effects of prenatal methylmercury exposure on brain monoamine oxidase activity and neurobehaviour of...
TRANSCRIPT

gy 28 (2006) 251–259www.elsevier.com/locate/neutera
Neurotoxicology and Teratolo
Effects of prenatal methylmercury exposure on brain monoamineoxidase activity and neurobehaviour of rats
Peter Beyrouty a,b, Christopher J. Stamler a,c,1, Jiun-Ni Liu a,c, Kovana Marcel Loua a,Stan Kubow c, Hing Man Chan a,b,c,⁎
a Centre for Indigenous Peoples’ Nutrition and Environment, Macdonald Campus of McGill University, Ste-Anne-de-Bellevue, Quebec, Canada, H9X-3V9b Natural Resource Sciences, Macdonald Campus of McGill University, Ste-Anne-de-Bellevue, Quebec, Canada, H9X-3V9
c The School of Dietetics and Human Nutrition, Macdonald Campus of McGill University, Ste-Anne-de-Bellevue, Quebec, Canada, H9X-3V9
Received 17 August 2005; received in revised form 14 November 2005; accepted 14 December 2005Available online 20 February 2006
Abstract
Monoamine oxidase (MAO) regulates levels of dopamine, serotonin, and noradrenaline in the nervous tissue and is required for properneuronal development. The purpose of this study was to determine if oral exposure of adult female rats to methylmercury (MeHg) at 0.5 or 1 mg/kg/day before and during pregnancy would affect MAO activity in various brain regions of the offspring. Offspring neurobehaviour performancewas also assessed. The brain MAO activity of female offspring was reduced at both MeHg doses with significantly lower values noted in thebrainstem region. No significant MeHg dose effects on MAO activity were observed in the male offspring. Neurobehavioural evaluationsindicated that MeHg exposure altered auditory startle in the female offspring. Rat whole embryos (gestational day 13.5) cultured with 750 μg/LMeHg in vitro significantly decreased total MAO activity by 15%. In conclusion, this study demonstrated that exposure to MeHg in rats beforeand/or during gestation resulted in a reduction of MAO activity in the developing embryo and brainstem of the female offspring withaccompanying changes in auditory startle response. Evaluation of MAO activity may serve as an indicator for neurotoxicity followingdevelopmental exposure to MeHg and should be further investigated.© 2006 Elsevier Inc. All rights reserved.
Keywords: Neurotoxicity; Auditory startle response; Brain development; Rat embryonic culture; Monoaminergic system
1. Introduction
Mercury (Hg) is a persistent pollutant that is widespread inthe environment. The dietary intake of the organic form of Hg,methylmercury (MeHg), in contaminated fish, seafood andaquatic mammals is the main route of human exposure [1]. Highdose accidental poisonings in Japan (Minamata) and Iraq haveindicated that the developing brain is particularly sensitive to theneurotoxic effects of MeHg [2,19]. Human offspring exposed tohigh concentrations of MeHg during gestation have exhibitedpostnatal symptoms similar to congenital cerebral palsy
⁎ Corresponding author. Community Health Program, University of NorthernBritish Columbia, 3333 University Way, Prince George, BC, Canada, V2N-4Z9.Tel.: +1 250 960 5237; fax: +1 250 960 5744.
E-mail address: [email protected] (H.M. Chan).1 Current address: Community Health Program, University of Northern
British Columbia, 3333 University Way, Prince George, BC, Canada, V2N-4Z9.
0892-0362/$ - see front matter © 2006 Elsevier Inc. All rights reserved.doi:10.1016/j.ntt.2005.12.007
including brain pathology involving severely damaged neuronsthroughout the cortex [41]. More recently, epidemiologicalstudies focusing on communities with a high fish consumptionhave suggested that subtle alterations in child motor andcognitive behaviour are related to low-level prenatal MeHgexposure [18].
In order to better evaluate the health risks and impact of low-level MeHg exposure, it is important to understand the potentialmechanisms of its toxicity. While some neurotoxic targets andmechanisms of high dose exposure are known [6,7], there existsa need to develop specific and functional methodologies toidentify subtle neurotoxic effects associated with low-level ex-posure to MeHg.
Monoamine neurotransmitters, including serotonin, dopa-mine, and noradrenaline, are involved in neurofunctionalbehaviours such as fight-or-flight response, emotion, arousal,motor activity, and cognition [4,11,20,23,43,44]. Disruption ofthese neuronal signaling pathways has been proposed as a

Table 1Summary of behavioural tests performed on offspring
Behavioural test Parameters Day oftesting(PND)
Functionalobservationalbattery (FOB)
Arena: rearing, gait abnormalities, overallgait score, bizarre behaviour, palpebralclosure, tremors, convulsions, piloerection,respiration, locomotor activity, arousal,grooming, defecation, urination, olfactoryresponse. Handling: lacrimation, pupilsize, salivation, urinary staining, diarrhea,body tone, extensor thrust, corneal reflex,pinna reflex, toe pinch, tail pinch, visualplacing, positional passivity, simple startle,air righting reflex. On surface: fore/hind-limb grip strength, hind-limb splay, bodytemperature, rotarod coordination, hotplate response and tail flick response.
21
Motor activity Total activity counts 17 and 21Auditory startleresponse
Average and maximum startle amplitude;time to reach maximum startle amplitude.
22
Swimmingperformance
Swimming ability in straight channel; time tocomplete multiple-T maze.
35 to 38
252 P. Beyrouty et al. / Neurotoxicology and Teratology 28 (2006) 251–259
mechanism for MeHg neurotoxicity using developing andadult rodent models [14,25,26]. Administration of Hgcompounds is known to increase noradrenaline and serotoninconcentrations in various brain regions of rats [25,26]. Inaddition, Hg exposure has been shown to increase rat striataldopamine release [13]. Brain monoamine neurotransmitters aredeaminated by a flavin-enzyme, monoamine oxidase (EC1.4.3.4, MAO) located in the outer mitochondrial membraneof astrocytes and neuronal cells [38]. Two isoenzymes havebeen identified, MAO-A and MAO-B, which differ in thespecificities for inhibitors, substrates and expression in tissue[38]. Pharmacological and genetic studies have shown thatMAO is important for serotonergic neuronal development andneurobehaviour [5,43]. Initial structure/function studies byGomes et al. [17] suggested that mercuric compounds interactwith functionally critical cysteine residues in MAO, resulting innon-competitive inhibition and permanent damage to theenzyme. In a more recent study, repeated exposure to MeHgresulted in MAO activity reduction in various brain regions ofadult Sprague–Dawley rats [8]. This literature suggests thatMAO may be a useful neurochemical marker for MeHg effectson the monoaminergic nervous system.
The objective of this study was to evaluate the use of MAOas a marker of developmental MeHg neurotoxicity. This wasperformed by determining the effects of low-level prenatalMeHg exposure on regional MAO brain activity levels in ratoffspring. A range of behavioural tests were also used to assessthe neurotoxic effects of the exposure. Additionally, the directeffects of MeHg on MAO levels in rat embryos cultured in vitrowere also evaluated.
2. Methods
2.1. Chemicals
Bovine serum albumin, Dulbecco's Modified Eagle'sMedium, horseradish peroxidase, kynauramine dihydrobromideresorufin, tyramine, 4-hydroxylquinoline and all other reagentswere purchased from Sigma-Aldrich (St. Louis, MO) and wereof analytical grade or higher. 10-Acetyl-3,7-dihydroxyphenox-azine (Amplex® Red) was purchased from Molecular Probes,Inc. (Eugene, OR). MeHg chloride (purityN95%; adjusted forchloride salt) was obtained from Alfa Aesar (Ward Hill, MA).
2.2. Animals
The protocol was approved by the Institutional Animal Careand Use Committee (IACUC) and McGill University, and thecare and use of animals were conducted in accordance withguidelines of the USA National Research Council and theCanadian Council on Animal Care. Adult female Sprague–Dawley rats (Charles River Canada, St. Constant, QC) wereapproximately 7.5 weeks of age at the start of treatment.Females used for embryo culture experiments were approxi-mately 10 weeks of age. All animals had ad libitum accessto a commercial laboratory diet (PMI Rodent Chow 5002)and water. Environmental conditions in the animal room
were set at 22±3 °C, 50±20% relative humidity, and 12h light/12 h dark.
2.3. In vivo phase
Eight adult female rats were randomly assigned to a control(deionized water), 0.5 or 1 mg MeHg/kg (body weight)/daygroup using a computerized randomization procedure. MeHgchloride was dissolved using a sonicator in nitrogen purgeddeionized water to achieve a final dose volume of 5 mL/kg bodyweight. The adult females were dosed by gavage once daily for4 weeks, and were then placed for mating with untreated adultmales of the same strain and source. The presence of sperm in avaginal lavage smear confirmed mating and was designated asgestation day (GD) 0. Treatment continued until GD 20. Theadult females were observed daily for general condition, andbody weight and food consumption were measured at least onceweekly. The dams were allowed to deliver and the generalcondition and body weight of the offspring were evaluatedperiodically. Each litter was culled to 4 males and 4 females onpostnatal day (PND) 4 using a procedure provided by acomputer-based random number generator. Pups were identifiedby tattooing (injection of India ink) of the paws. Weaningoccurred on PND 21.
Two male and two female offspring were randomly selectedfrom each litter for use in a series of behavioural tests (seeTable 1). The functional observational battery (FOB) wassimilar to the previously described method by Moser et al.[29]. The FOB consisted of a series of arena, handling and on-surface observations by an experimenter who was unaware ofoffspring group assignment. Rats were placed in a clear plasticarena (60 cm2) for 2 min observations primarily for activity,arousal and unusual behaviour (Table 1). Afterwards, theanimals underwent several handling evaluations, which

253P. Beyrouty et al. / Neurotoxicology and Teratology 28 (2006) 251–259
included body tone and various reflex tests (Table 1). Musclestrength was evaluated by measurement of fore and hind-limbgrip-strength. The animal was allowed to grip a wire-meshattached to a push–pull strain gauge (Chatillon®) and thenpulled until grip was released. Maximum strain was recorded ingrams for both the fore- and hind-limbs. The average value fromtwo assessments was used. Hind-limb foot splay was determinedby inking the hind paws and measuring the landing foot spreadfollowing a 30 cm drop. Rectal temperature was collected usinga probe attached to a digital recorder (Digitemp®). The rotarodperformance was assessed by recording the length of time (up to1 min) the animal remained on a rod rotating at 8 rpm up to amaximum of 60 s. Reactivity responsiveness to the presentationof heat was evaluated using a 55 °C hot plate (latency time to liftor lick paw) and tail analgesic meter (latency time to flick tail).
Animals were transferred to a testing room where motoractivities were measured. Motor activity levels were measuredindividually in Fig. 8 enclosures (San Diego Instruments®) byrecording the number of photo beam breaks during a 1 h testingsession. In the testing room, background white noise was pro-vided at a sound level of approximately 70 dBA using a whitenoise generator (Lafayette Instruments®) and the illuminationwas approximately 700–1000 Lux.
Auditory startle habituation response was evaluated individ-ually in a sound-attenuated chamber (San Diego Instruments®).The animals were given a 4-min acclimation period in thechamber enclosures during which time they were exposed tobackground noise level of 65 dBA. Animals were then sub-jected to brief sound pulses of 120 dBA on 50 identical trials(8 s separation between trials) and startle movement was mea-sured for each trial. Measurement of maximal startle amplitudeacross the recording window of 100 ms and the time to reachmaximum startle amplitude was recorded. The average startleamplitude across the recording window was also measured.
Swimming ability was evaluated on four consecutive trials(same day) by measuring the time to complete a straight waterchannel. The mean time in seconds was calculated from the fourtrials. Swimming performance was also assessed using theCincinnati water maze test [42], which was comprised of aseries of T-intersections. The animal had to choose the appro-priate swimming direction on multiple occasions in order tocomplete the path and locate the exit platform. Each animal wastested three times (approximately 15 min apart) and the timerequired to complete the path was recorded (up to a maximumof 300 s). The same procedure was performed on the next threeconsecutive days. The overall mean time required to completethe water maze was used for analysis.
The maternal rats were euthanized approximately 3 weeksafter weaning of the pups, and the offspring were euthanized atapproximately 6 weeks of age. The method of euthanasia wascarbon dioxide asphyxiation followed by exsanguinations fromthe abdominal aorta and/or decapitation. The brain regions(cortex, striatum, cerebellum, brainstem and hippocampus)were promptly dissected on ice from one male and one femaleoffspring randomly selected using a computer generatednumber system. The whole brain was used for Hg measurementfrom one male and one female offspring from five randomly
selected litters from each dose group. Tissues were immediatelyfrozen and maintained at −80 °C until for up to 3 months priorto MAO or total-Hg analysis.
2.4. In vitro phase
Embryos were obtained from pregnant rats and cultured basedon a previously described method [3] with modifications. On GD13.5, female ratswere euthanized by carbon dioxide asphyxiation.The uterus was immediately excised and harvested into a Petridish containing sterile Hanks' balanced salt solution (HBSS). Theuteruswas cut along the antimesometrial edge and each conceptuswas explanted. The remnants of the decidua, trophoblast andReichart's membrane were removed from the yolk sac. Embryoswere extruded into the medium still attached to the opened yolksac and placenta through the vitelline and umbilical vessels. Twoembryos were transferred to each culture bottle containing 10 mLof 15% rat serum (heat-inactivated), and 85% Dulbecco'sModified Eagle's Medium with either 0, 100 or 750 μg/L ofMeHg chloride. Bottles were gassed with 95% O2 and 5% CO2
for 2 min, incubated at 37 °C, rotated at 20 cycles/min, and re-gassed at a 6-h gassing interval. Following 12 h incubation,embryos were transferred into cryovials. Samples were stored at−80 °C for a maximum of a week prior to MAO analysis.
2.5. MAO analysis
MAO activity was measured in the five brain regions ofoffspring as previously described by Krajl [24] with modifica-tions [8]. Briefly, samples were homogenized in 10 volumes of50 mM phosphate buffer (pH 7.8) and mixed thoroughly in thepresence of 2% (v/v) Triton X-100. The homogenate was addedto 2.5 mL of phosphate buffer and 0.3 mL of 0.9% NaClsolution, and pre-incubated for 10 min at 37 °C. A singleconcentration of kynauramine dihydrobromide (22 μM) wasadded to the reaction mixture and incubated for 30 min at 37 °C.Preliminary experiments indicated that the enzyme activityduring this incubation period was linear with respect to time.The reaction was stopped by the addition of 0.2 mL of 5 Mperchloric acid followed by centrifugation at 1500 ×g for 10 minin a tabletop centrifuge (Sorvall RT6000D, Mandel ScientificCo. Ltd., Guelph, ON). An aliquot of the supernatant wasdiluted with 1 M NaOH solution and the fluorescence of theproduct, 4-hydroxylquinoline, was measured (excitation/emission; 318/380 nm) with a RF-551 Shimadzu spec-trofluorometric detector (Kyoto, Japan). The concentration ofproduct was determined from a standard curve of 4-hydro-xyquinoline and expressed as nmol product formed per 30 minper mg of protein.
Whole embryos were sonicated in 1 mL of sodium–potas-sium buffer (50 mM NaH2PO4, 5 mM KCl, 120 mM NaCl,pH 7.4) for 30 s. MAO activity in homogenates was measuredbased on a microplate method described by Zhou and Panchuk-Voloshina [45] and modified by Stamler et al. [40]. Embryosamples (50 μg of protein) were diluted with Sodium–potassiumbuffer and added to a 96-well plate. Reaction was initiated withthe addition of reaction buffer (100 μM Amplex® Red, 2 U/mL
CC Str Hip BS Cer
MA
O A
ctiv
ity (
nmol
/30m
in/m
g)
0
20
40
60
80 Females
*
Brain Region
Control0.5 mg/kg/d1.0 mg/kg/d
CC Str Hip BS Cer
MA
O A
ctiv
ity (
nmol
/30m
in/m
g)
0
20
40
60
80 Males
*
Control0.5 mg/kg/d1.0 mg/kg/d
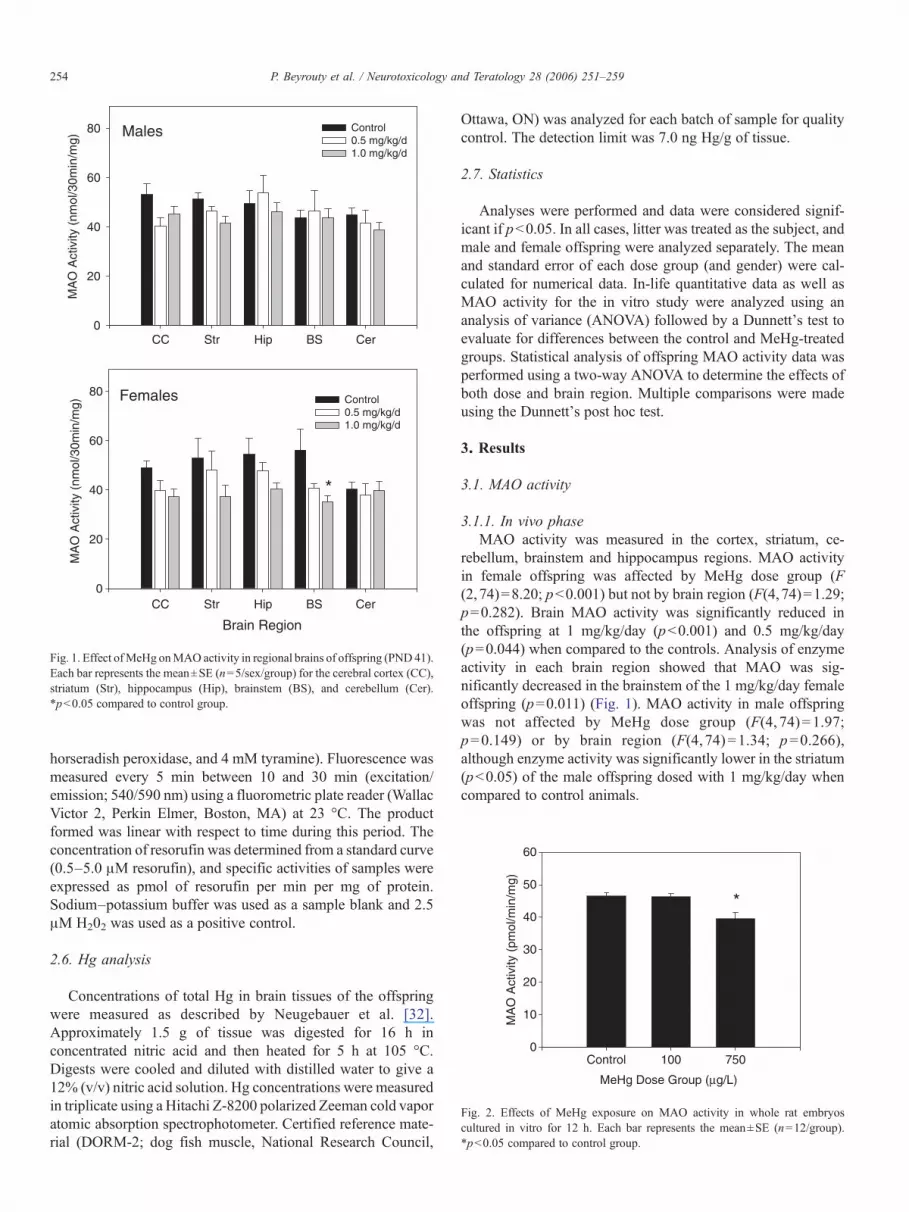
Fig. 1. Effect ofMeHg onMAO activity in regional brains of offspring (PND41).Each bar represents the mean±SE (n=5/sex/group) for the cerebral cortex (CC),striatum (Str), hippocampus (Hip), brainstem (BS), and cerebellum (Cer).*pb0.05 compared to control group.
MeHg Dose Group (µg/L)
Control 100 750
MA
O A
ctiv
ity (
pmol
/min
/mg)
0
10
20
30
40
50
60
*
Fig. 2. Effects of MeHg exposure on MAO activity in whole rat embryoscultured in vitro for 12 h. Each bar represents the mean±SE (n=12/group).*pb0.05 compared to control group.
254 P. Beyrouty et al. / Neurotoxicology and Teratology 28 (2006) 251–259
horseradish peroxidase, and 4 mM tyramine). Fluorescence wasmeasured every 5 min between 10 and 30 min (excitation/emission; 540/590 nm) using a fluorometric plate reader (WallacVictor 2, Perkin Elmer, Boston, MA) at 23 °C. The productformed was linear with respect to time during this period. Theconcentration of resorufin was determined from a standard curve(0.5–5.0 μM resorufin), and specific activities of samples wereexpressed as pmol of resorufin per min per mg of protein.Sodium–potassium buffer was used as a sample blank and 2.5μM H202 was used as a positive control.
2.6. Hg analysis
Concentrations of total Hg in brain tissues of the offspringwere measured as described by Neugebauer et al. [32].Approximately 1.5 g of tissue was digested for 16 h inconcentrated nitric acid and then heated for 5 h at 105 °C.Digests were cooled and diluted with distilled water to give a12% (v/v) nitric acid solution. Hg concentrations were measuredin triplicate using a Hitachi Z-8200 polarized Zeeman cold vaporatomic absorption spectrophotometer. Certified reference mate-rial (DORM-2; dog fish muscle, National Research Council,
Ottawa, ON) was analyzed for each batch of sample for qualitycontrol. The detection limit was 7.0 ng Hg/g of tissue.
2.7. Statistics
Analyses were performed and data were considered signif-icant if pb0.05. In all cases, litter was treated as the subject, andmale and female offspring were analyzed separately. The meanand standard error of each dose group (and gender) were cal-culated for numerical data. In-life quantitative data as well asMAO activity for the in vitro study were analyzed using ananalysis of variance (ANOVA) followed by a Dunnett's test toevaluate for differences between the control and MeHg-treatedgroups. Statistical analysis of offspring MAO activity data wasperformed using a two-way ANOVA to determine the effects ofboth dose and brain region. Multiple comparisons were madeusing the Dunnett's post hoc test.
3. Results
3.1. MAO activity
3.1.1. In vivo phaseMAO activity was measured in the cortex, striatum, ce-
rebellum, brainstem and hippocampus regions. MAO activityin female offspring was affected by MeHg dose group (F(2,74)=8.20; pb0.001) but not by brain region (F(4,74)=1.29;p=0.282). Brain MAO activity was significantly reduced inthe offspring at 1 mg/kg/day (pb0.001) and 0.5 mg/kg/day(p=0.044) when compared to the controls. Analysis of enzymeactivity in each brain region showed that MAO was sig-nificantly decreased in the brainstem of the 1 mg/kg/day femaleoffspring (p=0.011) (Fig. 1). MAO activity in male offspringwas not affected by MeHg dose group (F(4, 74)=1.97;p=0.149) or by brain region (F(4, 74)=1.34; p=0.266),although enzyme activity was significantly lower in the striatum(pb0.05) of the male offspring dosed with 1 mg/kg/day whencompared to control animals.
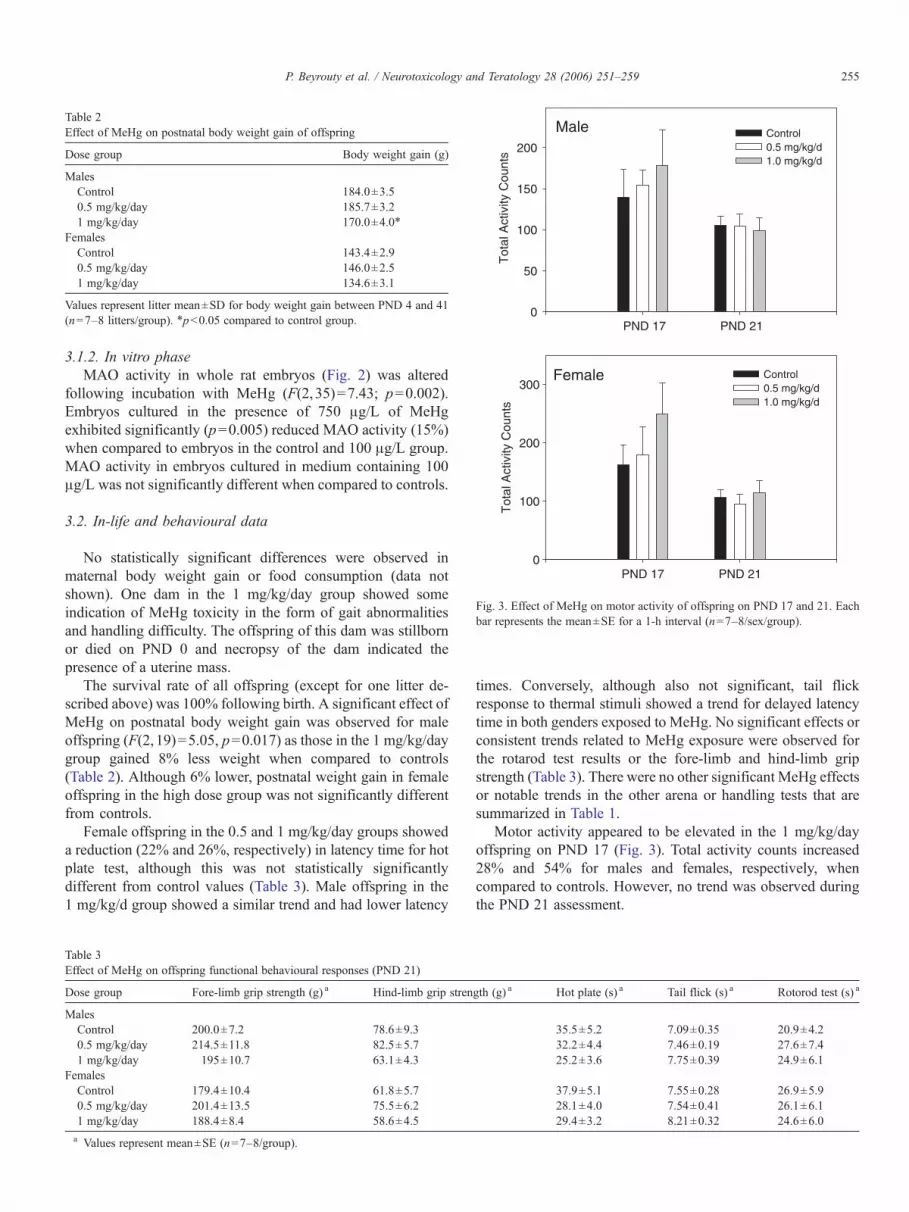
Table 2Effect of MeHg on postnatal body weight gain of offspring
Dose group Body weight gain (g)
MalesControl 184.0±3.50.5 mg/kg/day 185.7±3.21 mg/kg/day 170.0±4.0⁎
FemalesControl 143.4±2.90.5 mg/kg/day 146.0±2.51 mg/kg/day 134.6±3.1
Values represent litter mean±SD for body weight gain between PND 4 and 41(n=7–8 litters/group). ⁎pb0.05 compared to control group.
Male
PND 17 PND 21
Tot
al A
ctiv
ity C
ount
s
0
50
100
150
200Control 0.5 mg/kg/d1.0 mg/kg/d
PND 17 PND 21
Tot
al A
ctiv
ity C
ount
s
0
100
200
300Female Control
0.5 mg/kg/d1.0 mg/kg/d
Fig. 3. Effect of MeHg on motor activity of offspring on PND 17 and 21. Eachbar represents the mean±SE for a 1-h interval (n=7–8/sex/group).
255P. Beyrouty et al. / Neurotoxicology and Teratology 28 (2006) 251–259
3.1.2. In vitro phaseMAO activity in whole rat embryos (Fig. 2) was altered
following incubation with MeHg (F(2,35)=7.43; p=0.002).Embryos cultured in the presence of 750 μg/L of MeHgexhibited significantly (p=0.005) reduced MAO activity (15%)when compared to embryos in the control and 100 μg/L group.MAO activity in embryos cultured in medium containing 100μg/L was not significantly different when compared to controls.
3.2. In-life and behavioural data
No statistically significant differences were observed inmaternal body weight gain or food consumption (data notshown). One dam in the 1 mg/kg/day group showed someindication of MeHg toxicity in the form of gait abnormalitiesand handling difficulty. The offspring of this dam was stillbornor died on PND 0 and necropsy of the dam indicated thepresence of a uterine mass.
The survival rate of all offspring (except for one litter de-scribed above) was 100% following birth. A significant effect ofMeHg on postnatal body weight gain was observed for maleoffspring (F(2,19)=5.05, p=0.017) as those in the 1 mg/kg/daygroup gained 8% less weight when compared to controls(Table 2). Although 6% lower, postnatal weight gain in femaleoffspring in the high dose group was not significantly differentfrom controls.
Female offspring in the 0.5 and 1 mg/kg/day groups showeda reduction (22% and 26%, respectively) in latency time for hotplate test, although this was not statistically significantlydifferent from control values (Table 3). Male offspring in the1 mg/kg/d group showed a similar trend and had lower latency
Table 3Effect of MeHg on offspring functional behavioural responses (PND 21)
Dose group Fore-limb grip strength (g) a Hind-limb grip stren
MalesControl 200.0±7.2 78.6±9.30.5 mg/kg/day 214.5±11.8 82.5±5.71 mg/kg/day 195±10.7 63.1±4.3
FemalesControl 179.4±10.4 61.8±5.70.5 mg/kg/day 201.4±13.5 75.5±6.21 mg/kg/day 188.4±8.4 58.6±4.5a Values represent mean±SE (n=7–8/group).
times. Conversely, although also not significant, tail flickresponse to thermal stimuli showed a trend for delayed latencytime in both genders exposed to MeHg. No significant effects orconsistent trends related to MeHg exposure were observed forthe rotarod test results or the fore-limb and hind-limb gripstrength (Table 3). There were no other significant MeHg effectsor notable trends in the other arena or handling tests that aresummarized in Table 1.
Motor activity appeared to be elevated in the 1 mg/kg/dayoffspring on PND 17 (Fig. 3). Total activity counts increased28% and 54% for males and females, respectively, whencompared to controls. However, no trend was observed duringthe PND 21 assessment.
gth (g) a Hot plate (s) a Tail flick (s) a Rotorod test (s) a
35.5±5.2 7.09±0.35 20.9±4.232.2±4.4 7.46±0.19 27.6±7.425.2±3.6 7.75±0.39 24.9±6.1
37.9±5.1 7.55±0.28 26.9±5.928.1±4.0 7.54±0.41 26.1±6.129.4±3.2 8.21±0.32 24.6±6.0
Males Females
Ave
rage
Sta
rtle
Res
pons
e (m
V)
0
20
40
60
80
100
Males Females
Max
imum
Sta
rtle
Res
pons
e (m
V)
0
100
200
300
400
500
Tim
e to
Max
imum
Sta
rtle
(mill
i sec
onds
)
0
10
20
30Control0.5 mg/kg/d1.0 mg/kg/d
*
Males Females
Control0.5 mg/kg/d1.0 mg/kg/d
Control0.5 mg/kg/d1.0 mg/kg/d
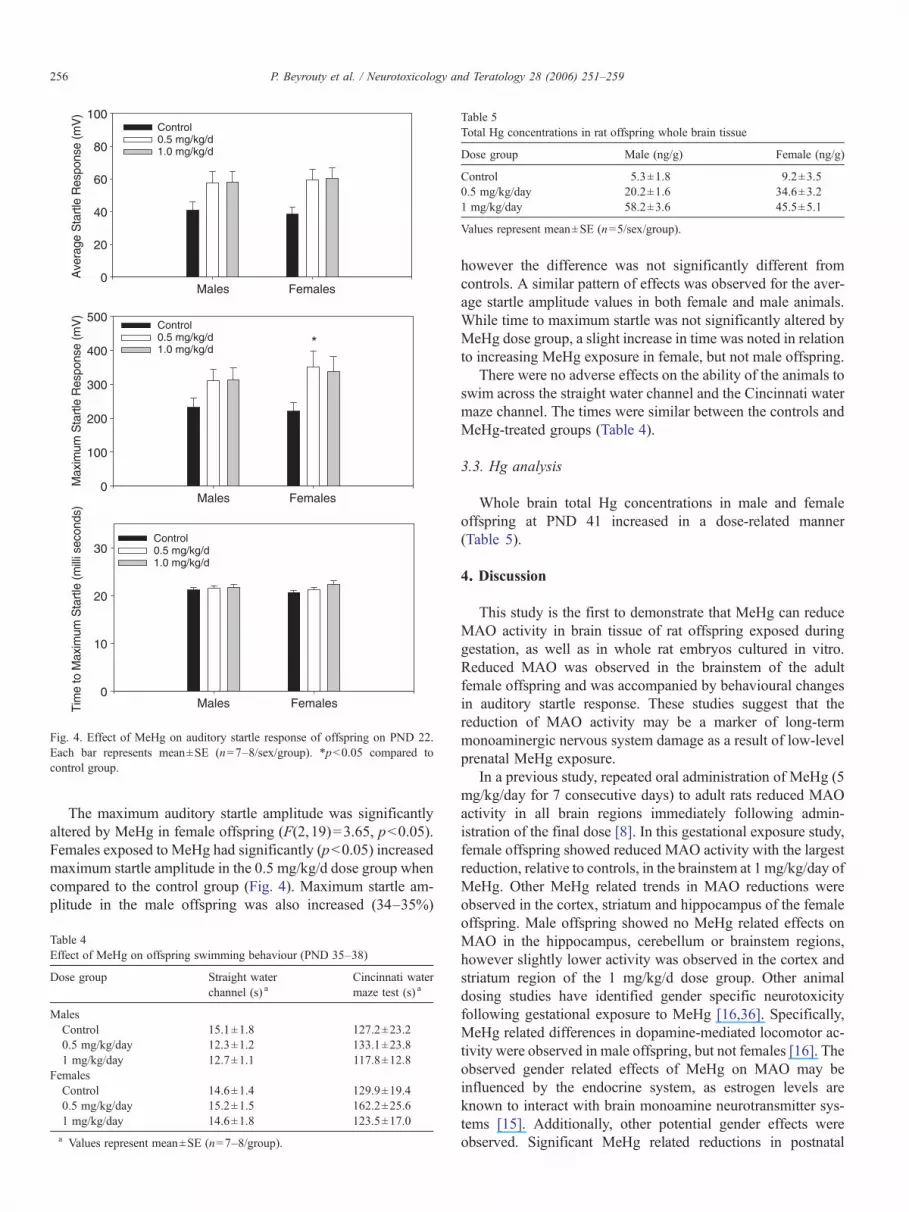
Fig. 4. Effect of MeHg on auditory startle response of offspring on PND 22.Each bar represents mean±SE (n=7–8/sex/group). *pb0.05 compared tocontrol group.
Table 5Total Hg concentrations in rat offspring whole brain tissue
Dose group Male (ng/g) Female (ng/g)
Control 5.3±1.8 9.2±3.50.5 mg/kg/day 20.2±1.6 34.6±3.21 mg/kg/day 58.2±3.6 45.5±5.1
Values represent mean±SE (n=5/sex/group).
256 P. Beyrouty et al. / Neurotoxicology and Teratology 28 (2006) 251–259
The maximum auditory startle amplitude was significantlyaltered by MeHg in female offspring (F(2,19)=3.65, pb0.05).Females exposed to MeHg had significantly (pb0.05) increasedmaximum startle amplitude in the 0.5 mg/kg/d dose group whencompared to the control group (Fig. 4). Maximum startle am-plitude in the male offspring was also increased (34–35%)
Table 4Effect of MeHg on offspring swimming behaviour (PND 35–38)
Dose group Straight waterchannel (s) a
Cincinnati watermaze test (s) a
MalesControl 15.1±1.8 127.2±23.20.5 mg/kg/day 12.3±1.2 133.1±23.81 mg/kg/day 12.7±1.1 117.8±12.8
FemalesControl 14.6±1.4 129.9±19.40.5 mg/kg/day 15.2±1.5 162.2±25.61 mg/kg/day 14.6±1.8 123.5±17.0a Values represent mean±SE (n=7–8/group).
however the difference was not significantly different fromcontrols. A similar pattern of effects was observed for the aver-age startle amplitude values in both female and male animals.While time to maximum startle was not significantly altered byMeHg dose group, a slight increase in time was noted in relationto increasing MeHg exposure in female, but not male offspring.
There were no adverse effects on the ability of the animals toswim across the straight water channel and the Cincinnati watermaze channel. The times were similar between the controls andMeHg-treated groups (Table 4).
3.3. Hg analysis
Whole brain total Hg concentrations in male and femaleoffspring at PND 41 increased in a dose-related manner(Table 5).
4. Discussion
This study is the first to demonstrate that MeHg can reduceMAO activity in brain tissue of rat offspring exposed duringgestation, as well as in whole rat embryos cultured in vitro.Reduced MAO was observed in the brainstem of the adultfemale offspring and was accompanied by behavioural changesin auditory startle response. These studies suggest that thereduction of MAO activity may be a marker of long-termmonoaminergic nervous system damage as a result of low-levelprenatal MeHg exposure.
In a previous study, repeated oral administration of MeHg (5mg/kg/day for 7 consecutive days) to adult rats reduced MAOactivity in all brain regions immediately following admin-istration of the final dose [8]. In this gestational exposure study,female offspring showed reduced MAO activity with the largestreduction, relative to controls, in the brainstem at 1 mg/kg/day ofMeHg. Other MeHg related trends in MAO reductions wereobserved in the cortex, striatum and hippocampus of the femaleoffspring. Male offspring showed no MeHg related effects onMAO in the hippocampus, cerebellum or brainstem regions,however slightly lower activity was observed in the cortex andstriatum region of the 1 mg/kg/d dose group. Other animaldosing studies have identified gender specific neurotoxicityfollowing gestational exposure to MeHg [16,36]. Specifically,MeHg related differences in dopamine-mediated locomotor ac-tivity were observed in male offspring, but not females [16]. Theobserved gender related effects of MeHg on MAO may beinfluenced by the endocrine system, as estrogen levels areknown to interact with brain monoamine neurotransmitter sys-tems [15]. Additionally, other potential gender effects wereobserved. Significant MeHg related reductions in postnatal

257P. Beyrouty et al. / Neurotoxicology and Teratology 28 (2006) 251–259
weight gain were only observed in males while significant in-creases in auditory startle response were only observed infemales. However, in both endpoints, similarMeHg related trendswere seen in the other gender. Therefore, it is possible thatthese differences may have been a result of insufficient statisticalpower and may not be directly due to gender differences.
There are several explanations for the observed MeHgrelated reductions in MAO activity in the offspring. In vitrostudies conducted with rat brain synaptosomes, reported thatMeHg inhibited MAO activity [8]. Structure and functionevaluations of MAO have demonstrated that several cysteineresidues are required for proper enzymatic function [21]. Hgcompounds have a high affinity for thiol groups [9], and itis likely that MeHg binds to cysteine residues resulting indisrupted structure and decreased MAO function as sug-gested by Gomes et al. [17]. Secondly, MeHg may decreaseMAO gene expression by directly disrupting transcriptionalprocesses. Exposure to MeHg has been shown to reducegene expression in Xenopus embryos [28]. However, it isalso possible that the observed decrease in enzyme activitymay reflect a MeHg-induced reduction in the number ofcells that express MAO relative to other brain cells. Thisreduction may be a result of cellular death or possibledevelopmental impairments caused by MeHg exposure duringdevelopment. In the current study, these effects of MeHg onMAO were observed when brain total-Hg concentrations werebelow 60 ng/g. These concentrations of Hg are lower than orequal to levels measured in autopsied brain tissue from victimsof the Minamata Bay poisoning episode [41]. It is alsoimportant to note that these biochemical changes weredetected 6 weeks after the final administration of MeHg tothe pregnant females, suggesting long-term effects. Futurestudies are required to evaluate temporal changes in MAOactivity following MeHg exposure during gestation.
Specific disruption of MAO function during developmentis associated with impairments in behaviour and neuronalgrowth in animal models [38,43]. In the current study, be-havioural tests were performed on the offspring to evaluateMeHg related abnormalities and monoaminergic nervoussystem dysfunction [11,20,23,43,44]. In this study, MeHg ap-peared to increase auditory startle response amplitude in theoffspring. MAO knockout mice have also been shown to havedisrupted auditory startle responses, although they show de-creased response activity following startle when compared towild-type mice [33]. The startle test is designed to investigatesimple sensorimotor neuronal circuitry in the lower brainstem,which involves monoamine neurotransmitters [23]. The markedreduction in brainstem MAO in female offspring, may suggestthe presence of a neurochemical change and monoaminergicneuron dysfunction. Developmental MeHg exposure may alterauditory startle response by impairing monoamine transmissionpathways in this brain region. These findings can be extrapolatedto observations in humans where gestational MeHg exposurewas correlated with delayed brainstem auditory-evoked-poten-tial latencies in adolescents from the Faroe Islands [31]. Thisis also consistent with studies showing that high develop-mental exposure to MeHg can induce severe lesions in the
brainstem of rats [22,37]. Collectively, these data suggest thatthe brainstem may be a significant target of developmentalMeHg neurotoxicity.
A previous study showed that the administration of specificinhibitors for both MAO-A and MAO-B (clorgyline anddeprenyl, respectively) to pregnant rats throughout gestation,resulted in a 50% reduction in brain MAO activity, increasedopen-field activity and a reduction in serotonergic neuroninnervations in the offspring [43]. In the current study, there wasa trend for increased motor activity, albeit transitory, for 1 mg/kg/day MeHg exposed male and female offspring. Thistransient MeHg-induced increase in motor activity waspreviously observed in offspring administered similar levelsof MeHg during gestation [16]. However, results from othermotor tests showed noMeHg related impairments in swimming,gait, and state of arousal behaviour. The increased motor activ-ity may suggest a transient and mild stage of developmentalMeHg neurotoxicity in these offspring.
Evaluations of the brains of MAO knockout mice indicatedimpairments in cortical somatosensory neuron barrel formation,suggesting a role for the enzyme in early neuronal development[5]. The nociceptive response pathways have been shown toinvolve the neurons of the somatosensory cortex [34]. Whilenot significant, hot plate test results suggest decreased res-ponse-time in offspring exposed to MeHg when compared tocontrols. Conversely, the tail flick test suggested that MeHgmay delay the response to thermal stimuli. Due to the lack ofsignificance and contradictory results in these tests, the effectsof MeHg on sensory pathways are inconclusive. MeHg devel-opmental neurotoxicity may occur through a wide range ofmechanisms, which may involve the disruption of variousproteins and neuronal cell dysfunction [6]. However, futuremechanistic studies should evaluate possible interactionsbetween MeHg exposure, MAO inhibition and impaired neu-ronal development.
The choice of MeHg dose in the embryo culture model wasbased on reported elevated concentrations in human cord bloodin Nunavik, Quebec [12,30] and Minamata, Japan [10]. Noeffect onMAO activity was observed at a dose level of 100 μg/L,which is higher than the average concentrations in the cord bloodcollected from the Faroe Islands where neurological effects werereported [18]. However, the lack of sensitivity may be due to thefact that rats are more resistant to MeHg toxicity (based on bodyweight) than humans [35]. Additionally, these embryos (GD13.5) were only cultured for a maximum of 12 h, as longerincubation periods resulted in hemorrhaging in embryos [27]. Ithas been estimated that gestational exposure to Hg during thepoisoning inMinamata may have been as high as 750 μg Hg/L inumbilical cord blood [10]. This concentration significantlyreducedMAO activity by 15% in whole rat embryos. In humans,this concentration resulted in severe neuronal damage andbehavioural abnormalities in the children, including hyperkine-sis, abnormal reflexes and ataxia [19]. Recently, a human studyconducted with fish consumers from Lac St. Pierre, Quebecshowed that Hg exposure (b20 μg/L blood Hg) was associatedwith reduced MAO activity in circulating blood platelets [39].These findings suggest that low-level exposure to MeHg from

258 P. Beyrouty et al. / Neurotoxicology and Teratology 28 (2006) 251–259
fish consumption may be sufficient to alter MAO activity inhumans and should be further investigated.
In conclusion, a low-level gestational exposure to MeHgsignificantly reduced MAO activity in female, but not in malerat offspring. Additionally, behavioural data suggest possiblegender-related differences with respect to MeHg neurotoxicity.While many of the behavioural tests were not significantlyaltered, MeHg exposure did alter the auditory startle responsesuggesting monoamine neurotransmission disruption. In vitroembryo culture experiments showed that MAO activity wasinhibited by MeHg concentrations that were similar to thoseobserved in Minamata Bay, however lower levels showed noeffects. This study provides evidence that MAO may serve as abiochemical marker of nervous system dysfunction related togestational MeHg exposure.
Acknowledgements
These studies were financially supported by the Collabo-rative Mercury Research Network (COMERN) and the Natu-ral Science and Engineering Research Council (NSERC)of Canada Discovery Grants to S. Kubow and H.M. Chan.ClinTrials BioResearch provided the facilities for the ani-mal experiments in-kind. Technical support from NiladriBasu, Donna Leggee, Laura Rostas and Leah Tivoli areacknowledged.
References
[1] Agency for Toxic Substances and Disease Registry (US) [ATSDR].Toxicological Profile for Mercury, 1999. Atlanta (GA): US Department ofHealth and Human Services, Public Health Service; 1999 Mar. 676 p.Available from: http://www.atsdr.cdc.gov/toxprofiles/ tp 46. html.
[2] F. Bakir, S.F. Damluji, L. Amin-Zaki, M. Murtadha, A. Khalidi, N.Y. al-Rawi, S. Tikriti, H.I. Dahahir, T.W. Clarkson, J.C. Smith, et al.,Methylmercury poisoning in Iraq, Science 181 (1973) 230–241.
[3] J.A. Blair, I.M. McGonnell, D.R. Newall, Strain difference in growth ofAHA and Han Wistar rat embryos in vitro and after extended culture invitro, Toxicol. In Vitro 10 (1996) 211–215.
[4] R. Brown, A.J. Silva, Molecular and cellular cognition; the unraveling ofmemory retrieval, Cell 117 (2004) 3–4.
[5] O. Cases, T. Vitalis, I. Seif, E. De Maeyer, C. Sotelo, P. Gaspar, Lack ofbarrels in the somatosensory cortex of monoamine oxidase A-deficientmice: role of a serotonin excess during the critical period, Neuron 16(1996) 297–307.
[6] A.F. Castoldi, T. Coccini, S. Ceccatelli, L. Manzo, Neurotoxicity andmolecular effects of methylmercury, Brain Res. Bull. 55 (2001) 197–203.
[7] A.F. Castoldi, T. Coccini, L. Manzo, Neurotoxic and molecular effects ofmethylmercury in humans, Rev. Environ. Health 18 (2003) 19–31.
[8] S.K. Chakrabarti, K.M. Loua, C. Bai, H. Durham, J.C. Panisset,Modulation of monoamine oxidase activity in different brain regions andplatelets following exposure of rats to methylmercury, Neurotoxicol.Teratol. 20 (1998) 161–168.
[9] T.W. Clarkson, The toxicology of mercury, Crit. Rev. Clin. Lab. Sci. 34(1997) 369–403.
[10] C. Dalgard, P. Grandjean, P.J. Jorgensen, P. Weihe, Mercury in theumbilical cord: implications for risk assessment for Minamata disease,Environ. Health Perspect. 102 (1994) 548–550.
[11] E. Dare, S. Fetissov, T. Hokfelt, H. Hall, S.O. Ogren, S. Ceccatelli, Effectsof prenatal exposure to methylmercury on dopamine-mediated locomotoractivity and dopamine D2 receptor binding, Naunyn-Schmiedeberg'sArch. Pharmacol. 367 (2003) 500–508.
[12] C. Després, A. Beuter, F. Richer, K. Poitras, A. Veilleux, P. Ayotte,E. Dewailly, D. Saint-Amour, G. Muckle, Neuromotor functions inInuit preschool children exposed to Pb, PCBs, and Hg, Neurotoxicol.Teratol. 27 (2005) 245–257.
[13] L.R. Faro, R. Duran, J.L. do Nascimento, M. Alfonso, C.W. Picanco-Diniz, Effects of methyl mercury on the in vivo release of dopamine and itsacidic metabolites DOPAC and HVA from striatum of rats, Ecotoxicol.Environ. Saf. 38 (1997) 95–98.
[14] L.R. Faro, R. Duran, J.L. Do Nascimento, D. Perez-Vences, M.Alfonso, Effects of successive intrastriatal methylmercury administra-tions on dopaminergic system, Ecotoxicol. Environ. Saf. 55 (2003)173–177.
[15] G. Fink, B.E. Sumner, R. Rosie, O. Grac, J.P. Quinn, Extrogen control ofcentral neurotransmission: effect on mood, mental state, and memory, Cell.Mol. Neurobiol. 16 (1996) 325–344.
[16] L. Gimenez-Llort, E. Ahlbom, E. Dare, M. Vahter, S. Ogren, S. Ceccatelli,Prenatal exposure to methylmercury changes dopamine-modulated motoractivity during early ontogeny: age and gender-dependent effects, Environ.Toxicol. Pharmacol. 9 (2001) 61–70.
[17] B. Gomes, H.G. Kloepfer, S. Oi, K.T. Yasunobu, The reaction ofsulfhydryl reagents with bovine hepatic monoamine oxidase. Evidence forthe presence of two cysteine residues essential for activity, Biochim.Biophys. Acta 438 (1976) 347–357.
[18] P. Grandjean, P. Weihe, R.F. White, F. Debes, S. Araki, K. Yokoyama, K.Murata, N. Sorensen, R. Dahl, P.J. Jorgensen, Cognitive deficit in 7-year-old children with prenatal exposure to methylmercury, Neurotoxicol.Teratol. 19 (1997) 417–428.
[19] M. Harada, Minimata disease: methylmercury poisoning in Japan causedby environmental pollution, Crit. Rev. Toxicol. 25 (1995) 1–24.
[20] D.P. Holschneider, K. Chen, I. Seif, J.C. Shih, Biochemical, behavioral,physiologic, and neurodevelopmental changes in mice deficient inmonoamine oxidase A or B, Brain Res. Bull. 56 (2001) 453–462.
[21] F. Hubalek, J. Pohl, D.E. Edmondson, Structural comparison of humanmonoamine oxidases A and B: mass spectrometry monitoring of cysteinereactivities, J. Biol. Chem. 278 (2003) 28612–28618.
[22] A. Kakita, K. Wakabayashi, M. Su, M. Sakamoto, F. Ikuta, H. Takahashi,Distinct pattern of neuronal degeneration in the fetal rat brain induced byconsecutive transplacental administration of methylmercury, Brain Res.859 (2000) 233–239.
[23] M. Koch, The neurobiology of startle, Prog. Neurobiol. 59 (1999)107–128.
[24] M. Krajl, A rapid microfluorometric determination of monoamine oxidase,Biochem. Parmacol. 14 (1965) 1683–1685.
[25] M.K. Lakshmana, T. Desiraju, T.R. Raju, Mercuric chloride-inducedalterations of levels of noradrenaline, dopamine, serotonin and acetylcho-line esterase activity in different regions of rat brain during postnataldevelopment, Arch. Toxicol. 67 (1993) 422–427.
[26] H. Lindstrom, J. Luthman, A. Oskarsson, J. Sundberg, L. Olson, Effects oflong-term treatment with methyl mercury on the developing rat brain,Environ. Res. 56 (1991) 158–169.
[27] J.-N. Liu, S. Kubow, H.M. Chan, Rat whole embryo culture (WEC) fromgestational day (GD) 13.5 to 14.5 using a roller bottle intermittent gassingsystem (manuscript in preparation).
[28] C.Monetti, D. Vigetti, M. Prati, E. Sabbioni, G. Bernardini, R. Gornati, Geneexpression in Xenopus embryos after methylmercury exposure: a search formolecular biomarkers, Environ. Toxicol. Chem. 21 (2002) 2731–2736.
[29] V.C. Moser, J.P. McCormick, J.P. Creason, R.C. MacPhail, Comparison ofchlordimeform and carbaryl using a functional observational battery,Fundam. Appl. Toxicol. 11 (1988) 189–206.
[30] G. Muckle, P. Ayotte, E.E. Dewailly, S.W. Jacobson, J.L. Jacobson,Prenatal exposure of the northern Quebec Inuit infants to environmentalcontaminants, Environ. Health Perspect. 109 (2001) 1291–1299.
[31] K. Murata, P. Weihe, E. Budtz-Jorgensen, P.J. Jorgensen, P. Grandjean,Delayed brainstem auditory evoked potential latencies in 14-year-oldchildren exposed to methylmercury, J. Pediatr. 144 (2004) 177–183.
[32] E.A. Neugebauer, G.L. Sans Cartier, B.J. Wakeford, Methods for thedetermination of metals in wildlife tissues using various atomic absorptionspectrophotometry techniques, Technical Report, Canadian Wildlife

259P. Beyrouty et al. / Neurotoxicology and Teratology 28 (2006) 251–259
Service, Environment Canada, Hull, Québec, Canada, 2000, 65 Availablefrom: http://www.cws-scf.ec.gc.ca/publications/Abstract Template.cfm?lang=e&id=322; CW69-5/337E.
[33] N.K. Popova, Y.A. Skrinskaya, T.G. Amstislavskaya, G.B. Vishnivets-kaya, I. Seif, E. de Meier, Behavioral characteristics of mice with geneticknockout of monoamine oxidase type A, Neurosci. Behav. Physiol. 31(2000) 597–602.
[34] D.D. Price, J.D. Greenspan, R. Dubner, Neurons involved in theexteroceptive function of pain, Pain 106 (2003) 215–219.
[35] D.C. Rice, Sensory and cognitive effects of developmental methylmercuryexposure in monkeys, and a comparison to effects in rodents,Neurotoxicology 17 (1996) 139–154.
[36] A.D. Rossi, E. Ahlbom, S.O. Ogren, P. Nicotera, S. Ceccatelli, Prenatalexposure to methylmercury alters locomotor activity of male but notfemale rats, Exp. Brain Res. 117 (1997) 428–436.
[37] M. Sakamoto, K. Wakabayashi, A. Kakita, T. Hitoshi, T. Adachi, A.Nakano, Widespread neuronal degeneration in rats following oraladministration of methylmercury during the postnatal developing phase:a model of fetal-type Minamata disease, Brain Res. 784 (1998) 351–354.
[38] J.C. Shih, Cloning, after cloning, knock-out mice, and physiologicalfunctions of MAO A and B, Neurotoxicology 25 (2004) 21–30.
[39] C.J. Stamler, N. Abdelouahab, C. Vanier, D. Mergler, H.M. Chan,Relationship between platelet monoamine oxidase-B (MAO-B) activityand mercury exposure in fish consumers from the Lac St. Pierre Region ofQuebec, Canada, Neurotoxicology (in press).
[40] C.J. Stamler, N. Basu, H.M. Chan, Biochemical markers of neurotoxicityin wildlife and human populations: considerations for method develop-ment, J. Toxicol. Environ. Health, A 68 (2005) 1413–1429.
[41] T. Takeuchi, K. Eto, The Pathology of Minimata Disease, KyushuUniversity Press, Hakozaki, Japan, 1999.
[42] C.V. Vorhees, Maze learning in rats: a comparison of performance in twowater mazes in progeny prenatally exposed to different doses of phenytoin,Neurotoxicol. Teratol. 9 (1987) 235–241.
[43] P.M. Whitaker-Azmitia, X. Zhang, C. Clarke, Effects of gestationalexposure to monoamine oxidase inhibitors in rats: preliminary behavioraland neurochemical studies, Neuropsychopharmacology 11 (1994)125–132.
[44] J.S. Yeomans, P.W. Frankland, The acoustic startle reflex: neurons andconnections, Brain Res. Brain Res. Rev. 21 (1995) 301–314.
[45] M. Zhou, N. Panchuk-Voloshina, A one-step fluorometric method for thecontinuous measurement of monoamine oxidase activity, Anal. Biochem.253 (1997) 169–174.