elevation of plasminogen activators in cerebrospinal fluid of mice with eosinophilic meningitis...
TRANSCRIPT

Elevation of plasminogen activators in cerebrospinal fluid of mice with
eosinophilic meningitis caused by Angiostrongylus cantonensis
Roger F. Houa, Wu-Chun Tua, Hsiu-Hsiung Leeb, Ke-Min Chenb,Hui-Lin Choub, Shih-Chan Laib,*
aDepartment of Entomology, National Chung-Hsing University, Taichung 402, Taiwan, ROCbDepartment of Parasitology, Chung Shan Medical University, 110, Section 1, Chien-Kuo North Road, Taichung 402, Taiwan, ROC
Received 23 July 2004; received in revised form 25 August 2004; accepted 27 August 2004
Abstract
A hallmark of parasitic meningitis is the infiltration of eosinophils into the subarachnoid space. Infection with Angiostrongylus
cantonensis in mice induced proteinase activity in parallel with the pathological changes of eosinophilic meningitis. Zymogram analysis
demonstrated that 70 and 55 kDa proteinases from cerebrospinal fluid (CSF) were active against the casein/plasminogen substrate. The
proteinase activities were clearly inhibited by phenylmethanesulphonyl fluoride but not by ethylenediamine tetraacetic acid, 1,10-
phenanthroline or leupeptin. Western blotting confirmed these enzymes to be tissue-type plasminogen activator and urokinase-type
plasminogen activator, respectively. High activities of tissue-type plasminogen activator and urokinase-type plasminogen activator were
detected in the CSF of mice with eosinophilic meningitis, and correlated positively with CSF eosinophil numbers and total protein,
respectively. Immunohistochemistry demonstrated that tissue-type plasminogen activator and urokinase-type plasminogen activator
localised in the endothelial cells of blood vessels, in blood clots and in infiltrated leukocytes. These results suggest that tissue-type
plasminogen activator and urokinase-type plasminogen activator may be play a role in the pathogenesis of eosinophilic meningitis of
angiostrongyliasis.
q 2004 Australian Society for Parasitology Inc. Published by Elsevier Ltd. All rights reserved.
Keywords: Angiostrongylus cantonensis; Eosinophilic meningitis; Plasminogen activator; Proteinase; Blood–brain barrier
1. Introduction
Mature adults of the zoonotic parasitic nematode
Angiostrongylus cantonensis reside in the pulmonary
arteries of the permissive hosts (rats) (Alicata and Jindrak,
1970). However, in non-permissive hosts (humans and
mice), the immature adults remain in the central nervous
system (CNS) of the host, this infection being the main
cause of eosinophilic meningitis and eosinophilic menin-
goencephalitis (Hsu et al., 1990; Ismail and Arsura, 1993).
In mice infected with A. cantonensis, the cerebrospinal fluid
(CSF) eosinophilia reaches a peak at around 3 weeks and
parallels the pathogenesis of eosinophilic meningitis
0020-7519/$30.00 q 2004 Australian Society for Parasitology Inc. Published by
doi:10.1016/j.ijpara.2004.08.010
* Corresponding author. Tel.: C886 4 2473 0022/1641; fax: C886 4 238
23381.
E-mail address: [email protected] (S.-C. Lai).
(Sugaya and Yoshimura, 1988; Sasaki et al., 1993). The
blood–brain barrier (BBB) serves to protect the CNS from
invasive agents, such as inflammatory cells and bacteria, as
well as from chemical agents. Elevation of CSF total protein
indicates damage to the BBB (Fryden et al., 1978) and such
an elevation has been reported in angiostrongylosis (Yii,
1976; Wan and Weng, 2004).
Plasminogen activators (PAs) are serine proteases that
convert the zymogen, plasminogen, into the active serine
protease, plasmin. There are two types—tissue-type PA
(tPA) and urokinase-type PA (uPA) (Vassalli et al., 1991).
In normal plasma and in tissue, they are inactive and
complexed to plasminogen activator inhibitors, of which
type 1 plasminogen activator inhibitor (PAI-1) is believed to
be the most important (Vassalli et al., 1991; Loskutoff et al.,
1993; Blasi, 1997). It is well known that tPA plays a primary
role in the plasmin generation required for fibrinolysis,
International Journal for Parasitology 34 (2004) 1355–1364
www.parasitology-online.com
Elsevier Ltd. All rights reserved.

R.F. Hou et al. / International Journal for Parasitology 34 (2004) 1355–13641356
including clot or thrombus lysis. It also promotes BBB
disruption and is involved in the pathophysiology of
bacterial meningitis (Busch et al., 1997). uPA is primarily
involved in cell surface proteolysis and, thus, is important in
extracellular matrix (ECM) degradation and cell invasion
(Blasi et al., 1987). Additionally, the uPA system has the
capacity to promote leukocyte recruitment and BBB
breakdown, and thus may play an important pathophysio-
logical role in bacterial meningitis (Winkler et al., 2002).
The induction of PAs in bacterial meningitis is well
known. However, the relationship between PAs and
parasitic meningitis is still unknown. The current study,
therefore, set out to measure the activity of tPA and uPA in
the CSF in A. cantonensis-infected mice, and to investigate
the correlation between eosinophilic meningitis and PAs in
angiostrongylosis.
2. Materials and methods
2.1. Experimental animals
Five-week-old male mice, BALB/c strain, were pur-
chased from the National Laboratory Animal Center,
Taipei, Taiwan. Mice were maintained at a 12 h light/dark
cycle photoperiod, provided with Purina Laboratory Chow
and water ad libitum, and kept in our laboratory for more
than 1 week before the experimental infection.
2.2. Larval preparation
L3 (infective) larvae of A. cantonensis were obtained
from naturally infected giant African snails, Achatina fulica,
collected from fields in Pingtung County, southern Taiwan.
The larvae within tissues were recovered using the method
of Parsons and Grieve (1990) with slight modifications.
Briefly, the shells were crushed, the tissues were homogen-
ised and digested in a pepsin–HCl solution (pH 1–2,
500 I.U. pepsin/g tissue), and incubated with agitation at
37 8C in a waterbath for 2 h. Host cellular debris was
removed from the digest by centrifugation at 1400 g for
10 min. The larvae in the sediment were observed under the
microscope. The morphological criteria for identification of
the L3 of A. cantonensis 425–524 mm in length and from 23
to 34 mm in width. The posterior end of the tail always
terminates in a fine point were provided by Ash (1970). To
confirm that the larvae found were A. cantonensis, 60 L3
were fed to five rats and then examined their brains (two
rats) 2–3 weeks later for evidence of infection. The other
rats were killed 5–6 weeks later they were found to harbour
the adults in their pulmonary arteries. The morphology of
the adult worms was consistent with that described for
A. cantonensis. The males measured 14–15 mm in length,
the tail with copulatory bursa and long spicules; females
24–26 mm in length, with characteristic barber-pole appear-
ance (Lindo et al., 2002).
2.3. Animal infection
A total of 90 male mice were randomly allocated to six
groups (D0, D5, D10, D15, D20, and D25) of 15 mice each.
They were prohibited food and water for 12 h before
infection. The mice of experimental groups (D5, D10, D15,
D20, and D25) were infected with 60 A. cantonensis larvae
by oral inoculation on day 0 and the groups sacrificed on
days 5, 10, 15, 20, and 25 p.i., respectively. The control
mice (D0) received only water and sacrificed on day 25 p.i.
The mice were sacrified by cervical dislocation, and the
brains and CSF samples were rapidly collected and frozen at
K70 8C before use.
2.4. Casein/plasminogen zymography
The CSF was centrifugated at 12,000 g for 10 min to
remove debris. The protein contents of supernatants were
loaded on 7.5% (mass/volume) SDS-polyacrylamide gels
that had been co-polymerised with 0.1% casein (Sigma,
USA) for plasmin activities, and plasminogen (13 mg/ml,
American Diagnostica) for PAs activities. Stacking gels
were 4% (mass/volume) polyacrylamide and did not contain
casein and plasminogen substrate. Electrophoresis was
performed in running buffer (25 mM Tris, 250 mM glycine,
1% SDS) at room temperature at 120 V for 1 h. The gel was
washed two times at room temperature for 30 min each in
2.5% Triton X-100, and then washed two times with double
distilled H2O for 10 min each. The gel was incubated in
reaction buffer (50 mM Tris–HCl, pH 8.0, containing
10 mM CaCl2, 0.01% NaN3) at 37 8C for 18 h. The gel
was stained with 0.25% Coomassie Brilliant Blue R-250
(Sigma, USA) for 1 h and destained in 15% methanol/7.5%
acetic acid. PAs activities were detected as unstained bands
on a blue background. Quantitative analysis of these
caseinolytic enzymes were performed with a computer-
assisted imaging densitometer system, UN-SCAN-ITe gel
Version 5.1 (Silk Scientific, USA).
2.5. Inhibition of proteinases on casein zymography
To explore the effects of various potential inhibitors on
the caseinolytic activities in the CSF samples, the samples
were run on SDS-polyacrylamide gels as described above.
Following electrophoresis, gels were soaked in 2.5% Triton-
X-100 to replace SDS, washed twice with water, then
incubated at 37 8C for 18 h in activation buffer (50 mM Tris,
pH 8.0, 10 mM CaCl2). For inhibitor studies, 10 mM
ethylenediamine tetraacetic acid (EDTA; Sigma, USA),
20 mM leupeptin (Sigma, USA), or 2 mM phenylmethane-
sulphonyl fluoride (PMSF; Sigma, USA), or 5 mM 1,10-
phenanthroline (Sigma, USA), was added to the Triton and
activation buffers. Zymography gels were stained with
Coomassie Brilliant Blue and destained in 15% metha-
nol/7.5% acetic acid. Proteins with casein activity were
revealed as clear bands on a blue background.

R.F. Hou et al. / International Journal for Parasitology 34 (2004) 1355–1364 1357
2.6. Western blot analysis
The CSF was centrifugated at 12,000 g for 10 min to
remove debris. The protein contents of supernatants were
determined with protein assay kits (Bio-Rad, USA) using
bovine serum albumin (BSA) as the standard. An equal
volume of loading buffer (62.5 mM Tris–HCl, pH 6.8, 10%
glycerol, 2% SDS, 5% 2-mercaptoethanol, 0.05% bromo-
phenol blue) was added to the samples, which contained
30 mg of brain tissue protein. The mixture was boiled for
5 min prior to electrophoresis on SDS-polyacrylamide gel
and electrotransferred to nitrocellulose membrane at a
constant current of 190 mA for 90 min. Afterwards, the
membrane was saturated with phosphate buffered saline
(PBS) containing 0.1% Tween 20 for 30 min at room
temperature. The membrane was allowed to react with
rabbit anti-mouse tPA and uPA polyclonal antibodies
(American Diagnostica, USA) diluted 1:100 at 37 8C for
1 h. Then, the membrane was washed three times with PBS
containing 0.1% Tween 20 (PBS-T), followed by incubation
with horseradish peroxidase (HRP)-conjugated goat anti-
rabbit IgG (Jackson ImmunoResearch Laboratories, USA)
diluted 1:5000 at 37 8C for 1 h to detect the bound primary
antibody. The reactive protein was detected by enhanced
chemiluminescence (Amersham, UK). To confirm equival-
ent protein loading, membranes were stripped by incubation
in 62.5 mM of Tris–HCl (pH 6.8), 2% SDS, and 100 mM 2-
mercaptoethanol at 55 8C, subsequently washed with PBS-
T, and reprobed with anti-b-actin antibody (dilution 1:500;
Sigma, USA).
2.7. Reverse transcriptase polymerase chain reaction
(RT-PCR) analysis
The CSF was centrifugated at 12,000 g for 10 min and
the liquid removed. Total RNA was isolated from the cell
pellets using Trizol reagent (Invitrogen, USA), according to
the manufacturer’s instructions. One microgram of total
RNA was used for first strand cDNA synthesis in 20 ml of
reaction volume using 50 units of Superscripte II reverse
transcriptase (Invitrogen, USA). PCR was performed under
standard conditions using Taq DNA polymerase (Invitro-
gen, USA) and primers. Forward (5 0–3 0) and reverse (5 0–3 0)
primers, respectively, were 5 0-GACATCAAGAAGGT-
GGTGAAGC-3 0 and 5 0-TGTCATTGAGAGCAATGC-
CAGC-3 0 for glyceraldehyde-3-phosphate dehydrogenase
(GAPDH), GGGAGGTTCAGAAGAGGAGCCCGG-3 0
and 5 0-GCGTTTCCCTACAAATCCATCAGGG-3 0 for
tPA (de Vries et al., 1995), 5 0-TGCCCAAGGAAATTC-
TGCCCAAGGAAATTCCAGGG-3 0 and 5 0-GCCAAT-
CTGCACATAGCACC-3 0 for uPA (de Vries et al., 1995),
5 0-CACAAGTCTGATGGCAGCAC-3 0 and 5 0-CAGG-
CATGCCCAACTTCTC-3 0 for PAI-1 (Yamamoto and
Loskutoff, 1996). PCR cycling conditions for GAPDH,
tPA, uPA and PAI-1 were denaturation at 94 8C for 45 s,
annealing at 55 8C for 1 min, primer extension at 72 8C for
2 min, and then holding at 4 8C; this was repeated for 30
cycles for tPA, uPA and PAI-1; 25 cycles for GAPDH. Ten
microlitre of the amplified product were then subjected to
electrophoresis in 1% agarose gels containing 20 mg/ml
ethidium bromide in Tris borate-EDTA buffer. Gels
were visualised on a UV transilluminator (Taiwan), and
digital images were taken using DGIS-5 Digital Gel
Image System (Taiwan). Quantitative analysis was per-
formed with a computer-assisted imaging densitometer
system, UN-SCAN-ITe gel Version 5.1 (Silk Scientific,
USA).
2.8. Cell counts in the CSF
The mice were sacrificed and their brains removed into a
35 mm dish. The cranial cavity and cerebral ventricles
(lateral, third and fourth ventricles) were rinsed with 1 ml
PBS each. The washing solution was collected into a
centrifuge to spin at 400g for 10 min. The resultant
sediments were then resuspended with 30 ml PBS from
each mouse for enumerating a total number of leukocytes on
hemacytometer. The differential cell count was assessed
with Wright–Giemsa staining (Sigma, Taufkirchen,
Germany) in 3 ml/smear. The percentages of eosinophils
were determined in 200 leukocytes/smear.
2.9. The measurement of CSF total protein
The CSF was centrifuged at 12,000 g at 4 8C for 10 min,
and the protein contents of the supernatants were deter-
mined with protein assay kits (Bio-Rad, USA) using BSA as
the standard. Protein concentration was determined by
absorbencies at 595 nm using a HITACHI U1100 spectro-
photometer (Japan).
2.10. Histology
The mouse brains were fixed separately in 10% neutral
buffered formalin for 24 h. The fixed specimens were
dehydrated in a graded ethanol series (50, 75, and 100%)
and xylene, then embedded in paraffin at 55 8C for 24 h.
Several serial sections were cut at a 5 mm thickness for each
organ from each mouse. Sections were deparaffinised,
stained with H&E using standard techniques and examined
under a light microscope.
2.11. Scanning electron microscopy
The mouse brains were fixed in 2.5% glutaraldehyde
(Electron Microscopy Science, USA) in 0.15 M PBS buffer,
pH 7.4, for 3 h at 4 8C, and post-fixed in 1% osmium
tetroxide (Electron Microscopy Science, USA) in the same
buffer for 1 h at 4 8C. The fixed specimens were dehydrated
in a graded ethanol series (30–100%) and dried in LADD
28000 critical point dryer (USA). The dried specimens were
mounted on stubs, coated with 20 nm gold in JBS E5150

R.F. Hou et al. / International Journal for Parasitology 34 (2004) 1355–13641358
sputter coater (UK), and photographed with a TOPCON
ABT-150S scanning electron microscope (Japan).
2.12. Transmission electron microscopy
The mouse brains were fixed in 2.5% glutaraldehyde
(Electron Microscopy Science, USA) in 0.15 M PBS buffer,
pH 7.4, for 3 h at 4 8C, and post-fixed in 1% osmium
tetroxide (Electron Microscopy Science, USA) in the same
buffer for 1 h at 4 8C. The fixed specimens were dehydrated
in a graded ethanol series (30–100%), and embedded in LR
White resin (Spi Supplies, USA) for 24 h at 54 8C. Ultrathin
sections were cut with an ultramicrotome (Reichert Ultracut
S, Austria) and were doubly stained in 2% uranyl acetate
(Merck, Germany) for 30 min and 1% lead citrate (Merck,
Germany) for 12 min. The sections were examined and
photographed using a 100 kV electron microscope (JOEL
1200 EX II, Japan).
2.13. Immunohistochemistry
The mouse brains were fixed separately in 10% neutral
buffered formalin for 24 h. The fixed specimens were
dehydrated in a graded ethanol series (50, 75, and 100%)
and xylene, then embedded in paraffin at 55 8C for 24 h. Ten
micrograms of paraffin-embedded sections were prepared
and mounted on glass slides. Serial sections were depar-
affinised with xylene and a graded series of ethanol.
Sections were treated with 3% H2O2 in methanol for
10 min to inativate endogenous peroxidase, and washed
three times with PBS, pH 7.4 for 5 min. Sections were
blocked non-specific reactions with 3% BSA at room
temperature for 1 h, incubated with primary antibodies
(rabbit anti-mouse tPA and uPA polyclonal antibodies;
American Diagnostica, USA) diluted 1:50 in 1% BSA at
37 8C for 1 h, and washed three times in PBS for 5 min each.
Sections were incubated with HRP-conjugated goat anti-
rabbit IgG (Jackson ImmunoResearch Laboratories, USA)
diluted 1:100 in 1% BSA at 37 8C for 1 h, and washed three
times in PBS for 5 min each. Sections were incubated in
DAB (3,3 0-diaminobenzidine; 0.3 mg/ml in 100 mM Tris
pH 7.5 containing 0.3 ml H2O2/ml) at room temperature for
3 min, and washed three times in PBS for 5 min each.
Mounted slides with 50% glycerol in PBS were examined
under a light microscope.
Fig. 1. Time-course studies for caseinolytic activity from CSF. (a) The
molecular mass 70 kDa bands were detected at all time points, and the
intensity increased gradually from days 5 to 25 p.i. The 55 kDa bands were
detected on day 10 p.i. and reached a high intensity from days 15 to 25 p.i.
but were undetectable in the uninfected control. (b) Quantitative analysis of
the proteolytic enzyme was performed with a computer-assisted imaging
densitometer system. The relative intensity of the bands in Angiostrongylus
cantonensis-infected mice showed significant increase (*P!0.05) com-
pared with uninfected control.
2.14. Statistical analysis
Results in the different groups of mice were compared
using the non-parametric Kruskal–Wallis test followed by
post-testing using Dunn’s multiple comparison of means.
Correlations between CSF laboratory parameters and PAs
were quantified using the Spearman’s ranking correlation
test. The best fitting regression curve was drawn using
Microsoftw Excel 2000 analysis software. All results were
presented as meanGstandard deviation (SD). P values of
!0.05 were considered statistically significant.
3. Results
3.1. Time-course studies for caseinolytic activity from CSF
Bands corresponding to 70 kDa were detected at all time
points tested, including in uninfected mice, and the intensity
increased gradually from days 5 to 25 p.i. 55 kDa bands
were detected on day 10 p.i. and reached a high intensity
from days 15 to 25 p.i. An increased activity of PAs was
observed in mice with meningitis (Fig. 1a). The relative
activity of PAs in A. cantonensis-infected mice showed a
significant increase (P!0.05) compared with uninfected
controls (Fig. 1b).
3.2. Identification of the proteinases
Casein/plasminogen zymography on day 20 p.i. showed
70 and 55 kDa proteinases present in mice infected with
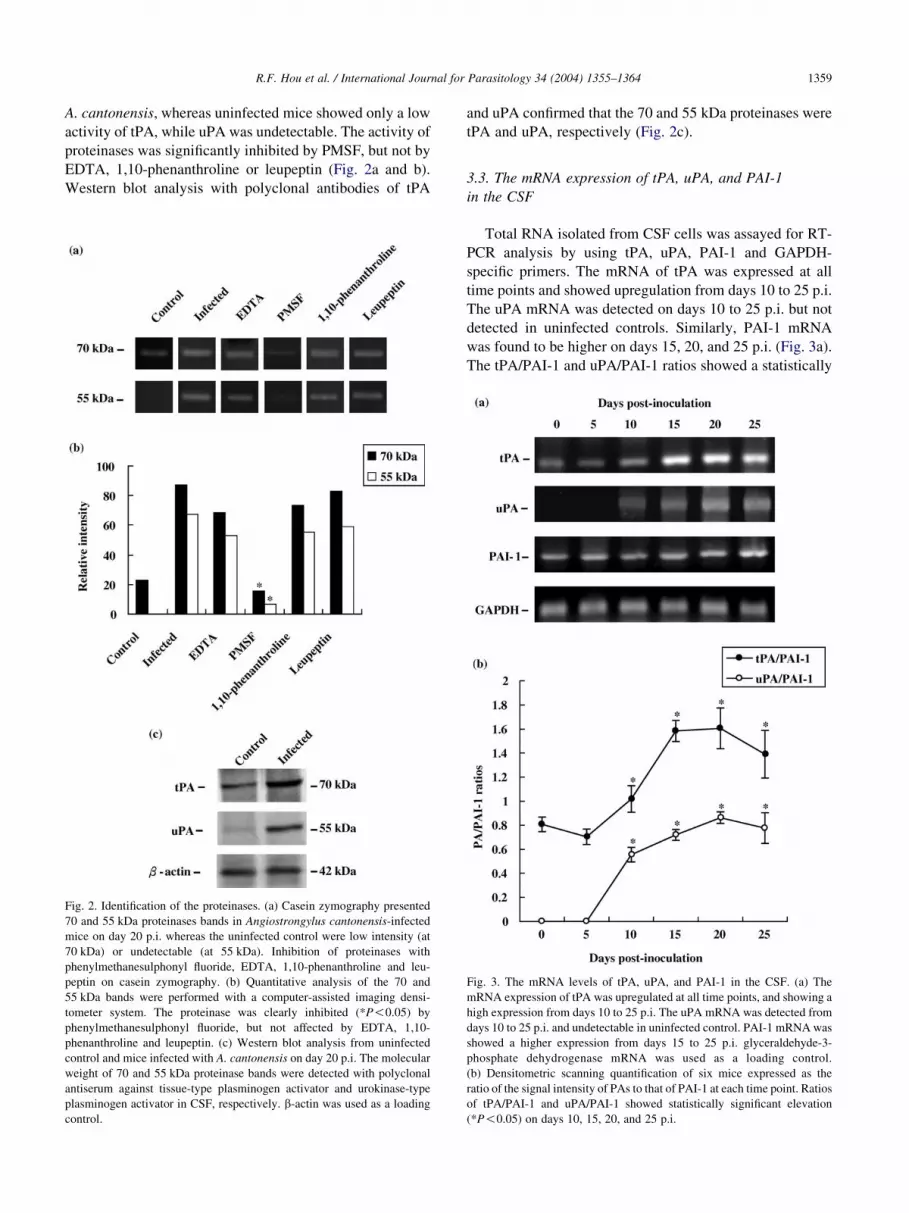
R.F. Hou et al. / International Journal for Parasitology 34 (2004) 1355–1364 1359
A. cantonensis, whereas uninfected mice showed only a low
activity of tPA, while uPA was undetectable. The activity of
proteinases was significantly inhibited by PMSF, but not by
EDTA, 1,10-phenanthroline or leupeptin (Fig. 2a and b).
Western blot analysis with polyclonal antibodies of tPA
Fig. 2. Identification of the proteinases. (a) Casein zymography presented
70 and 55 kDa proteinases bands in Angiostrongylus cantonensis-infected
mice on day 20 p.i. whereas the uninfected control were low intensity (at
70 kDa) or undetectable (at 55 kDa). Inhibition of proteinases with
phenylmethanesulphonyl fluoride, EDTA, 1,10-phenanthroline and leu-
peptin on casein zymography. (b) Quantitative analysis of the 70 and
55 kDa bands were performed with a computer-assisted imaging densi-
tometer system. The proteinase was clearly inhibited (*P!0.05) by
phenylmethanesulphonyl fluoride, but not affected by EDTA, 1,10-
phenanthroline and leupeptin. (c) Western blot analysis from uninfected
control and mice infected with A. cantonensis on day 20 p.i. The molecular
weight of 70 and 55 kDa proteinase bands were detected with polyclonal
antiserum against tissue-type plasminogen activator and urokinase-type
plasminogen activator in CSF, respectively. b-actin was used as a loading
control.
and uPA confirmed that the 70 and 55 kDa proteinases were
tPA and uPA, respectively (Fig. 2c).
3.3. The mRNA expression of tPA, uPA, and PAI-1
in the CSF
Total RNA isolated from CSF cells was assayed for RT-
PCR analysis by using tPA, uPA, PAI-1 and GAPDH-
specific primers. The mRNA of tPA was expressed at all
time points and showed upregulation from days 10 to 25 p.i.
The uPA mRNA was detected on days 10 to 25 p.i. but not
detected in uninfected controls. Similarly, PAI-1 mRNA
was found to be higher on days 15, 20, and 25 p.i. (Fig. 3a).
The tPA/PAI-1 and uPA/PAI-1 ratios showed a statistically
Fig. 3. The mRNA levels of tPA, uPA, and PAI-1 in the CSF. (a) The
mRNA expression of tPA was upregulated at all time points, and showing a
high expression from days 10 to 25 p.i. The uPA mRNA was detected from
days 10 to 25 p.i. and undetectable in uninfected control. PAI-1 mRNA was
showed a higher expression from days 15 to 25 p.i. glyceraldehyde-3-
phosphate dehydrogenase mRNA was used as a loading control.
(b) Densitometric scanning quantification of six mice expressed as the
ratio of the signal intensity of PAs to that of PAI-1 at each time point. Ratios
of tPA/PAI-1 and uPA/PAI-1 showed statistically significant elevation
(*P!0.05) on days 10, 15, 20, and 25 p.i.

R.F. Hou et al. / International Journal for Parasitology 34 (2004) 1355–13641360
significant elevation (P!0.05) on days 10, 15, 20, and 25
p.i. (Fig. 3b).
3.4. Correlation of CSF eosinophilia with tPA and uPA
Only infected mice showed CSF pleocytosis. The
leukocytes were identified as eosinophils cells by
Wright–Giemsa staining. The time-course studies showed
a mild eosinophilia on day 10 p.i. and a plateau response
from days 15 to 25 p.i. Using Spearman’s ranking
correlation test, the CSF eosinophilia showed a significant
correlation (P!0.05) with the activity of tPA (rZ0.93)
(Fig. 4a), and uPA (rZ0.91) (Fig. 4b). The relation was
best fitted using a regression curve, and the elative
intensity of PAs reached a plateau at 15–37% eosinophils
in CSF.
Fig. 4. Correlation of CSF eosinophil with tPA and uPA. The percentages of
CSF eosinophil significant correlated (*P!0.05) with the intensity of tPA
(a), and uPA (b) using the Spearman’s ranking correlation test.
3.5. Correlation of CSF total protein with tPA and uPA
The appearance of plasma proteins in CSF is a hallmark
of numerous CNS disorders with presumed or overt BBB
disruption. In this experimental eosinophilic meningitis of
angiostrongyliasis, CSF total protein significantly correlated
(P!0.05) with the activity of tPA (rZ0.81) (Fig. 5a), and
uPA (rZ0.82) (Fig. 5b) by Spearman’s ranking correlation
test. The relation was best fitted using a regression curve,
and the elative intensity of PAs reached a plateau at the total
protein concentration of 0.75–1.7 mg/ml.
3.6. Histopathological observations in
the subarachnoid space
In brain sections stained with haematoxylin and
eosin, uninfected mice had no inflammatory cells in
Fig. 5. Correlation of CSF total protein with tPA and uPA. The CSF total
protein significant correlated (*P!0.05) with the intensity of tPA (a), and
uPA (b) using the Spearman’s ranking correlation test.

Fig. 6. Histopathological observations in the subarachnoid space. (a) H&E stain showing severe inflammatory reaction (arrowheads) and hemorrhage (arrow).
C, cortex. (b) Enlargement of the portion shown in rectangle of (a). Inflammatory reaction consisting of polymorphonuclear (arrowhead) and mononuclear
(arrow) leukocytes. H, hemorrhage. (c) SEM showing the inflammatory cells (arrowheads) accumulate on the brain surface. (d) Red blood cells (arrowheads)
and inflammatory cells (arrows) aggregated on the brain surface by SEM. (e) Ultrastructural observations showing red blood cells (R) and lymphocytes (L)
aggregated in the subarachnoid space. E, endothelial cell of meninge. (f) TEM showing eosinophils with a bi-lobed nucleus (N) containing condensed
chromatin and the cytoplasm packed with many large, membrane-enclosed, dense crystalloid-containing ovoid granules (arrowheads).
R.F. Hou et al. / International Journal for Parasitology 34 (2004) 1355–1364 1361
the subarachnoid space and the meninges were normal (data
not shown). A gradual increase in pathological effects after
infection culminated in a severe infiltration of leukocytes,
edema and hemorrhage from days 15 to 25 p.i. Inflamma-
tory reaction consisting of polymorphonuclear and mono-
nuclear leukocytes were observed in the brain tissue on day
20 p.i. (Fig. 6a and b). Scanning electron micrographs
showed red blood cells and inflammatory cells accumulated
on the brain surface (Fig. 6c and d). Ultrastructural
observations showed red blood cells and eosinophils
aggregated on the subarachnoid space. The eosinophils
showed many crystalloid-containing secretory granules in
the cytoplasm (Fig. 6e and f).
3.7. Distribution of tPA and uPA in the subarachnoid space
Positive signals for tPA (Fig. 7a and b) and for uPA
(Fig. 7c and d) were localised in the endothelial cells of
blood vessels, in blood clot and in infiltrated polymorpho-
nuclear and mononuclear cells. No positive signal was
detected in these structures in uninfected mice (Fig. 7e).
4. Discussion
The activities of serine proteinases increased is associ-
ation with the inflammatory disease (Tarlton et al., 2000)

Fig. 7. Immunohistochemical distribution of tPA and uPA in the subarachnoid space. (a) tPA localised in endothelial cells (arrowhead) of blood vessel, in blood
clot (C) and in infiltrated leukocytes (arrows), and presented brown colour. (b) Enlargement of the portion shown in rectangle of (a). Polymorphonuclear
(arrowheads) and mononuclear (arrow) cells presented positive signal for tPA. (c) uPA localised brown colour in blood clot (C) and infiltrated leukocytes
(arrowheads). (d) The endothelial cells (arrowhead) and in infiltrated leukocytes (arrow) contained a positive signal for uPA. (e) No positive signal (brown
colour) could be detected with normal serum in the blood clot (C) nor in infiltrating leukocytes (arrowheads).
R.F. Hou et al. / International Journal for Parasitology 34 (2004) 1355–13641362
and meningitis (Winkler et al., 2002). Meningitis may be
caused by viruses or bacteria and less often by other
pathogens, such as rickettsia, fungi, and parasites (Zhang
and Tuomanen, 1999; Casadevall and Pirofski, 2000).
Angiostrongylosis of the meninges is a chronic meningitis
characterised by the aggregation of eosinophils in
the subarachnoid space (Reid and Wallis, 1984). The
distinct expression profiles found in the present study
indicate a role for uPA and tPA in the pathogenesis of
parasitic meningitis. The role of exaggerated extracellular

R.F. Hou et al. / International Journal for Parasitology 34 (2004) 1355–1364 1363
proteolysis in the CNS caused by PAs was further
strengthened by the high correlation between PAs and
eosinophil counts in the CSF.
Proteolysis becomes pathological when an imbalance
between proteinases and their inhibitors occurs. PAI-1,
which binds to and inactivates both tPA and uPA, is the
primary regulator of plasminogen activation in vivo
(Vassalli et al., 1991). Fibrinolysis and coagulation in
patients with infectious disease and sepsis showed that the
uPA level is markedly increased, but concomitant marked
PAI-1 upregulation (Philippe et al., 1991; Robbie et al.,
2000). Similarly, the present study showed that an
imbalance between tPA and PAI-1 and between uPA and
PAI-1 may be associated with eosinophilic pleocytosis in
the subarachnoid space in angiostrongylosis. Additionally,
the mRNA expression of the PAs coincided with proteolytic
activity, suggesting that increased PAs activity may be
transcriptionally regulated.
The breakdown of BBB is regarded as an important
pathophysiological event in bacterial meningitis. It causes
extravasation of different neurotoxic factors and results in
brain edema with consequent increased intracranial pressure
(Leib and Tauber, 1999). A possible role for uPA in BBB
breakdown was also found in a mouse model of brain
trauma, which showed that uPA deficiency resulted in
decreased extravasation of proteins into the CNS (Kataoka
et al., 2000). In the present study, the time of increase in PAs
corresponds with the time of CSF eosinophilia and total
protein. Increased activities of PAs might threaten the
integrity of BBB and may thereby lead to BBB damage, and
increased influx of inflammatory cells into subarachnoid
space. Therefore, it is plausible to assume that increased PA
activity may promote eosinophilic meningitis by disruption
of the BBB.
tPA, uPA, and PAI-1 have been implicated in fibrin
formation or removal and each are regulated during
inflammatory/thrombotic events. tPA is synthesised by
endothelial cells in normal blood vessels (Kristensen
et al., 1984), and functions in physiological thrombolysis
in vivo (Collen and Lijnen, 1991). Studies in uPA knockout
mice indicated that uPA is also involved in fibrinolysis
(Carmeliet et al., 1994). Additionally, elevations in PAI-1
activity have been demonstrated in a number of clinical
conditions associated with a predisposition to thrombosis
(Tabernero et al., 1989). The present study showed that tPA
and uPA localise in the endothelial cells of blood vessels
and blood clot. These data suggest that the imbalance
between PAs and PAI-1 in angiostrongyliasis may facilitate
cellular infiltration into the subarachnoid space and
thrombolysis.
Acknowledgements
We wish to thank Y.S. Lin and P.C. Chao, the
Instrumentation Center, National Chung Hsing University,
for technical assistance in electron microscopy. This study
was supported by a research grant NO. NSC 92-2314-B-
040-027 from the National Science Council, ROC.
References
Alicata, J.E., Jindrak, K., 1970. Angiostrongylosis in the Pacific and
Southeast Asia. Thomas, Springfield, IL.
Ash, L.R., 1970. Diagnostic morphology of the third-stage larvae of
Angiostrongylus cantonensis, Angiostrongylus vasorum, Aelurostron-
gylus abstrusus, and Anafilaroides rostratus (Nematoda: Metastrongy-
loidea). J. Parasitol. 56, 249–253.
Blasi, F., 1997. uPA, uPAR, PAI-1: key intersection of proteolytic,
adhesive and chemotacfic highways? Review. Immunol. Today 18,
415–417.
Blasi, F., Vassalli, J.D., Danø, K., 1987. Urokinase-type plasminogen
activator: proenzyme, receptor, and inhibitors. J. Cell Biol. 104,
801–804.
Busch, E., Kruger, K., Fritze, K., Allegrini, P.R., Hoehn-Berlage, M.,
Hossmann, K.A., 1997. Blood–brain barrier disturbances after rt-PA
treatment of thromboembolic stroke in the rat. Acta Neurochir. Suppl.
70, 206–208.
Carmeliet, P., Schoonjans, L., Kieckens, L., Ream, B., Degen, J.,
Bronson, R., de Vos, R., van den Oord, J.J., Collen, D.,
Mulligan, R.C., 1994. Physiological consequences of loss of plasmino-
gen activator gene function in mice. Nature 368, 419–424.
Casadevall, A., Pirofski, L.A., 2000. Host-pathogen interactions: basic
concepts of microbial commensalism, colonisation, infection, and
disease. Infect. Immun. 68, 6511–6518.
Collen, D., Lijnen, H.R., 1991. Basic and clinical aspects of fibrinolysis and
thrombolysis. Blood 78, 3114–3124.
de Vries, T.J., Kitson, J.L., Silvers, W.K., Mintz, B., 1995. Expression of
plasminogen activators and plasminogen activator inhibitors in
cutaneous melanomas of transgenic melanoma-susceptible mice.
Cancer Res. 55, 4681–4687.
Fryden, A., Link, H., Norrby, E., 1978. Cerebrospinal fluid and serum
immunoglobulins and antibody titers in mumps meningitis and aseptic
meningitis of other etiology. Infect. Immun. 21, 852–861.
Hsu, W.Y., Chen, J.Y., Chien, C.T., Chi, C.S., Han, N.T., 1990.
Eosinophilic meningitis caused by Angiostrongylus cantonensis.
Pediatr. Infect. Dis. J. 9, 443–445.
Ismail, Y., Arsura, E.L., 1993. Eosinophilic meningitis. West J. Med.
159, 623.
Kataoka, K., Asai, T., Taneda, M., Ueshima, S., Matsuo, O., Kuroda, R.,
Kawabata, A., Carmeliet, P., 2000. Roles of urokinase type plasmino-
gen activator in a brain stab wound. Brain Res. 887, 187–190.
Kristensen, P., Larsson, L.I., Nielsen, L.S., Grondahl-Hansen, J.,
Andreasen, P.A., Dano, K., 1984. Human endothelial cells contain
one type of plasminogen activator. Fed. Eur. Biochem. Soc. Lett. 168,
33–37.
Leib, S.L., Tauber, M.G., 1999. Pathogenesis of bacterial meningitis.
Infect. Dis. Clin. North. Am. 13, 527–548.
Lindo, J.F., Waugh, C., Hall, J., Cunningham-Myrie, C., Ashley, D.,
Eberhard, M.L., Sullivan, J.J., Bishop, H.S., Robinson, D.G., Holtz, T.,
Robinson, R.D., 2002. Enzootic Angiostrongylus cantonensis in rats
and snails after an outbreak of human eosinophilic meningitis, Jamaica.
Emerg. Infect. Dis. 8, 324–326.
Loskutoff, D.J., Sawdey, M., Keeton, M., Scheiderman, J., 1993.
Regulation of PAI-1 gene expression in vivo. Thromb. Haemost. 70,
135–137.
Parsons, J.C., Grieve, R.B., 1990. Effect of egg dosage and host genotype
on liver trapping in murine larval toxocariasis. J. Parasitol. 76, 53–58.

R.F. Hou et al. / International Journal for Parasitology 34 (2004) 1355–13641364
Philippe, J., Offner, F., Declerck, P.J., Leroux-Roels, G., Vogelaers, D.,
Baele, G., Collen, D., 1991. Fibrinolysis and coagulation in patients
with infectious disease and sepsis. Thromb. Haemost. 65, 291–295.
Reid, I.R., Wallis, W.E., 1984. The chronic and severe forms of
eosinophilic meningitis. Aust. N. Z. J. Med. 14, 163–165.
Robbie, L.A., Dummer, S., Booth, N.A., Adey, G.D., Bennett, B., 2000.
Plasminogen activator inhibitor 2 and urokinase-type plasminogen
activator in plasma and leucocytes in patients with severe sepsis. Br.
J. Haematol. 109, 342–348.
Sasaki, O., Sugaya, H., Ishida, K., Yoshimura, K., 1993. Ablation of
eosinophils with anti-IL-5 antibody enhances the survival of intracra-
nial worms of Angiostrongylus cantonensis in the mouse. Parasite
Immunol. 15, 349–354.
Sugaya, H., Yoshimura, K., 1988. T-cell-dependent eosinophilia in the
cerebrospinal fluid of the mouse infected with Angiostrongylus
cantonensis. Parasite Immunol. 10, 127–138.
Tabernero, M.D., Estelles, A., Vicente, V., Alberca, I., Aznar, J., 1989.
Incidence of increased plasminogen activator inhibitor in patients with
deep venous thrombosis and/or pulmonary embolism. Thromb. Res. 56,
565–570.
Tarlton, J.F., Whiting, C.V., Tunmore, D., Bregenholt, S., Reimann, J.,
Claesson, M.H., Bland, P.W., 1935. The role of up-regulated serine
proteases and matrix metalloproteinases in the pathogenesis of a murine
model of colitis. Am. J. Pathol. 157, 1927–1927.
Vassalli, J.D., Sappino, A.P., Belin, D., 1991. The plasminogen
activator/plasmin system. J. Clin. Invest. 88, 1067–1072.
Wan, K.S., Weng, W.C., 2004. Eosinophilic meningitis in a child raising
snails as pets. Acta Trop. 90, 51–53.
Winkler, F., Kastenbauer, S., Koedel, U., Pfister, H.W., 2002. Role of the
urokinase plasminogen activator system in patients with bacterial
meningitis. Neurology 59, 1350–1355.
Yamamoto, K., Loskutoff, D.J., 1996. Fibrin deposition in tissue from
endotoxin-treated mice correlates with decreases in the expression of
urokinase-type but not tissue-type plasminogen activator. J. Clin.
Invest. 97, 2440–2451.
Yii, C.Y., 1976. Clinical observations on eosinophilic meningitis and
meningoencephalitis caused by Angiostrongylus cantonensis on
Taiwan. Am. J. Trop. Med. Hyg. 25, 233–249.
Zhang, J.R., Tuomanen, E., 1999. Molecular and cellular mechanisms for
microbial entery into the CNS. J. Neurovirol. 5, 591–603.