicalsto be used with care or should be avoided likealdicarb, carbofuran, diazinon, dodine, methomyl,and various nematicides as they do not have anycompatible interact ion. However, specif icinteractions can vary based on the nematode andhost species and applicat ion rates. Theorganophosphate oxamyl increased S. carpocapsaeefficacy against Agrotis segetum synergistically, butonly in fumigated soil, probably by enhancing thenematodes nictation behavior (Ishibashi, 1993).Furthermore, even when a specific chemicalpesticide is not considered compatible, use of bothagents (chemical and nematode) can beimplemented by maintaining an appropriate intervalbetween applications (e.g., 1 – 2 weeks). Prior touse, compatibility and potential for tank-mixingshould be based on manufacturer recommendations.Combination of two nematode species may provideeffective control of two pests (Kaya et al., 1993) and

170 AGRICULTURAL REVIEWS
the nematode species may coexist (Koppenhofer andKaya, 1996) if the target differs in susceptibilityagainst the search strategies and/or pathogenicityof the two nematodes. Similarly, entomopathogenicnematodes are also compatible with many pathogens(Koppenhöfer and Grewal, 2005) like combinationof Bacillus thuringiensis (Bt) against severallepidopterous pests resulted in additive andantagonistic interactions. Interactions range fromantagonism to additivity or synergism depending onthe specific combination of control agents, targetpest, and rates and timing of application.Koppenhofer et al., (2000) documented synergismbetween Heterrhabditis spp. and S. glaseri andneonicotinoid imidacloprid in scarab larvae. Asimidacloprid reduces the grub’s defensive behaviorresulting in increased nematode attachment andpenetration. Nematodes are generally compatiblewith chemical fertilizers as well as composted manurethough fresh manure can be detrimental.
Commercial availability: Of the nearly eightysteinernematid and heterorhabditid nematodesidentified to date, at least twelve species have beencommercialized. A list of some commerciallyavailable entomopathogenic nematode products ininternational market are given in table 2 (Siddiquiet al., 2010). One billion nematodes per acre(250,000 per m2) is the rule-of-thumb against mostsoil insects (greenhouse soils tend to be treated athigher rates).
Conclusions: Entomopathogenic nematology hasa relatively short history dating back to the pioneeringresearch of R.W. Glaser and his coworkers in the1930s and 1940s (Gaugler R and Kaya HK, 1990).The primary emphasis of their research and of othersfollowing them focused on developing and usingthese nematodes as biological insecticides. Thereasons for success or lack of success in controllinginsect pests, particularly in the soil environment, oftenremains unknown, underscoring the need to obtainbasic information on the biology, behavior, ecology
and genetics of these nematodes. Recently advancesmade in nematode behavior and ecology clearlydemonstrates that they are not generalist pathogens;their behavior, for example, restricts much of theiractivity to certain soil stratum, eliminating manyinsects from infection. Understanding thesebehavioral patterns and their genetics will enhancethe use and production of the most adapted speciesfor insect control in the field.
Entomopathogenic nematodes (Rhabditida:Steinernematidae and Heterorhabditidae) inparticular have emerged as excellent biocontrolagents of soil-dwelling insect pests and henceattracted widespread commercial interest. Thesebiological control agents are endowed with manyadvantages including host seeking capability, highvirulence, ease of production, ease of application,mammalian safety and exception from registrationin many countries. They also possess a broad hostrange, are compatible with many other controlagents, are widespread in distribution and can beformulated and stored for reasonable length of time.However, the efficacy of these nematodes againstinsect pests varied, particularly in soil environment,are often unknown, emphasizing the need to obtainbasic information on the biology, behavioral ecologyand genetics of these nematodes. Indeed, there hasbeen a surge in research to obtain more basicinformation and in exploration to discover newnematode isolates. These nematodes are ubiquitousin nature, but their populations usually are notsufficiently high to cause epizootic and reduce pestpopulation. Research is needed to better understandthe factors that regulate their population and on howtheir population can be manipulated to initiateepizootic in insect-pest populations. Finally, thesefascinating animals may contribute more to sciencethan their use solely as biological control agents. Tobegin with, they may be useful tools in understandingthe evolution of parasitism and symbiosis andmechanisms by insect resistance to infection.
REFERENCESAkhurst, R. and K. Smith. (2002). Regulation and safety. In: Gaugler, R. (Ed.), Entomopathogenic Nematology. CABI,
New York, NY, pp-311-332.Akhurst, R.J., (1990). Safety to nontarget invertebrates of nematodes of economically important pests. In Safety of
Microbial pp. 234-238. Boca Raton, FL: CRC 259 pp.Akhurst, R.J., Bedding, R.A., Bull, R.M. and Smith, D.R.J. (1992). An epizootic of Heterorhabditis spp. (Heterorhabditidae:
Nematoda) in sugarcane scarabaeids (Coleoptera). Fund Appl Nematol., 15: 71-73.

171Vol. 34, No. 3, 2013
Alumai, A. and Grewal, P.S. (2004). Tankmix-comaptibility of the entomopathogenic nematodes, Heterorhabditisbacteriophora and Steinnernema carpocapsae, with selected chemical pesticides using in turfgrass. BiocontrolSci Tech., 14: 725-730.
Bedding, R.A. (1981). Low cost in vitro production of Neoplectana and Heterorhabditis species (Nematoda) for fieldcontrol of insect pests. Nematologica., 27: 109-114
Bedding, R.A. (1984). Large scale production, storage and transport of the insect-parasitic nematodes Neoaplectanaspp. and Heterorhabditis spp. Ann Appl Bio., 104: 117-120.
Bedding, R.A. (1990). Logistics and strategies for introducing entomopathogenic nematodes technology into developingcountries. In: Gaugler, R. and Kaya, H.K. (eds) EPN. Biol. Control., CRC Press, Boca Raton, Florida, pp-233-248.
Blaxter, M.L, DeLey, P. Gary, J.R., Liu, L.X., Scheldeman, P. and Vierstraete, A. 1998. A molecular evolutionaryframework for the phylum Nematoda. Nature., 392: 71-75.
Blinova, S.L. and Ivanova, E.S. (1987) Culturing the nematode-bacterial complex of Neoplectana carpocapsaeininsects. In: Sonin, M.D. (ed.) Helminthes of insects, Amerind Publishing, New Delhi, pp-22-26.
Boemare, N.E., Akhurst, R.J. and Mourant, R.G. (1993). DNA relatedness between Xenorhabdus spp. (Enterobacteriaceae),symbiotic bacteria of entomopathogenic nematodes, and a proposal to transfer Xenorhabdus luminescens to anew genus, Photorhabdus gen. nov. Int J Sys Bact., 43: 249-255.
Cabanillas, H.E and Raulston, J.R. (1996). Effects of furrow irrigation on the distribution and infectivity of Steinernemariobrave against corn earworm in corn. Fund Appl Nematol., 19: 273-281.
Cabanillas, H.E. and Raulston, J.R. (1995). Impact of Steinernema riobrave (Rhabditida: Steinernematidae) on theControl of Helicoverpa zea (Lepidoptera: Noctuidae) in Corn. J Econ Entomol., 88(1): 58-64.
Choo, H.Y., Kaya, H.K., Huh, H., Lee, D.W., Kim, H.H., Lee, S.M., Choo, Y.M., (2002). Entomopathogenic nematodes(Steinernema spp. and Heterorhabditis bacteriophora) and a fungus Beauveria brongniartii for biological controlof the white grubs, Ectinihoplia rufipes and Exomala orientalis, in Korean golf courses. BioControl., 47: 177–192.
Choo, H.Y., Kaya, H.K., Kim, J.B., Park, Y.D., (1991). Evaluation of entomopathogenic nematodes, Steinernemacarpocapsae (Steinernematidae) and Heterorhabditis bacteriophora (Heterorhabditidae) against rice stem borerChilo suppressalis (Walker) (Lepidoptera: Pyralidae). Korean J Appl Entomol., 30: 50–53.
Choo, H.Y., Kim, H.H., Lee, D.W., Park, Y.D., (1996). Microbial control of fly maggots with entomopathogenic nematodesand fungus in outhouses of farm houses. Korean J Appl Entomol., 35: 80–84.
Choo, H.Y., Lee, S.M., Chung, B.K., Park, Y.D., Kim, H.H., (1995). Pathogenicity of Korean entomopathogenicnematodes (Steinernematidae and Heterorhabditidae) against local agricultural and forest insect pests. KoreanJ Appl Entomol., 34: 314–320.
Divya, K. Sankar, M. and Marulasiddesha, K.N. (2010). Efficacy of entomopathogenic nematode, Heterorhabditisindica against three lepidopteran insect pests. Asian J Exp Biol Sci., 1 (1): 183-188.
Dutky, S.R., Thompson, J.V. and Cantwell, G.E. (1964). A technique for the mass propagation of the DD-136 nematode.J Insect Pathol., 6: 417-422.
Ehlers, R.U and Peters, A. (1995). Entomopathogenic nematodes in biological control: feasibility, perspectives andpossible risks. In: Hokkanen, HMT and Lynch JM (eds.) Biol. Control: Benefits and Risks. Cambridge UniversityPress, UK, pp- 119-136.
Finnegan, M.M., Downes, M.J., Regan, M.O. and Griffen, C.T., (1999). Effect of salt and temperature stress on survivaland infectivity of Heterorhabditis spp. Infective juveniles. Nematology., 1:69-78.
Finney, J.R and Walker, C. (1979). Assessment of a field trial using the DD-136 strain of Neoaplectana sp. for the controlof Scolytus scolytus. J Invertebr Patho., 33: 239-241.
Fitters, P.F.L, Patel, M.N, Griffin, C.T and Wright, D.J. (1999). Fatty acid composition of Heterorhabditis sp. duringstorage. Comp Biochem Physiol.,124: 81-88.
Flanders, K.L., Miller, J.M. and Shields, E.J. (1996). In vivo production of Heterorhabditis bacteriophora ‘Oswego’(Rhabditida: Heterorhabditidae), a potential biological control agent for soil-inhabiting insects in temperateregions. J Econ Entomol., 89:373-380.
Friedman, M.J. (1990). Commercial production and development. In: Gaugler, R. and Kaya, H.K. (eds) Entomopathogenicnematodes in Biological Control. CRC Press, Boca Raton, Florida, 153-172.
Gaugler, R. (1981). Biological control potential of neoaplectanid nematodes. J Nematol, 13: 241-249.Gaugler, R. (1988). Ecological considerations in the biological control of soil-inhabiting insect pests with entomopathogenic
nematodes. Agricult Ecosys Environ., 24:351-360.

172 AGRICULTURAL REVIEWS
Gaugler, R. (1991). Biological control potential of neoaplectanid nematodes. J Nematol, 13: 241-249.Gaugler, R. and Georgis, R. (1991). Culture method and efficacy of entomopathogenic nematodes (Rhabditida:
Steinernematids and Heterorhabditidae). Bio. Control., 1:269-274.Gaugler, R. and Han, R. (2002). Production technology. In: Gaugler, R. (ed.) Entomopathogenic Nematology. CAB
International, Wallingford, UK, 289-310.Gaugler, R. and Kaya, H.K. (1990). EPN. Bio Control. Boca Raton, FL: CRC Press.Georgis, R. and Poinar, G.O Jr. (1983). Effect of soil texture on the distribution and infectivity of Neoplectana carpocapsae
(Nematoda: Steinernematidae). J Nematol, 15: 308-311.Georgis, R. and R. Gaugler. (1991). Predictability in biological control using entomopathogenic nematodes. J Econ
Entomol., [Forum] 84: 713-20.Georgis, R., Dunlop, D.B and Grewal, P.S. (1995). Formulation of entomopathogenic nematodes. In FR Hall and JW
Barry (eds.), Biorational Pest Control Agents : Formulation and Delivery, ACS Symposium Series No. 595.Amer. Chem. Soc., Washington, DC, pp-197-205.
Georgis, R., H. Kaya, and Gaugler, R. (1991). Effect of steinernematid and heterorhabditid nematodes on nontargetarthropods. Environ Entomol., 20: 815-22.
Georgis, R., Koppenhoper, A.M., Lacey, L.A., Belair, G., Duncan, L.W., Grewal, P.S., Samish, M., Tan, L., Torr, P. andVanTol, R.W.H.M. 2006. Successes and failures in the use of parasitic nematodes for pest control. Biol Control.,38: 103–123.
Glaser, R.W., McCoy, E.E., Girth, H.B. (1940). The biology and economic importance of a nematode parasitic ininsects. J Parasitol. 26: 479-495.
Glazer, I. (2002). Survival biology. In: Gaugler R 9ed.) EPN., CAB International, Wallingford, UK, pp-169-187.Grewal, P. S., R-U, Ehlers, and D. I. Shapiro-Ilan. (2005). Nematodes as Biocontrol Agents. CABI, New York, NY. Grewal, P.S. (2002). Formulation and application technology. In: Gaugler R (ed.) EPN. Nematol.,CAB International,
Wallingford, UK pp-265-287.Griffin, C.P., Finnegon, M.M. and Downes, M.J. (1994). Environmental Tolerances and the dispersal of Heterorhabdites:
Survival and infectivity of European Heterorhabditis following prolonged immersion in seawater. Fundam andAppl Nematol., 17:415-421.
Han, R., Liu, X., Xu, J., Xie, R., (1996). Effects of entomopathogenic nematodes on the spatial distribution pattern oflitchi beetle, Aristobia testudo larvae in litchi orchards and the economic threshold. Natural Enemies of Insects18, 99–105.
Hazir, S., Kaya, H.K., Stock, S.P., Keskun, N. (2003). Entomopathogenic nematodes (Steinernematidae andHeterorhabditidae) for biological control of soil pests. Turk J Biol., 27: 181-202.
Head, J.,Walters, K.F.A., (2003). Augmentation biological control utilizing the entomopathogenic nematode, Steinernemafeltiae, against the South American leafminer, Liriomyza huidobrensis. Proc. First Int. Symp. Biol. Control, 13–18 January 2003, Hawaii, USA. USDA Forest Service, FHTET-03-05, pp-136–140.
Hominick, W. M. (2002). Biogeography. In: Gaugler, R. (Ed.), EPN., CABI, New York, NY, pp-115-143. Hussaini, S.S, Nagesh, M. and Shakeela, V. (2005). Survival of infective juvenile of entomopathogenic nematodes
under storage and their infectivity against Galleria mellonella and Spodoptera litura. Indian J Plant Prot., 33(1): 68-71.
Hussaini, S.S., (2003). Progress of research work on entomopathogenic nematodes in India. In: Hussaini, S.S., Rabindra,R.J., Nagesh, M. (Eds.), Current status of research on entomopathogenic nematodes in India. PDBC, Bangalore,India, pp. 27–68.
Hussaini, S.S., Singh, S.P., Parthasarathy, R. and Shakeela, V. (2000). Virulence of native entomopathogenic nematodesagainst black cutworms, Agrotis ipsilon (Hufnagel) and A. segetum (Noctuidae: Lepidoptera). Indian J Nematol.,30(1): 86-110.
Iiboshi, H, Izono, Y., (2003). Official test results of Steinernema carpocapsae for control of red palm weevil (Rhychopherousferrugineus). In: Report of Novel Pesticide Practical Examination. Japan Corp. Protection Association, Komagome,Tokyo, pp. 228–231. (In Japanese).
Ishibashi, N. (1993). Integrated control of insect pests by Steinernema carpocapsae. In: Bedding RA, Akhurst RJ andKaya HK (eds.) Nematodes and the Biological Control of Insects. CSIRO Publications, East Melbourne, Australia,pp-105-113.
Jagdale, G.B., Casey,M.L., Grewal, P.S., Lindquist, R.K., (2004). Eûects of application rate and timing, potting mediumand host plant on eûcacy of Steinernema feltiae against fungus gnat, Bradysia coprophila, in ûoriculture. BiolControl., 29: 296–305.

173Vol. 34, No. 3, 2013
Kang, Y.J., Lee, D.W., Choo, H.Y., Lee, S.M., Kweon, T.W., Shin, H.K., 2004. Biological control of Spodopteradepravata (Butler) (Lepidoptera: Noctuidae) using entomopathogenic nematodes. Korean J Appl Entomol.,43, 61–70.
Kaya, H. (2002). Natural enemies and other antagonists. In Gaugler R (ed.). EPN., CAB International, Wallingford, UK,pp-189-203.
Kaya, H.K. (1987). Diseases caused by nematodes. In Epizootiology of Insect Diseases, ed. JR Fuxa, Y tanada, pp. 453-470. New York: Wiley and Sons.
Kaya, H.K. and Gaugler, R. (1993). Entomopathogenic Nematodes. Annu Rev Entomol., 38: 181-206.Kaya, H.K. and Stock, P. (1997). Techniques in insect nematology. In: Lacey, L.A. (ed.) Manual of Techniques in Insect
pathology. Academic Press, San Diego, California, pp-282-324.Kaya, H.K., Burlando, T.M. and Thurston, G.S. (1993). Two entomopathogenic nematode species with different search
strategies for insect suppression. Environ Entomol., 22: 859-864.Khatri-Chhetri, H.B., Waeyenberg,e L., Manandhar, H.K. and Moens, M. (2010). Natural occurrence and distribution of
entomopathogenic nematodes (Steinernematidae and Heterorhabditidae) in Nepal. J Invertebr Pathol. 103:74-78.
Kim, H.H., Choo, H.Y., Kaya, H.K., Lee, D.W., Lee, S.M., Jeon, H.Y., (2004). Steinernema carpocapsae (Rhabditida:Steinernematidae) as a biological control agent against the fungus gnat Bradysia agrestis (Diptera: Sciaridae) inpropagation house. Biocontrol Sci Technol., 14: 171–183.
Kim, H.H., Choo, H.Y., Lee, D.W., Lee, S.M., Jeon, H.Y., Cho, M.R., Yiem, M.S., (2003). Control eYcacy of Koreanentomopathogenic nematodes against fungus gnat, Bradysia agrestis (Diptera: Sciaridae) and persistence inbed soil. J Korean Soc Hort Sci., 44:393–401.
Kim, H.H., Choo, H.Y., Lee, H.S., Park, C.G., Lee, D.W., Jin, B.R., Choo, Y.M., 2001a. Biological control of Lycoriellamali (Diptera: Sciaridae), a pest of oyster mushroom, Pleurotus ostreatus using entomopathogenic nematodes.Korean J Appl Entomol., 40: 59–67.
Kim, H.H., Choo, H.Y., Park, C.G., Lee, S.M., Choo, Y.M., 2001b. Biological control of cotton caterpillar, Palpitaindica Saunder (Lepidoptera: Pyralidae) with entomopathogenic nematodes. Korean J Appl Entomol., 40:245–252.
Klein, M.G., (1993). Biological control of scarabs with entomopathogenic nematodes. In: Bedding, R., Akhurst, R.,Kaya, H. (Eds.), Nematodes and the Biological Control of Insect Pests. CSIRO Press, East Melbourne, Australia,pp-49–58.
Koppenhöfer, A. M. and P. S. Grewal. (2005). Compatibility and interactions with agrochemicals and other biocontrolagents. In: Nematodes as Biocontrol Agents. CABI, New York, NY, pp-363-381.
Koppenhofer, A.M. and Kaya, H.K. (1996). Coexistence of two steinernematid nematode species (Rhabditida:Steinernematidae) in the presence of two host species. Appl Soil Ecol., 4: 221-230.
Koppenhöfer, A.M. and Kaya, H.K. (2001). Entomopathogenic nematodes and insect pest management. In: Advancesin Biopesticide research, Vol. 2 (O. Koul, Ed.), Harwood Academic Publishers, Amsterdam, The Netherlands.Pp-277-305.
Koppenhofer, A.M., Grewal, P.S. and Kaya, H.K. (2000). Synergism of imidacloprid and entomopathogenic nematodesagainst white grubs: the mechanism. Entomol Exp Appl., 94: 283-293.
Lalramliana, and Yadav, A.K. (2010). Laboratory evaluation of the pathogenicity of three entomopathogenic nematodesagainst larvae of cabbage butterfly, Pieris brassicae Linnaeus (Lepidoptera: Pieridae). Science Vision 9 (4): 166-173.
LeBeck, L.M., Gaugler, R., Kaya, H.K., Hara, A.H., Johnson, M.W., (1993). Host stage suitability of the leafminerLiriomyza trifolii (Diptera: Agromyzidae) to the entomopathogenic nematode Steinernema carpocapsae(Rhabditida: Steinernematidae). J Invertebr Pathol., 62:58–63.
Lee, S.M., Lee, D.W., Choo, H.Y., Kim, D.W., Kim, J.B., (1997). Pathogenicity of entomopathogenic nematodes tosome agro-forest insect pests. Korean J Soil Zoo., 2:76–82.
Lewis, E. (2002). Behaviooural ecology. In: Gaugler R (ed.) EPN. CAB International, Wallingford, UK, pp-205-223.Lewis, E.E. and. Grewal, P. S. (2005). Interactions with plant parasitic nematodes. In: Grewal, P.S., Ehlers, R.-U., and
Shapiro-Ilan, D.I. (Eds.), Nematodes as Biocontrol Agents. CABI, New York, NY., pp-349-362.Lewis, E.E., Campbell, J.F. and Gaugler, R. (1998). A conservation approach to using entomopathogenic nematodes in
turf and landscape. In: Barbosa P (ed.) Conserv. Biol. Control,. Academic Press, San Diego, California, pp-235-254.

174 AGRICULTURAL REVIEWS
Mannion, C.M., McLane, W., Klein, M.G., Moyseenko, J., Oliver, J.B., Cowan, D., 2001. Management of early-instarJapanese beetle (Cole- optera: Scarabaeidae) in ûeld-grown nursery crops. J Econ Entomol.,94: 1151–1161.
Mason, J.M. and Wright, D.J. (1997). Potential for the control of Plutella xylostella larvae with entomopathogenicnematodes. J of Invertebr Pathol., 70: 234-242.
Mathasoliya, J.B, Maghodia, A.B. and Vyas, R.V. (2004). Efficacy of Steinernema riobrave against Agrotis ipsilonHufnagel (Lepidoptera: Noctuidae) on Potato. Indian J Nematol., 34(2):177-179.
Mathur, S.B., Srivastava, R.P., Subba Rao, B.R.S., (1966). Records of nematodes parasitising Chilo zonellus Swinhoestem borer of maize. Indian J Entomol., 28: 414–415.
Narayanan, K. and Gopalakrishnan, C. (1987). Effect of entomopathogenic nematode, Steinernema feltiae (Rhabditida:Steinernematidae) to the pre-pupa, pupa and adult of Spodoptera litura (Noctuidae: Lepidoptera). Indian JNematol., 17(2): 273-276.
Nyasani, J.O., Kimenju, J.W., Olubayo, F.M. and Wilson, M.J. (2007). Potential of using entomopathogenic nematodesin the management of diamondback moth Plutella xylostella (L.) (Lepidoptera: Plutellidae) in Kenya. Afr CropSci Conference Proceedings Vol. 8: 1063-1066.
Park, S.H., Yu YS, Park, J.S., Choo HY, Bae S.D., and Nam M.H. (2001). Biological control of tobacco cutworm,Spodoptera litura Fabricius with entomopathogenic nematodes. Biotechnol. and Bioprocess Engr., 6: 139-143.
Piggott, S.J., Wright, D.J., Mathews, G.A., (2000). Polymeric formulation for the application of entomopathogenicnematodes against foliar pests. The BCPC Conference: Pests and diseases, Vol. 3. Proceedings of InternationalConference, Brighton, UK, 13–16 November 2000.
Poinar, G.O. (1979). Nematodes for biological Control of Insects. CRC Press, Boca Raton. Florida.Prabhuraj, A., Patil, B.V. and Shivaleela. (2006). Pathogenicity of Heterorhabditis indicus against Helicoverpa armigera
(Hubner). Indian J Entomol., 68(1): 36-38.Qiu, L. and Bedding, R. (2000). Energy metabolism and its relation to survival and infectivity of infective juveniles of
Steinernema carpocapsae under aerobic conditions. Nematology., 2: 551-559.Rao, Y.R.V.J., Prakash Rao, P.S., Verma, A., Israel, P., (1971). Tests with an insect parasitic nematode DD-136 against
the rice stem borer, Tryporyza incertulas. Indian J Entomol. 33:215–217.Raulston, J.R, Pair, S.D., Loera, J. and Cabanilla,s H.E. (1992). Prepupal and pupal parasitism of Helicoverpa zea and
Spodoptera frugiperda (Lepidoptera: Noctuidae) by Steinernema sp. In cornfields in the lower Rio Grandevalley. J Econ Entomol., 85(5): 1666-1670.
Salem, S.A., Abdel-Rahman, H.A., Zebitz, C.P.W, Saleh, M.M.E,, Ali Fawkia, I. and El-Kholy, M.Y. (2007). Evaluationof entomopathogenic nematodes in controlling some cabbage pests. J Appl Sci Res., 31 (4): 323-328.
Saravanapriya, B. and Subramanian, S. (2007). Pathogenicity of epn to certain foliar insect pests. Ann of Plant ProSci., 15(1): 219-222.
Schroede,r W.J. and Beavers, J.B. (1987). Movement of the entomopathogenous nematodes of the familiesHeterorhabitidae and Steinernematidae in soil. J Nematol,. 19: 257-259.
Schroeder, P.C, Ferguson, C.S., Shelton, A.M., Wilsey, W.T., Hoffmann, M.P. and Petzoldt, C. (1996). Greenhouse andfield evaluation of entomopathogenic nematodes (Nematoda: Heterorhabditidae and Steinernematidae) forcontrol of cabbage maggot (Diptera: Anthomyiidae) on cabbage. J Econ Entomol., 89(5): 1109-1115.
Selvan, S., Gaugler, R. and Lewis, E.E. (1993). Biochemical energy reserves of entomopathogenic nematodes. J ofParasitol., 79: 167-172.
Sexton, S.B. and Williams, P. (1981). A natural occurrence of parasitism of Graphagnathus luecoloma (Boheman) bythe nematode Heterorhabditis sp. J Austr Entomol Soc., 20: 253-255.
Shapiro-Ilan D. I. and R. Gaugler. (2002). Production technology for entomopathogenic nematodes and their bacterialsymbionts. J Ind Microbiol Biot., 28: 137-146.
Shapiro-Ilan, D. I., D. H. Gouge, and A. M. Koppenhöfer. (2002). Factors affecting commercial success: case studies incotton, turf and citrus. In: Gaugler, R. (Ed.), EPN., CABI, New York, NY, pp-333-356.
Shapiro-Ilan, D. I., T. E. Cottrell, R. F. Mizell, D. L. Horton, B. Behle, and C. Dunlap. (2010). Efficacy of Steinernemacarpocapsae for control of the lesser peachtree borer, Synanthedon pictipes: Improved aboveground suppressionwith a novel gel application. Bio Control., 54: 23–28.
Shapiro-Ilan, D.I., D. H. Gouge, S. J. Piggott, and J. Patterson Fife. (2006). Application technology and environmentalconsiderations for use of entomopathogenic nematodes in biological control. Bio Control., 38: 124-133.
Siddiqui, A.U., Pariha,r A. and Sharma, S.R. (2010). Importance and utilization of entom,opathogenic nematodes forthe suppression of insect pests. In: Gupta HCL, Siddiqui AV, Parihar A (eds.) Bio Pest Management(Entomopathogenic Nematodes, Microbes and Bioagents) Agrotech Publishing Academy, Udaipur, pp- 62-78.

175Vol. 34, No. 3, 2013
Singh, S.P., (1993). Effectiveness of an indigenous nematodes against citrus butterfly. J Insect Sci., 6: 107–108.Smitley, D.R., Warne,r W.R. and Bird, G.W. (1992). Influence of irrigation and Heterorhabditis bacteriophora on plant-
parasitic nematodes in turf. J Nematol., 24 (Suppl.): 637-641.Strauch, O, Niemann, I., Newmann, A., Schmidt, A.J., Peters, A, and Ehlers, R,U. (2000). Storage and formulation of
the entomopathogenic nematodes Heterorhabditis indica and H. bacteriophora. BioControl., 54: 483-500.Stuart, R. J., M. E. Barbercheck, P. S. Grewal, R.A.J. Taylor, and C. W. Hoy. (2006). Population biology of
entomopathogenic nematodes: Concepts, issues, and models. Biol Control., 38: 80-102.Thruston, G.S., Y.N. and Kaya, H.K. (1994). Influence of salinity on survival and infectivity of entomopathogenic
nematodes. J Nematol., 26:345-351.Wang, J., (1993). Control of the peach fruit moth, Carposina niponensis, using entomopathogenic nematodes. In:
Bedding, R., Akhurst, R., Kaya, H. (Eds.), Nematodes and the Biological Control of Insect Pests. CSIROPublications, East Melbourne, Australia, pp. 59–65.
Wang, Y. and Gaugler, R. (1999). Host and penetration site location by entomopathogenic nematodes against Japanesebeetle larvae. J Invertebr Pathol., 72: 313-318.
Wardlow, L.R., Piggott, S., Goldsworthy, R., (2001). Foliar application of Steinernema feltiae for the control of ûowerthrips. Medeelingen Faculteit Landbouwkundige Toegepaste Biol. Wetenschappen Univ. Gent 66, 285–291.
White, G.F. (1927). A method for obtaining infective nematode larvae from cultures. Science 66: 302-303.Williams, E.C., MacDonald, O.C., (1995). Critical factors required by the nematode Steinernema feltiae for the control
of the leaf miners, Liriomyza huidobrensis, Liriomyza bryoniae and Chromatomyia syngenesiae. Ann Appl Bio.127: 329–341.
Williams, E.C., Walters, K.F.A., (2000). Foliar application of the entomopathogenic nematode Steinernema feltiaeagainst leaf miners on vegetables. Biocontrol Sci and Tech., 10: 61–70.
Woodring, J.L. and Kaya, H.K. (1988). Steinernematid and Heterorhabditid Nematodes: A Handbook of Biology andTechniques. Southern Cooperative Bulltin 331, Arkansas Agricultural Experiment Station, Fayettville, Arkanas,30- pp.
Xu, J., Han, R., Liu, X., Cao, L., Yang, P., (1995). Application of codling moth nematode Steinernema carpocapsaeagainst the larvae of litchi longhorn beetle Aristobia testudo. Acta Phytophylacica Sinica 22: 12–16.
Yang, P., Liu, J., Liu, N., Han, R., Zhang, Y., Yu, C., (2000). Field control of the apple fruit moth Carposina nipponensis by using Steinernema carpocapsae nematodes. Nat Ene Insects., 22: 79–82.