dynamics of a subtropical plankton community, with emphasis on the copepod mesocyclops edax
TRANSCRIPT

Dynamics of a subtropical plankton community, with emphasis on the copepodMesocyclops edax
Grace A. Wyngaard', James L . Elmore 2 & Bruce C. CowellDepartment of Biology, University of South Florida, Tampa, FL 33620, U .S.A .'Department of Zoology, University of Maryland, College Park, MD 20742, U .S.A .2Environmental Sciences Division, Oak Ridge National Laboratory, Oak Ridge, TN 37830, U .S.A .
Keywords: subtropical zooplankton, zooplankton population dynamics, Mesocyclops edax, stationary agedistribution, Edmondson egg ratio model
Abstract
The zooplankton community of a subtropical, eutrophic lake was studied over an annual cycle .Mesocyclops edax, the numerically dominant crustacean, exhibited a stationary age distribution for most ofthe year (July-March) . Per capita growth rates (r) of M. edax approximated zero throughout most of thestudy, indicating a close correspondence between birth and death rates . High death rates in the spring andsummer probably were due to intense predation, since physiological mortality was low and predators wereabundant. Multiple regression analyses indicated that much of the variation in M. edax birth rates could beexplained by abundance of their animal prey. Other plankton in this lake exhibited considerable seasonality,but not to the marked degree characteristic of temperate populations . The number of zooplankton specieswas low and cladocerans were scarce, as is characteristic of many eutrophic Florida Lakes .
Introduction
Much of our understanding of zooplanktonecology is derived from temperate populations .Only a few studies characterize subtropical andtropical zooplankton dynamics (Burgis 1971, 1973 ;Gophen 1972 ; Cowell et al. 1975; Gaudy 1976 ;Richerson et al. 1977; Infante 1978 ; Young 1978 ;Lewis 1979 ; Elmore 1980) . In less seasonal climates,zooplankton life history tactics and species com-position may differ from those in temperate popu-lations . For example, climates with muted seasonalpulsing may favor dominance of copepods overcladocerans (Allan 1976) . Reproductive efforts oftropical plankters may exceed those of temperatespecies (Burgis & Walker 1972 ; Nilssen 1980) .However, we lack the necessary data describingsubtropical and tropical populations to verify orreject these ideas .
This study describes the zooplankton communityof a subtropical lake, with emphasis on the popula-tion dynamics of the cyclopoid copepod Meso-
Hydrobiologia 89,39-48(1982) . 0018-8158/82/0891-0039/$02 .00 .© Dr W. Junk Publishers, The Hague . Printed in The Netherlands .
cyclops edax (S .A . Forbes) . Probable factorsinfluencing M. edax population dynamics wereidentified by regressing chlorophyll a concentra-tion, abundance of animal prey, and temperature,on reproductive rate . Age structure and per capitabirth, death and growth rates were examined foradditional information on population dynamics .
Methods
Field and laboratory techniques
Lake Thonotosassa, Hillsborough County, Flo-rida, has a surface area of approximately 345 ha, is2.5 km long and 1 .5 km broad . Maximum depth is5 .1 m and mean depth is 3 .5 m . Detailedhydrographic, chemical and biological features ofthis eutrophic lake are given by Cowell et al. (1975),Young (1978), and Elmore (1980). Cowell et al.(1975) found no significant horizontal patchiness ofcopepods (nauplii, copepodids, or adults) and

40
cladocerans . Rotifer abundances were quantita-tively similar (p < 0 .05) at several of the deepersampling stations (> 3 .5 m) . Thus in this study, allsampling was conducted at one station in thedeepest part of the lake (4 .5-5 .0 m) .
Zooplankton samples were collected at weeklyintervals from July 1976 through June 1977 with ametered high-speed Miller plankton sampler(Miller 1961) equipped with No . 10 (158 pm) andNo. 20 (76 pm) nets . Two samples were collectedwith each net by towing the sampler obliquelythrough the water column . The volume of waterfiltered in each tow was usually between 2000 and3000 1 . Samples were narcotized with CO 2(Gannon& Gannon 1975) and preserved with modifiedLugol's solution (Edmondson 1959) . A preliminarydiel sampling indicated that M. edax copepodidsand adults remained in or near the bottom mudsduring the day and migrated to the surface at dusk(Wyngaard 1978) . Therefore all sampling wascommenced one half hour after sunset, to obtain anaccurate reflection of the population age structure .
Rotifers and copepod nauplii were enumeratedfrom the No . 20 samples, and all other organismsfrom the No . 10 samples . Three 1-ml subsamples(usually 0 .4% of field sample volume) of eachrotifer sample were counted in a Sedgwick-Raftercell at 40X magnification ; the entire cell wascounted. Crustacean zoo-plankton samples werediluted to a known volume and three 5 .5 mlsubsamples (usually 0.2% of field sample volume)were counted in a modified Ward rotary chamber .Over 1000 individuals were enumerated in eachsubsample. The coefficient of variation amongsubsamples was always less than 10% and replicatesamples differed significantly on only one occasion .
Egg sacs were dissected from M. edax females (insubsamples of collections with the No. 10 net) andthe number of eggs per female were determined .The number of sacs and eggs counted was variable,but usually over 1000 eggs were enumerated in eachsubsample. Some sample contained loose eggs, andthese eggs were assigned to M. edax since their eggsare released from the sac more easily than those ofDiaptomus dorsalis, the other possible source ofloose eggs . Smyly (1976) also made this assump-tion .
Development times of eggs of M. edax weredetermined at 30, 25, and 15 ° C using theregression method of Edmondson (1965) . Expe-
riments were conducted when the temperature ofthe lake was similar to the experimental tempera-ture. Each individual female with eggs was placed in10 ml of glass fiber (Whatman GF/C) filtered lakewater in a glass test tube and the time to hatchingwas noted. The number of females with eggs wasregressed against time and the time intercept wasused to estimate the mean egg development time . Atleast 100 individuals were used in each experiment,and observations were made every 2 h at 30 ° C and25 °C and every 4 h at 15 °C .Chlorophyll a content was used as an estimate of
phytoplankton abundance . Water samples fromthe surface, 2, and 4 m depths were collected with a2-1 PVC Van Dorn water bottle at the north andsouth points of plankton tows . We mixed samplesfrom each depth in equal parts before pigmentcontent determination . Usually 150-250 ml ofwater was filtered through glass fiber filter paper(Whatman GF/C) in darkness (A.P.H .A . 1976) .Samples were extracted with acetone in a darkfreezer overnight . Optical density was read on aBeckman DB spectrophotometer at 663 and750 nm wavelengths, before and after acidificationwith 4 N HCl (Golterman 1979) .
Temperature was recorded at 1 m intervals witha thermistor calibrated with a thermometer .
Statistical analyses
Least-squares, stepwise multiple regression ana-lyses (BMDP2R of the Biomedical ComputerPrograms of the University of California, Berkeley,1977 ed .) were used to determine the relationshipbetween independent variables and the repro-ductive rate of M. edax . The stepping algorithm ishierarchical. The most strongly correlated in-dependent variable is entered first, and additionalvariables are entered based on increasing themultiple R . Variables were entered based on anF-to-enter probability of 0.01 . All independentvariables were tested for normality and multi-colinearity .
Egg ratio model
M. edax is ideally suited to the Edmondson (1960)egg ratio model as the eggs are carried in sacs,allowing easy counting, and sexes may bedistinguished readily. This population reproduced

continuously and exhibited a stable age distribu-tion; both are important underlying assumptions ofthe model not usually met by temperate copepodpopulations .
Per capita reproductive rate, b, was calculatedaccording to Paloheimo (1974) :
females eggsb=ln
+1total females
D
where D = egg development time (days) and a 1 :1sex ratio of eggs is assumed . The total number offemales (N) is
N = 0 .5 -(no . nauplii) + proportion of females inadult population .(no. copepodids) + (no . maturefemales) .
The intrinsic rate of natural increase, r, wascalculated using:
(In N 2 - In N,)r = (t 2 - t1)
(1)
where Ni = total female density at time t 1 (days) .Death rates, d, were estimated using d = b - r. Sinceestimates of b correspond to actual sample datesand estimates of r to the midpoint between twodates, r values were interpolated linearly in thecalculation of d .
Results
Surface temperatures in Lake Thonotosassaranged from 11 .8°C to 31 °C (Fig . la). Summertemperatures (June-September) were fairly con-stant, ranging from 28 .1 to 31 °C . Short termthermal stratification occurred occasionally fromMay through October. The maximum differencebetween the surface and bottom was 3 .1 °C, but thegradient usually was less than 2 ° C . The lake wasessentially isothermal from 20 October through 20April .
Chlorophyll aconcentrations peaked (>60µg.1-1)during March and from May through August (Fig .l b) . Lower concentrations (< 40µg .1 -1 ) occurredfrom October through February (Fig. Ib) .
Rotifer densities were relatively constant fromJune through October, but declined appreciably
during the winter (Fig . lc) . During late April andearly May, rotifers increased markedly and reachedmaximum density. The rotifer populations consist-ed principally of three species : Brachionus havae-nensis Rousselet, Branchionus caudatus Barrois etDaday, and Conochiloides dossarius (Hudson) .These species comprised 48%, 41% and 3%, respec-tively, of total rotifer numbers and were presentthroughout the entire year . The remaining rotifersincluded 23 species found in very low densities .
Diaptomus dorsalis abundance followed apattern similar to that of the rotifers (Fig . Id) .Densities were high and relatively constant in thesummer months, and decreased through the falland winter with minimum values occurring inFebruary. A marked increase occurred in Marchwith maximum densities in May . The three majorlife stages of D. dorsalis showed similar seasonalpatterns of abundance .
Other crustaceans that were occasionallycollected in the samples were the cyclopoidsEuyclops agilis (Koch) and Tropocyclops prasinus(Fischer), and the cladocerans Daphnia ambigua(Scourfield), Bosmina longirostris (0 . F. Muller),and Diaphanosoma (possibly leuchtenbergianumFisher) .
Population dynamics of Mesocyclops edax
M. edax was the numerically dominantcrustacean and did not exhibit seasonal patterns ofabundance similar to that of D. dorsalis and therotifers . Population size, as well as percentages ofnauplii, copepodids, and adults, remained ratherconstant from 14 July 1976 through 9 March 1977,but considerable variation occurred from Marchthrough June (Figs. 2a and 2b) .Visual examinationof the age structure suggests a stationary age dis-tribution during the first 9 months of the study . Theadult sex ratio was quite constant from July throughMarch, with an average female : male sex ratio of1 :1 .5 . However, from March through May the sexratio approximated 1 : 1 .The mean and 95% confidence intervals for egg
hatching times used to calculate birth rates at eachtemperature (30, 25 and 15 °C) were 28 .0 ± 1 .27,48.9 ± 2 .39, and 130 ± 1 .95 h respectively . The plotof number of females versus time did not departappreciably from the calculated regression line,indicating continuous egg hatching . Egg hatching
4 1
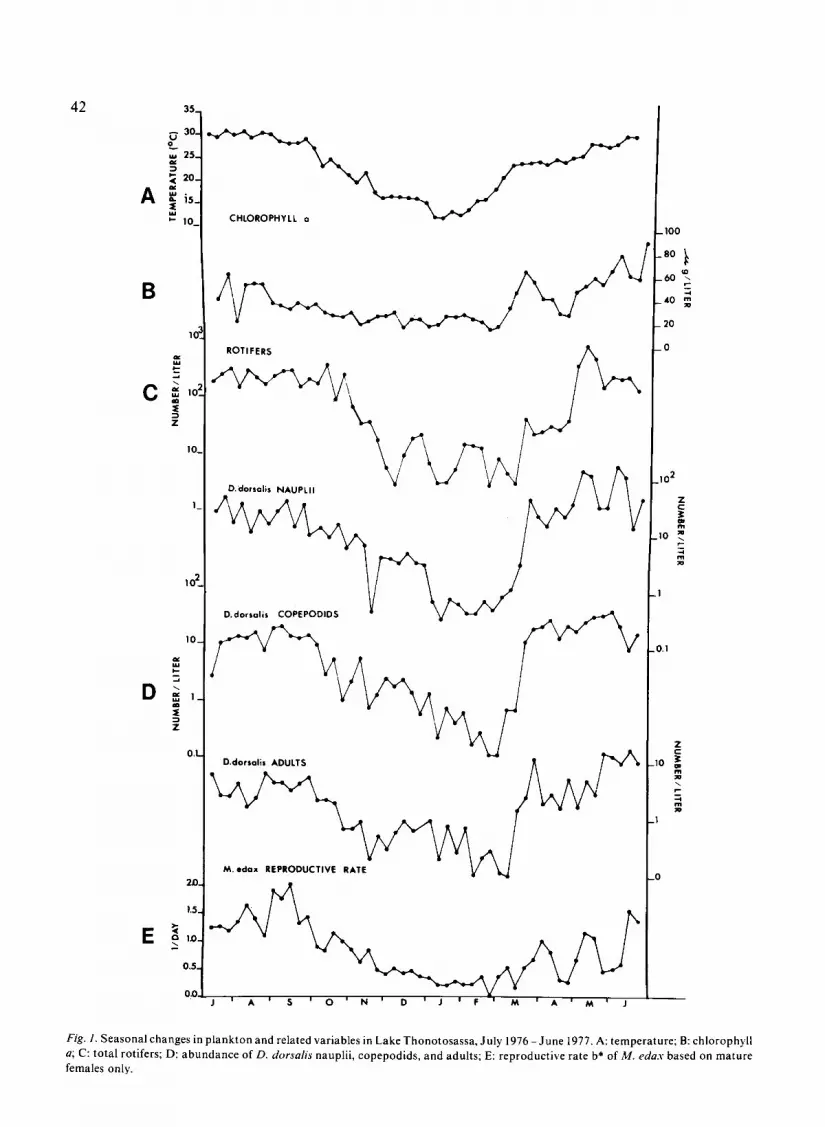
Fig. ] . Seasonal changes in plankton and related variables in Lake Thonotosassa, July 1976-June 1977 . A : temperature ; B : chlorophylla ; C : total rotifers ; D : abundance of D . dorsalis nauplii, copepodids, and adults ; E : reproductive rate b* of M . Max based on maturefemales only .

times at various temperatures were calculated fromthe linear relationship between log (egg develop-ment time) and temperature . For several dates inJanuary and February, when lake temperature wasbelow 15 'C, an estimate of egg hatching time wasobtained by extrapolation of the above relation-ship .
Population parameters, b, r, and d showedconsiderable seasonal variation . The per capitareproductive rate (b), in units of female birthsfemales -days ranged from 0 .01 in February to1.12 in June (Fig. 3a). The value of 1 .12 isnoticeably higher than other values obtained in thisstudy, and probably reflects an age structure effect .Estimates of b remained high during the summer,declined gradually during the fall and winter, andthen increased during the spring . Turnover times,calculated as 1 / b, were shortest during the summermonths of July-September (3 .0 days) and longest inthe winter months of January-March (13 .8 days) .
4 3
Fig. 2 . A: weekly densities of M. edax nauplii (A), copepod ids (0), and adults(*) in Lake Thonotosassa from July 1976 to June 1977 . B:percentage of mature females in Al . edax population .
The yearly mean turnover time was 7 .9 days. Theobserved per capita growth rate (r) approximatedzero from July to May, indicating an essentiallystationary population over this time period (Fig .3b) . During May and June, r values were morevariable . Per capita death rates closely balanced percapita birth rates on most sampling dates (Fig. 3a) .Death rates ranged from a high of 1 .27females -female I - day I in June to a low of -0 .03females . female- s .day s in March (the only nega-tive value obtained) .
Correlation of M . edax reproductive rate with preyand abundance temperature
Temperature and the abundance of animal prey(D. dorsalis and rotifers) were used as independentvariables to regress on reproductive rate of M. edaxduring the period of stable age structure (14 July-9March). For this analysis, mature females, rather

44
1 .3
11 .2
II
1I
1 .1
0.6
0.5
I
f
0.3
1
1
1'
I
I
I
1I
I
I0.2
'
IX
4
1
0.1
'1
0.0
-0.1
I
s
I
I
trY
I
I
Ir
1
IItI
1A' S
0
N
D
' J ' FI M
A
M
J
Fig. 3. A : Per capita birth rates (b) (-) and death rates (d) (---) ofM. eda .r in Lake Thonotosassa from July 1976 to June 1977 .
than total females, was used in equation (1), so that
Table 1 . Stepwise regression analysis, multiple correlation
effects of food and temperature would be examined
coefficients (R), and increase in coefficient of determination (R2)
only for those individuals capable of reproducing .
for reproductive rate, b*, of Mesocvclops edax. These statisticsare calculated for the time period during which the age structure
This reproductive rate (Fig . le) varied in a pattern
of M. edax appeared stationary (14 July 1976-9 March 1977) .very similar to b, which is to be expected since theproportion of mature females was quite constant(Fig . . 3a) . Regression analyses indicated thatabundance of D. dorsalis copepodids was the bestsingle predictor, accounting for 78% (p < 0 .01) ofthe variation in reproductive rate (Table 1) . Thesecond variable which contributed significantly toexplaining the variation in reproductive rate wastemperature, which combined with density of D .
Stepwisevariable
R R2
Diaptomus copepodids 0 .88 0 .78Temperature 0 .92 0 .86Rotifers 0.94 0 .88

-.20-
-.30-
B
I A' S' O' N' D' J' F' M ' A' M `JFig. 3. B : Per capita growth rate (r) of M. edax in Lake Thonotosassa .
dorsalis copepodids, accounted for 86.1% (p <0.01) of the variability in reproductive rate. Thefinal variable which significantly reduced theresidual variance was total rotifer density . Thesethree variables combined explained 88 .9% (p <0.01) of the variation in reproductive rate of M.edax.
The simple correlation matrix of reproductiverate, prey abundances and temperature indicatedmulticolinearity among most of the variables. Sincethe independent variables correlated with eachother, all interpretations of the influence ofindependent variables (prey abundances andtemperature) on reproductive rate must be viewedwith caution. However, relationships betweenreproductive rate and the independent variables didapproximate linear functions and the variableswere normally distributed .
Discussion
The relative constancy of the age structure of M.edax, compared to temperate copepod popula-tions, is striking. To our knowledge, only two othercopepod populations have been reported to exhibit
45
stable age distributions : M. leuckarti in subtropicalLake Kinneret (Gophen 1972), and Thermocyclopscrassus (Fischer, 1853) in tropical Lake George(Burgis 1971). However, generalizations aboutcyclopoid age structure in subtropical and tropicalareas will require more data . Explanation of thestationary age distribution of the M. edaxpopulation can be obtained from an understandingof factors affecting birth and death rates .
Temperature clearly influenced variation inreproductive rate . The seasonal temperature rangeof 11 .8'C is substantial for this warm waterspecies, and our data show that eggs require almostfive times as long to develop at 15 °C as at 30 °C .This alone could account for much of the variationin b throughout the study period . M. edax does notundergo diapause in Lake Thonotosassa as it doesin northern lakes (Comita 1972), probably becausethe winter temperature in Lake Thonotosassaremains relatively high . However, reproductionnearly ceases during the minimum temperatures inFebruary .
The influence of food on reproductive rate is lesseasily demonstrated by this study . The multi-colinearity of the variables makes these resultsdifficult to interpret. The regression analysis
.30-
.20-
.10..
ra0 0.0
- .10-

46
indicated that reproductive rate of M. edax variedsimilarly to the abundance of D . dorsaliscopepodids, and less so with rotifer numbers andtemperature . The use of a No . 20 net to samplerotifers in this study may have underestimated theabsolute rotifer densities by a substantial amount(Karabin 1978). However, this should not appre-ciably affect general trends in rotifer abundance northe regression analyses used here . The highproportion of variation in M. edax reproductiverate explained by abundance of D. dorsaliscopepodids does suggest that food is important .Theper capita birth rates of M. edax in this study
are comparable to values of other warm watercyclopoids (Burgis 1971 ; Gophen 1972 ; Burns 1979 ;but see Gophen 1978) . The b values of M. edax inthis study were somewhat lower than those of D .dorsalis, the calanoid copepod in Lake Thonoto-sassa (Elmore 1980) . Direct comparison of thesebirth rates to those of northern populations is notappropriate, since most temperate populationsexhibit a more or less pulsed reproduction duringsome portions of the year . This results from acohort age structure and inflates birth rateestimates . In contrast, the presence of alldevelopmental stages and reproducing individualsofM. edax throughout the year in this study resultsin a dramatically different age structure . Suchdifferences in age structure may bias comparison ofestimates of b .
Factors influencing M. edax death rates can beobtained from in situ life table studies of M. edax inLake Thonotosassa and knowledge of potentialpredators present . Culture of M. edax in 70 mlmesh cages suspended in Lake Thonotosassa(Wyngaard, in prep .) revealed 93% survivorship toadult and a physiological longevity of about onemonth. During summer months, M. edax exhibiteda stationary age distribution as well as high percapita birth rates. Since parasitism was nevernoticed and egg mortality was virtually zero, thesedata strongly suggest that an external mortalitysource such as predation is responsible for highsummer death rates . High populations of plankti-vores are known to exist in Lake Thonotosassa,including the phantom midge, Chaoborus puncti-pennis (Cowell & Vodopich 1980) and three fishspecies: gizzard shad Dorsoma cepedianum,threadfin shad Dorosoma petenense, and Tilapiaaurea (Florida Game and Freshwater Fish Comm .,
unpubl . data). Shad and tilapias feed onzooplankton only as juveniles, which are mostabundant during the spring and summer spawningperiods (Applegate & Mullan 1969 ; Moriarty et al.1972). Thus vertebrate planktivore abundancescorrelate positively with the high spring andsummer death rates and with low fall and winterdeath rates of M. edax. Chaoborus may be animportant predator on M. edax in Lake Thonoto-sassa . Lewis (1979) reported Thermocyclopshyalinus copepodids to suffer considerable pre-dation by Chaoborus in tropical Lake Lanao .
Two additional lines of evidence suggest that fishpredation on zooplankton is intense in this lake .Limnetic cladocerans are sparse and both M. edaxand D. dorsalis exhibit strong vertical migrations(Wyngaard 1978). Brooks & Dodson (1965) andZaret & Suffern (1976) have demonstrated similarpatterns in lakes with high fish predation.
Plankton populations in this study exhibitedseasonality in their dynamics, but not to the markeddegree observed in temperate populations . Thenarrower temperature range and more frequentmixing in shallow subtropical lakes relative todimictic temperate lakes may result in less variablenutrient resource distribution over the year (Lewis1976) . This, combined with a high trophic state mayreduce the intensity, frequency and duration offood limitation. The constancy of M. edax agestructure and population density in this study maybe a common occurrence in lowland tropical andsubtropical cyclopoid populations . The relativelysimple zooplankton species composition, con-tinuously reproducing copepod populations, anddepauparate limnetic cladoceran fauna of LakeThonotosassa are characteristic of many eutrophicFlorida lakes (Young 1978 ; Elmore, unpubl . data) .A few investigations suggest that these trends maycharacterize lowland tropical lakes as well (Burgis1973; Gophen 1972; Dumont 1979 ; Deevy 1979 ;Lewis 1979), but additional studies clearly areneeded to verify these generalizations .
Summary
The seasonal abundance of zooplankton wasstudied in a eutrophic lake in subtropical Florida .The dominant crustacean, M. edax, exhibited astationary age distribution from July to March .

The population growth rate, r, approximated zeroon most dates. Birth rates were estimated using theEdmondson egg ratio model . A stepwise multipleregression analysis was used to identify predictorvariables ofM. edax reproductive rate . Abundanceof D. dorsalis, a main prey of M. edax in this lake,explained 78% of the variation in reproductive rate .When temperature and rotifer abundance wereadded to the analysis, a total of 86 .1 % of variationin reproductive rate was explained . The highsummer death rates of M. edax were attributed tointense fish predation .
In contrast to M. edax, D. dorsalis abundance,total rotifer abundance, chlorophyll a concentra-tion, and temperature exhibited considerableseasonal fluctuations. Zooplankton species com-position in this lake was low relative to most northtemperate lakes and cladocerans were scarce. Morestudies of subtropical and tropical zooplanktoncommunities are needed to verify whether thespecies composition and seasonal dynamics ob-served in this study are typical of subtropical andtropical lakes .
Acknowledgments
We wish to thank V . Grey, E . Kosco, G . Lukos,C. Proffitt, R . Reese, and D. Vodopich forassistance in laboratory and field aspects of thisstudy . J . D. Allan, W . M . Lewis, C . E. King and G .B. Williamson commented on earlier drafts of themanuscript .
References
Allan, J . D ., 1976 . Life history patterns in zooplankton . Am .Natur . 971 : 165-180 .
A.P.H.A ., 1965 . American Public Health Association andothers . Standard Methods for Examination of Water andWaste-Water (12th edn .) New York, 769 p .
Applegate, R . L . & Mullan, J . W ., 1969 . Ecology of Daphnia inBull Shoals Reservoir . U . S . Bur. of Sport Fish and Wildl .Res . Rep . 74 . 23p .
Brooks, J . L . & Dodson, S . I ., 1965 . Predation, body size, andcomposition of plankton . Science 150 : 28-35 .
Burgis, M . J ., 1971 . The ecology and production of copepods,particularly Thermocyclops hyalinus, in the tropical LakeGeorge, Uganda . Freshwat . Biol . 1 : 169-192 .
Burgis, M . J ., 1973 . Observations on the Cladocera of LakeGeorge, Uganda . J . Zool . Lond . 170: 339-349 .
47
Burgis, M . J. & Walker, A . F., 1972 . A preliminary comparisonof the zooplankton in a tropical and a temperate lake (LakeGeorge, Uganda and Loch Leven, Scotland) . Verb int .Verein . Limnol . 18 : 647-655 .
Burns, C . W., 1979 . Population dynamics and production ofBoeckella dilatata (Copepoda : Calanoida) in Lake Hayes,New Zealand . Arch . Hydrobiol .(Suppl .) 54 : 409-465 .
Cowell, B . C ., Dye, C . W. & Adams, R. C ., 1975 . A synopticstudy of the limnology of Lake Thonotosassa, Florida . PartI . Effects of primary treated sewage and citrus wastes .Hydrobiologia 46 : 301-345 .
Cowell, B. C. & Vodopich, D . S ., 1981 . Distribution andseasonal abundance of benthic macroinvertebrates in asubtropical Florida lake . Hydrobiologia 78 : 97-105 .
Deevey, E . S ., 1980 . Structure of zooplankton communities inPeten Lake District . In: Kerfoot, W . C ., (ed .) The Evolutionand Ecology of Zooplankton Communities . Am. Soc .Limnol. Oceanogr. Spec . Symp. University Press of NewEngland, Vol . 3, pp . 669-678 .
Dumont, H. J., 1980. Zooplankton and the science ofbiogeography : the example of Africa . In : Kerfoot, W . C .(ed.) The Evolution and Ecology of ZooplanktonCommunities. Am. Soc . Limnol. Oceanogr. Spec. Symp .University Press of New England, vol . 3 pp . 685-698 .
Edmondson, W . T ., 1960 . Reproductive rates of rotifers innatural populations . Mem. Ist. Ital . Idrobiol. 12 : 21-77 .
Elmore, J . L ., 1980 . An experimental study of factors regulatingthe distribution of Diaptomus (Copepoda : Calanoida) insubtropical Florida . Ph . D . thesis, Univ . of South Florida,Tampa . 113 pp .
Gannon, J . E. & S. A. Gannon, 1975 . Observations on thenarcotization of crustacean zooplankton . Notes andobservations . Crustaceana 28 : 220-224 .
Gaudy, R ., 1976. Etude du plankton de la zone nord de la rade deVillefranch-sur-Mer a la fin du printemps (17 mai 71 au 16juin 71) . III . Production secondaire des copepbdespelagiques . Vie Milieu, Ser . B 26 : 77-106 .
Golterman, H . I ., 1969 . Methods for Chemical Analysis of FreshWaters . Int. Biol. Program Handbook 8 . BlackwellScientific Publications, Oxford . 112 pp .
Gophen, M ., 1972 . Zooplankton distribution in Lake Kinneret(Israel) 1969-1970 . Israel J . Zool . 21 : 17-27 .
Gophen, M ., 1978 . Errors in the estimation of recruitment ofearly stages of Mesocyclops leuckarti (Claus) caused by thediurnal periodicity of egg production . Hydrobiologia 57 :59-64 .
Infante, A ., 1978. The zooplankton of Lake Valencia(Venezuela) . 1 . Species composition and abundance . Verb .int . Verein Limnol . 20 : 1186-1191 .
Karabin, A ., 1978 . Studies on the usefulness of different meshsize plankton nets for thickening zooplankton . Ekol . pol . 26 :479-490 .
Kiefer, F ., 1929 . Crustacea copepoda - II . Cyclopoidagnathostoma . Das Tierreich . Lief . 53 : 51-102 .
Lewis, W. M . Jr ., 1976 . Observations on the superficial sedimenttemperatures of some lakes in the southeastern UnitedStates . Freshwat . Biol. 6 : 49-57 .
Lewis, W . M. Jr ., 1977 . Feeding selectivity of a tropicalChaoborus population . Freshwat . Biol . 7 : 311-325 .

48
Lewis, W. M. Jr., 1979 . Zooplankton Community Analysis .Studies on a Tropical System . Springer-Verlag, New York .163 pp .
Miller, D ., 1961 . A modification of the small Hardy planktonsampler for simultaneous high-speed plankton hauls . Bull .Mar. Ecol . 5 : 165-172 .
Moriarity, D . J . W., Darlington, J . R . E . C., Dunn, I . G .,Moriarity, C . M . & Tevlin, M . P ., 1973 . Feeding and grazingin Lake George, Uganda . Proc . R . Soc . Lond . 184 :299-319 .
Nilssen, J . P ., 1980 . When and how to reproduce : a dilemma forlimnetic cyclopoid copepods. In : Kerfoot, W . C ., (ed .) . TheEvolution and Ecology of Zooplankton Communities . Am .Soc. Limnol. Oceanogr. Spec . Symp. University Press ofNew England, Vol . 3, pp . 418-426 .
Paloheimo, J . E., 1974 . Calculation of instantaneous birth rates .Limnol. Oceanogr . 19 : 692-694 .
Richerson, P . J ., Widmer, C . & Kittel, T ., 1971 . The limnologyof Lake Titicaca (Peru-Bolivia) . A large, high altitudetropical lake . Inst . Ecology . Publ . No. 14. University ofCalifornia, Davis . 78 p .
Smyly, W . J . P ., 1976. The effects of enclosure on zooplanktonpopulation dynamics . Freshwat . Biol . 6 : 241-251 .
Wyngaard, G . A ., 1978 . The relationship between thereproductive rate of a predaceous copepod, prey itemabundance and temperature . M.S . thesis, University ofSouth Florida, Tampa, 57 pp .
Wyngaard, G. A . In situ life table of a subtropical copepod .(In prep.) .
Young, S . N ., 1978 . The relationship between abundance ofcrustacean zooplankton and trophic state indicators infourteen Central Florida lakes . M .S . thesis, University ofSouth Florida, Tampa . 105 pp .
Zaret, T. M & J. S . Suffern, 1976 . Vertical migration inzooplankton as a predator avoidance mechanism . Limnol.Oceanogr. 21 : 804-813 .
Received 2 March 1981 .