ecophysiology of abundant denitrifying bacteria in activated sludge: ecophysiology of denitrifying...
TRANSCRIPT

Ecophysiologyofabundantdenitrifying bacteria inactivated sludgeTrine Rolighed Thomsen, Yunhong Kong & Per Halkjær Nielsen
Section of Environmental Engineering, Department of Biotechnology, Chemistry and Environmental Engineering, Aalborg University, Denmark
Correspondence: Per H. Nielsen, Section of
Environmental Engineering, Department of
Biotechnology, Chemistry and Environmental
Engineering, Sohngaardsholmsvej 57,
DK-9000 Aalborg, Denmark.
Tel.: 145 96358503; fax: 145 96350558;
e-mail: [email protected]
Received 12 July 2006; revised 30 January
2007; accepted 30 January 2007.
First published online 28 March 2007.
DOI:10.1111/j.1574-6941.2007.00309.x
Editor: Michael Wagner
Keywords
denitrifiers; Betaproteobacteria ; activated
sludge; ecophysiology; microautoradiography.
Abstract
The abundance of potential denitrifiers in full-scale wastewater treatment plants
with biological nitrogen and phosphorus removal was investigated by FISH and
various oligonucleotide probes. The potential denitrifiers were characterized as
probe-defined populations that were able to consume radiolabelled substrate with
oxygen, nitrate and nitrite as electron acceptor as determined by microautoradio-
graphy. The most abundant potential denitrifiers were related to the genera
Aquaspirillum, Azoarcus, Thauera and Rhodocyclus, all within the Betaproteobacter-
ia. They made up 20–49% of all bacteria in most of the 17 nitrogen removal plants
investigated and were hardly present in four plants without denitrification. The
ecophysiology of Aquaspirillum, Azoarcus and Thauera-related bacteria was con-
sistent within each probe-defined group in the plants investigated. These three
groups showed distinct physiological differences, with the Aquaspirillum-related
bacteria appearing as the most specialized one, consuming only amino acids
among the substrates tested, and Thauera as the most versatile consuming some
volatile fatty acids, ethanol and amino acids. The coexistence of Aquaspirillum,
Azoarcus and Thauera-related bacteria in a range of treatment plants with
differences in wastewater, design and operation suggest that the populations
ensure a functional stability of the plants by occupying different ecological niches
related to the carbon transformation.
Introduction
Nitrogen removal from wastewater is usually performed by a
sequential nitrification-denitrification process in activated
sludge wastewater treatment plants (WWTP). Despite the
processes having been known, used and optimized for
decades, the most important nitrifying bacteria have only
recently been identified by culture-independent methods
(Juretschko et al., 1998; Daims et al., 2000), and still very
little is known about the identity of abundant denitrifiers
(Wagner & Loy, 2002; Wagner et al., 2002). The main reason
for this is that most studies of denitrifiers from activated
sludge systems have been carried out after isolation and
cultivation (e.g. Gorny et al., 1992; Scholten et al., 1999;
Gumaelius et al., 2001), and these isolates do not necessarily
represent the important denitrifiers actually present in the
WWTP (Wagner & Loy, 2002).
Some potential abundant denitrifiers have recently been
identified by using culture-independent methods in a
WWTP treating industrial wastewater (Wagner & Loy,
2002; Wagner et al., 2002) and in treatment plants treating
municipal wastewater (Thomsen et al., 2004). In the two
types of plants, the abundant denitrifiers were Azoarcus-
related bacteria and Aquaspirillum-related bacteria, respec-
tively. The abundance of these bacteria was evaluated using
FISH with specific oligonucleotide probes, and it was shown
that in some plants Aquaspirillum-related bacteria consti-
tuted 20–30% of all bacteria (Thomsen et al., 2004). Their
proposed denitrifying activity was shown by applying a
combination of microautoradiography (MAR) and FISH.
Also, bacteria belonging to the genera Thauera and Zoogloea
are assumed to be important denitrifiers in some WWTP,
as they are often present in WWTP with denitrification
(Juretschko et al., 2002; Rosello-Mora et al., 1995), and
because isolates are capable of performing full denitrifica-
tion (Harder, 1997; Foss & Harder, 1998; Scholten et al.,
1999; Mechichi et al., 2002). Their actual abundance and
importance in the denitrification process in full-scale plants
have, however, not yet been evaluated. Finally, polypho-
sphate-accumulating organisms (PAOs) have also been
suggested as potential denitrifiers in WWTP with combined
nitrogen (N) removal and enhanced biological phosphorus
removal (EBPR) (van Loosdrecht et al., 1997; Seviour et al.,
2003). Recently, it has been shown that uncultured Rhodo-
cyclus-related PAOs grown in lab-scale reactors were able to
denitrify (Zeng et al., 2003) and that in full-scale plants they
FEMS Microbiol Ecol 60 (2007) 370–382c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved

can also most likely perform denitrification (Kong et al.,
2004).
The physiology of these potentially important denitrifiers
is generally not very well known because most of them are
uncultured and because the isolates, which are the closest
relatives, are rather distantly related, making extrapolations
of the physiology difficult. Isolates of Azoarcus and Thauera
from activated sludge are able to grow on a large number of
organic and aromatic compounds or monoterpenes under
aerobic and denitrifying conditions (Anders et al., 1995;
Harder, 1997; Song et al., 1998, 2001; Scholten et al., 1999;
Mechichi et al., 2002). Isolates closely related to the newly
identified Aquaspirillum-related bacteria have not been
obtained so far, although they were very abundant in several
municipal treatment plants (Thomsen et al., 2004). Among
numerous substrates tested in situ by MAR, they could only
consume a mixture of amino acids and no common
substrate such as acetate, ethanol, or glucose. Thus, they
were suggested to be involved mainly in the degradation of
protein in the WWTP investigated (Thomsen et al., 2004).
The physiology of the Rhodocyclus-related PAOs is better
described than the other denitrifiers because intensive
studies in lab-scale reactors have been conducted (Seviour
et al., 2003). Largely, detailed studies of their actual ecophy-
siology in full-scale plants confirm the observations ob-
tained in lab-scale reactors (Kong et al., 2004).
The bacterial groups mentioned above all belong to the
Betaproteobacteria, which seem to include the main deni-
trifiers in activated sludge systems. Betaproteobacteria is an
abundant group in many WWTP (e.g. Juretschko et al.,
2002; Wagner et al., 2002; Klausen et al., 2004), and may
reflect that denitrification is a dominant process in these
WWTP. In this study, we have by quantitative FISH investi-
gated the abundance of potential denitrifiers targeted by
different oligonucleotide probes. Although the probes ap-
plied are not completely genus-specific, the targeted bacteria
will in this study be mentioned as Zoogloea (probe ZRA,
Rosello-Mora et al., 1995), Azoarcus (probe Azo644, Hess
et al., 1997), Thauera (probe Thau646, Lajoie et al., 2000),
Aquaspirillum (probe Aqs997, Thomsen et al., 2004) and
Rhodocyclus-related (probe PAOmix, Crocetti et al., 2000) in
several WWTP. Furthermore, we have studied various
aspects of the ecophysiology of these probe-defined groups
in order to see whether they have any physiological differ-
ences, which can be related to different ecological niches in
the WWTP investigated.
Materials and methods
Activated sludge sampling
The ecophysiology experiments were carried out with acti-
vated sludge from Aalborg East and Horsens WWTP,
Denmark. Aalborg East has carbon removal, nitrification,
denitrification, chemical and biological phosphorus re-
moval, and a mean cell residence time (sludge age) of
20–30 days. Horsens WWTP has carbon removal, nitrifica-
tion, denitrification, chemical phosphorus removal and a
sludge age of 20–25 days. The activated sludge samples were
collected from April until December 2004 from aeration
tanks and brought to the laboratory within 1–2 h.
FISH and probe specificity
The FISH procedure was performed on fixed sludge as de-
scribed by Amann (1995), and the following oligonucleotide
probes were used: EUB338, EUB338-II and EUB338-III, called
EUBmix (all Bacteria; Amann et al., 1990; Daims et al., 1999),
BET42a (Betaproteobacteria; Manz et al., 1992), BONE23a and
competitor (Betaone subgroup; Amann et al., 1996), Azo644
(targeting most Azoarcus; Hess et al., 1997), ZRA (Zoogloea
ramigera; Rosello-Mora et al., 1995), PAOmix consisting of
probe PAO462, PAO651 and PAO846 (Candidatus ‘Accumuli-
bacter phosphatis’, a polyphosphate-accumulating bacterium,
also called Rhodocyclus-related PAO; Crocetti et al., 2000),
OTU4-645 (targeting Alcaligenes latus and some activated
sludge clones; Juretschko et al., 2002) and OTU6-178 (targeting
Brachymonas denitrificans and some activated sludge clones;
Juretschko et al., 2002). Details about the probes can be found
in probeBase (Loy et al., 2003). In addition Thau646 (Thauera
spp.; Lajoie et al., 2000) and Aqs997, including competitors
(targeting Aquaspirillum-like sequences; Thomsen et al., 2004),
were used. Probe Aqs997 has a perfect match with the Aqua-
spirillum-like sequences found by Thomsen et al. (2004);
Aquaspirillum delicatum ATCC14667 (recently reclassified as
Curvibacter delicatus comb. nov.; Ding & Yokota, 2004),
Pseudomonas lanceolata AB021390 (recently reclassified as
Curvibacter lanceolatus comb. nov.; Ding & Yokota, 2004) and
other bacteria in the Curvibacter genus. Also, a few other
sequences of Comamonas sp. and clones related within the
family Comamonadaceae are targeted (some Rhodoferax, Acid-
ovorax and Comamonas). Probe Thau644 targets different
Thauera but also sequences in other genera within the Rhodo-
cyclales order and sequences within the Burkholderiales, Hydro-
genophilales and Nitrosomonadales orders. Many of these are
sequences of uncultured bacteria. Probe Azo644 targets most
bacteria in the Azoarcus genus but also a few uncultured
bacteria in other genera within the family Rhodocyclaceae (some
Rhodocyclus and Dechloromonas). PAOmix targets Candidatus
‘Accumulibacter phosphatis’ and other primarily uncultured
bacteria within the Rhodocyclales order. The cells were simulta-
neously hybridized with EUBmix, a probe targeting the Beta-
proteobacteria, as well as a more specific probe. In Fig. 1 the
specificity of probe ZRA, Azo644, Thau646, Aqs997 and
PAOmix is illustrated, showing representative isolates from the
various genera targeted by the probes. Azoarcus sp., AF011344 is
FEMS Microbiol Ecol 60 (2007) 370–382 c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
371Ecophysiology of denitrifying bacteria in activated sludge

targeted by both probe Thau646 and Azo644, which is illu-
strated with a � and an imprecise border between the two
probes in Fig. 1.
Oligonucleotides were 50-labelled with 5(6)-carboxyfluores-
cein-N-hydroxy-succinimide ester (FLUOS) or with sulphoin-
docyanine dyes (Cy5 and Cy3) (Thermo Hybaid, Germany).
For the study of the ecophysiology, the identity was always
linked to the function. FISH was combined either by MAR,
enzyme-labelled fluorescence (ELF), microspheres’ adhesion to
cells (MAC) or staining for storage products (PHA and
polyphosphate). This was conducted by dual staining, where
FISH was performed either before (PHA) or after (MAR, ELF,
MAC and polyphosphate) the different methods mentioned.
An automatic stage controller in the LSM 510 software allowed
saved positions on a cover glass to be relocated.
Autoradiography
MAR experiments were performed according to the proce-
dure described by Nielsen & Nielsen (2002a) with 2 mM
final concentration of each substrate, 10 mCi [3H]-labelled
substrate (propionate, formate, and pyruvate were 14C-
labelled) and 2 mL of diluted activated sludge (1 g SS L�1).
Under aerobic conditions, uptake of the following substrates
was investigated: formate, acetate, propionate, lactate, pyr-
uvate, oleic acid, ethanol, glucose, galactose, mannose,
glycine, leucine, aspartate, glutamate and thymidine (Amer-
sham, NEN/Perkin-Elmer, American radiolabelled chemi-
cals). Furthermore, an [3H]-labelled amino acid mixture
(ICN Biomedicals) was investigated with each of the 16
amino acids (alanine, arginine, aspartic acid, glutamic acid,
glycine, histidine, isoleucine, leucine, lysine, phenylalanine,
proline, serine, threonine, tyrosine and valine) in a concen-
tration of 0.1 mM each, and the specific activity varied from
6.8 to 28.8mCi mL�1. The uptake of acetate incubated with
unlabelled amino acid mixture and thymidine with unla-
belled acetate was also tested under aerobic conditions. The
potential use of electron acceptors other than oxygen was
investigated by studying the uptake of selected substrates
with nitrate (2 mM) or nitrite (0.5 mM) as electron acceptor
Aqs997
Azo644
ZRAZoogloea sp., DQ413151
Zoogloea ramigera, D14257
Azoarcus evansii, X77679
Azoarcus tolulyticus, AF229857
Azoarcus anaerobius, Y14701
Azoarcus sp., AF011344*
Thauera sp., AF110005
Thauera aromatica, AF170281
Thauera aromatica, AF229881
Thauera terpenica, AJ005817
Azonexus sp., AB166882
Dechloromonas sp., AF170356
Candidatus ”Accumulibacter phosphatis”, AY962316
uncultured Betaproteobacterium, AY062125
Leptothrix sp., AF385528Aquabacterium parvum, AF035052
Aquabacterium sp., AF089857
Comamonas sp., AF188304
Acidovorax sp., AY258065
Comamonas sp., AF501881
Aquaspi clone C, AY322153
Aquaspi clone B, AY322152
Aquaspi clone A, AY322151
Pseudomonas lanceolata (reclassified as Curvibacter lanceolata), AB021390
Aquaspirillum delicatum (reclassified as Curvibacter delicatum), AF078756
0.10
Thau646
PAOmix
Fig. 1. A phylogenetic tree illustrating the specificity of the applied gene probes Azo644, Thau646, ZRA, Aqs997 and PAOmix. Pure culture
representatives of the different genera targeted by the probes are shown. The tree was calculated using default settings for the maximum likelihood
algorithm in ARB. The scale bar corresponds to 10% estimated sequence divergence. �Azoarcus sp. (AF011344) is targeted by both probe Azo644 and
Thau646.
FEMS Microbiol Ecol 60 (2007) 370–382c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
372 T.R. Thomsen et al.

in the absence of oxygen or under anaerobic conditions
without nitrate/nitrite present. In experiments with nitrate
or nitrite as electron acceptor and in anaerobic experiments,
a pre-incubation step of 2 h with unlabelled organic sub-
strate (and nitrate/nitrite when tested as electron acceptor)
was included in order to ensure that substrate uptake could
be ascribed to direct growth or very large storage capabil-
ities. In some experiments the simultaneous uptake of two
substrates was investigated as described by Kong et al.
(2004). As a control for chemography, sludge was pasteur-
ized at 70 1C for 10 min just before the incubation. After 3 h
incubation, the activated sludge was fixed, and the FISH
procedure was performed before applying the photographic
emulsion (Lee et al., 1999).
Storage compounds
The lipidic storage granules of the different bacteria investi-
gated were studied by Nile Blue staining of polyhydroxyalk-
anoates (PHA) (Ostle & Holt, 1982). Polyphosphate
granules were stained with methylene blue (Neisser stain)
as described in Eikelboom & van Buijsen (1983). Fixed
sludge was spread out on gelatine-coated cover slides,
hybridized with specific oligonucleotide probes, and images
were taken. Subsequently, the slides were washed for 10 min
with 70% ethanol, and the Nile Blue or Neisser stain was
performed. Both stains were performed on sludge directly
from the WWTP. In addition, Nile Blue stain was performed
on sludge incubated for 3 h with acetate or amino acid
mixture under aerobic conditions with nitrate or nitrite as
electron acceptor, and under anaerobic conditions. Forma-
tion of intracellular PHA granules by probe Azo644,
Thau646 and Aqs997 probe-defined cells was then investi-
gated. Furthermore, selected substrates (alanine, arginine,
aspartic acid, glutamic acid, histidine, isoleucine, leucine,
lysine, phenylalanine, proline, serine, threonine, tyrosine
and valine) were incubated (2 mM) with sludge for 3 h
followed by dual staining with the Aqs997 oligonucleotide
probe and Nile Blue.
ELF and MAC
The presence of exo-enzyme activity was determined using
ELF (ELF-97s, Molecular Probes; www.probes.invitrogen.
com), where, after enzymatic cleavage, substrates form a
fluorescent precipitate on the surfaces of bacteria. The same
procedure as described by Nielsen et al. (2002) and Krage-
lund et al. (2005) was used. The following enzymes were
evaluated: ELFs 97 esterase (ELFs 97 acetate), ELFs 97
lipase (ELFs 97 palmitate), ELFs 97 b-D-galactosidase
(ELFs 97 b-D-galactopyranoside), ELFs 97 b-D-glucuroni-
dase (ELFs 97 b-D-glucuronide), ELFs 97 chitinase/N-
acetylglucosaminidase (ELFs 97 N-acetylglucosaminide;
ELFs 97 NAG) and an ELFs 97 Endogenous Phosphatase
detection kit.
The hydrophobicity of the bacteria was investigated by
MAC with a modified version of Zita & Hermansson, (1997)
and Nielsen et al. (2001) as described by Kragelund et al.
(2005).
Microscopy
A model LSM 510-Meta confocal laser scanning microscope
(Carl Zeiss, Oberkochen, Germany) was used for the detec-
tion of oligonucleotide probe positive cells combined with
various in situ methods. For quantitative FISH, 32 images
were taken of each of the five probe-defined populations in
all sludge samples investigated and the images were subse-
quently analysed using IMAGE ANALYSIS software (ImageJ
1.33 s, Rasband W, National Institutes of Health, USA,
http://rsb.info.nih.gov/ij/) with a designed macro. Numbers
are given as average � standard deviation. For ecophysio-
logical studies, a minimum of 20 microcolonies of Aqua-
spirillum, Azoarcus or Thauera-related bacteria were
investigated visually to estimate the percentage of cells
exhibiting a certain physiological property. Most experi-
ments concerning ecophysiology were performed with
sludge from both WWTP investigated (see Table 2).
Results
Abundance and morphology revealed by FISHprobing
Bacteria related to Aquaspirillum, Azoarcus, Thauera and
Rhodocyclus-related PAOs were present in most WWTP
investigated (Table 1). In the 16 plants treating mainly
domestic wastewater with biological nitrogen (N) and/or
phosphorus (P) removal, the predominant group was the
Aquaspirillum-related bacteria, which constituted 11–29%
of all bacteria detected with the EUBmix. Azoarcus and
Thauera were also abundant, representing 3–16% of the
biovolume. Rhodocyclus-related PAOs were present in EBPR
plants only, where they constituted 3–9%.
In treatment plants without denitrification, only very
small numbers of Thauera, Azoarcus, Aquaspirillum-related
bacteria and Rhodocyclus-related PAOs were observed. In
contrast, these groups of bacteria constituted the vast
majority of the Betaproteobacteria present in most munici-
pal denitrifying WWTP, where they constituted up to 49%
of the total biovolume (Table 1). Other potential denitri-
fiers, Zoogloea-related bacteria, were observed but only in
small numbers (Table 1).
The Aquaspirillum-like bacteria showed quite distinct
morphology, being relatively large, coccoid cells with a
diameter of 1–1.5 mm. The size of the microcolonies varied,
with the majority being 10–15 mm in diameter, but colonies
FEMS Microbiol Ecol 60 (2007) 370–382 c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
373Ecophysiology of denitrifying bacteria in activated sludge
of up to 40 mm in diameter were observed. In addition, the
specific probe also targeted some filamentous bacteria in
some WWTP, which have been investigated in detail in
another study (Thomsen et al., 2006). Thauera-related
bacteria made characteristic zoogloeal clusters of various
sizes (5–40mm). The cell size was typically 1.0–2.5 mm, and
the cell morphology was rod-like. Zoogloea-related bacteria
made characteristic clusters similar to the Thauera. Most
cells were rod-like (1–1.5 mm� 1 mm). Different Azoarcus
morphotypes were observed in most WWTP, usually as
rather large (1.0mm� 2.0 mm) or small (0.5mm� 1.0 mm)
rods growing either as single cells or in relatively small
microcolonies (5–20mm). Rhodocyclus-related bacteria were
coccobacilli with a diameter of 0.4–1.2 mm. Only very few
cells hybridizing with probe OUT4-645 (Alcaligenes latus)
and probe OTU6-178 (Brachymonas denitrificans) were
found.
Ecophysiology
Two WWTP, Aalborg East and Horsens, were studied in
detail to reveal possible physiological differences and simila-
rities among bacteria from the three gene probe-defined
groups investigated (Aquaspirillum, Azoarcus, Thauera). The
studies included substrate uptake profiles under different
electron acceptor conditions, presence of storage products
and exoenzymes, and presence of specific surface properties.
Rhodocyclus-related PAOs have previously been studied
(Kong et al., 2004) and Zoogloea–related organisms were
not abundant enough in the WWTP to be investigated.
The substrate uptake pattern, as determined by MAR, was
almost identical for each probe-defined population in the
two WWTP investigated. The pattern was, however, differ-
ent for the three populations investigated (Tables 2 and 3).
The Aquaspirillum-related bacteria were tested for uptake of
16 different substrates under aerobic, denitrifying and
anaerobic conditions (Thomsen et al., 2004; Tables 2 and 3).
Only the amino acid mixture was taken up by some, but not
all, microcolonies (20–30%). Substrate was taken up with
oxygen, nitrate or nitrite as electron acceptor, but not under
anaerobic conditions without the addition of external
electron acceptors, thus confirming their denitrifying po-
tential. Several amino acids were also tested as single
substrates, but no uptake was observed by MAR. All
substrates were taken up by other bacteria in the flocs
investigated, hence serving as positive controls. The uptake
of amino acids was also confirmed by PHA staining (Nile
blue) after incubation with the amino acid mixture for 3 h
(Table 4). Large PHA inclusions were observed in many of
the Aquaspirillum-related bacteria. These cells clearly con-
tained more PHA than cells in fresh sludge taken directly
from the WWTP, where Aquaspirillum-related bacteria only
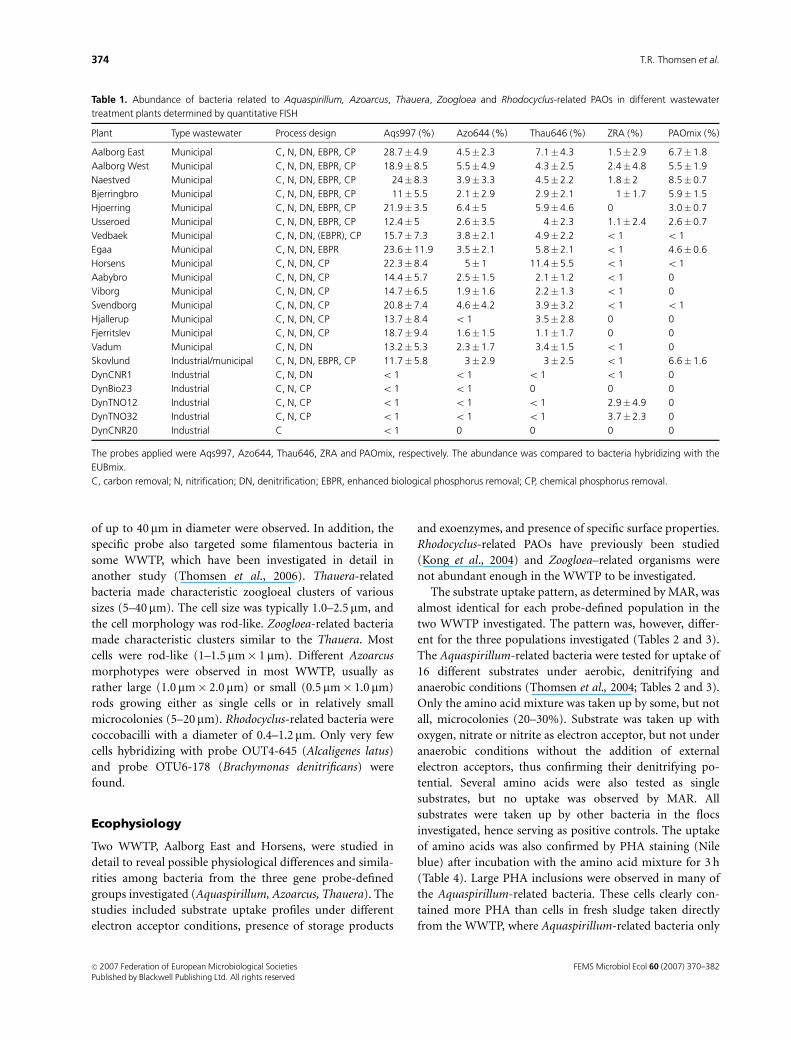
Table 1. Abundance of bacteria related to Aquaspirillum, Azoarcus, Thauera, Zoogloea and Rhodocyclus-related PAOs in different wastewater
treatment plants determined by quantitative FISH
Plant Type wastewater Process design Aqs997 (%) Azo644 (%) Thau646 (%) ZRA (%) PAOmix (%)
Aalborg East Municipal C, N, DN, EBPR, CP 28.7� 4.9 4.5�2.3 7.1� 4.3 1.5� 2.9 6.7�1.8
Aalborg West Municipal C, N, DN, EBPR, CP 18.9� 8.5 5.5�4.9 4.3� 2.5 2.4� 4.8 5.5�1.9
Naestved Municipal C, N, DN, EBPR, CP 24� 8.3 3.9�3.3 4.5� 2.2 1.8� 2 8.5�0.7
Bjerringbro Municipal C, N, DN, EBPR, CP 11� 5.5 2.1�2.9 2.9� 2.1 1� 1.7 5.9�1.5
Hjoerring Municipal C, N, DN, EBPR, CP 21.9� 3.5 6.4�5 5.9� 4.6 0 3.0�0.7
Usseroed Municipal C, N, DN, EBPR, CP 12.4� 5 2.6�3.5 4� 2.3 1.1� 2.4 2.6�0.7
Vedbaek Municipal C, N, DN, (EBPR), CP 15.7� 7.3 3.8�2.1 4.9� 2.2 o 1 o 1
Egaa Municipal C, N, DN, EBPR 23.6� 11.9 3.5�2.1 5.8� 2.1 o 1 4.6�0.6
Horsens Municipal C, N, DN, CP 22.3� 8.4 5�1 11.4� 5.5 o 1 o 1
Aabybro Municipal C, N, DN, CP 14.4� 5.7 2.5�1.5 2.1� 1.2 o 1 0
Viborg Municipal C, N, DN, CP 14.7� 6.5 1.9�1.6 2.2� 1.3 o 1 0
Svendborg Municipal C, N, DN, CP 20.8� 7.4 4.6�4.2 3.9� 3.2 o 1 o 1
Hjallerup Municipal C, N, DN, CP 13.7� 8.4 o 1 3.5� 2.8 0 0
Fjerritslev Municipal C, N, DN, CP 18.7� 9.4 1.6�1.5 1.1� 1.7 0 0
Vadum Municipal C, N, DN 13.2� 5.3 2.3�1.7 3.4� 1.5 o 1 0
Skovlund Industrial/municipal C, N, DN, EBPR, CP 11.7� 5.8 3�2.9 3� 2.5 o 1 6.6�1.6
DynCNR1 Industrial C, N, DN o 1 o 1 o 1 o 1 0
DynBio23 Industrial C, N, CP o 1 o 1 0 0 0
DynTNO12 Industrial C, N, CP o 1 o 1 o 1 2.9� 4.9 0
DynTNO32 Industrial C, N, CP o 1 o 1 o 1 3.7� 2.3 0
DynCNR20 Industrial C o 1 0 0 0 0
The probes applied were Aqs997, Azo644, Thau646, ZRA and PAOmix, respectively. The abundance was compared to bacteria hybridizing with the
EUBmix.
C, carbon removal; N, nitrification; DN, denitrification; EBPR, enhanced biological phosphorus removal; CP, chemical phosphorus removal.
FEMS Microbiol Ecol 60 (2007) 370–382c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
374 T.R. Thomsen et al.
occasionally contained a few PHA granules. Subsequently,
the potential of utilizing one or more of the specific amino
acids present in the mixture was tested by incubating
activated sludge for 3 h with single amino acids followed by
dual staining with oligonucleotide probes and Nile Blue.
Only incubations with phenylalanine resulted in large
intracellular inclusions of PHA in many Aquaspirillum-
related bacteria, strongly indicating that this compound
was a suitable sole substrate. Aquaspirillum-related bacteria
in sludge from Aalborg East did not contain polyphosphate,
but a few did in Horsens WWTP. Surface-associated exoen-
zymatic activity was only exhibited by a few Aquaspirillum-
related bacteria as detected by the applied ELF-substrates.
A few galactosidase-positive cells were found in sludge from
Aalborg East WWTP, while none were found in the two
plants with esterase, lipase, chitinase, glucoronidase or
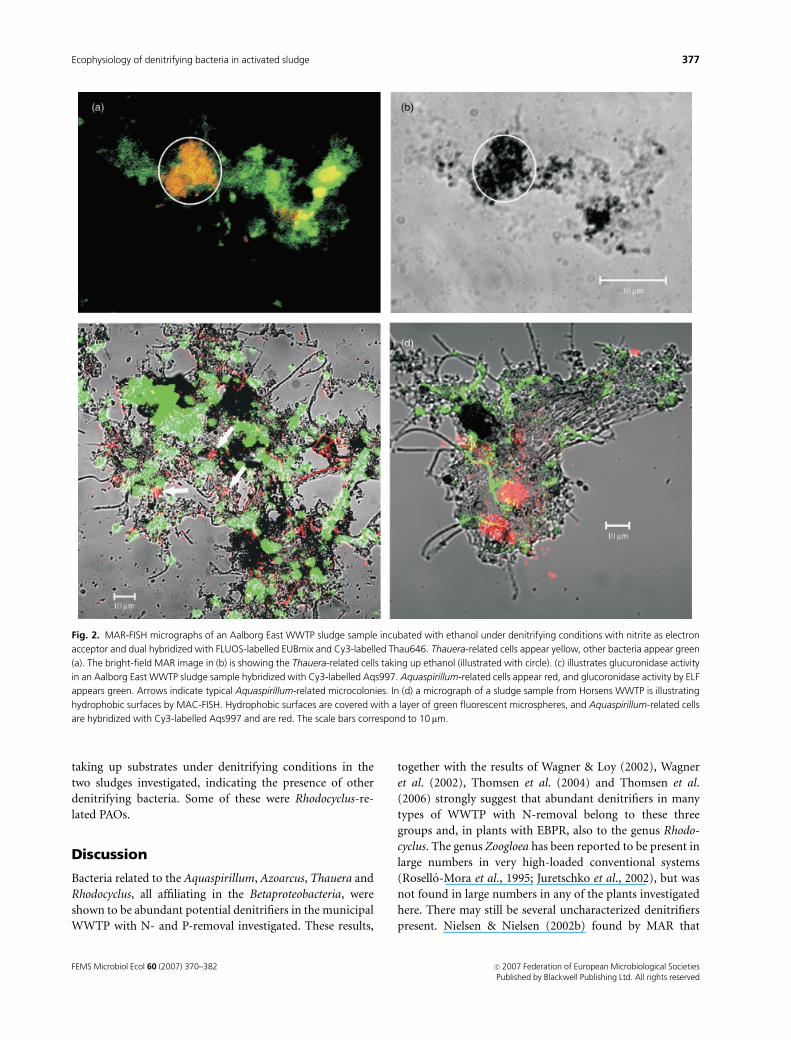
phosphatase activity (Fig. 2c). When the surface properties
were investigated using MAC, both hydrophobic and hydro-
philic structures were found in the flocs as indicated by
many and few attached fluorescent hydrophobic micro-
spheres, respectively. No hydrophobic microspheres were
adsorbed on the Aquaspirillum-related bacteria in any of
the two sludges, indicating a rather hydrophilic surface
(Fig. 2d).
The probe-defined Azoarcus bacteria were only able to
take up a limited number of substrates with oxygen as
electron acceptor (Table 2). Acetate, ethanol and the amino
acid mixture were taken up by 20–60% of the bacteria, while
fewer took up pyruvate. No uptake of formate, propionate
or lactate was observed. Leucine could only be taken up
when acetate was added as a cosubstrate. The same sub-
strates were generally taken up with nitrate or nitrite as
electron acceptor, although the active fraction of probe-
defined bacteria was lower (Table 3), indicating that some of
them may not be able to perform full denitrification. No
Azoarcus bacteria were able to take up substrate under
anaerobic conditions, all were Neisser-negative (no poly-
phosphate granules), and they did not contain PHA gran-
ules in samples taken directly from the treatment plants.
Only a few Azoarcus cells containing PHA granules were
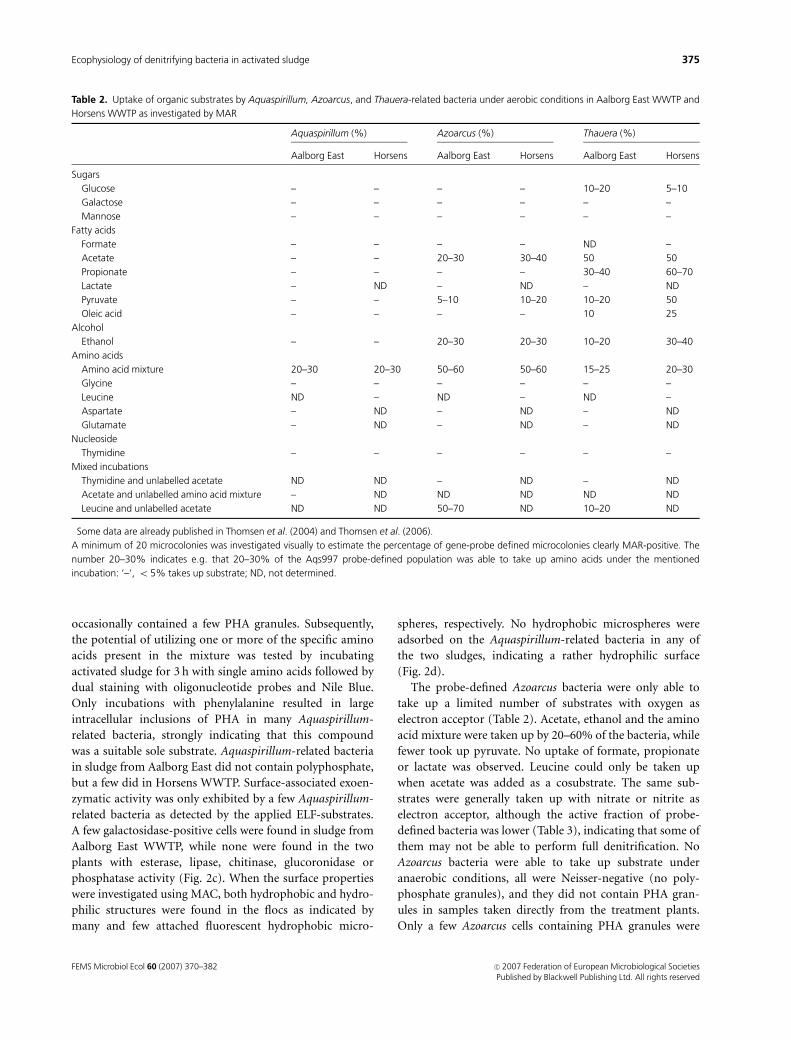
Table 2. Uptake of organic substrates by Aquaspirillum, Azoarcus, and Thauera-related bacteria under aerobic conditions in Aalborg East WWTP and
Horsens WWTP as investigated by MAR
Aquaspirillum (%)� Azoarcus (%) Thauera (%)
Aalborg East Horsens Aalborg East Horsens Aalborg East Horsens
Sugars
Glucose – – – – 10–20 5–10
Galactose – – – – – –
Mannose – – – – – –
Fatty acids
Formate – – – – ND –
Acetate – – 20–30 30–40 50 50
Propionate – – – – 30–40 60–70
Lactate – ND – ND – ND
Pyruvate – – 5–10 10–20 10–20 50
Oleic acid – – – – 10 25
Alcohol
Ethanol – – 20–30 20–30 10–20 30–40
Amino acids
Amino acid mixture 20–30 20–30 50–60 50–60 15–25 20–30
Glycine – – – – – –
Leucine ND – ND – ND –
Aspartate – ND – ND – ND
Glutamate – ND – ND – ND
Nucleoside
Thymidine – – – – – –
Mixed incubations
Thymidine and unlabelled acetate ND ND – ND – ND
Acetate and unlabelled amino acid mixture – ND ND ND ND ND
Leucine and unlabelled acetate ND ND 50–70 ND 10–20 ND
�Some data are already published in Thomsen et al. (2004) and Thomsen et al. (2006).
A minimum of 20 microcolonies was investigated visually to estimate the percentage of gene-probe defined microcolonies clearly MAR-positive. The
number 20–30% indicates e.g. that 20–30% of the Aqs997 probe-defined population was able to take up amino acids under the mentioned
incubation: ‘–’, o 5% takes up substrate; ND, not determined.
FEMS Microbiol Ecol 60 (2007) 370–382 c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
375Ecophysiology of denitrifying bacteria in activated sludge
observed after 3-hour incubations with amino acid mixture
(Table 4). Esterase activity was observed for some Azoarcus
bacteria in sludge from Aalborg East and Horsens, whereas
no other exoenzymes were detected. All Azoarcus investi-
gated had hydrophilic surfaces.
Probe-defined Thauera bacteria were versatile in their
uptake of organic substrates under aerobic conditions as
they were able to use glucose, acetate, propionate, pyruvate,
oleic acid, ethanol and the amino acid mixture (Table 2, Fig.
2a and b). Leucine could only be taken up when acetate was
added as a cosubstrate. However, for several substrates, only
a fraction of the bacteria was active, ranging from 5–10% for
glucose to 60–70% for propionate. Nitrate or nitrite could
be used as electron acceptor for uptake of most substrates,
except glucose and oleic acid. For propionate, an identical
fraction of Thauera was active with oxygen and nitrate as
electron acceptor, while only less than half of them were
active with nitrite as electron acceptor, indicating that some
of them were unable to perform full denitrification. Under
anaerobic conditions, pyruvate was consumed by a few of
the Thauera in Horsens WWTP. Thauera had only a small
amount of polyphosphate granules in a few cells, but
contained some PHA granules in samples taken directly
from the WWTP. However, a much higher PHA storage
potential was observed when the sludge was incubated with
amino acid mixture or acetate for 3 h (Table 4). Surface-
associated esterase and glucoronidase activity was observed
for only very few Thauera in sludge from Aalborg East.
All microcolonies belonging to Thauera had hydrophilic
surfaces.
Other bacteria than the probe-defined Aquaspirillum-,
Azoarcus- and Thauera-related bacteria were observed
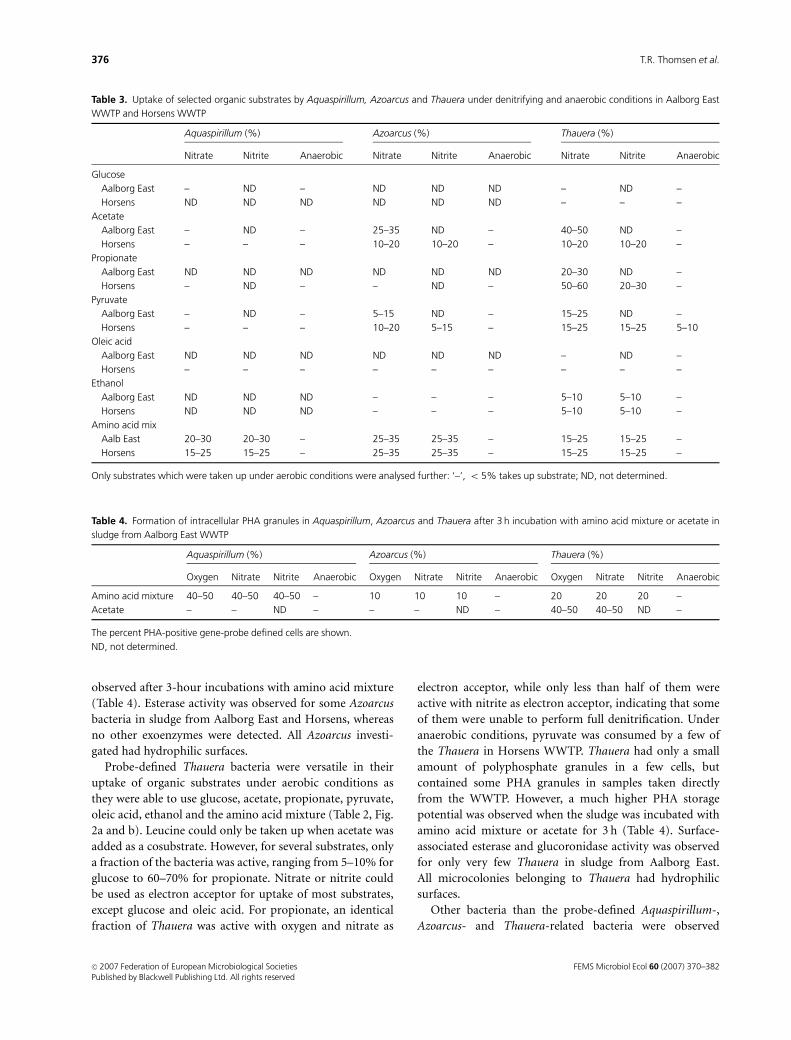
Table 4. Formation of intracellular PHA granules in Aquaspirillum, Azoarcus and Thauera after 3 h incubation with amino acid mixture or acetate in
sludge from Aalborg East WWTP
Aquaspirillum (%) Azoarcus (%) Thauera (%)
Oxygen Nitrate Nitrite Anaerobic Oxygen Nitrate Nitrite Anaerobic Oxygen Nitrate Nitrite Anaerobic
Amino acid mixture 40–50 40–50 40–50 – 10 10 10 – 20 20 20 –
Acetate – – ND – – – ND – 40–50 40–50 ND –
The percent PHA-positive gene-probe defined cells are shown.
ND, not determined.
Table 3. Uptake of selected organic substrates by Aquaspirillum, Azoarcus and Thauera under denitrifying and anaerobic conditions in Aalborg East
WWTP and Horsens WWTP
Aquaspirillum (%) Azoarcus (%) Thauera (%)
Nitrate Nitrite Anaerobic Nitrate Nitrite Anaerobic Nitrate Nitrite Anaerobic
Glucose
Aalborg East – ND – ND ND ND – ND –
Horsens ND ND ND ND ND ND – – –
Acetate
Aalborg East – ND – 25–35 ND – 40–50 ND –
Horsens – – – 10–20 10–20 – 10–20 10–20 –
Propionate
Aalborg East ND ND ND ND ND ND 20–30 ND –
Horsens – ND – – ND – 50–60 20–30 –
Pyruvate
Aalborg East – ND – 5–15 ND – 15–25 ND –
Horsens – – – 10–20 5–15 – 15–25 15–25 5–10
Oleic acid
Aalborg East ND ND ND ND ND ND – ND –
Horsens – – – – – – – – –
Ethanol
Aalborg East ND ND ND – – – 5–10 5–10 –
Horsens ND ND ND – – – 5–10 5–10 –
Amino acid mix
Aalb East 20–30 20–30 – 25–35 25–35 – 15–25 15–25 –
Horsens 15–25 15–25 – 25–35 25–35 – 15–25 15–25 –
Only substrates which were taken up under aerobic conditions were analysed further: ‘–’, o 5% takes up substrate; ND, not determined.
FEMS Microbiol Ecol 60 (2007) 370–382c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
376 T.R. Thomsen et al.
taking up substrates under denitrifying conditions in the
two sludges investigated, indicating the presence of other
denitrifying bacteria. Some of these were Rhodocyclus-re-
lated PAOs.
Discussion
Bacteria related to the Aquaspirillum, Azoarcus, Thauera and
Rhodocyclus, all affiliating in the Betaproteobacteria, were
shown to be abundant potential denitrifiers in the municipal
WWTP with N- and P-removal investigated. These results,
together with the results of Wagner & Loy (2002), Wagner
et al. (2002), Thomsen et al. (2004) and Thomsen et al.
(2006) strongly suggest that abundant denitrifiers in many
types of WWTP with N-removal belong to these three
groups and, in plants with EBPR, also to the genus Rhodo-
cyclus. The genus Zoogloea has been reported to be present in
large numbers in very high-loaded conventional systems
(Rosello-Mora et al., 1995; Juretschko et al., 2002), but was
not found in large numbers in any of the plants investigated
here. There may still be several uncharacterized denitrifiers
present. Nielsen & Nielsen (2002b) found by MAR that
Fig. 2. MAR-FISH micrographs of an Aalborg East WWTP sludge sample incubated with ethanol under denitrifying conditions with nitrite as electron
acceptor and dual hybridized with FLUOS-labelled EUBmix and Cy3-labelled Thau646. Thauera-related cells appear yellow, other bacteria appear green
(a). The bright-field MAR image in (b) is showing the Thauera-related cells taking up ethanol (illustrated with circle). (c) illustrates glucuronidase activity
in an Aalborg East WWTP sludge sample hybridized with Cy3-labelled Aqs997. Aquaspirillum-related cells appear red, and glucoronidase activity by ELF
appears green. Arrows indicate typical Aquaspirillum-related microcolonies. In (d) a micrograph of a sludge sample from Horsens WWTP is illustrating
hydrophobic surfaces by MAC-FISH. Hydrophobic surfaces are covered with a layer of green fluorescent microspheres, and Aquaspirillum-related cells
are hybridized with Cy3-labelled Aqs997 and are red. The scale bars correspond to 10 mm.
FEMS Microbiol Ecol 60 (2007) 370–382 c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
377Ecophysiology of denitrifying bacteria in activated sludge

about 70% of the biomass in Aalborg East WWTP is likely to
denitrify, which is in accordance with observations in this
study that many, but not all, nitrate and nitrite-respiring
organisms have been identified.
The denitrifying capacity of the probe-defined popula-
tions was shown by MAR-FISH in the study. It is difficult to
link the denitrifying activity to specific species in environ-
mental samples (Philippot & Hallin, 2005), so their poten-
tial denitrifying activities were here defined as being able to
take up organic substrates in the presence of nitrate as well
as nitrite as sole electron acceptor. Furthermore, they were
unable to consume any substrates under strictly anaerobic
conditions without nitrite or nitrate present. The fraction of
bacteria able to use nitrite was identical for Aquaspirillum-
related bacteria and Azoarcus to the fraction using nitrate as
electron acceptor, suggesting a full denitrification. This was
also the case for Thauera except with propionate as electron
acceptor, where only half of them were able to use nitrite
(Table 3). This may indicate a partial denitrification (to
nitrite) due to the presence of several subpopulations as
discussed below.
Ecophysiology of potential denitrifiers
Some interesting physiological differences between the three
groups were observed. The results showed, in agreement
with previous studies (Thomsen et al. 2004, 2006), that
Aquaspirillum-related bacteria apparently are very specia-
lized, only consuming very few substances. Surprisingly,
none of the substrates assumed to be commonly present in
activated sludge systems, such as acetate, propionate and
lactate (Nielsen et al., 1992; Lie & Welander, 1997; Henze
et al., 2002), could be consumed. Bacteria related to
Azoarcus were slightly more versatile and able to consume
acetate and a few other substrates, but not propionate or
glucose. Bacteria related to Thauera were the most versatile
ones and able to consume most substrates tested. It is,
however, remarkable that none of the groups studied were
able to assimilate glycine, leucine or formate, which are
likely to be present in many types of wastewater as hydrolysis
or fermentation products from protein and other macro-
molecules. However, leucine could be taken up when acetate
was added as a co-substrate, as has also been observed by the
Rhodocyclus-related PAOs (Kong et al., 2004). It is unclear
whether the uptake of leucine under these conditions is
mainly as an N or C-source. The overall results suggest that
acetate and some amino acids potentially are key com-
pounds for the removal of nitrate in many WWTP.
Rhodocyclus-related PAOs were present in the EBPR
plants, constituting 3–9% of the total biovolume. Some of
these PAOs are also able to denitrify (Kong et al., 2004) and
may constitute a significant part of the denitrifiers. Their
substrate uptake profile with oxygen, nitrate or nitrite as
electron acceptor in Aalborg East WWTP and two other
plants is slightly more diverse than that of Azoarcus, but less
than that of Thauera; there is uptake of acetate, propionate
and pyruvate, but not glycine, leucine, ethanol or oleic acid
(Kong et al., 2004). In contrast to the other denitrifiers they
are also able to take up substrate under anaerobic condi-
tions, store it as PHA and subsequently take up orthopho-
sphate under denitrifying and aerobic conditions for storage
of polyphosphate.
In common for all probe-defined populations was the
observation that a fraction of the bacteria appeared inactive
and did not take up labelled substrate. Some FISH-positive
cells may have been viable but not active, or, as the three
probes applied in this study are not very specific and
hybridize with several species within the genera described
(see ‘Material and methods’), variations in physiology may
be due to an undescribed microdiversity covered by the
probes. For Azoarcus and Thauera, it seems most plausible
that several subpopulations with different substrate uptake
profiles were present, as different morphologies existed.
However, a clone library study of Aalborg East sludge
showed a rather low diversity of bacteria targeted by probe
Thau644, indicating the probe seems not to target many
unspecific bacteria in this specific sludge (data not shown).
For the microcolony-forming Aquaspirillum-related bacter-
ia, which all had a very characteristic morphology, 30–50%
were metabolically active (in MAR and PHA experiments),
indicating that a certain fraction was inactive or that
morphologically identical subpopulations were present.
Alternatively, we have not yet found the primary substrates,
especially for the Aquaspirillum-related bacteria, but possi-
bly also for subpopulations of the two other groups. Some of
these might uptake substrates not tested in this study, e.g.
aromatic compounds and monoterpenes, which are usually
not present in high concentration in influent to most
WWTP (Henze et al., 2002). However, we also observed that
a certain fraction of probe-defined filamentous Alphapro-
teobacteria had a very low activity in a WWTP (Nielsen
et al., 2003) and that 10–50% of denitrifying Rhodocyclus-
related PAOs were inactive in different EBPR plants (Kong
et al., 2004). It was suggested that the operation of the
treatment plants caused inactivity for a part of the bacteria
due to poor growth conditions, but whether this explana-
tion is also likely for the denitrifiers investigated here is
unknown.
Intracellular storage of PHA is a well-known feature of
several activated sludge bacteria (Seviour & Blackall, 1999),
but PHA was hardly present in any of the probe-defined
bacteria studied here in fresh sludge. Rhodocyclus-related
PAOs usually have it when sampled from the anaerobic
tanks (Kong et al., 2004). However, Aquaspirillum and
Thauera had a high PHA storage potential, as shown when
they were incubated with suitable substrates, so under the
FEMS Microbiol Ecol 60 (2007) 370–382c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
378 T.R. Thomsen et al.

dynamic conditions that occur in most full-scale N-removal
plants this metabolic feature can be a competitive advantage.
Bacteria from all three groups besides the PAOs only showed
storage capacity when incubated under aerobic and deni-
trifying conditions and not under anaerobic conditions,
which correlates well with the MAR results. Furthermore,
the three groups had no or very few Neisser-positive
granules, indicating an absence of large amounts of poly-
phosphate, thus confirming that they were not PAOs.
The presence of specific surface-associated exoenzymes
on different probe-defined bacteria has been observed in
activated sludge flocs (Kloeke & Geesey, 1999; Nielsen et al.,
2002). Such enzymes were only detected to a very small
extent in the three groups investigated. This indicates that all
three groups were mainly involved in the consumption of
soluble, easily consumable compounds and not in the
hydrolysis of macromolecules.
The microcolonies of Aquaspirillum, Azoarcus and
Thauera all had hydrophilic surfaces. Other bacteria in the
floc had hydrophobic surfaces, revealing some heterogeneity
in accordance with earlier studies. Cell surface properties are
important to the floc strength and stability (Zita & Her-
mansson, 1997; Olofsson et al., 1998; Nielsen et al., 2004),
and they can be important to the accessibility of substrates
to the cells (Nielsen et al., 2002). There was no detectable
difference between the surface properties of the three groups
investigated. It is known that Thauera can form gels with a
high content of water (Lajoie et al., 2000), but it was not
further investigated in this study.
Comparison with physiological data from purecultures
A number of isolates of Azoarcus and Thauera are available.
Most of them are not isolated from activated sludge, so their
identity, and thereby their physiology, may be different from
those in the sludges investigated. Azoarcus isolates can grow
on many aromatic compounds, some amino acids such as
alanine, on acetate, lactate, pyruvate, methanol and ethanol
(Anders et al., 1995; Song et al., 2001; Mechichi et al., 2002).
This generally corresponds well to the findings in this study,
although lactate could not be used. Furthermore, several
isolates of Azoarcus can denitrify with nitrate and nitrite,
and they can accumulate and grow on PHA (Mechichi
et al., 2002). There are no reports of the presence of
polyphosphate.
Recently, Ginige et al. (2005) showed that Thauera and
other members of the families Comamonadaceae and Rho-
docyclaceae are acetate-utilizing denitrifiers in activated
sludge by combining stable isotope probing and a full-cycle
rRNA analysis, and it corresponds well with the data
obtained in this study. Isolates of Thauera use nitrate, nitrite
or oxygen as electron acceptor, and many utilize aromatic
compounds, monoterpenes, amino acids, organic substrates
such as sugars, acetate, lactate, pyruvate and ethanol (Foss &
Harder, 1998; Song et al., 1998, 2001; Mechichi et al., 2002).
The relatively high versatility is in accordance with the data
obtained in this study. Interestingly, however, lactate was not
taken up in situ, although isolates can grow on this substrate.
This indicates the presence of several uncultured subpopu-
lations of this genus, or it illustrates differences between
bacteria growing in pure culture and in situ. Some isolates of
Thauera accumulate PHA (Anders et al., 1995; Scholten
et al., 1999), which was also observed in this study. T.
linealoolentis and T. terpenica are able to use sugar com-
pounds under aerobic conditions, and in accordance with
data obtained in our study these isolates are not able to grow
on sugars under denitrifying conditions (Foss & Harder,
1998). Such a restriction of the range of substrate to be
consumed with nitrate or nitrite as electron acceptor instead
of oxygen was also recently observed in situ by some
filamentous Alphaproteobacteria in activated sludge (Krage-
lund et al., 2005).
Recently 199 denitrifying bacteria were isolated from
activated sludge, where the majority belonged to the Beta-
proteobacteria (Heylen et al., 2006). Some Thauera, one
Zoogloea and one Aquaspirillum-related bacterium were
isolated. The isolated Aquaspirillum bacterium was distantly
related to the Aquaspirillum-related sequences used for the
design of probe Aqs997 (Thomsen et al., 2004), and the
probe does not target the isolate. No other pure culture of
the Aquaspirillum-related bacteria exists, so this study pre-
sents the first more detailed study of the physiology of these
organisms. The closest isolated relatives are Aquaspirillum
delicatum and Pseudomonas lanceolata, which grow on a
variety of substrates under aerobic conditions, but are
unable to perform full denitrification (the strains can only
make a partial reduction of nitrate to nitrite; Leifson, 1962;
Krieg, 1984; Pot et al., 1991). This physiology is very
different from that found in this in situ study of the
Aquaspirillum-related bacteria, where only uptake of pheny-
lalanine and perhaps a few other amino acids were observed
and they most likely carried out full denitrification.
Implications for full-scale treatment plant
It was shown that the abundant denitrifiers in a wide range
of WWPT with biological N and P-removal belonged to
relatively few genera; Aquaspirillum, Azoarcus, Thauera and
Rhodocyclus. Also, relatively few genera are dominant for
other groups of functional importance in full-scale activated
sludge systems, e.g. for the ammonium oxidizers (Purkhold
et al. 2000) and the PAO (Kong et al., 2005). It is assumed
that a large diversity ensures a better functional stability and
robustness against changes, leading to a more stable treat-
ment plant. The species richness of nitrifiers, for example,
FEMS Microbiol Ecol 60 (2007) 370–382 c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
379Ecophysiology of denitrifying bacteria in activated sludge

varies in different WWTP, where, in some cases, it is
dominated by one single species, in others by at least four
different species (Purkhold et al. 2000). Factors such as
differences in substrate affinity and growth rates may con-
trol their presence. Among PAOs typically three (or more, so
far unknown) groups are usually present and differences in
substrate utilization capabilities seems to determine their
presence (Kong et al., 2005). It seems also to be the case for
the groups of denitrifiers investigated in this study. The
physiology was rather consistent within each probe-defined
group, and each group showed distinct differences, with the
Aquaspirillum–related bacteria as the most specialized and
Thauera as the most versatile (generalist). These differences
indicate that they occupy different nutritional niches related
to the carbon flow in the WWTP although differences in
storage capabilities (PHA and polyphosphate) and maybe
also in other kinetic parameters were present. Usually, the
heterogeneity in space and/or time can explain the degree of
diversity in an ecosystem (Kassen & Rainey, 2004). The
heterogeneity in space (within the floc) is probably limited,
but the dynamic nature of substrate supply, aerobic and
anaerobic phases, and a high degree of physiological differ-
ences supported the presence of four to five coexisting
denitrifying groups with an unknown microdiversity in
most WWTP investigated. Most likely several other species
of denitrifiers were present in low numbers in the WWTP
investigated indicating that the total diversity was large, but
it seems that the diversity of the functional abundant species
was much smaller.
Besides being important to the denitrification process, the
dominating species are, simply due to their high abundance
(up to 49% of all biovolume), also important to floc
formation, floc properties and thus solid–liquid separation.
Thauera and Zoogloea, for example, are known to produce
high amounts of water-containing extracellular polymeric
substances (EPS) that can cause problems with sludge
compaction and dewatering (Easson et al., 1987; Lajoie
et al., 2000). The Aquaspirillum–related bacteria, on the
other hand, seem not to be related to poor floc properties
(Thomsen & Nielsen, unpublished results). Therefore, better
knowledge about the ecophysiology of the different species
might help to control excessive growth of Thauera and
Zoogloea to ensure presence of denitrifiers with good floc
properties.
Acknowledgements
The Danish Technical Research Council supported this
study under the framework program ‘Activity and Diversity
in Complex Microbial Systems’. We thank M. Stevenson,
M. Fredsgaard and S. Bielidt for their valuable technical
assistance. S. Fuereder from Vienna University is greatly
acknowledged for investigating targets of gene probes used
in this study on a clone library on Aalborg East sludge.
References
Amann RI (1995) In situ identification of microorganisms by
whole cell hybridization with rRNA- targeted nucleic acid
probes. Molecular Microbial Ecological Manual (Akkermans
ADL, van Elsas JD & de Bruijn FJ, eds), pp. 1–15. Kluwer
Academic Publications, London, UK.
Amann RI, Binder BJ, Olson RJ, Chisholm SW, Devereux R &
Stahl D (1990) Combination of 16S rRNA-targeted
oligonucleotide probes with flow cytometry for analyzing
mixed microbial populations. Appl Environ Microbiol 56:
1919–1925.
Amann R, Snaidr J, Wagner M, Ludwig W & Schleifer K-H (1996)
In situ visualization of high genetic diversity in a natural
microbial community. J Bacteriol 178: 3496–3599.
Anders H, Kaetzke A, Kampfer P, Ludwig W & Fuchs G (1995)
Taxonomic position of aromatic-degrading denitrifying
pseudomonad strains K 172 and KB 740 and their description
as new members of the genera Thauera, as Thauera aromatica
sp. nov., and Azoarcus, as Azoarcus evansii sp. nov.,
respectively, members of the beta subclass of the
Proteobacteria. Int J Syst Bacteriol 45: 327–333.
Crocetti GR, Hugenholtz P, Bond PL, Schuler A, Keller J, Jenkins
D & Blackall LL (2000) Identification of polyphospate-
accumulating organisms and design of 16S rRNA-directed
probes for their detection and quantification. Appl Environ
Microbiol 66: 1175–1182.
Daims H, Bruhl A, Amann R, Schleifer K-H & Wagner M (1999)
The domain-specific probe EUB338 is insufficient for the
detection of all Bacteria: development and evaluation of a
more comprehensive probe set. Syst Appl Microbiol 22:
434–444.
Daims H, Nielsen PH, Nielsen JL, Juretschko S & Wagner M
(2000) Novel Nitrosospira-like bacteria as dominant nitrite-
oxidizers in biofilms from wastewater treatment plants:
diversity and in situ physiology. Wat Sci Technol 41: 85–90.
Ding L & Yokota A (2004) Proposals of Curvibacter gracilis gen.
nov., sp. nov. and Herbaspirillum putei sp. nov. for bacterial
strains isolated from well water and reclassification of
[Pseudomonas] huttiensis, [Pseudomonas] lanceolata,
[Aquaspirillum] delicatum and [Aquaspirillum] autotrophicum
as Herbaspirillum huttiense comb. nov., Curvibacter lanceolatus
comb. nov., Curvibacter delicatus comb. nov. and
Herbaspirillum autotrophicum comb. nov. Int J Syst Evol
Microbiol 54: 2223–2230.
Easson DD, Peoples OP & Sinskey AJ (1987). Biopolymer
engineering: genetic control of exopolysaccharide biosynthesis.
Industrial Polysaccharides: Genetic Engineering, Structure/
Property Relations and Applications: 57–64.
Eikelboom DH & van Buijsen HJJ (1983) Microscopic Sludge
Investigation Manual, 2nd edn. TNO Research Institute for
Environmntal Hygiejne, Delft.
FEMS Microbiol Ecol 60 (2007) 370–382c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
380 T.R. Thomsen et al.

Foss S & Harder J (1998) Thauera linaloolentis sp. nov. and
Thauera terpenica sp. nov., isolated on oxygen-containing
monoterpenes (linealool, menthol, and eucalyptol) and
nitrate. Syst Appl Microbiol 21: 365–373.
Ginige MP, Keller J & Blackall LL (2005) Investigation of an
acetate-fed denitrifying microbial community by stable
isotope probing, full-cycle rRNA analysis, and fluorescent in
situ hybridization-microautoradiography. Appl Environ
Microbiol 71: 8683–8691.
Gorny N, Wahl G, Brune A & Schink B (1992) A strictly anaerobic
nitrate-reducing bacterium growing with resorcinol and other
aromatic compounds. Arch Microbiol 158: 48–53.
Gumaelius L, Magnusson G, Pettersson B & Dalhammer G (2001)
Comamonas denitrificans sp. nov., an efficient denitrifying
bacterium isolated from activated sludge. Int J Syst Evol
Microbiol 51: 999–1006.
Harder J (1997) Anaerobic degradation of cyclohexane-1,2-diol
by a new Azoarcus species. Archives Microbiol 168: 199–204.
Henze M, Harremoes P, Jansen JlC & Arvin E (2002) Wastewater
Treatment. Biological and Chemical Processes, 3rd ed. Springer,
Berlin, Germany.
Hess A, Zarda B, Hahn D, Haner A, Stax D, Hohener P & Zeyer J
(1997) In situ analysis of denitrifying toluene- and m-xylen-
degrading bacteria in a diesel fuel-contaminated laboratory
aquifer column. Appl Environ Microbiol 63: 2136–2141.
Heylen K, Vanparys B, Wittebolle L, Verstraete W, Boon N & De
Vos P (2006) Cultivation of denitrifying bacteria: optimization
of isolation conditions and diversity study. Appl Environ
Microbiol 72: 2637–2643.
Juretschko S, Timmermann G, Schmid M, Schleifer K-H,
Pommerening-Roser A, Koops H-P & Wagner M (1998)
Combined molecular and conventional analysis of nitrifying
bacterium diversity in activated sludge: Nitrosococcus mobilis
and Nitrosospira-like bacteria as dominant populations. Appl
Environ Microbiol 64: 3042–3051.
Juretschko S, Loy A, Lehner A & Wagner M (2002) The microbial
community composition of a nitrifying-dentrifying activated
sludge from an industrial sewage treatment plant analyzed by
the full-cycle rRNA approach. Syst Appl Microbiol 25: 84–99.
Kassen R & Rainey PB (2004) The ecology and genetics of
microbial diversity. Annu Rev Microbiol 58: 207–231.
Klausen MM, Thomsen TR, Nielsen JL, Mikkelsen LH & Nielsen
PH (2004) Variations in microcolony strength of probe-
defined bacteria in activated sludge flocs. FEMS Microbiol Ecol
50: 123–132.
Kloeke FO & Geesey GG (1999) Localization and identification of
populations of phosphatase-active bacterial cells associated
with activated sludge flocs. Microbial Ecol 38: 201–214.
Kong Y, Nielsen JL & Nielsen PH (2004) Microautoradiographic
study of Rhodocyclus-related poly-P accumulating bacteria in
full-scale EBPR plants. Appl Environ Microbiol 70: 5383–5390.
Kong Y, Nielsen JL & Nielsen PH (2005) Identity and
ecophysiology of uncultured actinobacterial polyphosphate-
accumulating organisms in full-scale enhanced biological
phosphorus removal plants. Appl Environ Microbiol 71:
4076–4085.
Kragelund C, Nielsen JL, Thomsen TR & Nielsen PH (2005) In
situ physiology of the filamentous Alphaproteobacterium
Meganema perideroedes in activated sludge. FEMS Microbiol
Ecol 54: 111–122.
Krieg NR (1984) Aerobic/microaerophilic, motile, helical/vibroid
gram-negative bacteria. Bergey’s Manual of Systematic
Bacteriology, Vol. 1 (Krieg NR & Holt JG, eds), pp. 104–110.
Williams & Wilkins, Baltimore, MD.
Lajoie CA, Layton AC, Gregory IR, Sayle GS, Taylor DE & Meyers
AJ (2000) Zoogleal clusters and sludge dewatering potential in
an industrial activated-sludge wastewater treatment plant. Wat
Environ Res 72: 56–64.
Lee N, Nielsen PH, Andreasen KH, Juretschko S, Nielsen JL,
Schleifer KH & Wagner M (1999) Combination of fluorescent
in situ hybridization & microautoradiography – a new tool for
structure function analyses in microbial ecology. Appl Environ
Microbiol 65: 1289–1297.
Leifson E (1962) The bacterial flora of distilled and stored water.
III. New species of the genera Corynebacteria, Flavobacterium,
Spirillum and Pseudomonas. Int Bull Bacteriol Nomencl Taxon
12: 161–170.
Lie E & Welander T (1997) A method for determination of the
readily fermentable organic fraction in municipal wastewater.
Wat Res 31: 1269–1274.
Loy A, Horn M & Wagner M (2003) probeBase – an online
resource for rRNA-targeted oligonucleotide probes. Nucleic
Acids Res 31: 514–516.
Manz W, Amann R, Ludwig W, Wagner M & Schleifer K-H
(1992) Phylogenetic oligodeoxynucleotide probes for the
major subclasses of proteobacteria: problems and solutions.
Syst Appl Microbiol 15: 593–600.
Mechichi T, Stackebrandt E, Gad’on N & Fuchs G (2002)
Phylogenetic and metabolic diversity of bacteria degrading
aromatic compounds under denitrifying conditions, and
description of Thauera phenylacetica sp. nov., Thauera
aminoaromatica sp. nov., and Azoarcus buckelii sp. nov. Arch of
Microbiol 178: 26–35.
Nielsen JL & Nielsen PH (2002a) Enumeration of acetate-
consuming bacteria by microautoradiography under oxygen-
and nitrate respiring conditions in activated sludge. Wat Res
36: 421–428.
Nielsen JL & Nielsen PH (2002b) Quantification of functional
groups in activated sludge by microautoradiography. Wat Sci
Technol 46: 389–395.
Nielsen PH, Raunkjær K, Norsker NH, Jensen NA & Hvitved-
Jacobsen T (1992) Transportation of wastewater in sewer
systems – a review. Wat Sci Technol 25: 17–31.
Nielsen JL, Mikkelsen LH & Nielsen PH (2001) In situ detection
of cell surface hydrophobicity of probe-defined bacteria in
activated sludge. Water Sci Technol 43: 97–103.
Nielsen PH, Roslev P, Dueholm TE & Nielsen JL (2002)
Microthrix parvicella, a specialized lipid consumer in
FEMS Microbiol Ecol 60 (2007) 370–382 c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
381Ecophysiology of denitrifying bacteria in activated sludge

anaerobic-aerobic activated sludge plants. Water Sci Technol
46: 73–80.
Nielsen JL, Christensen D, Kloppenborg M & Nielsen PH (2003)
Quantification of cell-specific substrate uptake by probe-
defined bacteria under in situ conditions by
microautoradiography and fluorescence in situ hybridization.
Environ Microbiol 5: 202–211.
Nielsen PH, Thomsen TR & Nielsen JL (2004) Bacterial
composition of activated sludge–importance for floc and
sludge properties. Water Sci Technol 49: 51–58.
Olofsson AC, Zita A & Hermansson M (1998) Floc stability and
adhesion of green-fluorescent-protein-marked bacteria to
flocs in activated sludge. Microbiol 144: 519–528.
Ostle AG & Holt JG (1982) Nile blue A as a fluorescent stain for
poly-b-hydroxybutyrate. Appl Environ Microbiol 44: 238–241.
Philippot L & Hallin S (2005) Finding the missing link between
diversity and activity using denitrifying bacteria as a model
functional community. Curr Opinion Microbiol 8: 1–6.
Pot B, Gillis M & de Ley J (1991) The genus Aquaspirillum. The
Prokaryotes: A Handbook of the Biology of Bacteria:
Ecophysiology, Isolation, Identification, Application (Balows A,
Truper HG, Dworkin M, Harder W & Schleifer KH, eds), pp
2569–2582. Springer-Verlag, New York.
Purkhold U, Pommerening-Roser A, Juretschko S, Koops HP &
Wagner M (2000) Phylogeny of all recognized species of
ammonia-oxidizers based on comparative 16S rRNA and
amoA sequence analysis: implications for molecular diversity
surveys. Appl Environ Microbiol 66: 5368–5382.
Rosello-Mora RA, Wagner M, Amann R & Schleifer K-H (1995)
The abundance of Zoogloea ramigera in sewage treatment
plants. Appl Environ Microbiol 61: 702–707.
Scholten E, Lukow T, Auling G, Kroppenstedt R, Rainey F &
Diekmann H (1999) Thauera mechernichensis sp. nov., an
aerobic denitrifier from a leachate treatment plant. Int J Syst
Bacteriol 49: 1045–1051.
Seviour RJ & Blackall LL (1999) The Microbiology of Activated
Sludge. Kluwer academic publisher, Dordrecht.
Seviour RJ, Mino T & Onuki M (2003) The microbiology of
biological phosphorus removal in activated sludge systems.
FEMS Microbiol Rev 27: 99–127.
Song B, Young L & Palleroni N (1998) Identification of denitrifier
strain T1 as Thauera aromatica and proposal for emendation
of the genus Thauera definition. Int J Syst Bacteriol 48:
889–894.
Song B, Palleroni NJ, Kerkhof LJ & Haggblom MM (2001)
Characterization of halobenzoate-degrading, denitrifying
Azoarcus and Thauera isolates and description of
Thauera chlorobenzoica sp. nov. Int J Syst Evol Microbiol 51:
589–602.
Thomsen TR, Nielsen JL, Ramsing NB & Nielsen PH (2004)
Micromanipulation and further identification of FISH-
labelled microcolonies of a dominant denitrifying bacterium
in activated sludge. Environ Microbiol 6: 470–479.
Thomsen TR, Kragelund C & Nielsen PH (2006) Abundance and
physiology of Aquaspirillum-related filamentous bacteria in
activated sludge. Wat Sci Technol 54: 237–245.
van Loosdrecht MCM, Smolder GJ, Kuba T & Heijnen JJ (1997)
Metabolism of micro-organisms responsible for enhanced
biological phosphorus removal from wastewater. Antonie van
Leeuwenhoek 71: 109–116.
Wagner M & Loy A (2002) Bacterial community composition and
function in sewage treatment systems. Current Opinion
Biotechnol 13: 218–227.
Wagner M, Loy A, Nogueira R, Purkhold U, Lee N & Daims H
(2002) Microbial community composition and function in
wastewater treatment plants. Antonie van Leeuwenhoek 81:
665–680.
Zeng RJ, Saunders AM, Yuan Z, Blackall LL & Keller J (2003)
Identification and comparison of aerobic and denitrifying
polyphosphate-accumulating organisms. Biotechnol Bioeng 83:
140–148.
Zita A & Hermansson M (1997) Effects of bacterial cell surface
structures and hydrophobicity on attachment to activated
sludge. Appl Environ Microbiol 63: 1168–1170.
FEMS Microbiol Ecol 60 (2007) 370–382c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
382 T.R. Thomsen et al.