does motoneuron adaptation contribute to muscle fatigue
TRANSCRIPT

INVITED REVIEW ABSTRACT: To help reduce the gap between the cellular physiology ofmotoneurons (MNs) as studied “bottom-up” in animal preparations and the“top-down” study of the firing patterns of human motor units (MUs), thisarticle addresses the question of whether motoneuron adaptation contrib-utes to muscle fatigue. Findings are reviewed on the intracellularly recordedelectrophysiology of spinal MNs as studied in vivo and in vitro using animalpreparations, and the extracellularly recorded discharge of MUs as studiedin conscious humans. The latter “top-down” approach, combined with kineticmeasurements, has provided most of what is currently known about theneurobiology of muscle fatigue, including its task and context dependencies.It is argued that although the question addressed is still open, it should nowbe possible to design new “bottom-up” research paradigms using animalpreparations that take advantage of what has been learned with the use ofrelatively noninvasive quantitative procedures in conscious humans.
Muscle Nerve 35: 135–158, 2007
DOES MOTONEURON ADAPTATION CONTRIBUTETO MUSCLE FATIGUE?
MICHAEL A. NORDSTROM, PhD,1 ROBERT B. GORMAN, PhD,2 YIANNIS LAOURIS, MD, PhD,3
JOHN M. SPIELMANN, PhD, MD,4 and DOUGLAS G. STUART, PhD5
1 Discipline of Physiology, School of Molecular and Biomedical Science, University of Adelaide, Adelaide,South Australia, Australia
2 Advanced Bionics, Valencia, California, USA3 Department of Neural Control, Cyprus Neuroscience and Technology Institute, Nicosia, Cyprus4 White Oak Anesthesiology Associates, Buffalo Hospital, Buffalo, Minnesota, USA5 Department of Physiology, University of Arizona College of Medicine, Tucson, Arizona 85724-5051, USA
Accepted 24 October 2006
It is generally recognized that muscle fatigue (here-after referred to as fatigue) denotes an increase inthe perceived effort necessary to exert a desiredforce and an eventual inability to produce thisforce.52 Both conceptually and operationally, thisdefinition accommodates another well-accepted def-inition of fatigue as any reduction in the force-gen-erating capacity of the total neuromuscular systemregardless of the force required in any given situa-tion.14 For both definitions, it is necessary to add thatrest reverses fatigue.61
It has been known for at least a century thatfatigue can be caused by an impairment of numer-ous physiological processes distributed throughout
the central nervous system (CNS) and peripheralneuromuscular system.62 Among these processes, farmore is known about those occurring external to theCNS, although much is still to be learned aboutthem.33 The case for CNS processes is no less com-pelling.45,61,144 Among these, Gandevia61 recently dis-cussed an intrinsic property of motoneurons termedspike-frequency adaptation (hereafter termed adap-tation). Experimentally, it involves a decline in firingrate in the presence of a sustained, constant depo-larizing pressure exerted on the motoneuron (MN)by intracellular (IC) current injection. Gandevia cau-tioned that the possibility of MN adaptation influ-encing fatigue is diminished when extrinsic influ-ences are exerted on the MN, particularly byconductance changes that produce a persistent in-ward (depolarizing) current (PIC). This creates aplateau potential in the MN that may offset its adap-tation.
The purpose of this article, which was written foran interdisciplinary movement neuroscience reader-ship, is to evaluate the possibility that MN adaptationcontributes to muscle fatigue from the perspective ofan emerging need in movement neuroscience. It is
Abbreviations: 5HT, serotonin; AHP, afterhyperpolarization; AP, adductorpollicis muscle; CNS, central nervous system; EC, extracellular; EHL, exten-sor hallucis muscle; EMG, electromyography/electromyographic; IC, intracel-lular; MN, motoneuron; MVC, maximum voluntary contraction; MU, motorunit; NE, norepinephrine; PIC, persistent inward current; SO, slow-twitchoxidative muscle fiber and motor unitKey words: motoneuron adaptation; muscle fatigue; spike-frequency adap-tation; spinal motoneuron dischargeCorrespondence to: D.G. Stuart; e-mail: [email protected]
© 2006 Wiley Periodicals, Inc.Published online 28 December 2006 in Wiley InterScience (www.interscience.wiley.com). DOI 10.1002/mus.20712
Motoneurons and Muscle Fatigue MUSCLE & NERVE February 2007 135

important to reduce the ever-widening gap betweenthe cellular physiology of MNs, as studied “bottom-up” in surgically reduced in vitro and in vivo animalpreparations, and the “top-down” study of the firingpatterns of the motor units (MUs) supplied by MNsin both intact conscious animals and the human. Ascautioned by Loeb,119 it is important that the formerapproach using animal preparations not result in“an almost random collection of bits of data in mediasres” which are often then used “to extrapolate to‘principles’ having no necessary place in the greaterscheme of the organism.” Conversely, the latter stud-ies using humans provide the possibility of learningthe control strategies for a given type of movementand, as such, provide a challenge to the designers ofreductionist studies on animal preparations under-taken at the cellular/molecular level of analysis.
For the present purposes, we are unaware of anystudies in which MU firing patterns have been stud-ied during the voluntary (operant-trained), fatigue-inducing contractions of nonhuman subjects. Thus,the gap we are addressing is primarily between ICmicroelectrode work undertaken on surgically re-duced animal preparations and tissue and extra-cellular (EC) macroelectrode electromyographic(EMG) recordings made in intact humans. The lat-ter approach has provided most of what is currentlyknown about the neurobiological aspects of musclefatigue and its context and task dependencies.
Ideas on our topic appeared in abstracts sometime ago101,102,154 but much has developed sincethose presentations.79,80,82,103,130,142,143
CELLULAR MECHANISMS UNDERLYINGMOTONEURON ADAPTATION AND PERSISTENTINWARD CURRENTS
First, it is helpful to review the ionic conductancesunderlying the mammalian MN action potential andits descending CNS control, MN adaptation, PICsand their functional significance, and then the ex-tent to which the latter can offset adaptation. Al-though the emphasis below is almost exclusively onmammalian spinal MNs, mention is briefly made ofresults obtained on mammalian hypoglossal MNsand the spinal MNs of nonmammalian vertebrateswhen the results are particularly relevant to this ar-ticle’s intent.
MN Action Potential Conductances. Figure 1 is anupdate and modification of a previously published84
schematic using conventional terminology for spinalMNs.142 It shows the conductances mediated by theion channels now known to generate the spike and
afterhyperpolarization (AHP) phases of the genericmammalian MN action potential, including (pre-sumably) that of humans. Not shown is the slow (s)AHP that can occur in other types of mammalianneurons.146 For the present purposes, note that thegeneration of a single action potential generationinvolves at least 13 conductances. There are evenmore conductances that become active during andimmediately after repetitive MN discharge.
Ionotropic vs. Metabotropic Actions. The Figure 1conductances are influenced by two types of postsyn-aptic action on MNs: ionotropic and metabotropiceffects. Eccles and the McGeers51,128 coined theseterms. Eccles’ group showed ionotropic (voltage-regulated) effects on mammalian spinal MNs intheir classic articles of the early 1950s.28 They aretriggered by excitatory and inhibitory postsynapticpotentials, which operate directly on ion channels todepolarize and hyperpolarize their parent mem-brane, respectively. Metabotropic effects were first
FIGURE 1. Conductances underlying the spike and AHP of theaction potential of the generic adult mammalian MN. They areshown in bold italics if known to be the primary contributor to theaction-potential phase in question. Substances in parenthesesare the better-known blockers of these conductances. For anearlier version of this figure, see Hornby et al.84
136 Motoneurons and Muscle Fatigue MUSCLE & NERVE February 2007

observed over two decades later.72,115 They involvesynaptic or hormonal inputs acting in many cases viaG-protein–coupled receptors embedded in themembrane. They set in motion “second messenger”IC cascades that can then indirectly modify the be-havior of the voltage-sensitive ion channels. In thesecond (1987) edition of the McGeer et al. vol-ume,128 metabotropic action was extended to in-clude second-messenger systems that could translo-cate to the neuron’s nucleus and act on DNAtranscription. This latter effect, which is fundamen-tal to the study of learning and memory within theCNS and MN plasticity, is beyond the purview of thisarticle, however.
Descending and Sensory Feedback Effects on MN Con-ductances. In anesthetized in vivo mammalian prep-arations, the postsynaptic effects of descending com-mand signals and sensory input via spinal reflexpathways on MNs are largely ionotropic.21 Whendescending command signals and sensory feed-back are studied in unanesthetized (largely decere-brate) preparations, however, a different pictureemerges.80,82 A variety of neuromodulators (i.e.,agents that trigger metabotropic effects) are nowknown to act on spinal MNs, including the mono-amines serotonin (5HT) and norepinephrine (NE),thyroid-releasing hormone, adenosine, substance P,and several other neuropeptides. In addition, neu-rotransmitters first shown to have ionotropic effectsare now known to have additional metabotropic ef-fects (e.g., acetylcholine, glutamate, GABA82).Therefore, as emphasized by Heckman,78 “. . .thefiring pattern produced by the ionotropic inputfrom a motor command or a reflex can be dramati-cally different, depending on the level of neuro-modulatory input to the MN.”
Activation of Neuromodulators during Movement.The most studied descending tract neuromodulatorsthat affect spinal MN conductances are 5HT and NE.These monoamines are released in the spinal cordlargely by reticulospinal tract axons whose cells oforigin are in the brainstem’s diffuse raphe nuclei for5HT83,125 and the much more localized locus coer-uleus for NE.5
Figure 2 shows schematically the concept ofHeckman et al.82 that the command for voluntarymovement has two components that operate tempo-rally in parallel after their activation by a generalcommand from the sensorimotor cortex (and pre-sumably its connections with the basal ganglia, cer-ebellum, thalamus, and brainstem).
As reviewed in several chapters in Mori et al.132 (seealso Lemon and Griffiths113), the Figure 2 conceptproposes that there is a specific command that
emerges from the motor cortex to engage largely cor-ticospinal and selected reticulospinal (and often otherextrapyramidal) tract axons. The latter’s spinal effectsare largely on spinal interneuronal circuits that, inturn, produce ionotropic effects in spinal MNs. In par-allel, the concept proposes a concomitant excitabilitycommand, which presumably emerges from the senso-rimotor cortex, basal ganglia, and midbrain to actlargely on other reticulospinal tract axons that exertboth direct and indirect (via other spinal interneuro-nal circuitry) metabotropic (including 5HT and NE)effects on the same MNs. Thus, this concept proposesthat the specific command for voluntary movement ismediated in the spinal cord largely by (1) corticospinaltract axons and (2) non-5HT and non-NE componentsof reticulospinal and other descending extrapyramidalpathways. In contrast, (3) the descending excitabilitycommand is mediated in the spinal cord largely by wayof 5HT and NE components of the reticulospinal tract.Recent work suggests, however, that at least somemetabotropic input to MNs arises in the spinal corditself (Miles and Brownstone, unpublished).
The excitability command is also activated duringcontrolled near-natural or fictive (in paralyzed ani-mals) locomotion of the high decerebrate cat, suchas to also adjust cardiorespiratory activity and re-
FIGURE 2. Descending and sensory feedback control of MNsduring voluntary movement.82 Left-side connections show a cor-ticospinal command emanating from the motor cortex (and otherbrain regions) and sensory input, which converge largely onspinal interneuronal circuits, which exert ionotropic effects onMNs. Right-side connections show a concomitant excitabilitycommand via reticulospinal and other extrapyramidal tracts thatexerts metabotropic effects on spinal INs and MNs. The excit-ability pathways may activate selected spinal INs, which exertother metabotropic effects on MNs by activating the latter’smetabotropic (m) glutamate and muscarinic ACh receptors.Question marks (?) and dashed arrows (- - - 3) indicate under-studied possibilities. Reprinted from Heckman et al.82 with per-mission of Elsevier.
Motoneurons and Muscle Fatigue MUSCLE & NERVE February 2007 137

flexes to the demands of the motor task.166 For suchcontrolled locomotion, electrical stimulation is ap-plied to a specific diencephalic, midbrain, or cere-bellar site.74,133,160 These comments on controlledlocomotion serve to emphasize that the Figure 2schema is a component of an even broader CNSfunction: somatovisceral integration during the elab-oration of movement.
When evaluating the Figure 2 concept it is nec-essary to keep in mind that its applicability to rapidlyalternating voluntary movements is yet to be deter-mined. Nonetheless, for the present discourse onmuscle fatigue it warrants serious consideration.
An emerging literature80,82 on the behavior offreely moving animals suggests that the intensity andhence force-producing capacity of a specific motorbehavior is determined by both the depolarizationpressure exerted on spinal INs and MNs by ionotropiceffects induced by specific descending motor com-mands and the intensity/prevalence of metabotropiceffects attributable to 5HT, NE, and other neuromodu-lators.76,92,145 Like the voluntarily commanded iono-tropic effects on spinal MNs, the metabotropic effectsare task dependent, such that, for example, the preva-lence/intensity of both 5HT92 and NE145 effects pre-sumably exerted in the spinal cord vary according tothe motor and allied psychological behavior of the testsubject (for additional complexities, which are stillemerging, the reader is referred elsewhere6,57,67). Anexample of the powerful behaviorally/task-dependentneuromodulatory effect is the 5HT-induced openingof the spinal MN’s CaL (specifically Cav1.3) channel toproduce a PIC-based plateau potential even when theionotropic depolarizing pressure is less than the MN’sthreshold for repetitive discharge.87 The functional sig-nificance of this action, which occurs largely, but notexclusively, in the MN’s dendrites22,81 is discussed be-low.
Finally, metabotropic effects on all of the Figure 1conductances are presumably influenced to varyingextents during the repetitive discharge associated withfatigue. With the exception of the remarkable findingsof Fornal et al.,57 however, which are discussed below,very little specific information is available about suchchanges during fatigue, including the fatigue-taskdependencies of such presumed changes.
MN Adaptation. This phenomenon has usually beenstudied when MNs are responding to sustained stim-ulation for �30 s. Functionally, however, it is just asimportant to consider what is known about adapta-tion to repetitive stimulation.
Adaptation to Sustained Stimulation. Figure 3shows schematically that MN adaptation to sustained
stimulation has at least two phases, and possibly sev-eral others that may be animal species–specific (e.g.,cat vs. turtle) or cell type–specific (e.g., spinal vs.hypoglossal MN).11,12,28,70,130,143 For the present pur-poses, the focus is on the initial and late adaptationof spinal MNs as studied in adult cat reduced in vivopreparations.29,96,97,101,102,116,155 MN adaptation insuch preparations is likely to resemble that in thehuman. Reference is also made to the adaptation ofspinal MNs studied in spinal slice in vitro prepara-tions of the young mouse130 and adult turtle70 be-cause their adaptation patterns are at least qualita-tively like those in the cat.87 It is also likely thatmechanisms underlying the adaptation of superiorcolliculus cells of the adult guinea pig117,118 andhypoglossal MNs of the young rat147,148,176 give adegree of insight into analogous properties of spinalMNs in the adult cat and human.
Initial Adaptation to Sustained Stimulation. In theadult cat, this phase of spinal MN adaptation occurswithin the first second of repetitive MN dischargeand it has usually been attributed to AHP summa-tion. This is thought to be brought on by increasedCa2� entry (the 1[Ca]i in Fig. 3) leading to agreater activation of the Ca2�-dependent K� chan-nel responsible for the medium AHP: i.e., primarilythe KCa-SK conductance (reviews of the early litera-ture supporting this possibility are provided else-where7–9,71,91,98).
In contrast to the above work on adult cat MNs, arecent carefully undertaken in vitro study on spinalMNs of the young mouse showed that a change in themedium AHP could not account for initial adaptation.Rather, it was best explained by a slow inactivation ofNaT, the fast, transient Na� conductance.28,130 Interest-ingly, this mechanism was suggested previously for thelate, but not initial, adaptation of spinal MNs in theadult cat.116 At this stage, it is not clear if the timecourse of initial and late adaptation is similar for thespinal MNs of the adult cat and young mouse. Thelatter may have an initial adaptation that lasts �100 msand a late adaptation that is fully expressed within thefirst second of repetitive discharge (see fig. 6 in Miles etal.130). For the moment, then, it seems best to considerthat the mechanism underlying the initial adaptationof mammalian spinal MNs is still a partially open issue,with the evidence favoring some species-dependentcombination of AHP summation and reduced NaT
conductance.Later Phases of Adaptation to Sustained Stimulation.
Figure 3 shows that more mechanisms have beenproposed to underlie the later phases of adaptationthan is the case for initial adaptation. For adult catMNs, the most frequently emphasized ones are (1)
138 Motoneurons and Muscle Fatigue MUSCLE & NERVE February 2007
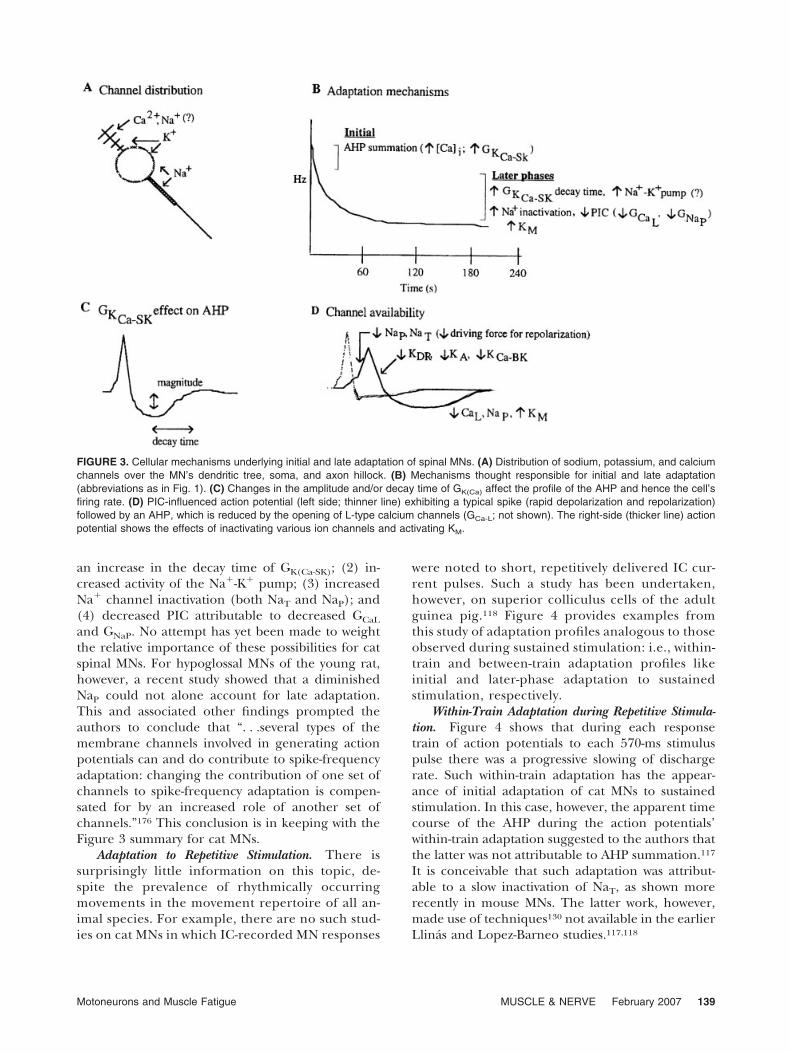
an increase in the decay time of GK(Ca-SK); (2) in-creased activity of the Na�-K� pump; (3) increasedNa� channel inactivation (both NaT and NaP); and(4) decreased PIC attributable to decreased GCaL
and GNaP. No attempt has yet been made to weightthe relative importance of these possibilities for catspinal MNs. For hypoglossal MNs of the young rat,however, a recent study showed that a diminishedNaP could not alone account for late adaptation.This and associated other findings prompted theauthors to conclude that “. . .several types of themembrane channels involved in generating actionpotentials can and do contribute to spike-frequencyadaptation: changing the contribution of one set ofchannels to spike-frequency adaptation is compen-sated for by an increased role of another set ofchannels.”176 This conclusion is in keeping with theFigure 3 summary for cat MNs.
Adaptation to Repetitive Stimulation. There issurprisingly little information on this topic, de-spite the prevalence of rhythmically occurringmovements in the movement repertoire of all an-imal species. For example, there are no such stud-ies on cat MNs in which IC-recorded MN responses
were noted to short, repetitively delivered IC cur-rent pulses. Such a study has been undertaken,however, on superior colliculus cells of the adultguinea pig.118 Figure 4 provides examples fromthis study of adaptation profiles analogous to thoseobserved during sustained stimulation: i.e., within-train and between-train adaptation profiles likeinitial and later-phase adaptation to sustainedstimulation, respectively.
Within-Train Adaptation during Repetitive Stimula-tion. Figure 4 shows that during each responsetrain of action potentials to each 570-ms stimuluspulse there was a progressive slowing of dischargerate. Such within-train adaptation has the appear-ance of initial adaptation of cat MNs to sustainedstimulation. In this case, however, the apparent timecourse of the AHP during the action potentials’within-train adaptation suggested to the authors thatthe latter was not attributable to AHP summation.117
It is conceivable that such adaptation was attribut-able to a slow inactivation of NaT, as shown morerecently in mouse MNs. The latter work, however,made use of techniques130 not available in the earlierLlinas and Lopez-Barneo studies.117,118
FIGURE 3. Cellular mechanisms underlying initial and late adaptation of spinal MNs. (A) Distribution of sodium, potassium, and calciumchannels over the MN’s dendritic tree, soma, and axon hillock. (B) Mechanisms thought responsible for initial and late adaptation(abbreviations as in Fig. 1). (C) Changes in the amplitude and/or decay time of GK(Ca) affect the profile of the AHP and hence the cell’sfiring rate. (D) PIC-influenced action potential (left side; thinner line) exhibiting a typical spike (rapid depolarization and repolarization)followed by an AHP, which is reduced by the opening of L-type calcium channels (GCa-L; not shown). The right-side (thicker line) actionpotential shows the effects of inactivating various ion channels and activating KM.
Motoneurons and Muscle Fatigue MUSCLE & NERVE February 2007 139
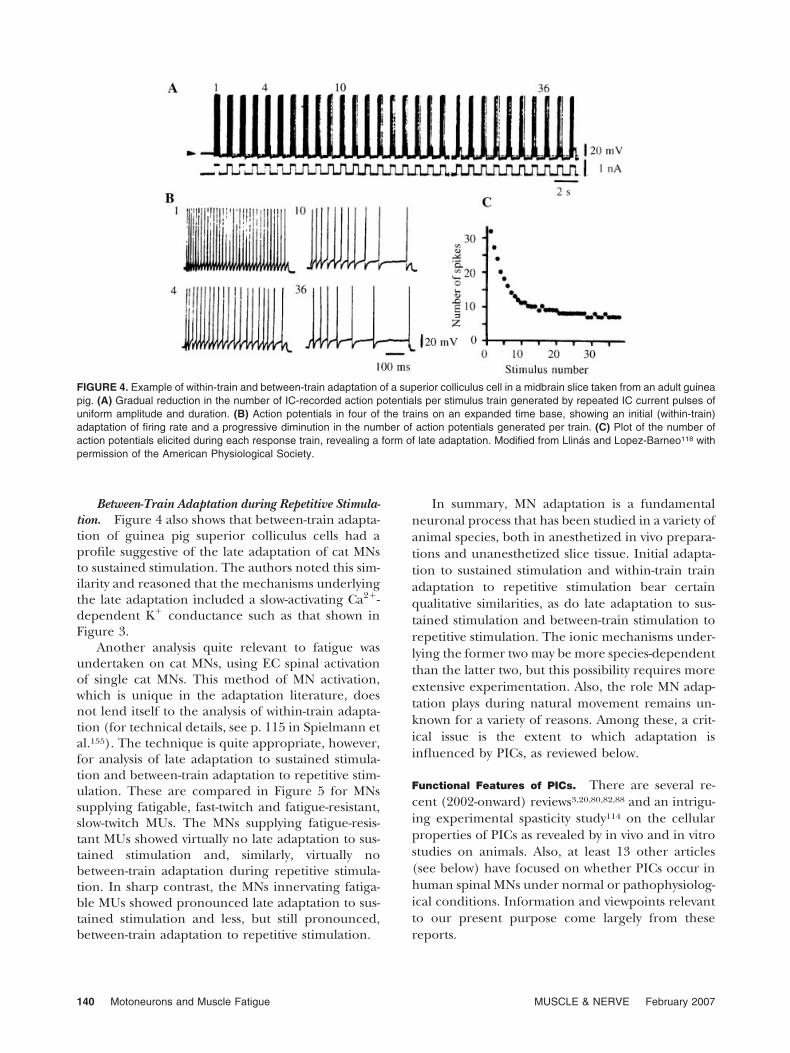
Between-Train Adaptation during Repetitive Stimula-tion. Figure 4 also shows that between-train adapta-tion of guinea pig superior colliculus cells had aprofile suggestive of the late adaptation of cat MNsto sustained stimulation. The authors noted this sim-ilarity and reasoned that the mechanisms underlyingthe late adaptation included a slow-activating Ca2�-dependent K� conductance such as that shown inFigure 3.
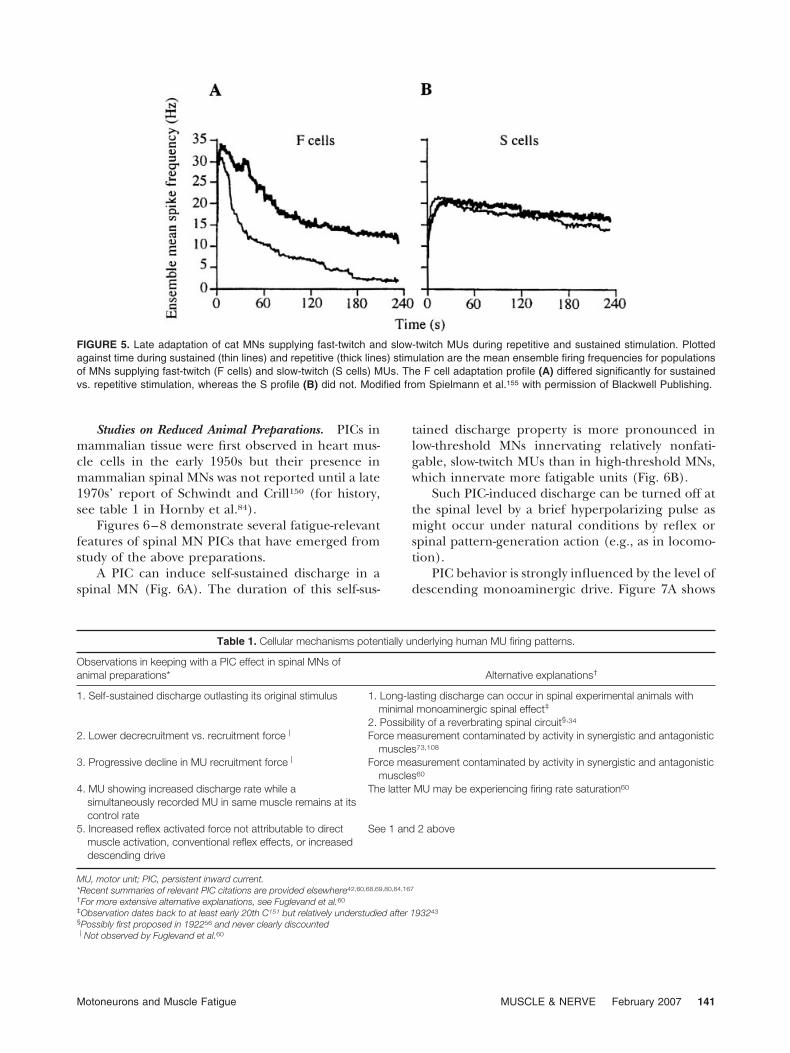
Another analysis quite relevant to fatigue wasundertaken on cat MNs, using EC spinal activationof single cat MNs. This method of MN activation,which is unique in the adaptation literature, doesnot lend itself to the analysis of within-train adapta-tion (for technical details, see p. 115 in Spielmann etal.155). The technique is quite appropriate, however,for analysis of late adaptation to sustained stimula-tion and between-train adaptation to repetitive stim-ulation. These are compared in Figure 5 for MNssupplying fatigable, fast-twitch and fatigue-resistant,slow-twitch MUs. The MNs supplying fatigue-resis-tant MUs showed virtually no late adaptation to sus-tained stimulation and, similarly, virtually nobetween-train adaptation during repetitive stimula-tion. In sharp contrast, the MNs innervating fatiga-ble MUs showed pronounced late adaptation to sus-tained stimulation and less, but still pronounced,between-train adaptation to repetitive stimulation.
In summary, MN adaptation is a fundamentalneuronal process that has been studied in a variety ofanimal species, both in anesthetized in vivo prepara-tions and unanesthetized slice tissue. Initial adapta-tion to sustained stimulation and within-train trainadaptation to repetitive stimulation bear certainqualitative similarities, as do late adaptation to sus-tained stimulation and between-train stimulation torepetitive stimulation. The ionic mechanisms under-lying the former two may be more species-dependentthan the latter two, but this possibility requires moreextensive experimentation. Also, the role MN adap-tation plays during natural movement remains un-known for a variety of reasons. Among these, a crit-ical issue is the extent to which adaptation isinfluenced by PICs, as reviewed below.
Functional Features of PICs. There are several re-cent (2002-onward) reviews3,20,80,82,88 and an intrigu-ing experimental spasticity study114 on the cellularproperties of PICs as revealed by in vivo and in vitrostudies on animals. Also, at least 13 other articles(see below) have focused on whether PICs occur inhuman spinal MNs under normal or pathophysiolog-ical conditions. Information and viewpoints relevantto our present purpose come largely from thesereports.
FIGURE 4. Example of within-train and between-train adaptation of a superior colliculus cell in a midbrain slice taken from an adult guineapig. (A) Gradual reduction in the number of IC-recorded action potentials per stimulus train generated by repeated IC current pulses ofuniform amplitude and duration. (B) Action potentials in four of the trains on an expanded time base, showing an initial (within-train)adaptation of firing rate and a progressive diminution in the number of action potentials generated per train. (C) Plot of the number ofaction potentials elicited during each response train, revealing a form of late adaptation. Modified from Llinas and Lopez-Barneo118 withpermission of the American Physiological Society.
140 Motoneurons and Muscle Fatigue MUSCLE & NERVE February 2007
Studies on Reduced Animal Preparations. PICs inmammalian tissue were first observed in heart mus-cle cells in the early 1950s but their presence inmammalian spinal MNs was not reported until a late1970s’ report of Schwindt and Crill150 (for history,see table 1 in Hornby et al.84).
Figures 6–8 demonstrate several fatigue-relevantfeatures of spinal MN PICs that have emerged fromstudy of the above preparations.
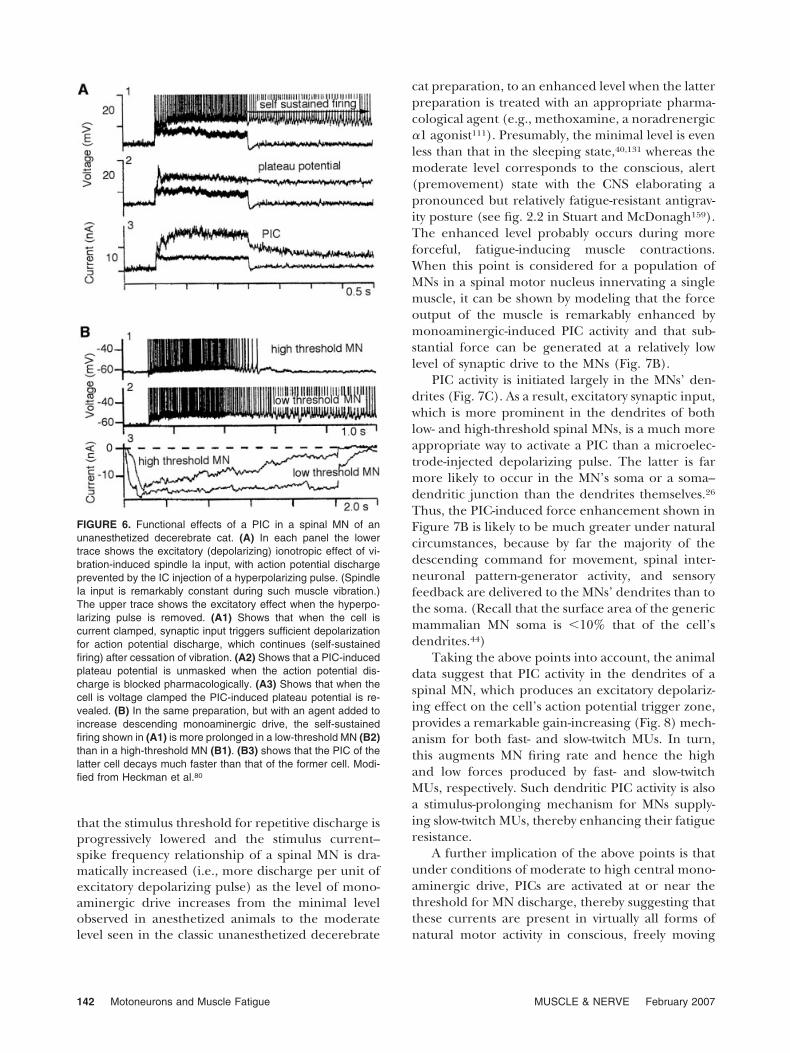
A PIC can induce self-sustained discharge in aspinal MN (Fig. 6A). The duration of this self-sus-
tained discharge property is more pronounced inlow-threshold MNs innervating relatively nonfati-gable, slow-twitch MUs than in high-threshold MNs,which innervate more fatigable units (Fig. 6B).
Such PIC-induced discharge can be turned off atthe spinal level by a brief hyperpolarizing pulse asmight occur under natural conditions by reflex orspinal pattern-generation action (e.g., as in locomo-tion).
PIC behavior is strongly influenced by the level ofdescending monoaminergic drive. Figure 7A shows
FIGURE 5. Late adaptation of cat MNs supplying fast-twitch and slow-twitch MUs during repetitive and sustained stimulation. Plottedagainst time during sustained (thin lines) and repetitive (thick lines) stimulation are the mean ensemble firing frequencies for populationsof MNs supplying fast-twitch (F cells) and slow-twitch (S cells) MUs. The F cell adaptation profile (A) differed significantly for sustainedvs. repetitive stimulation, whereas the S profile (B) did not. Modified from Spielmann et al.155 with permission of Blackwell Publishing.
Table 1. Cellular mechanisms potentially underlying human MU firing patterns.
Observations in keeping with a PIC effect in spinal MNs ofanimal preparations* Alternative explanations†
1. Self-sustained discharge outlasting its original stimulus 1. Long-lasting discharge can occur in spinal experimental animals withminimal monoaminergic spinal effect‡
2. Possibility of a reverbrating spinal circuit§,34
2. Lower decrecruitment vs. recruitment forceP Force measurement contaminated by activity in synergistic and antagonisticmuscles73,108
3. Progressive decline in MU recruitment forceP Force measurement contaminated by activity in synergistic and antagonisticmuscles60
4. MU showing increased discharge rate while asimultaneously recorded MU in same muscle remains at itscontrol rate
The latter MU may be experiencing firing rate saturation60
5. Increased reflex activated force not attributable to directmuscle activation, conventional reflex effects, or increaseddescending drive
See 1 and 2 above
MU, motor unit; PIC, persistent inward current.*Recent summaries of relevant PIC citations are provided elsewhere42,60,68,69,80,84,167
†For more extensive alternative explanations, see Fuglevand et al.60
‡Observation dates back to at least early 20th C151 but relatively understudied after 193243
§Possibly first proposed in 192256 and never clearly discountedPNot observed by Fuglevand et al.60
Motoneurons and Muscle Fatigue MUSCLE & NERVE February 2007 141
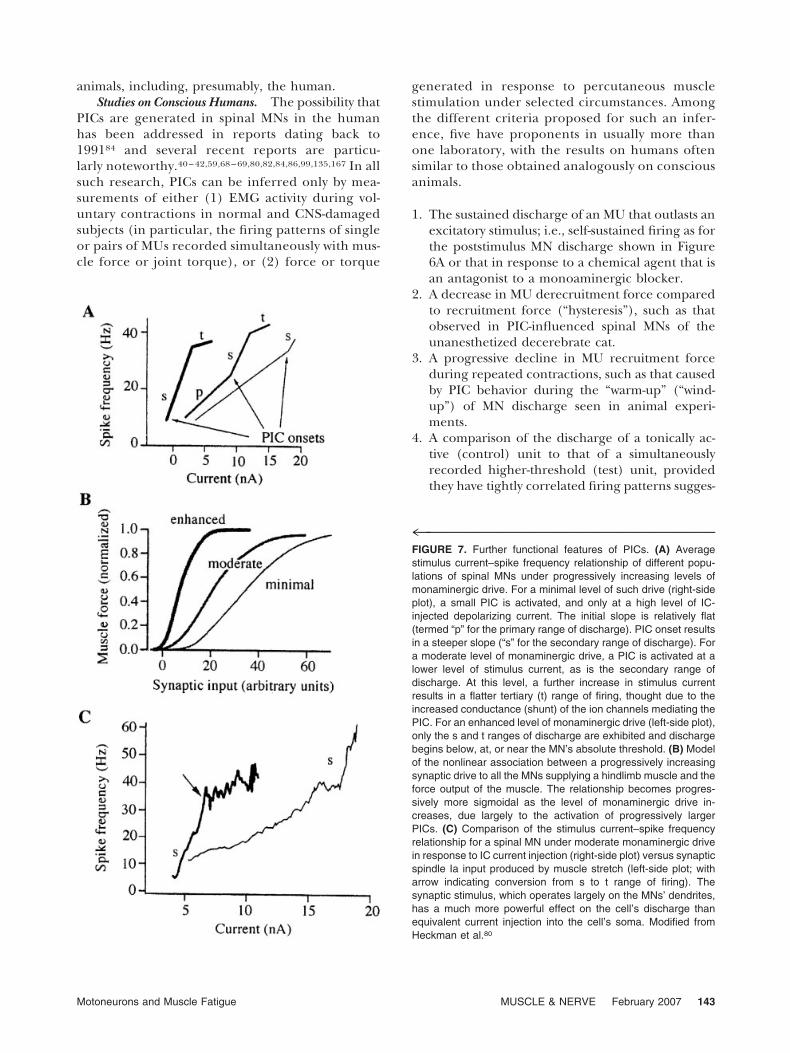
that the stimulus threshold for repetitive discharge isprogressively lowered and the stimulus current–spike frequency relationship of a spinal MN is dra-matically increased (i.e., more discharge per unit ofexcitatory depolarizing pulse) as the level of mono-aminergic drive increases from the minimal levelobserved in anesthetized animals to the moderatelevel seen in the classic unanesthetized decerebrate
cat preparation, to an enhanced level when the latterpreparation is treated with an appropriate pharma-cological agent (e.g., methoxamine, a noradrenergic�1 agonist111). Presumably, the minimal level is evenless than that in the sleeping state,40,131 whereas themoderate level corresponds to the conscious, alert(premovement) state with the CNS elaborating apronounced but relatively fatigue-resistant antigrav-ity posture (see fig. 2.2 in Stuart and McDonagh159).The enhanced level probably occurs during moreforceful, fatigue-inducing muscle contractions.When this point is considered for a population ofMNs in a spinal motor nucleus innervating a singlemuscle, it can be shown by modeling that the forceoutput of the muscle is remarkably enhanced bymonoaminergic-induced PIC activity and that sub-stantial force can be generated at a relatively lowlevel of synaptic drive to the MNs (Fig. 7B).
PIC activity is initiated largely in the MNs’ den-drites (Fig. 7C). As a result, excitatory synaptic input,which is more prominent in the dendrites of bothlow- and high-threshold spinal MNs, is a much moreappropriate way to activate a PIC than a microelec-trode-injected depolarizing pulse. The latter is farmore likely to occur in the MN’s soma or a soma–dendritic junction than the dendrites themselves.26
Thus, the PIC-induced force enhancement shown inFigure 7B is likely to be much greater under naturalcircumstances, because by far the majority of thedescending command for movement, spinal inter-neuronal pattern-generator activity, and sensoryfeedback are delivered to the MNs’ dendrites than tothe soma. (Recall that the surface area of the genericmammalian MN soma is �10% that of the cell’sdendrites.44)
Taking the above points into account, the animaldata suggest that PIC activity in the dendrites of aspinal MN, which produces an excitatory depolariz-ing effect on the cell’s action potential trigger zone,provides a remarkable gain-increasing (Fig. 8) mech-anism for both fast- and slow-twitch MUs. In turn,this augments MN firing rate and hence the highand low forces produced by fast- and slow-twitchMUs, respectively. Such dendritic PIC activity is alsoa stimulus-prolonging mechanism for MNs supply-ing slow-twitch MUs, thereby enhancing their fatigueresistance.
A further implication of the above points is thatunder conditions of moderate to high central mono-aminergic drive, PICs are activated at or near thethreshold for MN discharge, thereby suggesting thatthese currents are present in virtually all forms ofnatural motor activity in conscious, freely moving
FIGURE 6. Functional effects of a PIC in a spinal MN of anunanesthetized decerebrate cat. (A) In each panel the lowertrace shows the excitatory (depolarizing) ionotropic effect of vi-bration-induced spindle Ia input, with action potential dischargeprevented by the IC injection of a hyperpolarizing pulse. (SpindleIa input is remarkably constant during such muscle vibration.)The upper trace shows the excitatory effect when the hyperpo-larizing pulse is removed. (A1) Shows that when the cell iscurrent clamped, synaptic input triggers sufficient depolarizationfor action potential discharge, which continues (self-sustainedfiring) after cessation of vibration. (A2) Shows that a PIC-inducedplateau potential is unmasked when the action potential dis-charge is blocked pharmacologically. (A3) Shows that when thecell is voltage clamped the PIC-induced plateau potential is re-vealed. (B) In the same preparation, but with an agent added toincrease descending monoaminergic drive, the self-sustainedfiring shown in (A1) is more prolonged in a low-threshold MN (B2)than in a high-threshold MN (B1). (B3) shows that the PIC of thelatter cell decays much faster than that of the former cell. Modi-fied from Heckman et al.80
142 Motoneurons and Muscle Fatigue MUSCLE & NERVE February 2007
animals, including, presumably, the human.Studies on Conscious Humans. The possibility that
PICs are generated in spinal MNs in the humanhas been addressed in reports dating back to199184 and several recent reports are particu-larly noteworthy.40 – 42,59,68 – 69,80,82,84,86,99,135,167 In allsuch research, PICs can be inferred only by mea-surements of either (1) EMG activity during vol-untary contractions in normal and CNS-damagedsubjects (in particular, the firing patterns of singleor pairs of MUs recorded simultaneously with mus-cle force or joint torque), or (2) force or torque
generated in response to percutaneous musclestimulation under selected circumstances. Amongthe different criteria proposed for such an infer-ence, five have proponents in usually more thanone laboratory, with the results on humans oftensimilar to those obtained analogously on consciousanimals.
1. The sustained discharge of an MU that outlasts anexcitatory stimulus; i.e., self-sustained firing as forthe poststimulus MN discharge shown in Figure6A or that in response to a chemical agent that isan antagonist to a monoaminergic blocker.
2. A decrease in MU derecruitment force comparedto recruitment force (“hysteresis”), such as thatobserved in PIC-influenced spinal MNs of theunanesthetized decerebrate cat.
3. A progressive decline in MU recruitment forceduring repeated contractions, such as that causedby PIC behavior during the “warm-up” (“wind-up”) of MN discharge seen in animal experi-ments.
4. A comparison of the discharge of a tonically ac-tive (control) unit to that of a simultaneouslyrecorded higher-threshold (test) unit, providedthey have tightly correlated firing patterns sugges-
4™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™FIGURE 7. Further functional features of PICs. (A) Averagestimulus current–spike frequency relationship of different popu-lations of spinal MNs under progressively increasing levels ofmonaminergic drive. For a minimal level of such drive (right-sideplot), a small PIC is activated, and only at a high level of IC-injected depolarizing current. The initial slope is relatively flat(termed “p” for the primary range of discharge). PIC onset resultsin a steeper slope (“s” for the secondary range of discharge). Fora moderate level of monaminergic drive, a PIC is activated at alower level of stimulus current, as is the secondary range ofdischarge. At this level, a further increase in stimulus currentresults in a flatter tertiary (t) range of firing, thought due to theincreased conductance (shunt) of the ion channels mediating thePIC. For an enhanced level of monaminergic drive (left-side plot),only the s and t ranges of discharge are exhibited and dischargebegins below, at, or near the MN’s absolute threshold. (B) Modelof the nonlinear association between a progressively increasingsynaptic drive to all the MNs supplying a hindlimb muscle and theforce output of the muscle. The relationship becomes progres-sively more sigmoidal as the level of monaminergic drive in-creases, due largely to the activation of progressively largerPICs. (C) Comparison of the stimulus current–spike frequencyrelationship for a spinal MN under moderate monaminergic drivein response to IC current injection (right-side plot) versus synapticspindle Ia input produced by muscle stretch (left-side plot; witharrow indicating conversion from s to t range of firing). Thesynaptic stimulus, which operates largely on the MNs’ dendrites,has a much more powerful effect on the cell’s discharge thanequivalent current injection into the cell’s soma. Modified fromHeckman et al.80
Motoneurons and Muscle Fatigue MUSCLE & NERVE February 2007 143
tive of the sharing of synaptic inputs. If in re-sponse to an excitatory stimulus the test unitshows relatively more excitatory discharge thanthe control unit, it is argued that the test unit’sMN is generating a PIC, whereas the MN of thecontrol unit has either reached its maximum PICor is not generating one.
5. A muscle force or torque response to percutane-ous muscle stimulation, the strength of whichcannot be explained by (1) direct muscle activa-tion by motor axons, (2) non-PIC–assisted reflexaction, and (3) increased descending voluntarydrive.
In all five of the above cases, there is certainlyclear-cut information in support of a PIC inferencederived from experimental work on animal prepara-tions. In parallel, however, Table 1 shows alternativeexplanations. Note that the latter are based on acombination of animal experimentation and often-neglected biomechanical considerations.
Table 1 suggests that the case for PIC develop-ment in human spinal MNs in now quite strong,albeit slightly overemphasized by the technical diffi-
culty of including other possibilities. Such overem-phasis has often been the case in human experimen-tation on normal and CNS- or peripheral nervoussystem–damaged subjects when a new spinal mech-anism has been recently established by rigorous ex-perimentation on surgically reduced animal prepa-rations. Previous examples dating back to at least the1960s include, for example, the fusimotor innerva-tion of muscle spindles, recurrent inhibition via Ren-shaw cells, spinal presynaptic inhibition, complexalternative spinal reflex circuitry, spinal neurophar-macology, and neuromuscular junction malfunctionin myasthenia gravis. In these cases, examples ofinitial overemphases have been reviewed and evalu-ated.30,95,119,141 Similar initial overemphases have oc-curred in the study of Parkinson’s disease25 andspasticity109,162 and, presumably, many other neuro-logical deficits. In all these and presumably all futurecases, however, animal experimentation is absolutelyessential for providing insight into potential mecha-nisms underlying human motor behavior and foradvancing the scientific basis and efficacy of drugand rehabilitation therapies for human movementdisorders.
Despite the alternative possibilities summa-rized in Table 1, the case for PIC involvement inspinal MN behavior is compelling on evolutionarygrounds alone, given that PICs are ubiquitousthroughout the mammalian CNS, including hu-man neocortical neurons.149 The problem re-mains, however, how to determine the relativecontribution that a variety of other spinal mecha-nisms make to the firing patterns of MNs in con-scious human subjects.
Does MN Adaptation Occur in the Presence of a PIC?
There is surprisingly little consideration in the liter-ature about whether MN adaptation is obviatedwhen an MN is exhibiting a PIC. Figure 9 shows thatinferences are possible, however.
Figure 9A,B shows some results obtained byBrownstone and colleagues27,29 and Spielmann’s153
additional measurements (Fig. 9C,D) on some oftheir data. Brownstone compared the firing proper-ties of cat hindlimb MNs when the unanesthetizedparalyzed high decerebrate cat was in its controlstate, and when it was elaborating fictive locomotionbrought on by brainstem stimulation. The formerstate is suggestive of the moderate level of monoam-inergic drive shown in Figure 6, whereas the fictivelocomotive state involved an enhanced drive. Notethat initial adaptation (within repetitive stimulustrains) was present in the control (Fig. 9B) but notthe fictive locomotor (Fig. 9A) state. Subsequently,
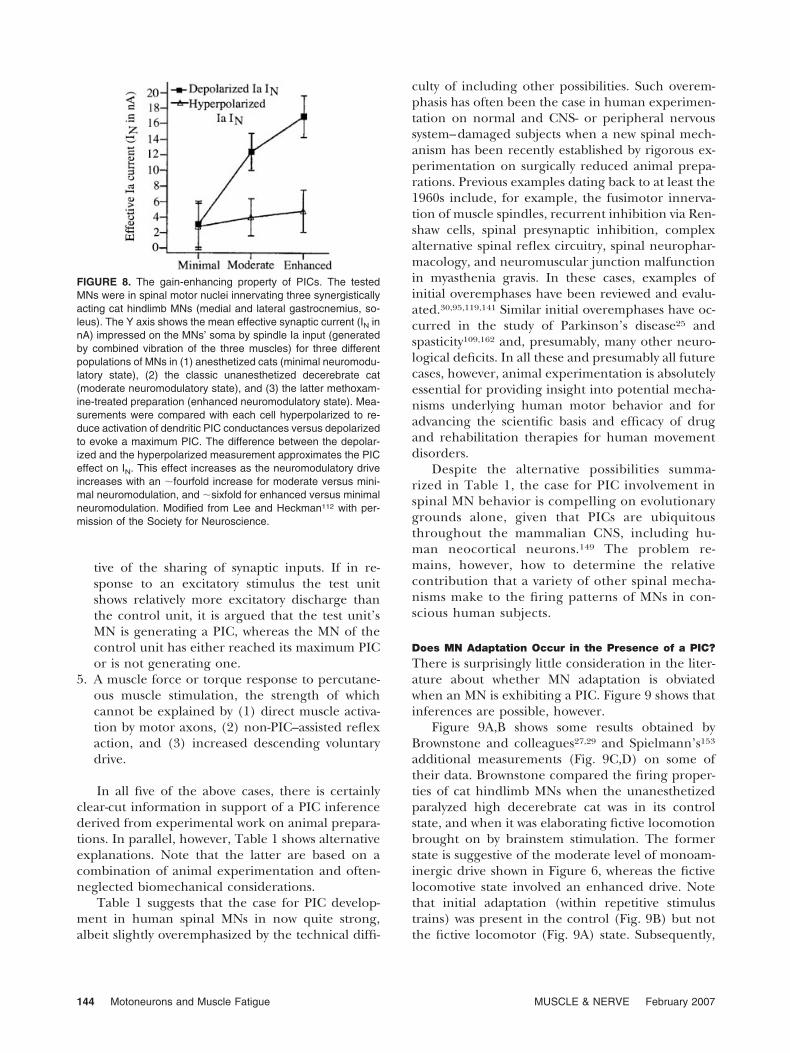
FIGURE 8. The gain-enhancing property of PICs. The testedMNs were in spinal motor nuclei innervating three synergisticallyacting cat hindlimb MNs (medial and lateral gastrocnemius, so-leus). The Y axis shows the mean effective synaptic current (IN innA) impressed on the MNs’ soma by spindle Ia input (generatedby combined vibration of the three muscles) for three differentpopulations of MNs in (1) anesthetized cats (minimal neuromodu-latory state), (2) the classic unanesthetized decerebrate cat(moderate neuromodulatory state), and (3) the latter methoxam-ine-treated preparation (enhanced neuromodulatory state). Mea-surements were compared with each cell hyperpolarized to re-duce activation of dendritic PIC conductances versus depolarizedto evoke a maximum PIC. The difference between the depolar-ized and the hyperpolarized measurement approximates the PICeffect on IN. This effect increases as the neuromodulatory driveincreases with an �fourfold increase for moderate versus mini-mal neuromodulation, and �sixfold for enhanced versus minimalneuromodulation. Modified from Lee and Heckman112 with per-mission of the Society for Neuroscience.
144 Motoneurons and Muscle Fatigue MUSCLE & NERVE February 2007
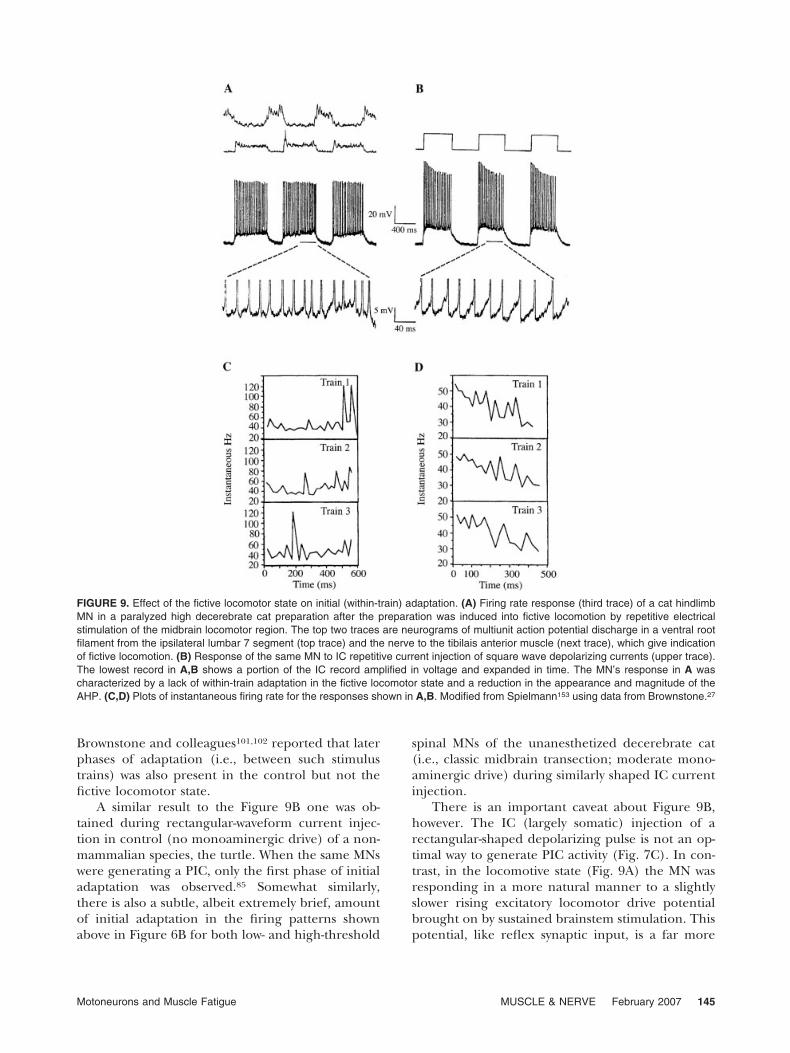
Brownstone and colleagues101,102 reported that laterphases of adaptation (i.e., between such stimulustrains) was also present in the control but not thefictive locomotor state.
A similar result to the Figure 9B one was ob-tained during rectangular-waveform current injec-tion in control (no monoaminergic drive) of a non-mammalian species, the turtle. When the same MNswere generating a PIC, only the first phase of initialadaptation was observed.85 Somewhat similarly,there is also a subtle, albeit extremely brief, amountof initial adaptation in the firing patterns shownabove in Figure 6B for both low- and high-threshold
spinal MNs of the unanesthetized decerebrate cat(i.e., classic midbrain transection; moderate mono-aminergic drive) during similarly shaped IC currentinjection.
There is an important caveat about Figure 9B,however. The IC (largely somatic) injection of arectangular-shaped depolarizing pulse is not an op-timal way to generate PIC activity (Fig. 7C). In con-trast, in the locomotive state (Fig. 9A) the MN wasresponding in a more natural manner to a slightlyslower rising excitatory locomotor drive potentialbrought on by sustained brainstem stimulation. Thispotential, like reflex synaptic input, is a far more
FIGURE 9. Effect of the fictive locomotor state on initial (within-train) adaptation. (A) Firing rate response (third trace) of a cat hindlimbMN in a paralyzed high decerebrate cat preparation after the preparation was induced into fictive locomotion by repetitive electricalstimulation of the midbrain locomotor region. The top two traces are neurograms of multiunit action potential discharge in a ventral rootfilament from the ipsilateral lumbar 7 segment (top trace) and the nerve to the tibilais anterior muscle (next trace), which give indicationof fictive locomotion. (B) Response of the same MN to IC repetitive current injection of square wave depolarizing currents (upper trace).The lowest record in A,B shows a portion of the IC record amplified in voltage and expanded in time. The MN’s response in A wascharacterized by a lack of within-train adaptation in the fictive locomotor state and a reduction in the appearance and magnitude of theAHP. (C,D) Plots of instantaneous firing rate for the responses shown in A,B. Modified from Spielmann153 using data from Brownstone.27
Motoneurons and Muscle Fatigue MUSCLE & NERVE February 2007 145
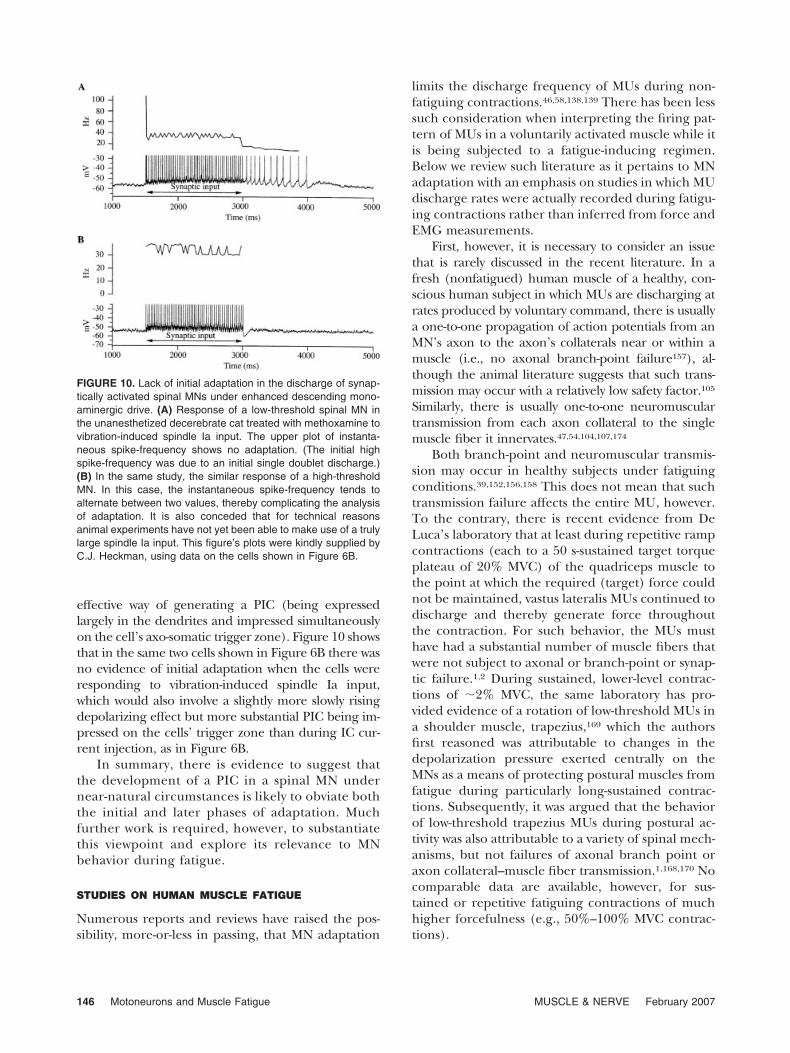
effective way of generating a PIC (being expressedlargely in the dendrites and impressed simultaneouslyon the cell’s axo-somatic trigger zone). Figure 10 showsthat in the same two cells shown in Figure 6B there wasno evidence of initial adaptation when the cells wereresponding to vibration-induced spindle Ia input,which would also involve a slightly more slowly risingdepolarizing effect but more substantial PIC being im-pressed on the cells’ trigger zone than during IC cur-rent injection, as in Figure 6B.
In summary, there is evidence to suggest thatthe development of a PIC in a spinal MN undernear-natural circumstances is likely to obviate boththe initial and later phases of adaptation. Muchfurther work is required, however, to substantiatethis viewpoint and explore its relevance to MNbehavior during fatigue.
STUDIES ON HUMAN MUSCLE FATIGUE
Numerous reports and reviews have raised the pos-sibility, more-or-less in passing, that MN adaptation
limits the discharge frequency of MUs during non-fatiguing contractions.46,58,138,139 There has been lesssuch consideration when interpreting the firing pat-tern of MUs in a voluntarily activated muscle while itis being subjected to a fatigue-inducing regimen.Below we review such literature as it pertains to MNadaptation with an emphasis on studies in which MUdischarge rates were actually recorded during fatigu-ing contractions rather than inferred from force andEMG measurements.
First, however, it is necessary to consider an issuethat is rarely discussed in the recent literature. In afresh (nonfatigued) human muscle of a healthy, con-scious human subject in which MUs are discharging atrates produced by voluntary command, there is usuallya one-to-one propagation of action potentials from anMN’s axon to the axon’s collaterals near or within amuscle (i.e., no axonal branch-point failure157), al-though the animal literature suggests that such trans-mission may occur with a relatively low safety factor.105
Similarly, there is usually one-to-one neuromusculartransmission from each axon collateral to the singlemuscle fiber it innervates.47,54,104,107,174
Both branch-point and neuromuscular transmis-sion may occur in healthy subjects under fatiguingconditions.39,152,156,158 This does not mean that suchtransmission failure affects the entire MU, however.To the contrary, there is recent evidence from DeLuca’s laboratory that at least during repetitive rampcontractions (each to a 50 s-sustained target torqueplateau of 20% MVC) of the quadriceps muscle tothe point at which the required (target) force couldnot be maintained, vastus lateralis MUs continued todischarge and thereby generate force throughoutthe contraction. For such behavior, the MUs musthave had a substantial number of muscle fibers thatwere not subject to axonal or branch-point or synap-tic failure.1,2 During sustained, lower-level contrac-tions of �2% MVC, the same laboratory has pro-vided evidence of a rotation of low-threshold MUs ina shoulder muscle, trapezius,169 which the authorsfirst reasoned was attributable to changes in thedepolarization pressure exerted centrally on theMNs as a means of protecting postural muscles fromfatigue during particularly long-sustained contrac-tions. Subsequently, it was argued that the behaviorof low-threshold trapezius MUs during postural ac-tivity was also attributable to a variety of spinal mech-anisms, but not failures of axonal branch point oraxon collateral–muscle fiber transmission.1,168,170 Nocomparable data are available, however, for sus-tained or repetitive fatiguing contractions of muchhigher forcefulness (e.g., 50%–100% MVC contrac-tions).
FIGURE 10. Lack of initial adaptation in the discharge of synap-tically activated spinal MNs under enhanced descending mono-aminergic drive. (A) Response of a low-threshold spinal MN inthe unanesthetized decerebrate cat treated with methoxamine tovibration-induced spindle Ia input. The upper plot of instanta-neous spike-frequency shows no adaptation. (The initial highspike-frequency was due to an initial single doublet discharge.)(B) In the same study, the similar response of a high-thresholdMN. In this case, the instantaneous spike-frequency tends toalternate between two values, thereby complicating the analysisof adaptation. It is also conceded that for technical reasonsanimal experiments have not yet been able to make use of a trulylarge spindle Ia input. This figure’s plots were kindly supplied byC.J. Heckman, using data on the cells shown in Figure 6B.
146 Motoneurons and Muscle Fatigue MUSCLE & NERVE February 2007

In summary, under normal (nonpathophysi-ological) circumstances the literature on fatigue-induced, axonal branch-point, and axon-collateralneuromuscular junction failure is quite sparse ex-cept for low-force contractions, and that on fatigue-induced neuromuscular transmission failure is quitecontroversial.33,62 Presumably, both are task-depen-dent and if they do occur, either singly or together,they are more likely to involve the higher-threshold,more fatigable glycolytic MUs than the lower thresh-old, fatigue-resistant ones.39,62,152
Even in fatigue-inducing tasks in which axonalbranch point- and neuromuscular transmission fail-ure may not be implicated, evidence of MN adapta-tion during human muscle contractions is quitescanty and subject to alternative explanations, asreviewed below.
Sustained Contractions. For the question at hand, itis convenient to consider such contractions bothwhen an MVC was undertaken and when the proto-col required the elaboration of lesser contractionsset at various levels of the MVC.
Sustained MVCs. In this type of experiment, theinvestigators typically use highly motivated trainedsubjects in order to ensure that they (1) can indeedelicit maximum contractions by voluntary effort, and(2) sustain this effort for the duration of a fatiguetest.14,61 It is well known that it is far more difficult toachieve the latter in the conscious human as com-pared to IC current injection into MNs in experi-ments on surgically reduced animals or slices of theirCNS. Note, however, that in the latter case therehave been understandably (i.e., for technical rea-sons) no studies in which the IC current injectionwas of sufficient strength to raise MN firing rates tothe level required for a peak sustained (tetanic)contraction of the supplied MU. Another problemwith MVCs, whether sustained or repetitive, iswhether the discharge of single MUs can be mea-sured unambiguously, due to interference from thedischarge of other MUs. In a few select cases thereappears to be general consensus that faithful unitaryrecording was achieved during sustained MVCs: e.g.,the unit in figure 9 of Marsden et al.,124 and thestudies of certain other investigators.24,75,121In mostother cases, however, an element of doubt is everpresent.
Bigland-Ritchie and her co-workers16,18,175 stimu-lated interest to a considerable degree in the possi-bility that MN adaptation contributes to fatigue dur-ing sustained MVCs in humans. In a sense, theirwork extended the earlier findings of Marsden et al.,which were first reported in a widely read abstract123
before a full-length account124 appeared over a de-cade later.
Figure 11A shows that as a human hand muscle,adductor pollicis (AP), fatigued during a voluntarilycommanded 40-s MVC, there was a decline in bothmuscle force (by �33%) and the mean activationrate of populations of MUs within the muscle (abouta 45% reduction; from �27 Hz to �15 Hz). Asargued below, it seems likely that the MUs exhibitingsuch a decline in discharge rate were composed ofglycolytic (fast-twitch type II) muscle fibers, which inthis muscle presumably make up �20% of the totalfiber content (see tables 2 and 4 in Johnson et al.93).
In a subsequent report, Bigland-Ritchie et al.16
proposed that “. . .during fatigue, motoneurone fir-ing rates may be regulated by a peripheral reflexoriginating in response to fatigue-induced changeswithin the muscle.” The authors cautioned, however,that “. . .the decline in motoneurone firing rates . . .may result primarily from changes in central mo-toneurone excitability (i.e., adaptation); the timecourse of frequency changes is quite similar to thatreported by Kernell and colleagues for changes inthe discharge rates of cat single motoneurones inresponse to constant current injection.” Further ex-
FIGURE 11. Comparison of force and firing rate decline in theconscious human and the anesthetized cat. (A) The upper panelshows force developed by the human hand muscle, adductorpollicis, during a 40-s MVC. Throughout the contraction the force(in Newtons, N) reduced by �33%. The lower panel shows theconcomitant drop in on-average firing rate (HZ) of MUs. Eachdata point is the mean rate of a single spike train recorded fromgroups of MUs, as based on a single 1-s average at various timesthroughout the contraction. The on-average firing rate dropped by�45%. (B) The upper panel shows the force developed by afast-twitch fatigable MU in a cat hindlimb muscle, medial gastroc-nemius, during 40-s of IC current injection into the unit’s MN. Thelower panel shows the reduction in the MNs’ firing rate, which canclearly be attributed to adaptation. This reduction was also�45%. The fall in force (80%) was greater, however. Redrawnfrom parts of figures in Bigland-Ritchie et al.18 (A) and Kernell andMonster97 (B) with permission of Blackwell Publishing (A) andSpringer-Verlag (B).
Motoneurons and Muscle Fatigue MUSCLE & NERVE February 2007 147

periments, with added controls, led Bigland-Ritchieand colleagues175 to assert more strongly that:“. . .motoneuron firing rates are inhibited by somereflex originating from the muscle.” They went on toreflect that: “It is unclear what additional rolechanges in central motoneuron excitability may playin regulating motoneuron discharge rates duringvoluntary contractions.”
Figure 11B is from a study on deeply anesthetizedcats.97 In this work, MNs of a hindlimb muscle, me-dial gastrocnemius, were activated by a sustained ICcurrent injection set 5 nA above the threshold forsteady-state discharge.96,97 As in Figure 11A, the sus-tained stimulus produced a decline in both MUforce and MN discharge rate, with the latter clearlyattributable to adaptation.
Presumably, the relative similarity in the dis-charge rate declines shown in Figure 11A and B wasa key factor when the possibility was first raised bythe Bigland-Ritchie group that adaptation mightcontribute to fatigue in conscious humans. Thereare two caveats about this comparison, however.First, in Figure 11A the period of MN/MU activationin the human study began clearly with an MVC,which was sustained as fatigue ensued. In Figure11B, however, an initial MN firing rate of 29 Hz foran MN supplying a fast-twitch MU in a study onanesthetized cats would not produce a maximumMU contraction, but rather about a 40% MVC (forreferences on cat medial gastrocnemius MN/MUproperties, see Burke31). A more valid comparisonwould require either activating cat MNs by synapticactivation, diminishing the forcefulness of the hu-man sustained contraction (to �40% MVC), or rais-ing the initial stimulus strength of the IC currentinjection to produce an initial firing rate of �100 Hzin an MN supplying a fast-twitch MU in the cat.Unfortunately, such requisite information is not yetavailable.
A second caveat about the Figure 11 comparisonis that it is highly likely that the unanesthetizedhuman MNs of Figure 11A were generating PICs(i.e., due to relatively high monoaminergic “tone”),whereas the anesthetized cat MNs were clearly not.As such, the decline in human MN firing rates shownin Figure 11A were possibly not attributable to MNadaptation, whereas this was indeed the cause of thedecline shown in Figure 11B for the cat MN.
Early on, Bigland-Ritchie et al.16 thought the in-hibitory reflex lowering MU discharge rate during asustained MVC took origin in the fatigue-inducedactivation of Group III and IV metabolic muscleafferents: “They . . . seem ideally suited to mediate areflex which may modulate motoneurone firing rate
in fatigue.” In that report, and subsequently,175 how-ever, the possibility was not discounted that the af-ferent limb of the inhibitory reflex involved sensoryinput from low-threshold mechanoreceptors, musclespindles, and Golgi tendon organs. Recently, Butleret al.32 have again raised the latter possibility formuscle spindle input. Their study involved notingthe responses of MUs in human elbow flexor mus-cles to corticospinal stimulation during MVCs andischemia of the test muscle group. Their resultsruled out an inhibitory reflex attributable to GroupIII–IV afferents. Rather, they proposed that the ob-served reduction in MU firing rate was due to (1)MN adaptation; (2) dysfacilitation of MN dischargevia reduced muscle spindle discharge, which hasbeen observed in humans during fatigue23,122; or (3)both.
Macefield et al.121 provided de novo informationon the firing rates of MUs in a human toe muscle,EHL (extensor hallucis longus), during a fatiguingMVC that was sustained for up to 120–150 s. In thisstudy it was indeed possible to record unambigu-ously the discharge of single MUs during a full MVC.It was shown that the mean firing rate did notchange appreciably as force dropped. One of thereasons proposed by the authors for this finding,which is unique in the human MU literature, was thelack of adaptation of MNs supplying slow-twitchMUs, as observed previously in hindlimb MNs of thecat.97,155 The number of slow-twitch (SO or type I)fibers in EHL has not yet been counted. In a previ-ous study, however, those authors showed that thecontractile properties of such MUs were like those ofthe human soleus muscle,120 which is composed of�86%–89% SO fibers.93
It seems likely that there is no inconsistency be-tween the findings on AP, a human hand muscle,18,19
and EHL, a human toe muscle.121 There is evidence,albeit limited, of differences in the organization andproperties of the segmental control system (spinalcircuitry, reflex connectivity, neuromechanical prop-erties of motor units) for upper and lower limbs, andproximal and distal limb muscles. Among severalexamples, two striking ones are (1) the greaterstrength of Renshaw cell effects on MNs supplyingproximal than distal MUs in the cat163,171 and thenonhuman primate and human,90 and (2) fastertwitch contraction times for the human AP muscle59
than EHL120 and soleus11 MUs even though the rel-ative percentage of SO fibers is similar in all threemuscles. The latter comparison must also take intoaccount, however, that the corticospinal tract hasmore prevalent monosynaptic connections with APMNs than with MNs supplying EHL and soleus.
148 Motoneurons and Muscle Fatigue MUSCLE & NERVE February 2007

In summary, there is no clear-cut evidence thatMN adaptation contributes to fatigue during sus-tained MVCs. To the contrary, there is hard evidencethat such adaptation does not occur in a toe musclewith a high proportion of SO MUs.120,121 Also,fatigue-induced MU firing-rate declines in the hu-man upper limb18 are now better explained by othersegmental control mechanisms.32
It also deserves mention that in the MN study ofKernell and Monster96 on anesthetized cats “. . .sev-eral cells were encountered that were unable to de-liver a maintained discharge of several seconds inresponse to constant stimulation.” This type of be-havior has also been seen in human MUs (see, e.g.,fig. 7.3 in McComas127), but it is rarely commentedon. Similarly, some MNs in the Kernell and Mon-ster96 sample, which discharged for �10 s but �4min in response to constant-current IC injection set5 nA above the threshold for repetitive discharge,resumed their firing after a short “rest” period inresponse to IC pulses set at �3 nA above the initialstimulus strength. This type of behavior resemblesthat observed in some of the human forearm MUsstudied by Peters and Fuglevand.140 In the lattercase, however, the authors could not exclude thepossibility that “. . .excitatory drive may diminishduring maximal voluntary effort.” These variousfindings suggest that much is yet to be learned aboutMN adaptation during sustained stimulation in bothanesthetized animal preparations and conscious hu-mans.
Sustained, Submaximal Voluntary Contractions. Asmentioned earlier, the study of MN adaptation inanimal preparations by noting a decline in firing ratefollowing the IC injection of a constant-strength de-polarizing current into an MN supplying a fast-twitchMU has never involved an initial firing rate thatwould produce peak tetanic force in the MU. Rather,the published literature suggests that the maximumsuch force would be �40% of the unit’s peak force.Therefore, it would be of interest to know whetherMN adaptation is a feature of human MU firingpatterns during submaximal voluntary contractions(sub-MVCs) sustained until the requested (target)force begins to decline. In evaluating this literature,it is helpful to remember that in such contractionsthere is evidence of motor unit substitution but notrotation. Rotation requires that the same motor unitbe recruited again subsequently, whereas substitu-tion means that a motor unit is derecruited andother motor units are recruited.169,170 It is still some-what uncertain, however, whether rotation is possi-ble once force has begun to decline.
At first glance, it would seem that a sustainedsub-MVC (at least an isometric one) involves a pro-gressive increase in the excitatory drive to the rele-vant MNs and an in-parallel reduction in MU firingrate, which may or may not be linear. Althoughliterature supporting these generalities is indeedavailable, the situation becomes more complex if thefull literature is examined.1,2,4,39,49,63–66,106,126,129,137
Although a declining MU firing rate is the mostprevalent finding in the above studies, there are justas valid examples in them of MUs whose firing rateeither remained unchanged or increased through-out, or during part of the sub-MVCs. Several of thesereports emphasize that the variable MU dischargebehavior is attributable in part to different fatigue-inducing muscle activation regimens (e.g., differenttest muscles with different percentages of the differentmuscle fiber types; different activation protocols; andhigh- vs. low-force sub-MVCs). This is not surprisingbecause fatigue mechanisms in the CNS and pe-ripheral neuromuscular system are task depen-dent.52,61,89,134
Common to virtually all of the above literature isthe lack of claim that the excitatory drive to theactive MNs remained constant throughout the dura-tion of the sustained contraction. As such, it is notpossible to assert assuredly that MN adaptation ac-counted in part for a declining MN firing rate, eventhough it may well be a factor, even in the face of achanging excitatory drive to the active MNs.
Repetitive Contractions. Figure 5 shows that within-train (like initial) and between-train (like late) MNadaptation are clearly evident in studies on MNssupplying fast-twitch but not slow-twitch cat MUswhen subjected to repetitive IC current injection.Such results raise the question as to whether compa-rable data (i.e., MU firing rate) are available in thehuman literature on repetitive contractions that fea-tured epochs of constant or near-constant force. Ac-cordingly, we searched in the literature for suchstudies, looking in particular for fatiguing protocolsin which the repeated contractions were sustained ata constant force for 1-s to 10-s periods and wererepeated at intervals �5 s. In such studies, it isconventional to use the term “endurance time” forthe time throughout which the requested (target)%MVC can be maintained. This time also signals thetime in the fatigue test when the force of a MVCequals the target force. It has invariably been foundthat the surface EMG of the active muscle(s) in-creases progressively throughout the fatigue test,thereby indicating that the excitatory drive to theactive MNs is also progressively increasing. This is
Motoneurons and Muscle Fatigue MUSCLE & NERVE February 2007 149
not the case, of course, for experimental work onMN adaptation in animal preparations, which hasinvariably involved use of constant-strength IC stim-ulation of the tested MNs.
Repetitive MVCs. To our knowledge, no suchstudies have been undertaken. There are, however,repetitive MVC studies in the literature that madeuse of measurements of force (or torque) and whole-muscle EMG but not MU firing patterns.94,136
Repetitive Sub-MVCs. There are several studieson the force (or torque) and EMG during suchcontractions,50 but not including measurements onMU firing patterns. One study included the lattermeasurements but they were not reported for themaintained portions of each contraction, but ratherduring the contractions’ ramp-up and ramp-downphases.53 Thus, for the present purposes, it seemssufficient to comment on five studies that have pro-vided data relevant to the question at hand. In noneof these studies were measurements provided on MUfiring rates during the steady-state phase of the indi-vidual contractions (i.e., a test like that for within-train MN adaptation). Rather, one of the foci inthese studies was the change in MU firing ratethroughout the sustained phases of the fatiguingtask, i.e., like the between-train MN adaptation dis-cussed above.
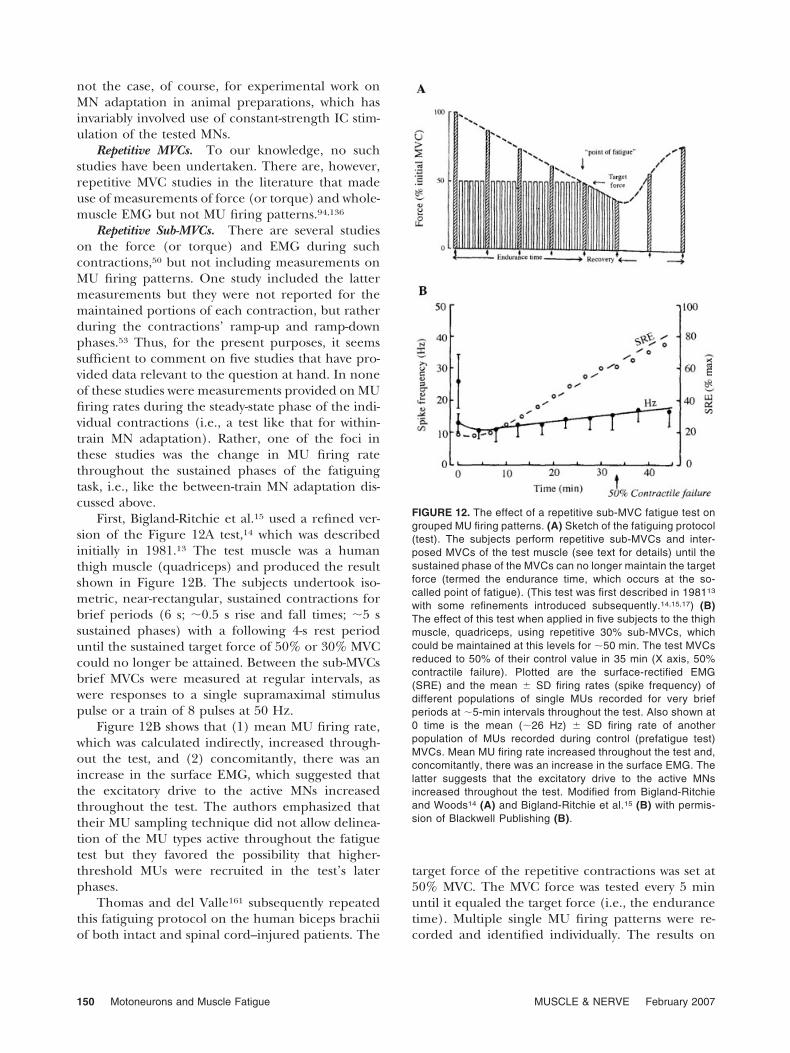
First, Bigland-Ritchie et al.15 used a refined ver-sion of the Figure 12A test,14 which was describedinitially in 1981.13 The test muscle was a humanthigh muscle (quadriceps) and produced the resultshown in Figure 12B. The subjects undertook iso-metric, near-rectangular, sustained contractions forbrief periods (6 s; �0.5 s rise and fall times; �5 ssustained phases) with a following 4-s rest perioduntil the sustained target force of 50% or 30% MVCcould no longer be attained. Between the sub-MVCsbrief MVCs were measured at regular intervals, aswere responses to a single supramaximal stimuluspulse or a train of 8 pulses at 50 Hz.
Figure 12B shows that (1) mean MU firing rate,which was calculated indirectly, increased through-out the test, and (2) concomitantly, there was anincrease in the surface EMG, which suggested thatthe excitatory drive to the active MNs increasedthroughout the test. The authors emphasized thattheir MU sampling technique did not allow delinea-tion of the MU types active throughout the fatiguetest but they favored the possibility that higher-threshold MUs were recruited in the test’s laterphases.
Thomas and del Valle161 subsequently repeatedthis fatiguing protocol on the human biceps brachiiof both intact and spinal cord–injured patients. The
target force of the repetitive contractions was set at50% MVC. The MVC force was tested every 5 minuntil it equaled the target force (i.e., the endurancetime). Multiple single MU firing patterns were re-corded and identified individually. The results on
FIGURE 12. The effect of a repetitive sub-MVC fatigue test ongrouped MU firing patterns. (A) Sketch of the fatiguing protocol(test). The subjects perform repetitive sub-MVCs and inter-posed MVCs of the test muscle (see text for details) until thesustained phase of the MVCs can no longer maintain the targetforce (termed the endurance time, which occurs at the so-called point of fatigue). (This test was first described in 198113
with some refinements introduced subsequently.14,15,17) (B)The effect of this test when applied in five subjects to the thighmuscle, quadriceps, using repetitive 30% sub-MVCs, whichcould be maintained at this levels for �50 min. The test MVCsreduced to 50% of their control value in 35 min (X axis, 50%contractile failure). Plotted are the surface-rectified EMG(SRE) and the mean � SD firing rates (spike frequency) ofdifferent populations of single MUs recorded for very briefperiods at �5-min intervals throughout the test. Also shown at0 time is the mean (�26 HZ) � SD firing rate of anotherpopulation of MUs recorded during control (prefatigue test)MVCs. Mean MU firing rate increased throughout the test and,concomitantly, there was an increase in the surface EMG. Thelatter suggests that the excitatory drive to the active MNsincreased throughout the test. Modified from Bigland-Ritchieand Woods14 (A) and Bigland-Ritchie et al.15 (B) with permis-sion of Blackwell Publishing (B).
150 Motoneurons and Muscle Fatigue MUSCLE & NERVE February 2007
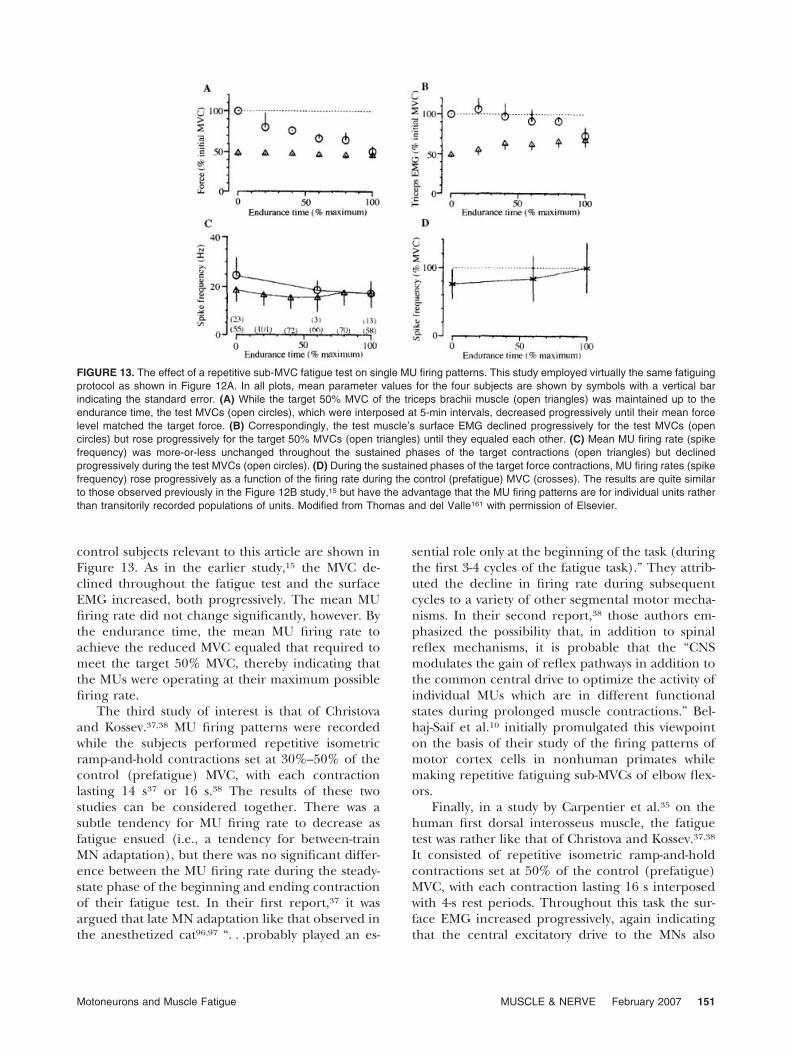
control subjects relevant to this article are shown inFigure 13. As in the earlier study,15 the MVC de-clined throughout the fatigue test and the surfaceEMG increased, both progressively. The mean MUfiring rate did not change significantly, however. Bythe endurance time, the mean MU firing rate toachieve the reduced MVC equaled that required tomeet the target 50% MVC, thereby indicating thatthe MUs were operating at their maximum possiblefiring rate.
The third study of interest is that of Christovaand Kossev.37,38 MU firing patterns were recordedwhile the subjects performed repetitive isometricramp-and-hold contractions set at 30%–50% of thecontrol (prefatigue) MVC, with each contractionlasting 14 s37 or 16 s.38 The results of these twostudies can be considered together. There was asubtle tendency for MU firing rate to decrease asfatigue ensued (i.e., a tendency for between-trainMN adaptation), but there was no significant differ-ence between the MU firing rate during the steady-state phase of the beginning and ending contractionof their fatigue test. In their first report,37 it wasargued that late MN adaptation like that observed inthe anesthetized cat96,97 “. . .probably played an es-
sential role only at the beginning of the task (duringthe first 3-4 cycles of the fatigue task).” They attrib-uted the decline in firing rate during subsequentcycles to a variety of other segmental motor mecha-nisms. In their second report,38 those authors em-phasized the possibility that, in addition to spinalreflex mechanisms, it is probable that the “CNSmodulates the gain of reflex pathways in addition tothe common central drive to optimize the activity ofindividual MUs which are in different functionalstates during prolonged muscle contractions.” Bel-haj-Saif et al.10 initially promulgated this viewpointon the basis of their study of the firing patterns ofmotor cortex cells in nonhuman primates whilemaking repetitive fatiguing sub-MVCs of elbow flex-ors.
Finally, in a study by Carpentier et al.35 on thehuman first dorsal interosseus muscle, the fatiguetest was rather like that of Christova and Kossev.37,38
It consisted of repetitive isometric ramp-and-holdcontractions set at 50% of the control (prefatigue)MVC, with each contraction lasting 16 s interposedwith 4-s rest periods. Throughout this task the sur-face EMG increased progressively, again indicatingthat the central excitatory drive to the MNs also
FIGURE 13. The effect of a repetitive sub-MVC fatigue test on single MU firing patterns. This study employed virtually the same fatiguingprotocol as shown in Figure 12A. In all plots, mean parameter values for the four subjects are shown by symbols with a vertical barindicating the standard error. (A) While the target 50% MVC of the triceps brachii muscle (open triangles) was maintained up to theendurance time, the test MVCs (open circles), which were interposed at 5-min intervals, decreased progressively until their mean forcelevel matched the target force. (B) Correspondingly, the test muscle’s surface EMG declined progressively for the test MVCs (opencircles) but rose progressively for the target 50% MVCs (open triangles) until they equaled each other. (C) Mean MU firing rate (spikefrequency) was more-or-less unchanged throughout the sustained phases of the target contractions (open triangles) but declinedprogressively during the test MVCs (open circles). (D) During the sustained phases of the target force contractions, MU firing rates (spikefrequency) rose progressively as a function of the firing rate during the control (prefatigue) MVC (crosses). The results are quite similarto those observed previously in the Figure 12B study,15 but have the advantage that the MU firing patterns are for individual units ratherthan transitorily recorded populations of units. Modified from Thomas and del Valle161 with permission of Elsevier.
Motoneurons and Muscle Fatigue MUSCLE & NERVE February 2007 151
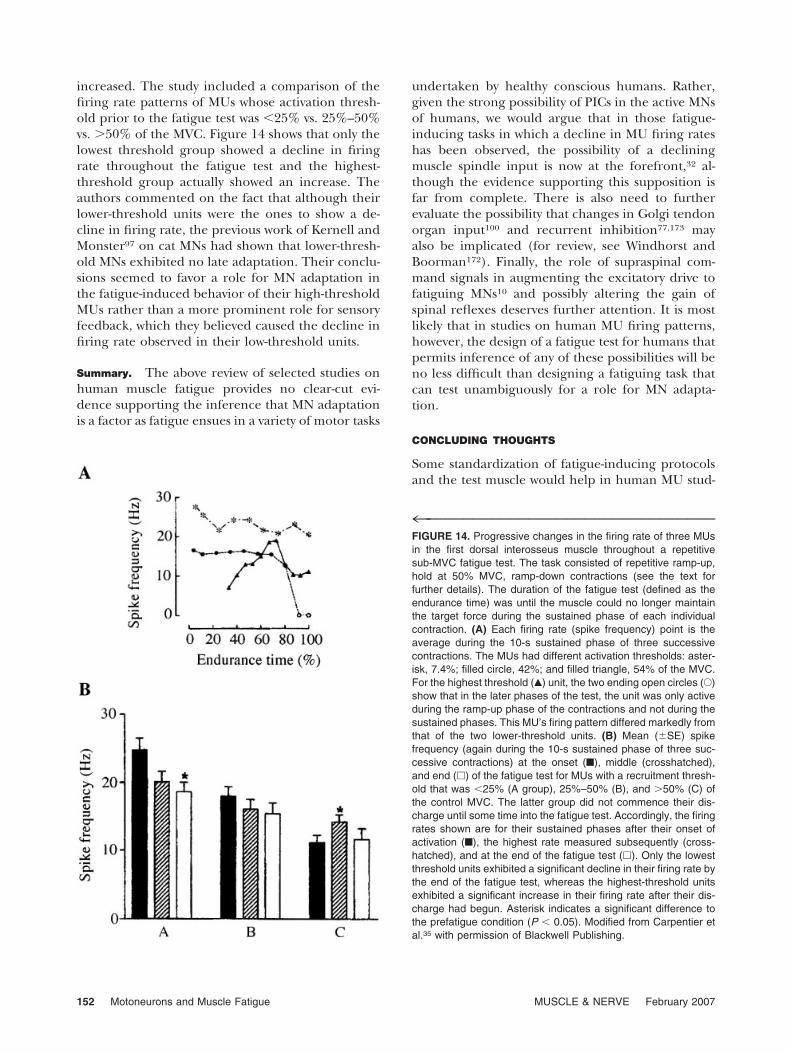
increased. The study included a comparison of thefiring rate patterns of MUs whose activation thresh-old prior to the fatigue test was �25% vs. 25%–50%vs. �50% of the MVC. Figure 14 shows that only thelowest threshold group showed a decline in firingrate throughout the fatigue test and the highest-threshold group actually showed an increase. Theauthors commented on the fact that although theirlower-threshold units were the ones to show a de-cline in firing rate, the previous work of Kernell andMonster97 on cat MNs had shown that lower-thresh-old MNs exhibited no late adaptation. Their conclu-sions seemed to favor a role for MN adaptation inthe fatigue-induced behavior of their high-thresholdMUs rather than a more prominent role for sensoryfeedback, which they believed caused the decline infiring rate observed in their low-threshold units.
Summary. The above review of selected studies onhuman muscle fatigue provides no clear-cut evi-dence supporting the inference that MN adaptationis a factor as fatigue ensues in a variety of motor tasks
undertaken by healthy conscious humans. Rather,given the strong possibility of PICs in the active MNsof humans, we would argue that in those fatigue-inducing tasks in which a decline in MU firing rateshas been observed, the possibility of a decliningmuscle spindle input is now at the forefront,32 al-though the evidence supporting this supposition isfar from complete. There is also need to furtherevaluate the possibility that changes in Golgi tendonorgan input100 and recurrent inhibition77,173 mayalso be implicated (for review, see Windhorst andBoorman172). Finally, the role of supraspinal com-mand signals in augmenting the excitatory drive tofatiguing MNs10 and possibly altering the gain ofspinal reflexes deserves further attention. It is mostlikely that in studies on human MU firing patterns,however, the design of a fatigue test for humans thatpermits inference of any of these possibilities will beno less difficult than designing a fatiguing task thatcan test unambiguously for a role for MN adapta-tion.
CONCLUDING THOUGHTS
Some standardization of fatigue-inducing protocolsand the test muscle would help in human MU stud-
4™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™FIGURE 14. Progressive changes in the firing rate of three MUsin the first dorsal interosseus muscle throughout a repetitivesub-MVC fatigue test. The task consisted of repetitive ramp-up,hold at 50% MVC, ramp-down contractions (see the text forfurther details). The duration of the fatigue test (defined as theendurance time) was until the muscle could no longer maintainthe target force during the sustained phase of each individualcontraction. (A) Each firing rate (spike frequency) point is theaverage during the 10-s sustained phase of three successivecontractions. The MUs had different activation thresholds: aster-isk, 7.4%; filled circle, 42%; and filled triangle, 54% of the MVC.For the highest threshold (Œ) unit, the two ending open circles (E)show that in the later phases of the test, the unit was only activeduring the ramp-up phase of the contractions and not during thesustained phases. This MU’s firing pattern differed markedly fromthat of the two lower-threshold units. (B) Mean (�SE) spikefrequency (again during the 10-s sustained phase of three suc-cessive contractions) at the onset (■), middle (crosshatched),and end (�) of the fatigue test for MUs with a recruitment thresh-old that was �25% (A group), 25%–50% (B), and �50% (C) ofthe control MVC. The latter group did not commence their dis-charge until some time into the fatigue test. Accordingly, the firingrates shown are for their sustained phases after their onset ofactivation (■), the highest rate measured subsequently (cross-hatched), and at the end of the fatigue test (�). Only the lowestthreshold units exhibited a significant decline in their firing rate bythe end of the fatigue test, whereas the highest-threshold unitsexhibited a significant increase in their firing rate after their dis-charge had begun. Asterisk indicates a significant difference tothe prefatigue condition (P � 0.05). Modified from Carpentier etal.35 with permission of Blackwell Publishing.
152 Motoneurons and Muscle Fatigue MUSCLE & NERVE February 2007

ies such that different results from different labora-tories could not be attributed simply to the task-dependency of the fatigue process. Even if such astandardization of human MU research effort was tooccur, however, it is likely that the present gap willcontinue to widen between cellular physiology un-dertaken in animal preparations and systems physi-ology undertaken on MU firing patterns in con-scious humans. One strategy that would help narrowthis gap would be for the cellular approach in animalpreparations to accommodate the difficulties thatare inherent in noninvasive human studies and totake advantage of what the latter studies have con-tributed to current understanding of the neurobiol-ogy of muscle fatigue. Three such approaches aresuggested below.
Stimulus Profile for Studying MN Adaptation. Cur-rently, the animal literature on MN adaptation isrestricted to experiments in which the IC-injectedcurrent to produce repetitive MN discharge hasbeen rectangular in shape. Such abruptly risingcurrents cannot be simulated in human experi-mentation. The reverse is quite feasible, however,in animal research, using both in vitro slices ofspinal cord and in vivo, surgically reduced animalsin both an unanesthetized (decerebrate) and anes-thetized state. Such research can take advantage ofagents that block the mechanisms that are cur-rently thought responsible for the initial and laterphases of adaptation (Fig. 3).
Effects of Descending Monoaminergic Effects on MN
Adaptation. It would be of great interest to repeatthe Kernell and Monster stimulus protocol,96,97
using a decerebrate cat preparation in the absenceand presence of a descending monoaminergicdrive comparable to that degree known to occurduring active muscle contractions (for relevantapproaches, see Heckman et al.82). Such studiesshould include analyses of IC-recorded MN firingpatterns during fictive locomotion that extend onBrownstone’s seminal findings.29,102 In the ab-sence of descending monoaminergic drive, it islikely that the discharge of high-threshold cathindlimb MNs would be like that shown above inFigure 11B (i.e., exhibiting both initial and lateadaptation), whereas no such adaptation would beseen when descending monoaminergic drive gen-erated sufficient PIC to offset it (as in Fig. 10).
Another approach, which is already under way, isto explore the nature of descending monoaminergicinput to the spinal cord during various motor tasksundertaken by freely moving cats,164,165 including
one that produces muscle fatigue.57 Admittedly,these studies have to infer that EC recordings madein brainstem structures are indeed from serotoner-gic neurons and that the recorded cells’ projectionsinclude one to the spinal cord. Nonetheless, therecent finding that the activity of medullary neuronsin n. raphe and n. pallidus diminishes when the catsrun on a treadmill to the point of fatigue is worthy ofspecial consideration.57 Such a result raises the pos-sibility that as fatigue ensues not only does a reduceddescending serotonergic drive dysfacilitate motoroutput but, in addition, it enables MN spike fre-quency adaptation and thereby exacerbates fatigue.The overall exercise-induced role of serotonin andother monoamines in the control of movement isquite multifaceted, however.36,45,48
Effects of Fatigue on the Firing Patterns of MNs and
MUs in Nonhuman Primates. In recent years twoapproaches have considerably advanced the possi-bility of studying cellular mechanisms as the non-human primate elaborates fatiguing contractions.First, the Fetz laboratory showed that it is possibleto obtain EC recordings of spinal interneuron dis-charge and relate such firing patterns to both thefiring patterns of motor cortex cells corticopetallyand MUs corticofugally.55 Second, the Maton lab-oratory has showed that operant conditioning canbe used to generate fatigue in elbow flexor mus-cles and study the firing patterns of motor cortexcells as fatigue ensues.10 A combination of thesetechniques, together with appropriate neurophar-macology to vary the level of descending monoam-inergic drive as monkeys undergo fatiguing con-tractions, could provide answers to several of theissues raised above.
Presented in part by D.G.S. on February 3, 2005 at the SatelliteMotor Control Symposium of the annual meeting of the Austra-lian Neuroscience Society, Perth, West Australia, The authorsthank Drs. Robert Brownstone and Andrew Fuglevand for theirreview of a near-final draft of the article. We also thank Drs. CarloDeLuca, Roger Enoka, Charles Heckman, and Randall Powers fortheir input on selected issues and Drs. David Burke, WilliamLandau, Erik Stålberg, and Robert Young for clinical insights.Finally, we thank Patricia Pierce for technical and editorial help.
NOTE ADDED IN PROOF
Recently, it has been reported that provisionalPICs can be demonstrated in MNs supplying hind-limb muscles in adult rats anesthetized with a mix-ture of ketamine and xylazine (Button et al. JPhysiol (Lond) 2006;573:663– 667) and that thereis an inverse relation between PIC amplitude andthe extent of MN adaptation (fig. 3 in Button et al.
Motoneurons and Muscle Fatigue MUSCLE & NERVE February 2007 153

J Appl Physiol 2006; in press). Future studies willbe required to determine whether these importantfindings generalize across vertebrates.
REFERENCES
1. Adam A, De Luca CJ. Recruitment order of motor units inhuman vastus lateralis muscle is maintained during fatiguingcontractions. J Neurophysiol 2003;90:2919–2927.
2. Adam A, De Luca CJ. Firing rates of motor units in humanvastus lateralis muscle during fatiguing isometric contrac-tions. J Appl Physiol 2005;99:268–280.
3. Alaburda A, Perrier JF, Hounsgaard J. Mechanisms causingplateau potentials in spinal motoneurones. Adv Exp MedBiol 2002;508:219–226.
4. Aoki S, Uchida S, Inoue H. Fatigue-related changes in dis-charge patterns of motor units in the inferior head of thelateral pterygoid muscle in humans. Arch Oral Biol 2005;50:727–737.
5. Aston-Jones G. Locus coeruleus. In: Adelman G, Smith BH,editors. Encyclopedia of neuroscience, 2nd ed. (CD-ROM).Amsterdam: Elsevier BV; 2003.
6. Aston-Jones G, Cohen JD. Adaptive gain and the role of thelocus coerulus-norepinephrine system in optimal perfor-mance. J Comp Neurol 2005;493:99–110.
7. Baldissera F, Gustafsson B. Regulation of repetitive firing inmotoneurones by the afterhyperpolarization conductance.Brain Res 1971;30:431–434.
8. Baldissera F, Gustafsson B. Firing behaviour of a neuronemodel based on the afterhyperpolarization conductancetime course and algebraic summation. Acta Physiol Scand1974;92:27–47.
9. Baldissera F, Gustafsson B, Parmiggiani F. Adaptation in asimple neurone model compared to that in spinal motoneu-rones. Brain Res 1973;52:382–384.
10. Belhaj-Saif A, Fourment A, Maton B. Adapatation of theprecentral cortical commands to elbow muscle fatigue. ExpBrain Res 1996;111:405–416.
11. Bellemare F, Woods JJ, Johansson R, Bigland-Ritchie B. Mo-tor-unit discharge rates in maximal voluntary contractions ofthree human muscles. J Neurophysiol 1983;50:1380–1392.
12. Benda J, Herz AV. A universal model for spike-frequencyadaptation. Neural Comput 2003;15:2523–2564.
13. Bigland-Ritchie B. EMG and fatigue of human voluntary andstimulated contractions. In: Porter R, Whelan J, editors.Human muscle fatigue: physiological mechanisms. London:Pitman Medical; 1981. p. 130–148.
14. Bigland-Ritchie B, Woods JJ. Changes in muscle contractileproperties and neural control during human muscular fa-tigue. Muscle Nerve 1984;7:691–699.
15. Bigland-Ritchie B, Cafarelli E, Vollestad NK. Fatigue of sub-maximal static contractions. Acta Physiol Scand 1986;128(Suppl 556):137–148.
16. Bigland-Ritchie BR, Dawson NJ, Johansson RS, Lippold OC.Reflex origin for the slowing of motoneurone firing rates infatigue of human voluntary contractions. J Physiol (Lond)1986;379:451–459.
17. Bigland-Ritchie BR, Furbush F, Woods JJ. Fatigue of inter-mittent submaximal voluntary contractions: central and pe-ripheral factors. J Appl Physiol 1986;61:421–429.
18. Bigland-Ritchie B, Johansson R, Lippold OC, Smith S,Woods JJ. Changes in motoneurone firing rates during sus-tained maximal voluntary contractions. J Physiol (Lond)1983;340:335–346.
19. Bigland-Ritchie B, Johansson R, Lippold OC, Woods JJ.Contractile speed and EMG changes during fatigue ofsustained maximal voluntary contractions. J Neurophysiol1983;50:313–324.
20. Binder MD. Integration of synaptic and intrinsic dendriticcurrents in cat spinal motoneurons. Brain Res Brain Res Rev2002;40:1–8.
21. Binder MD, Heckman CJ, Powers RK. Relative strengths anddistributions of different sources of synaptic input to themotoneurone pool. Adv Exp Biol Med 2002;508:207–212.
22. Binder MD, Davenport C, Margolis D, Powers RK. Multi-photon fluorescence imaging of calcium fluxes in the den-drites of rat hypoglossal motoneurons. In: Heckman CJ,Enoka RM, editors. Abstracts, International symposium onactive dendrites in motor neurons (http://www.colora-do.edu/intphys/news/ActiveDendrites.html). Boulder: Uni-versity of Colorado; 2004.
23. Bongiovanni LG, Hagbarth K-E. Tonic vibration reflexeselicited during fatigue from maximum voluntary contrac-tions in man. J Physiol (Lond) 1990;423:1–14.
24. Borg J, Grimby L, Hannerz J. The fatigue of voluntary con-traction and peripheral electrical propogation of single mo-tor units in man. J Physiol (Lond) 1983;340:435–444.
25. Braak H, Del Tredici K, Rub U, de Vos RA, Jansen Steur EN,Braak E. Staging of brain pathology related to sporadicParkinson’s disease. Neurobiol Aging 2003;24:197–211.
26. Brock LG, Coombs JS, Eccles JC. The recording of potentialsfrom motoneurones with an intracellular electrode.J. Physiol (Lond) 1952;117:431–460.
27. Brownstone RM. The regulation of the repetitive firing oflumbar motoneurones during fictive locomotion in the cat.PhD Thesis. Winnipeg: University of Manitoba; 1989.
28. Brownstone RM. Beginning at the end: repetitive firingproperties in the final common pathway. Prog Neurobiol2006;78:156–172.
29. Brownstone RM, Jordan LM, Kriellaars DJ, Noga BR, Shef-chyk SJ. On the regulation of repetitive firing in lumbarmotoneurones during fictive locomotion in the cat. ExpBrain Res 1992;90:441–455.
30. Burke D. Critical examination of the case for or againstfusimotor involvement in disorders of muscle tone. In: Des-medt JE, editor. Motor control mechanisms in health anddisease. New York: Raven Press; 1983. p 133–150.
31. Burke RE. Motor units: anatomy, physiology and functionalorganization. In: Brookhart JM, Mountcastle VB, editors.Handbook of physiology, sec. 1, vol. II, pt 1. The nervoussystem: motor control. Bethesda, MD: American Physiologi-cal Society; 1981. p 345–422.
32. Butler JE, Taylor JL, Gandevia SC. Responses of humanmotoneurons to corticospinal stimulation during maximumvoluntary contractions and ischemia. J Neurosci 2003;23:10224–10230.
33. Callister RJ, Sesodia S, Enoka RM, Nemeth PM, ReinkingRM, Stuart DG. Fatigue of rat hindlimb motor units: bio-chemical-physiological associations. Muscle Nerve 2004;30:714–726.
34. Carp JS, Rymer WZ. Enhancement by serotonin of tonicvibration and stretch reflexes in the decerebrate cat. ExpBrain Res 1986;62:111–122.
35. Carpentier A, Duchateau J, Hainaut K. Motor unit behaviorand contractile changes during fatigue in the human firstdorsal interosseus. J Physiol (Lond) 2001;543.3:903–912.
36. Chalouf F. Effects of acute physical exercise on central sero-tonergic systems. Med Sci Sports Exerc 1997;29:58–62
37. Christova P, Kossev A. Motor unit activity during long-lastingintermittent muscle contractions in humans. Eur J ApplPhysiol Occup Physiol 1998;77:379–387.
38. Christova P, Kossev A. Human motor unit recruitment andderecruitment during long lasting intermittent contractions.J Electromyogr Kinesiol 2001;11:189–196.
39. Clamann HP, Robinson AJ. A comparison of electromyo-graphic and mechanical fatigue properties in motor units ofthe cat hindlimb. Brain Res 1985;327:203–219.
40. Collins DF, Burke D, Gandevia SC. Large involuntary forcesconsistent with plateau-like behavior of human motoneu-rons. J Neurosci 2001;21:4059–4065.
154 Motoneurons and Muscle Fatigue MUSCLE & NERVE February 2007

41. Collins DF, Burke D, Gandevia SC. Sustained contractionsproduced by plateau-like behaviour in human motoneu-rones. J Physiol (Lond) 2002;538:289–301.
42. Collins DF, Gorassini M, Bennett D, Burke D, Gandevia SC.Recent evidence for plateau potentials in human motoneu-rones. Adv Exp Med Biol 2002;508:227–235.
43. Creed RS, Denny-Brown D, Eccles JC, Liddell EGT, Sher-rington CS. Reflex activity of the spinal cord. London: Ox-ford University Press; 1932.
44. Cullheim S, Fleshman JW, Glenn LL, Burke RE. Membranearea and dendritic structure in type-identified triceps suraealpha motoneurons. J Comp Neurol 1987;255:68–81.
45. Davis JM, Bailey SP. Possible mechanisms of central nervoussystem fatigue during exercise. Med Sci Sports Exerc 1997;29:45–57.
46. De Luca CJ. Control properties of motor units. J Exp Biol1985;329:125–136.
47. Denny-Brown D, Pennybacker JB. Fibrillation and fascicula-tion in voluntary muscle. Brain 1938;61:311–334.
48. Dishman RK. Brain monamines, exercise, and behavioralstress: animal models. Med Sci Sports Exerc 1997;29:63–74.
49. Dorfman LJ, Howard JE, McGill KC. Triphasic behavioralresponse of motor units to submaximal fatiguing exercise.Muscle Nerve 1990;13:621–628.
50. Duchateau J, Balestra C, Carpentier A, Hainaut K. Reflexregulation during sustained and intermittent submaximalcontractions in humans. J Physiol (Lond) 2002;541.3:959–967.
51. Eccles JC, McGeer PL. Ionotropic and metabotropic synaptictransmission. Trends Neurosci 1979;2:39–40.
52. Enoka RM, Stuart DG. Neurobiology of muscle fatigue.J Appl Physiol 1992;72:1631–1648.
53. Enoka RM, Robinson GA, Kossev AR. Task and fatigue ef-fects on low-threshold motor units in human hand muscle.J Neurophysiol 1989;62:1344–1359.
54. Fatt P, Katz B. An analysis of the end-plate potential re-corded with an intra-cellular electrode. J Physiol (Lond)1951;115:320–370.
55. Fetz EE, Perlmutter SI, Prut Y, Seki K, Votaw S. Roles ofprimate spinal interneurons in preparation and execution ofvoluntary hand movement. Brain Res Brain Res Rev 2002;40:53–65.
56. Forbes A. The interpretation of spinal reflexes in terms ofpresent knowledge of nerve conduction. Physiol Rev 1922;2:361–414.
57. Fornal C, Martın-Cora FJ, Jacobs BL. “Fatigue” of medullarybut not mesencephalic raphe serotonergic neurons duringlocomotion in cats. Brain Res 2006;1072:55–61.
58. Freund H-J. Motor unit and muscle activity in voluntarymotor control. Physiol Rev 1983;63:387–436.
59. Fuglevand AJ, Keen DA. Re-evaluation of muscle wisdom inthe human adductor pollicis using physiological rates ofstimulation. J Physiol (Lond) 2003;549:865–875.
60. Fuglevand AJ, Dutoit AP, Johns RK, Keen DA. Evaluation ofplateau-potential mediated ’warm up’ in human motorunits. J Physiol (Lond) 2006;571:683–693.
61. Gandevia SC. Spinal and supraspinal factors in muscle fa-tigue. Physiol Rev 2001;81:1725–1789.
62. Gandevia SC, Enoka RM, McComas AJ, Stuart DG, ThomasCK. Neurobiology of muscle fatigue: advances and issues.Adv Exp Biol Med 1995;384:515–525.
63. Gantchev GN, Gatev P, Ivanova T, Tankov N. Motor unitactivity during different functional states of the neuromus-cular system. Biomed Biochim Acta 1986;45:S69–S75.
64. Garland SJ, Enoka RM, Serrano LP, Robinson GA. Behaviorof motor units in human biceps brachii during a submaximalfatiguing contraction. J Appl Physiol 1994;76:2411–2419.
65. Garland SJ, Griffin L, Ivanova T. Motor unit discharge rate isnot associated with muscle relaxation time in sustained sub-maximal contractions in humans. Neurosci Lett 1997;239:25–28.
66. Gatev P, Ivanova T, Gantchev GN. Changes in the firingpattern of high-threshold motor units due to fatigue. Elec-tromyogr Clin Neurophysiol. 1986;26:83–93.
67. Gerin C, Privat A. Direct evidence of a link betweenmonaminergic descending pathways and motor activity. II. Astudy with microdialysis probes implanted in the ventralhorn of the spinal cord. Brain Res 1988;794:169–173.
68. Gorassini M, Yang JF, Siu M, Bennett DJ. Intrinsic activationof human motoneurons: possible contribution to motor unitexcitation. J Neurophysiol 2002;87:1850–1858.
69. Gorassini M, Yang JF, Siu M, Bennett DJ. Intrinsic activationof human motoneurons: reduction of motor unit recruit-ment thresholds by repeated contractions. J Neurophysiol2002;87:1859–1866.
70. Gorman RB, McDonagh JC, Hornby TG, Reinking RM, Stu-art DG. Measurement and nature of firing rate adaptation inturtle spinal neurons. J Comp Physiol A 2005;191:583–603.
71. Granit R, Kernell D, Shortess GK. Quantitative aspects ofrepetitive firing of mammalian motoneurones, caused byinjected currents. J Physiol (Lond) 1963;168:911–931.
72. Greengaard P. Possible role for cyclic nucleotides and phos-phorylated membrane proteins in postsynaptic actions ofneurotransmitters. Nature 1976;160:101–108.
73. Gribble PL, Mullin LI, Cothros N, Mattar A. Role of cocon-traction in arm movement accuracy. J Neurophysiol 2003;89:2396–2405.
74. Grillner S, Wallen P. Innate versus learned movements—afalse dichotomy? Prog Brain Res 2004;143:3–12.
75. Grimby L, Hannerz J, Hedman B. The fatigue and voluntarydischarge properties of single motor units in man. J Physiol(Lond) 1981;316:545–554.
76. Hammar I, Bannatyne BA, Maxwell DJ, Edgley SA,Jankowska E. The actions of monoamines and distribution ofnoradrenergic and serotoninergic contacts on different sub-populations of commissural interneurons in the cat spinalcord. Eur J Neurosci 2004;19:1305–1316.
77. Hayward L, Breitbach D, Rymer WZ. Increased inhibitoryeffects on close synergists during muscle fatigue in the de-cerebrate cat. Brain Res 1988;440:199–203.
78. Heckman CJ. Introduction to motor neuron properties andthe generation of firing patterns. In: Heckman CJ, EnokaRM, editors. Abstracts, International symposium on activedendrites (http://www.colorado.edu/intphys/news/Active-Dendrites.html). Boulder: University of Colorado; 2004.
79. Heckman CJ, Enoka RM, editors. International symposiumon active dendrites in motor neurons (http://www.colo-rado. edu/intphys/news/ActiveDendrites.html). Boulder:University of Colorado; 2004.
80. Heckman CJ, Gorassini MA, Bennett DJ. Persistent inwardcurrents in motoneuron dendrites: implications for motoroutput. Muscle Nerve 2005;31:135–156.
81. Heckman CJ, Kuo JJ, Ballou EW, Johnson MD, HyngstromAJ. Roles of dendritic and somatic components of the per-sistent inward currents in controlling motoneuron firing. In:Heckman CJ, Enoka RM, editors. Abstracts, Internationalsymposium on active dendrites (http://www.colorado.edu/intphys/news/ActiveDendrites.html). Boulder: University ofColorado; 2004.
82. Heckman CJ, Lee RH, Brownstone RM. Hyperexcitable den-dites in motoneurons and their neuromodulatory controlduring motor behavior. Trends Neurosci 2003;26:688–695.
83. Hochman S, Garraway SM, Machacek DW, Shay BL. 5-HTreceptors and the neuromodulatory control of spinal cordfunction. In: Cope TC, editor. Motor neurobiology of thespinal cord. Boca Raton, FL: CRC Press; 2001. p 47–87.
84. Hornby TG, McDonagh JC, Reinking RM, Stuart DG. Mo-toneurons: a preferred firing range across vertebrate spe-cies? Muscle Nerve 2002;25:632–648.
85. Hornby TG, McDonagh JC, Reinking RM, Stuart DG. Effectsof excitatory modulation on intrinsic properties of turtlemotoneurons. J Neurophysiol 2002;88:86–97.
Motoneurons and Muscle Fatigue MUSCLE & NERVE February 2007 155

86. Hornby TG, Rymer WZ, Benz EN, Schmit BD. Windup offlexion reflexes in chronic human spinal cord injury: amarker for neuronal plateau potentials? J Neurophysiol2003;89:416–426.
87. Hounsgaard J. Motoneurons do what motoneurons have todo. J Physiol (Lond) 2002;538.1:4.
88. Hultborn H, Brownstone RB, Toth TI, Gossard JP. Keymechanisms for setting the input-output gain across themotoneuron pool. Prog Brain Res 2004;143:77–95.
89. Hunter SK, Duchateau J, Enoka RM. Muscle fatigue and themechanisms of task failure. Exerc Sport Sci Rev 2004;32:44–49.
90. Illert M, Kummel H. Reflex pathways from large musclespindle afferents and recurrent axon collaterals to motoneu-rones of wrist and digit muscles: a comparison in cats, mon-keys and humans. Exp Brain Res 1999;128:13–19.
91. Ito M, Oshima T. Temporal summation of after-hyperpolar-ization following a motoneurone spike. Nature 1962;195:910–911.
92. Jacobs BL, Martin-Cora FJ, Fornal CA. Activity of medullaryserotonergic neurons in freely moving animals. Brain ResBrain Res Rev 2002;40:45–52.
93. Johnson MA, Polgar J, Weightman D, Appleton D. Data onthe distribution of fibre types in thirty-six human muscles —an autopsy study. J Neurol Sci 1973;18:111–129.
94. Kawakami Y, Amemiya K, Kanehisa H, Ikegawa S, FukunagaT. Fatigue responses of human triceps surae muscles duringrepetitive maximal isometric contractions. J Appl Physiol2000;88:1969–1975.
95. Keesey JC. Myasthenia gravis, an illustrated history. Roseville,CA: Publishers Design Group; 2002.
96. Kernell D, Monster AW. Time course and properties of lateadaptation in spinal motoneurones of the cat. Exp Brain Res1982;46:191–196.
97. Kernell D, Monster AW. Motoneurone properties and motorfatigue. An intracellular study of gastrocnemius motoneu-rones of the cat. Exp Brain Res 1982;46:197–204.
98. Kernell D, Sjoholm H. Repetitive impulse firing: compari-sons between neurone models based on ’voltage clamp equa-tions’ and spinal motoneurones. Acta Physiol Scand 1973;87:40–56.
99. Kiehn O, Eken T. Functional role of plateau potentials invertebrate motor neurons. Curr Opin Neurobiol 1998;8:746–752.
100. Kirsch RF, Rymer WZ. Neural compensation for muscularfatigue: evidence for significant force regulation in man.J Neurophysiol 1987;57:1893–1910.
101. Krawitz S, Brownstone RM. Late adaptation in cat motoneu-rones during fictive locomotion. Soc Neurosci Abstr 1994;241.8.
102. Krawitz S, Brownstone RM, Noga BR, Jordan LM. Can thenervous system overcome a possible central fatigue process— late adaptation? Muscle Nerve Suppl 1996;4:S52.
103. Krawitz S, Fedirchuk B, Dai Y, Jordan LM, McCrea DA.State-dependent hyperpolarization of voltage threshold en-hances motoneurone excitability during fictive locomotionin the cat. J Physiol (Lond) 2001;532:271–281.
104. Krnjevic K, Miledi R. Failure of neuromuscular propagationin rats. J Physiol (Lond) 1958;140:440–461.
105. Krnjevic K, Miledi R. Presynaptic failure of neuromuscularpropagation in rats. J Physiol (Lond) 1959;149:1–22.
106. Kuchinad RA, Ivanova TD, Garland SJ. Modulation of motorunit discharge rate and H-reflex amplitude during submaxi-mal fatigue of the human soleus muscle. Exp Brain Res2004;158:345–355.
107. Kudina LP, Person RS. “Split” motor unit potentials of thehuman muscle. Fiziol Zhurnal SSSR 1971;57:38–54.
108. Laidlaw DH, Hunter SK, Enoka RM. Nonuniform activationof the agonist muscle does not covary with index fingeracceleration in old adults. J Appl Physiol 2002;93:1400–1410.
109. Landau WM. Re editorial: clinical management of spasticity.J Neurol Neurosurg Psychiatry. E-letter to editor, August 26,2005.
110. Lee RH, Heckman CJ. Bistability of spinal motoneurons invivo: systematic variations in rhythmic firing patterns. J Neu-rophysiol 1998;80:572–582.
111. Lee RH, Heckman CJ. Enhancement of bistability in spinalmotoneurons in vivo by the noradrenergic �1 agonist me-thoxamine. J Neurophysiol 1999;81:2164–2174.
112. Lee RH, Heckman CJ. Adjustable amplification of synapticinput in the dendrites of spinal motoneurons in vivo. J Neu-rosci 2000;20:6734–6740.
113. Lemon RN, Griffiths J. Comparing the function of the cor-ticospinal system in different species: organizational differ-ences for motor specialization? Muscle Nerve 2005;32:261–279.
114. Li Y, Gorassini MA, Bennett DJ. Role of persistent sodiumand calcium currents in motoneuron firing and spasticity inchronic spinal rats. J Neurophysiol 2004;91:767–783.
115. Libet B, Kobayashi H, Tanaka T. Synaptic coupling into theproduction and storage of a neuronal memory trace. Nature1975;258:155–157.
116. Lindsay AD, Heckman CJ, Binder MD. Analysis of late adap-tation in cat motoneurons. Soc Neurosci Abstr 1986; p 247.
117. Llinas R, Lopez-Barneo J. Electrophysiology of mammaliantectal neurons in vitro. I. Transient ionic conductances.J Neurophysiol 1998;60:853–867.
118. Llinas R, Lopez-Barneo J. Electrophysiology of mammaliantectal neurons in vitro. II. Long-term adaptation. J Neuro-physiol 1988;60:869–878.
119. Loeb GE. Hard lessons in motor control from the mamma-lian spinal cord. Trends Neurosci 1987;10:108–113.
120. Macefield VG, Fuglevand AJ, Bigland-Ritchie B. Contractileproperties of single motor units in human toe extensorsassessed by intrneural motor axon stimulation. J Neuro-physiol 1996;75:2509–2519.
121. Macefield VG, Fuglevand AJ, Howell JN, Bigland-Ritchie B.Discharge behavior of single motor units during maximalvoluntary contractions of a human toe muscle. J Physiol(Lond) 2000;528:227–234.
122. Macefield VG, Hagbarth KE, Gorman R, Gandevia SC, BurkeD. Decline in spindle support to alpha-motoneurones dur-ing sustained voluntary contractions. J Physiol (Lond) 1991;440:497–512.
123. Marsden CD, Meadows JC, Merton PA. Isolated single motorunits in human hand muscle and their rate of dischargeduring maximum voluntary effort. J Physiol (Lond) 1971;217:12–13P.
124. Marsden CD, Meadows JC, Merton PA. “Muscular wisdom”that minimizes fatigue during prolonged effort in man: peakrates of motoneuron discharge and slowing of dischargeduring fatigue. Adv Neurol 1983;39:169–211.
125. Martin RF, Jordan LM, Willis WD. Differential projections ofcat medullary raphe neurons demonstrated by retrogradelabelling following spinal cord lesions. J Comp Neurol 1978;182:77–88.
126. Maton B, Gamet D. The fatigability of two antagonisticmuscles in human isometric voluntary submaximal con-traction: an EMG study. II. Motor unit firing rate andrecruitment. Eur J Appl Physiol Occup Physiol 1989;58:369 –374.
127. McComas AJ. Neuromuscular function and disorders. Lon-don: Butterworths; 1987.
128. McGeer PL, Eccles JC, McGeer EG. Molecular neurobiologyof the mammalian brain. New York: Plenum Press; 1978.
129. Meyers BM, Caferelli E. Caffeine increases time to fatigue bymaintaining force and not by altering firing rates dringsubmaximal isometric contractions. J Appl Physiol 2005;99:1056–1063.
130. Miles GB, Dai Y, Brownstone RM. Mechanisms underlyingthe early phase of spike frequency adaptation in mouse
156 Motoneurons and Muscle Fatigue MUSCLE & NERVE February 2007

spinal motoneurones. J Physiol (Lond) 2005;566:519 –532.
131. Morales F, Chase MH. Postsynaptic control of lumbar mo-toneuron excitability during active sleep in the chronic cat.Brain Res 1981;225:279–295.
132. Mori S, Stuart DG, Wiesendanger M (eds.). Brain mecha-nisms for the integration of posture and movement. Progressin brain research, vol. 143. Amsterdam: Elsevier Science;2004.
133. Mori S, Nakajima K, Mori F, Matsuyama K. Integration ofmultiple motor segments for the elaboration of locomotion:role of the fastigial nucleus of the cerebellum. Prog BrainRes 2004;143:341–351.
134. Mottram CJ, Jakobi JM, Semmler JG, Enoka RM. Motor-unitactivity differs with load type during fatiguing contractions.J Neurophysiol 2005;93:1381–1392.
135. Nickolls P, Collins DF, Gorman RB, Burke D, Gandevia SC.Forces consistent with plateau-like behaviour of spinal neu-rons evoked in patients with spinal cord injuries. Brain 2004;127:660–670.
136. Nordlund MM, Thorstensson A, Cresswell AG. Central andperipheral contributions to fatigue in relation to level ofactivation during repeated maximal voluntary isometricplantar flexions. J Appl Physiol 2004;96:218–225.
137. Nordstrom MA, Miles TS. Instability of motor unit firingrates during prolonged isometric contractions in humanmasseter. Brain Res 1991;549:268–274.
138. Person RS, Kudina LP. The frequency of human motorneurons impulsation during voluntary muscle contraction.Neirofiziologiia 1971;3:200–209.
139. Person RS, Kudina LP. Discharge frequency and dischargepattern of human motor unit during voluntary contractionof muscle. Electroencephalogr Clin Neurophysiol 1972;32:471–483.
140. Peters EJ, Fuglevand AJ. Cessation of human motor unitdischarge during sustained maximal voluntary contraction.Neurosci Lett 1999;274:66–70.
141. Pierrot-Deseilligny E, Burke D. The circuitry of the humanspinal cord: its role in motor control and movement disor-ders. Cambridge, UK: Cambridge University Press; 2005.
142. Powers RK, Binder MD. Input-output functions of mamma-lian motoneurons. Rev Physiol Biochem Pharmacol 2001;143:137–263.
143. Powers RK, Sawczuk A, Musick JR, Binder MD. Multiplemechanisms of spike-frequency adaptation in motoneu-rones. J Physiol (Paris) 1999;93:101–114.
144. Presland JD, Dowson MN, Cairns SP. Changes of motordrive, cortical arousal and perceived exertion following pro-longed cycling to exhaustion. Eur J Appl Physiol 2005;95:42–51.
145. Rajkowski J, Majczynski H, Clayton E, Aston-Jones G. Activa-tion of locus coeruleus neurons varies with difficulty andperformance in a target slection task. J Neurophysiol 2004;92:361–371.
146. Sah P. Ca2�-activated K� currents in neurones: types, phys-iological roles and modulation. Trends Neurosci 1996;19:150–154.
147. Sawczuk A, Powers RK, Binder MD. Spike frequency adap-tation studied in hypoglossal motoneurons of the rat. J Neu-rophysiol 1995;73:1799–1810.
148. Sawczuk A, Powers RK, Binder MD. Contribution of out-ward currents to spike-frequency adaptation in hypoglos-sal motoneurons of the rat. J Neurophysiol 1997;78:2246 –2253.
149. Sayer RJ, Brown AM, Schwindt PC, Crill WE. Calcium cur-rents in acutely isolated neocortical neurons. J Neurophysiol1993;69:1596–1606.
150. Schwindt PC, Crill WE. A persistent negative resistance in catlumbar motoneurons. Brain Res 1977;120:173–178.
151. Sherrington CS. Integrative action of the nervous system.New York: Charles Scribner’s Sons; 1906.
152. Sieck GC, Prakash YS. Fatigue at the neuromuscular junc-tion: branch point vs. presynaptic vs. postsynaptic mecha-nisms. Adv Exp Biol Med 1995;384:83–100.
153. Spielmann JM. Quantification of motor-neuron adaptationto sustained and intermittent stimulation. PhD Thesis. Tuc-son: University of Arizona; 1991.
154. Spielmann JM, Laouris Y, Nordstrom MA, Reinking RM,Stuart DG. The relation between motor-neuron adaptationand muscle fatigue. Abstracts, Third IBRO World CongrNeurosci, Montreal, Canada, 1991; p 407.
155. Spielmann JM, Laouris Y, Nordstrom MA, Robinson GA,Reinking RM, Stuart DG. Adaptation of cat motoneurons tosustained and intermittent extracellular activation. J Physiol(Lond) 1993;464:75–120.
156. Stålberg E, Thiele B. Transmission block in terminal nervetwigs: a single fibre electromyographic finding in man.J Neurol Neurosurg Psychiatry 1972;35:52–59.
157. Stålberg E, Trontelj JV. Demonstration of axon reflexes inhuman motor nerve fibres. J Neurol Neurosurg Psychiatry1970;33:571–579.
158. Stålberg E, Trontelj JV. Single fiber electromyography —studies in healthy and diseased muscle, 2nd ed. New York:Raven Press; 1994.
159. Stuart DG, McDonagh JC. Reflections on a Bernsteinianapproach to systems neuroscience: the controlled treadmilllocomotion of high decerebrate cats. In: Latash ML, editor.Progress in motor control: Bernstein’s traditions in move-ment studies. Champaign, IL: Human Kinetics; 1998. p21–49.
160. Takakusaki K, Oohinata-Sugimoto J, Saitoh K, Habaguchi T.Role of basal ganglia-brainstem systems in the control ofpostural muscle tone and locomotion. Prog Brain Res 2004;143:231–237.
161. Thomas CK, del Valle A. The role of motor unit modulationversus recruitment in repeated submaximal voluntary con-tractions performed by control and spinal cord injured sub-jects. J Electromyogr Kinesiol 2001;11:217–229.
162. Thompson AJ, Jarrett L, Lockley L, Marsden J, StevensonVL. Clinical management of spasticity. J Neurol NeurosurgPsychiatry 2005;76:459–463.
163. Turkin VV, Monroe KS, Hamm TM. Organization of re-current inhibition and facilitation in motor nuclei inner-vating ankle muscles of the cat. J Neurophysiol 1998;79:778 –790.
164. Veasey SC, Fornal CA, Metzler CW, Jacobs BL. Responses ofserotonergic caudal raphe neurons in relation to specificmotor activities in freely moving cats. J Neurosci 1995;15:5346–5359.
165. Veasey SC, Fornal CA, Metzler CW, Jacobs BL. Single-unitresponses of serotonergic dorsal raphe neurons to specificmotor challenges in freely moving cats. Neuroscience 1997;79:161–169.
166. Waldrop TG, Eldredge FL, Iwamoto G, Mitchell JH. Centralneural control of respiration and circulation during exer-cise. In: Rowell LB, Shepherd JT, editors. Handbook ofphysiology, sect. 12. Exercise: regulation and integration ofmultiple systems. New York: Oxford University Press; 1996. p333–380.
167. Walton C, Kalmar JM, Cafarelli E. Effect of caffeine onself-sustained firing in human motor units. J Physiol (Lond)2002;545.2:671–679.
168. Westad C, Westgaard RH, De Luca CJ. Motor unit recruit-ment and derecruitment induced by brief increase in con-traction amplitude of the human trapezius muscle. J Physiol(Lond) 2003;552.2:645–656.
169. Westgaard RH, De Luca CJ. Motor unit substitution in longduration contractions of the human trapezius muscle. J Neu-rophysiol 1999;82:501–504.
170. Westgaard RH, De Luca CJ. Motor control of low-thresholdmotor units in the human trapezius muscle. J Neurophysiol2001;85:1777–1781.
Motoneurons and Muscle Fatigue MUSCLE & NERVE February 2007 157

171. Windhorst W. On the role of recurrent inhibitory feedbackin motor control. Prog Neurobiol 1996;49:517–587.
172. Windhorst U, Boorman G. Overview: potential role of seg-mental motor circuitry in muscle fatigue. Adv Exp Med Biol1995;384:241–258.
173. Windhorst U, Kokkoroyiannis T. Interaction of recurrentinhibitory and muscle spindle afferent feedback during mus-cle fatigue. Neuroscience 1991;43:249–259.
174. Wood SJ, Slater CR. Safety factor at the neuromuscularjunction. Prog Neurobiol 2001;64:393–429.
175. Woods JJ, Furbush F, Bigland-Ritchie B. Evidence for a fa-tigue-induced reflex inhibition of motoneuron firing rates.J Neurophysiol 1987;58:125–137.
176. Zeng J, Powers RK, Newkirk G, Yonkers M, Binder MD. Contribu-tion of persistent sodium currents to spike-frequency adaptation inrat hypoglossal motoneurons. J Neurophysiol 2005;93:1035–1041.
158 Motoneurons and Muscle Fatigue MUSCLE & NERVE February 2007