electrodynamics of microtubular motors: the building blocks of a new model
TRANSCRIPT

BioSystems 39 (1996) 187-226
Electrodynamics of microtubular motors: the building blocks of a new model
Ezio M. Insinna*” Patrick Zaborskib Jack Tuszynski’ “Bioelectronics Research Association, It? allet? des F&es Lurk&e, 77600 Bussy SI Georges, France
bCentre National de la Recherche Scientifique, CNRS - UPR 420 Molec. Genet. & Dev. Biol., BP 8, 94801 Villeju(f Cede.x France ‘Department of Physics, University qf Alberta, Edmonton, Alberta, T6G 2JI Canada
Received 2 January 1996; revised 21 March 1996; accepted 21 March 1996
Abstract
Microtubules are ubiquitous components of the cytoskeleton. They participate in many motility processes ranging from intracellular transport or chromosome movement during mitosis to ciliary and flagellar beating. The biophysical mechanism inherent in the generation and control of movement in all these motility phenomena has not yet been entirely elucidated. The authors propose a new model based on a charge transfer mechanism capable of shedding a new light on the molecular foundations of all motility processes. Electron transfer along the microtubular lattice is responsible for activation and control of all microtubule-associated ATPases (i.e. force generating enzymes). Microtubules are thus shown to be the basic motors of cell dynamics. The model is first applied to intracellular transport and ciliary and flagellar beating. Through two additional examples, the authors show the heuristic capabilities of the suggested hypothesis. The application of charge transfer control to the Protozoan Euglena gracilis leads to a plausible model capable of accounting for its phototactic response mechanism. Furthermore, the model allows a new interpretation of the electrophysiological response in vertebrate photoreceptors.
Keywords: Microtubules; Cytoskeleton; Electron transfer; ATPase activation; Conformational changes; Ciliary beating; Intracellular transport; Protozoan; Euglena gradis; Vertebrate photoreceptors
1. Introduction
During the last two decades and through the extensive use of electron microscopy, the cyto- skeleton has been shown to possess a very com- plex dynamical structure. One of the major
* Corresponding author, Tel.: + 33 1 64 66 37 68; fax: + 33 1 64 66 37 86; e-mail: [email protected].
components of the cytoplasm, besides actin filaments, intermediate filaments and a network of tiny fibrils called the microtrabecular lattice (MTL) (Bershadsky, 1988) are microtubules (MT). MT are ubiquitous in the eukaryatic cells and contribute to most motility mechanisms within and outside the cell via intracellular trans- port (axoplasmic transport), ciliary and Bagellar movement, chromosome movement during mito-
0303-2647/96/$15.00 0 1996 Elsevier Science Ireland Ltd. All rights reserved PII SO303-2647(96)01616-4

188 E.M. Isinna et al. / BioSystems 39 (1996) 187-226
sis, etc. (Warner, 1974; Amos, 1976, 1979 and 1982; Allen, 1985a/b; for review, see Roberts, 1979; Dustin, 1984; Vale; 1987, 1993).
MT are polymeric hollow cylinders with a di- ameter of 25 nm and a variable length ranging from pm to several mm. The cylinder is usually made up of 13 linearly arranged protofilaments forming the MT wall lattice (Fig. 1).
The MT lattice is a polymer formed of dimers containing one c(- and one /?-tubulin monomer. The dimensions of each dimer are: 4 x 5 x 8 nm (Amos, 1976, 1979, 1982; Tucker, 1979). In the wall lattice, the dimers are arranged into helical patterns together with microtubule-associated proteins (MAPS). The latter contribute to both MT stability and mechanical rigidity (Amos, 1976).
MT either assemble (or disassemble) sponta- neously from a pool of tubulin dimers in the cytoplasm (labile MT), such as the mitotic spindle MT, or they are stable and are not replaced during the lifetime of the structure they belong to. The latter situation occurs e.g., in centrioles or flagella.
In vitro, nucleation of tubulin cr-8 het- erodimers into MT requires the presence of GTP (with subsequent hydrolysis of this nucleotide and production of GDP + inorganic phosphate), Mg’ + and low Ca2+ concentrations (for review see David-Pfeuty et al., 1977; Kirschner, 1978; Scheele, 1979; Weisenberg, 198 1; Dustin, 1984). The presence of microtubule associated proteins (MAPS) also seems (under certain conditions) to regulate MT assembly (Olmsted, 1986). In vitro, at critical free tubulin concentrations, elongation of the polymer slows down until a steady state is reached (dynamic equilibrium), wherein the lattice subunits are exchanged with the free pool of tubulins, to the extent that GTP is available.
MT growth exhibits a finite polarity, which means that polymerization of tubulin occurs pref- erentially at one end, called A, the net assembly end, or the + end. The rate of addition of dimers at the + end is several times greater than at the opposite D, the net disassembly end, or the - end, where tubulin dimers preferentially disassem- ble. Incorporated tubulin dimers have, under spe- cific conditions, been seen to wander from the +
to the - end (treadmilling) (Kirschner, 1980; Margolis, 198 1; Cleveland, 1982). Currently, in vivo observations have generated an instability model of MT dynamics: at low concentrations of GTP-bound tubulin dimers (below the steady state concentration), growing and shrinking MT populations co-exist and fluctuate between these two phases. The difference between phases lies in the presence or absence of a GTP-liganded cap at one end (the + end). The growing populations have a cap of GTP tubulins which stabilizes the entire structure, whereas the shrinking popula- tions miss such a cap. The length of the cap is a function of the free tubulin concentration. Such phases rarely interconvert (Mitchison and Kirschner, 1984; Horio and Hotani, 1986; Sam- mak et al., 1987).
The mechanism underlying the dynamics of MT and related microtubular structures is still an un- solved mystery. The present situation is probably due to the absence of a heuristic hypothesis capa- ble of opening the way to new experimental inves- tigations. Some suggestions concerning MT have already been made in connection with informa- tion processing and sensory transduction but they have, by and large, provided no practical predic- tion (Atema, 1973; Hameroff and Watt, 1982; Hameroff et al., 1984; Hameroff, 1987). One of the authors has already proposed a model based
dins
Fig. 1. Structure of a microtubule. The a- and /3- globular tubulins are assembled in the form of a hollow cylinder about 240 A in diameter. The structure also contains MAPS (micro- tubule associated proteins).

E.M. Isinna et al. / BioSystrms 39 (1996) 187-226 189
on a charge transfer mechanism and capable of explaining some dynamic properties of MT in flagellar movement (Insinna, 1989). However, during the last few years, it has become clear to the present authors that additional theoretical investigations and application examples are needed in order to find experimental evidence for the existence of the proposed charge transfer mechanism.
This paper presents a new theoretical approach in which charge transfer is hypothesized to occur through a dissipative mechanism involving GTP hydrolysis and a self-trapping mechanism some- what reminiscent of the Davydov model.
As will be shown in the following, the model can lead to a new view of dynamic processes inherent in microtubular complex structures and to the consideration of microtubular motors (MTM) as active electronic components of cell dynamics. The hypothesis of charge transfer oc- curring along the microtubular lattice helps un- derstand some functional specificities of MT dynamics, ranging from intracellular transport and ciliary or flagellar beating to vertebrate pho- toreceptors. In Part I of this paper we propose the charge transfer mechanism and apply it to intra- cellular transport and ciliary/flagellar beating. In Part II, we present two more examples of MT structures manifestly supporting the postulated model.
2. Part I - Theoretical considerations and general applications
2.1. Microtubule dynamics and charge transfer
2.1.1. Charge/electron trunsfer Ever since Szent-Gybrgyi predicted that biolog-
ical materials could have semiconductor proper- ties, electronic interactions such as charge and energy transfer between proteins have been recog- nized as viable at the biological level (Szent-Gybr- gyi, 1941, Pethig and Szent-Gyiirgyi, 1977, for review see Pethig, 1979). However, most investiga- tions have been concerned with electron transfer occurring in metalloproteins as cytochrome c (ox- idative phosphorylation) and photosynthesis
(Hopfield, 1974; Marcus and Sutin, 1985). The main problem, in biological systems, remains the existence of relatively wide energy gaps between proteins. This represents an insurmountable bar- rier for direct electron transfer at the temperature ranges and energy levels existing in living organ- isms. Several physical mechanisms have been pro- posed to overcome this problem, such as quantum mechanical tunnelling, electron superconductivity or soliton waves (Devault, Parkes and Chance, 1967; Cope, 1974; Miller; 1975, Davydov, 1982; Devault, 1984).
As for microtubules, few suggestions have been made with respect to the possibility of charge transfer within protein subunits. Hameroff and Watt have suggested that Ca2 + ions could greatly enhance charge and energy transfer between tubu- lin subunits by pulling electrons from their reso- nance orbitals in hydrophobic regions; the Ca’ + could act as acceptor ‘impurities’, creating ‘holes’, as in semiconductors (Hameroff and Watt, 1982; Hameroff et al., 1984).
Referring to the amino acids classification of Sneddon (1988), Koruga and Simic-Krstic (1990) advanced a hypothesis that for a- and ,0-tubulin, the relative amount of n-type amino acids (con- taining carbon impurities) is 2.5 times greater than that of p-type amino acids. Tubulins would, thus, have a surplus of electrons allowing for a charge transfer mechanism.
Hameroff et al. (1984) as well as Tuszynski et al. (1995) have also mentioned the possibility of the propagation of soliton waves in MT. These localized excitations are seen to arise from a coupling between tubulin’s conformational changes and electronic degrees of freedom. In fact, Davydov (1982), suggested that energy trans- fer (electron transfer) between donor-acceptor protein pairs can easily be explained by assuming that electrons are captured by supersonic acoustic solitons, much like waves travelling along the lattice structures of crystals (proteins), trapping electrons on their way (electrosolitons). In con- nection with MT, propagation of such energy waves could act as a clocking mechanism, ,switch- ing the dimer dipoles from one conformational state to the other and triggering conformational changes.

190 E.M. Isinna et al. / BioSystems 39 (1996) 187-226
Still another model was proposed by Friihlich (Frohlich, 1968, 1970, 1975a, 1975b) and has been applied to MT to account for coherent charge displacement in MT (Hameroff et al., 1984). Al- though Bose condensation of oscillating modes is a phenomenon observed only at low temperatures (superfluidity and superconductivity), Frohlich, more than 20 years ago, suggested that biological molecules, if maintained in a non-equilibrium state at constant temperature and subjected to a steady external energy supply, would start behav- ing in coherent fashion under the effect of longitu- dinal frequency dipolar modes. Through strong excitation, some polar groups of these molecules would, in fact, be stretched, producing large dipole moments and entering a metastable polar- ized state. The oscillating dipoles would condense into a momentum-ordered state, displaying coher- ent oscillations of a single mode (coherent polar- ization waves) as well as long-range interactions and long-range correlations. The Friihlich’s mech- anism is, however, very improbable to occur at body temperatures and we have mentioned it here only for the sake of completeness.
2.1.2. MT dissipative dynamics and charge transfer
Our basic premise here is that the entire MT may be physically viewed as a triangular array of dipole moments which arise due to the presence of mobile electrons in hydrophobic pockets of the tubulin dimer. These electrons can switch their location from the a- to the P-monomer, thereby reversing the orientation of the associated dipole moment of the tubulin dimer. The above reversal mechanism is most likely triggered by the GTP- GDP hydrolysis. More specifically, the a-state of the tubulin unit has its mobile electron in the top dimer (dipole moment up) while the p-state has its mobile electron in the bottom dimer (dipole moment down). The former state corresponds to the GTP-bound tubulin while the latter to the GDP-bound tubulin. Therefore, a single hydroly- sis event results in electron motion which is also coupled to dipole reversal and an associated con- formational change. It is also conceivable that, under the influence of strong electric fields (which may arise in the cell’s interior) or due to doping
with impurity ions or, finally, as a result of ther- mal effects, these mobile electrons can become completely delocalized and form a conduction band. This, then, would directly lead to current flows along MT protofilaments. Otherwise, i.e. when the mobile electrons have only a limited range of motion, the corresponding dipole mo- ments of each tubulin dimer interact with their immediate tubulin neighbours via dipole-dipole forces. Each lattice site is assumed to possess a dipole moment proportional to the magnitude of the mobile charge unit (one or two elementary charges) and the distance over which it hops (2-4 nm).
Monte Carlo simulations have been performed for the resulting dipolar lattice (which is triangu- lar and has well-known dimensions) on the sur- face of a microtubule and the results obtained indicate the possibility of an emergence of at least three distinct dipolar phases: (a) the low-tempera- ture ferroelectric phase which is strongly polar- ized, (b) the intermediate temperature spin-glass phase which is very irregular in behaviour and (c) the high-temperature paraelectric phase which is disordered. The question of the temperature val- ues delineating these various phases is still open but it is suspected that room temperature magni- tudes are involved. From the point of view of energy and charge transfer potential the ferroelec- tric phase deserves the greatest attention. We describe its properties below.
In the ferroelectric phase, the MT system has a strong uniaxial dielectric anisotropy so that the array of individual dipole oscillators located at each tubulin site can be effectively described in terms of only one degree of freedom. This degree of freedom corresponds to the value of the net polarization along the protofilament axis. Athen- staedt (1974) experimentally demonstrated that a tubulin dimer undergoes a conformational change induced by the GTP-GDP hydrolysis in which one monomer shifts its orientation by 29” from the vertical axis. This fact was already mentioned briefly above. Here we also deduce on this basis that the single degree of freedom is also related to the projection on the MT cylinder’s axis of the dimer’s displacement from its undistorted posi- tion. When the cross section of a MT is viewed

E.M. lsinna et al. i BioSystems 39 (1996) 187-226 191
using electron microscopy, the outer surfaces of a MT are surrounded by a ‘clear-zone’ of several nm which apparently represents the oriented molecules of cytoplasmic water and enzymes (Stebbings and Hunt, 1982). This could be ex- plained by the presence of an electric field pro- duced by a MT. Therefore, it is assumed that together with the polarized water surrounding it, a MT generates a nearly uniform intrinsic electric field parallel to its axis. This is an important fact which plays a role in the modelling of the propa- gation of energy modes along the MT axis. Since these modes are associated with electronic mo- tion, they are naturally affected by any electric fields acting on them. It is worth emphasizing that the low-temperature ferroelectric phase is stabi- lized by both external and internal electric fields and is characterized by an almost perfect align- ment of dipole moments along the protofilament axis. It is also characterized by long-range order and hence it eagerly supports the formation of domain walls between the two stable orientations of dipole moments: up and down the protofila- ment axis. A domain wall is a small segment of the protofilament over which the dipole orienta- tion changes from predominantly upward to pre- dominantly downward. This region of the MT lattice stores both electronic and mechanical en- ergy which can, in fact, propagate with little loss as will be discussed below. For example, the ap- plication of an external electric field preferentially directs kink-like domain wall excitation towards the properly aligned end of the protofilament which may cause a disassembly of the protofila- ment due to the energy released by the kink as it unloads its energy at its destination point. On the other hand, when local impurities are encountered by kinks on their way down the prototilament, they slow their motion down, eventually stopping kink motion altogether for sufficiently large impu- rity potentials. This mechanism may elucidate the role of MAPS in the capture of GTP-generated energy which is known to disappear from the positive end’s so-called lateral cap.
A realistic equation of motion for the above described dynamic degree of freedom has been derived (Sataric, 1993) which includes the viscos- ity of the cytoplasm, whose resistance these sur-
face modes encounter, as well as the associated force due to the electric fields which would other- wise lead to the acceleration of the moving do- main walls. As a result of the balance between these two competing mechanisms a constant ve- locity solution is obtained for this equation. In fact, it has been shown analytically that a unique bounded solution exists in the form of a kink giving the boundary between the two electronic states characterized by the preferential pasition in the top or bottom monomer, respectively. It is interesting to note that this domain wall moves with a unique terminal velocity which is nearly proportional to the electric field intensity and inversely proportional to the viscosity.
Taking as an example a moderately long MT consisting of approximately lo* dimers with length L g 10 ~ 6 m, the effective charge Q z 1.5.10- I6 C is estimated so that the intrinsic electric field in the vicinity of the middle point of a MT is found to be on the order of E 2 lo6 V/m. Accounting for the dielectric effects of the sur- rounding water molecules, this value must be reduced to approximately E z 10’ V/m. Thus taking E r 10’ V/m as a representative average of the electric field, the propagation velocity of a kink is on the order of 2 m/s. Assuming a.smooth journey from one end of the MT to the other, the average time of propagation for a single kink should be 10 - ’ s. These numbers are somewhat unrealistic and they overestimate the speed of propagation. In reality, the fields may be even further reduced due to screening effects of water and ions in the cytoplasm. Furthermore, the pres- ence of MAPS and lattice irregularities could sig- nificantly reduce the above values (by several orders of magnitude) which would then fall in the range of typical micrometer/second values for protein transport along filaments.
It should be emphasized that the effects of discreteness of the MT lattice (which have been ignored in the past by using the continuum ap- proximation) may play an important role. For example Kimball (1980) performed an anailysis of kink dynamics for a discrete lattice inferring that there may exist a threshold value of the external field E required to sustain kink motion.

192 E.M. Isinna et al. / BioSystems 39 (1996) 187-226
We should also emphasize that both possibilities of linear and helical kink propagation exist. The first, i.e. propagation of kinks along the protofila- ment axis might exist in MT lattices with a seam (Mandelkow, 1992; Kikkawa et al., 1994). The other is a natural mode of propagation in seamless MT lattices which actually are less abundant in vitro. Theoretically, both possibilities have also been confirmed for acoustic phonon modes. In the context of electronic degrees of freedom, the helical pathway would slow down the net speed of prop- agation by at least one order of magnitude bring- ing it closer to the observed values of l-2 pm/s.
It is apparent that the intrinsic electric field governs the rate of propagation of kinks. By adding an external electric field to the Hamiltonian one can introduce a new control mechanism in the MT dynamics. An applied electric field will result in a faster moving population of kinks and, thus, a greater stability against thermal fluctuations; if, on the other hand, the intrinsic and applied electric fields are oriented in opposite directions, the kink’s motion may be slowed or stopped altogether. Each kink within a MT can be viewed as a energy unit (for example, arising from GTP hydrolysis) whose propagation can be controlled by an external electric field or by the concentration of GTP molecules in the cytoplasm. It should be mentioned that another group of authors (Chou, Zhang and Maggiora, 1994) predicted the emergence of soli- tary wave excitations in MT’s but they did not link these non-linear waves with dielectric polarization. Instead, they only considered structural dislocation motion. Due to the pyroelectric effect these two degrees of freedom are coupled and hence these may just be two aspects of the same phenomenon.
Fig. 2. Model showing a microtubule constituted of thirteen protofilaments. A three-start helical family means that the microtubule is constituted of three helices stacked together. The pitch of a single helix is, in this case, 120 A. Although different lattice types have been observed (A and B type, corresponding to the A and B subfibers of axonemal MT doublets), it is probable that both configurations allow charge transfer to occur along the lattice (see text).
2.1.3. MT stability requirements Following the above considerations on the exis-
tence of electron transfer along the MT lattice, we suggest that microtubule organizing centers (MTOCs) should be divided into electron sources and sink devices, that is, into electron donors and electron acceptors. In our model, we assume that both devices are needed to maintain charge trans- fer and ensure the necessary stability.
In the following we will now briefly outline the Under conditions of sufficiently high tubulin stability requirements for microtubular motors, i.e. concentration, MT nucleate preferentially from a the necessary requirements to sustain kink motion MTOC, as for example the centrosome, which we (charge transfer) in the MT lattice. For the sake of assimilate to a source device, i.e. electron donor simplicity, we have based our model on the helical (Mitchison and Kirschner, 1984; Mitchison et al., mode of propagation by assuming that charge 1986). Disconnection from a source device like transfer follows a helical pathway according to a the centrosome causes disassembly (Kitanishi-Yu- 3-start helical family as shown in Fig. 2. Although mura and Fukui, 1987) because of the lack of A and B lattice types have been observed (Man- electron supply at the - end. But the same effect delkow et al., 1986) we believe that both allow can be produced in the absence of a sink at the + charge transfer either following a linear or a helical end like the kinetochores of chromosomes in the path along the MT lattice. mitotic spindle, which we assimilate to an electron

E.M. Isinna et al. I BioSystems 39 (1996) 187-226 193
acceptor. Spindle MT, in fact, nucleate during mitosis from centrosome and tubulin dimers are added distally at the kinetochores by an unknown capping mechanism (Margolis and Wilson, 1981; Mitchison, 1986; Huitorel and Kirschner; 1988). The arrangement probably ensures the continuous addition of GTP-carrying tubulins (the GTP cap) at the + end to avoid catastrophic depolymeriza- tion in the absence of a current sink.
Cytoplasmic MT growing from the centrosome reach the cell membrane by progressive addition of dimers at the -I- end (Mitchison et al., 1986). When they finally attain the membrane, special capping proteins attach at their + end and most likely produce an electric field (potential differ- ence) to maintain the necessary sink (bias) and stabilize the MT fibers. Stabilization, in this case, can be achieved only if MT stay connected to the electron source, i.e. the centrosome.
During the development of ciliated epithelial cells, ciliary MT assembly is nucleated when the MT issued from the basal bodies reach the plasma membrane (Portman and Dentler, 1987, 1989). Therefore, it is plausible to think that the mem- brane potential is used for MT stabilization through special capping proteins that reach into the membrane domain (which should be positive with respect to the cytoplasm).
Perhaps the most suggestive hint at the probable role of electron suppliers for source MTOCs is the replacement of basal bodies by mitochondria in the flagella of the alga Chlorogonium. During cell division, the cell needs the basal bodies to recover their primary role in the cell spindle, i.e. to act as centrosomes. They, thus, detach from the flagella and move toward the cell center. The transition zone, though, establishes a connection to an apical mitochondrion (Hoops and Witman, 1985). Be- cause there are no available free electrons in the cytoplasm, the cell has to look for the next best source. There is, in the cell, no better electron supplier than a mitochondrion, provided the MT are capable to divert some electrons from the oxidation capabilities of NADH (the reduced form of nicotinamide adenine dinucleotide). When glu- cose is converted to pyruvate in the cytoplasm, two NAD+ (oxidised form) are reduced to NADH. Subsequently, the electrons resulting
from oxidation of NADH are transferred into the mitochondrion through reduced electron shuttles. Somehow, the MT structure is capable of tapping the electron transport system and diverting the charges for its use before they enter the mitochon- drial membrane.
Summing up, we believe that MT structures need a potential difference between the -I- and the - end as well electron source and sink devices in order to become stable through electron transfer. We already mentioned the centrosome and the basal bodies in cilia and flagella as electron sources. We also assumed the kinetochores to be electron sink devices. But are there other biologi- cal structures which can be considered as probable candidates for the role of current sink devices?
In our view, it is not an accident that the most sophisticated capping structures have been imple- mented by the cell in cilia and flagella (for review see Dentler, 1990). The distal tips of the nine A fibers from axonemal MT doublets, as well as the central MT pair, are all connected to rather com- plex microtubule capping structures at the distal end of the axoneme in most cilia and flagella (see Fig. 3a). The MT are capped, either by means of central MT caps, for the MT central pair, or through distal filament plugs for the A tubules. The plugs insert into the MT lumen indicating a probable conductive role for the connection. The B subfibers of the doublet MT are not capped.
Capping structures stop MT growth ,in vitro (Dentler and Rosenbaum, 1977, DentIer and LeCluyse, 1982) and, in vivo, they regulate MT growth by allowing for MT addition at the + end. The mechanism possibly involves phosphory- lation of the capping proteins (Dentler, 1986) perhaps in a way similar to the model proposed by Mitchison et al. (1986) and Huitonel and Kirschner (1988) consisting in a sliding collar around the MT fibers.
Apart from their growth-regulating features, capping structures have been thought to work as ball bearings protecting the MT ends to avoid damage when they slide past one another during ciliary (flagellar) beating. The fact that B subfiber MT are not capped has been interpreted as a confirmation of this hypothesis for they terminate proximally to the A tubule (Dentler, 199tY).
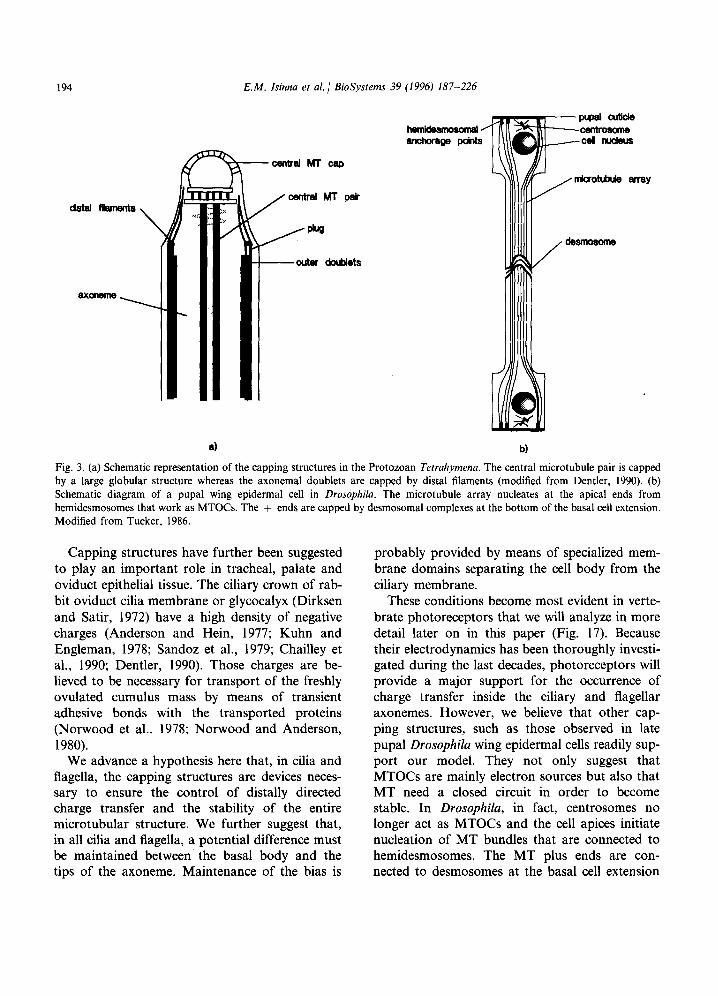
194 E.M. Isinna et al. 1 BioSystems 39
a) b)
Fig. 3. (a) Schematic representation of the capping structures in the Protozoan Tetrahymena. The central microtubule pair is capped by a large globular structure whereas the axonemal doublets are capped by distal filaments (modified from Dentler, 1990). (b) Schematic diagram of a pupal wing epidermal cell in Drosophila. The microtubule array nucleates at the apical ends from hemidesmosomes that work as MTOCs. The + ends are capped by desmosomal complexes at the bottom of the basal cell extension. Modified from Tucker, 1986.
Capping structures have further been suggested to play an important role in tracheal, palate and oviduct epithelial tissue. The ciliary crown of rab- bit oviduct cilia membrane or glycocalyx (Dirksen and Satir, 1972) have a high density of negative charges (Anderson and Hein, 1977; Kuhn and Engleman, 1978; Sandoz et al., 1979; Chailley et al., 1990; Dentler, 1990). Those charges are be- lieved to be necessary for transport of the freshly ovulated cumulus mass by means of transient adhesive bonds with the transported proteins (Norwood et al., 1978; Nor-wood and Anderson, 1980).
We advance a hypothesis here that, in cilia and flagella, the capping structures are devices neces- sary to ensure the control of distally directed charge transfer and the stability of the entire microtubular structure. We further suggest that, in all cilia and flagella, a potential difference must be maintained between the basal body and the tips of the axoneme. Maintenance of the bias is
probably provided by means of specialized mem- brane domains separating the cell body from the ciliary membrane.
These conditions become most evident in verte- brate photoreceptors that we will analyze in more detail later on in this paper (Fig. 17). Because their electrodynamics has been thoroughly investi- gated during the last decades, photoreceptors will provide a major support for the occurrence of charge transfer inside the ciliary and flagellar axonemes. However, we believe that other cap- ping structures, such as those observed in late pupal Drosophila wing epidermal cells readily sup- port our model. They not only suggest that MTOCs are mainly electron sources but also that MT need a closed circuit in order to become stable. In Drosophila, in fact, centrosomes no longer act as MTOCs and the cell apices initiate nucleation of MT bundles that are connected to hemidesmosomes. The MT plus ends are con- nected to desmosomes at the basal cell extension

E.M. Isirzna et al. / BioSystems 39 (1996) 187-226 195
(Tucker et al., 1986; Mogesen and Tucker, 1987; Mogesen et al., 1989) (Fig. 3b).
In the following, will apply our charge transfer model to the dynamics of intracellular transport then to flagellar and ciliary beating and finally, in Part II, to some control mechanisms inherent in vertebrate photoreceptors. We will see how the electrodynamics of some MT configurations sup- plies much evidence that electron transfer does exist inside MT and related complex structures such as cilia and flagella and that they are dynam- ically active systems truly deserving the name of microtubular motors (MTM).
2.2. Dynamics of intracellular transport
All dynamic cell processes involving motility-re- lated proteins such as dyneins, kinesins or actins and myosins, presently lack a plausible model capable of accounting for synchronisation of the contraction/motility process (for review see Gib- bons, 1988; Vale, 1987 and 1993).
Why shouldn’t, for example, the ATPase (the dynein arms) in the MT doublets of a ciliary axoneme contract in an anarchic fashion along the entire axonemal structure when ATP is present in sufficient concentration? Why should cytoplasmic dyneins transport along MT fibers in nerve axons or chromosome movement during mitosis in all dividing cells occur as a very ordered process? Which mechanism lies at the basis of the sequential activation of all those motility-related proteins? To date there is no available hypothesis capable of giving a plausible answer to those questions.
The motor protein family englobes several types of molecules capable of anterograde (toward the MT - end) or retrograde (toward the MT + end) transport (Brady, 1985; Brady et al., 1990; Hirokawa et al., 1991; Hirokawa, 1993; Vale, 1993; Niclas et al., 1994; Nangaku et al., 1994, Okada et al., 1995). In retrograde protein trans- port we suggest that the sequential activation of motor protein-mediated is due to a mechanism involving charge transfer. Conformational changes are thus induced by electron transfer in the ATP binding domain of the motor protein. This, in turn, causes binding of a new ATP
molecule, disconnection of the dynein head from the MT binding domain and recovery of the initial conformational state. The entire cycle can be described as follows (see Fig. 4):
The flow of an electron inside the helical MT lattice induces conformational changes within the ATP binding domain (motor domain) of the force-generating enzyme (site number 2) allowing an ATP molecule to bind to the enzyme. Hydroly- sis of the ATP molecule subsequently disconnects one of the protein heads from the MT. This one head then binds very rapidly to the next site (number 3) before the electron activates the ATP binding site of the second head (site number 1). The protein then waits for the next activation to continue its retrograde gliding walk along the track. A recovery time inherent in the mechanism prevents the head from re-attaching to the same site immediately after ATP hydrolysis.
This model accounts for the necessity of the presence of multiple heads in motor proteins. In fact, two heads are the ideal solution in order to keep the contact with the MT track. A single head might result in a random walk on the MT surface, although one-headed kinesins do exist (Nangaku et al., 1994; Okada et al., 1995). The presence of multiple heads in force-generating enzymes, on the other hand, ensures a stable attachment to the binding site on the MT and the production of higher forces. Additionally, multiple heads also allow the mechanism to work at higher frequen- cies. In cilia and flagella, dynein arms have in fact multiple heads (Gibbons, 198 1).
The mechanism inherent in anterograde trans- port observed for some kinesin families (Vale, 1993) probably involves different characteristics of the motor protein. A tentative mechanism as- sumes slower ATP hydrolysis and a transiently active binding domain on the MT lattice. The cycle for a specific one-headed + end (antero- grade) directed kinesin type would, thus, be as follows (see Fig. 5):
The electron transfer activates the binding of ATP on site number 1 and allows the motor protein head to disconnect. Because the process of ATP hydrolysis is slower and the activation of the binding sites number 0 and 1 is only a transient phenomenon the head can only bind to site num-
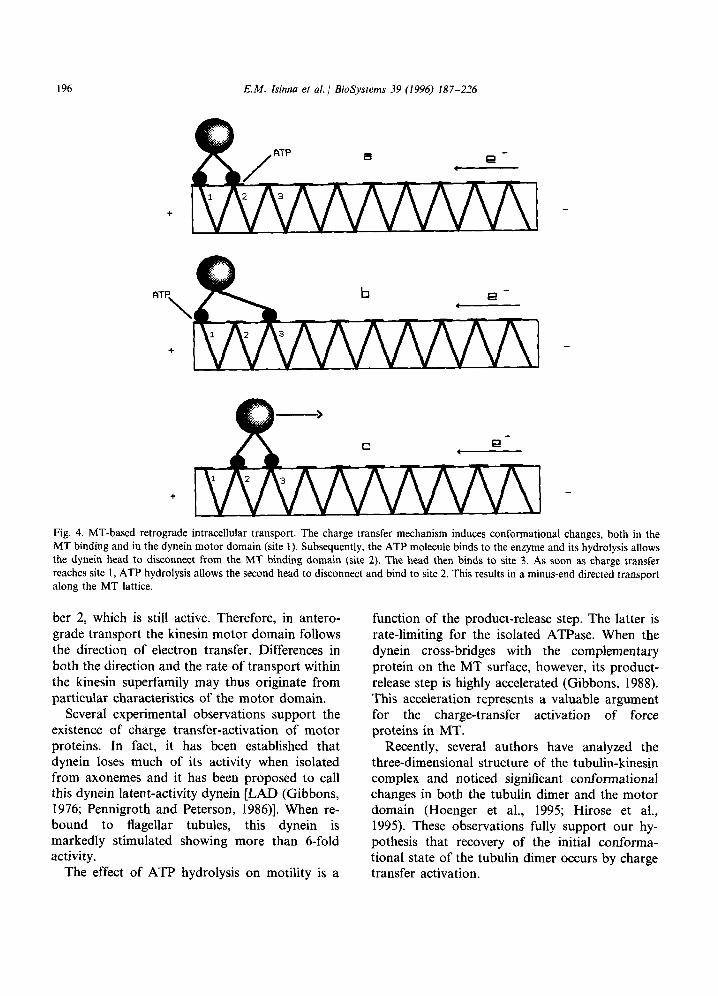
196 E.M. Isinna et al. 1 BioSystems 39 (1996) 187-226
ATP
\
+
Fig. 4. MT-based retrograde intracellular transport. The charge transfer mechanism induces conformational changes, both in the MT binding and in the dynein motor domain (site 1). Subsequently, the ATP molecule binds to the enzyme and its hydrolysis allows the dynein head to disconnect from the MT binding domain (site 2). The head then binds to site 3. As soon as charge transfer reaches site 1, ATP hydrolysis allows the second head to disconnect and bind to site 2. This results in a minus-end directed transport along the MT lattice.
ber 2, which is still active. Therefore, in antero- grade transport the kinesin motor domain follows the direction of electron transfer. Differences in both the direction and the rate of transport within the kinesin superfamily may thus originate from particular characteristics of the motor domain.
Several experimental observations support the existence of charge transfer-activation of motor proteins. In fact, it has been established that dynein loses much of its activity when isolated from axonemes and it has been proposed to call this dynein latent-activity dynein [LAD (Gibbons, 1976; Pennigroth and Peterson, 1986)]. When re- bound to flagellar tubules, this dynein is markedly stimulated showing more than 6-fold activity.
The effect of ATP hydrolysis on motility is a
function of the product-release step. The latter is rate-limiting for the isolated ATPase. When the dynein cross-bridges with the complementary protein on the MT surface, however, its product- release step is highly accelerated (Gibbons, 1988). This acceleration represents a valuable argument for the charge-transfer activation of force proteins in MT.
Recently, several authors have analyzed the three-dimensional structure of the tubulin-kinesin complex and noticed significant conformational changes in both the tubulin dimer and the motor domain (Hoenger et al., 1995; Hirose et al., 1995). These observations fully support our hy- pothesis that recovery of the initial conforma- tional state of the tubulin dimer occurs by charge transfer activation.

E.M. Isinna et al. / BioSystems 39 (1996) 187-226 197
e- *
e- <
e- 4
Fig. 5. MT-based anterograde intracellular transport. In this case we additionally assume that activation of the MT binding site is transient and that ATP hydrolysis is slightly slower. The head then binds to site number 2 which is still active and follows the direction of charge transfer. _ - _
Additional support for motor protein activation dependent on the (probably helical) motion of electrons in the MT lattice comes from experi- ments with Tetrahymena 14 S single-headed dynein isolated from axonemes. This motor protein has been observed to cause rotation of the axonemes on a glass surface (Marchese-Ragona and Johnson, 1985: Vale and Toyoshima, 1988). According to our model, the resulting torque is generated by the helical paths followed by the electrons in the axoneme.
The discovery of a seam in the MT lattice (Linck and Langevin, 1981; Linck et al., 1981; Mandelkow et al., 1986; Chretien and Wade, 1991; Kikkawa et al., 1994) appears to be an additional argument in favor of our hypothesis. The seam, constituted by MAPS, has been inter- preted to be a discontinuity in the MT wall lattice.
We believe that the seam is the control path on which the charge transfer dependent interactions between MT dimers and motor proteins occur. Especially in connection with flagellar MT, one of the protofilaments of the doublet MT, has been shown to be composed of special MAPS, i.e. tektins (Linck and Langevin, 1981; Linck et al., 1981; Nojima et al., 1995). Those stable filaments most probably play an important role as activa- tion sites in the charge transfer-dependent control mechanism. In fact their axial periodicities exactly match the binding sites of axonemal components (dynein, nexin, radial spokes) on the tubulin lat- tice (Pirner and Linck, 1994). The various ar- rangements of tubulins observed in the lattice represent, in our opinion, different solutions (elec- tron or control pathways) for different control requirements.

198 E.M. Isinna et al. / BioSystems 39 (1996) 187-226
In the following and in connection with other motility-related proteins like actin, we will show that charge transfer-dependent protein activation is an important process capable of shedding a new light on ciliary and flagellar dynamics as well as on axonemal growth and vertebrate photoreceptor’s membrane renewal. This mechanism is at the base of all MT-driven motility processes MT are the clocking devices capable of co-ordinating the ac- tivity of all force-generating enzymes.
2.3. Ciliary and flagellar beating
One of the authors has proposed a model of MT dynamics and by extension, of flagellar and ciliary beating (Insinna, 1989) based on charge transfer. Here we will consider the basic features of that early model, but we will also give an updated account of it, for an introduction to cilia and flagella, the reader is asked to refer to (Sleigh, 1974; Goldman et al., 1976; Dustin, 1984, Wit- man, 1990).
2.3.1. Cilia andj7agella as active devices In cilia and flagella, displacement of charges is
suggested to occur, in the normal state, distally inside all MT fibers composing the axoneme. In motile ciliary structures, a potential difference has to be maintained between the tip of the axoneme and the cell’s interior for correct functioning of the beating mechanism. In most cilia and flagella, such a potential difference (bias) seems to be main- tained by special membrane domains separating the ciliary membrane from the negative potential in the cell’s interior. The capping structures we previously mentioned probably play a significant role in the maintenance of this polarization. This fact, as we will see, is a necessary and universal prerequisite in all ciliary systems, and most partic- ularly in photoreceptors. The existence of a bias in MT structures, leads naturally to a comparison between MT and semiconductors. The electrical parameters of MT, however, are likely to be very different. For instance, the reverse voltage V,, if any, may be expected to be very low in compari- son to silicon devices and its value is probably highly influenced by several factors such as tem- perature, ionic concentrations, etc.
Because of such dissimilarities, charge transfer probably occurs in both directions along the MT fibers of cilia and flagella. Experimental data from ciliary and flagellar activity seem to confirm this possibility. The dynamics of ciliary beating implic- itly indicates the necessity for charge transfer reversal during the recovery stroke (Machemer, 1974, Naitoh and Eckert, 1974) and bending waves of opposite direction have been observed to propagate along irradiated flagella (Holwill, 1965; Goldstein, 1972; Sleigh, 1974). In cilia, it has been shown that the power stroke is much faster than the recovery stroke (Brokaw, 1974). This is proba- bly due to the different conduction characteristics when the MTM is forced to work in reverse mode. The above observations suggest that, unless the applied forward voltage V, is much larger than the reverse voltage V,, charge transfer may be expected to take place in an unsteady fashion inside the MTM and its direction may sometimes change in absence of a finite polarization.
Since the beginning of the century, many exper- imental data from studies of responses to electric stimuli in Protozoa seem to support the possibility of a predominantly distally directed charge trans- fer inside cilia and flagella (for review see Naitoh and Eckert, 1974). When Paramecium is placed in the path of a galvanic current, ciliary reversal occurs on the cell surface facing the cathode. Increase of the beating frequency of the cilia in the normal direction manifests instead on the surface of the cell facing the anode. In Opalina, ciliary reversal takes place also at the surface facing the cathode. Stimulation experiments by intracellular electrodes in Opalina, confirm that outward cur- rent through the plasma membrane (depolariza- tion, i.e. membrane interior positive) induces ciliary reversal. Inward current (hyperpolarization, i.e. membrane interior more negative) induces, instead, an increase in beat frequency of the cilia in the normal direction. Moreover, the beating frequency also increases in high calcium concen- trations (Naitoh and Eckert, 1974).
This last fact need not be confusing. In fact, external Ca2 + may act locally on the exterior of the ciliary membrane (for instance on the capping structures) and increase the potential difference

E.M. Isinna et al. 1 BioSysrems 39 (1996) 187-226 199
between the tips of the axoneme and the cell’s interior. This, in turn, causes an acceleration of charge transfer inside the ciliary axoneme with subsequent increase in beating frequency. Gualtieri et al. (1986) have succeeded in evaginat- ing the reservoir membrane in Euglenu gracilis by using high external concentrations of CaCl,. The flagellum beating frequency is highly accelerated and the entire flagellar apparatus is displaced out of the reservoir. This observation is totally in accordance with the model we propose here.
ATP hydrolysis (in the presence of Mg2 +) is the main source of energy for the axonemal motil- ity proteins (dyneins). It has already been ob- served that frequency of beating is mainly a function of ATP concentration, but that the speed of bend propagation is independent from it (Goldstein, 1974; Sleigh, 1974). From the view- point of our model, it may be inferred that the beating frequency is proportional to both the number of hydrolysed ATP molecules and the speed of charge transfer due to the potential difference existing between the cell’s interior and the distal part of the axoneme. The speed of bend propagation, however, should be a function of charge transfer only. The above results seem once more to confirm our suggestions.
Observations concerning ciliary reversal phe- nomena have been at the base of the calcium current hypothesis. According to it, all stimuli that evoke ciliary reversal, electrical or mechani- cal, are at the origin of a net influx of calcium from the external medium into the cell (Eckert, 1972; Naitoh and Eckert, 1974). This hypothesis involves membrane conductivity changes through activation of ion channels. We believe that this hypothesis needs still to be complemented with another mechanism involving MT-membrane in- teractions. We will, therefore, expand on this sub- ject later on in Part II, in connection with the dynamics of Euglena’s photoreceptor.
2.3.2. Charge transfer activation of axonernal ATPases and radial spoke sating mechanism
In cilia and flagella, we suggest that the sequen- tial activation of motility proteins is due to the presence of electron transfer in the MTB subfibers to which the multiple heads of the dynein arms
are attached. The motion of charges induces spe- cific conformational changes in the motor protein head and in its site of attachment. Only after this charge transfer-induced change has occurred can the ATPase bind an ATP molecule, and the dynein arms disconnect from the adjacent MT subfiber via ATP hydrolysis.
In ciliary and flagellar beating, the smooth propagation of the bending wave along the axon- eme presupposes, not only the sequential activa- tion of the dynein arms on each of the doublets, but also a sequential co-ordination of the entire force-generating mechanism.
The current model, based on the sliding MT hypothesis (Satir, 1968; Sale and Satir, 1977) can- not account for such a co-ordination, so we pro- pose here a new model of motility-related protein dynamics capable of shedding a new light on the co-ordination of the beating process.
Our starting point is the fact that Clumy- domonas mutants, lacking either the radial spokes or the central MT pair, are immobilized (Warr et al., 1966; Witman et al., 1976, 1978; Luck, 1984; Huang, 1986; Witman, 1990). On the other hand, ATPase activity has been detected in the center of the axoneme (Warner and Satir, 1974; Warner, 1976) and Witman et al. (1976) have advanced that ATPase activity of the radial spoke head is probable.
ATPase activation in dynein arms is suggested to occur through charge transfer signalling at the site of attachment of the head on the MTB sub- fiber. Additionally, we advance that this activa- tion of the proximal part (the head), is accom- panied by a conformational change occurring at its distal part. This distal conformational change in motility proteins has been widely implemented by the cell to solve all synchronization problems inherent in ciliary and flagellar dynamics.
The ATPase in the radial spoke-head is needed for signalling to and control of the MT doublet to which the radial spoke is attached distally. The conformational change occurring in the distal part of the radial spokes works as a gating mechanism as in semiconductors devices. When inactivated, the radial spokes block the current flow inside the corresponding MTB subfibers, as long as the ra- dial spoke head is not disconnected from the
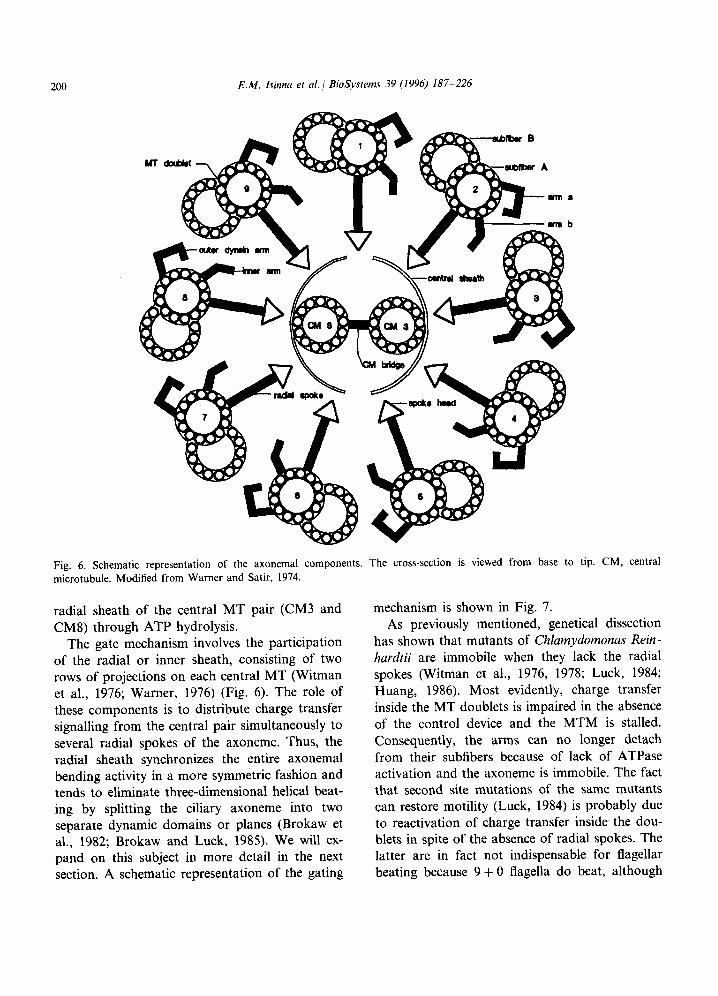
200 E.M. Isinna et al. / BioSystems 39 (1996) 187-226
Fig. 6. Schematic representation of the axonemal components. The cross-section is viewed from base to tip. CM, central mi&otubule. Modified from Warner and Satir, 1974.
radial sheath of the central MT pair (CM3 and CM8) through ATP hydrolysis.
The gate mechanism involves the participation of the radial or inner sheath, consisting of two rows of projections on each central MT (Witman et al., 1976; Warner, 1976) (Fig. 6). The role of these components is to distribute charge transfer signalling from the central pair simultaneously to several radial spokes of the axoneme. Thus, the radial sheath synchronizes the entire axonemal bending activity in a more symmetric fashion and tends to eliminate three-dimensional helical beat- ing by splitting the ciliary axoneme into two separate dynamic domains or planes (Brokaw et al., 1982; Brokaw and Luck, 1985). We will ex- pand on this subject in more detail in the next section. A schematic representation of the gating
mechanism is shown in Fig. 7. As previously mentioned, genetical dissection
has shown that mutants of Chlamydomonas Rein- hardtii are immobile when they lack the radial spokes (Witman et al., 1976, 1978; Luck, 1984; Huang, 1986). Most evidently, charge transfer inside the MT doublets is impaired in the absence of the control device and the MTM is stalled. Consequently, the arms can no longer detach from their subfibers because of lack of ATPase activation and the axoneme is immobile. The fact that second site mutations of the same mutants can restore motility (Luck, 1984) is probably due to reactivation of charge transfer inside’ the dou- blets in spite of the absence of radial spokes. The latter are in fact not indispensable for flagellar beating because 9 + 0 flagella do beat, although
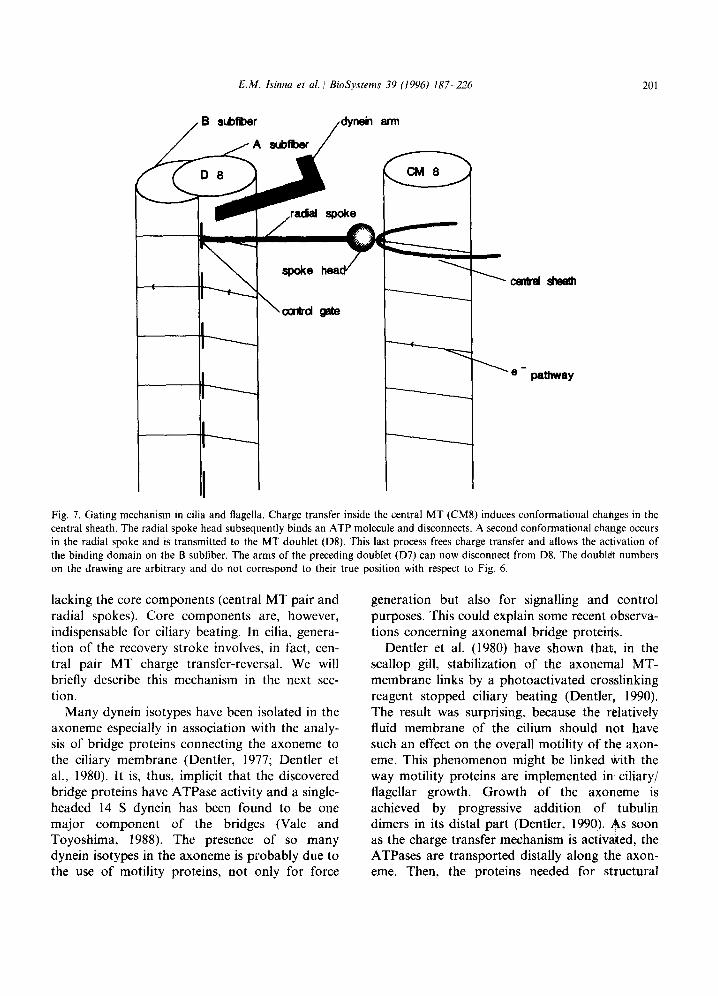
E.M. Isinna et al. / BioSystems 39 (1994) 187-226
/
B s&fiber
/* /
dynein arm
201
Fig. 7. Gating mechanism in cilia and flagella. Charge transfer inside the central MT (CM8) induces conformational changes in the central sheath. The radial spoke head subsequently binds an ATP molecule and disconnects. A second conformational change occurs in the radial spoke and is transmitted to the MT doublet (D8). This last process frees charge transfer and allows the activation of the binding domain on the B subfiber. The arms of the preceding doublet (D7) can now disconnect from D8. The doublet numbers on the drawing are arbitrary and do not correspond to their true position with respect to Fig. 6.
lacking the core components (central MT pair and radial spokes). Core components are, however, indispensable for ciliary beating. In cilia, genera- tion of the recovery stroke involves, in fact, cen- tral pair MT charge transfer-reversal. We will briefly describe this mechanism in the next sec- tion.
Many dynein isotypes have been isolated in the axoneme especially in association with the analy- sis of bridge proteins connecting the axoneme to the ciliary membrane (Dentler, 1977; Dentler et al., 1980). It is, thus, implicit that the discovered bridge proteins have ATPase activity and a single- headed 14 S dynein has been found to be one major component of the bridges (Vale and Toyoshima, 1988). The presence of so many dynein isotypes in the axoneme is probably due to the use of motility proteins, not only for force
generation but also for signalling and control purposes. This could explain some recent observa- tions concerning axonemal bridge proteins.
Dentler et al. (1980) have shown that, in the scallop gill, stabilization of the axonemal MT- membrane links by a photoactivated crosslinking reagent stopped ciliary beating (Dentler, 1990). The result was surprising, because the relatively fluid membrane of the cilium should not have such an effect on the overall motility of the axon- eme. This phenomenon might be linked with the way motility proteins are implemented in ciliary/ flagellar growth. Growth of the axoneme is achieved by progressive addition of tubulin dimers in its distal part (Dentler, 1990). As soon as the charge transfer mechanism is activated, the ATPases are transported distally along the axon- eme. Then, the proteins needed for structural

202 E.M. Isinna et al. / BioSystems 39 (1996) 187-226
purposes progressively bind to the axonemal dou- blets forming cross-bridges with the membrane. However, because their role is only structural, once they are bound, their charge transfer activa- tion mechanism is somehow inhibited and they remain as stable membrane bridges. Although they do no longer detach from the doublets it is probable that they are still partially involved in the charge transfer circuit. Stabilization as de- scribed by Dentler et al. (1980) probably stops charge transfer along the doublets and the cilium is immobilized.
This last example implies that proteins (dyneins) used for structural purposes become immobilized through inhibition of the ATPase mechanism. The cell has probably found a way to modulate conformational changes adapting them to different uses.
2.3.3. The beating mechanism of cilia andjagella We are now in possession of the necessary tools
to supply a new model for the motility of cilia and flagella. The dynamics of a bending cycle can now be described as follows:
To begin with, we imagine the flagellum in its rigor state, i.e., immobilized because of the at- tachment of all dynein arms to the adjacent MT pairs. If we consider that the arms on all the MT fibers are disposed on a helical path and if we assume the arms on MTlA to be at the very bottom of the axoneme, then, at the end of one period, the dynein arms A9a and A9b protruding from MT9A will be located higher up along the helix with respect to those of MTlA. Thus, when attached, the arms A9a and b will pull on the opposite subfiber MTlB distally, or either MT9 will tend to slide proximally (See Fig. 6).
At the beginning of the cycle, in the proximal zone of the flagellum, Ala and b start disconnect- ing from MT2B. Because the arms A9a and A9b of MT9 are still attached to MTlB and pull on it (they are located higher up in the axoneme), the doublet MT1 will have the tendency to slide dis- tally. Now it will be the turn of A2a and b to disconnect from MT3B. Yet, this cannot happen without previous signalling from the central MT CM3. This occurs by means of the radial sheath protruding from it and the radial spoke connected
to MT3. When charges move along CM3, they activate the radial sheath and consequently the ATPase on the radial spoke head. The head will bind an ATP molecule and will disconnect itself from. the radial sheath through ATP hydrolysis. The hydrolysis simultaneously produces a confor- mational change that is transmitted distally to the area where the radial spoke segment is connected to MT3A and B. This connection has the purpose of controlling the conduction circuit of MT3, that is, of the subfiber MT3B connected in series. This is necessary to prevent the release of the dynein arms A2a and b without previous signalling from CM3. This mechanism is comparable to a semi- conductor gating device. Therefore, the current inside MT3B starts flowing only after activation of the radial spoke head ATPase, subsequent ATP hydrolysis and disconnection of the radial spoke from the radial sheath. Charge transfer release in MT3 in turn activates the site of attachment of the dynein ATPases of arms A2a and A2b (on MT2A). The activation allows the binding of a new ATP molecule so as to disconnect them from MT3B. When this occurs, it is the turn of MT2 to be displaced distally and so on.
As already mentioned, the dynein arms and the radial spokes are helically disposed along the ax- onemal structure. This helical disposition results in a progressive spreading apart of a winding of the axonemal components under the action of MTl-5 (Fig. 8). Because of the mechanical con-
a 1s dLectlon of bon&g
Fig. 8. Spreading apart of a winding of the helically disposed axonemal components. The bending follows a helical path along the axoneme and tends to generate a purely helical beating in flagella with only one central MT and in those lacking central components. In cilia and flagella with a central MT pair instead, the helical beating is attenuated by the separation existing between the central sheaths (see Fig. 6).
E.M. Isinna et al. 1 BioSystems 39 (1996) 187-226 203
al
b)
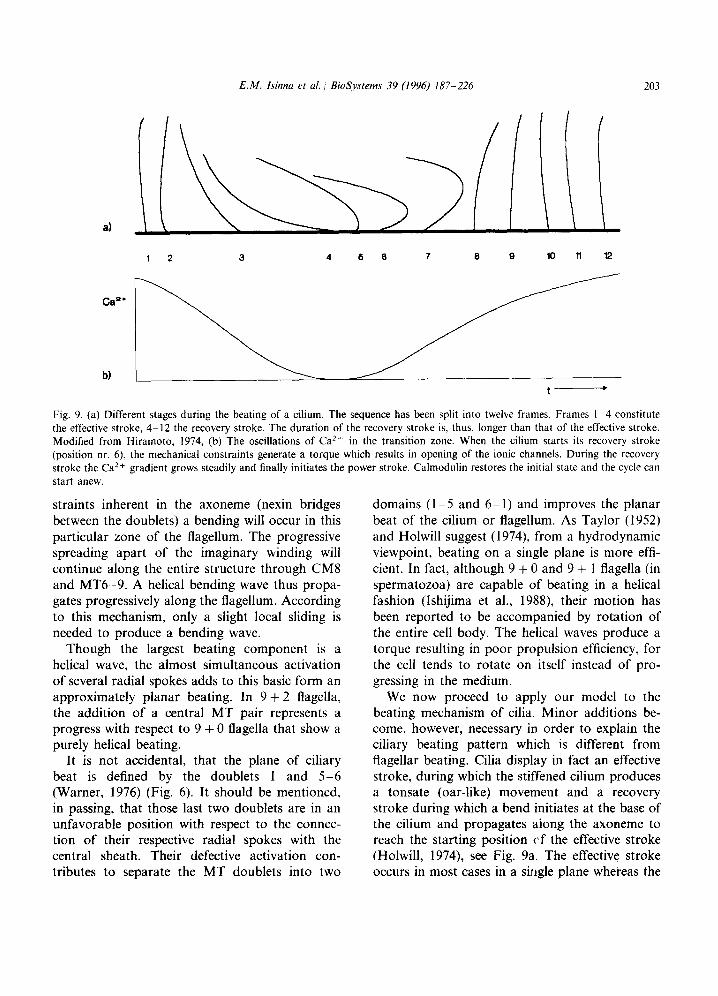
Fig. 9. (a) Different stages during the beating of a cilium. The sequence has been split into twelve frames. Frames I-4 constitute the effective stroke, 4-12 the recovery stroke. The duration of the recovery stroke is. thus, longer than that of the effective stroke. Modified from Hiramoto, 1974, (b) The oscillations of Ca *+ in the transition zone. When the cilium starts its recovery stroke (position nr. 6), the mechanical constraints generate a torque which results in opening of the ionic channels. During the recovery stroke the Ca*+ gradient grows steadily and finally initiates the power stroke. Calmodulin restores the initial state and the cycle can start anew.
straints inherent in the axoneme (nexin bridges between the doublets) a bending will occur in this particular zone of the flagellum. The progressive spreading apart of the imaginary winding will continue along the entire structure through CM8 and MT6-9. A helical bending wave thus propa- gates progressively along the flagellum. According to this mechanism, only a slight local sliding is needed to produce a bending wave.
Though the largest beating component is a helical wave, the almost simultaneous activation of several radial spokes adds to this basic form an approximately planar beating. In 9 + 2 flagella, the addition of a central MT pair represents a progress with respect to 9 + 0 flagella that show a purely helical beating.
It is not accidental, that the plane of ciliary beat is defined by the doublets 1 and 5-6 (Warner, 1976) (Fig. 6). It should be mentioned, in passing, that those last two doublets are in an unfavorable position with respect to the connec- tion of their respective radial spokes with the central sheath. Their defective activation con- tributes to separate the MT doublets into two
domains (l-5 and 6- 1) and improves the planar beat of the cilium or flagellum. As Taylor (1952) and Holwill suggest (1974) from a hydrodynamic viewpoint, beating on a single plane is more effi- cient. In fact, although 9 + 0 and 9 + 1 flagella (in spermatozoa) are capable of beating in a helical fashion (lshijima et al., 1988) their motion has been reported to be accompanied by rotation of the entire cell body. The helical waves produce a torque resulting in poor propulsion efficiency, for the cell tends to rotate on itself instead of pro- gressing in the medium.
We now proceed to apply our model to the beating mechanism of cilia. Minor additions be- come, however, necessary in order to explain the ciliary beating pattern which is different from flagellar beating. Cilia display in fact an effective stroke, during which the stiffened cilium produces a tonsate (oar-like) movement and a recovery stroke during which a bend initiates at the base of the cilium and propagates aiong the axoneme to reach the starting position cf the effective stroke (Holwill, 1974), see Fig. 9a. The effective stroke occurs in most cases in a siligle plane whereas the
204 E.M. Isinna et al. / BioSystems 39 (1996) 187-226
_/--central MT pair t
MT doublets
7 axoneme
transition
zone
transitional fiber
i
basal body
diary necklace k axoscinal plate
\ transitiial fiber
basal plate
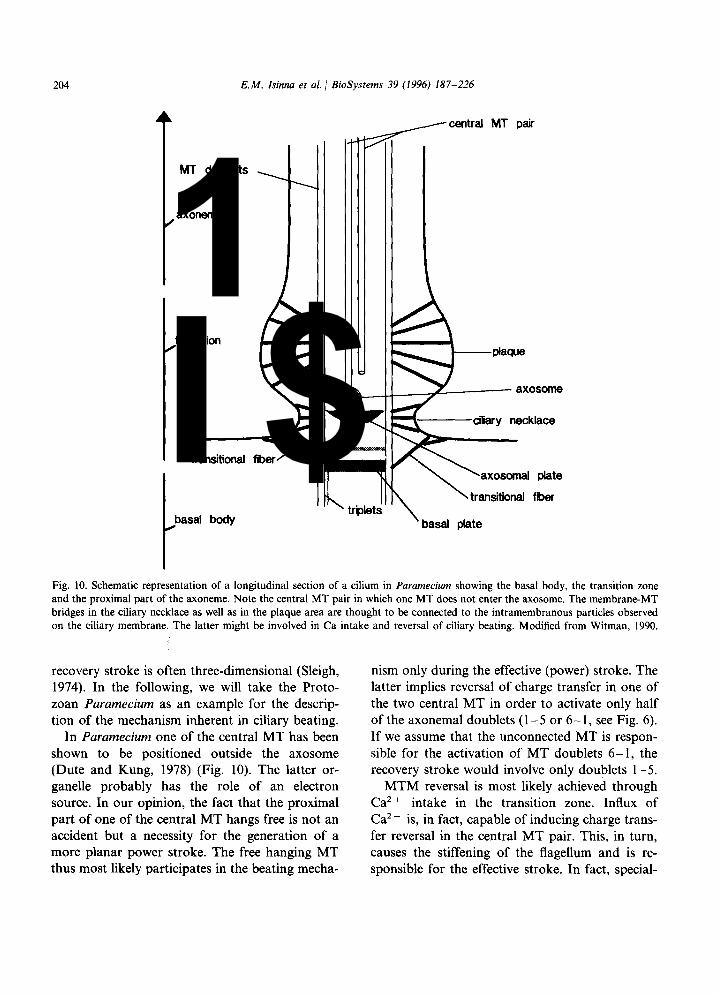
Fig. 10. Schematic representation of a longitudinal section of a cilium in Paramecium showing the basal body, the transition zone and the proximal part of the axoneme. Note the central MT pair in which one MT does not enter the axosome. The membrane-MT bridges in the ciliary necklace as well as in the plaque area are thought to be connected to the intramembranous particles observed on the ciliary membrane. The latter might be involved in Ca intake and reversal of ciliary beating. Modified from Witman, 1990.
recovery stroke is often three-dimensional (Sleigh, 1974). In the following, we will take the Proto- zoan Paramecium as an example for the descrip- tion of the mechanism inherent in ciliary beating.
In Paramecium one of the central MT has been shown to be positioned outside the axosome (Dute and Kung, 1978) (Fig. 10). The latter or- ganelle probably has the role of an electron source. In our opinion, the fact that the proximal part of one of the central MT hangs free is not an accident but a necessity for the generation of a more planar power stroke. The free hanging MT thus most likely participates in the beating mecha-
nism only during the effective (power) stroke. The latter implies reversal of charge transfer in one of the two central MT in order to activate only half of the axonemal doublets (l-5 or 6- 1, see Fig. 6). If we assume that the unconnected MT is respon- sible for the activation of MT doublets 6- 1, the recovery stroke would involve only doublets l-5.
MTM reversal is most likely achieved through Ca’ + intake in the transition zone. Influx of Ca’ + is, in fact, capable of inducing charge trans- fer reversal in the central MT pair. This, in turn, causes the stiffening of the flagellum and is re- sponsible for the effective stroke. In fact, special-

E.M. Isinna et al. / BioSystems 39 (1996) 187-226 205
ized membrane domains with intramembranous particles have been observed to exist in the ciliary necklace (for review see Bardele, 1981) (Fig. 10). Some cilia possess an additional specialised mem- brane domain in the transition zone above the ciliary necklace, the plaque, in which arrays of intramembranous particles are bound to the MT doublets by thin fibers (Dute and Kung, 1978; Allen, 1978; Watanabe, 1990; Witman, 1990; Chailley et al., 1990). It has been suggested that the specialized membrane domains in the necklace as well as the ciliary plaques may bear ionic channels for Ca2 + intake that may be opened by mechanical stimuli (Naitoh and Eckert, 1969, 1974; Allen, 1978; Ogura and Machemer, 1980). We think that the last suggestion is very plausible and that the proteins linking the MT doublets to the membrane (inactivated ATPases) are involved in the control of the Ca ’ + channels for a localized depolarization. The gating mechanism we previ- ously hypothesized in connection with the radial spokes of the axoneme is based on conforma- tional changes of the motility proteins. However, in the case of the Ca2 + gates of the necklace area, simple mechanical forces may induce such confor- mational changes and allow for intake of cations.
Let us try to describe the entire beating cycle by starting at the moment where the cilium is in the middle of its recovery stroke (frame 6 in Fig. 9a). Tentatively, we suggest that mechanical stress of the necklace initiates Ca2+ intake. Due to the three-dimensional helical form of the beating. the ciliary necklace area is, in fact, submitted to a strong torque during the beginning of the recov- ery stroke. Such a stress might be responsible for the opening of the Ca2+ channels. However, oscil- lation of intracellular cyclic nucleotides acting on specific receptors contained in the necklace and plaque area can also be envisaged.
During the recovery stroke, when the cilium is in position 6, Ca2+ starts accumulating in the necklace area in which the two central MT origi- nate. The gradient steeply rises through the regen- erative response of the membrane to Ca2 + while the cilium fulfils its recovery stroke (see Figs. 9b and 15). The more Ca2+ accumulates in the neck- lace area, the slower the bending wave propagates along the cilium because the speed of charge
transfer is directly dependent on the Ca’+ con- centration. The time for the recovery stroke is, thus, much longer than for the effective stroke and may represent almost 2/3 of the total beating cycle. The cilium starts now stiffening because the charge transfer in the central MT is almost stopped. Thus, most axonemal components are inactivated, i.e., in a state of rigor. When the gradient reaches a critical threshold the stiffened cilium abruptly starts its effective stroke which is very short compared to the recovery stroke. The Ca2 + gradient is now quickly lowered by the intervention of cyclic nucleotides and/or calmod- ulin, which are part of the mechanism as sug- gested by several authors (Neitoh and Eckert; Bonini et al., 1991, for review on the role of Ca2 + in ciliary motion see Preston and Saimi, 1990).
In special environments requesting higher propulsion forces through a more powerful effec- tive stroke, we believe that more Ca2+ is neces- sary to achieve faster MT conduction reversal. The ciliary plaques existing in some cilia above the ciliary necklace are probably needed to ensure this additional Ca*+ intake.
The charge transfer hypothesis puts in question the need for central pair rotation which seems to have been observed by several authors (Jarosch and Fuchs, 1975; Kamiya, 1982; Omoto and Kung, 1980; Kamiya et al., 1982). Except perhaps for the alga Micromonas, which has a very spe- cialized configuration of the central pair MT (Omoto and Witman, 1981) we believe that cen- tral pair rotation may not necessarily be an indis- pensable primary distribution mechanism, as suggested by Omoto and Kung (1979). Rotation of the central pair is inconsistent with the exis- tence of the capping structures previously de- scribed. If the central pair should rotate, then it could only be a limited secondary effect produced by a torque which is itself induced by the helical wave generated in the axoneme. Helical motion is, however, what the central pair attempts to coun- teract by producing a planar beating form.
A final comment concerns the dynein arms in both cilia and flagella. As previously suggested, dyneins are used for motility as well as for control and structural purposes. Proximal ATPase activa-

206 E.M. Isinna et al. / BioSystems 39 (1996) 187-226
tion (in the head of the motor protein), is accom- panied in our model by a distal conformational change used as control mechanism for charge transfer. In the case of the dynein arms we do not know if the distal part of the protein is capable of influencing charge transfer in the MT outer dou- blet to which it is attached. Brokaw and Kamiya (1987), however, have observed that mutants flag- ella of Chlumydomonas lacking the internal dynein arms are immobile whereas those lacking the outer arms are mobile but the beating frequency is halved. The only suggestion we have at this stage of theoretical analysis, is that the outer dynein arms are not attached to the MTA from which they protrude in the same fashion as the inner arms, and that they are not in a position to influence current flow. This assumption allows us to understand why mutants lacking the inner arms are immobile. In mutants flagella lacking the internal arms the outer doublets have a defective charge transfer mechanism and outer dynein arms cannot detach from the associated MT.
We think that this is not the place for an exhaustive comparison between our and existing models of axonemal motility and we wish to leave it for future publications. However, we would like to express a few comments mainly in connection with the sliding filament hypothesis of Satir (1968; Warner and Satir, 1974; Sale and Satir, 1977). Firstly, the present model gives a solution for the presence of helical (three-dimensional) waves ob- served in cilia and flagella, a phenomenon the sliding hypothesis cannot account for (for a recent review see Witman, 1990). Secondly, it proposes an alternative answer concerning mechanical in- teractions between axonemal MT tips and the capping structures of cilia and flagella. As Kuhn and ‘Englemann (1978) stated, the sliding mecha- nism raises serious questions in regard to the dimensions of the capping structures. Measure- ments made by Dentler and LeCluyse (1982), have shown that in rabbit and chick tracheal cilia, MT displacement values are not consistent with the amount of sliding predicted by the sliding filament model. In our model, the displacement requested for bending is relatively small and is distributed along the helically disposed axonemal components. Thirdly and lastly, we think that
charge transfer-activation of motor proteins and related distal conformational changes is the only mechanism capable of accounting for the long searched synchronization mechanism responsible for the progressive propagation of the bending wave along the axoneme.
3. Part II - two application examples of MTM: Euglene gracilis’ phototaxis and vertebrate photoreceptors
The main features of our model described in Part I will now help us to shed a new light on other ciliary structures, such as vertebrate pho- toreceptors. However, before we proceed with the analysis of the role of cilia in vertebrate photore- ceptors, we should first consider how the complex dynamics of MTM has been implemented in the ‘primitive’ ciliary photoreceptor of a protozoan alga, Euglena gracilis. Although of simple archi- tecture, it contains some important basic features and components.
3.1. Euglena gracilis
3.1.1. Euglena’s photoreceptor and phototaxis Invertebrate photoreceptors may be composed
of arrays of microvilli protruding from an exten- sion of the outer cell membrane (the rhabdom) or they consist of one or more ciliary arrangements forming the membrane array. Cilia occur in many eyes of invertebrate organisms and ciliary pho- toreceptors have been investigated in mollusca, annelids, coelenterates, echinoderms (for review see Eakin, 1972; Barber, 1974).
Yet more primitive organisms, such as algae, show simple perceptive functions like phototaxis implying the use of a functional photoreceptor organ. The unicellular Euglena gracilis has been the object of studies because of its phototactic capabilities (for review see Feinleib and Curry, 1971; Piccinni and Omodeo, 1975; Buetow, 1968a, 1968b, 1982, 1989).
Euglena moves by the propulsive force of the 50 pm long beating flagellum in which roughly heli- cal bending waves propagate along its structure. The 50 pm long and 10 pm wide cell thus moves in a helical fashion by spinning along its axis with
E.M. Isinna et al. 1 BioSystems 39 (1996) 187-226 207
paraflagdar swellhg____
contractile vaato@_
emergent flag&n
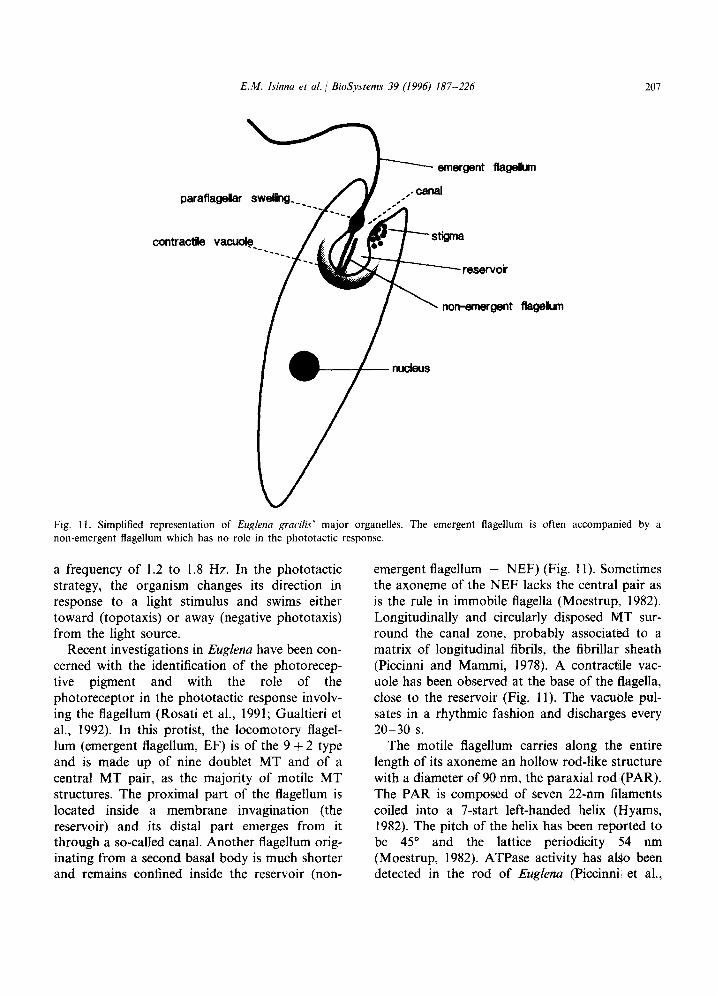
Fig. Il. Simplified representation of Euglena gracih major organelles. The emergent flagellum is often accompanied by a non-emergent flagellum which has no role in the phototactic response.
a frequency of 1.2 to 1.8 Hz. In the phototactic strategy, the organism changes its direction in response to a light stimulus and swims either toward (topotaxis) or away (negative phototaxis) from the light source.
Recent investigations in Euglena have been con- cerned with the identification of the photorecep- tive pigment and with the role of the photoreceptor in the phototactic response involv- ing the flagellum (Rosati et al., 1991; Gualtieri et al., 1992). In this protist, the locomotory flagel- lum (emergent flagellum, EF) is of the 9 + 2 type and is made up of nine doublet MT and of a central MT pair, as the majority of motile MT structures. The proximal part of the flagellum is located inside a membrane invagination (the reservoir) and its distal part emerges from it through a so-called canal. Another flagellum orig- inating from a second basal body is much shorter and remains confined inside the reservoir (non-
emergent flagellum - NEF) (Fig. 11). Sometimes the axoneme of the NEF lacks the central pair as is the rule in immobile flagella (Moestrup, 1982). Longitudinally and circularly disposed MT sur- round the canal zone, probably associated to a matrix of longitudinal fibrils, the fibrillar sheath (Piccinni and Mammi, 1978). A contractile vac- uole has been observed at the base of the flagella, close to the reservoir (Fig. 11). The vacuole pul- sates in a rhythmic fashion and discharges every 20-30 s.
The motile flagellum carries along the entire length of its axoneme an hollow rod-like structure with a diameter of 90 nm, the paraxial rod (PAR). The PAR is composed of seven 22-nm filaments coiled into a 7-start left-handed helix (Hyams, 1982). The pitch of the helix has been reported to be 45” and the lattice periodicity 54 nm (Moestrup, 1982). ATPase activity has also been detected in the rod of Euglena (Piccinni et al.,
208 E.M. Isinna et al. / BioSystems 39 (1996) 187-226
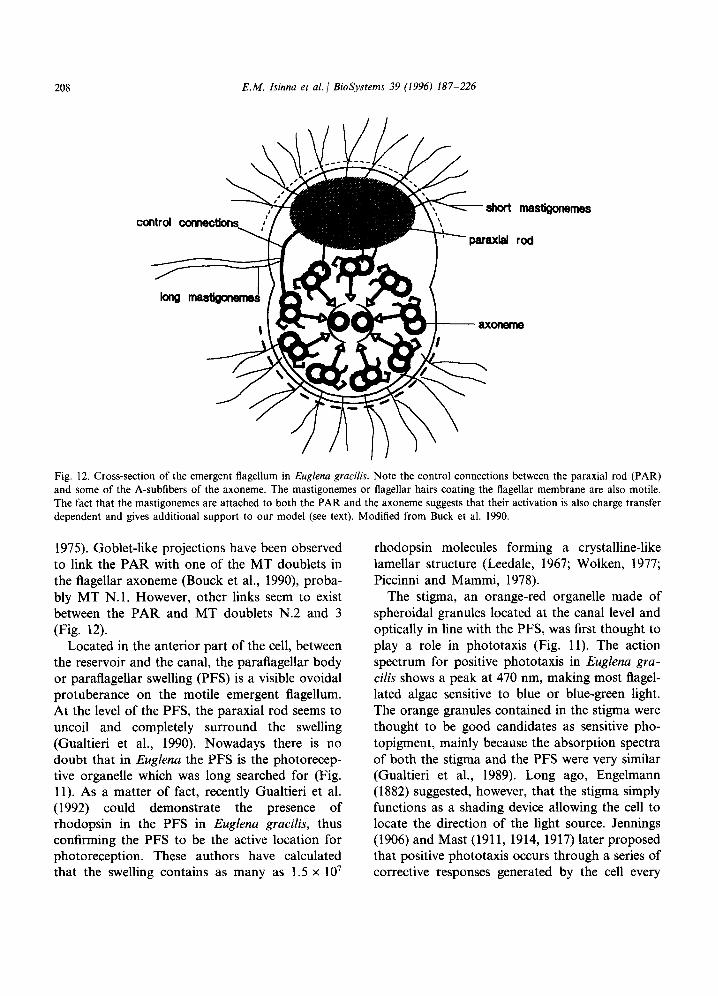
Fig. 12. Cross-section of the emergent flagellum in Euglena gracilis. Note the control connections between the paraxial rod (PAR) and some of the A-subfibers of the axoneme. The mastigonemes or flagellar hairs coating the flagellar membrane are also motile. The fact that the mastigonemes are attached to both the PAR and the axoneme suggests that their activation is also charge transfer dependent and gives additional support to our model (see text). Modified from Buck et al. 1990.
1975). Goblet-like projections have been observed to link the PAR with one of the MT doublets in the flagellar axoneme (Bouck et al., 1990), proba- bly MT N.l. However, other links seem to exist between the PAR and MT doublets N.2 and 3 (Fig. 12).
Located in the anterior part of the cell, between the reservoir and the canal, the paraflagellar body or paraflagellar swelling (PFS) is a visible ovoidal protuberance on the motile emergent flagellum. At the level of the PFS, the paraxial rod seems to uncoil and completely surround the swelling (Gualtieri et al., 1990). Nowadays there is no doubt that in Euglena the PFS is the photorecep- tive organelle which was long searched for (Fig. 11). As a matter of fact, recently Gualtieri et al. (1992) could demonstrate the presence of rhodopsin in the PFS in Euglena grucilis, thus confirming the PFS to be the active location for photoreception. These authors have calculated that the swelling contains as many as 1.5 x 10’
rhodopsin molecules forming a crystalline-like lamellar structure (Leedale, 1967; Wolken, 1977; Piccinni and Mammi, 1978).
The stigma, an orange-red organelle made of spheroidal granules located at the canal level and optically in line with the PFS, was first thought to play a role in phototaxis (Fig. 11). The action spectrum for positive phototaxis in Euglena gra- cilis shows a peak at 470 nm, making most flagel- lated algae sensitive to blue or blue-green light. The orange granules contained in the stigma were thought to be good candidates as sensitive pho- topigment, mainly because the absorption spectra of both the stigma and the PFS were very similar (Gualtieri et al., 1989). Long ago, Engelmann (1882) suggested, however, that the stigma simply functions as a shading device allowing the cell to locate the direction of the light source. Jennings (1906) and Mast (1911, 1914, 1917) later proposed that positive phototaxis occurs through a series of corrective responses generated by the cell every

E.M. Isinna et al BioSystems 39 (1996) 187-226 209
rotation
a b
Fig. 13. Phototactic mechanism of Euglena grads according to Mast (1917). As long as the incoming light is within the rotation cone, the cell swims toward the light source. When the stigma casts a shadow on the photoreceptor, the flagellum is erected and the cell changes its direction accordingly. Modified from Piccinni and Omodeo, 1975.
time the stigma casts a shadow on the photore- ceptor. The cell usually swims leaving its photore- ceptor permanently illuminated (Fig. 13). When the light beams are shaded by the stigma, the cell responds with an erection of the flagellum (shock or bending reaction) which results in a change of direction. Bancroft (1913) noted that the stiffen- ing of the flagellum is proportional to the shading time. These last suggestions have, today, been widely accepted (Feinleib and Curry, 1971; Fein- leib, 1985).
Although, today, there is no doubt of the role of the PFS in the photoreceptor mechanism, the dynamics of the entire structure has not yet been elucidated.
Several suggestions have been made trying to connect Euglena’s various phototactic strategies with the photoreceptor mechanism. Surek and Melkonian (1986) advanced a hypothesis that the MT of the stigma membrane somehow establish a
connection between the stigma and the PFS, thus modulating the light impinging on the photore- ceptor.
Piccinni and Omodeo (1975) suggested instead the existence of a synaptic type junction between the non-emerging flagellum and the plasma mem- brane of the motile flagellum. A comparator was further assumed to exist at the base of the.roots of the flagella. The latter would give its infarmation to the effector located in the canal zone. When the stigma casts a shadow on the PFS, the ,effector composed of the MT and associated fibrils in the canal induces a contraction and modifies the beat- ing angle of the motile flagellum allowing’ the cell to remain on the light track. In a way, it acts as a rudder by modifying the inclination of the flagel- lum with respect to the cell body. Such junctions have, however, not been confirmed (Osafune and Schiff, 1980; Gualtieri et al., 1990; Rosati et al., 1991).

210 E.M. Isinna ef al. / BioSystems 39 (1996) 187-226
A more sophisticated suggestion involving piezoelectric features of the MT in the axoneme has been made by Bovee and Jahn (1972). In their model, the paraxial rod would receive electronic information (a potential) from the PFS when the latter is illuminated. Rosati et al. (1991), on the other hand, have based their model on an idea of Piccinni et al. (1975), as well as on the previously mentioned piezoelectric theory of Bovee and Jahn (1972) and Hameroffs MT information process- ing properties (Hameroff, 1982, 1984). They pro- pose that the PAR might be contractile and that the signal transduced by the PFS would propa- gate distally by means of PAR contraction spreading through the entire MT system, i.e. the cytoskeleton. They further suggest the possible involvement of the non-emergent flagellum in the phototactic response through contraction of the flagellar root system.
Eakin (1968) advanced that the sophisticated vertebrate photoreceptor is the result of an evolu- tive pathway implementing ciliary structures which started with Euglena. If Eakin is right, by investigating Euglena, we should find in this Pro- tozoan some of the basic elements constituting the ciliary photoreceptors of animals ranking higher in the evolutive pathway such as vertebrates. If viewed from the angle of the model for flagellar motility we previously exposed, Euglena indeed possesses some important general features.
3.1.2. Euglena’s phototactic mechanism a. The phototactic mechanism. The analysis of
Euglena’s sophisticated phototactic mechanism will lead us to additional characteristics of MTM dynamics and particularly to the probable exis- tence of MTM-membrane interactions capable of influencing cell polarization.
We believe that the basic principle of charge transfer control we previously postulated for the radial spokes of cilia and flagella has been imple- mented in Euglena’s photoreceptor architecture in a similar way. We, thus, suggest that the goblet shaped projections connected to the doublets 1 to 3 of the axoneme (Figs. 12 and 14) work as gates for the control of the axonemal current. We will henceforth call them gate projections. The activa- tion of the gate projections happens most proba-
bly through distally directed charge transfer occurring inside the helically disposed PAR filaments. We now outline Euglena’s phototactic capabilities as follows (Fig. 14):
The incoming light bleaches the rhodopsin molecules contained in the paraflagellar swelling. Isomerization of 1 I-cis-retinal group to all-trans- retinal, by means of an enzymatic cascade, in- duces changes in the charge transfer capability of PAR filaments. Whether this happens through previous changes in PAR membrane permeability or not remains an open question.
As previously mentioned, ATPase activity has been detected in the rod of Euglena by Piccinni et a:. (1975). Charge transfer activates the ATPase of the gate projection on the paraxial rod. ATP hydrolysis induces a conformational change in the distal part of the gate projection attached to the axonemal MT doublet and the conduction path is made free. The current flow on axonemal doublet MT1 may thus occur as long as electron flow in the PAR continuously activates ATP hydrolysis. Charge transfer inside the PAR filaments and subsequent ATP hydrolysis in the rod are thus light-dependent.
In order to achieve more efficient response, the control pathway probably involves more than one MT doublet as shown in Fig. 12. Every time light impinges on the detector, the current flow in the axonemal MT doublets is proportionally in- creased and so is the beat frequency of the flagel- lum. Casting a shadow on the stigma produces instead a decrease in ATP hydrolysis and current flow in the MT doublets with a subsequent pro- gressive stiffening of the corresponding axonemal components. The beating pattern of the flagellum and, thus, the cell’s swimming path are modified accordingly. The stiffened fibers actually result in transforming the beat form of the flagellum so that it can work like a rudder. Therefore, shadow- ing of the PFS by the stigma helps Euglena find the direction of the light beam. The right trajec- tory is kept by means of successive corrections involving the flagellum beat direction.
This model exactly supports the intuitive sug- gestion of Piccinni and Omodeo (1975) who, how- ever, had envisioned that the rudder function of the flagellum was to be found in the contraction

E.M. Isinna et al. 1 BioSystems 39 (1996) 187-226 211
Fig. 14. Phototactic response control mechanism in Euglena gracilis. For the sake of snnplicity, the paraxial rod is shown with only three fibers instead of seven. Charge transfer inside the PAR is suggested to depend on the bleaching of rhodopsin molecules contained in the paraflagellar swelling. This in turn controls the activity of the axoneme via the goblet-shaped projections connected to the B-subfiber. The cell can, thus, react to light changes. The phototactic response is based on a completely automatic mechanism.
of the matrix of longitudinal fibrils located in the canal zone. Those fibrils have a more natural function: they control the periodic contraction of the canal and relative vacuole.
The mechanism inherent in the vacuole dynam- ics becomes also understandable if we envisage a connection between the MT and the fibrillar sheath. The contractile vacuolar region is encir- cled by two MT arrays. One consists of circular, the other of longitudinal MT arranged in pairs (Piccinni and Omodeo, 1975; Willey and Wibel, 1987). The latter are indispensable if the contrac- tion must progressively spread from the bottom toward the top of the canal to empty it. In fact, the longitudinal MT set is most likely to com- mand the progressive contraction of the canal and vacuolar region through interactions with the fibrillar sheath. This sheath probably consists of motor or actin-like proteins. The connection of
MT with arrays of actin is a major point which we will consider again later on in this paper in connection with vertebrate photoreceptors.
6. Negative phototaxis and MTM-membrane in- teractions. Euglena’s phototactic response to a sharp increase in light intensity results in a nega- tive phototaxis. The flagellum is first completely straightened out and the cell subsequently moves backward during a short period (Bancroft, 1913). Two major implications are contained in negative phototaxis. The first concerns the capability of arresting charge transfer inside the MT doublets in order to induce an artificial state of rigor in the doublets controlled by the PAR gate projec- tions.
Upon intense illumination, the sudden, bleach- ing of the rhodopsin pigments completely stops charge transfer in the PAR. Consequently, all the gates connected to the relative MT doublets are

212 EM. Isinna et al. / BioSystems 39 (1996) 187
Mechanical stress
226
T ca PunDed out~Kali,
I 1 1 Reversed beat Normsl beat
Fig. 15. In the original hypothesis (Eckert, 1972), a depolarizing stimulus such as an outward current induces a transient increase in the conductance of the cell membrane to Ca*+(g Ca). The subsequent rise of Ca’+(I Ca), on the other hand, results in reversal of the ciliary beating. Active processes (such as the activation of calmodulin) are able to restore the initial state. The arrow between I Ca and depolarization shows the positive feed-back mechanism. In our model, the increase in membrane conductance is dependent on mechanical stress (as in the cilium) and not on an external depolarizing stimulus. Additionally, it is probable that changes in the conduction state of the MTM also indirectly affect the polarization of the cell membrane (probably via cyclic nucleotides). Modified from Naitoh and Eckert, 1974.
closed, i.e., all electronic circuits along the entire length of the concerned MT axonemal doublets are switched off. This results in complete stiffen- ing of the flagellum (phototaxis).
The second implication concerns MTM reversal needed for negative phototaxis. This implies that somehow the cell membrane is depolarized in order to produce the necessary reverse voltage VR and allow charge transfer to occur in the opposite sense along the entire axoneme.
Observations in other Protozoa have, in fact, shown that the direction of phototaxis in the alga Platymonas is directly dependent on the concen- tration of cations in the medium (Halldal 1957, 1959). When the ratio of Mg* + to Ca*+ (with K + very low) is around 6: 1, the direction is random; above this value the cell displays positive and below, negative phototaxis. When the cul- tures are illuminated at a stage where orientation
is close to random, the sign may be reversed by cell illumination (quoted in Feinleib and Curry, 1971). Additionally, it has been noted that, at high light intensities, the response in Chlamy- domonas changes from positive to negative in a few seconds (Feinleib and Curry, 1971).
Taking into account the present Ca-current hy- pothesis (Eckert, 1972; Naitoh and Eckert, 1974, see Fig. 15) the only mechanism we can envisage for negative phototaxis is that membrane depolar- ization in Euglena is somehow connected with the changes in current flow occurring inside the MTM. Any perturbation of the MTM most prob- ably affects the entire cytoskeleton of the cell. This may in turn influence, more or less, the permeability of the membrane to cations, perhaps via cyclic nucleotides and/or membrane-MT in- teractions. As a consequence of the intake of Ca*+ the MTM, i.e., the flagellum, starts work- ,

E.M. Isinna et al. i BioSystems 39 (1996) 187-226
kM
M Gate P P -
- T4 A 4 - F 4
M R s t-------
4
C
213
M u Fig. 16. Schematic diagram of the putative control mechanism in Euglena grucilis. In the normal state (positive phototaxis), the more light impinges on the paraflagellar swelling (PFS), the more the charge transfer mechanism is activated inside the paraxial rod (PAR). As a result, the membrane (CM) conductance to cations decreases and the cell hyperpolartzes. This results in an increase in the beating frequency of the flagellum and in an acceleration of the cell in the medium (toward the light source). In negative phototaxis, instead, proximity of the light source induces the complete bleaching of the rhodopsin molecules. Charge transfer inside the PAR is stopped and the MTM is stalled. The membrane conductance to calcium increases and Ca*+ enters the cell. The flagellum is first in a state of rigor, then as soon as the Ca’+ concentration is htgh enough it starts beating in reverse until an regulating process (probably calmodulin) sets in and eliminates the surplus in Caz+.
ing in reverse over a short distance. Euglena now swims away from the light source until the mem- brane slowly recovers its usual conductance con- ditions through Ca*+ removal by active processes. In the normal state of positive photo- taxis, a feedback phenomenon is instead likely to occur: the more light reaches the PFS, the more current flows throughout the MTM and the more the membrane hyperpolarizes because the Ca-gat- ing mechanism is impaired. This in turn increases the beating frequency of the flagellum (Fig. 16).
However, only future investigations will enable us to answer the question of the existence of MTM-membrane interactions.
3.2. Vertebrate photoreceptors
The presence of ciliary structures in sense or- gans is still an unsolved mystery. The function of the cilium, connecting the inner to the outer seg- ment in vertebrate photoreceptors, has not yet been elucidated. Its role in protein transport is unclear and its participation in the electrodynam- ics of generator potentials awaits elucidation.
We have gone a long way, from the molecular basis of charge transfer to axonemal dynamics in flagellar beat via charge transfer-activation of mo- tor proteins. This detour was, however, necessary in order to, at least partly, elucidate the role of the cilium in vertebrate photoreceptors.
We are of the opinion that the need for faster response in the photoreceptors of vertebrates has imposed the use of a cilium. In fact, ciliary pho- toreceptors of invertebrates are usually faster al- though less sensitive compared to those bearing microvilli. The presence of an MTM generates, in turn, some specificities imposed by the MTM electrodynamics. Those specificities can be sum- marized as follows: a. Resting potential of the cell membrane adapted to the requirements of the MTM; b. Ciliary transition zone as a potential barrier between the cilium and the cell’s interior and as isolating medium between ciliary and cell plasma membrane; c. Indirect link between MTM and cell membrane for polarization control;

214 E.M. Isinna et al. / BioSystems 39 (1996) 187-226
d. Response to light stimuli dependent on MTM conditions; e. Interactions between MT and motility-related proteins.
This is what we intend to investigate in the following sections.
3.2.1. General structure Vertebrate photoreceptors are divided into rods
and cones (for review see Cohen, 1972, Crescitelli, 1972, Schnapf and Baylor, 1987, McNaughton, 1990). Rods, with a diameter of 1 pm and a length of 40 pm, are very sensitive to low light stimuli and are capable of detecting single pho- tons. They are involved in monochromatic vision with a broad sensitivity spectrum. Cones are much less sensitive and are involved in color vision requiring bright light. In the development of our hypothesis we will consider the features and dynamics of rods only, for the differences between these photoreceptor types are not always relevant for our investigation (see Fig. 17).
Rods are composed of an outer segment (OS) containing a stack of 160 A thick disks (up to ZOOO), originating from invaginations of the cell membrane and bearing photoreceptor pigments (rhodopsin) responsible for light absorption and response. The disk membranes are completely iso- lated from the plasma OS membrane. They are continually renewed and have a lifetime of about 1 month (Young, 1970).
The inner segment (IS) contains in its distal part the usual Golgi complex with endoplasmic reticulum, and a relatively high number of mito- chondria located near the base of the cilium. In the nuclear portion we find the cell nucleus, and in its proximal part the synaptic body connected together by the inner fibre or axon, very similar to the axons of nerve cells. MT are present in the IS as in other cell types. Particularly in the axon, MT are very dense and regularly ordered as in nerve cells. Some specific type of MT-like structures has recently been discovered in the distal part of the OS between the ends of the axonemal MT and the cell membrane (Roof et al., 1991). The cilium connecting the IS with the OS is of the 9 + 0 type, with nine pairs of MT fibers in the axoneme. The central pair of MT, present in mobile cilia and
flagella, is lacking and so are the dynein arms (the ATPase responsible for the beating) located be- tween the MT doublets. The cilium stems from a centriole (the basal body) to which sometimes a striated root is connected. A second centriole is usually located near the first similarly to somatic cell diplosomes (De Robertis and Lasansky, 1961).
3.2.2. Electrophysiology Intracellular recordings have shown that during
the dark adapted state, the photoreceptor’s IS is depolarized with a resting potential of about - 10 to - 40 mV (Tomita, 1971). Hagins (1970) has found that the cell’s energy consumption is much higher in the dark than during illumination. A dark current of ionic nature flows from the IS to the OS through the surrounding extracellular space, such as to make the distal tips negative.
Fig. 17. Schematic representation of a vertebrate rod photore- ceptor. The outer segment containing the disks carrying the rhodopsin molecules is connected to the inner segment via an immobile cilium lacking the central MT pair.

EM. Isinna et al. 1 BioSystems 39 (1996) 187-226 215
Consequently the inner of the OS becomes positive with respect to the IS inner. This current is main- tained through a gradient generated by an active Na + /K + -ATPase pump located in the rod IS and involves light sensitive Na+ -Ca2 + channels and an Na+~ -Ca2+, K+ exchange mechanism in the OS (Tomita, 1971; Penn and Hagins; 1972; Mc- Naughton, 1990; Perry and McNaughton, 1991 and 1993).
Upon illumination, the cell responds with a hyperpolarization ( - 1 to - 20 mV depending on the intensity of the stimulus). The dark current is abruptly reduced by a resistance change in the OS membrane. When exposed to light, the OS mem- brane resistance increases (i.e. its conductance decreases) by several MR up to a saturation point.
The mechanism leading to the cell’s hyperpolar- ization, i.e. decrease in membrane conductance of Na+ and Ca2+ ions has been thoroughly investi- gated during the last decade and the original hypothesis of Hagins (1970) has been experimen- tally confirmed. (Stryer, 1986; Fesenko et al., 1985; Perry and McNaughton, 1991 and 1993).
At the danger of oversimplification, the mecha- nism can be described as follows: Rhodopsin molecules in the OS membrane are composed of retinal and opsin. Light induces the isomerization of the 1 I-cis-retinal group to all- trrtns-retinal leading to a cascade of transient photolytic intermediate forms of rhodopsin. This, in turn, triggers an enzymatic cascade involving transducin (T-GTP) and activated phosphodi- esterase (PDE) with subsequent hydrolysis of cyclic GMP (guanylate). The drop in GMP level closes the cation specific channels (mainly Na + ) in the OS membrane thus inducing a decrease in conductance with subsequent hyperpolarization of the photoreceptor’s IS.
In the IS, during the dark adapted state, a steady release of neurotransmitter (L-glutamate) prevents the connected neurons (bipolar cells) from firing. The photoreceptor’s response is said to be inhibitory because the hyperpolarization inhibits the Ca2+ influx at the synapse and thus induces a decrease in transmitter release. This results in the excitation of the post-synaptic neu- rons and signalling of the light stimulus upward into the neural pathway.
3.2.3. MTM dynamics in vertebrate photoreceptors Investigations of photoreceptors response
started with measurements in the retinae of inver- tebrate eyes because some of them possess pho- toreceptors large enough to permit direct electrical monitoring of a single cell. The generator potential of most microvilli-based invertebrate photorecep- tors was found to be generally a depolarization in many animals. As soon as the development of special microtechniques allowed the measurement of vertebrate ciliary photoreceptors, they were shown to respond with a hyperpolarization of the cell membrane. This was indeed a surprise, for the pigment (rhodopsin) contained in both vertebrate and invertebrate photoreceptors had been proven to be consistently the same. Why should vertebrate photoreceptors hyperpolarize in response to light stimuli and use a far more complicated electro- physiological response scheme?
a. MTA4 requirements and OSpolarizadion in the dark-adapted state. In the dark adapted state, the vertebrate photoreceptor cell consumes more en- ergy than during illumination. This is not a logical choice unless some important reasons have com- pelled evolution to such an energy-consuming so- lution. From the viewpoint of our model, however, this is the only way to ensure proper functioning of the MTM located between the two segments. The MTM requires, in fact, a potential difference to exist between the IS and OS. This OS depolarization, i.e., ciliary polarization, has been achieved by the use of active ion transport in the IS (the Na + /K + -ATPase) coupled with a complex interplay of ion exchanges in the OS membrane (the Na + -Ca’+, K + exchange mechanism) (Perry and McNaughton, 1993). Because of this distally directed current, the photoreceptor’s OS acts as a current sink and is, in the dark, several mV more positive than the IS. The dark current generates a polarizing potential (bias) allowing the negative charges (e - ) to be preferentially transferred dis- tally along the MT structure, thus flowing from the basal body toward the tips of the axoneme. A bias of a few mV suffices to ensure disk renewal as well as transport of cytosolic components to and from the OS. The OS thus acts as a huge,capping
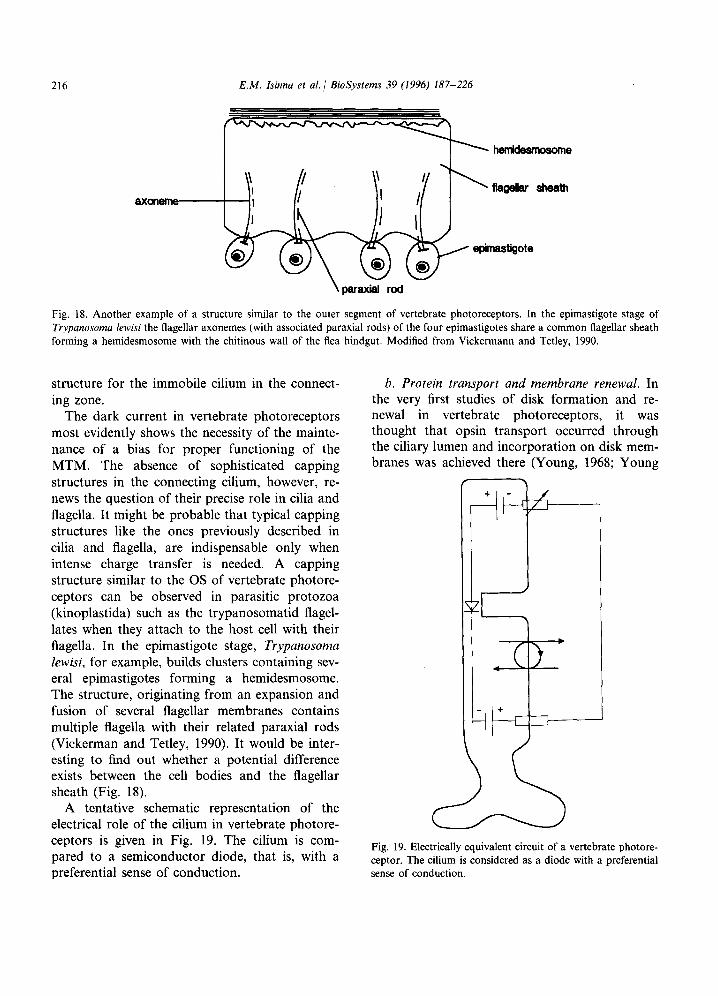
216 E.M. Isinna et al. / BioSystems 39 (1996) 187-226
axame
Fig. 18. Another example of a structure similar to the outer segment of vertebrate photoreceptors. In the epimastigote stage of Trypanosoma lewisi the flagellar axonemes (with associated paraxial rods) of the four epimastigotes share a common flagellar sheath forming a hemidesmosome with the chitinous wall of the flea hindgut. Modified from Vickermann and Tetley, 1990.
structure for the immobile cilium in the connect- ing zone.
The dark current in vertebrate photoreceptors most evidently shows the necessity of the mainte- nance of a bias for proper functioning of the MTM. The absence of sophisticated capping structures in the connecting cilium, however, re- news the question of their precise role in cilia and flagella. It might be probable that typical capping structures like the ones previously described in cilia and flagella, are indispensable only when intense charge transfer is needed. A capping structure similar to the OS of vertebrate photore- ceptors can be observed in parasitic protozoa (kinoplastida) such as the trypanosomatid flagel- lates when they attach to the host cell with their flagella. In the epimastigote stage, Trypanosoma Zewisi, for example, builds clusters containing sev- eral epimastigotes forming a hemidesmosome. The structure, originating from an expansion and fusion of several flagellar membranes contains multiple flagella with their related paraxial rods (Vickerman and Tetley, 1990). It would be inter- esting to find out whether a potential difference exists between the cell bodies and the flagellar sheath (Fig. 18).
A tentative schematic representation of the electrical role of the cilium in vertebrate photore- ceptors is given in Fig. 19. The cilium is com- pared to a semiconductor diode, that is, with a preferential sense of conduction.
6. Protein transport and membrane renewal. In the very first studies of disk formation and re- newal in vertebrate photoreceptors, it was thought that opsin transport occurred through the ciliary lumen and incorporation on disk mem- branes was achieved there (Young, 1968; Young
Fig. 19. Electrically equivalent circuit of a vertebrate photore- ceptor. The cilium is considered as a diode with a preferential sense of conduction.

E.M. Isinna et al. /( BioSystems 39 (1996) 187-226 217
d b)
Fig. 20. Schematic representation of the process leading to membrane renewal in vertebrate photoreceptors. The actin bundles elongate through interactions with alpha-actinin in the presence of Ca. Progressive elongation of the actin bundle is accompanied by distally directed displacement via MAP-mediated intracellular transport. The membrane “blob” grows and is finally inserted as a disk into the outer segment. However MTM-activated actin elongation (via MAPS) should not be excluded.
and Droz, 1968). Later studies have suggested, (Besharse, 1986). As for the formation of the however, that opsin is not transferred through disks, Steinberg et al. (1980) have proposed a the ciliary pathway and that it is incorporated model in which the disks are formed through an into the membrane in the IS. (Papermaster et evagination process of the ciliary membrane al., 1975; Besharse and Pfenninger, 1980; starting in the proximal part of the transition Besharse et al., 1985). The transport of opsin zone. Although it is not our intention to expand molecules from the site of synthesis in the endo- on opsin transport, which should be the subject plasmic reticulum to the distal part of the IS, in of further experimental investigations, our model the ellipsoid, has not yet been elucidated. Ac- can contribute to shed some light on the process cording to recent investigations, MT seem not to of disk formation and delivery of cytosolic com- be involved (Vaughan et al., 1989). ponents into the OS.
Presently, the most plausible model involves transport of opsin molecules in the form of vesi- cles that are incorporated as buds into the plasma membrane. Opsin fuses later with the cil- iary plasma membrane which evaginates to form OS disks (Besharse, 1986) Fig. 20a. Alterna- tively, this same author has suggested that extra- cellular vesicle transport might be possible. However, as he notes, such vesicles have never been observed in the extracellular space
Concerning disk formation, we propose here that MTM-actin interactions are at the core of disk/membrane renewal process.
Discrete concentrations of short segments of filamentous actin (F-actin) and cc-actinin have, in fact, been discovered at the site of disk mor- phogenesis within the membrane of the connect- ing cilium but are absent in the OS between mature disks (Arikawa and Williams, 1988; Chaitin, 1992).

218 E.M. Isinna et al. 1 BioSystems 39 (1996) 187-226
Interactions between cr -actinin and F-actin are probable and it has been observed that a-actinin is able to bundle actin filaments in vitro (Jockusch and Isenberg, 1981; Stossel et al., 1985). This phenomenon has already been observed to occur in brain and other non-muscle cells and seems to be dependent on calcium concentrations (Duhaiman and Bamburg, 1984).
Both previously quoted models of Besharse (1986) and Steinberg (1980), become more plausi- ble if combined together with the ciliary MTM into a tentative model that can roughly be de- scribed as follows:
Opsin vesicles are incorporated in the plasma membrane intracellularly in the ellipsoid region. The short actin filaments attach to the ciliary (opsin carrying) membrane via cl-actinin. At the same time, they are elongated in the form of parallel bundles by interaction with cr-actinin ac- cording to the concentration gradient of Ca molecules present along the ciliary structure in the transition zone. The fluid membrane follows the elongation of actin filaments while it is continu- ously renewed in the ellipsoid. The more mem- brane is available, the longer becomes the bundle of actin filaments that elongates horizontally. Ad- ditionally, we believe that an MTM/motor protein-based intracellular transport mechanism ensures the distally directed displacement of the membrane evagination (blob) (see Fig. 20b). This results in a progressive displacement of the initial blob which transforms itself into a larger and flattened disk during its distal motion. The new disk is finally inserted into the OS at the bottom of the stack and the entire stack is shifted upward (Fig. 20a).
The fact that in cells treated with cytochalasin D the lateral growth of new disks is not inhibited (Williams et al., 1988) may be explained by consid- ering that though the drug affects the elongation of actin filaments, it does not stop the process of membrane synthesis nor the MTM activity. The serial insertion of membrane components, which is also partly responsible for the outward grow of the blob, is likely to continue and the existing blobs form into new although defective disk-like struc- tures. New membrane evaginations are however inhibited (Chaitin, 1992; Arikawa, 1988; Williams et al., 1988).
To sum up, MTM-dependent intracellular transport (through kinesin-like MAPS) and elon- gation of bundles of short F-actin filaments are, thus, probably responsible for shaping new disks by controlling the horizontal growth while pro- gressively shifting the disk upwards along the axoneme. However, other factors than Ca and a-actinin may also be involved in actin filament elongation such as direct MTM/MAPs-actin inter- actions (Selden and Pollard, 1986; Schmit and Lambert, 1985).
Finally, a last few words should be spent con- cerning the transport of all enzymes and proteins synthesized in the IS. The distally directed charge transfer ensures their dynamic transport through the axonemal lumen into the OS. As long as the MTM is working forward, it creates a natural barrier against back diffusion of the delivered components.
Immunocytochemical studies have shown that rapid shift in immunoreactivity occur in both OS and IS (Broekhuyse et al., 1985; for discussion see Besharse, 1990). The changes in immunoreactivity for several proteins seems to be light-dependent. In connection with our model, this means that transport between OS and IS is indeed likely to occur through the connecting cilium and is a function of the MTM conduction state, a subject we will expand on in the next subsection.
Proper functioning of the MTM and related transport, as well as accelerated protein synthesis in the IS requires large amounts of energy (ATP). This energy is supplied by an unusually large number of mitochondria tightly packed in the ellipsoid region. The association of MTM with mitochondria is a general rule imposed by the MTM functional requirements and can be ob- served in many cell types.
c. Hyperpolarizing response to light. We fully agree with Besharse and Horst, (1990) who ad- vanced a hypothesis that one of the main functions of the cilium is the maintenance of discrete mem- brane domains. In vertebrate photoreceptors, the efficiency of the photosensitive mechanism includ- ing membrane renewal and protein transport is based on such a separation between the inner and outer segment. It is a demarcation zone electrically isolating two completely different domains.

E.M. lsinna et al., Bidystems 39 (1996) 187-226 219
The existence of such distinct membrane spe- taneously, does not need to be based on the same cializations has already been advanced in connec- principles as OS hyperpolarization. We tentatively tion with cilia and flagella of protozoa (Musgrave suggest that, in vertebrate photoreceptors the hy- et al., 1986; Bloodgood, 1990). The membrane perpolarizing response of the IS is only subse- near the transition zone, is such a specialized quently induced through indirect MTM signalling domain separating the ciliary membrane from the to the IS membrane. During cell illumination, cell body plasma membrane. Machemer and hyperpolarization of the OS has the result of Ogura (1979) have found that ciliary membranes affecting the conduction state of the MTM be- have very high transmembrane resistances (proba- cause of changes in the forward bias. The new bly the highest of all) and that for this reason they charge transfer conditions induce changes in the are capable of conducting electrical signals cytoskeleton and subsequently in the IS mem- (Machemer, 1986; Witman, 1990). We think in- brane which in turn hyperpolarizes. The involved stead that such very high resistance is particularly lag phase between OS and IS hyperpolarization is well suited to isolate the cilium from the cell probably too short to be detected. Even if de- membrane and this factor, in our view, plays a tected, such a propagation delay time could be fundamental role in vertebrate photoreceptor elec- interpreted as being the result of membrane ca- trophysiology. pacitances.
We propose here that the resulting isolation prevents direct passive propagation of the hyper- polarizing response from the OS to the IS and, by extension, to the synaptic area. Additionally, such a separation is needed for subsequent hyperpolar- ization of the IS by means of the more or less indirect MTM-plasma membrane interactions previously suggested in connection with Euglena’s negative phototaxis. There, such interactions be- tween MTM and cell membrane may explain MTM reversal, i.e. membrane depolarization. Therefore, it is probable that a similar mechanism exists in vertebrate photoreceptors as well. How- ever, in this case, the change in the MTM conduc- tion state should induce a hyperpolarization by affecting some ionic channels located in the IS membrane.
We have seen how the requirements of the MTM have conditioned the cell’s electrical ground state (the dark current or OS bias). This does not explain, however, why vertebrate pho- toreceptors are forced to hyperpolarize in re- sponse to light stimuli. Compared to invertebrate microvilli-based motors, whose response is, in most cases a depolarization, this behavior appears to be a rather unorthodox way of responding to light. We believe that hyperpolarization is a means for the cell to avoid massive MTM reversal and produce damage in the disk renewal process.
In the present paradigm, hyperpolarization of the OS is passively transmitted by the plasma membrane to the IS according to the cable prop- erties of cell membranes (Hagins, 1970; Tomita, 1971, Fuertes and O’Bryan, 1972). Yet, as we have seen, the OS is separated from the IS by the ciliary connective which displays discrete mem- brane domains. Those domains, in our view, must play a significant role in separating the signal pathway into two distinct domains: the OS with the light induced membrane hyperpolarization on the one side and the IS with MTM-dependent membrane hyperpolarization on the other. Hyper- polarization of the IS, although occurring simul-
To sum up, we believe that further experimental work is needed in order to fully elucidate the role of the cilium in vertebrate photoreceptors. As McNaughton (1990) states, ‘it would perhaps be unwise to conclude that there are no other surprises awaiting us’.
4. Conclusion
Far from having the pretention of giving an exhaustive account of MTM electrodynamics, the scope of this paper was limited to expose the reader to the heuristic capabilities of the charge transfer model. In this first tentative approach, we also hope to have brought sufficient theoretical evidence for the existence of specific basic .require- ments for the proper functioning of MTM and, thus, open the way to experimental investigations.

220 E.M. Isinna et al. / BioSystems 39 (1996) 187-226
We wish that this model will give access to new, still unexplored pathways in cell biology. Cur- rently, the biology of the cytoskeleton is, in fact, almost exclusively based on a biochemical view of cell processes. Therefore, we are profoundly con- vinced that consideration of the existence of elec- trodynamical phenomena in cytoskeletal components may help to elucidate some of the many still unsolved problems.
The examples of possible applications of our model are legion. From the synchronization of proteins responsible for cell motility to the func- tioning of sensory neurons, from mitosis to brain dynamics, the existence of charge transfer in the cytoskeleton may challenge the currently rather restrictive view of protein-protein interactions. However, besides more or less local protein- protein interactions, our model might also shed a new light on global cell dynamics and information processing inside the cell.
For example, it should be possible to envisage experimental investigations concerning the infor- mation transfer processes occurring between the cytoskeleton and the cell nucleus. It is a well known fact that, in eukaryotic cells, a potential difference exists between the interior of the cell nucleus and the cytoplasm. From the viewpoint of our model, this simple fact acquires a totally new dimension and can be interpreted as an additional evidence for the universality of MTM require- ments we describe in this paper.
Another domain of applications concerns cell sensitivity to magnetic and electromagnetic fields. Presently, no plausible hypothesis exists which could account for such sensitivity. Electromag- netic phenomena and interactions are thus consid- ered as mere curiosities and have largely been ignored within present science. The hypothesis of charge transfer in the cytoskeleton represents a possible solution for some of those inexplicable phenomena based on long range interactions. Such interactions might be, in fact, the result of high frequency coupling through GHz oscillations generated by the charge transfer phenomenon in- side the cytoskeleton.
Still another field is mechanosensory perception involving both MT - as well as microvilli -
based sensory mechanisms. In fact, mechanical as well as chemical stimuli may influence electron transfer occurring inside MT and probably also inside microvilli (made of actin) and, subsequently the electrical response of the cell. Thus, the charge transfer hypothesis might also concur to explain some still unsolved problems in this domain.
To summarize, we hope that our suggestions will lead to further investigations in this new research field. Experimental work cannot effi- ciently progress without models and hypotheses which show new pathways for solving old ques- tions.
References
Allen, R.D., 1978, Particle arrays in the surface membrane of Paramecium: junctional and possible sensory sites. J. Ultra- struct. Res. 63, 64-78.
Allen, R.D. et al., 1985a, Gliding movement of and bidirec- tional transport along single native microtubules from squid axoplasm: evidence for active role of microtubules in cyto- plasmic transport. J. Cell Biol. 100, 1736-1752.
Allen, R.D. et al., 1985b, The dynamic microtubules theory of axonal and cytoplasmic transport, and a hypothesis regard- ing the mechanism. Microtubules and microtubules in- hibitors. M. De Brabander and J. De Mey (eds.) Elsevier, Amsterdam.
Amos, L.A. et al., 1976, Molecular structure of flagella micro- tubules, in: Cell Motility, R. Goldman, T. Pollard and J. Rosenbaum (eds.) Cold Spring Harbor Laboratory, pp. 847-868.
Amos, L.A., 1979, Structure of microtubules, in: Micro- tubules, K. Roberts and J.S. Hyams (eds.) Academic Press, London.
Amos, L.A., 1982, Tubulin and associated proteins, in: Elec- tron Microscopy of Proteins, Vol. 3, I.R. Harris (ed.) Aca- demic Press, London-N.Y., pp. 207-250.
Anderson, R.G.W. and Hein, C.E., 1977, Distribution of anionic sites on the oviduct ciliary membrane. J. Cell Biol. 72, 482-492.
Arikawa, K. and Williams, D.S., 1989, Organization of actin filaments and immunocolocalization of a-actinin in the con- necting cilium of rat photoreceptors. J. Comp. Neurol. 288, 640-646.
Athenstaedt, H., 1974, Pyroelectric and piezoelectric properties of vertebrates. Ann. Acad. Sci. 238, 68-94.
Bancroft, F.W., 1913, Heliotropism, differential sensibility and galvanotropism in Euglena. J. Exp. 2001. 15, 3833428.
Barber, V.C., 1974, Cilia in sense organs, in: Cilia and Flag- ella, M.A. Sleigh (ed.) Academic Press, London.
Bershadsky, A.D. and Vasiliev, J.M., 1988, Cytoskeleton Plenum Press, N.Y.

E.M. lsinna et ul. / BioSystems 39 (1996) 187-226 221
Besharse, J.C.. 1986, Photosensitive membrane turnover: dif- ferentiated membrane domains and cell-cell interaction, in: The Retina: a Model for Cell Biological Studies. Part 1. R. Adler and D. Farber (eds.) Academic Press, N.Y., pp. 297-352.
Besharse, J.C., and Pfenninger, K.H., 1980, Membrane assem- bly in retinal photoreceptors. I. Freeze fracture analysis of cytoplasmic vesicles in relationship to disc assembly. J. Cell Biol. 87, 451.-463.
Besharse, J.C. et al., 198.5, Membrane assembly in retinal photoreceptors. III. Distinct membrane domains of the con- necting cilium of developing rods. J. Neurosci. 5, lO35- 1048.
Besharse. J.C. and Horst, C., 1990, The photoreceptor con- necting cilium. In: Ciliary and Flagellar Membranes, R.A. Bloodgood (ed.) Plenum Press, N.Y.
Bonini, N. et al., 1991, The regulation of ciliary motility in Paramecium by Ca’+ and cyclic nucleotides. Adv. Second Messenger and Phosphoprotein Res. 23, 2277272.
Bouck, G.B. et al., 1990, Euglena gracilis: a model for flagellar surface assembly, with reference to other cells that bear flagellar mastigonemes scales. In: Ciliary and Flagellar Membranes, R.A. Bloodgood (ed.) Plenum Press, N.Y.
Bovee, E.C. and Jahn, T.L., 1972, A theory of piezoelectric activity and ion-movements in the relation of flagellar struc- tures and their movements to the phototaxis of Euglenu. J. Theor Biol. 35, 259-273.
Brady, ST., 1985, A novel brain ATPase with properties expected for the fast axonal transport motor. Nature 317, 13375.
Brady et al., 1990, A monoclonal antibody against kinesin inhibits both anterograde and retrograde fast axonal trans- port in squid axoplasm. Proc. Natl. Acad. Sci. USA 87, 1061- 1065.
Broekhuyse, R.M., et al., 1985, Light induced shift and bind- ing of S-antigen in retinal rods. Curr. Eye Res. 4, 6133618.
Brokaw, C.J., 1974, Movement of the flagellum of some marine invertebrate spermatozoa. In: Cilia and Flagella, M.A. Sleigh, (ed.) Academic Press.
Brokaw, C.J. et al., 1982, Analysis of the movement of Chlamidumonas flagella. 111. The function of the radial spoke system is revealed by comparison of wild-type and mutant flagella. J. Cell Biol. 92, 7222732.
Brokaw, C.J. and Luck, D.J.L., 1985, Bending patterns of Chlamidomonas flagella. III. A radial spoke head deficient mutant and a central pair deficient mutant. Cell Motil. 5, 1955208.
Brokaw, C.J. and Kamiya, R., 1987, Bending patterns of Chlamidomonas flagella: IV. Mutants with defects in inner and outer dynein arms indicate differences in dynein arm function. Cell Motil. 8, 68- 75.
Buetow, D.E., 1968a, The Biology of Euglena. Vol. I. Academic Press, N.Y.
Buetow, D.E., 1968b, The Biology of Euglena. Vol. II, Academic Press, N.Y.
Buetow, D.E., 1982, The Biology of Eug/ena. Vol. 111. Academic Press. N.Y.
Buetow, D.E., 1989, The Biology of Euglenu. Vol. IV, Aca- demic Press, N.Y.
Chailley, B. et al., 1990, Structure and assembly of the oviduct ciliary membrane, in: Ciliary and Flagellar Membranes, R.A. Bloodgood (ed.).
Chaitin, M.H., 1992, Double immunogold localization of opsin and actin in the cilium of developing mouse photore- ceptors. Exp. Eye Res. 54, 261-267.
Chou, K.-C. Zhang, C-T. and Maggiora, G.M., 1994, Solitary wave dynamics as a mechanism for explaining the internal motion during microtubule growth. Biopolymers, 34, 143.
Chretien, D. and Wade, R.H., 1991, New data on the micro- tubule surface lattice. Biol. Cell 71, 161-174.
Cleveland, D.W. 1982, Treadmilling of tubulin and actin. Ceil 28. 6899691.
Cohen, A.I., 1972, Rods and cones, in: Handbook of Sensory Physiology-Vol. VII/2-Physiology of Photoreceptor Organs. Fuortes, M.G.F. ed. Springer, Berlin.
Cope, F.W., 1974, Solid state physical mechanism of biologi- cal energy transduction, Ann. NY Acad. Sci., 227, 74-97.
Crescitelli. F., 1972, The visual cells and visual pigments of the vertebrate eye, in: Handbook of Sensory Physiology, Vol. VII/l - Photochemistry of Vision, H.J.A. Dartnall (ed.) Springer Verlag. Berlin.
David-Pfeuty, T. et al., 1977, GTPase activity of tubulin associated with microtubule assembly. Proc. Natl. Acad. Sci. USA 14, 5372-5376.
Davydov, A.S., 1982. The migration of energy and electrons in biological systems, in: Biology and Quantum Mechanics. A.V. Holden and B. Winlow (eds.) Springer Verlag. Berlin.
Dentler, W.L., 1977, Fine structural localization of phospatases in cilia and basal bodies of Tetrullymrrzu pyri-
fbrmis. Tissue Cell 9, 2099222. Dentler. W.L., 1981, Microtubule-membrane interactions in
cilia and flagella. Int. Rev. Cytol. 72. I-47. Dentler, W.L., 1986. Isolation of capped cilia from bovine
trachea and the effect of caps on microtubule assembly in vitro. J. Cell Biol. 103, 279a.
Dentler. W.L., 1987, Cilia and flagella. Int. Rev. Cytol. Suppl. 17, 391-456.
Dentler, W.L., 1990, Microtubule-membrane interactions, in: Ciliary and Flagellar Membranes, R.A. Bloodgood (ed.) N.Y., Plenum Press.
Dentler, W.L. and Rosenbaum, J.L., 1977, Flagellas elonga- tion and shortening in Chlamydomonus. 111. Structures at- tached to the tips of flagellar microtubules and their relationship to the directionality of microtubule assembly. J. Cell Biol. 74. 7477759.
Dentler, W.L. et al., 1980, Microtubule-membrane interactions in cilia. II. Photochemical crosslinking of bridge structures and the identification of a membrane-associated ATPase. J. Cell Biol. 84, 381-403.
Dentler. W.L. and Le Cluyse, E.L., 1982, The effects of structures attached to the tips of tracheal ciliary micro- tubules on the nucleation of microtubules assembly in vitro. Cell Motil. Suppl. I. 13.- 18.

222 E.M. Isinna et al. 1 BioSystems 39 (1996) 187-226
De Robertis, E. and Lasansky, A., 1961, Ultrastructure and chemical organization of photoreceptors, in: The Structure of the Eye. Academic Press, N.Y.
DeVault, D., 1984, Quantum-Mechanical Tunnelling in Bio- logical Systems. Cambridge Univ. Press.
DeVault, D. et al., 1967, Electron tunnelling in cytochromes. Nature, Lond. 215, 6422644.
Dirksen, E.R. and Satir, P., 1972, Ciliary activity in the mouse oviduct as studied by transmission and scanning electron microscopy. Tissue Cell 4, 389-404
Duhaiman, A.S and Bamburg, J.R., 1984, Isolation of brain r-actinin: its characterization and a comparison of its prop- erties with those of muscle E-actinins. Biochemistry 23, I600- 1608.
Dustin, P., 1984, Microtubules. Springer, Berlin-N.Y. Dute, R. and Kung, C., 1978, Ultrastructure of the proximal
region of somatic cilia in Paramecium tetraurelia, J. Cell Biol. 78, 451-464.
Eakin, R.M., 1968, Evolution of photoreceptors, in: Evolu- tionary Biology, Vol. II. T. Dobzhanski, M.K. Hecht, W.C. Steere (eds.), Appleton-Century-Crofts, N.Y.
Eakin, R.M., 1972, Structure of Invertebrate Photoreceptors, in: Handbook of Sensory Physiology, Vol. VII/l. H.J.A. Dartnall (ed.) Springer Verlag, Berlin.
Eckert, R., 1972, Bioelectric control of ciliary activity. Science, 176, 473-481.
Engelmann, T.W., 1882. Ueber Licht-und Farbenperception niederster Organismen. Plliigers Arch. 29, 387-400.
Feileib, M.E. and Curry, GM., 1971, The nature of the photoreceptor in phototaxis, in: Handbook of Sensory Physiology, Vol. 1. Springer Verlag, Berlin.
Feileib, M.E., 1985, Behavioural studies of free-swimming photoresponsive organisms, in: Sensory Perception and Transduction in Aneural Organisms, Cl. Colombetti et al. (eds.) Plenum Press, N.Y.
Fesenko, E.E. et al., 1985, Induction by cyclic GMP of cationic conductance in plasma membrane of retinal rod outer segment. Nature 313, 310-313.
Frohlich, H., 1968, Long-range coherence and energy storage in biological systems. Int. J. Quantum Chem. 2, 64-649.
Friihlich, H., 1970, Long range coherence and the action of enzymes. Nature, 228, 1093.
Frohlich, H., 1975a, The extraordinary dielectric properties of biological materials and the action of enzymes. Proc. Nat]. Acad. Sci. USA 72, 4211-4215.
Friihlich, H., 1975b, Evidence for Bose condensation-like exci- tations of coherent modes in biological systems. Phys. Let- ters, Vol. 51, A-l.
Fuertes, M.G.F. and O’Bryan, P.M., 1972, Generator poten- tial in invertebrate photoreceptors, in: Handbook of Sensory Physiology - Vol. VII/2 - Physiology of Photoreceptor Organs. M.G.F. Fuorte (ed.) Springer, Berlin. pp. 279-321.
Gibbons, I.R., 1981, Cilia and flagella of eukaryotes. J. Cell Biol. 91, 107s-124s.
Gibbons, I.R. et al., 1976, Multiple forms of dynein in sea urchin sperm flagella, in: Cell Motility, R. Goldman, T.
Pollard and J. Rosenbaum (eds.) Cold Spring Harbor Labo- ratory, pp. 847-868.
Gibbons, I.R., 1988, Dynein ATPases as microtubule motors. J. Biol. Chem., 263, 15837-15840.
Goldman, R. et al., 1976, Cell Motility, R. Goldman, T. Pollard and J. Rosenbaum (eds.) Cold Spring Harbor Labo- ratory.
Goldstein, SF., 1974, Isolated, reactivated and laser-irradiated cilia and flagella, in: Cilia and Flagella, M.A. Sleigh (ed.) Academic Press.
Gualtieri, P. et al., 1986, Isolation of the photoreceptor (paraflagellar body) of the phototactic flagellate Euglena gracilis. Arch. Microbial. 145, 303-305.
Gualtieri, P. et al., 1989, Absorption spectrum of a single isolated paraflagellar swelling of Euglena gracilis. Biochim. Biophys. Acta. 993, 293-296.
Gualtieri, P. et al., 1990, A look into the reservoir of Euglena gracilis: S.E.M. investigations of the flagellar apparatus. Micron Microsc. Acta, 21, 131-138
Gualtieri, P. et al., 1992, Identification of a rhodopsin pho- toreceptor in EugLena gracilis. Biochim. Biophys. Acta, 1117, 55-59.
Hagins, W.A. et al., 1970, Dark current and photocurrent in retina1 rods. Biophys. J. IO, 380-412.
Halldal, P., 1957, Importance of calcium and magnesium ions in phototaxis of motile green algae. Nature 179, 2155216.
Halldal, P., 1959, Factors affecting light response in phototac- tic algae. Physiol. Plant. 12, 742-752.
Hameroff, S. R., 1987, ‘Ultimate Computing: Biomolecular Consciousness and Nanotechnology’. Elsevier - North Holland, Amsterdam.
Hameroff, S.R., Watt, R.C., 1982, Information processing in microtubules. J. Theor. Biol. 98, 549-561.
Hameroff, S.R., Smith, S., Watt, R.C., 1984, Non-linear elec- trodynamics in cytoskeletal protein lattices, in: Nonlinear Electrodynamics in Biology and Medicine, F.A. Lawrence and W.R. Adey (ed.) Plenum Press. N.Y.
Hiramoto, Y., 1974, Mechanics of ciliary movement, in: Cilia and Flagella, M.A. Sleigh, (ed.), Academic Press.
Hirokawa, N. et al., 1991, Kinesin associates with antero- gradely transported membranous organelles in vivo. J. Cell Biol. 114, 295-302.
Hirokawa, N., 1993, Axonal transport and the cytoskeleton. Curr. Opin. Neurobiol. 3, 724-731.
Hirose, K. et al., 1995, Nucleotide-dependent angular change in kinesin motor domain bound to tubulin. Nature 376, 277-279.
Hitt, A. et al., 1990, Microtubule solutions display nematic liquid crystalline structure. J. BioI. Chem. 265, 1639-1647.
Hoenger, A. et al., 1995, Three-dimensional structure of a tubulin-motor-protein complex. Nature 376, 27 l-274.
Hoops, H.J. and Witman, G.B., 1985, Basal bodies and associ- ated structures are not required for normal flagellar motion or photaxis in the green alga Chorogonium elongatum. J. Cell Biol. 100, 297-309.
Holwill, M.E.J., 1974, Hydrodynamic aspects of ciliary and flagellar movement, in: Cilia and Flagella, M.A. Sleigh (ed.) Academic Press.

E.M. lsinna et al. / BioSystems 39 (1996) 187-226 223
Hopfield, J.J.. 1974, Electron transfer between biological molecules by thermally activated tunnelling. Proc. Nat. Acad. Sci, USA, Vol. 71, No. 9, 3640-3644.
Horio, T. and Hotani, H., 1986, Visualisation of the dynamic instability of individual microtubules by darkfield mi- croscopy. Nature (Lond.) 321. 6055607.
Huang. B.P.-H.. 1986, Chlamydomonas Reinhardtii: a model system for the genetic analysis of flagellar structure and motility. Int. Rev. Cytol. 99, I81 -215.
Huitorel, P. and Kirschner, M.W., 1988, The polarity and stability of microtubule capture by the kinetochore. J. Cell Biol. 106. 151-159.
Hyams, J.S., 1982, The Euglena paraflagellar rod: structure, relationship to the other flagellar components and prelimi- nary biochemical characterization. J. Cell Sci. 55, 199-210.
Insinna, E.M., 1989, Jungian Synchronicity and Biological Morphogenesis. An Application to the Study of Micro- tubules. Ph.D. Thesis, Universite Paris X, Nanterre. ISSN: 0294p 1767.
Ishijima, S. et al., 1988, Comparative study of the beat pat- terns of American and Asian horseshoe crab sperm: evi- dence for a role of the central pair complex in forming planar waveforms in flagella. Cell Motil. 9, 264-270.
Jarosch, R. and and Fuchs, B., 1975, Zur Fibrillenrotation in der Synura-Geissel, Protoplasma 85, 285-290.
Jennings. H.S., 1906, Behavior of the Lower Organisms. N.Y. Columbia University Press.
Jockush, B.M. and Isenberg. G., 1981, Interaction of I-actinin and vinculin with actin: opposite effects on filament network formation. Proc. Natl. Acad. Sci. USA 78, 3005-3009.
Kamiya, R.. 1982, Extension and rotation of the central-pair microtubules in detergent-treated Chlamydomonas flagella. Cell Motil. Suppl. 5, 1699173.
Kamiya, R. et al., 1982, Rotation of the central pair micro- tubules in Chlam~~domonas flagella, in: Biological Functions of Microtubules and Related Structures. H. Sakai, H. Mohri and G.G. Borisy (eds.) Academic Press, N.Y., pp. 189-198.
Kikkawa, M. et al.. 1994, Direct visualization of the micro- tubule lattice seam both in vitro and in vivo. J. Cell Biol. 127, 196551971.
Kimball, J.C., 1980, Kinks, solitons and nonlinear transport in solids. Phys. Rev. B21, 2104-2107.
Kirchner, K and Mandelkow, E., 1985. EMBO J. 4, 2397- 2402.
Kirschner, M.W. 1978, Microtubule assembly and nucleation. Int. Rev. Cytol. 54, I-71.
Kirschner, M.W. 1980, Implication of treadmilling for the stability and polarity of actin and tubulin polymers in vivo. J. Cell Biol. 86, 330-334.
Kitanishi-Yumura, T. and Fukui, Y., 1987, Re-organisation of microtubules during mitosis in Dyctyostelium: dissociation from MTOC and selective assembly/disassembly in situ. Cell Motil. 8, lO6- I 17.
Koruga, D., 1990, Semiconductor and crystal symmetry as- sessment of microtubule proteins as molecular machines. J. Mol. Electron,, Vol. 6, 1677173.
Kuhn, C. and Engleman, W., 1978, The structure of the tips of mammalian respiratory cilia. Cell Tissue Res. 186, 491-498.
Leedale, G.F., 1967, The Euglenoid Flagellates. Englewood Cliffs, N.J. Prentice Hall.
Linck, R.W. and Langevin, G.L., 1981, Reassembly of flagel- lar B (ab) tubulin into singlet microtubules: consequences for cytoplasmic microtubule structure and assembly. J. Cell Biol. 89, 323-337.
Linck, R.W. et al., 1981, Arrangement of tubulin subunits and microtubule-associated proteins in the central-pair micro- tubule apparatus of squid (Loligo pealei) sperm flagella. J. Cell Biol. 89, 3099322.
Luck, D.J.L., 1984, Genetic and biochemical dissection of the eukaryotic flagellum. J. Cell Biol. 98, 7899794.
Luduena, R.F.. 1979, Biochemistry of tubulin, in Micro- tubules, K. Roberts and J.S. Hyams (eds.) Academic Press, London.
Machemer, H., 1974, Ciliary activity and metachronism in Protozoa, in: Cilia and Flagella, M.A. Sleigh (ed.) Academic Press.
Machemer, H., 1986, Electromotor coupling in cilia, in: Mem- brane Control of Cellular Activity, Vol. 33. C. Luttgau (ed.) Fortschritte der Zoologie, Fischer Verlag, Stuttgart, pp. 205-250.
Machemer, H. and Ogura, A., 1979, Ionic conductances of membranes in ciliary and deciliated Paramecium. J. Physiol. 296, 49960.
Mandelkow, E.-M and Mandelkow, E., 1992, Microtubule oscillations. Cell Motil. Cytoskel. 22, 235244.
Mandelkow, E. et al., 1985, Domains, antigenicity and sub- structure of tubulin, in: Microtubules and Microtubules Inhibitors. M. De Brabander and J. De Mey (eds.) Elsevier, Amsterdam.
Mandelkow et al., 1986, On the surface lattice of microtubules: helix starts, protofilament number, seam and handedness. J. Cell Biol. 102, 1067-1073.
Mandelkow, E. et al., 1989, Spatial patterns from oscillating microtubules. Science, 246, l29l- 1293.
Marchese-Ragona, S.P. and Johnson, 1985. Localization of 14s and 22s dyneis in Tetrahymena cilia. J. Cell Biol. 101, 278a.
Marcus, R.A. and Sutin, N., 1985, Electron transfers in chem- istry and biology. Biochim. Biophys. Acta, 81 I, 265-322.
Margolis, R.L. and Wilson, L., 1981, Microtubule treadmills - possible molecular machinery. Nature 293, 7055711.
Mast, S.O., I91 1, Light and the Behavior of Organisms, N.Y. John Wiley and Sons.
Mast, S.O., 1914, Orientation in Euglena with some remarks on tropism. Biol. Zbl. 34, 64-664.
Mast, SO., 1917, The relation between spectral color and stimulation in the lower organisms. J. Exp. Zool. 22, 47ll 528.
McNaughton, P.A., 1990, Light response of vertebrate pho- toreceptors. Physiol. Rev., 70, 8477883.
Miller, J.R., 1975, Reactions of trapped electrons by quantum mechanical tunnelling observed by pulse radiolysEs of an aqueous glass. J. Phys. Chem. 79, 1070-1078.

224 E.M. Isinna et al. 1 BioSystems 39 (1996) 187-226
Mitchison, T. and Kirschner, M., 1984, Dynamic instability of microtubules growth. Nature 312, 237-242.
Mitchison, T. et al., 1986, Sites of microtubule assembly and disassembly in the mitotic spindle. Cell, 45, 115-527.
Mogesen, M.M. and Tucker, J.B., 1987, Evidence for micro- tubule nucleation at plasma membrane-associated sites in Drosophila. J. Cell Sci., 88, 95-107.
Mogesen, M.M. et al., 1989, Microtubule polarities indicate that nucleation and capture of microtubules occurs at cell surfaces in Drosophila. J. Cell Biol. 108, 1445-1452.
Moestrup, O., 1982, Flagellar structure in algae: a review, with new observations particularly on the Crysophyceae, Phaenophyceae (Fucophyceae), Euglenophyceae and Reck- ertia. Phycologica, 21, 427-528.
Murray, J.M., 1984, Three-dimensional structure of a mem- brane-microtubule complex. J. Cell Biol. 98, 283-295.
Musgrave, A., et al., 1986, Evidence for a functional mem- brane barrier in the transition zone between the flagellum and the cell body of Chlamydomonas eugametos gametes. Planta 167, 544553.
Naitoh, Y. and Eckert, R., 1969, Ionic mechanisms controlling behavioral responses of Paramecium to mechanical stimula- tion. Science 164, 963-965.
Naitoh, Y. and Eckert, R., 1974, The control of ciliary activity in Protozoa, in: Cilia and Flagella, M.A. Sleigh (ed.) Aca- demic Press.
Nangaku, M., 1994, KIFIB, a novel microtubule plus end-di- rected monomeric motor protein for transport of mitochon- dria. Cell 79, 1209-1220.
Niclas, J. et al., 1994, Cloning and localization of a conven- tional kinesin motor expressed exclusively in neurons. Neu- ron 21, 1059-1072.
Nojima, D. et al., 1995, At least one of the protofilamnets in flagellar microtubules is not composed of tubulin. Curr. Biol. 5, 158-167.
Norwood, J.T. et al., 1978, Polycationic macromolecules in- hibit cilia-mediated ovum transport in the rabbit oviduct. Proc. Natl. Acad. Sci. USA 75, 44134416.
Norwood, J.T. and Anderson, R.G.W., 1980, Evidence that adhesive sites on the tips of oviduct cilia membranes are required for ovum pickup in situ. Biol. Reprod. 23, 788- 791.
Ogura, A. and Machemer, H., 1980, Distribution of mechanoreceptor channels in the Paramecium surface mem- brane. J. Comp. Physiol. 135, 233-242.
O’Hara, P.T., 1970, Spiral tilt of triplet fibers in human leukocyte centrioles. J. Ultrastruct. Res. 31, 195-198.
Okada, Y. et al., 1995, The neuron-specific kinesin superfamily protein KIFIA is a unique monomeric motor for antero- grade axonal transport of synaptic vesicle precursors. Cell 81, 769-780.
Olmsted, J. B., 1986, Microtubule-associated proteins. Ann. Rev. Cell. Biol. 2, 421-457.
Omoto, C.K. and Kung, C., 1979, The pair of central tubules rotates during ciliary beat in Paramecium. Nature 279, 532-534.
Omoto, C.K. and Kung, C., 1980, Rotation and twist of the central-pair microtubules in the cilia of Paramecium. J. Cell Biol. 87, 33-46.
Osafune, T. and Schiff, J.A., 1980, Stigma and flagellar swelling in relation to light and carotenoids in Euglena gracilis var. Bacillaris. J. Ultrastruct. Res. 73, 336-349.
Papermaster, D.S. et al., 1975, Membrane biosynthesis in the retina: opsin transport in the photoreceptor cell. Biochem- istry 14, 134331352.
Penn, R.D. and Hagins, W.A., 1972, Kinetics of the photocur- rent of retinal rods. Biophys. J. 12, 1073-1094..
Penningroth, S. M. and Petersons, D.D., 1986, Evidence for functional differences between two flagellar dynein ATPases. Cell Motil. 6, 586-594.
Perry, R.J. and McNaughton, P.A., 1991, Response properties of cones from the retina of the Tiger salamander. J. Physiol. 433, 561-587.
Perry, R.J. and McNaughton, P.A., 1993, The mechanism of ion transport by the Na + -Ca2+, K + exchange in rods isolated from the Salamander retina. J. Physiol. 466, 443- 480.
Pethig, R., 1979,, Dielectric and Electronic Properties of Bio- logical Materials. Wiley, Chichester.
Pethig, R. and Szent-Gyorgyi, A., 1977, Electronic properties of the casein methylglyoxal complex. Proc. Nat. Acad. Sci, USA, 74, 226-228.
Piccinni, E. and Omodeo, P., 1975, Photoreceptors and photo- tactic programs in Protista. Boll. Zool. 42, 57-79.
Piccinni, E. et al., 1975, ATPase activity in flagella from Euglena gracilis. Localization of the enzyme and effects of detergents. J. Protozool. 22, 331-335.
Picinni, E. and Mammi, M., 1978, Motor apparatus of Eu- glena gracilis: ultrastructure of the basal portion of the flagellum and the para-flagellar body. Boll. Zool. 45, 405- 414.
Pirner, M.A. and Linck, R.W., 1994, Tektins are het- erodimeric polymers in flagellar microtubules with axial periodicities matching the tubulin lattice. J. Biol. Chem. 269, 31800-31806.
Portman, R.W. et al., 1987, Development of microtubule capping structures in ciliated epithelial cells. J. Cell Sci. 87 (Pt. I), 85-94.
Preston, R.R. and Saimi, Y., 1990, Calcium ions and the regulation of motility in Paramecium, in: Ciliary and Flagel- lar Membranes, R.A. Bloodgood (ed.).
Rasmussen, S. et al., 1990, Computational connectionism within neurons: a model of cytoskeletal automata subserving neural networks. Physica D42, 428-449.
Roberts, K. and Hyams, J.S., 1979, Microtubules, Academic Press, London - N.Y.
Rosati, G. et al., 1991, Ultrastructure of the apical zone of Euglena gracilis: photoreceptors and motor apparatus. Elec- tron Microsc. Rev. 4, 319-342.
Roth, L.E. et al., 1970, Microtubules in the Heliozoan Ax- opodium. I. The gradion hypothesis of allosterism in struc- tural proteins. J. Ultrastr. Res., 30, 7-37.

E.M. Isinna et al. / BioSystems 39 (1996) 187-226 225
Roth, L.E. and Pihlaja, D.J., 1977, Gradionation: hypothesis Tomita, T., 1971, Electrical activity of vertebrate photorecep- for positioning and patterning. J. Protozool., 24, 2-9. tors. Q, Rev. Biophys. 3, 179-222.
Sale, W.S. and Satir, P., 1977, Direction of active sliding of Tucker, J.B., 1979, Spatial organization of microtubules, in: microtubules in Tetrahymena cilia, Proc. Natl. Acad. Sci. Microtubules, Roberts, K. and Hyams. J. S. (eds.) Aca- USA 74, 204552049. demic Press, London.
Sammak, P.J. et al., 1987, Mechanism of microtubules dynam- ics ‘in vivo’. J. Cell Biol. 104, 4022405.
Sandoz. D., et al. 1979, Relationship between intramembrane particles and glycoconjugates in the ciliary membrane of the quail oviduct. Biol. Cell. 39. 267-279.
Sataric, M.V. et al., 1993, Kinklike excitations as an energy transfer mechanism in microtubules. Phys. Rev. E 48, 589.
Satir, P., 1968. Studies on cilia 111. Further studies on the cilium tip and a ‘sliding filament’ model of ciliary motility. J. Cell Biol. 39, 77794.
Tucker, J.B., 1986, Centrosomal microtubule-organizing cen- tres and a switch in the control of protofilament [number for cell surface-associated microtubules during Drosnphila wing morphogenesis. Eur. J. Cell Biol. 41. 2799289.
Tuszynski, J. A. et al., 1995, Ferroelectric behavior in micro- tubule dipole lattices: implications for information process- ing, signaling and assembly/disassembly. J. Theor. Biol. (1995) 174, 371-380.
Scheele, R.B. and Borisy. G.G., 1979, In vitro assembly of microtubules, in: Microtubules, K. Roberts and J.S. Hyams (eds.) Academic Press, London.
Schmit, A-C. and Lambert. A-M., 1985, F-Actin is associated to microtubule function during cytokinesis in higher plants. A revised concept, in: Microtubules and Microtubule In- hibitors, M. De Brabander and J. De Mey (eds.) Elsevier, Amsterdam. pp. 243-246.
Vale, R. D..l987, tntracellular transport using microtubules- based motors. Ann. Rev. Cell. Biol. 3, 3477378.
Vale, R.D. and Toyoshima, Y.Y., 1988, Rotation and translo- cation of microtubules in vitro induced by dyneins from Tetrahymena cilia. Cell 52, 4599469.
Vale, R.D.. 1993, Guidebook to the Cytoskeletal and Motor Proteins, Thomas, K. and Vale R. eds., 1755212. Oxford Univ. Press.
Schnapf, J.L. and Baylor, D.A., 1987, How photoreceptor cells respond to light. Sci. Am. 256, 40-47.
Selden, S.C. and Pollard, T.D., 1986, Interaction of actin filaments with microtubules is mediated by microtubule-as- sociated-proteins and regulated by phosphorylation. Ann. N.Y. Acad. Sci. 466, 803. 812.
Sleigh, M.A., 1974, Metachronism of cilia of Metazoa, in: Cilia and Flagella, M.A. Sleigh (ed.) Academic Press.
Sneddon, S.F. et al., 1988, A new classification of the amino acid side chains based on doublet acceptor energy levels. Biophys. J., 53, I.
Vaughan, D.K. et al., 1989, Evidence that microtubules do not mediate opsin vesicle transport in photoreceptors. J. Cell Biol. 109. 3053-3062.
Spiegelman. B.L. et al., 1977, Turnover of tubulin and the jV-site GTP in CHO cells. Cell 12, 5877600.
Stebbings, H. and Hunt. C., 1982, The nature of the clear zone around microtubules. Cell Tiss. Res. 227, 609.
Steinberg, R.H., 1980, Disc morphogenesis in vertebrate pho- toreceptors. J. Comp. Neurol. 190, 501-518.
Stossel, T.P. et al., 1985, Non-muscle actin-binding proteins. Ann. Rev. Cell Biol. I, 353-402.
Stryer. L., 1986, Cyclic GMP cascade of vision. Annu. Rev. Neurosci. 9. 87- 119.
Vickerman, K. and Tetley, L., 1990, Flagellar surfaces of parasitic Proro:oa and their role in attachment, in: Ciliary and Flagellar Membranes, R.A. Bloodgood (ed.).
Warner, F.D.. 1974, The fine structure of the ciliary and flagellar axoneme, in: Cilia and Flagella, M.A. Sleigh (ed.) Academic Press, London. pp. 11~ 37.
Warner, F.D. and Satir, P., 1974, The structural basis of ciliary bend formation. radial spoke positional changes ac- companying microtubule sliding. J. Cell Biol. 63, 35.
Warner, F.D., 1976, Cross-bridge mechanism in ciliary motil- ity: the sliding-bending conversion, in: Cell Motility, R. Goldman, T. Pollard and J. Rosenbaum eds. Cold Spring Harbor Laboratory, pp. 891-914.
Warr. J.R. et al., 1966, Genetic control of flagellar structure in Chlamydomonas Reinhardiii. Genet. Res. 7. 3355351.
Watanabe, T., 1990, The role of ciliary surfaces in mating in Paramecium, in: Ciliary and Flagellar Membranes. R.A. Bloodgood (ed.).
Surek, B. and Melkonian, M., 1986, A cryptic cytostome is present in Euglenrr. Protoplasma 133, 39-59.
Szent-Gyorgyi, A.. 1941. Towards a new biochemistry. Science 93, 6099611.
Weisenberg, R.C., 1981, The role of nucleotide triphosphate in actin and tubulin assembly and function. Cell Motil. I, 485-498.
Wheatley, D.N., 1982, The Centriole: A Central Enigma of Cell Biology. Elsevier, N.Y.
Tabony. J. and Job, D., 1990, Spatial structures in microtubu- lar solutions requiring a sustained energy source. Nature, Vol. 346, 448-451.
Tabony, J. and Job, D., 1992. Microtubular dissipative struc- tures in biological auto-organization and pattern formation. Nanobiology, Vol. I, Nr 2, 131, 147.
Taylor, G., 1952, The action of waving cylindrical tails in propelling microscopic organisms. Proc. Sot. A. 21 I, 225- 239.
Willey, R.L. and Wibel, R.G., 1987, Flagellar roots and the reservoir cytoskeleton of Colacium libellae (Eu- glenophyceae). J. Phycol. 23, 283-288.
Williams, D.S. et al., 1988, Disruption of microfilaments orga- nization and deregulation of disk membrane morphogenesis by cytokalasin D in rod and cone photoreceptors. J. Comp. Neurol. 272, 161-172.
Witman, G.B. et al., 1976, Chlamydomonas mutants: evidence for the roles of specific axonemal components in flagellar movement, in: Cell Motility, R. Goldman, T. Pollard and J.

226 E.M. Isinna et al. / BioSystems 39 (1996) 187-226
Rosenbaum (eds.) Cold Spring Harbor Laboratory, pp. 9699986.
Witman, G.B. et al., 1978, Chlamydomonas flagellar mutants lacking radial spokes and central tubules. structure, compo- sition and function of specific axonemal components. J. Cell Biol. 76, 729-747.
Witman, G.B., 1990, Introduction to cilia and flagella, in: Ciliary and Flagellar Membranes, R.A. Bloodgood (ed.).
Wolken, J.J., 1977, Euglena: the phoreceptor system for pho- toxasis. J. Protozool. 24, 518-522.
Young, R.W., 1968, Passage of newly formed protein through the connecting cilium of retinal rods in the frog. J. Ultra- struct. Res. 23, 462-473.
Young, R.W., 1970, Visual cells. Sci. Am. 223, 80-91 Young, R.W. and Droz, B., 1968, The renewal of protein in
retinal rods and cones. J. Cell Biol. 39, 169-184.