drosophila hox complex downstream targets and the function of homeotic genes
TRANSCRIPT

Yacine Graba, Denise Aragnol and Jacques Pradel
Summary
Hox complex genes are key developmental regulators highly conserved throughout evolution. The encoded proteins share a 60-amino-acid DNA-binding motif, the homeodomain, and function as transcription factors to control axial patterning. An important question concerns the nature and function of genes acting downstream of Hox proteins. This review focuses on Drosophila, as little is known about this question in other organisms. The noticeable progress gained in the field during the past few years has significantly improved our current understanding of how Hox genes control diversified morphogenesis. Here we summarise the strategies deployed to identify Hox target genes and discuss how their function contributes to pattern formation and morphogenesis. The regulation of target genes is also considered with special emphasis on the mechanisms underlying the specificity of action of Hox proteins in the whole Accepted
20 January 1997 animal.
Introduction The origin of pattern diversity within metazoans is one of the most intriguing questions to challenge biologists. Genetic analysis in certain model systems has led to the identifica- tion of mutations that dramatically affect pattern elements, and has provided the framework for a better understanding of the biological events controlling pattern differentiation. One particular class of mutations often transforms one part of the body into another one. The discovery of these ‘homeotic’ transformations led to identification in Drosophila of the homeotic (Hox) genes(’$*), which were subsequently found in a wide range of other organisms, from hydra and nematodes to vertebrates, including mouse and human(3). Disrupting Hox gene function causes dramatic morphologi- cal alterations in various tissue types. In Drosophila, Hox genes are expressed and required for accurate pattern for- mation in the epidermis, the central and peripheral nervous systems, the somatic and visceral musculature and the end~derm(~) . In the mouse, the loss or gain of Hox gene function also provokes severe morphogenetic defects in the axial skeleton, neural crest and hind braid5). Hox genes therefore specify which structure forms along the antero- posterior embryonic axis, controlling the diversified morpho- genesis of the various body parts.
As long ago as 1975, A. Garcia-Bellido predicted that the
homeotic loci do not act directly to specify morphological dif- ferences between the metameric unitd6). He proposed instead that Hox ‘selector’ genes control a battery of subor- dinate target genes, the ‘realizator’ genes, encoding cellular functions directly required in differentiation processes. The amount of molecular data obtained since then on Hox com- plex genes has largely substantiated this view. The discov- ery of the homeodomain (HD) provided a key molecular clue to the function of the encoded proteins. The HD, a 60- amino-acid motif, mediates sequence-specific DNA bind- ing(’~*), and Hox proteins have been shown to act as positive or negative transcription factordg). It is therefore thought that each metamer will enter a specific pathway and develop a unique shape, form and function, depending on the target genes that were transcriptionally regulated by the set of Hox proteins expressed in it. While the question of target gene identity is not a novel problem, significant progress in this area has been gained only recently.
In this paper we first summarise the various strategies deployed to identify Hox target genes. After next considering the nature of the identified targets, we examine how their function contributes to pattern and morphogenesis. Finally, we discuss how the work on target genes has led to the question of Hox gene specificity and how it has improved our current understanding of Hoxcomplex gene function.

Approaches for target gene identification The various strategies that have been deployed in the search for target genes are briefly presented below, together with their respective advantages and shortcom- ings, particularly concerning the isolation of directly regu- lated targets. This is obviously an important point, since the goal is to identify functions that are required to translate the positional information delivered by Hox proteins into diversi- fied morphogenetic programmes.
Most of the known targets correspond to Drosophila genes that had been previously characterised(10-20). The expression patterns of many of these genes were sugges- tive of a homeotic regulation, because their spatial domains were actually found to be changed upon Hox gene inactiva- tion. While this approach, centering on candidate genes, has proved to be efficient, it confines the search to genes that have already been identified and cloned. Moreover, it is not certain from the genetic results whether the regulation by Hox genes is direct or indirect. In one case only, that of decapentaplegic (dpp), has definitive proof been obtained that regulation occurs directly(21 ,22). An alternative approach is the detection of gene regulatory regions driving expression of a reporter in a pattern consistent with homeotic regulation, using enhancer trap method^(^^!^^). But, here again, such results do not establish whether the homeotic control occurs directly or not.
Two additional methods have been recently proposed. The first one uses a substractive hybridisation procedure to clone sequences that are differentially transcribed in response to the overexpression of a Hox gene(25), but such data do still not establish whether there is direct control. The second method consists of the selection of genomic DNA
fragments that are able to activate a reporter gene in yeast cells expressing a Hox gene product(26) (Fig. 1A). While in principle the search is limited to positively regulated sequences at non-physiological concentrations of the Hox trans-regulator and in the probable absence of cofactors known to assist homeoproteins in the whole animal, the method does have the significant advantage to select directly controlled targets in an in vivo context.
lmmunoprecipitation of DNA fragments bound in vivo to Hox proteins (with or without stabilisation of chromatin com- plexes, see Fig. 16) and cloning the neighbouring transcrip- tion units has also been a successful a p p r o a ~ h ( ’ ~ > ~ ~ - ~ ~ ) . Fur- ther evidence is required to establish that the selected sequence actually corresponds to a target gene, after show- ing that its expression actually depends on Hox gene activity. The advantages of the direct ‘molecular in vivo’ approach are multiple. Firstly, it appears to be specially designed to clone direct effectors of homeoproteins. Sec- ondly, it identifies cis-acting elements in the targets, which are key tools for the analysis in molecular terms of the mode of action of Hox proteins. Finally, it allows the isolation of tar- get genes that had not previously been cloned through clas- sical genetic screens.
Function of Hox downstream target genes: distinct contributions to pattern and morphogenesis So far, 19 candidate Hox target genes have been identified in Drosophila (Table 1). With the exception of refrofranspo- son 4 12, whose role in development remains elusive, three major functional classes can be defined from the nature of the encoded proteins and from genetic data that are avail-
Fig. 1. Direct approaches for the identification of Hox downstream target genes involve cloning of genomic DNA fragments containing Hox binding sites (A) Selection in yeast of genomic fragments containing Hox binding sites (B) lmmunopurification of genomic DNA fragments associated m vwo with Hox proteins
embryos or tissues
f preparation of chromatin
previously fixed or not
cleavage of chromatin
f immunoprecipitation of
a given homeotic protein chromatin complexes containing
f cloning of DNA fragments associated
in vivo with a Hox protein
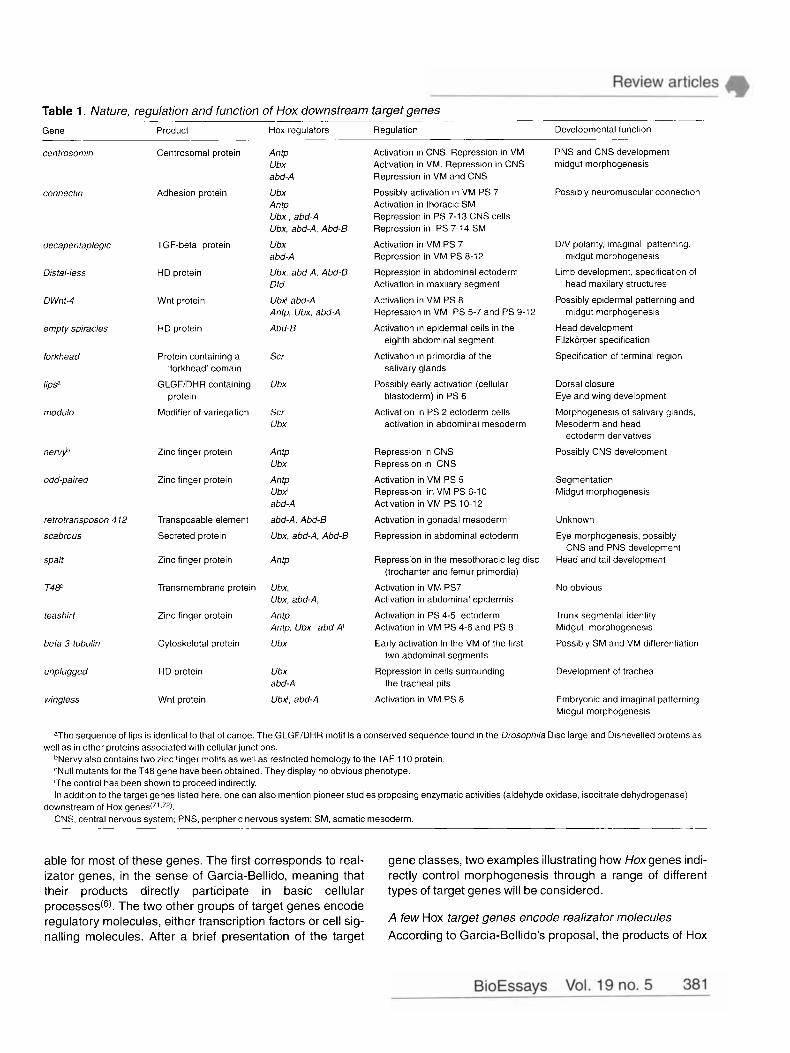
Table 1. Nature, regulation and function of Hox downstream target genes Gene
centrosomin
~~ ~
Connectin
decapentaplegic
Distal-less
DWnt-4
empty spiracles
forkhead
lip.+
modulo
nervyb
odd-paired
retrotransposon 4 12
scabrous
spalt
T4ff
teashirt
beta 3-tubulin
unplugged
wingless
Product Hox regulators Regulation Developmental function
Centrosomal protein
Adhesion Drotein
TGF-beta protein
HD protein
Wnt protein
HD protein
Protein containing a 'forkhead' domain
GLGF/DHR containing protein
Modifier of variegation
Zinc finger protein
Zinc finger protein
Transposable element
Secreted protein
Zinc finger protein
Transmembrane protein
Zinc finger protein
Cytoskeletal protein
HD protein
Wnt protein
Antp Ubx abd-A
Ubx Antp Ubx , abd-A Ubx, abd-A. Abd-B
Ubx abd-A
Ubx, abd-A, Abd-B Dfd
UbX abd-A Antp, Ubx, abd-A
Abd-B
Scr
Ubx
Scr Ubx
Antp Ubx
Antp Ubx' abd-A
abd-A, Abd-B
Ubx, abd-A, Abd-B
Antp
Ubx, Ubx, abd-A,
Antp Antp, Ubx abd-A'
Ubx
Ubx abd-A
UbX, abd-A
Activation in CNS. Repression in VM Activation in VM. Repression in CNS Repression in VM and CNS
Possibly activation in VM PS 7 Activation in thoracic SM Repression in PS 7-13 CNS cells Repression in PS 7-14 SM
Activation in VM PS 7 Repression in VM PS 8-12
Repression in abdominal ectoderm Activation in maxilary segment
Activation in VM PS 8 Repression in VM PS 5-7 and PS 9-12
Activation in epidermal cells in the eighth abdominal segment
Activation in primordia of the salivary glands
Possibly early activation (cellular blastoderm) in PS 6
Activation in PS 2 ectoderm cells activation in abdominal mesoderm
Repression in CNS Repression in CNS
Activation in VM PS 5 Repression in VM PS 6-10 Activation in VM PS 10-1 2
Activation in gonadal mesoderm
Repression in abdominal ectoderm
Repression in the mesothoracic leg disc
Activation in VM PS7 Activation in abdominal epidermis
Activation in PS 4-5 ectoderm Activation in VM PS 4-6 and PS 8
Early activation in the VM of the first two abdominal segments
Repression in cells surrounding the tracheal pits
Activation in VM PS 8
(trochanter and femur primordia)
PNS and CNS development midgut morphogenesis
Possibly neuromuscular connection
D/V polarity imaginal patterning
Limb development, specification of
Possibly epidermal patterning and
Head development Filzkorper specification
Specification of terminal region
midgut morphogenesis
head maxilary structures
midgut morphogenesis
Dorsal closure Eye and wing development
Morphogenesis of salivary glands Mesoderm and head
ectoderm derivatives
Possibly CNS development
Segmentation Midgut morphogenesis
Unknown
Eye morphogenesis, possibly
Head and tail development CNS and PNS development
No obvious
Trunk segmental identity Midgut morphogenesis
Possibly SM and VM differentiation
Development of trachea
Embryonic and imaginal patterning Midgut morphogenesis
aThe sequence of lips is identical to that of canoe. The GLGFiDHR motif is a conserved sequence found in the Drosophila Disc large and Dishevelled proteins as
bNervy also contains two zinc finger motifs as well as restricted homology to the TAF 110 protein. CNull mutants for the T48 gene have been obtained. They display no obvious phenotype. 'The control has been shown to proceed indirectly. In addition to the target genes listed here, one can also mention pioneer studies proposing enzymatic activities (aldehyde oxidase, isocitrate dehydrogenase)
CNS. central nervous system; PNS, peripheric nervous system; SM, somatic mesoderm.
well as in other proteins associated with cellular junctions.
downstream of Hox ge r~es (~ ' .~ * ) .
able for most of these genes. The first corresponds to real- izator genes, in the sense of Garcia-Bellido, meaning that their products directly participate in basic cellular types of target genes will be considered. processes@). The two other groups of target genes encode regulatory molecules, either transcription factors or cell sig- nalling molecules. After a brief presentation of the target
gene classes, two examples illustrating how Hoxgenes indi- rectly control morphogenesis through a range of different
A few Hox target genes encode realizator rno'ecules According to Garcia-Bellido's proposal, the products of Hox

downstream targets directly specify at a cellular level char- acteristics that are central in the process of morphogenesis, such as the rate and orientation of mitoses, the size and shape of cells, cell-cell adhesion and communication and, eventually, expression of terminal differentiation productd6). Three of the identified targets correspond to such realizator genes: P3-tubulin and centrosomin (cnn), which encode structural proteins, and connectin (con), a cell adhesion
Three isoforms of P-tubulin, cytoskeleton-associated proteins, are known in Drosophila. Several data, however, suggest that the P3 isoform has a more specialised function than PI and p2. Firstly, p3 is the less evolutionary con- served isoform, with 87% of homology with the major ver- tebrate isoforms, in comparison to 95% for p1 and p2. Sec- ondly, p3 is preferentially found in highly elongated microtubule arrays while 01 and p2 are present in all micro- tubule types(35). Finally, P3-tubulin is expressed, transiently, in one cell lineage only, the mesoderm. These data are consistent with a role of P3-tubulin in changes of cellular morphology occurring during morphogenetic events con- trolled by Hoxgenes.
cnn encodes an essential centrosomal protein with sev- eral coiled-coil domains and leucine zipper motifs(32). These structural features suggest that Cnn oligomerizes with itself or with other proteins to carry out its function. Cnn might control the assembling and/or stabilisation of microtubules at the centrosome, and therefore be required for proper cell division and proliferation(33). Cnn participates in multiple developmental processes, such as oogenesis, CNS and midgut morphogenesis. Further investigations should clarify how Cnn organises microtubules during axonal outgrowth and midgut constriction formation, and what are the proteins contacted during these two events influenced by the activity of Hoxgenes.
con encodes a GPI-linked cell surface protein, which acts as a homophilic cell adhesion molecule and probably has a role in the formation of neuromuscular ~onnec t ions (~~8~~) . con therefore functions as a realizator gene, controlling cell adhesion during morphogenetic events. At the end of Drosophila embryogenesis, each segment contains a stereotyped array of 30 muscles, each one specifically innervated by motoneurons. The adhesion protein Con appears during neuron outgrowth at the surface of glial cells, along the pathways leading to eight target muscles, and at the tip of motoneurons and muscle fibres as neuro- muscular connection proceeds(37). The protein is no longer found in glial cells once motoneurons have reached target muscles, nor at synaptic sites after neuromuscular junction. The dynamics of this expression and the effects of ectopic Con expression on growth cone trajectory and synapse for- mation suggest a role for Con as either an attractive(37) or a repulsive in motoneuron migration and targeting. Definitive evidence on this point should be obtained once loss-of-function mutants are isolated.
molecule(’8,29,32.33).
Most Hox target genes encode regulatory molecules About one half of the known targets, eight out of 19, produce likely transcriptional regulators. This is the case for the homeobox-containing genes Distal-less(’ 1-12), empty spir- acle(’ 3, and ~nplugged2~); n e r ~ # ~ ~ ) , odd-paired’g), ~ p a l 1 ( ~ ~ ) and teashirt (tsh)(l7), coding for zinc finger containing pro- teins; forkheadiO), which encodes an evolutionary con- served DNA binding motif; and modulo (mod)(20), a dominant suppressor of position effect variegation, which is thought to modify the structure and activity of chromatin domains. For- mally, Hoxgenes themselves belong to this group since they were the first to be identified as potential targets(39). The sec- ond group, of five genes, produces cell signalling molecules involved in transduction pathways, downstream of which transcription factors function. The Scabrous (Sca) protein(16) is thought to be involved in lateral inhibition during neurogen- e~is(~O). dpp, wingless (wg) and Dwnt-4, members of the TGF-fl and Wnt gene families, produce signalling glycopro- te in~ ( ’~13~) . lipd25) is identical to canoe; it encodes a GLGF/DHR containing protein, which participates with mem- bers of the Notch pathway in regulating cell adhesion, affect- ing cell fate determination(41). It appears therefore that most of the identified targets encode regulatory molecules, indi- cating that the genetic cascade that controls pattern forma- tion extends further, with most of the Hox target genes being themselves at the top of a genetic hierarchy.
The identification of signalling molecules downstream of Hoxgenes sheds some light to cell non-autonomous effects of homeotic mutations. Hoxgenes are indeed thought to act cell-autonomously, namely strictly within the cells where they are transcribed. This does not, however, preclude any con- sequential effects of their expression in neighbouring cells. Rather, since morphogenesis requires the functional con- nection of different cell types, it is logical to assume that Hox proteins determine not only individual cell fate but also neigh- bouring cell fates, through signalling effects. This is best exemplified by Hox gene function in Drosophila midgut mor- phogenesis. Firstly, Sex combs reduced (Scr) is essential for the development of four tubular evaginations at the anterior end of the midgut, also called ‘gastric caeca’; it itself is not expressed in caeca precursor cells, but just posterior to them(l5). Secondly, Antennapedia (Antp) and Abd-A expression activate tsh, even in cells where they are not present(42). Also, Screxpression, which does not overlap the Antp domain, is reduced in Antpdeficient animals(’5). Most likely, non-autonomous effects of Hoxgenes are mediated by secreted signalling molecules produced by target genes. This has been demonstrated in the regulation of tsh by Dpp and Wg, themselves controlled by Ubx and Abd-A(42).
Functional cooperation of target genes: a case study of morphogenesis To date, very little is known on precisely how target genes work together to control proper organogenesis. The best documented example with regard to the mechanistic inte-
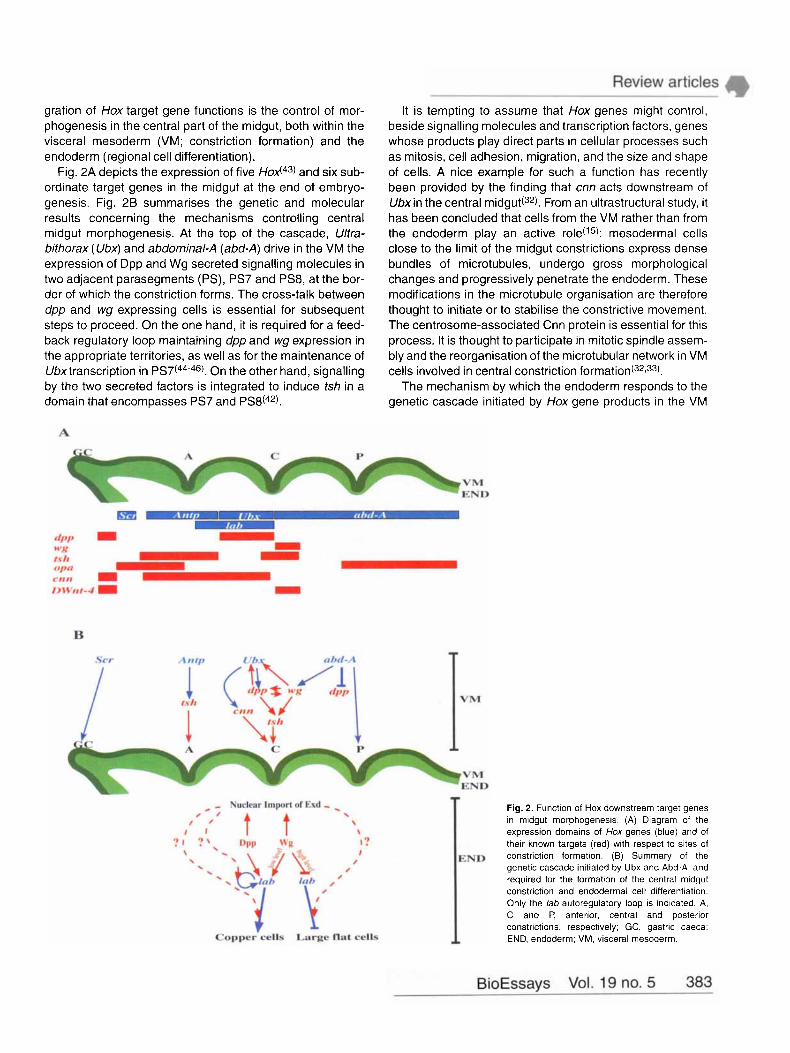
gration of Hox target gene functions is the control of mor- phogenesis in the central part of the midgut, both within the visceral mesoderm (VM; constriction formation) and the endoderm (regional cell differentiation).
Fig. 2A depicts the expression of five and six sub- ordinate target genes in the midgut at the end of embryo- genesis. Fig. 2B summarises the genetic and molecular results concerning the mechanisms controlling central midgut morphogenesis. At the top of the cascade, Ultra- bithorax (Ubx) and abdominal-A (abd-A) drive in the VM the expression of Dpp and Wg secreted signalling molecules in two adjacent parasegments (PS), PS7 and PS8, at the bor- der of which the constriction forms. The cross-talk between dpp and wg expressing cells is essential for subsequent steps to proceed. On the one hand, it is required for a feed- back regulatory loop maintaining dpp and wg expression in the appropriate territories, as well as for the maintenance of Ubxtranscription in PS7(44-46). On the other hand, signalling by the two secreted factors is integrated to induce tsh in a domain that encompasses PS7 and PS8(42).
It is tempting to assume that Hox genes might control, beside signalling molecules and transcription factors, genes whose products play direct parts in cellular processes such as mitosis, cell adhesion, migration, and the size and shape of cells. A nice example for such a function has recently been provided by the finding that cnn acts downstream of Ubxin the central m i d g ~ t ( ~ ~ ) . From an ultrastructural study, it has been concluded that cells from the VM rather than from the endoderm play an active role(15): mesodermal cells close to the limit of the midgut constrictions express dense bundles of microtubules, undergo gross morphological changes and progressively penetrate the endoderm. These modifications in the microtubule organisation are therefore thought to initiate or to stabilise the constrictive movement. The centrosome-associated Cnn protein is essential for this process. It is thought to participate in mitotic spindle assem- bly and the reorganisation of the microtubular network in VM cells involved in central constriction f o r m a t i ~ n ( ~ ~ , ~ ~ ) .
The mechanism by which the endoderm responds to the genetic cascade initiated by Hox gene products in the VM
A
VM END
B
VM
- Nuclear Import of Exd - . t t 0 / ,
/ / \ I 1 \
I? I
/ 0
0 /
0
Copper cells Large tlat cells
Fig. 2. Function of Hox downstream target genes in midgut rnorphogenesis. (A) Diagram of the expression domains of Hox genes (blue) and of their known targets (red) with respect to sites of
END constriction formation. (9) Summary of the genetic cascade initiated by Ubx and Abd-A, and required for the formation of the central midgut constriction and endodermal cell differentiation. Only the lab autoregulatory loop is indicated A, C and P, anterior, central and posterior constrictions, respectively; GC, gastric caeca: END, endoderm; VM, visceral mesoderm.

has been further substantiated by other findings. Signalling by Dpp and Wg, proteins secreted from the mesoderm to the endoderm, is required for the coordinated development of the two germ layers. Firstly, the pathway results in a differ- ential activation of the Hoxgene lab in a domain of the endo- derm lying between the anterior and the central constric- tions. The anteroposterior gradient of Lab, with a stronger accumulation at the posterior edge, determines the mor- phology of the ‘copper cells’, which are large cuprophilic cup-shaped cells located at a peripheral position within the central part of the midgut epithelium. High Lab levels induce large and highly differentiated copper cells, while low levels generate smaller and poorly differentiated ones(47). The graded expression of lab, and therefore the copper cell differentiation, depends on Wg signalling: the closer cells are to the Wg source (PS8 of the VM), the greater transcrip- tion of lab occurs and the stronger the copper cell pheno- type is expressed(48). Posterior to the copper cell domain, cells differentiate as long and thin cells, the so-called ‘large flat cells’. There, a higher Wg input has the opposite effect on lab transcription, down-regulating the gene and allowing large flat cells to differentiate. Secondly, it has recently been shown that Dpp and Wg signalling pathways also control the nuclear import of Extradenticle (Exd) protein in the central part of the midgut e n d ~ d e r m ( ~ ~ ) . exdencodes a homeobox- containing protein, homologous to the f b x l human proto- oncogene, which provides additional specificity to Hox pro- t e i n ~ ( ~ ~ ) . Thus, within the central part of the midgut, the nuclear translocation of Exd allows it to interact with Lab, an interaction that is presumably required for lab autoregula- tion and for the regulation of downstream target genes involved in endodermal cell differentiation.
These findings illustrate how Ubxand abd-A, through the control of dpp, wg and cnn downstream target genes, instruct cells of the VM and of the underlying endoderm for their commitment in the formation of the central midgut con- striction and regional cell differentiation. This constitutes an exciting example of how Hox-dependent coordination between cell signalling, transcription regulation and the dynamics of cellular morphology, can result in the induction of morphogenetic processes.
Stable maintenance of homeotic decisions Evidence is accumulating that the maintenance of Hoxgene expression and function in the appropriate metameric units requires the activity of distinct classes of genes. The first one corresponds to the Hox genes themselves, which con- trol both the levels and patterns of Hox gene expression through self-regulatory and cross-regulatory interac- t i o n ~ ( ~ ~ ) . The second class comprises the evolutionarily conserved Polycomb and trithorax groups of genes (Pc-G and trx-G). Pc-G and Trx-G proteins are responsible for the progressive opening of the Hox complex chromatin and maintenance throughout development of individual homeotic loci in a conformation state which is either open or
closed to tran~cription(~l). The third class corresponds to Hox downstream target genes. The best example of target genes involved in the control of Hox gene activity is the maintenance of Ubx expression, in PS7 of the VM, by dpp and wg, genes themselves controlled in this germ layer by Ubx and Abd-A, r e s p e ~ t i v e l y ( ~ ~ , ~ ~ ) .
Another target gene, the modifier of variegation mod, is involved in maintaining homeotic decisions. The products of such genes, from their roles in position effect variegation, participate in the formation and clonal inheritance of higher order chromatin structures(52). Unlike Pc-G and trx-G gene products, Mod does not regulate or maintain Hox gene expression. Acting downstream, it is instead thought to imprint locally specific chromatin structural states and thereby control genes critical for the process of morphogen- esis, in response to Hox gene activity(20). Since chromoso- mal proteins encoded by modifiers of variegation are believed to form multimeric complexes required for chro- matin assembly, mod is probably a pioneer example of a class of genes acting in concert to confer stable inheritance of developmental decisions, under instruction by Hox genes.
Regulation of Hox target genes: clues to the mechanisms of homeotic gene action Closely related in vitro DNA-binding of Hox proteins and common downstream targets HD-containing proteins display a poor in vitro DNA-binding specificity. They recognise nearly the same nucleotide sequence, which is rather ill-defined except that it contains an ATTA core with flanking nucleotides that are variable in nature(*). The same situation presumably occurs in vivo, since DNA protein cross-linking experiments performed on whole embryos have shown that Even-skipped and Futzi- tarazu, proteins that contain distantly related HDs, never- theless have the same DNA-binding activity(53).
One must wonder, considering the highly specific action of homeotic genes in the whole animal, about the functional significance of the poor DNA-binding specificity their prod- ucts exhibit. An interesting clue lies in the fact that, in many cases, one Hox protein can be replaced by another and still achieve repression. For example, Antp, Ubx and Abd-A, which control the differential expression of DWnt-4 in the VM, down-regulate the gene in PS5-6, PS7 and PS9-12, respectively. In mutant embryos, ectopic Antp can substitute for Ubx to achieve repression in PS7, and ectopic Ubx for Abd-A in PS9-1 2(34). Comparable situations, where the repression of a common target is alternately assumed by different Hox proteins, have been described: Antp and/or Ubx down-regulate n e r ~ f i ~ ~ ) and Ubx/Abd-A down-regulate unplugged24); Ubx can substitute for Abd-A, and Abd-A for Abdominal-B (Abd-B), in the silencing of con(27,29), Distal- less(11) and sca (I6) in abdominal segments. Analysis of the Antp P2 promoter repression by Ubx and Abd-A led to the

proposition that inactivation results from a steric blockade due to the fixation of Hox gene products on the ATTA core- containing binding sites, rather than from specific interac- tions with trans-activating factor(^)(^^). Two aspects in this model deserve to be underlined. Firstly, it provides a simple and attractive explanation for the interchangeability of Hox proteins in the repression of a common target gene. Sec- ondly, it integrates the poor nucleotide specificity of the HD with Hox gene specificity, showing that this property might explain some aspects of homeotic gene function during development.
Although Ubx and Abd-A can assume a similar function in somatic muscle and appendages s p e c i f i c a t i ~ n ( ~ ~ % ~ ~ ) , the interchangeability of Hox proteins for transcriptional activa- tion has so far not been reported at the level of downstream target gene regulation. For instance, Ubx activates dpp(21) and represses DWnt-4(34) in PS7 of the VM. In embryos deficient for Ubx, Antp becomes ectopically expressed in this PS and is able to replace Ubx in DWnt-4 silencing, but not in the activation of dpp. One cannot argue that these results reflect intrinsic properties of Antp and Ubx, with the first acting only as a repressor and the second only as an activator, since the two Hox proteins are known to act as either positive or negative trans-regulators. As Ubx and Antp recognise identical nucleotide sequences in vitro(8), it is conceivable that the two proteins bind the same sites on DNA, but that once bound, Antp is not able to promote tran- scription in a developmental context where Ubx does. Thus, the different effects of Antp and Ubx on the dpp enhancer activity do not necessarily result from differential DNA bind- ing. One could rather assume that transcriptional activation by Ubx requires conditions that repression by Antp does not, such as specific interaction with colfactors like Exd or with the basic transcription machinery. Understanding how Hox proteins interact with the transcription machinery obviously needs further work, especially on adapter proteins which are involved in the process.
Common downstream targets, functional hierarchy and phenotypic suppression Functional hierarchy and phenotypic suppression are closely related phenomena described from gain of Hox gene function experiments, performed in mouse and fly as
A general derepression of Hoxgenes is observed in Drosophila upon mutation of the Pc-G gene extra sexcombs (esc); in such embryos, cuticular structures that are nor- mally found in the Abd-B domain differentiate all over the body(58). Thus, when several Hox genes are simultaneously derepressed, the posteriormost acting one is prevalent in its effects, indicating the existence of a functional hierarchy among the Hox genes. In slightly different experiments, Hox genes were ubiquitously expressed under the control of a heat-shock promotor. In such conditions, homeotic transfor- mations are observed only in segments anterior to the nor- mal domain of action of the overproduced Hox proteid3). In
other words, posterior-expressed genes impose their mor- phogenetic programmes over those of resident anterior ones. This phenotypic suppression therefore follows the functional hierarchy between homeotic genes.
The molecular mechanisms underlying functional hierar- chy and phenotypic suppression are not well understood. However, some interesting indications are provided by look- ing at Hox target genes. On the one hand, target genes con- stitute models for a molecular analysis of these phenomena. For instance, mod is downstream of Ubx and Scr, with Ubx enhancing its transcription in the abdominal mesoderm and Scr activating the gene in the ectoderm of PS2(20). Upon Ubx ubiquitous expression, mod transcription occurs, as expected, at the same high level in the thoracic and abdomi- nal parts of the mesoderm. At the same time, mod tran- scripts are no longer detected in PS2 ectoderm, indicating that ectopic Ubx in this region prevents Scr from activating mod. The regulation of mod therefore exemplifies the func- tional hierarchy between Ubx and Scr and the phenotypic suppression of Scr by the Ubx gene product; it should be possible to dissect this situation in molecular terms by look- ing at the interactions between the two Hox transactivators and the response elements in the control region of the target gene.
On the other hand, the data listed in Table 1 indicate that most of the target genes are regulated by several Hox pro- teins. One can therefore hypothesise that Hox trans-regula- tors, depending on their relative abundance and affinity for the cognate DNA sequences, compete for the control of par- tially overlapping sets of target genes. Supporting this view, it has been shown that the homeotic transformations observed in Drosophila embryos expressing Ubx and Abd-B simultaneously under the control of a heat-shock promotor, depend on the ratio between the two overproduced pro- t e i n ~ ( ~ ~ ) . To account for the phenotypic suppression and functional hierarchy phenomena, the more posterior Hox product must be the more efficient in the competition. One possibility is that posterior prevalence relies on intrinsic dif- ferences in the DNA-binding activity, with a higher affinity of the posterior Hox proteins to common target sequences. Unexpectedly, in vitro studies have rather led to the opposite conclusion, indicating that the intrinsic affinity with DNA tends to decrease the more posteriorly the homeotic protein is expressed(60). This result favours a second possibility: that other genetic functions are required in vivo to assist homeotic genes. For instance, the interaction with specific cofactors may improve the ability of posterior Hox gene products to compete for the regulation of common down- stream target genes.
Understanding the functional specificity of Hox proteins As noted above, Hox proteins recognise similar nucleotide sequences in vitro, a property that stands in contrast with the diversity of the morphogenetic programmes they initiate in vivo. This paradox raises the question of the mechanisms

by which Hox proteins achieve highly specific action in the developing animal. The functional specificity of Hox proteins presumably results from a complex interplay of differential affinity, competition for common targets, subtle DNA-binding preferences@) and interactions with assisting cofactors. Strong evidence supporting the view that Hox cofactors con- tribute to the specificity of Hox proteins emerged from the genetic and molecular analyses of the control of dpp, the most thorough case study of regulation of a Hox down- stream target gene(2’ .Z2,6l).
dpp is transcribed under the positive control of Ubx and exd in PS7 of the VM and is repressed more posteriorly by abd-A. This expression pattern depends on the integrity of several enhancers, in which functional binding sites for Ubx, Abd-A and Exd proteins have been characterised. In vitro DNA-binding experiments indicate that Exd and Ubx bind cooperatively to minimal dpp enhancers(22162). Although the details of the Ubx/Exd interactions still need to be dissected, the process involves an evolutionary conserved motif, the hexapeptide (or YPWM motif), as well as residues within the Ubx HD and just C-terminal to it(50). Surprisingly, functional Ubx binding sites often do not contain the ATTA core sequence recognised in vitro by Hox proteins and strikingly differ from the Ubx consensus binding sequence defined in binding site selection experiments(*). This also holds for the only other well-studied homeotic responsive element, a 20- base-pair autoregulatory fragment of the mammalian Hoxb- 7 gene, whose activity also depends on Hox/Exd interac-
This highlights the fact that Hox binding sites defined in vifro might not accurately represent the sequences recognised in the living animal. The apparent paradox between the ill-defined DNA-binding and the high biological specificity of Hox proteins might well result from the absence of such cofactors.
At the molecular level, the analysis of the regulation of the dpp gene leaves unanswered several questions concerning the mode of action of homeotic proteins. Abd-A, which represses dpp expression in the posterior midgut, binds the same sequences in vifro as Ubx, within the identified enhancers. Although a homeotic cofactor contributing to the activation of dpp by Ubx was identified, why Ubx and Abd-A, which both interact with EX^@^), have opposite effects on dpp regulation was not explained. In the VM, exd is ubiqui- tously expressed and the protein seems to be located in the nuclei in all cells. Thus differential subcellular localisation, as is observed within the e n d ~ d e r m ( ~ ~ ) , cannot account for Exd interacting only with Ubx. Distinct but non-exclusive hypotheses can be formulated. Firstly, the same cofactor, Exd, might interact in different ways with Ubx or Abd-A, and with DNA, ultimately driving distinct regulatory effects. Sup- port for this idea has been obtained from molecular studies on Hoxb- 7 regulation(63). Functional Pbxl and Hoxb-1 bind- ing sites were identified within an enhancer responsible for Hoxb- 7 autoregulation; cooperative binding of Hoxb-1 with Pbxl (or Exd) was observed. In the same conditions, no
cooperation between Ubx and Pbxl (or Exd) occurs. These observations, corroborated by recent data(66), argue that the target sequence allows the cooperative interaction of some Hox/Pbxl or Exd combinations while it forbids others. Sec- ondly, some locally induced post-translational modification of the Exd protein (possibly controlled by Hox downstream target genes) might provide additional complexity. Finally, it may well be that other as yet unidentified proteins are required to distinguish Ubx from Abd-A function. Consistent with this view, several sequences, evolutionarily conserved in some cases, which do not contain a HD consensus bind- ing site nor bind in vitroto Ubx and Exd, influence the activity of the dpp enhancer^(^^,^').
The study of dpp, lab and Hoxb-7 regulation illustrates how important it is to get biologically relevant target sequences. When Exd/Pbx and Hox protein consensus sequences defined in vitro are used, Exd/Pbx and several hexapeptide-containing Hox proteins cooperatively bind DNA with almost no selectivity. In contrast, when relevant in vivo binding sites are used, selectivity in interaction is observed. Overall, it is now obvious that elucidation of the details of homeotic transcriptional specificity will emerge from the comparative analysis of a few well-analysed situ- ations. In each case, the determinant of specificity should be precisely mapped within the Hox protein, the interaction with cofactor(s) characterised in detail and the DNA sequence requirement established. The integration of these data should provide us with precise regulatory models which, when compared, should elucidate the molecular mecha- nisms underlying the specificity of action of homeotic genes.
Conclusions Despite the use of varied and distinct approaches, the search for Hox target genes has turned out to be a difficult task and only a few have been identified so far. The compila- tion presented in Table 1 deserves some general com- ments. Firstly, target genes fulfil essential functions in devel- opmental processes other than just being downstream targets of Hox genes. Most are expressed early in develop- ment and play critical roles before being subjected to homeotic control. This feature could explain, at least in part, the difficulty in designing genetic screens for target genes. As a corollary, looking for targets constitutes per se a genetic screen for new developmental functions. Secondly, a reduced number of the identified targets corresponds to realizator genes; most of them encode regulatory mol- ecules, either transcription factors or cell signalling proteins. The genetic cascade that controls pattern formation there- fore further extends the Hoxcomplex genes, before the acti- vation of realizator genes thought to provide the basic cellu- lar or ‘house keeping’ functions required for differentiation processes. Significant in this context is the role of signalling proteins in the connection of embryonic germ layers. Germ layers do indeed develop almost independently during early

embryogenesis, but become functionally interconnected as morphogenesis proceeds under the control of Hox proteins. The best example of such a mechanism concerns Wg and Dpp, molecules which are secreted from the mesoderm, that control lab activity and cell differentiation in the midgut endoderm. Thirdly, the majority of downstream targets are regulated by several Hox genes. Hox protein specificity in the whole animal could therefore be gained from an inter- play of relative abundance, differential affinity, interaction with cofactor proteins and competition for common targets. Highly significant in this context is the discovery that interac- tion with Exd/Pbx cofactor radically changes the binding properties of Hox proteins on dpp or Hoxb-I enhancers. This emphasises the importance of analysing bona fide in vivo Hox response elements.
The high degree of structural, and to some extent func- tional, conservation of homeotic genes during evolution indi- cates that different species use the same master regulatory molecules to control cell fate and body plan organisation. Not only have the Hoxmaster genes been conserved during the course of evolution, but so have the mechanisms con- trolling their expression and modulating their function: genes of the Pc-G and trx-G groups assume the stable maintenance of their expression domains, and the Exd/Pbx family of proteins assists Hox proteins to achieve specificity. A still open question is whether or not the conservation of the Hox ‘nexus’ during evolution includes downstream tar- get genes. In this respect, one can note that several Drosophila Hox downstream targets have been conserved. For example, the dpp, wg, nervy and empty spiracles genes have their counterparts in vertebrates, namely BMPs, Wnts, MGGB proto-oncogene and E mx 1-2, respectively(68), although nothing is yet known about their regulation by ver- tebrate Hoxgenes.
Despite such conservation of genes, the morphological outcomes of the developmental programmes that are instructed by homeotic genes are very different between metazoan phyla. The origin of pattern diversity should result from changes in Hoxgene activity. This could in principle be assumed at several levels. A possibility, which has received some experimental support, is that diversity finds its primary origin upstream of the Hox complex genes, by redesigning their expression d y n a m i c ~ ( ~ ~ 1 ~ ~ ) . Another and not exclusive possibility is that pattern diversity results from the introduc- tion, suppression or functional modification of Hox cofac- tors, so that homeotic proteins will select distinct combina- tions of downstream targets. Future work aiming to identify and analyse the regulation and function of more down- stream targets, not only in Drosophila but also in other species, should clarify many aspects of Hox complex gene function. It should also allow an investigation of the degree of conservation of the regulatory circuits controlling mor- phogenesis and highlight differences that might explain how evolutionarily conserved master genes control diverse mor- phogenetic events among species.
References 1 Lewis, E. 8. (1978). A gene complex controlling segmentation in Drosophila Nature 276, 565-570. 2 Kaufman, T. C., Lewis, R., and Wakimoto, B. (1980). Cytogenetic analysis of chromosome 3 in Drosophila melanogaster. the homeotic gene complex in polytene chromosome interval 84A-8. Genetics94. 11 5-1 33. 3 McGinnis, W. and Krumlauf, R. (1992). Homeobox genes and axial patterning. Cel/68,283-302. 4 Morata, G. (1993). Homeotic genes in Drosophila. Curr Opin. Genet. Dev 3. 606-614. 5 Krumlauf, R. (1994). Hox genes in vertebrate development. Cell78, 191-200 6 Garcia-Bellido, A. (1975). Genetic control of the wing disc development in Drosophila. In Cell Patterning, Ciba Foundation Symposium 29, 161 -1 78 7 Desplan, C., Theis, J. and O’Farrell, P. H. (1988). The sequence specificity of homeodomain-DNA interaction. Cell54, 1081 -1090. 8 Ekker, S. C. eta/. (1994). The degree of variation in DNA sequence recognition among four Drosophila homeotic proteins EMBOJ. 13, 3551-3560. 9 Krasnow, M., Saffman, E. E., Kornfeld, K. and Hogness, D. S. (1989). Transcriptional activation and repression by Ultrabithorax proteins in cultured Drosophila cells. Cell57. 1031 -1 043. 10 Panzer, S., Weigel, D. and Beckendorf, S. K. (1992). Organogenesis in Drosophila. embryonic salivary gland determination is controlled by homeotic and dorso-ventral patterning genes. Development1 14.49-57. 11 Vachon, G., Cohen, B., Pfeifle, C., McGuffin, M. E., Botas, J. and Cohen, S. M. (1992). Homeotic genes of the Bithorax complex repress limb development in the abdomen of the Drosophila embryo through the target gene Distal-less. Cell 71, 437-450. 12 O’Hara, E., Cohen, B., Cohen, S. M. and McGinnis, W. (1 993). Distal-less is a downstream gene of Deformed required for ventral maxillary identity. Development 117,847-856. 13 Jones, B. and McGinnis, W. (1993). The regulation of empty spiracles by Abdominal-6 mediates an abdominal segment identity function. Genes Dev 7. 229-240. 14 Immergluck, K., Lawrence, P. A. and Bienz, M. (1990). Induction across germ layers in Drosophila mediated by a genetic cascade. Cell62, 261 -268. 15 Reuter, R. and Scott, M. P. (1990). Expression and function of the homeotic genes Antennapedia and Sex combs reduced in the embryonic midgut of Drosophila. Development 109, 289-303. 16 Graba, Y. eta/. (1992). Homeoticcontrol in Drosophila: the scabrousgene is an in vivo target of Ultrabithorax proteins. EMBOJ. 9. 3375-3384 17 McCormick, A,, Core, N., Kerridge, S. and Scott, M. P. (1995). Homeotic response elements are tightly linked to tissue specific elements in a transcriptional enhancer of the teashirtgene. Development121,2799-2812. 18 Hinz, U., Wolk, A. and Renkawitz-Pohl, R. (1992) Ultrabithorax is a regulator of /33-lubulin expression in the Drosophila visceral mesoderm. Development 116. 543-554. 19 Cimbora, D., M. and Sakonju, S. (1995). Drosophila midgut morphogenesis requires the function of the segmentation gene odd-paired. Dev. B i d 169. 580-95. 20 Graba, Y., Laurenti, P., Perrin, L., Aragnol, D. and Pradel, J. (1994). The suppressor of PEV modulo gene acts downstream selector genes and is required for morphogenesis. Dev Biol. 166, 704-715. 21 Capovilla, M., Brandt, M. and Botas, J. (1994). Direct regulation of decapentaplegic by Uitrabithorax and its role in Drosophila midgut morphogenesis. Cell76.461-475. 22 Sun, B., Hursh, D., Jakson, D. and Beachy, P. A. (1995). Ultrabithorax protein is necessary but not sufficient for full activation of decapenfaplegic expression in the visceral mesoderm. EMBO. J. 14,520-535. 23 Wagner-Bernholz, J. T., Wilson, C., Gibson, G., Schuh, R. and Gehring, W. (1991). Identification of target genes of the homeotic gene Antennapedia by enhancer detection. Genes Dev. 5,2467-2480. 24 Chiang C., Young, K. E. and Beachy, P. A. (1995). Control of Drosophila tracheal branching by the novel homeodomain gene unplugged. a regulatory target for genes of the bithorax complex. Development 121,3901-3912. 25 Feinstein, P. G., Kornfeld, K., Hogness, D. S. and Mann, R. S. (1995) Identification of homeotic target genes in Drosophila melanogaster including nervy, a proto-oncogene homologue. Genetics 140,573-586. 26 Mastick, G. S., McKay, R., Oligino, T., Donovan, K. and Lopez A. J. (1995). Identification of target genes regulated by homeotic proteins in Drosophila melanogaster through genetic selection of Ultrabithorax protein binding sites in yeast. Genetics 139,349-363. 27 Gould, A. P., Brookman, J. J., Strutt, D. 1. and White, R. A. H. (1990). Targets of homeotic gene control in Drosophila Nature348.308-312. 28 Tomotsume, D., Shogi, H., Wakamutsu, Y., Kondoh, H. and Takahashi, N. (1993). A mouse homologue of the Drosophila tumor suppressor gene l(2)gl controlled by HOX-C8 in vivo. Nature365 69-72. 29 Gould, A. P. and White, R. A. H. (1992). Connectin a target of homeotic gene control in Drosophila. Developmentll6, 1163-1 174

30 Strutt, D. I. and White, R. A. H. (1994) Characterisation of T48. a target of homeotic gene regulation in Drosophila embryogenesis. Mech. Dev. 46,27-39. 31 Brookman, J. J.,Toosy, A.T., Shashidhara, L. S. and White, R. A. H. (1992). The 412 retrotransposon and the development of gonadal mesoderm in Drosophila. Development 11 6, 1 185-1 192. 32 Heuer J. G., Li, K. and Kaufman, T. C. (1 995) The Drosophila homeotic target gene centrosomin encodes a novel protein with leucine zippers and maps to a genomic region required for midgut morphogenesis. Development 121, 3861 - 3876. 33 Li, K. and Kaufman, T. C. (1996). The homeotic target gene centrosomin encodes an essential centrosomal component. Cel/85,585-596. 34 Graba, Y. eta/. (1 995). DWnt-4. a new Drosophila Wntgene acts downstream homeotic complex genes in the visceral mesoderm. Development121,209-218. 35 Kimble, M., Dettman, R.,W. and Raff, E. C. (1990). The 133 tubulin gene of Drosophila melanogaster is essential for viability and fertility. Genetics 126, 991- 1005. 36 Meadows, L. A,, Gell, D., Broadie, K., Gould, A. P. and White, R. A. H. (1994). The cell adhesion molecule Connectin and the development of the Drosophila neuromuscular systhem. J. CellSci. 107,321-328. 37 Nose, A,, Mahajan, V. 8. and Goodman, C. S. (1992). Connectin : A homophilic cell adhesion molecule expressed in a subset of muscles and the motoneurons that innervate them in Drosophila. Cel/70,553-567. 38 Nose, A,, Takeichi, M. and Goodman, C. S. (1994). Ectopic expression of connecbn reveals a repulsive function during growth cone guidance and synapse formation. Neuron 13, 525-539. 39 Andrew, D. and Scott, M. P. (1992). Downstream of the homeotic genes. The NewBiologist4,5-15. 40 Baker, N. E., Mlodzik, M. and Rubin, G. M. (1990). Spacing differentiation in the developing Drosophila eye: A fibrinogen-related lateral inhibitor encoded by scabrous. Science 250, 1370-1377. 41 Miyamoto, H. et a/. (1995). canoe encodes a novel protein containing a GLGFiDHR motif and functions with Notch and scabrous in common developmental pathways in Drosophila. Genes Dev. 9. 612-25. 42 Mathies, L. D., Kerridge, S. and Scott, M. P. (1994). Role of the teashirfgene in Drosophila midgut morphogenesis: secreted proteins mediate the action of homeotic genes. Development 120,2799-2809. 43 Tremml, G. and Bienz, M. (1989). Homeotic gene expression in the visceral mesoderm of Drosophila embryos. EMBO J. 8, 2677-85. 44 Thiiringer, F., Cohen, S. M. and Bienz, M. (1993). Dissection of an indirect autoregulatory response of a homeotic Drosophila gene. EM60 J. 12,2419-2430. 45 Hursh, D. A,, Padgett, R. W. and Gelbart, W. (1993). Cross regulation of decapentaplegic and Ultrabithorax transcription in the embryonic visceral mesoderm of Drosophila. Development 117,121 1-1 222. 46 Xiang , Y., Hoppler, S., Eresh, S. and Bienz, M. (1996). decapentaplegic, a target gene of the Wingless signalling pathway in the Drosophila midgut. Developmenf 122,849-858. 47 Hoppler, S. and Bienz, M. (1994). Specification of a single cell type by a Drosophila homeotic gene. Cell76. 689-702. 48 Hoppler, S. and Bienz, M. (1995). Two different threshold of Wingless signalling with distinct developmental consequences in the Drosophila midgut. EMBOJ 14,5016-5026. 49 Mann R. S. and Abu-Shaar, M. (1996). Nuclear import of the homeodomain protein Extradenticle in response to Wg and Dpp signalling. Nature 383. 630-633. 50 Mann, R. S. and Chan, S. K. (1996) Extra specificity from extradenticle: the partnership between Hox and PBXiEXD homeodomain proteins. Trends Genet 12,258-262. 51 Orlando , V. and Paro, R. (1995). Chromatin multiprotein complexes involved in the maintenance of transcription patterns Cur[ Opin. Genet. Dev. 5, 174-179. 52 Garzino, V. eta/. (1992). Cell lineage specific expression of modulo. a dose- dependent modifier of variegation in Drosophila. EM60 J. 11,4471 -4479. 53 Walter, J., Dever, C. A. and Biggin, M. D. (1994). Two homeodomain proteins bind with similar specificity to a wide range of DNA sites in Drosophila embryo. Genes Dev 8.1678-1692. 54 Appel, B. and Sakonju, S. (1993) Cell-type-specific mechanisms of
transcriptional repression by the homeotic gene products Ubx and Abd-A in Drosophilaembryos. EM60 J. 12, 1099-1 109. 55 Michelson, M. (1994). Muscle pattern diversification in Drosophila is determined by the autonomous function of homeotic genes in the embryonic mesoderm. Development 120,755-768. 56 Casares, F., Calleja, M. and Sanchez-Herrero, E. (1 996). Functional similarity in appendage specification by the Ulfrabithorax and abdominal-A Hox genes.
57 Duboule, D. and Morata, G. (1994). Colinearity and functional hierarchy among genes of the homeotic complexes Trends Genet 10,358-364 58 Struhl, G. (1983). Role of the esc gene product in ensuring the selective expression of segment specific homeotic genes in Drosophila. J. Embryo/ exp. Morph. 76,297-331. 59 Lamka, M. L., Boulet, A. M. and Sakonju, S. (1992). Ectopic expression of Ubx and Abd-B proteins during Drosophila embryogenesis. competition. not a functional hierarchy, explains phenotypic suppression. Development 116, 841 - 854. 60 Pellerin, I., Schnabel, C., Catron, K. M. and Abate, C. (1994). Hox proteins have different affinities for a consensus DNA site that correlate with the position of their genes on the Hox cluster. Mol. Cell Biol. 14,4532-4545. 61 Rauskolb, C. and Wieschaus, E. (1994). Coordinate regulation of downstream genes by extradenticle and a homeotic selector proteins EM60 J 13, 3561-3569. 62 Chan, S. K., Jaffe, L., Capovilla, M., Botas, J. and Mann, R. S. (1994). The DNA-binding specificity of Ultrabithorax is modulated by cooperative interactions with Extradenticle. another homeoprotein. Cell78. 603-61 5. 63 Popperl, H. eta/. (1995). Segmental expression of Hoxb-1 is controlled by a highly conserved autoregulatory loop dependent upon ExdiPbx. Cell 81. 1031- 1042. 64 Chan, S. K., Popperl, H., Krumlauf, R. and Mann, R. S. (1996) An Extradenticle induced conformational change in a Hox protein overcomes an inhibitory function of the conserved hexapeptide motif. EMBO J. 15, 2476-2487. 65 Van Dijk, M. A. and Murre, C. (1994). Extradenicle raises the DNA-binding specificity of homeotic selector gene products. Cell78,617-624 66 Chan, S. K. and Mann, R. S. (1 996). A structural model for a homeotic protein- extradenticle-DNA complex accounts for the choice of Hox protein in the heterodimer. Proc. Natl Acad. Sci. USA 93, 5223-5228. 67 Manak, J. R., Mathies, L. D. and Scott, M. P. (1994). Regulation of a decapentaplegic midgut enhancer by homeotic proteins. Development 120, 3605- 3619. 68 Simeone, A. M., Gulisano, M., Acampora, A., Stornaiuolo, A,, Rambaldi, M. and Boncinelli, E. (1992). Two vertebrate homeobox genes related to the Drosophila empty spiracles gene are expressed in the embryonic cerebral cortex. EMBO. J. 11,2541 -2550. 69 Carroll, S. B. (1 994). Developmental regulatory mechanisms in the evolution of insect diversity. Development Supplement, 21 7-223. 70 Burke, A. C., Nelson, C. E., Morgan, 8. A. and Tabin, C. (1 995). Hox genes and the evolution of vertebrate morphology. Development 121, 333-346. 71 Kuhn, D. T. and Cunningham, G. M. (1977) Aldehyde oxydase distribution in haltere discs in homeotic bithorax mutants in Drosophila melanogaster. Mol Gen Genet. 150,37-42. 72 Kuhn, D. T. and Cunningham, G. M. (1986). lsocitrate dehydrogenase in Drosophila melanogaster imaginal discs: pattern development and alteration by homeotic mutant genes. Dev. Genet. 7,21-34.
EM60 J. 15,3934-3942.
Yacine Graba, Denise Aragnol and Jacques Pradel* are at the Laboratoire de Genetique et de Biologie du Developpernent, lnstitut de Biologie du Developpernent de Marseille, Parc Scientifique de Lurniny, CNRS Case 907, 13288 Marseille cedex 9, France. *Author for correspondence.