enhanced enzymatic degradation resistance of plasmid dna in ionic liquids
TRANSCRIPT

RSC Advances
PAPER
Publ
ishe
d on
07
May
201
5. D
ownl
oade
d by
Mon
ash
Uni
vers
ity o
n 01
/06/
2015
07:
15:1
6.
View Article OnlineView Journal | View Issue
Enhanced enzym
aDepartment of Chemical Engineering, Mo
Australia. E-mail: Wenlong.Cheng@monashbSchool of Chemistry, Monash University ClcMonash Institute of Pharmaceutical Scienc
3052, Australia
† Electronic supplementary informa10.1039/c5ra05518k
Cite this: RSC Adv., 2015, 5, 43839
Received 28th March 2015Accepted 7th May 2015
DOI: 10.1039/c5ra05518k
www.rsc.org/advances
This journal is © The Royal Society of C
atic degradation resistance ofplasmid DNA in ionic liquids†
Romiza R. Mazid,a Alexandra Cooper,a Ying Zhang,a R. Vijayaraghavan,b
Douglas R. MacFarlane,b Christina Cortez-Jugoc and Wenlong Cheng*a
Plasmid deoxyribonucleic acid (pDNA) is a promising therapeutic in gene therapy and as a DNA vaccine, but
it is susceptible to degradation by nucleases in the extracellular environment. In this study, we report on the
enhanced enzymatic degradation resistance of pDNA, when stored in a hydrated buffered ionic liquid (BIL)
based on choline dihydrogen phosphate (CDHP). The stability of pDNA stored in buffered CDHP (bCDHP) in
the presence of Turbo DNase was studied using agarose gel electrophoresis, which showed a prolonged
shelf life at room temperature over a period of 28 days in bCDHP compared to within 10 minutes for
phosphate buffered saline (PBS). In addition, the biological activity of pDNA was maintained under such
conditions, as demonstrated by the expression of yellow fluorescent protein (YFP).
Introduction
Plasmid DNA (pDNA) is a powerful tool in genetics for multi-plying or expressing particular genes or therapeutic agents.However, pDNA is susceptible to nucleases particularly in vivoand during long term storage.1 It has been reported that thehalf-life of pDNA in vivo can be as short as 5–15 min.1,2 It wouldalso be advantageous to store plasmid DNA at room tempera-ture in non-aseptic conditions, which would enable ease ofglobal transportation and ease of administration. Presently, theideal storage conditions for pDNA are in a dry solid state atroom temperature, at �20 �C, �80 �C or cryogenic preservationat �196 �C for prolonged storage and stability of stocksolutions.3
One strategy that has been used for improving the stability ofDNA is by storage in BILs.4 One type of proton buffered ILs isachieved by introduction of a conjugate acid/base pair such asH2PO4
�/HPO42� ion components. These materials are typically
water miscible and can be formulated at various water contentsfrom low water levels (when they are oen referred to ashydrated ionic liquids) to relatively high water contents wheretheir properties approach those of a traditional aqueous saltsolution. ILs, particularly the systems based on choline dihy-drogen phosphate (CDHP) and its buffered version (bCDHP),have demonstrated their great potential as storage media forseveral biomolecules to date, including siRNA,5 linear DNA,6–11
nash University Clayton, Victoria, 3800,
.edu
ayton, Victoria, 3800, Australia
es, Monash University Parkville, Victoria
tion (ESI) available. See DOI:
hemistry 2015
and several enzymes/proteins, which were found to displayenhanced stability and activity in the presence of the IL.12–18
Despite this, storage of pDNA in BILs has not yet demonstratedto the best of our knowledge.
Here, we thoroughly investigated the biological, thermal andstructural activity of pDNA in bCDHP. The pDNA studiedconsists of 5 kbps with the yellow uorescent protein (YFP) asthe reporter gene. It is representative of a typical plasmid thatmay be used for DNA vaccines or gene therapy (see ESI Fig. S1†).When stored at 25 �C or 37 �C in the presence of Turbo DNase(TD), we found a signicant improvement in the thermal andstructural stability of pDNA using bCDHP as a storage buffer incomparison to PBS. In addition, the biological activity of theplasmid was maintained, as expression of YFP was observed inTD treated pDNA stored in bCDHP for up to one month at 25 �Cor up to one week at 37 �C, highlighting the benet of storingtherapeutic plasmids in a BIL.
Results & discussionStructural stability of pDNA in bCDHP
We rst investigated the structural stability of YFP-pDNAstored in PBS, 20% (w/w) bCDHP or 50% (w/w) bCDHP in theabsence and presence of TD using agarose gel electrophoresis(Fig. 1). TD offers 50 times more activity and 350% greatercatalytic activity than DNase I. It is also capable of maintainingits activity in the presence of high salt environments andcleaves double stranded DNA (dsDNA) and single strandedDNA (ssDNA) non-specically.19,20 It can be seen in Fig. 1 thatthe plasmid (in PBS) displays the typical three distinct forms ofpDNA: relaxed circular (20 kbps); linear (7 kbps); and super-coiled, which made up the majority of pDNA present. In both20% (w/w) bCDHP and 50% (w/w) bCDHP however, only two
RSC Adv., 2015, 5, 43839–43844 | 43839
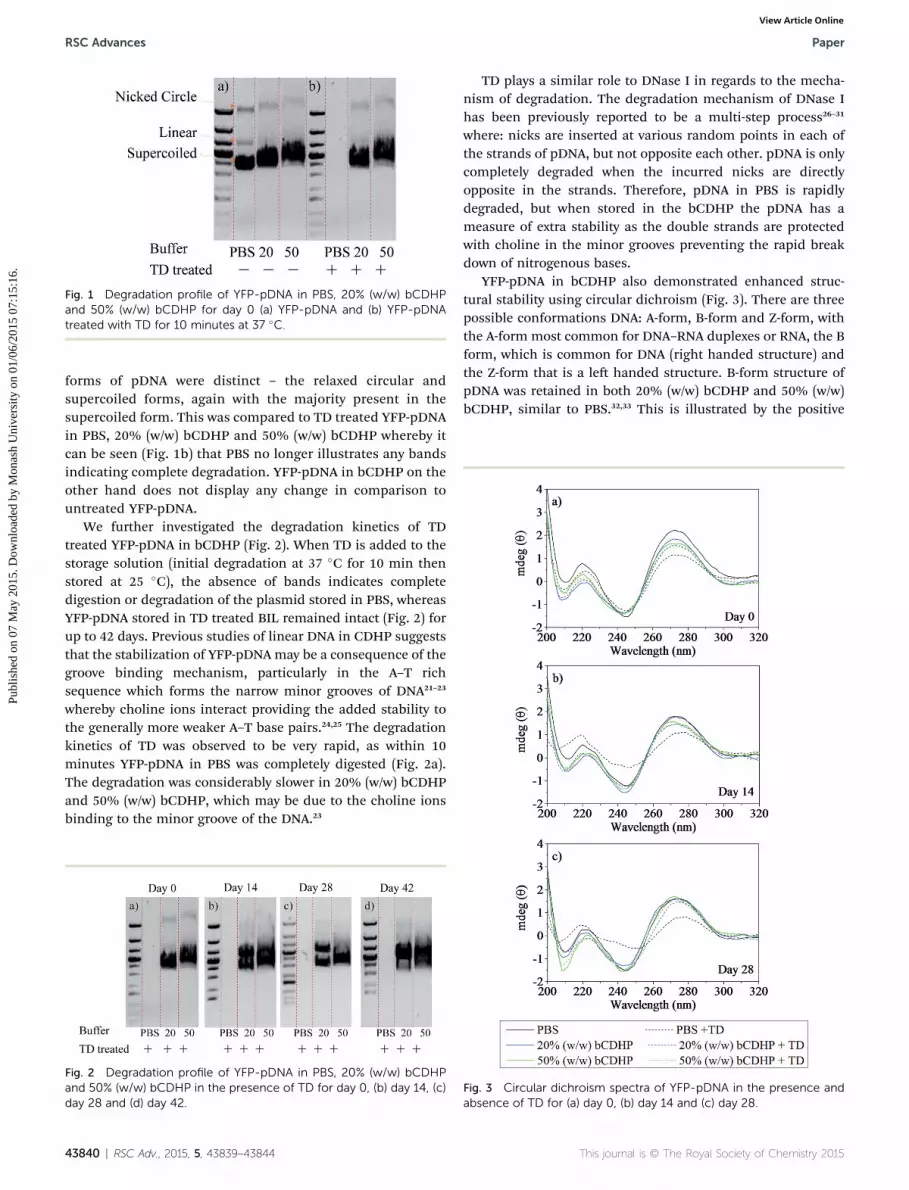
Fig. 1 Degradation profile of YFP-pDNA in PBS, 20% (w/w) bCDHPand 50% (w/w) bCDHP for day 0 (a) YFP-pDNA and (b) YFP-pDNAtreated with TD for 10 minutes at 37 �C.
RSC Advances Paper
Publ
ishe
d on
07
May
201
5. D
ownl
oade
d by
Mon
ash
Uni
vers
ity o
n 01
/06/
2015
07:
15:1
6.
View Article Online
forms of pDNA were distinct – the relaxed circular andsupercoiled forms, again with the majority present in thesupercoiled form. This was compared to TD treated YFP-pDNAin PBS, 20% (w/w) bCDHP and 50% (w/w) bCDHP whereby itcan be seen (Fig. 1b) that PBS no longer illustrates any bandsindicating complete degradation. YFP-pDNA in bCDHP on theother hand does not display any change in comparison tountreated YFP-pDNA.
We further investigated the degradation kinetics of TDtreated YFP-pDNA in bCDHP (Fig. 2). When TD is added to thestorage solution (initial degradation at 37 �C for 10 min thenstored at 25 �C), the absence of bands indicates completedigestion or degradation of the plasmid stored in PBS, whereasYFP-pDNA stored in TD treated BIL remained intact (Fig. 2) forup to 42 days. Previous studies of linear DNA in CDHP suggeststhat the stabilization of YFP-pDNA may be a consequence of thegroove binding mechanism, particularly in the A–T richsequence which forms the narrow minor grooves of DNA21–23
whereby choline ions interact providing the added stability tothe generally more weaker A–T base pairs.24,25 The degradationkinetics of TD was observed to be very rapid, as within 10minutes YFP-pDNA in PBS was completely digested (Fig. 2a).The degradation was considerably slower in 20% (w/w) bCDHPand 50% (w/w) bCDHP, which may be due to the choline ionsbinding to the minor groove of the DNA.23
Fig. 2 Degradation profile of YFP-pDNA in PBS, 20% (w/w) bCDHPand 50% (w/w) bCDHP in the presence of TD for day 0, (b) day 14, (c)day 28 and (d) day 42.
43840 | RSC Adv., 2015, 5, 43839–43844
TD plays a similar role to DNase I in regards to the mecha-nism of degradation. The degradation mechanism of DNase Ihas been previously reported to be a multi-step process26–31
where: nicks are inserted at various random points in each ofthe strands of pDNA, but not opposite each other. pDNA is onlycompletely degraded when the incurred nicks are directlyopposite in the strands. Therefore, pDNA in PBS is rapidlydegraded, but when stored in the bCDHP the pDNA has ameasure of extra stability as the double strands are protectedwith choline in the minor grooves preventing the rapid breakdown of nitrogenous bases.
YFP-pDNA in bCDHP also demonstrated enhanced struc-tural stability using circular dichroism (Fig. 3). There are threepossible conformations DNA: A-form, B-form and Z-form, withthe A-form most common for DNA–RNA duplexes or RNA, the Bform, which is common for DNA (right handed structure) andthe Z-form that is a le handed structure. B-form structure ofpDNA was retained in both 20% (w/w) bCDHP and 50% (w/w)bCDHP, similar to PBS.32,33 This is illustrated by the positive
Fig. 3 Circular dichroism spectra of YFP-pDNA in the presence andabsence of TD for (a) day 0, (b) day 14 and (c) day 28.
This journal is © The Royal Society of Chemistry 2015

Paper RSC Advances
Publ
ishe
d on
07
May
201
5. D
ownl
oade
d by
Mon
ash
Uni
vers
ity o
n 01
/06/
2015
07:
15:1
6.
View Article Online
maxima at �273 nm and a negative maxima at �243 nm, with acrossover occurring at �252 nm.33,34
It can be seen in Fig. 3 that YFP-pDNA demonstrates small tonegligible changes in its structure over the duration of 28 daysfor both untreated YFP-pDNA and those treated with TD,initially at 37 �C for 10 minutes and subsequently stored atroom temperature, in 20% (w/w) bCDHP and 50% (w/w)bCDHP. Any slight decrease in the circular dichroism signalunder these conditions may be the result of the choline ionsinteracting with the DNA.35 On the other hand, YFP-pDNA inPBS displays a signicant change in its structural integrity overthe 28 day period, particularly for TD treated YFP-pDNA. YFP-pDNA stored in PBS displayed limited structural stability andintegrity, exhibiting a reduction in its structural stability as thepeak at 273 nm is reduced for the control samples aer fourteendays and further by 28 days. Similarly, those treated with TDshowed pronounced loss of structural integrity, with animmediate decrease at 273 nm that further decreased at day 14and 28, and a concurrent positive increase of the negativemaxima, which indicates unwinding of the DNA helix.36
Thermal stability of pDNA in bCDHP
As further investigation into the stability of YFP-pDNA, we alsotested the potential thermal stability provided by bCDHP. TDtreated and non TD treated YFP-pDNA in PBS, 20% (w/w)bCDHP and 50% (w/w) bCDHP were measured for thermalstability. The presence of the BIL enhanced the thermal stabilityof YFP-pDNA relative to PBS, with up to 7 �C greater in meltingtemperature in 20% (w/w) bCDHP and 17 �C in 50% (w/w)bCDHP (Table 1 and see ESI Fig. S2†). Upon thermodynamicanalysis using the two state van't Hoff model as an approxi-mation of the pDNA denaturation, we also demonstrateenhanced binding stability in bCDHP represented by the Gibbsfree energy (DG0
25). YFP-pDNA binding stability stored in bCDHPincreases by almost 2 fold and 3 fold for 20% (w/w) bCDHP and50% (w/w) bCDHP respectively. This suggests much greaterthermal and thermodynamic stability of YFP-pDNA in bCDHP.
Following treatment with TD, the melting temperature ofYFP-pDNA in PBS was not possible to determine, indicating
Table 1 Thermodynamic parameters for the melting temperature of YF
DH0 [kcal mol�1] TDS0 [kc
PBSpDNA �34.79 � 0.8 �21.13 �pDNA + TD n.dc n.dc
20%(w/w) bCDHPpDNA �98.71 � 2.9 �73.26 �pDNA + TD �40.88 � 0.9 �22.32 �
50% (w/w) bCDHPpDNA �142.0 � 2.9 �101.1 �pDNA + TD �41.86 � 0.9 �23.43 �a All experiments were conducted in 0.01 M PBS and 20% (w/w) bCDHP anCary UV Thermal soware using van't Hoffmodelling. b Melting temperatumelting temperature was not able to be determined due to complete degr
This journal is © The Royal Society of Chemistry 2015
degradation of the plasmid, which supports the gel electro-phoresis data seen in Fig. 1 and 2. YFP-pDNA in the bCDHPremains very much intact, as the presence of TD only changesthe Tm by a degree or two (Table 1 and see ESI Fig. S3†). Inaddition, we also investigated the thermodynamic parametersof TD treated YFP-pDNA in bCDHP. The Gibbs free energy(DG0
25) gives an indication of the stability of the plasmid, andthe greater the negative value, the greater the stability. The DG0
25
of TD treated YFP-pDNA in bCDHP decreases in both 20% (w/w)bCDHP and 50% (w/w) bCDHP. Such an occurrence wasexpected as the binding stability is affected by the presence ofTD as it attempts to disrupt the choline cation binding in theminor grooves. Thus despite a loss in binding stability in thepresence of TD is seen in bCDHP, the melting temperature isretained.
Biological stability of pDNA in bCDHP
While the data above has demonstrated enhanced structuraland thermal stability of pDNA in bCDHP, a key requirement forpDNA is its ability to still be biological active and confer geneexpression in host cells. In this study, we have used pDNA withan inserted YFP reporter gene, but it is a typical plasmid thatmay be applicable as a pDNA vaccine by replacing the YFP genewith another gene of interest.37
The transfection with treated or non-treated YFP-pDNA wastested in cultured HEK 293T cells, using polyethylenimine (PEI)as the transfection agent. An increase in the uorescenceintensity of 293T cells, which is a result of the production ofYFP, is indicative of successful transfection. Typically,untreated 293T cells are non-uorescent and the uorescenceintensity of these cells was used as the base value. As expected,transfection from YFP-pDNA stored in PBS, resulted insubstantial expression of YFP (see ESI Fig. S4a†), with themajority of cells (65%) increasing in uorescence intensity.Similarly, successful transfection was achieved from YFP-pDNAstored in the bCDHP mixtures, with transfection efficiencies of31% and 21%, respectively, at day 0 (see Table 2 and ESI Fig. S4band c†).
P-pDNA in PBS, 20% (w/w) bCDHP and 50% (w/w) bCDHPa
al mol�1] DG025 [kcal mol�1] Tm
b [�C]
2.4 �13.66 � 0.9 72.54n.dc n.dc
8.0 �25.41 � 2.4 80.183.4 �18.57 � 0.5 81.10
18 �35.38 � 2.0 89.482.5 �18.45 � 0.8 87.15
d 50% (w/w) bCDHP. Thermodynamic parameters were evaluated fromre was calculated at a strand concentration of 0.2 mM. c n.d denotes thatadation.
RSC Adv., 2015, 5, 43839–43844 | 43841

Table 2 Transfection efficiency of FACS data for aged transfections(see ESI Fig. S4)a
Transfection efficiency [%]
Day 0 Day 14 Day 28
PBSpDNA 64.5 � 1.5 54 � 2.2 39.7 � 5.0pDNA + TD 3.4 � 2.4 1 � 0.2 0.1 � 0.5
20% (w/w) bCDHPpDNA 31.3 � 0.5 42.6 � 0.5 42.5 � 1.7pDNA + TD 29.4 � 0.8 40 � 4.3 1.1 � 0.9
50% (w/w) bCDHPpDNA 20.9 � 0.5 39.3 � 5.6 42 � 0.9pDNA + TD 20.6 � 0.4 42.6 � 0.2 40.1 � 0.9
a Each sample was conducted in triplicates.
RSC Advances Paper
Publ
ishe
d on
07
May
201
5. D
ownl
oade
d by
Mon
ash
Uni
vers
ity o
n 01
/06/
2015
07:
15:1
6.
View Article Online
In the presence of TD, however, there is no longer YFP geneexpression from YFP-pDNA stored in PBS. Meanwhile, YFP-pDNA stored in bCDHP maintained YFP gene expressionlevels similar to that of untreated plasmid for both 20% (w/w)bCDHP and 50% (w/w) bCDHP up to day 14 (see ESI Fig. S4(iand ii)†). Storage in latter mixture appeared to be superior to theformer as the gene expression of TD treated YFP-pDNA wassimilar to that of untreated YFP-DNA for up to 28 days, whereasin 20% (w/w) bCDHP, there is a gradual decrease in transfectionefficiency with incubation time and by day 28, the TD treatedYFP-pDNA is no longer expressing the YFP gene indicating lossof biological activity.
Fig. 4 Flow cytometry analysis of PEI-facilitated YFP-pDNA transfectionbCDHP, over a period of 7 days at 37 �C storage. The black line in the histoblue line represents the fluorescence intensity of untreated YFP-pDNA anby storage at 37 �C.
43842 | RSC Adv., 2015, 5, 43839–43844
The expression of YFP-pDNA stored in various buffers wasalso tested over a seven day period with continual storagetemperature of 37 �C. Aliquots were taken at 0 hours (no TDtreatment) and then TD treated samples at 10 minutes, 4 hours,12 hours, 24 hours, 48 hours and 7 days (Fig. 4). Similar to thetrend seen in Fig. S4,† it can be seen in Fig. 4 that TD treatedYFP-pDNA stored in PBS is degraded rapidly within 10 minutes(Fig. 4) and no transfection is observed. TD treated pDNA in20% (w/w) bCDHP and 50% (w/w) bCDHP continue to increasetheir level of gene expression or up to 12 hours at 37 �C, aerwhich the YFP expression from 20% (w/w) bCDHP begins todecrease while for 50% (w/w) bCDHP the pDNA maintains asimilar level of YFP production continually for 7 days (see ESIFig. S5† for transfection efficiencies).
Cytotoxicity of bCDHP
As a means of testing the toxicity of the bCDHP we diluted thestock concentrations of pDNA in PBS and bCDHP, and con-ducted cell viability tests. The cell viability tests were carried outusing ow cytometry at the same concentrations used fortransfections. The viability of cells in the presence of bCDHPnormalized against PBS treated HEK 293T cells (see ESIFig. S6†). Under the conditions used for transfection of pDNA,there was insignicant difference in the cell viability levelsbetween cells treated with PBS and those treated with bCDHP.Our results are supported by previous ndings on the cytotox-icity levels and the biocompatibility of bCDHP,5 which exhibitsminimal cytotoxicity in comparison to PBS. There was also verylittle difference in the toxicity levels between 20% (w/w) bCDHPand 50% (w/w) bCDHP. Thus the negligible cytotoxicity allowsfor the elimination of removing bCDHP from pDNA prior to
from plasmid stored in (a) PBS, (b) 20% (w/w) bCDHP and (c) 50% (w/w)gram represents fluorescence intensity of cells treatedwith buffer only,d the red line represents YFP-pDNA treatedwith TD for 10min followed
This journal is © The Royal Society of Chemistry 2015
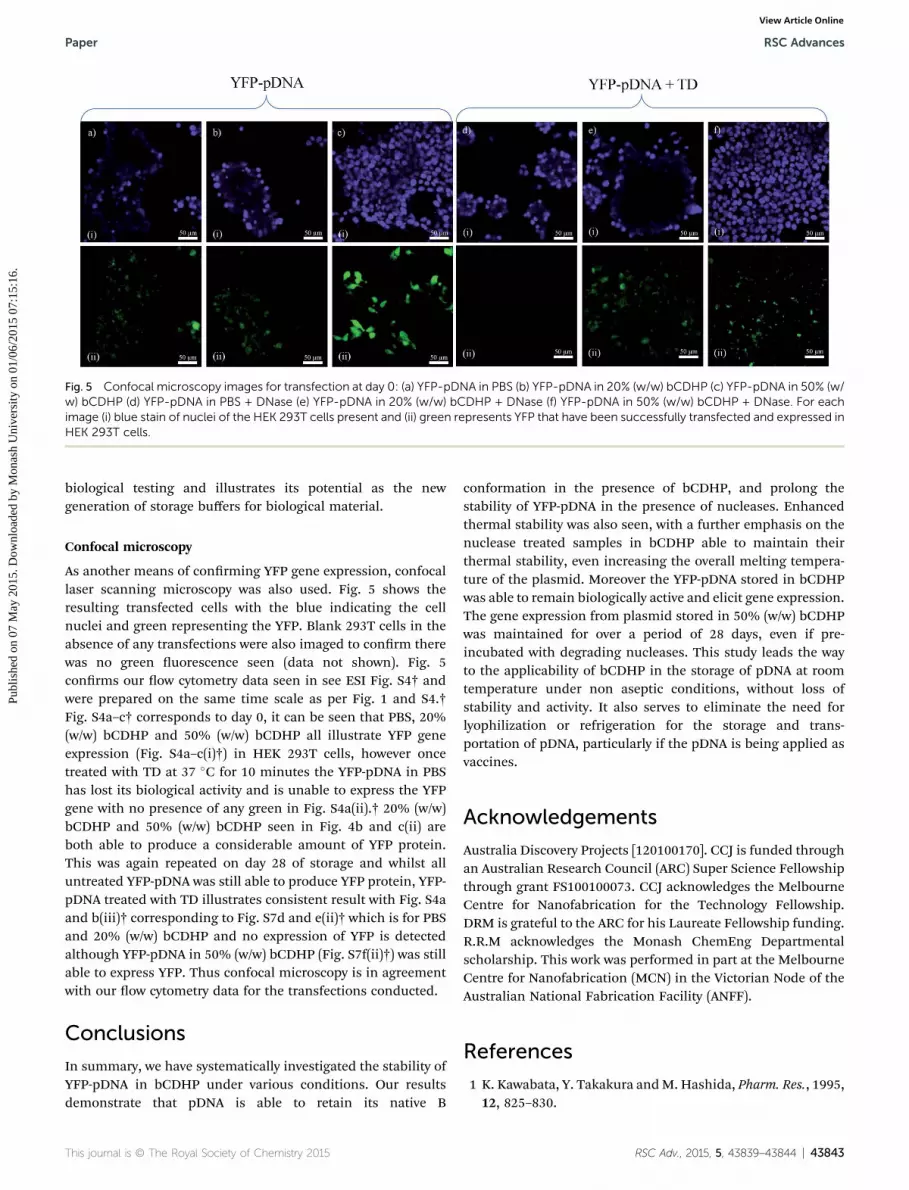
Fig. 5 Confocal microscopy images for transfection at day 0: (a) YFP-pDNA in PBS (b) YFP-pDNA in 20% (w/w) bCDHP (c) YFP-pDNA in 50% (w/w) bCDHP (d) YFP-pDNA in PBS + DNase (e) YFP-pDNA in 20% (w/w) bCDHP + DNase (f) YFP-pDNA in 50% (w/w) bCDHP + DNase. For eachimage (i) blue stain of nuclei of the HEK 293T cells present and (ii) green represents YFP that have been successfully transfected and expressed inHEK 293T cells.
Paper RSC Advances
Publ
ishe
d on
07
May
201
5. D
ownl
oade
d by
Mon
ash
Uni
vers
ity o
n 01
/06/
2015
07:
15:1
6.
View Article Online
biological testing and illustrates its potential as the newgeneration of storage buffers for biological material.
Confocal microscopy
As another means of conrming YFP gene expression, confocallaser scanning microscopy was also used. Fig. 5 shows theresulting transfected cells with the blue indicating the cellnuclei and green representing the YFP. Blank 293T cells in theabsence of any transfections were also imaged to conrm therewas no green uorescence seen (data not shown). Fig. 5conrms our ow cytometry data seen in see ESI Fig. S4† andwere prepared on the same time scale as per Fig. 1 and S4.†Fig. S4a–c† corresponds to day 0, it can be seen that PBS, 20%(w/w) bCDHP and 50% (w/w) bCDHP all illustrate YFP geneexpression (Fig. S4a–c(i)†) in HEK 293T cells, however oncetreated with TD at 37 �C for 10 minutes the YFP-pDNA in PBShas lost its biological activity and is unable to express the YFPgene with no presence of any green in Fig. S4a(ii).† 20% (w/w)bCDHP and 50% (w/w) bCDHP seen in Fig. 4b and c(ii) areboth able to produce a considerable amount of YFP protein.This was again repeated on day 28 of storage and whilst alluntreated YFP-pDNA was still able to produce YFP protein, YFP-pDNA treated with TD illustrates consistent result with Fig. S4aand b(iii)† corresponding to Fig. S7d and e(ii)† which is for PBSand 20% (w/w) bCDHP and no expression of YFP is detectedalthough YFP-pDNA in 50% (w/w) bCDHP (Fig. S7f(ii)†) was stillable to express YFP. Thus confocal microscopy is in agreementwith our ow cytometry data for the transfections conducted.
Conclusions
In summary, we have systematically investigated the stability ofYFP-pDNA in bCDHP under various conditions. Our resultsdemonstrate that pDNA is able to retain its native B
This journal is © The Royal Society of Chemistry 2015
conformation in the presence of bCDHP, and prolong thestability of YFP-pDNA in the presence of nucleases. Enhancedthermal stability was also seen, with a further emphasis on thenuclease treated samples in bCDHP able to maintain theirthermal stability, even increasing the overall melting tempera-ture of the plasmid. Moreover the YFP-pDNA stored in bCDHPwas able to remain biologically active and elicit gene expression.The gene expression from plasmid stored in 50% (w/w) bCDHPwas maintained for over a period of 28 days, even if pre-incubated with degrading nucleases. This study leads the wayto the applicability of bCDHP in the storage of pDNA at roomtemperature under non aseptic conditions, without loss ofstability and activity. It also serves to eliminate the need forlyophilization or refrigeration for the storage and trans-portation of pDNA, particularly if the pDNA is being applied asvaccines.
Acknowledgements
Australia Discovery Projects [120100170]. CCJ is funded throughan Australian Research Council (ARC) Super Science Fellowshipthrough grant FS100100073. CCJ acknowledges the MelbourneCentre for Nanofabrication for the Technology Fellowship.DRM is grateful to the ARC for his Laureate Fellowship funding.R.R.M acknowledges the Monash ChemEng Departmentalscholarship. This work was performed in part at the MelbourneCentre for Nanofabrication (MCN) in the Victorian Node of theAustralian National Fabrication Facility (ANFF).
References
1 K. Kawabata, Y. Takakura andM. Hashida, Pharm. Res., 1995,12, 825–830.
RSC Adv., 2015, 5, 43839–43844 | 43843

RSC Advances Paper
Publ
ishe
d on
07
May
201
5. D
ownl
oade
d by
Mon
ash
Uni
vers
ity o
n 01
/06/
2015
07:
15:1
6.
View Article Online
2 R. Mazid, M. X. Tan and M. K. Danquah, Curr. Pharm.Biotechnol., 2013, 14, 615–622.
3 J. G. Baust, Biopreserv. Biobanking, 2008, 6, 251–252.4 D. R. MacFarlane, R. Vijayaraghavan, H. N. Ha, A. Izgorodin,K. D. Weaver and G. D. Elliott, Chem. Commun., 2010, 46,7703–7705.
5 R. R. Mazid, U. Divisekera, W. Yang, V. Ranganathan,D. R. MacFarlane, C. Cortez-Jugo and W. Cheng, Chem.Commun., 2014, 50, 13457–13460.
6 K. Fujita and H. Ohno, Chem. Commun., 2012, 48, 5751–5753.7 D. Mondal, M. Sharma, C. Mukesh, V. Gupta and K. Prasad,Chem. Commun., 2013, 49, 9606–9608.
8 H. Tateishi-Karimata, M. Nakano and N. Sugimoto, Sci. Rep.,2014, 4, 3593.
9 R. Vijayaraghavan, A. Izgorodin, V. Ganesh,M. Surianarayanan and D. R. MacFarlane, Angew. Chem.,Int. Ed., 2010, 49, 1631–1633.
10 C. Zhao, J. Ren and X. Qu, Langmuir, 2013, 29, 1183–1191.11 I. Mamajanov, A. E. Engelhart, H. D. Bean and N. V. Hud,
Angew. Chem., Int. Ed., 2010, 49, 6310–6314.12 D. Constatinescu, C. Herrmann and H. Weingartner, Phys.
Chem. Chem. Phys., 2010, 12, 1756–1763.13 D. M. Foureau, R. M. Vrikkis, C. P. Jones, K. D. Weaver,
D. R. Macfarlane, J. C. Salo, I. H. McKillop andG. D. Elliott, Cell. Mol. Bioeng., 2012, 5, 390–401.
14 K. Fujita, D. R. MacFarlane and M. Forsyth, Chem. Commun.,2005, 4804–4806, DOI: 10.1039/b508238b.
15 K. Fujita, D. R. MacFarlane, M. Forsyth, M. Yoshizawa-Fujita,K. Murata, N. Nakamura and H. Ohno, Biomacromolecules,2007, 8, 2080–2086.
16 K. Fujita and H. Ohno, Biopolymers, 2010, 93, 1093–1099.17 R. M. Vrikkis, K. J. Fraser, K. Fujita, D. R. Macfarlane and
G. D. Elliott, J. Biomech. Eng., 2009, 131, 074514.18 K. D. Weaver, R. M. Vrikkis, M. P. Van Vorst, J. Trullinger,
R. Vijayaraghavan, D. M. Foureau, I. H. McKillop,D. R. MacFarlane, J. K. Krueger and G. D. Elliott, Phys.Chem. Chem. Phys., 2012, 14, 790–801.
43844 | RSC Adv., 2015, 5, 43839–43844
19 E. Melgar and D. A. Goldthwait, J. Biol. Chem., 1968, 243,4401–4408.
20 E. Melgar and D. A. Goldthwait, J. Biol. Chem., 1968, 243,4409–4416.
21 T. E. Haran and U. Mohanty, Q. Rev. Biophys., 2009, 42, 41–81.
22 R. Rohs, S. M. West, A. Sosinsky, P. Liu, R. S. Mann andB. Honig, Nature, 2009, 461, 1248–1253.
23 B. Nguyen, S. Neidle andW. D. Wilson, Acc. Chem. Res., 2009,42, 11–21.
24 M. Nakano, H. Tateishi-Karimata, S. Tanaka andN. Sugimoto, J. Phys. Chem. B, 2013, 118, 379–389.
25 H. Tateishi-Karimata and N. Sugimoto, Angew. Chem., Int.Ed., 2012, 51, 1416–1419.
26 A. F. Weir, in Enzymes of Molecular Biology, ed. M. Burrell,Humana Press, 1993, vol. 16, ch. 2, pp. 7–16.
27 C. A. Thomas, J. Am. Chem. Soc., 1956, 78, 1861–1868.28 E. T. Young 2nd and R. L. Sinsheimer, J. Biol. Chem., 1965,
240, 1274–1280.29 V. N. Schumaker, E. G. Richards and H. K. Schachman, J. Am.
Chem. Soc., 1956, 78, 4230–4236.30 F. W. Studier, J. Mol. Biol., 1965, 11, 373–390.31 G. Bernardi and C. Sadron, Biochemistry, 1964, 3, 1411–1418.32 J. D. Watson and F. H. C. Crick, Nature, 1953, 171, 737–738.33 C. A. Sprecher, W. A. Baase and W. C. Johnson, Biopolymers,
1979, 18, 1009–1019.34 J. Brahms andW. F. H. M.Mommaerts, J. Mol. Biol., 1964, 10,
73–88.35 A. Chandran, D. Ghoshdastidar and S. Senapati, J. Am. Chem.
Soc., 2012, 134, 20330–20339.36 J. Zhu, M. Lin, D. Fan, Z. Wu, Y. Chen, J. Zhang, Y. Lu and
Z. Guo, Dalton Trans., 2009, 10889–10895, DOI: 10.1039/b913236h.
37 F. N. Al-Deen, J. Ho, C. Selomulya, C. Ma and R. Coppel,Langmuir, 2011, 27, 3703–3712.
This journal is © The Royal Society of Chemistry 2015