effect of iontophoresis on the in vitro trans-scleral transport of three single stranded...
TRANSCRIPT

European Journal of Pharmaceutical Sciences 49 (2013) 142–147
Contents lists available at SciVerse ScienceDirect
European Journal of Pharmaceutical Sciences
journal homepage: www.elsevier .com/ locate /e jps
Effect of iontophoresis on the in vitro trans-scleral transport of three singlestranded oligonucleotides
Silvia Pescina a, Maxim Antopolsky b, Patrizia Santi a, Sara Nicoli a, Lasse Murtomäki b,c,⇑a Department of Pharmacy, University of Parma, Parco Area delle Scienze 27/A, 43124 Parma, Italyb Centre for Drug Research, University of Helsinki, PO Box 56, FI-00014 Helsinki, Finlandc Aalto University, Department of Chemistry, PO Box 16100, FI-00076 Aalto, Finland
a r t i c l e i n f o
Article history:Received 3 September 2012Received in revised form 21 December 2012Accepted 3 February 2013Available online 26 February 2013
Keywords:OligonucleotideIontophoresisScleraPosterior eye
0928-0987/$ - see front matter � 2013 Elsevier B.V. Ahttp://dx.doi.org/10.1016/j.ejps.2013.02.010
⇑ Corresponding author at: Aalto University, DeBox 16100, FI-00076 Aalto, Finland. Tel.: +358 50 570
E-mail address: [email protected] (L. Murt
a b s t r a c t
Oligonucleotides represent a subject of clinical interest due to their potential ability to treat several dis-eases, including those affecting the posterior segment of the eye. Unfortunately, therapeutic oligonucle-otides are currently administered by means of highly invasive approaches, such as intravitreal injections.The aim of the present work was to study in vitro, across isolated bovine sclera, the effect of iontophoresison the transport of three single stranded oligonucleotides (ssDNA), 12-, 24- and 36-mer, selected as ref-erence compounds in view of a non-invasive drug delivery to the back of the eye.
All the three sequences were able to cross bovine sclera in vitro without iontophoresis. When anodaliontophoresis was applied, no change in flux was observed, while in the presence of cathodal iontopho-resis the permeability coefficients increased four-fold compared to passive conditions. This behavior canbe ascribed to the electrorepulsive mechanism, due to the negative charge of the nucleic acid backbone. Itwas also observed that the molecular weights of the three sequences did not affect trans-scleral trans-port, neither in passive, nor in current assisted permeation. Furthermore, increasing the current intensityfrom 1.75 mA to 3 mA, no effect on the trans-scleral transport of the 24-mer was noticed.
Although preliminary, the results demonstrate that cathodal iontophoresis enhances trans-scleraltransport of single stranded oligonucleotides and suggest its use as a novel non-invasive approach forthe treatment of diseases affecting the posterior segment of the eye.
� 2013 Elsevier B.V. All rights reserved.
1. Introduction
Nucleic acids are characterized by a wide therapeutic potential,since they are the tool of gene therapy, but can also play the role oftherapeutic agents (therapeutic oligonucleotides), as proposed forthe first time at the end of the 1970s by Zamecnik and Stephenson(1978) and Stephenson and Zamecnik (1978). The eye is particu-larly proper for this type of therapeutic approach, due to its peculiarcharacteristics. In fact, the eye bulb has a smaller size and is moreattainable than other organs; furthermore, several genes are in-volved in ocular pathologies (Fattal and Bochot, 2006; Colellaet al., 2009). Therapeutic oligonucleotides were introduced in clinicat the end of the 90s, when FDA and EMA approved fomivirsen(Vitravene™), an antisense oligonucleotide of 21 bases for thetreatment of cytomegalovirus retinitis in patients with acquiredimmunodeficiency (de Smet et al., 1999). Later, the same regulatoryauthorities have authorized the use of pegaptanib (Macugen�),an RNA aptamer with anti-VEGF165, in the treatment of the wet
ll rights reserved.
partment of Chemistry, PO6352; fax: +358 9 470 22580.omäki).
form of macular degeneration (Ng et al., 2004). Currently, severalclinical trials already completed or in development, involve secondgeneration antisense drugs such as siRNA bevasiranib (NationalInstitutes of Health, 2008) and iCo-007 (National Institutes ofHealth, 2012). Yet, the route of administration remains a barrier:since the target is generally the retina, in order to have an effect,therapeutic oligonucleotides are introduced into the eye by meansof injections (i.e. subconjunctival and intravitreal), slightly toler-ated and sometimes responsible for severe side effects, such asendophthalmitis and retinal detachment (Sampat and Garg, 2010).
In order to restrict the invasiveness, thanks to the relativelyhigh permeability of the sclera to macromolecules (Maurice andPolgar, 1977; Olsen et al., 1995; Ambati et al., 2000), trans-scleralroute has been suggested as a non-invasive or minimally invasiveapproach for the treatment of the retinal diseases. Several enhanc-ing strategies have been proposed, including microneedles (Jianget al., 2007, 2009; Patel et al., 2011), therapeutic ultrasound (Che-ung et al., 2010) and iontophoresis application. The latter has dem-onstrated its in vitro ability to promote trans-scleral transport ofmacromolecules such as proteins (Chopra et al., 2010) or monoclo-nal antibodies (Chopra et al., 2010; Pescina et al., 2010). Interestingresults were also reached by applying trans-scleral iontophoresis

S. Pescina et al. / European Journal of Pharmaceutical Sciences 49 (2013) 142–147 143
in vivo to deliver genetic material (Asahara et al., 2001; Voigt et al.,2002; Souied et al., 2008) or proteins (Molokhia et al., 2009).
The purpose of this work was to study in vitro the effect ofiontophoresis on the permeation of three single stranded oligonu-cleotides, chosen as model compounds of antisense oligonucleo-tides, across isolated bovine sclera. Both anodal and cathodaliontophoresis were used in order to determine whether electroos-mosis or electrorepulsion was the main mechanism of theiontophoretic flux enhancement. Additionally, the effect of two dif-ferent current densities was evaluated by employing the 24-mersequence.
2. Materials and methods
2.1. Materials
Hepes buffer (HEPES): 5.96 g/l Hepes (4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid, purchased from Sigma (St. Louis,MO, USA), 9.0 g/l NaCl; pH 7.4 with 5 N NaOH. Three singlestranded DNAs(ssDNA) were synthesized and labeled at the 50
end with fluorescein: a 12-mer (50-GTT CCA TTC ATA-30, MW4193.4 Da), a 24-mer (50-ACC TGG GAC ATC GTT CCA TTC ATA-30,MW 7885.8 Da) and a 36-mer (50-ACC TGG GAC ATC GTT CCATTC ATA GTT CCA TTC ATA-30, MW 11543.2 Da). All the oligonucle-otides were synthesized with ASM-800 DNA/RNA synthesizerfollowing standard DNA synthetic protocol, using monomers andUS-II solid support purchased from Glen Research. Cleavage/depro-tection was performed following standard US-II solid supportcleavage protocol employing 3.5 M ammonia solution in methanolfollowed with 28% aqueous ammonia solution treatment. Crudeoligonucleotides were purified by i.e. HPLC (TSKgel DEAE-2SW5 lm, 4.6 � 250 mm column and linear gradient from 0% to 60%of B within 30 min (A – 0.1 M sodium acetate, NaAc, in 20% aceto-nitrile, ACN, pH = 8.0; B – 0.1 M NaAc and 0.4 M NaClO4 in 20%ACN, pH = 8.0) at 1 ml/min flow rate), desalted by gel permeationchromatography (Sephadex G-10 10 � 150 mm column, elutionwith water at flow rate 0.5 ml/min) and kept frozen in water solu-tion at �18 �C. Purity of all oligonucleotides was not less than 95%according to RP HPLC (XBridghe™ C18 5 lm, 4.6 � 150 mm col-umn and linear gradient from 0% to 50% of B within 30 min (A –0.05 M triethyl ammonium acetate, TEAA, in water, B – 0.05 MTEAA in 70% ACN) at 1 ml/min flow rate).
2.2. Tissue preparation
Bovine eyes were obtained from a slaughterhouse (Atria Ltd.,Kuopio, Finland). Eye bulbs were transported in an ice bath tothe laboratory and, after removal the muscular portion, the sclerawas isolated from other ocular tissues and dissected with the scal-pel. Fragments of sclera, having an area of about 1 cm2, were frozenat �80 �C, and then stored at �20 �C until used. Freezing and stor-age procedures did not damage the tissue and consequently didnot affect the sclera permeability towards macromolecules, as pre-viously demonstrated (Pescina et al., 2011).
2.3. Permeation experiments
Permeation experiments were performed in Franz-type diffu-sion cells (permeation area 0.6 cm2), with the conjunctiva side ofbovine sclera facing the donor compartment. The donor compart-ment contained 0.3 ml of a 250 lg/ml of oligonucleotide (either12, 24 or 36-mer) in Hepes buffer, while the receptor compartmentwas filled with 4 ml of Hepes buffer, thermostated at 37 �C andstirred with a magnetic stirrer (350 rpm). Each experiment lastedfor 5 h, during which the receptor solution was sampled at
predetermined times for the quantification of oligonucleotidepermeated.
In the iontophoretic experiments, silver–silver chloride elec-trodes and salt bridges (agar 2% w/w in 1 M KCl) were used. Thecurrent (intensity 1.75 mA, density 3.0 mA/cm2; anodal or cath-odal) was applied for 2 h by a constant current generator (6614C,Hewlett Packard, Palo Alto, Ca, USA). In the case of anodal ionto-phoresis Ag electrode (anode) was put in contact with the donorcompartment, while AgCl electrode (cathode) was connected tothe receiving phase. When cathodal iontophoresis was applied,the previous set-up was inverted. After breaking the applicationof iontophoresis, passive diffusion through the tissue was followedfor 3 h.
By using the 24-mer oligonucleotide, chosen as reference of thethree sequences, the effect of the applied current (current intensity3.0 mA, current density 5 mA/cm2) was evaluated.
2.4. Data processing
Trans-scleral flux (J, lg/cm2 h) was calculated as the slope of theregression line at pseudo steady-state, while the apparent perme-ability coefficients (P, cm/s) were obtained by dividing flux perdrug donor concentration (CD, lg/ml). Lag time (min) was deter-mined by extrapolation of the linear part of the permeation profilesto the time axis. Furthermore, passive data were processed byapplying the appropriate solution of Fick’s law in Eq. (1) (Crank,1956):
Q ¼ KCvehhDt
h2 �16� 2
p2
X1n¼1
ð�1Þn
n2 exp �n2p2Dt
h2
� �" #ð1Þ
where Q (lg) is the cumulative amount of drug permeated per unitarea at time t, Cveh (lg/ml) is the concentration of the drug in the do-nor vehicle, K is the sclera/vehicle partition coefficient, D (cm2/h) thediffusion coefficient and h (cm) the diffusion path length. The fitswere done with Microsoft Excel solver, resulting in the values of K,D and h; the first four terms of the series were included in the fits.The average error associated with each fitted value was in the range0.01–0.004. The permeability coefficient P (cm/s) was calculated asK � D/h. The enhancement factor (EF) due to the iontophoresisapplication was calculated as the ratio between Piontophoresis andPpassive.
2.5. Oligonucleotides quantification
The concentration of oligonucleotides in the receiver phasewere determined by fluorescence (excitation wavelength 485 nm,emission wavelength 535 nm) with multi-label plate reader (Vik-tor2 1420, Wallac). Three different calibration curves were builtby employing standard solutions of each oligonucleotide; theirrange of concentrations was between 8.29 and 238.47 nM for 12-mer, between 0.95 and 95.11 nM for 24-mer and between 2.62and 104.82 nM for 36-mer. Blank permeation experiments wereperformed by filling both donor and receiving compartments withHepes buffer, in order to check the interference from the tissueitself.
2.6. Statistical analysis
Each experiment was replicated at least 3 times and the resultswere expressed as the mean ± standard error of the mean (sem).Statistical differences were determined by ANOVA followed byBonferroni test.

144 S. Pescina et al. / European Journal of Pharmaceutical Sciences 49 (2013) 142–147
3. Results and discussion
3.1. Permeation experiments
Three single stranded oligodeoxynucleotides of different length(12, 24 and 36-mer) were selected as model for antisense oligonu-cleotides. At the beginning, in vitro permeation experiments wereconducted in passive conditions. As shown in Fig. 1, bovine sclerawas permeable to all oligonucleotides tested; the correspondingpermeation parameters are reported in Table 1.
In all three cases, the lag time was greater than 2 h which israther high if compared to neutral macromolecules, such as bev-acizumab (anti-VEGF monoclonal antibody; MW 149 kDa) appliedon human sclera or FD-40 (fluorescently labeled dextran; MW39 kDa) through porcine sclera, for which the lag time in passivepermeation was 30 min (Pescina et al., 2010) and 40 min (Pescinaet al., 2011), respectively. This behavior probably depends on thepolyanionic nature of the oligonucleotides, which induces repul-sion towards the charges of the same sign present in the extracel-lular matrix of the sclera (Cyenge et al., 2003; Taylor and Parker,2003), as well as to the different origin of the tissues employed(human/porcine/bovine). Shuler et al. (2004) studied the passivetransport in vitro through isolated human sclera using a fluores-cently labeled single stranded oligonucleotide, consisting of thesame sequence of the 24-mer as employed in the present work.The permeability coefficient, calculated after 24 h, was equal to(7.6 ± 1.8) � 10�7 cm/s, comparable to that obtained in this workfor the bovine sclera (8.47 ± 0.51) � 10�7 cm/s, after 5 h of perme-ation. The application of Eq. (1) has led to the results shown inTable 2. P values calculated result comparable to those previouslycalculated (Table 1), indicating the quality of the fitting. K valuesobtained can be considered plausible due to the hydrophilic natureof the oligonucleotides and extensive hydration of the tissue. Con-cerning D, a negative trend as function of the molecular weight wasobserved (Table 2); the diffusion coefficients fit with the Mark–Houwink equation as shown in Fig. 3 (Tanford, 1961), withD = 3.34 � 10�5 N�0.500 cm2/s, where N is the number of bases,even if the differences were not statistical. That is in agreementwith results reported in literature and referred to single strandedDNA with length between 5 and 50 bases (Reineck et al., 2010).The exponent �0.5 indicates that the conformation of the three se-quences is a random coil (Kontturi et al. 2002).
In order to evaluate the possible effect of promoting the trans-scleral transport of the oligonucleotides, in vitro permeation
0 60 120 180 240 300 3600.0
0.5
1.0
1.5
2.0
2.5
time (min)
olig
onuc
leot
ide
perm
eate
d (µ
g/cm
2 )
Fig. 1. Passive permeation profiles of 12-mer (d), 24-mer (s) and 36-mer (j)oligonucleotides across isolated bovine sclera.
Fig. 2. Permeation profiles referred to 12-mer (panel a), 24-mer (panel b) and 36-mer (panel c); permeation conditions: passive (d), anodal iontophoresis (j),cathodal iontophoresis (�). In iontophoretic conditions current was applied for 2 h(0–1200) and after current breaking, post-iontophoretic diffusion was followed for3 h.
experiments were conducted in the presence of iontophoresis, ap-plied for 2 h. As expected, the application of the anodal iontopho-resis did not led to any enhancement in the oligonucleotidestransport, while the use of opposite polarity allowed a significantpromotion of the transport across the barrier (Fig. 2 and Table 1).Sclera is a porous and negatively charged membrane at physiolog-ical pH: due to the presence of negatively charged polymers (hyal-uronic acid and glycosaminoglycans), its isoelectric point isbetween 3 and 4 (Nicoli et al., 2009; Güngör et al., 2010). Therefore,the application of anodal iontophoresis causes a convective flow(electroosmosis) in the anode-to-cathode direction, which en-hances the transport towards cathode, while in cathodal iontopho-resis, electroosmosis opposes it. But since the oligonucleotidebackbone has many phosphate groups and is negatively charged

Table 1Permeation parameters of oligonucleotides through isolated bovine sclera.
Oligonucleotide Permeation condition J (lg/cm2h) P (10�7 cm/s) Lag time (min) EF
12-mer Passivea 0.68 ± 0.17 7.51 ± 1.93 171.1 ± 10.3 –Anodal 1.75 mAa 1.12 ± 0.21 12.40 ± 2.38 152.2 ± 1.2 1.65 ± 0.32Cathodal 1.75 mAb 2.47 ± 0.59 27.44 ± 5.71 60.4 ± 3.5 3.65 ± 0.76
24-mer Passivea 0.76 ± 0.05 8.47 ± 0.51 142.8 ± 7.5 –Anodal 1.75 mAa 0.83 ± 0.14 9.27 ± 1.58 158.4 ± 2.1 1.09 ± 0.19Anodal 3.0 mAa 0.62 ± 0.18 6.93 ± 2.00 168.5 ± 6.3 0.65 ± 0.24Cathodal 1.75 mAb 3.04 ± 0.77 33.80 ± 8.56 61.8 ± 6.7 3.99 ± 1.01Cathodal 3.0 mAb 2.07 ± 0.68 23.00 ± 7.60 53.4 ± 8.9 2.72 ± 0.90
36-mer Passivea 0.56 ± 0.15 6.27 ± 1.69 152.1 ± 13.8 –Anodal 1.75 mAa 0.50 ± 0.17 5.56 ± 1.94 170.3 ± 10.6 0.89 ± 0.31Cathodal 1.75 mAb 2.64 ± 1.11 29.3 ± 12.4 74.3 ± 13.2 4.67 ± 1.97
Data are mean ± sem.a Parameters calculated between 180 and 300 min.b Parameters calculated between 80 and 120 min.
Table 2Parameters calculated by applying the solution of Fick’s law (Eq. (1)) to data collectedfrom passive permeation experiments.
Oligonucleotide D (10�6 cm2/s) K P (10�7 cm/s)
12-mer 9.70 ± 4.04 1.17 ± 0.17 14.53 ± 5.3224-mer 6.54 ± 0.21 1.00 ± 0.01 11.90 ± 0.4836-mer 5.77 ± 0.55 1.00 ± 0.01 10.28 ± 1.38
Data are mean ± sem.
Fig. 3. Diffusion coefficients for the three oligonucleotides, calculated by applyingEq. (1) (see also Table 2), fit with the Mark–Houwink equation.
S. Pescina et al. / European Journal of Pharmaceutical Sciences 49 (2013) 142–147 145
at pH 7.4, electrorepulsion is predominant over electroosmosis,and it is therefore possible to observe a promotion of the trans-scleral transport in the presence of cathodal iontophoresis.
It is interesting to note that there were no substantial differ-ences in the transport of the three oligonucleotides through the bo-vine sclera, neither in passive conditions, nor in the presence ofanodal or cathodal iontophoresis (Fig. 2). In the case of passivediffusion, the similarity in P can be ascribed to the relatively smalldifferences in MW between the oligonucleotides tested; the subtleeffect of MW on the diffusion coefficient is analyzed in Fig. 3.Concerning the iontophoretic results, the charge/mass ratio, thereal driving-force of the permeation, might be considered constantfor all three sequences. This is also corroborated by the study(Kontturi et al. 2002) where it was found that the effective chargenumber of 15-, 22- and 30-mer remained at ca. 25% of the
stoichiometric one. Thus, it is reasonable to assume that the molec-ular weight as such is not a critical parameter. This may be attrib-uted mainly to the high porosity of the sclera, but also to thedifferent sequence of bases of the three oligonucleotides whichmight be responsible of the assumption of different conformations.In fact, as reported in literature, like proteins, also single-strandednucleic acids can adopt complex secondary and tertiary structures(Goodchild, 2011), with consequent effects on the surface chargedensity, as well as on their shape and hydrodynamic radii.
The evidence that the molecular weight did not represent a dis-criminating factor for the trans-scleral diffusion of the oligonucle-otides is in contrast with some results reported in literature wherecathodal iontophoresis is applied for promoting transdermal trans-port of three oligonucleotides, having a comparable length to thosestudied in this work (10-, 20-, 30-mer). In fact, the flux at steady-state conditions was found to be inversely proportional to thelength of the oligonucleotide sequence (Oldenburg et al., 1995).However, it is important to emphasize that the two tissues, dueto their different structure, have substantially different permeabil-ity. Furthermore, it is worth noting that factors other than sizeinfluence oligonucleotides transport, such as the base sequence(Brand et al., 1997). In fact, as has been demonstrated by the sameauthors, for the smaller compounds transport generally decreasedwith increasing size, but there were several oligonucleotides thatdid not follow this scheme.
Finally, the post-iontophoretic transport across sclera was ana-lyzed. As shown in Fig. 2, following the cathodal iontophoresis,there was an immediate reduction in the slope of the permeationprofile as soon as the current was turned off. Post-iontophoreticflux became comparable to the passive one, indicating that nossDNA accumulation took place in the sclera during current appli-cation. This behavior is different from that previously observed inthe case of neutral macromolecule (40 kDa dextran), where a sig-nificant ‘‘reservoir effect’’ was detected (Pescina et al., 2011), andcan be ascribed to the negative charges on the oligonucleotidebackbone.
3.2. Effect of current intensity
By increasing the current intensity from 1.75 to 3 mA (currentdensity from 2.9 to 5 mA/cm2), no further increase of the 24-mertrans-scleral transport was detected, neither in anodal, nor in cath-odal conditions, as shown in Fig. 4 and reported in Table 1. A sim-ilar behavior was observed by using a 40 kDa dextran, although inthat case, the mechanism of enhancement was electroosmotic,rather than electrorepulsive, due to the neutral nature of the per-meant (Pescina et al., 2011). Anyway, this result was in contrast
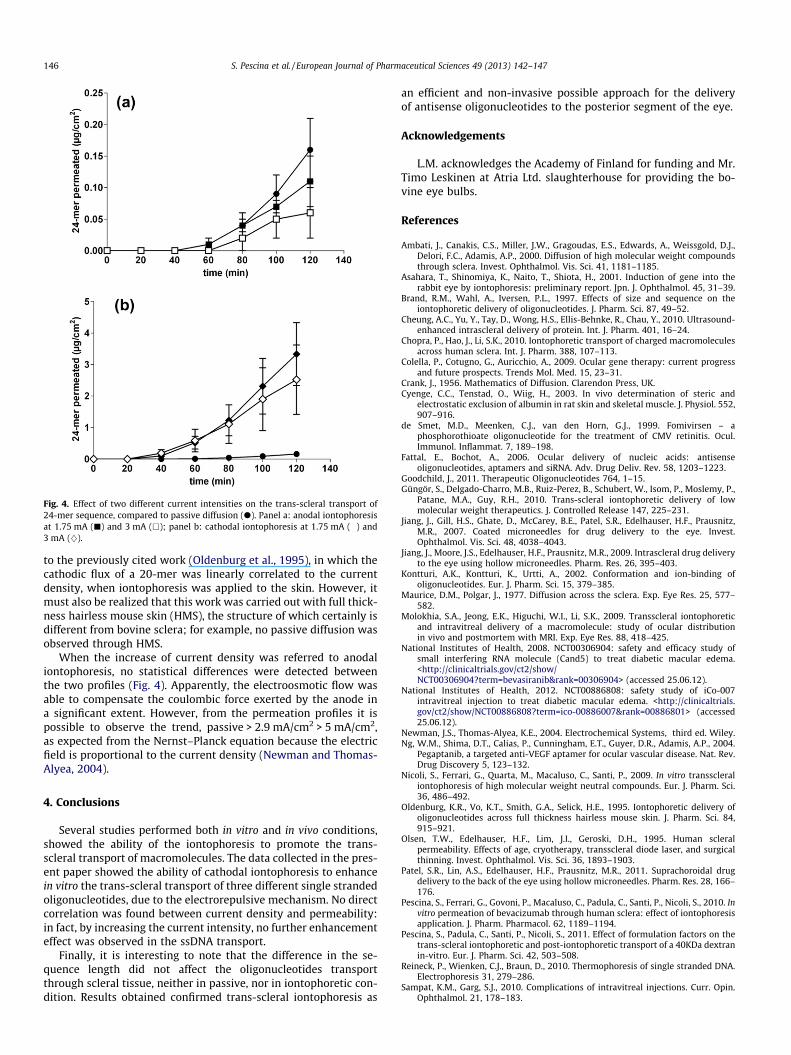
Fig. 4. Effect of two different current intensities on the trans-scleral transport of24-mer sequence, compared to passive diffusion (d). Panel a: anodal iontophoresisat 1.75 mA (j) and 3 mA (h); panel b: cathodal iontophoresis at 1.75 mA (�) and3 mA (}).
146 S. Pescina et al. / European Journal of Pharmaceutical Sciences 49 (2013) 142–147
to the previously cited work (Oldenburg et al., 1995), in which thecathodic flux of a 20-mer was linearly correlated to the currentdensity, when iontophoresis was applied to the skin. However, itmust also be realized that this work was carried out with full thick-ness hairless mouse skin (HMS), the structure of which certainly isdifferent from bovine sclera; for example, no passive diffusion wasobserved through HMS.
When the increase of current density was referred to anodaliontophoresis, no statistical differences were detected betweenthe two profiles (Fig. 4). Apparently, the electroosmotic flow wasable to compensate the coulombic force exerted by the anode ina significant extent. However, from the permeation profiles it ispossible to observe the trend, passive > 2.9 mA/cm2 > 5 mA/cm2,as expected from the Nernst–Planck equation because the electricfield is proportional to the current density (Newman and Thomas-Alyea, 2004).
4. Conclusions
Several studies performed both in vitro and in vivo conditions,showed the ability of the iontophoresis to promote the trans-scleral transport of macromolecules. The data collected in the pres-ent paper showed the ability of cathodal iontophoresis to enhancein vitro the trans-scleral transport of three different single strandedoligonucleotides, due to the electrorepulsive mechanism. No directcorrelation was found between current density and permeability:in fact, by increasing the current intensity, no further enhancementeffect was observed in the ssDNA transport.
Finally, it is interesting to note that the difference in the se-quence length did not affect the oligonucleotides transportthrough scleral tissue, neither in passive, nor in iontophoretic con-dition. Results obtained confirmed trans-scleral iontophoresis as
an efficient and non-invasive possible approach for the deliveryof antisense oligonucleotides to the posterior segment of the eye.
Acknowledgements
L.M. acknowledges the Academy of Finland for funding and Mr.Timo Leskinen at Atria Ltd. slaughterhouse for providing the bo-vine eye bulbs.
References
Ambati, J., Canakis, C.S., Miller, J.W., Gragoudas, E.S., Edwards, A., Weissgold, D.J.,Delori, F.C., Adamis, A.P., 2000. Diffusion of high molecular weight compoundsthrough sclera. Invest. Ophthalmol. Vis. Sci. 41, 1181–1185.
Asahara, T., Shinomiya, K., Naito, T., Shiota, H., 2001. Induction of gene into therabbit eye by iontophoresis: preliminary report. Jpn. J. Ophthalmol. 45, 31–39.
Brand, R.M., Wahl, A., Iversen, P.L., 1997. Effects of size and sequence on theiontophoretic delivery of oligonucleotides. J. Pharm. Sci. 87, 49–52.
Cheung, A.C., Yu, Y., Tay, D., Wong, H.S., Ellis-Behnke, R., Chau, Y., 2010. Ultrasound-enhanced intrascleral delivery of protein. Int. J. Pharm. 401, 16–24.
Chopra, P., Hao, J., Li, S.K., 2010. Iontophoretic transport of charged macromoleculesacross human sclera. Int. J. Pharm. 388, 107–113.
Colella, P., Cotugno, G., Auricchio, A., 2009. Ocular gene therapy: current progressand future prospects. Trends Mol. Med. 15, 23–31.
Crank, J., 1956. Mathematics of Diffusion. Clarendon Press, UK.Cyenge, C.C., Tenstad, O., Wiig, H., 2003. In vivo determination of steric and
electrostatic exclusion of albumin in rat skin and skeletal muscle. J. Physiol. 552,907–916.
de Smet, M.D., Meenken, C.J., van den Horn, G.J., 1999. Fomivirsen – aphosphorothioate oligonucleotide for the treatment of CMV retinitis. Ocul.Immunol. Inflammat. 7, 189–198.
Fattal, E., Bochot, A., 2006. Ocular delivery of nucleic acids: antisenseoligonucleotides, aptamers and siRNA. Adv. Drug Deliv. Rev. 58, 1203–1223.
Goodchild, J., 2011. Therapeutic Oligonucleotides 764, 1–15.Güngör, S., Delgado-Charro, M.B., Ruiz-Perez, B., Schubert, W., Isom, P., Moslemy, P.,
Patane, M.A., Guy, R.H., 2010. Trans-scleral iontophoretic delivery of lowmolecular weight therapeutics. J. Controlled Release 147, 225–231.
Jiang, J., Gill, H.S., Ghate, D., McCarey, B.E., Patel, S.R., Edelhauser, H.F., Prausnitz,M.R., 2007. Coated microneedles for drug delivery to the eye. Invest.Ophthalmol. Vis. Sci. 48, 4038–4043.
Jiang, J., Moore, J.S., Edelhauser, H.F., Prausnitz, M.R., 2009. Intrascleral drug deliveryto the eye using hollow microneedles. Pharm. Res. 26, 395–403.
Kontturi, A.K., Kontturi, K., Urtti, A., 2002. Conformation and ion-binding ofoligonucleotides. Eur. J. Pharm. Sci. 15, 379–385.
Maurice, D.M., Polgar, J., 1977. Diffusion across the sclera. Exp. Eye Res. 25, 577–582.
Molokhia, S.A., Jeong, E.K., Higuchi, W.I., Li, S.K., 2009. Transscleral iontophoreticand intravitreal delivery of a macromolecule: study of ocular distributionin vivo and postmortem with MRI. Exp. Eye Res. 88, 418–425.
National Institutes of Health, 2008. NCT00306904: safety and efficacy study ofsmall interfering RNA molecule (Cand5) to treat diabetic macular edema.<http://clinicaltrials.gov/ct2/show/NCT00306904?term=bevasiranib&rank=00306904> (accessed 25.06.12).
National Institutes of Health, 2012. NCT00886808: safety study of iCo-007intravitreal injection to treat diabetic macular edema. <http://clinicaltrials.gov/ct2/show/NCT00886808?term=ico-00886007&rank=00886801> (accessed25.06.12).
Newman, J.S., Thomas-Alyea, K.E., 2004. Electrochemical Systems, third ed. Wiley.Ng, W.M., Shima, D.T., Calias, P., Cunningham, E.T., Guyer, D.R., Adamis, A.P., 2004.
Pegaptanib, a targeted anti-VEGF aptamer for ocular vascular disease. Nat. Rev.Drug Discovery 5, 123–132.
Nicoli, S., Ferrari, G., Quarta, M., Macaluso, C., Santi, P., 2009. In vitro transscleraliontophoresis of high molecular weight neutral compounds. Eur. J. Pharm. Sci.36, 486–492.
Oldenburg, K.R., Vo, K.T., Smith, G.A., Selick, H.E., 1995. Iontophoretic delivery ofoligonucleotides across full thickness hairless mouse skin. J. Pharm. Sci. 84,915–921.
Olsen, T.W., Edelhauser, H.F., Lim, J.I., Geroski, D.H., 1995. Human scleralpermeability. Effects of age, cryotherapy, transscleral diode laser, and surgicalthinning. Invest. Ophthalmol. Vis. Sci. 36, 1893–1903.
Patel, S.R., Lin, A.S., Edelhauser, H.F., Prausnitz, M.R., 2011. Suprachoroidal drugdelivery to the back of the eye using hollow microneedles. Pharm. Res. 28, 166–176.
Pescina, S., Ferrari, G., Govoni, P., Macaluso, C., Padula, C., Santi, P., Nicoli, S., 2010. Invitro permeation of bevacizumab through human sclera: effect of iontophoresisapplication. J. Pharm. Pharmacol. 62, 1189–1194.
Pescina, S., Padula, C., Santi, P., Nicoli, S., 2011. Effect of formulation factors on thetrans-scleral iontophoretic and post-iontophoretic transport of a 40KDa dextranin-vitro. Eur. J. Pharm. Sci. 42, 503–508.
Reineck, P., Wienken, C.J., Braun, D., 2010. Thermophoresis of single stranded DNA.Electrophoresis 31, 279–286.
Sampat, K.M., Garg, S.J., 2010. Complications of intravitreal injections. Curr. Opin.Ophthalmol. 21, 178–183.

S. Pescina et al. / European Journal of Pharmaceutical Sciences 49 (2013) 142–147 147
Shuler, R.K., Dioguardi, P.K., Henjy, C., Nickerson, J.M., Cruysberg, L.P., Edelhauser,H.F., 2004. Scleral permeability of a small, single-stranded oligonucleotide. J.Ocul. Pharmacol. Ther. 20, 159–168.
Souied, E.H., Reid, S.N.M., Piri, N.I., Lerner, L.E., Nusinowitz, S., Farber, D.B., 2008.Non-invasive gene transfer by iontophoresis for therapy of an inherited retinaldegeneration. Exp. Eye Res. 87, 168–175.
Stephenson, M.L., Zamecnik, P.C., 1978. Inhibition of Rous sarcoma viral RNAtranslation by a specific oligodeoxynucleotide. Proc. Nat. Acad. Sci. USA 75,285–288.
Tanford, C., 1961. Physical Chemistry of Macromolecules. Wiley, New York.
Taylor, A.E., Parker, J.C., 2003. Interstitial excluded volumes: the effect of charge. J.Physiol. 553, 333.
Voigt, M., de Kozak, Y., Halhal, M., Courtois, Y., Behar-Cohen, F., 2002. Down-regulation of NOSII gene expression by iontophoresis of anti-senseoligonucleotide in endotoxin-induced uveitis. Biochem. Biophys. Res.Commun. 295, 336–341.
Zamecnik, P.C., Stephenson, M.L., 1978. Inhibition of rous sarcoma virus replicationand cell transformation by a specific oligodeoxynucleotide. Proc. Nat. Acad. Sci.USA 75, 280–284.